
Molecular techniques for pathogen identification and ... · Molecular techniques for pathogen...
13
ARTICLE VOLUME 2 · NO. 2 177 © 2011 International Mycological Association You are free to share - to copy, distribute and transmit the work, under the following conditions: Attribution: You must attribute the work in the manner specified by the author or licensor (but not in any way that suggests that they endorse you or your use of the work). Non-commercial: You may not use this work for commercial purposes. No derivative works: You may not alter, transform, or build upon this work. For any reuse or distribution, you must make clear to others the license terms of this work, which can be found at http://creativecommons.org/licenses/by-nc-nd/3.0/legalcode. Any of the above conditions can be waived if you get permission from the copyright holder. Nothing in this license impairs or restricts the author’s moral rights. INTRODUCTION Fungi represent the greatest eukaryotic diversity on earth and they are among the primary decomposers in ecosystems. It is conservatively estimated that 1.5 million species of fungi exist (Hawksworth 1991). Many fungal species are important plant and human pathogens (Agrios 2005). Rapid and accurate detection of fungal pathogens to species or strain level is often essential for disease surveillance and implementing a disease management strategy. Developing direct detection assays is challenging because fungal pathogens can exist as multiple species complexes or at very low concentration in clinical and natural environments. Different molecular genotypes/varieties can also exist within species, and may have different pathogenic profiles and virulence levels to the host. In addition, unculturable and non-sporulating fungi remain a major challenge when studying biotrophic, endophytic, and mycorrhizal groups. Therefore, novel techniques are required when attempting to detect fungi in the environment. Increasingly molecular techniques are employed in studies requiring the detection of fungi in the environment. In this paper, based on presentations at a Special Interest Group meeting convened during the International Mycological Congress (IMC9) in Edinburgh, UK, August 2010, some of the latest diagnostic techniques employed in the detection of fungi, including fluorescence in situ hybridisation (FISH), DNA array technology, Multiplex tandem PCR, and Padlock probe technology with rolling circle amplification and loop-mediated isothermal amplification (LAMP) were discussed. The importance of DNA extraction and sampling methodologies were also briefly discussed. Molecular techniques for pathogen identification and fungus detection in the environment Clement K.M. Tsui 1 , James Woodhall 2 , Wen Chen 3 , C. André Lévesque 3 , Anna Lau 4 *, Cor D. Schoen 5 , Christiane Baschien 6** , Mohammad J. Najafzadeh 7,8 , and G. Sybren de Hoog 7 1 Department of Forest Sciences, The University of British Columbia, Vancouver, BC, V6T 1Z4, Canada; corresponding author e-mail: [email protected] 2 The Food and Environment Research Agency, Sand Hutton, York YO41 1LZ, UK 3 Central Experimental Farm, Agriculture and Agri-Food Canada, Ottawa, Canada, K1A OC6 4 Centre for Infectious Diseases and Microbiology and the University of Sydney, Westmead Hospital, Westmead, NSW 2145, Australia 5 Plant Research International, Business Unit Bio-Interactions and Plant Health, PO Box 16, 6700 AA, Wageningen, The Netherlands 6 Technische Universität Berlin, Environmental Microbiology, Sekr. FR1-2, Franklinstrasse 29, 10587 Berlin, Germany 7 CBS-KNAW Fungal Biodiversity Centre, Uppsalalaan 8, 3584 CT Utrecht, The Netherlands 8 Department of Parasitology and Mycology, School of Medicine, Mashhad University of Medical Sciences, Mashhad, Iran *Current mailing address: Department of Laboratory Medicine, 10 Center Drive, National Institutes of Health, Bethesda, MD 20892, USA **Current mailing address: Federal Environment Agency Germany, Corrensplatz 1, 14195 Berlin, Germany Abstract: Many species of fungi can cause disease in plants, animals and humans. Accurate and robust detection and quantification of fungi is essential for diagnosis, modeling and surveillance. Also direct detection of fungi enables a deeper understanding of natural microbial communities, particularly as a great many fungi are difficult or impossible to cultivate. In the last decade, effective amplification platforms, probe development and various quantitative PCR technologies have revolutionized research on fungal detection and identification. Examples of the latest technology in fungal detection and differentiation are discussed here. Article info: Submitted 17 October 2011;Accepted 3 November 2011; Published 18 November 2011. Key words: FISH LAMP macroarray medical mycology molecular diagnostics molecular ecology padlock probe pathogenic fungi plant pathology rolling circle amplification doi:10.5598/imafungus.2011.02.02.09 IMA FUNGUS · VOLUME 2 · NO 2: 177–189
Transcript of Molecular techniques for pathogen identification and ... · Molecular techniques for pathogen...

ARTIC
LE
v o l u m e 2 · n o . 2 177
© 2011 International Mycological Association
You are free to share - to copy, distribute and transmit the work, under the following conditions:Attribution: Youmustattributetheworkinthemannerspecifiedbytheauthororlicensor(butnotinanywaythatsuggeststhattheyendorseyouoryouruseofthework). Non-commercial: Youmaynotusethisworkforcommercialpurposes.No derivative works: Youmaynotalter,transform,orbuilduponthiswork.For any reuse or distribution, you must make clear to others the license terms of this work, which can be found at http://creativecommons.org/licenses/by-nc-nd/3.0/legalcode. Any of the above conditions can be waived if you get permission from the copyright holder. Nothing in this license impairs or restricts the author’s moral rights.
INtroductIoN
Fungi represent the greatest eukaryotic diversity on earth and theyareamongtheprimarydecomposersinecosystems.Itisconservativelyestimatedthat1.5millionspeciesoffungiexist(Hawksworth1991).Manyfungalspeciesareimportantplantand human pathogens (Agrios 2005). Rapid and accuratedetection of fungal pathogens to species or strain level is often essential for disease surveillance and implementing a diseasemanagement strategy. Developing direct detectionassays is challenging because fungal pathogens can existasmultiplespeciescomplexesoratvery lowconcentrationin clinical and natural environments. Different moleculargenotypes/varieties canalsoexistwithin species,andmayhave different pathogenic profiles and virulence levels tothe host. In addition, unculturable and non-sporulatingfungi remain a major challenge when studying biotrophic,
endophytic, and mycorrhizal groups. Therefore, noveltechniques are required when attempting to detect fungi in theenvironment.
Increasingly molecular techniques are employed instudiesrequiringthedetectionoffungiintheenvironment.In this paper, based on presentations at a SpecialInterestGroupmeetingconvenedduringtheInternationalMycological Congress (IMC9) in Edinburgh, UK, August2010,someof the latestdiagnostic techniquesemployedin the detection of fungi, including fluorescence in situ hybridisation (FISH), DNA array technology, Multiplextandem PCR, and Padlock probe technology withrolling circle amplification and loop-mediated isothermalamplification (LAMP)were discussed.The importanceofDNA extraction and sampling methodologies were alsobrieflydiscussed.
Molecular techniques for pathogen identification and fungus detection in the environmentClementK.M.Tsui1, James Woodhall2,WenChen3,C.AndréLévesque3,AnnaLau4*,CorD.Schoen5,ChristianeBaschien6**, MohammadJ.Najafzadeh7,8,andG.SybrendeHoog7
1Department of Forest Sciences, The University of British Columbia, Vancouver, BC, V6T 1Z4, Canada; corresponding author e-mail:[email protected],SandHutton,YorkYO411LZ,UK3CentralExperimentalFarm,AgricultureandAgri-FoodCanada,Ottawa,Canada,K1AOC64CentreforInfectiousDiseasesandMicrobiologyandtheUniversityofSydney,WestmeadHospital,Westmead,NSW2145,Australia5PlantResearchInternational,BusinessUnitBio-InteractionsandPlantHealth,POBox16,6700AA,Wageningen,TheNetherlands6TechnischeUniversitätBerlin,EnvironmentalMicrobiology,Sekr.FR1-2,Franklinstrasse29,10587Berlin,Germany7CBS-KNAWFungalBiodiversityCentre,Uppsalalaan8,3584CTUtrecht,TheNetherlands8DepartmentofParasitologyandMycology,SchoolofMedicine,MashhadUniversityofMedicalSciences,Mashhad,Iran*Currentmailingaddress:DepartmentofLaboratoryMedicine,10CenterDrive,NationalInstitutesofHealth,Bethesda,MD20892,USA**Currentmailingaddress:FederalEnvironmentAgencyGermany,Corrensplatz1,14195Berlin,Germany
Abstract: Manyspeciesoffungicancausediseaseinplants,animalsandhumans.Accurateandrobustdetection and quantification of fungi is essential for diagnosis, modeling and surveillance.Also directdetection of fungi enables a deeper understanding of natural microbial communities, particularly as a great manyfungiaredifficultorimpossibletocultivate.Inthelastdecade,effectiveamplificationplatforms,probedevelopmentandvariousquantitativePCRtechnologieshaverevolutionizedresearchonfungaldetectionandidentification.Examplesofthelatesttechnologyinfungaldetectionanddifferentiationarediscussedhere.
Article info:Submitted17October2011;Accepted3November2011;Published18November2011.
Key words: FISHLAMPmacroarraymedical mycologymolecular diagnosticsmolecular ecologypadlock probepathogenic fungi plant pathologyrollingcircleamplification
doi:10.5598/imafungus.2011.02.02.09 IMA FuNgus · voluMe 2 · No 2: 177–189

ARTICLE
178
tsui et al.
i m a f u n G u S
geNerAl coNsIderAtIoNs
Suitable methodologies to isolate nucleic acids from theenormous range of habitats that fungi can occupy are crucial toallmoleculardetectionmethods.Criticaltothesuccessfulisolationofnucleicacidsarethemethodsusedforextractionand sample preparation, as well as the sampling strategy employed.
dNA extraction methods An enormous variety of nucleic acid extraction methodsis available. For many applications, commercial kits areavailable,butthesearenotalwayssuitable.Forexample,inthe case of soil, none of the kits presently available are able to extract efficiently using sample sizes typically required(see below). Therefore, with unusual and difficult sampletypes, customized methods are typically employed. Key tothis is the cell disruption/homogenisation step in the presence of an appropriate buffer (typically a CTAB or guanidiniumthiocyanatebasedbuffer).Awidevarietyofmethodscanbeused, including specialist equipments such as a planetary ball mill (Brierley et al. 2009), appropriate bead beaters,pressurecyclingtechnology(Okubaraet al.2007),andnovelapproaches using equipment such as a paint shaker for soil samples(Reelederet al.2003),orevenaconventionalfoodblenderforplantandfoodsamples.Allcelldisruptionstepsarepotentially damaging to the nucleic acid quantity and quality, either through direct shearing of the nucleic acid, or through theco-extractionofhumicacids(aninhibitorofPCR)presentinplantandsoilsamples.Therefore,methodswhichprovideanadequate level of cell disruption to enable satisfactory nucleic acidextraction,butwithouttoomuchnucleicaciddamageorreleaseofpotentialPCRinhibitors,arerequired.
Followingcelldisruptionandsubsequentextractionsteps,the nucleic acid can be purified using standard methods,including pelleting, silica membrane spin filter, and silica-magnetic particle separation. When selecting a purificationstep, consideration needs to be given to the nucleic acid quality/quantityrequired,cost,speedofextractionandabilitytoautomateanyprocess.Themethodofanalysingthenucleicacid will determine what purification method is used. Forexample,DNAprofilingandmetagenomicstudiesusingnextgeneration sequencing approaches will require pure, high quality nucleic acid extracts. Conversely, cruder, less pureextractsmaybeusedwherereal-timePCRisusedforroutinediagnostic purposes where the emphasis is on nucleic acid quantity,speedandcostofextractionbecausethedownstreamapplicationismoretolerantofimpuritiesintheDNAextract.
sample preparation Innovative sample preparation can enhance chances ofdetection. For example, organic matter can be removedfrom soil and processed separately for fungal targets which survive solely in that component of the soil. Sclerotium-forming pathogens may be separated from the rest of the soil bysievingorfloatingpriortonucleicacidextraction.These
approaches effectively concentrate the fungal target, thereby increasing the chance of successful detection. Whereappropriate, a baiting approach could be used in conjunction with nucleic acid extraction and PCR.This has the addedbenefitofconfirmingthetargetisviable.
sampling strategy Inaddition,anappropriateandstatisticallyrobustsamplingstrategy should be utilised. This will vary between fungaltargets, depending upon the biology (and possiblyepidemiology)oftheorganismtobedetected.Forexample,a‘W’ sampling strategy across an area may not be appropriate for a target with a highly clustered distribution, and a grid sampling approach should beused instead.The individualsample size processed for DNA extraction should also belargeenoughtobebiologicallyrelevant(Ophel-Kelleret al. 2008).Awidevarietyofmethodsexist,andnooneapproachislikelytobesuitableforalltargets.Knowledgeofthebiologyof the target species is essential for designing and determining the optimum sampling and extraction methodology in anyparticularcase.
sequeNcINg INdepeNdeNt Methods
DNAsequencingoftheinternaltranscribedspacer(ITS)andlargesubunit(LSU)regionsofrRNA,followedbycomparativesequence analysis, has been the ‘gold standard’ for molecular identificationofmostfungi,particularlyofculturablefungi.Thisstrategy is fast and accurate, but is dependent on sequence quality in existing reference databases. In many recentlyevolved fungal groups, however, the ribosomal genes are insufficiently variable, and sequencing of additional genesis necessary. Costs could also be a challenge for routinediagnostic laboratories, and therefore various sequencing independentmethodsthatareavailablecanbeused.
Fluorescent in situ hybridisationSincefungiareubiquitous,itisimportanttounderstandtheirbiodiversity and abundance, as well as their ecological roles in different habitats, such as soil, decaying leaves and wood, andalsoindoorenvironmentsorhumans.Indeed,itiscriticalto identify the metabolically active species (‘real players’)in communities or ecosystems in order to understand the ecosystem processes that involve fungi. PCR basedmethods,suchasfingerprintingormolecularcloning,donotdiscriminatebetween‘realplayers’andfungithataredormant.Also,DNAandRNAextractionprotocolsmaybebiasedduetorigidcellwallsoffungalhyphae.Immuno-stainingprovidesa means to identify species in situbutisextremelylaboriousintheexperimentalpreparation. Allthesekindsoftechnicaldifficulties, can, however, be overcome by fluorescence in situhybridisation(FISH).
FISH is a powerful method for the in situ detection of activegrowingorganismsinenvironmentalsamples(Amannet al.1995).Thetechniquecanvisualizethepreciselocation

Molecular identification and detection of fungi ARTIC
LE
179v o l u m e 2 · n o . 2
of particular DNA or RNA sequences in the cytoplasm,organelles,ornucleiofbiologicalmaterials.Asaresult,thetechnique can detect metabolically active fungi directly in the environment without cultivation when RNA is present. Thespatial distribution of growing mycelia on or within colonized substratacanalsobeinvestigated(Liet al.1996,Baschienet al.2001,McArthuret al.2001,Robinet al.1986).
The major step of FISH involves the preparation ofbiological materials or environmental samples, and the labelling (incorporation of a fluorescent label/marker) of anucleicacidsequencetoformaprobe.Then,undercontrolledexperimentalconditions,theprobeishybridizedtotheDNAor RNA in biological materials to form a double-strandedmolecule. Finally, the sites of hybridization are detectedand visualized. FISH is commonly used in the ‘top-bottomapproach’ research followed with feedback to the top level (Amann et al. 1995). The molecular biology and geneticsof targeted organism is well-understood, and the probe is designed based on the nucleotide sequence which has been wellcharacterized.
FISH probes often target sequences of ribosomalRNAor mitochondrial genes because they are abundant in sequencedatabasesandinmultiplecopiesineachcell.Theprobes can be designed by computer-assisted search from target organisms for “signatures”. These signature regionscan be specific at different phylogenetic levels dependingon the variability of the target molecule sequences. Theprobe comprises a short sequence, ranging from 15 to20 nucleotides, that is specific for one or several taxa atspecies, genus, or higher taxonomic ranks. The probe oroligonucleotide is then labelled with a fluorochrome, forexampleacarboindocyaninedye(CY3).
AparticularattractionisthatFISHprobescanbeappliedtoformaldehydeorethanolfixedenvironmentalsamples,ortocultures.Attheribosomes,theprobewillspecificallyannealto itscomplementarysequenceresulting inaheteroduplex.Afterincubation,awashingstepiscrucialtodiscriminatethetarget from non-target organisms. Probes that do not bindto an rRNA sequence will be washed away, resulting in afluorescentsignalofexclusivelytargetorganisms.Theprobeconferred signal is correlated with the ribosome content, and thereforeincreasesincellswithhighermetabolicactivity.
The first FISH probe targeting a living fungus wasdesigned by Li et al. (1996) for Aureobasidium pullulans on the phylloplane of apple seedlings; this was the firsttime that a living fungus had been visualized by FISH.Examplesofsituationsinwhichthemethodhasbeenusedare very varied. Fungi belonging to Eurotiomycetes were demonstrated to be more abundant than Dothideomycetes in anextremelyacidicminedrainageusing18SrRNA-targetedprobes(Bakeret al.2004).Baschienet al.(2008)designednine taxon-specific probes targeting the 18S or 28S rRNAfor thedetectionofaquatichyphomycetes in leaf litter(Fig.1A, B). Domain specific probeswere also used to detectactivefungiinmice(Scuphamet al.2006)andinwastewatersewagegranule biofilms (Weberet al. 2007).Most probeshavebeendesignedtotargetthe18Sor28SrRNAgene,andtheirspecificityneedstobekeptunderreviewassequencedatabasesexpand.
Several factorscan influence theefficiencyofFISH, forexample thestericalandelectrostaticalpropertiesof rRNA,the features of the probe, and hybridisation conditions such as the fixation method, buffers, the stringency of probebinding,andincubationtime.Inacioet al.(2003)investigated
Fig. 1.Examplesoffluorescence in situhybridisation.A. Clavariopsis aquaticagrowingonaquatic leaf litter,probedwith18rRNA-targetedMY1574domainspecificprobe(fromBaschienet al.2008).B.AccessibilityofTetracladium marchalianum conidiaforFISH28SrRNA-targetedspeciesspecificprobeTmarchB10(modifiedfromBaschienet al.2008).

ARTICLE
180
tsui et al.
i m a f u n G u S
the different accessibility of the rRNA molecule for FISHprobes.Theydesigned32differentprobesthattargetedthe26S(largesubunit)ofrRNAforyeasts,andinvestigatedtheconferredsignal intensitygenerated from theprobes.Theyconcluded that the prerequisite for successful FISH is thespecificityofprobes(Inácioet al.2003).
Limitations of theFISHmethod can include fungal andsubstrateinherentautofluorescence,insufficientpermeabilityofcellwalls,non-specificbindingofprobes,andlowribosomecontents.Autofluorescence of fungi can also lead to falsepositivefluorescencesignals.Afluorescencescanconductedby confocal laser scanning microscopy revealed that many freshwaterfungiemitagreenautofluorescence(Baschienet al. 2001), and, consequently, a green-labelled FISH probecould not be used. It is, therefore, important to check forautofluorescence of both target and non-target organismsbefore selecting the fluorochrome for probe labelling.Environmentalsamplesandsubstratessuchasleaves,wood,and animal tissues, also emit strong autofluorescence inseveral wavelengths at the same time, interfering with probe signals.However,thepin-holetechniqueofalaserscanningmicroscope is helpful in reducing background emissions compared to conventional epifluorescence (Baschienet al.2001).
Low or no permeability of fungal cell walls can lead toweakornosignalsbecauseofthefailureofFISHprobestopenetraterigidcellwalls(Brulet al.1997).Onepossiblewayto overcome this problem is to use cell wall lysing enzymes, suchaschitinasesandglucanases.Amoreelegantmethodistheuseofpeptidenucleicacid(PNA)probes.PNAprobesare mimics in which the negatively charged sugar-backbone ofDNAisreplacedwithanon-chargedpolyamidebackbone.PNA probes share the same nucleotide bases as theconventionalDNAprobes.However,PNAprobespenetratecell walls more effectively due to their neutrality and they do not have to overcome the destabilizing electrostatic repulsion duringhybridisation(Wilsonet al.2005,Shepardet al.2008).
Lowribosomecontentscanarisefromeitherascarcityofpotential substrate in oligotrophic habitats, or the presence of componentsinhibitingfungalgrowth.However,theCatalysedReporterDepositionFISH (CARD-FISH) isa variantof theFISHmethoddesignedtoenhanceprobeconferredsignals.Themajordifferenceisthattheprobesarelabelledwithhorse-radish-oxidase instead of a fluorochrome. Fluorescence-labelledtyramideisthenaddedtothecellsafterhybridisation.The horse-radish-oxidase catalyses the deposition of thereporter,andthisreactionleadsto20to30foldstrongerFISHsignalsthanconventionalFISH.Consequently,thenumberofactiverRNAmoleculesnecessarytodetectaprobeconferredsignal decreases by using CARD FISH (Pernthaler et al.2002).
dNA array hybridization DNA array hybridization, also known as Reverse Dot BlotHybridization (RDBH) ormacroarray, is a techniquebased on hybridization of amplified and labelled genome regionsof interest to immobilized oligonucleotides spotted on a
solidsupportplatform. It was originally developed to detect mutations of human genes, and is still an important diagnostic tool in clinical research (Chehab &Wall 1992,Yang et al.2001, Zhanget al. 1991). It is now considered a powerfuland practical technique for the detection and identificationoffungiandothermicrobes,suchasbacteria,fromcomplexenvironmental samples without the need for isolation in culture(Chenet al.,2009,Ehrmannet al.1994,Lévesqueet al. 1998,Tambonget al. 2006,Ueharaet al.1999,Wuet al.2007,Zhanget al. 2007,2008).Forthistypeofapplication,oligonucleotides,ormicrocodes(Summerbellet al.2005),aredesignedfromataxonomicallycompletedatasetofsuitablegenomeregion(s)(Chenet al.2009,Tambonget al.2006).Theoligonucleotidescanbeselectedmanually,byanalysingmulti-sequence alignments, or through computer programs, such as SigOli (Zahariev et al. 2009) andArray Designer(Premier Biosoft International, PaloAlto, CA). Synthesizedoligonucleotideswith5’-aminemodificationsarethenspottedonto a supporting platform, such as a nylon membrane or glass slide, eithermanually or robotically. Robotic spottingcan significantlyincreasethemaximumdensityofthearraywhichcanfavourthedetectionofbroadertaxonomicgroups(Chenet al. 2009).Amplicons of the target gene region(s)areamplifiedbyPCR, labelledwithdigoxygenin (DIG)andsubjected to the DNA hybridization procedure previouslydescribed(Fessehaieet al.2003).Apositivereactionbetweenan amplicon and a perfectlymatched (PM) oligonucleotidegenerates a chemiluminescent signal which can be detected by X-ray film or a digital camera in dark rooms. Capturedimages are then analysed on a computer program such as GenePixPro(MolecularDevices,Sunnyvale,CA).
Thedesignofspeciesorgroup-specificoligonucleotidesisacrucialstepinthisprocesssinceitdefinesthespecificityandsensitivityof theassay(Lievenset al.2006,Pozhitkovet al. 2006, Urakawa et al. 2003). It is generally agreedthat the length of the oligonucleotide, the number, type and positionofSNPscontainedinanoligonucleotide,determineitsdiscriminatorypotentialforDNAhybridization.Adesirableoligonucleotide should: (1) have suitable thermodynamiccharacters such as melting temperature; (2) contain asmany polymorphic sites as possible located close to the centreor on the3’-half of the sequence; and (3) have theleastprobabilityofhairpinordimerformation(Kawasakiet al.1993,Lievenset al.2006).Oligonucleotide lengthsrangingfrom25to35merhavedisplayedthebestbalancebetweenspecificityandsensitivity(Chenet al.2009).Longerlengths(>35mer)ofoligonucleotidesreducetheratioofmismatchedto matched base pairs, yet increase the number of bases available for hybridization, providing lower specificity buthighersensitivitytoanarray(Dorriset al.2003,Lievenset al.2006).Oligonucleotidesup to70mer,however,havebeenused in macroarrays for the detection of plant viruses using gene sequences that are widely different between the closely related pathogens (Agindotan & Perry 2007, 2008). It hasalso been demonstrated that more complicated parameters, such as actual sequence arrangement and the mismatched duplextype,andtheir interactions,canplayimportantroles

Molecular identification and detection of fungi ARTIC
LE
181v o l u m e 2 · n o . 2
in affecting the discriminatory power of an oligonucleotide (Urakawaet al. 2003,Lievenset al. 2006,Pozhitkovet al.2006).Despitenumerousattempts topredict thebehaviourofoligonucleotidesinDNAhybridization,thethermodynamicinteraction between the probes and oligonucleotides remains poorlyunderstood(Pozhitkovet al.2007).
Most DNAmacroarrays that have been developed arebased on a single region for the detection of a specifictaxonomicgroup.Amongtheseregions,16SribosomalDNAhas been used for the detection of bacteria (Fessehaieet al.2003,Xionget al.2006).Variousgenomeregions,suchasribosomalDNAspacers(ITS),mitochondrialgenes(e.g.cytochrome oxydase c subunit 1, cox1) and some proteincodingregions(β-tubulin,EF-1α,etc),werechosentotargetfungiandfungus-likeorganisms(Chenet al.2009,Gilbertet al.2008,Harperet al.2011,Ninget al.2007,2008,Seifert& Lévesque 2004).Oligonucleotideswith higher specificityare often designed from polymorphic sites located at indels presentinmulti-sequencealignments(Robideauet al.2008,Tambong et al. 2006). Oligonucleotides extracted from alocus that can be well aligned with low sequence divergence tends to cross-react with non-target amplicons and display strong false positive signals even when tested with pure cultures because of the low frequency of polymorphisms in thespecificoligonucleotides.Thishasbeenobserved inarrays using cox1 in Penicillium subgen.Penicillium (Chenet al. 2009, Seifert et al. 2007). In a recent study, DNAarrays were constructed from multiple loci of Phytophthora species, including ITS, cox1andtheintergenicregion(cox2-1 spacer, CS)betweencytochromecoxidase2(cox2)andcox1 (W. Chen et al. pers. comm.). In comparison to thecox1 region, the length variation and the presence of indels in the sequence alignments of ITS and CS provided better opportunities to select highly specificoligonucleotides.Thecombination of all three arrays increased the discrimination potential,detectionaccuracy,andredundancyoftheassay.
DNA arrays have been developed for the detection ofplant pathogens in a wide range of environmental samples, suchasgreenhousecrops(LeFlochet al.2007,Lievenset
al.2003),potatoes(Fessehaieet al.2003),ginseng(Punjaet al.2007),andfruits(Robideauet al.2008,Sholberget al.2005,2006).Macroarraysarealsoeffectivediagnostictoolsfor thedetectionofphytopathogenicbacteria(Fessehaieet al.2003),fungiandfungus-likeorganisms(Chenet al.2009,Lévesque et al. 1998, Tambong et al. 2006), nematodes(Ueharaet al.1999),andviruses(Agindotan&Perry2007,2008).
DNAarray hybridization is highly sensitive as aremostPCR-based approaches. With the unlimited capacity forthe accommodation of oligonucleotides on one membrane and the reusability of the membranes, it shows superior multiplexing detection capability at a lower cost over otherPCR-based methods. In a biodiversity study, the speciesprofilecouldbe revealedbyhybridizing theoligonucleotidearray with a mixed pool of DIG-labelled PCR productsamplifiedfromthetotalDNAofaquerysample.Thisassaywould be especially valuable for the simultaneous detection ofmultipleplantpathogenswhichcoverabroadtaxonomicrange but are specific to a certain host. As an example,Robideauet al.(2008)developedaDNAarrayforcranberryfruitrot(CFR)fungalpathogenswithover2000fieldsamplesbeingtested.Thisarrayhadtheabilitytodetectspeciesfrom24 genera of fungi known on cranberry with a single test.Fig. 2 shows that thisDNAmacroarraywasable todetectspecies of Phyllosticta, Cladosporium, Epicoccum, Godronia, Alternaria, Pestalotia, and Pilidium from a single frozen cranberryfruitsample(Robideauet al.2008).
Themacroarraytechnologyisnowavailablecommerciallyin four European countries under the nameDNAMultiscan(http://www.dnamultiscan.com) and at the Guelph PestDiagnosticClinic(http://www.guelphlabservices.com/) forthe test of plant pathogens in greenhouse nutrient solutions androotsofcrops.Thereareverysignificantadvancesinnextgeneration sequencing technology that have reduced the need of DNA array technology in functional genomics. However,there are developments in lab-on-a-chip array technology for very quick and for large-scale testing. A recent example isa low density array for Phytophthora detection, where amino-
Fig. 2.DNAmacroarrayhybridizationresultsfromfrozencranberryfruitsamplecollectedfromMassachusetts,USA(adaptedfromRobideauet al.2008).Thex-rayfilmisoverlaidwiththeoligonucleotidespottingpatternonDNAarraymembraneforcranberryfruitrotfungi.Thetopleftpairofspotsandbottomrightpairofspotsareoligonucleotideswhichservedaspositivecontrols.ThisarrayisshowingpositivesignalforPhyllosticta, Coleophoma, Epicoccum, Godronia, Alternaria, Pestalotia, and Pilidium.

ARTICLE
182
tsui et al.
i m a f u n G u S
labelled oligonucleotides are spotted over a gap between two electrodesinsideamicrochip(Julichet al.2011).Hybridizationis detected by a current passing through a silver deposit over thegap.Becauseofsuchdevelopments,array-basedassaysarelikelytoremainapopularoptionfordiagnostics.
Multiplex tandem pcr Multiplexed-tandemPCR(MT-PCR)isatechnologyplatformdeveloped for highly multiplexed gene expression profilingandtherapididentificationofclinically importantpathogens(Stanley et al. 2005). The platform consists of two roundsofamplification(Fig.3).Inthefirststep,amultiplexPCRisperformedat10 to15cycles toallowenrichmentof targetDNAwithout creating competition between amplicons (Lauet al.2009).Thisproductisdilutedandusedastemplateforthesecondamplification that consistsofmultiple individualquantitative PCR reactions with primers nested withinthose used in the multiplex PCR. Up to 72 different PCRreactionscanbemultiplexedandperformedsimultaneously.Fluorescence is measured by SYBR green technology atthe end of each extension cycle, and melt-curve analysisprovides species-specific or gene-specific identification.Theincorporationoftwosetsofspecies-specificprimersforeachtargetensurescorrectamplificationanddetection,thusavoidingtheexpenseofDNAprobes.SYBRgreendetectionalso increases the multiplexing and quantitative capacityof real-timePCRsystems,whichareusually limitedby theavailabilityoffluorescentchannelsandtheneedtooptimizeeachindividualmultiplexPCR.
In the clinical setting, invasive fungal infections (IFIs)remain a leading cause of morbidity and mortality in immune-compromised hosts. MT-PCR was considered asuitablealternative for therapiddetectionand identification
of fungal pathogens directly from clinical specimens, thus circumventing the need for gold standard culture and histology,whichisslow,insensitiveandnon-specific.Fasterand accurate molecular identification would also enablebetter guidance and earlier administration of targeted antifungal therapies.As such, the laboratory atWestmeadhospital, in conjunction with AusDiagnostics (Alexandria,NSW, Australia), developed several MT-PCR assays todetectthe16majorcausesoffungalbloodstreaminfections.The targets included11 species ofCandida, Cryptococcus neoformans complex, Saccharomyces cerevisiae, pan-Fusarium, F. solani, and Scedosporium prolificans.Primerswere designed using sequence variations within the ITSregions, elongation factor 1-a(EF1-a),andb-tubulingenes.Fungal targets were selected according to their frequency of causing infections, their potential resistance to frontline antifungal agents (especially fluconazole), and their highattributablemortality.
TheMT-PCRplatformwasinitiallyevaluatedon70bloodculturesinwhichayeastormouldwasseeninGram’sstainpreparations,aswellas200bacterialbloodculturesand30sampleswhichdidnotflagpositive(Lauet al.2008a).Thesensitivityandspecificityoftheassaywas100%.Thisincludedthecorrect identificationoffungi infivecaseswithbacterialco-infection. In addition, no interference was observed insimulatedcasesofpolyfungalinfection.Unfortunately,threeraredisease-causingyeasts(Candida lambica, C. nivariensis and Kodamaea ohmeri) were not detected byMT-PCR astargetswerenotavailableonthefungalpanel.Nevertheless,this study demonstrated the diagnostic usefulness of the platform to rapidly identify common fungal pathogens within four hours of blood culture flagging (including automatednucleicacidextraction),whichisconsiderablyfasterthanthe48-96hrequiredbygoldstandardmethods.
AnexpandedversionoftheMT-PCRfungalplatformwasthenevaluatedon255EDTAwholeblood,29serum,and24plasmaspecimensobtained from109patientswith provencandidemia(Lauet al.2010)with theaimofcircumventingthe technical and sensitivity issues inherent in routine blood culture diagnosis. Although the MT-PCR assay was lesssensitivethanbloodculture(75%sensitivity), thediagnosisofcandidemiaandpathogenidentificationwasexpeditedbyup to four days.The results also supported the contentionthat serum and plasma samples were better than whole bloodsamplesforthemoleculardetectionofcandidemia.
Using a technique known as colony MT-PCR (Lau et al. 2008b), the fungal assaywas also able to provide rapid(1.5h)andunambiguous identificationof yeastsdirect fromprimaryisolationplateswithouttheneedforDNAextractionorseparationfrommixedfungal/bacterialcultures.Assuch,colonyMT-PCRofferedafasterandbetteralternativetobiochemicalassayswhichareoftensubjective,pronetomisidentification,anddependentonapureculturebeingobtained.Currentworkis aimed at integrating identification panels with targets todetectmolecularmechanismsofantifungalresistance.
Overall, thefungalMT-PCRassaycomparesfavourablyto other commercial platforms and integrates well into the
Fig. 3. Schematic representation of the multiplex tandem–PCRprocedureillustratingspecificdetectionandidentificationfromabloodsample containing Candida albicans and C. glabrata. ReproducedfromLauet al.(2009)withthepermissionofFutureMedicine.
Molecular identification and detection of fungi ARTIC
LE
183v o l u m e 2 · n o . 2
routine workflow of diagnostic laboratories. Automatedoperations and use of commercial reagents further enables standardizedprocedurestobeestablished.Nevertheless,themajor limitation of all molecular diagnostic assays is target availability, which is often dictated by an effort to balance out costsand turn-around timewithmaximum throughput.Theincreased sensitivity of molecular assays and its ability to detect viable and non-viable cells should also place heavy emphasis on interpreting results with other microbiological dataandclinicalinformation.
IsotherMAl sYsteMs
Isothermalsystemsdonotrequireathermalcyclertoproducerapid temperature changes, but require only a simple platform such as heating blocks or water bath. Isothermal systemsincluderollingcircleamplification(RCA)and loop-mediatedisothermalamplification(LAMP).
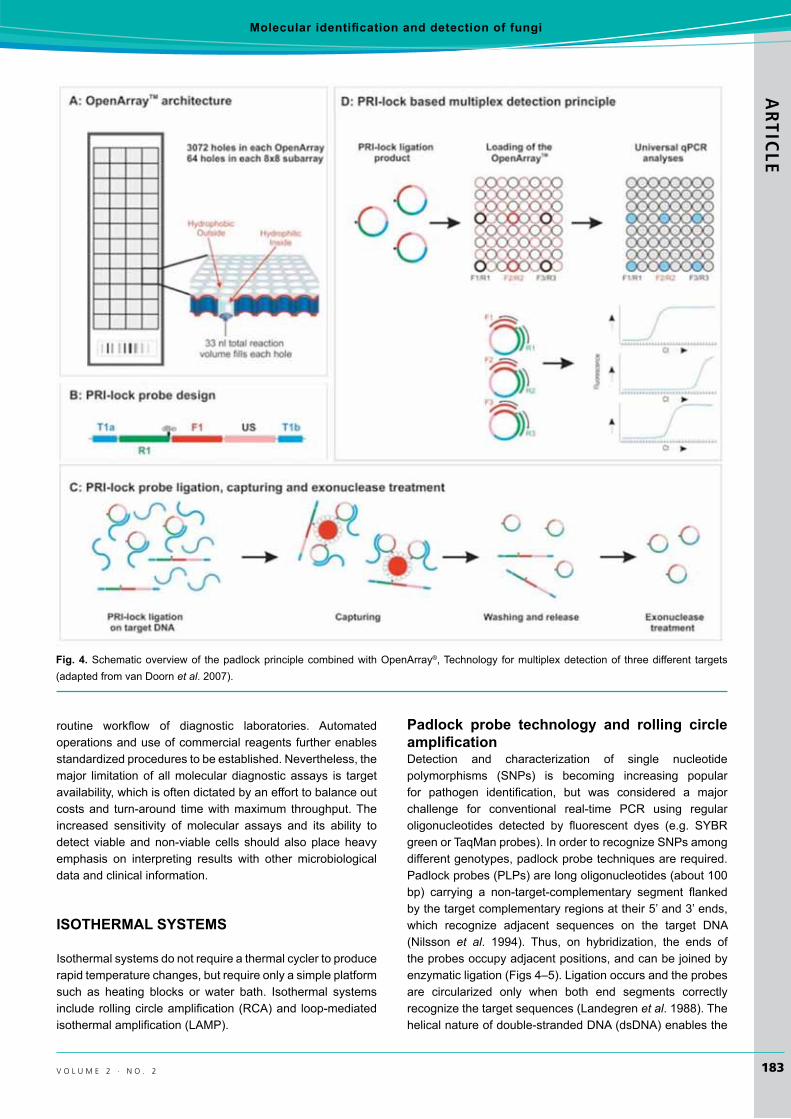
padlock probe technology and rolling circle amplification Detection and characterization of single nucleotidepolymorphisms (SNPs) is becoming increasing popularfor pathogen identification, but was considered a majorchallenge for conventional real-time PCR using regularoligonucleotides detected by fluorescent dyes (e.g. SYBRgreenorTaqManprobes).InordertorecognizeSNPsamongdifferentgenotypes,padlockprobetechniquesarerequired.Padlockprobes(PLPs)arelongoligonucleotides(about100bp) carrying a non-target-complementary segment flankedbythetargetcomplementaryregionsattheir5’and3’ends,which recognize adjacent sequences on the target DNA(Nilsson et al. 1994). Thus, on hybridization, the ends ofthe probes occupy adjacent positions, and can be joined by enzymaticligation(Figs4–5).Ligationoccursandtheprobesare circularized only when both end segments correctly recognizethetargetsequences(Landegrenet al.1988).Thehelicalnatureofdouble-strandedDNA(dsDNA)enablesthe
Fig. 4.Schematicoverviewof thepadlockprinciplecombinedwithOpenArray®,Technology formultiplexdetectionof threedifferent targets(adaptedfromvanDoornet al.2007).

ARTICLE
184
tsui et al.
i m a f u n G u S
probe to topologically bind to the target strand and the probe can’tbedisplaced(Nilssonet al.1994).
Padlock probes were initially introduced for in situ DNA localization and detection (Nilssonet al. 1994). Theywere developed originally for discrimination of centromeric sequence variation in human chromosomes (Nilsson et al. 1997). However, the method has now been applied todetection of genetically modified organisms (Prins et al.2008). This concept also provides extensive multiplexpotential for pathogen detection as the interaction between padlock probes does not give rise to circular molecules,
which can be easily removed from the detection system (Nilssonet al.1994,1996).Recently,padlockprobe-basedapplications formultiplexquantitative targetsdetectionandfor genotyping fungal and microbial community analysis usinghigh throughput real-timePCRonOpenArrays® have been developed (van Doorn et al. 2007). Advantages ofPLP-baseddiagnosticapplicationsdevelopedareaflexibleand easily adaptable design, specificity, and multiplexing,universal downstream processing after ligation, and high-throughput formatwith real-timeanalysis (Tsuiet al. 2012)(Fig.4).
Briefly, various padlock probes are designed to targetorganismsand ligated toDNAextracted fromenvironmentalsamples or cultures (van Doorn et al. 2007). Targets forligationpresent incomplexDNAsamplessuchassoilorre-circulatedwatercanbegeneratedbyPCRpre-amplification,through Phi29polymeraseandwholegenomeamplificationtoensureefficientdetection.Real-timequantificationformultipletargetsisperformedinOpenArrays®, which can accommodate up to3072x33nlPCRamplificationwithpreloadedprobe-specificprimers(Fig.4).Multiplexpadlockligationisfollowedbysingle-plexamplificationusinguniqueprobe-specificprimerpairs and SYBR green based detection in nano-litre PCRarray(vanDoornet al.2007).Theperformanceofthepadlockprobe detection system has been demonstrated using 13 probestargetingseveralplantpathogensatvarioustaxonomiclevels (Szemes et al. 2005, van Doorn et al. 2007). Allprobesspecificallydetected theircorresponding targets,andprovided perfect discrimination against non-target organisms with very similar target sites. Pathogen quantification wasrobust in single target versusmixedtargetassays.Thisnovelassay enables very specific, high-throughput, quantitativedetection of multiple pathogens over a wide range of target concentrations, and should be easily adaptable for versatile diagnosticpurposes(vanDoornet al.2007).
Also,padlockprobescontainingzip-codesequenceorabiotin-labelled moiety and internal endonuclease cleavage site, in conjunction with quantitative PCR and LuminexTM technology or microarray technology, can be used for multiplexpathogendetectionandquantification(Szemeset al.2005,Erikssonet al.2009,vanDoornet al.2009).
Alternatively, the signal by which the target hybridizesperfectly to the padlock probes can be amplified byhyperbranched rolling circle amplification (H-RCA) (Banéret al. 1998).Rolling circle amplification (RCA) is basedontherollingreplicationofshortsinglestrandedDNA(ssDNA)circularmolecules(Lizardiet al.1998, Fire&Xu1995).RCAinvolves a single forward primer complementary to the linker regionofthepadlockprobeandaDNApolymerasewithstranddisplacement activity in an isothermal condition (Pickeringet al. 2002). Asa result, thepadlockprobesignal canbeamplified several thousand-fold because the polymeraseextendstheboundprimeralongthepadlockprobesformanycycles and displaces upstream sequences, producing a long ssDNA molecule comprising multiple repeats of the probesequence.Twoprimersareemployed:afirstforwardprimerthat binds to thepadlockprobeand initializesRCA,anda
Fig. 5. Schematic representation of the steps in padlock probetechnology coupled with hyperbranched rolling circle amplification(H-RCA)forSNPsdetection.1. Thehybridizationofpadlockprobes(containingthecomplementarysequencesatthe5’and3’ends)tothe target templates.2.Duringaperfectmatch, theprobeformsacircularmolecularwith theaidofDNA ligase;while in thecaseofmismatch,nocircularmoleculesformed.3. Non-hybridizedtemplatewill be removed during the exonucleolysis reactions (digestionby exonucleases I and III).4. H-RCA is performedusing twopre-designed primers and Bstpolymerase,butnoamplificationwilltakeplaceintheabsenceofacircularmolecular.5. TheaccumulationofdsDNAproductsduringisothermalrollingcircleamplificationofDNAminicircles ismonitored in a real timePCR thermocycler with theadditionofSYBRgreen.

Molecular identification and detection of fungi ARTIC
LE
185v o l u m e 2 · n o . 2
secondprimerthattargetstherepeatedssDNAsequenceoftheprimaryRCAproduct,finallygeneratinglargenumbersofcopiesoftheDNAfragments.ThisiscalledhyperbranchingRCA(H-RCA)(Lizardiet al.1998)(Fig.5).
Padlock probe coupled with H-RCA offers a significantadvantage for thedetectionofSNPs (Tsuiet al. 2012).Theformation of circular probes via ligation occurs when both ends of the padlock probes perfectly hybridize to the target at juxtaposition(Fig.5).ThesubsequentH-RCAamplificationofa target probe could be carried out when circularized probes becomeavailable.Thesetwostrictconditionscreateanidealdetection platform for highly sensitive and specific SNPsdetection. By increasing the hybridization temperature andshortening the 3’ arm (below the reaction temperature), thediscriminationofSNPcanbe further improved (Pickeringet al.2002, Faruqi et al. 2001).ThismethodforSNPsdetectionbasedonDNAligase-mediatedsinglenucleotidediscriminationwith consecutive signal amplification by H-RCA has beendeveloped for various groups of pathogenic organisms, includingfungi,bacteria,andviruses(Konget al.2008,Zhouet al.2008,Kaocharoenet al.2008,Wanget al.2009,2010). Recently,thetechnologyhasbeenusedto differentiate and to detect two closely related conifer pathogens vectored by the mountainpinebeetles(Tsuiet al.2010).
Loop mediated isothermal amplificationLoop-mediatedisothermalamplification(LAMP)isapowerfuland novel nucleic acid amplification method that amplifiesa fewcopiesof targetDNAwith high specificity, efficiency,and rapidity under isothermal conditions, using a set of four speciallydesignedprimersandaDNApolymerasewithstranddisplacementactivity(Notomiet al.2000,Paridaet al.2008,Tomita et al. 2008).Thecycling reactionscan result in theaccumulationof109to1010-fold copies of target in less than anhour.Considering theadvantagesof rapidamplification,simple operation and easy detection, LAMP has potentialapplications for clinical diagnosis as well as surveillance of infectious diseases in developing countries without requiring sophisticatedequipmentorskilledpersonnel(Mori&Notomi 2009, Parida et al.2008).
Thetechniquewasfirstdescribedandinitiallyevaluatedfor detection of hepatitis B virus DNA by Notami et al.(Notomiet al. 2000).LAMPassayshavebeenmostlyusedfor the diagnostics of bacteria (Chen et al. 2011, Han et al. 2011), virus (Wanget al. 2011,Zhaoet al. 2011), andparasites (Ikadaiet al. 2004, Isekiet al. 2007), but it hasalso been developed for the rapid detection of pathogenic orallergenicfungi.Ohoriet al.(2006)usedthetechniqueforrapid identification ofOchroconis gallopava, an emerging fungal pathogen and causative agent of zoonotic infections, whileSunandco-workers,useditfortherapiddiagnosisofPenicillium marneffeiinarchivedtissuesamples(Sunet al. 2010a)andofFonsecaea agents of chromoblastomycosis (Sunet al. 2010b).SimilarlyEndoet al.(2004)andTatibanaet al. (2009) detected the presence of the gp43 gene in Paracoccidioides brasiliensis by LAMP, and Lucas et al.(2010) used LAMP for differentiation of Cryptococcus neoformans varieties from C. gattii based on CAP59sequences.Recently,Luet al.(2011)utilizedthetechnologyfortheidentificationofPseudallescheria and Scedosporium species.
TwoinnerandtwoouterprimersarerequiredforLAMP(Fig. 6A). In the initial stepsof theLAMP reaction, all fourprimers are employed, but in the later cycling steps only the inner primers are used for strand displacement DNAsynthesis.Theouterprimersarereferred toasF3andB3,while the inner primers are forward inner primer (FIB) andbackwardinnerprimer(BIP).BothFIPandBIPcontainstwodistinct sequences corresponding to the sense and antisense sequences of the target DNA, one for priming in the firststageandtheotherforself-priminginlaterstages(Notomiet al.2000).Thesizeandsequenceoftheprimerswaschosensothattheirmeltingtemperature(Tm)isbetween60-65°C,the optimal temperature for Bst polymerase. The F1c andB1cTmvaluesshouldbealittlehigherthanthoseofF2andB2 to form the looped-out structure.TheTmvaluesof theouterprimersF3andB3havetobelowerthanthoseofF2andB2toassurethattheinnerprimersstartsynthesisearlierthantheouterprimers.Additionally,theconcentrationsoftheinner primers are higher than the concentrations of the outer primers(Notomiet al.2000,Tomitaet al.2008).Furthermore,itiscriticalforLAMPtoformastem-loopDNAfromadumb-
Fig. 6. Schematic representation of the mechanism of LAMP. A. GenerallocationoftheLAMPprimersetinrelationtodefinedregionsof the targetDNA.Forward (F3) andbackward (B3) outer primersand forward (FIP)andbackward (BIP) innerprimersare indicated.B, c. Basic principle and amplification steps in LAMP. In generalnewDNAstrandsaresynthesizesfromtheF3andB3primers,andthesestrandsarerecognizedbyFIPandBIPtostartloop-mediatedautocycling amplifications. The final products are stem-loop DNAswithseveralinvertedrepeatsofthetargetDNA,andcauliflower-likestructuresbearingmultipleloops(modifiedfromdiagramsat<http://loopamp.eiken.co.jp/e/lamp/principle.html>©EikenChemical.

ARTICLE
186
tsui et al.
i m a f u n G u S
bell structure. Various sizes of the loop between F2c andF1candbetweenB2candB1cwereexamined,andthebestresults are obtained when loops of 40 nucleotides (40nt)or longerareused (Notomiet al.2000).Thesizeof targetDNA is an important factor that LAMP efficiency dependson,becausetheratelimitingstepforamplificationisstranddisplacementDNAsynthesis.
LAMP reliesonauto-cycling stranddisplacementDNAsynthesis in the presence of BstDNApolymerase,specificprimers and the target DNA template. Themechanism ofthe LAMP amplification reaction includes three steps:production of starting material, cycling amplification, andrecycling(Notomiet al.2000,Tomitaet al.2008)(Fig.6B,C).Toproducethestartingmaterial,innerprimerFIBhybridizestoF2cinthetargetDNAandinitiatescomplementarystrandsynthesis.OuterprimerF3hybridizes toF3c in the targetandinitiatesstranddisplacementDNAsynthesis,releasingaFIP-linkedcomplementarystrand,whichformsalooped-outstructureatoneend(DNAamplificationproceedswithBIPinasimilarmanner).ThissinglestrandedDNAservesastemplateforBIP-initiatedDNAsynthesisandsubsequentB3-primed strand displacement DNA synthesis leading tothe production of a dumb-bell form DNAwhich is quicklyconverted to a stem loop DNA. This then serves as thestartingmaterialforLAMPcycling,thesecondstageoftheLAMPreaction.Duringcyclingamplification,FIPhybridizesto the loop in the stem-loop DNA and primes stranddisplacementDNAsynthesis,generatingasanintermediateonegappedstemloopDNAwithanadditionalinvertedcopyof the target sequence in the stem, and a loop formed at the opposite end via theBIP sequence.Subsequent self-primed strand displacement DNA synthesis yields onecomplementarystructureoftheoriginalstem-loopDNA,andonegaprepairedstem-loopDNAwithastemelongatedtotwiceaslongandaloopattheoppositeend.Bothoftheseproducts then serve as templates for BIP-primed stranddisplacement in the subsequent cycles, the elongation and recyclingstep.Thefinalproduct isamixtureofstem loopDNAwithvariousstemlengthandcauliflower-likestructureswith multiple loops formed by annealing between alternately inverted repeats of the target sequence in the same strand (Notomi et al.2000,Tomitaet al.2008).
LAMPproductscanbedirectlyobservedbythenakedeyeorusingaUVtransilluminatorinthereactiontubebyadding2.0µlof10folddilutedSYBRGreenIstaintothereactiontube separately. Under UV illumination, the gel shows aladder-likestructurefromtheminimumlengthoftargetDNAup to the loading well, which are the various length stem-loop productsoftheLAMPreaction.
coNclusIoN
Numerous detectionmethodologies are now available, butregardless of the approach, important questions need to be answered prior to their inclusion into experiments. Theseinclude sensitivity, accuracy, robustness, frequency of testing,
andcost.Despitemanynoveltechnologiesbeingavailable,challenges remain to identify as yet unculturable fungi, to detect cryptic species, and to characterize the assemblage and diversity of fungal communities in different environments without bias.There is always a need to characterize fungiquickly and accurately. No one knows how many fungalspecies exist, but sequencing of environmental DNA mayimprove the accuracy of current estimates (Hawksworth2001). Next-generation sequencing and pyrosequencingapproaches will also provide promising ways of enlarging the scope of molecular-detection studies.
AcKNowledgeMeNts
WewouldliketothanktheIMC9organizersforsupportingthisSIGsymposium.WearethankfultoSatokoYamamoto(EikenChemical,Japan)forpermissiontousediagramsofLAMPfrom<http://loopamp.eiken.co.jp/e/lamp/principle.html>.
reFereNces
Agindotan B, Perry KL (2007)Macroarray detection of plant RNAviruses using randomly primed and amplified complementaryDNAsfrominfectedplants.Phytopathology 97:119–127.
Agindotan B, Perry KL. (2008) Macroarray detection of elevenpotato-infecting viruses and Potato spindle tuber viroid. Plant Disease 92:730–740.
AgriosG(2005)Plant Pathology.5thedn.London:ElsevierAcademicPress.
Amann RI, Ludwig W and Schleifer K-H (1995) Phylogeneticidentification and in situ detection of individual microbial cells withoutcultivation.Microbiology and Molecular Biology Reviews 59:143–169.
Baker BJ, Lutz MA, Dawson SC, Bond PL, Banfield JF (2004)Metabolicallyactiveeukaryoticcommunitiesinextremelyacidicmine drainage. Applied and Environmental Microbiology 70: 6264–6271.
BanérJ,NilssonM,Mendel-HartvigM,LandegrenU(1998)Signalamplification of padlock probes by rolling circle replication.Nucleic Acids Research 26:5073–5078.
BaschienC,ManzW,NeuTR,SzewzykU(2001)Fluorescence in situ hybridization of freshwater fungi. International Review of Hydrobiology 86:371–384.
BaschienC,ManzW,NeuTR,MarvanováL,SzewzykU(2008)In situ detection of freshwater fungi in an alpine stream by new taxon-specificfluorescencein situ hybridization probes. Applied and Environmental Microbiology 74:6427–6436.
BrierleyJL,StewartJA,LeesAK(2009)QuantifyingpotatopathogenDNAinsoil.Applied Soil Ecology41:234–238.
BrulS,NussbaumJ,DielbandhoesingSK(1997)Fluorescentprobesforcellwall16porosityandmembrane integrity infilamentousfungi.Journal of Microbiological Methods 28:169–178.
Chehab FF, Jeff W (1992) Detection of multiple cystic fibrosismutations by reverse dot blot hybridization: a technology for carrierscreening.Human Genetics 89:163–168.

Molecular identification and detection of fungi ARTIC
LE
187v o l u m e 2 · n o . 2
ChenJ,MaX,YuanY,ZhangW(2011)Sensitiveandrapiddetectionof Alicyclobacillus acidoterrestris using loop-mediated isothermal amplification.Journal of the Sciemce of Food and Agriculture 91: 1070–1074.
Chen W, Seifert K, Lévesque CA (2009) A high density COX1barcodeoligonucleotidearrayforidentificationanddetectionofspecies of Penicillium subgenus Penicillium.Molecular Ecology Resources 9:114–129.
EndoS,KomoriT,RicciG,SanoA,YokoyamaK,OhoriA,KameiK,FrancoM,MiyajiM,NishimuraK (2004)Detectionof gp43of Paracoccidioides brasiliensis by the loop-mediated isothermal amplification (LAMP)method.FEMS Microbiology Letters 234: 93–97.
Eriksson R, Jobs M, Ekstrand C, Ullberg M, Herrmann B,Landegren U, Nilsson M, Blomberg J (2009) Multiplex andquantifiable detection of nucleic acid from pathogenic fungiusing padlock probes, generic real time PCR and specificsuspensionarrayreadout.Journal of Microbiological Methods 78:195–202.
FaruqiAF,HosonoS,DriscollMD,DeanFB,AlsmadiO,BandaruR,KumarG,GrimwadeB,ZongQ,SunZ,DuY,KingsmoreS,KnottT,LaskenRS(2001)High-throughputgenotypingofsinglenucleotidepolymorphismswithrollingcircleamplification.BMC Genomics 2:4.
FessehaieADeBoerSH,LévesqueCA(2003)Anoligonucleotidearray for the identification and differentiation of bacteriapathogeniconpotato.Phytopathology 93:262–269.
Fire A, Xu SQ (1995) Rolling replication of short DNA circles.Proceedings of the National Academy of Sciences, USA 92: 4641–4645.
GilbertCA,ZhangN,HutmacherRB,DavisRM,SmartCD(2008)DevelopmentofaDNA-basedmacroarrayforthedetectionandidentificationofFusarium oxysporumf.sp.vasinfectumincottontissue.Journal of Cotton Science 12:165–170.
Han HJ, Jung SJ, Oh MJ, Kim DH (2011) Rapid and sensitivedetection of Streptococcus iniae by loop-mediated isothermal amplification(LAMP).Journal of Fish Disease 34:395–8.
HarperKA,SmartCD,MichaelDavisR (2011)Development of aDNA-based macroarray for the detection and identification ofAmanitaspecies.Journal of Forensic Science 56:1003–1009.
Hawksworth DL (1991) The fungal dimension of biodiversity:magnitude, significance, and conservation. Mycological Research 95:641–655.
HawksworthDL (2001)Themagnitude of fungal diversity: the 1.5million species estimate revisted. Mycological Research 105: 1422–1432.
Ikadai H, Tanaka H, Shibahara N, Matsuu A, Uechi M, Itoh N,Oshiro S, Kudo N, Igarashi I, Oyamada T (2004) Molecularevidence of infections with Babesia gibsoni parasites in Japan and evaluation of the diagnostic potential of a loop-mediated isothermalamplificationmethod.Journal of Clinical Microbiology 42:2465–2469.
InácioJ,BehrensS,FuchsBM,FonsecaÁ,Spencer-MartinsI,AmannR(2003)In situ accessibility of Saccharomyces cerevisiae 26SrRNA tocy3-labeledoligonucleotideprobescomprising theD1andD2domains.Applied and Environmental Microbiology 69: 2899–2905.
Iseki H, AlhassanA, Ohta N, Thekisoe OM, Yokoyama N, InoueN,NambotaA,Yasuda J, Igarashi I (2007)Development of amultiplex loop-mediated isothermal amplification (mLAMP)method for the simultaneous detection of bovine Babesia parasites.Journal of Microbiological Methods 71:281–287.
JulichS,RiedelM,KielpinskiM,UrbanM,KretschmerR,WagnerS,FritzscheW,HenkelT,MöllerR,WerresS(2011)Developmentof a lab-on-a-chip device for diagnosis of plant pathogens. Biosensors and Bioelectronics 26:4070–4075.
KaocharoenS,WangB,TsuiKM,TrillesL,KongF,MeyerW(2008)HyperbranchedrollingcircleamplificationasarapidandsensitivemethodforspeciesidentificationwithintheCryptococcus species complex.Electrophoresis 29:3183–3191.
Kawasaki E, Saiki R, Erlich H (1993) Genetic analysis usingpolymerase chain reaction-amplified DNA and immobilizedoligonucleotide probes: reverse dot-blot typing. Methods in Enzymology 218:369–381.
KongF,TongZ,ChenX,SorrellT,WangB,WuQ,EllisD,ChenSC(2008)RapididentificationanddifferentiationofTrichophyton species, based on sequence polymorphisms of the ribosomal internaltranscribedspacerregions,byrolling-circleamplification.Journal of Clinical Microbiology 46:1192–1199.
LandegrenU,KaiserR,SandersJ,HoodL(1988)Aligase-mediatedgenedetectiontechnique.Science 241:1077–1080.
LauA,ChenS,SleimanS,SorrellT(2009)Currentstatusandfutureperspectives on molecular and serological methods in diagnostic mycology.Future Microbiology 4:1185–1222.
LauA,SorrellTC,ChenS,StanleyK,IredellJ,HallidayC(2008a)MultiplextandemPCR:anovelplatformforrapiddetectionandidentificationoffungalpathogensfrombloodculturespecimens.Journal of Clinical Microbiology 46:3021–3027.
LauA, Sorrell TC, Lee O, Stanley K, Halliday C (2008b) Colonymultiplex-tandemPCRforrapid,accurateidentificationoffungalcultures.Journal of Clinical Microbiology 46:4058–4060.
LauA,HallidayC,ChenSCA,PlayfordEG,StanleyK,SorrellTC(2010)Comparisonofwholeblood,serumandplasmaforearlydetection of candidemia bymultiplex-tandem PCR. Journal of Clinical Microbiology 48:811–816.
Le FlochG, Tambong J, Vallance J, Tirilly Y, LévesqueA, Rey P(2007) Rhizosphere persistence of three Pythium oligandrum strainsintomatosoillesscultureassessedbyDNAmacroarrayandreal-timePCR.FEMS Microbiology Ecology 61:317–326.
Lévesque CA, Harlton CE, de Cock AWAM (1998) Identificationof some oomycetes by reverse dot blot hybridization.Phytopathology 88:213–222.
LiS,CullenD,HjortM,SpearR,AndrewsJH(1996)Developmentof an oligonucleotide probe for Aureobasidium pullulans based on the small subunit rRNA gene. Applied and Environmental Microbiology 62:1514–1518.
LievensB,BrouwerM,VanachterACRC,LévesqueCA,CammueBPA,ThommaBPHJ(2003)DesignanddevelopmentofaDNAarrayfor rapiddetectionand identificationofmultiple tomatovascularwiltpathogens.FEMS Microbiology Letters 223:113–122.
LievensB,ClaesL,VanachterAC,CammueBP,ThommaBP(2006)Detecting single nucleotide polymorphisms using DNA arraysfor plant pathogendiagnosis.FEMS Microbiology Letters 255: 129–139.

ARTICLE
188
tsui et al.
i m a f u n G u S
LizardiPM,HuangX,ZhuZ,Bray-WardP,ThomasDC,WardDC(1998) Mutation detection and single-molecule counting usingisothermalrolling-circleamplification.Nature Genetics 19:225–232.
LuQ,Gerrits van den EndeAH, Bakkers JM, Sun J, LacknerM,Najafzadeh MJ, Melchers WJ, Li R, de Hoog GS (2011).Identification of Pseudallescheria and Scedosporium species bythreemolecularmethods.Journal of Clinical Microbiology 49: 960-7
LucasS, da LuzMartinsM, FloresO,MeyerW,Spencer-MartinsI, Inácio J (2010) Differentiation ofCryptococcus neoformans varieties and Cryptococcus gattii using CAP59-based loop-mediated isothermal DNA amplification. Clinical Microbiology and Infection 16:711–714.
McArthur FA, Bärlocher MO, McLean NAB, Hiltz MD, BärlocherF (2001) Asking probing questions: can fluorescent in situhybridization identify and localize aquatic hyphomycetes on leaf litter? International Review of Hydrobiology 86:429-438.
Mori Y, Notomi T (2009) Loop-mediated isothermal amplification(LAMP):arapid,accurate,andcost-effectivediagnosticmethodfor infectiousdiseases.Journal of Infection and Chemotherapy 15:62–69.
NilssonM,MalmgrenH,SamiotakiM,KwiatkowskiM,ChowdharyBP, Landegren U (1994) Padlock probes: circularizingoligonucleotides for localized DNA detection. Science 265: 2085–2088.
Nilsson M, Krejci K, Koch J, Kwiatkowski M, Gustavsson P,Landegren U (1997) Padlock probes reveal single-nucleotidedifferences, parent of origin and in situ distribution of centromeric sequencesinhumanchromosomes13and21.Nature Genetics 16:252–255.
Nilsson M, Dahl F, Larsson C, Gullberg M, Stenberg J (2006)Analyzinggenesusingclosingandreplicatingcircles.Trends in Biotechnology 24:83–88.
Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K,AminoN,HaseT(2000)Loop-mediatedisothermalamplificationofDNA.Nucleic Acids Research 28:E63.
OhoriA,EndoS,SanoA,YokoyamaK,YaritaK,YamaguchiM,KameiK,MiyajiM,NishimuraK(2006)RapididentificationofOchroconis gallopava by a loop-mediated isothermal amplification (LAMP)method.Veterinary Microbiology 114:359–365.
OkubaraPA,SchroederKL,LiC,SchumacherRT,LawrenceNP
(2007)ImprovedextractionofRhizoctonia and PythiumDNAfrom wheat roots and soil samples using pressure cycling technology.Canadian Journal of Plant Pathology 29: 304–310.
Ophel-KellerK,McKayA,HartleyD,HerdinaA,Curran J (2008)Development of a routine DNA-based testing service forsoilborne diseases inAustralia.Australasian Plant Pathology 37:243–253.
ParidaM,SannarangaiahS,DashPK,RaoPV,MoritaK(2008)Loopmediatedisothermalamplification(LAMP):anewgenerationofinnovativegeneamplificationtechnique;perspectivesinclinicaldiagnosisofinfectiousdiseases.Reviews in Medical Virology 18: 407–421.
Pernthaler A, Pernthaler J, Amann R (2002) Fluorescence in situ hybridization and catalyzed reporter deposition for the
identification of marine bacteria. Applied and Environmental Microbiology 68:3094–3101.
PickeringJ,BamfordA,GodboleV,BriggsJ,ScozzafavaG,RoeP,WheelerC,GhouzeF,CussS(2002)IntegrationofDNAligationand rollingcircleamplification for thehomogeneous,end-pointdetection of single nucleotide polymorphisms. Nucleic Acids Research 30:e60.
PozhitkovA,NoblePA,Domazet-LosˇoT,NolteAW,SonnenbergR,StaehlerP,BeierM,TautzD(2006)TestsofrRNAhybridizationto microarrays suggest that hybridization characteristics of oligonucleotide probes for species discrimination cannot be predicted.Nucleic Acids Research 34:e66.
PozhitkovAE,TautzD,NoblePA(2007)Oligonucleotidemicroarrays:widely applied – poorly understood. Briefings in Functional Genomics & Proteomics 6:141–148.
PrinsTW,vanDijkJP,BeenenHG,VanHoefAA,VoorhuijzenMM,SchoenCD,AartsHJ,KokEJ(2008)Optimisedpadlockprobeligation and microarray detection of multiple (non-authorised)GMOsinasinglereaction. BMC Genomics 9:584.
PunjaZK,WanA,GoswamiRS,VermaN,RahmanM,BarasubiyeT,SeifertKA,LévesqueCA(2007)DiversityofFusarium species associated with discolored ginseng roots in British Columbia.Canadian Journal of Plant Pathology 29:340–353.
Reeleder RD, Capella BB, Tomlinson LD, HickeyWJ (2003) Theextraction of fungal DNA from multiple large soil samples.Canadian Journal of Plant Pathology 25:182–191.
RobideauGP,CarusoFL,OudemansPV,McManusPS,RenaudMA,AuclairME,BilodeauGJ,YeeD,DésaulniersNL,DeVernaJW,LévesqueCA(2008)DetectionofcranberryfruitrotfungiusingDNAarray hybridization.Canadian Journal of Plant Pathology 30:226–240.
RobinBJ,ArffaRC,AvniI,RaoNA(1986)Rapidvisualizationofthreecommonfungiusingfluorescein-conjugatedlectins.Investigative Ophthalmology & Visual Science 27:500–506.
Scupham AJ, Presley LL, Wei B, Bent E, Griffith N, McPhersonM,ZhuF,OluwadaraO,RaoN,Braun J,BornemanJ (2006)Abundantanddiversefungalmicrobiotainthemurineintestine.Applied and Environmental Microbiology 72:793–801.
Seifert,KA,LévesqueCA(2004)Phylogenyandmoleculardiagnosisof mycotoxigenic fungi. European Journal of Plant Pathology 110:449–471.
SeifertKA,SamsonRA,deWaardJR,HoubrakenJ,LévesqueCA,MoncalvoJ-M,Louis-SeizeGandHebertPDN(2007)Prospectsfor fungus identification using CO1 DNA barcodes, withPenicilliumasatestcase.Proceedings of the National Academy of Sciences, USA 104:3901–3906.
Shepard JR, Addison RM, Alexander BD, Della-Latta P, GhernaM, Haase G, Hall G, Johnson JK, Merz WG, Peltroche-LlacsahuangaH,StenderH,VeneziaRA,WilsonD.,ProcopGW,WuF,FiandacaMJ(2008)MulticenterevaluationoftheCandida albicans/Candida glabratapeptidenucleicacidfluorescentinsituhybridizationmethodforsimultaneousdual-coloridentificationofC. albicans and C. glabratadirectly frombloodculturebottles.Journal of Clinical Microbiology 46:50–55.
SholbergPL,O’GormanD,BedfordK,LévesqueCA(2005)DevelopmentofaDNAmacroarrayfordetectionandmonitoringofeconomicallyimportantapplediseases.Plant Disease 89:1143–1150.

Molecular identification and detection of fungi ARTIC
LE
189v o l u m e 2 · n o . 2
SholbergPL,O’GormanD,BedfordKE (2006)MonitoringErwinia amylovora in pome fruit orchards using reverse dot-blot hybridization.Acta Horticulturae 704:91-98.
Stanley KK, Szewczuk E (2005) Multiplexed tandem PCR: geneprofiling from small amounts of RNA using SYBR Greendetection.Nucleic Acids Research 33:e180.
SummerbellRC,LévesqueCA,SeifertKA,BoversM,FellJW,DiazMR, Boekhout T, de Hoog GS, Stalpers J, Crous PW (2005)Microcoding: thesecondstep inDNAbarcoding.Philosophical Transactions of the Royal Scoiety, B 360:1897–1903.
Sun J, Li X, Zeng H, Xie Z, Lu C, Xi L, de Hoog GS (2010a)Development and evaluation of loop-mediated isothermalamplification (LAMP) for the rapid diagnosis of Penicillium marneffei in archived tissue samples.FEMS Immunology and Medical Microbiology 58:381–388.
Sun J, Najafzadeh MJ, Vicente V, Xi L, de Hoog GS (2010b)Rapid detection of pathogenic fungi using loop-mediatedisothermal amplification, exemplified by Fonsecaea agents of chromoblastomycosis. Journal of Microbiological Methods 80: 19–24.
SzemesM,BonantsP,deWeerdtM,BanerJ,LandegrenU,SchoenCD(2005)Diagnosticapplicationofpadlockprobes–multiplexdetectionofplantpathogensusinguniversalmicroarrays.Nucleic Acids Research 33:e70.
Tambong JT, de Cock AWAM, Tinker NA, Lévesque CA (2006)OligonucleotidearrayforidentificationanddetectionofPythium species. Applied and Environmental Microbiology 72: 2691–2706.
TatibanaBT,SanoA,UnoJ,KameiK,IgarashiT,MikamiY,MiyajiM,NishimuraK,ItanoEN(2009)DetectionofParacoccidioides brasiliensis gp43 gene in sputa by loop-mediated isothermal amplificationmethod.Journal of Clinical Laboratory Analysis 23: 139–143.
Tomita N, Mori Y, Kanda H, Notomi T (2008) Loop-mediatedisothermalamplification(LAMP)ofgenesequencesandsimplevisualdetectionofproducts.Nature Protocols 3:877–882.
Tsui CKM, Wang B, Khadempour L, Alamouti SM, BohlmannJ, Murray BW, Hamelin RC (2010) Rapid identification anddetection of pine pathogenic fungi associated with mountain pine beetlesbypadlockprobes.Journal of Microbiological Methods 83:26–33.
TsuiCKM,WangB,SchoenC,HamelinRC(2012)Padlockprobefor the detection of pathogenic fungi. In Laboratory Protocols in Fungal Biology: Current Methods in Fungal Biology (GuptaVA,TuohyM,AyyachamyM,TurnerKM,O’DonovanA,eds):inpress.Springer-Verlag.
Uehara T, Kushida A, Momota Y (1999) Rapid and sensitiveidentification of Pratylenchus spp. using reverse dot blothybridization.Nematology 1:549–555.
UrakawaH, Fantroussi SE, SmidtH, Smoot JC,TribouEH, KellyJJ,NoblePA,StahlDA(2003)Optimizationofsingle-base-pairmismatchdiscriminationinoligonucleotidemicroarrays.Applied and Environmntal Microbiology 69:2848–2856.
van Doorn R, Szemes M, Bonants P, Kowalchuk GA, Salles JF,OrtenbergE,SchoenCD(2007)Quantitativemultiplexdetectionof plant pathogens using a novel ligation probe-based system coupled with universal, high-throughput real-time PCR on
OpenArrays®.BMC Genomics 8:276.van Doorn R, Slawiak M, Szemes M, Dullemans AM, Bonants
P, Kowalchuk GA, Schoen CD (2009) Robust detection andidentificationofmultipleoomycetesand fungi inenvironmentalsamples by using a novel cleavable padlock probe-based ligation detection assay. Applied and Environmental Microbiology 75: 4185–4193.
WangB,DwyerDE,ChewCB,KolC,HeZP,JoshiH,SteainMC,CunninghamAL,SaksenaNK(2009)SensitivedetectionoftheK103Nnon-nucleosidereversetranscriptaseinhibitorresistancemutationintreatment-naiveHIV-1infectedindividualsbyrollingcircleamplification.Journal of Virological Methods 161:128–135.
Wang B, Dwyer DE, Blyth CC, Soedjono M, Shi H, Kesson A,RatnamohanM,McPhieK,CunninghamAL,SaksenaNK(2010)DetectionoftherapidemergenceoftheH275YmutationassociatedwithoseltamivirresistanceinseverepandemicinfluenzavirusA/H1N109infections.Antiviral Research 87:16–21.
WangY,KangZ,GaoH,GaoY,QinL,LinH,YuF,QiX,WangX(2011)Aone-stepreversetranscriptionloop-mediatedisothermalamplificationfordetectionanddiscriminationofinfectiousbursaldiseasevirus.Virology Journal 8:108.
Weber SD, Ludwig W, Schleifer K-H, Fried J (2007) Microbialcompositionandstructureofaerobicgranularsewagebiofilms. Applied and Environmental Microbiology 73:6233–6240.
Wilson DA, Joyce MJ, Hall LS, Reller LB, Roberts GD, Hall GS,Alexander BD, Procop W (2005) Multicenter evaluation ofa Candida albicans peptide nucleic acid fluorescent in situhybridization probe for characterization of yeast isolates from bloodcultures.Journal of Clinical Microbiology 43:2909–2912.
XiongL,KongF,YangY,ChengJ,GilbertGL(2006)UseofPCRandreverse line blot hybridization macroarray based on 16S-23SrRNA gene internal transcribed spacer sequences for rapididentification of 34Mycobacterium species. Journal of Clinical Microbiology 44:3544–3550.
Yang YP, Corley N, Garcia-Heras J (2001) Reverse dot-blothybridization as an improved tool for the molecular diagnosis of point mutations in congenital adrenal hyperplasia caused by 21-hydroxylasedeficiency.Molecular Diagnostics 6:193–199.
Zhang N, Geiser MD, Smart CD (2007) Macroarray detection ofsolanaceous plant pathogens in the Fusarium solani species complex.Plant disease 91:1612–1620.
ZhangN,McCarthyML,SmartCD(2008)Amacroarraysystemforthe detection of fungal and oomycete pathogens of solanaceous crops.Plant disease 92:953–960.
ZhangY,CoyneMY,WillSG,LevensonCH,KawasakiES (1991)Single-basemutationalanalysisofcancerandgeneticdiseasesusingmembraneboundmodifiedoligonucleotides.Nucleic Acids Research 19:3929–3933.
ZhaoK,ShiW,HanF,XuY,ZhuL,ZouY,WuX,ZhuH,TanF,Tao S, Tang X (2011). Specific, simple and rapid detection ofporcine circovirus type 2 using the loop-mediated isothermalamplificationmethod.Virology Journal 8:126.
Zhou X, Kong F, Sorrell TC, Wang H, Duan Y, Chen SC (2008)Practical method for detection and identification of Candida, Aspergillus, and Scedosporium spp. by use of rolling-circleamplification.Journal of Clinical Microbiology 46:2423–2427.


















