
Molecular Phylogeny of Cercomonadidae and Kinetid Patterns ... · Molecular Phylogeny of...
34
Protist, Vol. 157, 125—158, April 2006 http://www.elsevier.de/protis Published online date 26 April 2006 Molecular Phylogeny of Cercomonadidae and Kinetid Patterns of Cercomonas and Eocercomonas gen. nov. (Cercomonadida, Cercozoa) Serguei A. Karpov a , David Bass b , Alexander P. Mylnikov c , and Thomas Cavalier-Smith b,1 a Department of Zoology, Herzen State Pedagogical University of Russia, Moika emb. 48, 191186, St. Petersburg, Russian Federation b Department of Zoology, University of Oxford, South Parks Road, Oxford OX1 3PS, UK c Institute for the Biology of Inland Waters RAS, Borok 152742, Russian Federation Submitted February 28, 2005; Accepted January 24, 2006 Monitoring Editor: Michael Melkonian Cercomonads are among the most abundant and widespread zooflagellates in soil and freshwater. We cultured 22 strains and report their complete 18S rRNA sequences and light microscopic morphology. Phylogenetic analysis of 51 Cercomonas rRNA genes shows in each previously identified major clade (A, B) two very robust, highly divergent, multi-species subclades (A1, A2; B1, B2). We studied kinetid ultrastructure of five clade A representatives by serial sections. All have two closely associated left ventral posterior microtubular roots, an anterior dorsal root, a microtubule-nucleating left anterior root, and a cone of microtubules passing to the nucleus. Anterior centrioles ( ¼ basal bodies, kinetosomes) of A1 have cartwheels; the posterior centriole does not, suggesting it is older, and implying flagellar transformation similar to other bikonts. Strain C-80 (subclade A2) differs greatly, having a dorsal posterior microtubule band, but lacking the A1-specific fibrillar striated root, nuclear extension to the centrioles, centriolar diaphragm, extrusomes; both mature centrioles lack cartwheels. For clade A2 we establish Eocercomonas gen. n., with type Eocercomonas ramosa sp. n., and for clade B1 Paracercomonas gen. n. (type Paracercomonas marina sp. n.). We establish Paracercomonas ekelundi sp. n. for culture SCCAP C1 and propose a Cercomonas longicauda neotype and Cercomonas ( ¼ Neocercomonas) jutlandica comb. n. and Paracercomonas ( ¼ Cercomonas) metabolica comb. n. & 2006 Elsevier GmbH. All rights reserved. Keywords: centriolar transformation; Cercomonas; Eocercomonas; kinetid structure; Paracercomonas; 18S rRNA phylogeny. Introduction The recently established phylum Cercozoa ( Cavalier- Smith 1998) has turned out to be one of the most morphologically diverse of all in the kingdom Protozoa (Cavalier-Smith and Chao 2003; Nikolaev et al. 2004; Polet et al. 2004) as well as one of the most speciose (Bass and Cavalier- Smith 2004). The classical genus Cercomonas ARTICLE IN PRESS 1 Corresponding author. e-mail [email protected] (T. Cavalier-Smith). & 2006 Elsevier GmbH. All rights reserved. doi:10.1016/j.protis.2006.01.001
Transcript of Molecular Phylogeny of Cercomonadidae and Kinetid Patterns ... · Molecular Phylogeny of...

ARTICLE IN PRESS
http://www.elsevier.de/protisPublished online date 26 April 2006
1
Correspondine-mail tom.ca
& 2006 Elsevdoi:10.1016/j
157, 125—158, April 2006
Protist, Vol.Molecular Phylogeny of Cercomonadidaeand Kinetid Patterns of Cercomonasand Eocercomonas gen. nov.(Cercomonadida, Cercozoa)
Serguei A. Karpova, David Bassb, Alexander P. Mylnikovc, and Thomas Cavalier-Smithb,1
aDepartment of Zoology, Herzen State Pedagogical University of Russia, Moika emb. 48, 191186,St. Petersburg, Russian Federation
bDepartment of Zoology, University of Oxford, South Parks Road, Oxford OX1 3PS, UKcInstitute for the Biology of Inland Waters RAS, Borok 152742, Russian Federation
Submitted February 28, 2005; Accepted January 24, 2006Monitoring Editor: Michael Melkonian
Cercomonads are among the most abundant and widespread zooflagellates in soil and freshwater. Wecultured 22 strains and report their complete 18S rRNA sequences and light microscopic morphology.Phylogenetic analysis of 51 Cercomonas rRNA genes shows in each previously identified major clade(A, B) two very robust, highly divergent, multi-species subclades (A1, A2; B1, B2). We studied kinetidultrastructure of five clade A representatives by serial sections. All have two closely associated leftventral posterior microtubular roots, an anterior dorsal root, a microtubule-nucleating left anteriorroot, and a cone of microtubules passing to the nucleus. Anterior centrioles ( ¼ basal bodies,kinetosomes) of A1 have cartwheels; the posterior centriole does not, suggesting it is older, andimplying flagellar transformation similar to other bikonts. Strain C-80 (subclade A2) differs greatly,having a dorsal posterior microtubule band, but lacking the A1-specific fibrillar striated root,nuclear extension to the centrioles, centriolar diaphragm, extrusomes; both mature centrioleslack cartwheels. For clade A2 we establish Eocercomonas gen. n., with type Eocercomonas ramosasp. n., and for clade B1 Paracercomonas gen. n. (type Paracercomonas marina sp. n.). Weestablish Paracercomonas ekelundi sp. n. for culture SCCAP C1 and propose a Cercomonaslongicauda neotype and Cercomonas ( ¼ Neocercomonas) jutlandica comb. n. and Paracercomonas( ¼ Cercomonas) metabolica comb. n.& 2006 Elsevier GmbH. All rights reserved.
Keywords: centriolar transformation; Cercomonas; Eocercomonas; kinetid structure; Paracercomonas; 18SrRNA phylogeny.
Introduction
The recently established phylum Cercozoa (Cavalier-Smith 1998) has turned out to be one of the most
g [email protected] (T. Cavalier-Smith).
ier GmbH. All rights reserved..protis.2006.01.001
morphologically diverse of all in the kingdomProtozoa (Cavalier-Smith and Chao 2003;Nikolaev et al. 2004; Polet et al. 2004) as well asone of the most speciose (Bass and Cavalier-Smith 2004). The classical genus Cercomonas

ARTICLE IN PRESS
126 S.A. Karpov et al.
(Dujardin 1841) is the second most commonly andwidely encountered zooflagellate genus in soil(Ekelund and Patterson 1997; Foissner 1991;Sandon 1927); they are also very common infreshwater (Arndt et al. 2000). Excluding the doubt-ful taxa of Skvortzov (1977), at least 49 Cercomonasspecies have been named (Mylnikov and Karpov2004), but sequences of 18S rRNA genes amplifiedfrom environmental DNA extracts suggest that thereal number is well over a hundred (Bass andCavalier-Smith 2004). Moreover, the status of many— probably the majority — of the named species isuncertain, and the taxonomy of the genus urgentlyneeds revision (Al-Qassab et al. 2002; Foissner1991). Only a few species have been named fromclonal cultures allowing proper appreciation of theirrange of variation and identification of cysts andplasmodial stages, if present (Mylnikov 1992c).Recently it became clear that there are two verydistinct clades of Cercomonas on rRNA trees,which only sometimes group together as sisters(Bass and Cavalier-Smith 2004, Bass et al. 2005;Cavalier-Smith and Chao 2003).
A sounder Cercomonas taxonomy will comefrom integrated studies of numerous clonal cul-tures by light microscopy and DNA sequencing,and sampling a representative selection for thin-section electron microscopy. Accordingly we haveisolated 21 new, mostly clonal Cercomonascultures from freshwater and soil in Russia,Western Europe, Panama, and New Zealand,completely sequenced their 18S rRNA genes,and that of C. longicauda CCAP 1910/2, andcarried out phylogenetic analysis to place them onthe rapidly growing Cercomonas phylogenetic tree(51 sequences, including 10 additional previouslyunpublished sequences from environmental genelibraries to provide a more comprehensive tree).We also show phase contrast light micrographs ofthe general morphology of the cultured strains.Only a small number can be assigned to de-scribed species, but we shall give new names tomost of those that cannot be identified thus onlyafter studying still more strains.
Ekelund et al. (2004) recently confirmed thedistinctiveness of Cercomonas clades A and B(their ‘‘type 1’’ is part of our clade B, and ‘‘type 2’’part of clade A), described a new speciesNeocercomonas jutlandica within clade A, andapplied the new generic name Neocercomonas toall ‘‘type 2’’ species. They introduced the usefulidea of using signature sequences to improvecercomonad taxonomy, and distinguished Neo-cercomonas from Cercomonas solely by 18S rRNAsignature sequences; however, as this was based
on a very small database (7 sequences), theirNeocercomonas signatures do not apply to allclade A, being absent from C. sp. E (Cavalier-Smith and Chao 2003). However, the culturedesignated as neotype for Cercomonas longicau-da Dujardin (1841) by Ekelund et al. (2004) is notactually that species, and for this and otherreasons is invalid as a neotype for it. Our presentsequence signature analysis based on over 100Cercomonas sequences and phylogenetic treesfor 51 Cercomonas sequences clearly show thatclade A is divisible into two major subclades, A1(including N. jutlandica) and A2 (including C. sp. E),of which only clade A1 possesses the Neocerco-monas signatures. As those cercomonads mor-phologically most resembling C. longicauda(Dujardin 1841), the type species of Cercomonas,are all in A1 and have the ‘Neocercomonas’signatures, Neocercomonas is probably a juniorsynonym of Cercomonas and thus is not adoptedhere. The early assumption that the classicalgenus Cercomonas is paraphyletic (Cavalier-Smithand Chao 1996/7) or polyphyletic (Ekelund et al.2004) may be invalid, and a confusion caused bylong-branch attraction artefacts (clade B havinglonger branches than clade A); in agreement withour earlier maximum likelihood analyses (Cavalier-Smith and Chao 2003) the best-sampled recentanalyses consistently show classical Cercomonasas holophyletic by both distance and likelihood,albeit with weak support (Bass and Cavalier-Smith2004; Bass et al. 2005).
Although there have been several ultrastructuralstudies of Cercomonas (Karpov 1997; Mignot andBrugerolle 1975; Mylnikov 1986b, c, 1987, 1989a,b, 1990, 1992a,b, 1995, 2000, 2002; Mylnikov andMylnikova 2001; Mylnikov et al. 2000; Schuster andPollack 1978), many were on unidentified species,none were of strains characterized by molecularphylogeny, and none used serial section recon-struction of flagellar roots, which is important forcharacterizing protist cytoskeletons, and of greatphylogenetic significance (Karpov 2000a; Moestrup2000). The fragmentary studies of cercozoankinetids to date suggest a very considerablevariation in flagellar roots among different majorgroups (reviewed briefly in Cavalier-Smith 2002). Itis therefore possible that even though someCercomonas strains that are genetically verydistinct can look remarkably similar by lightmicroscopy they differ internally quite substantially.
To test this we have serially sectioned andreconstructed kinetid ultrastructure of five phylo-genetically diverse representatives of Cercomo-nas clade A, chosen from among those described

ARTICLE IN PRESS
127Cercomonad Kinetids and Phylogeny
by light microscopy in this paper. This revealscommon features of their cytoskeleton that onlypartially resemble those of the heteromitid Katabiagromovi (also order Cercomonadida and classSarcomonadea), the only phagotrophic cercozoanpreviously with a detailed serial-section study ofits kinetid ultrastructure (Karpov et al. 2003a), andprobable synapomorphies for Sarcomonadea(cercomonads plus heteromitids). Furthermore,the most marked differences among clade Acytoskeletons are between those of subcladesA1 and A2, indicating excellent correlation be-tween kinetid and rRNA divergence. These andmarked non-cytoskeletal ultrastructural differ-ences between clades A1 and A2, plus their evensharper contrast with clade B, clearly justify threeseparate genera for the classical Cercomonas.Accordingly, we establish a new genus Eocerco-monas for clade A2, and a new genus Paracerco-monas for clade B1, and provide more definitivemorphological and genetic signature diagnosesfor Cercomonas sensu stricto. For brevity we usethe term centriole throughout, rather than thesynonyms basal body or kinetosome.
A striking new observation is that in Cercomo-nas sensu stricto (clade A1) the posterior centrioleonly lacks the cartwheel that is invariably presentin the anterior centrioles. Since early developingcentrioles have a central cartwheel as in othereukaryotes (Cavalier-Smith 1974), this shows thatthe posterior centriole is older than the anteriorone and loses the cartwheel after assembly,providing the first evidence in Cercozoa, or anyRhizaria, for the typical bikont pattern of flagellartransformation (Cavalier-Smith 2002). In Eocerco-monas (clade A2) neither mature centriole hascartwheels and in Paracercomonas (clade B1)both do.
Results
We first describe the molecular phylogeny toprovide the evolutionary context for the micro-scopical data, enabling broad patterns to bediscerned.
Phylogenetic Analysis
Figure 1 shows a Bayesian tree for 51 Cercomo-nas sequences; the 21 new strains and 10 newenvironmental sequences are spread widelyacross the tree, 24 in clade A and 7 in cladeB. Two Russian strains (C-56 and C-72) areidentical in sequence, but the other cultured strain
sequences differ so substantially from each otherand previous sequences that they are probablydistinct species. Two other Russian strains clusterfairly closely together as a robust clade (C-59 and C-84), as do two Canadian environmental sequences(13-2.6 and 9-6.2) and two UK environmentalsequences (10-3.6 and 8-3.1), all from differentlibraries. Apart from these, most clades containstrains from widely separate geographic locations.Within clade A there are two very divergentsubclades, each with strong bootstrap support:A1, with 32 taxa and including C. plasmodialis, C.longicauda (CCAP 1910/2), C. ( ¼ N.) jutlandica andC. alexeieffi; A2 comprises only five unidentifiedstrains, for one of which we establish below a newspecies, Eocercomonas ramosa.
The speciose clade A1 itself comprises twodistinct subclades (A1a, A1b). Although A1b lackssignificant bootstrap support, it is supported by atleast one unique signature (Table 1). Signaturesidentified for the A1b subclades (Table 1) stronglysupport the topology shown and indicate thatC. jutlandica lacks close relatives among thepresent data set. The weak bootstrap supportfor clade A1b2, despite strong Bayesian support,is attributable to long-branch attraction pulling thevery long branch of Cercomonas sp. 23 to thebase of clade A1, an artifact to which Bayesian-likelihood methods seem more resistant. Apartfrom this, and a similar tendency for the lone C.jutlandica sequence to move towards the base ofA1 in distance trees, there is excellent agreementbetween the distance and Bayesian analyses.
Clade B also consists of three major subclades(B1a, B1b and B2), all reproducibly holophyletic.Clade B1a is the most speciose, with 9 verydistinct lineages and a fully resolved topology.Note that the two species that most resemble C.cometa in having highly branched filopodia are notmutually related: C-80 is in clade A2 and C-71 inclade B, showing that this morphotype is con-vergent. The two strains, Cs-4 and AZ-6, inde-pendently identified as C. plasmodialis are clearlynot the same species, belonging to subclades A1aand A1b2 respectively; Cs-4 is actually the typestrain (Mylnikov et al. 2000). Paracercomonas sp.‘small’ (morphologically indistinguishable fromFig. 5 of Ekelund et al. 2004) is more closelyrelated to P. ekelundi sp. n. (SCCAP C1, as‘Cercomonas longicauda’ sensu Ekelund et al.(2004); see discussion below) than to P. marina sp.n. (the new name proposed below for the ATCC50344 culture also misidentified as C. longicauda).Most of the few differences (especially all theunique single nucleotide indels) between P. sp.

ARTICLE IN PRESS
Figure 1. Consensus Bayesian likelihood tree for 51 Cercomonadidae sequences using 1013 nucleotidepositions of the 18S rRNA gene, with Heteromitidae (Cercomonadida) as outgroup. A gamma intersitevariation, covarion and autocorrelation GTR substitution model was used: see Methods for details. Thefigures and blobs at the nodes are distance tree bootstrap percentages for this dataset (upper) and Bayesianposterior probabilities (lower). All bootstrap percentages and posterior probabilities X75% are shown; otherslower than this are shown for clades of particular interest to this study. Black blobs indicate 100% bootstrapsupport. Dashes indicate nodes for clades not found in the consensus distance tree. Italicized numericalcodes refer to sequences obtained from Cercozoa-specific environmental gene libraries. The strains whoseultrastructure is described here are shaded in grey; asterisks indicate strains illustrated in Figures 3 and 4.The 32 new sequences are shown in bold. f/w ¼ fresh water.
128 S.A. Karpov et al.
small and P. ekelundi (SCCAP C1) could besequencing errors; some of those that distinguishP. marina (‘Cercomonas longicauda’ ATCC 50344)from P. ekelundi (SCCAP C1) are genuine, havingbeen confirmed by two independent laboratories(Cavalier-Smith and Chao 2003), but others aremore doubtful and probably sequencing errors
(mainly in the AF101052 sequence). However, thefact that these two distinct sequences eachformerly identified as longicauda are in clade B1,whereas the CCAP longicauda is in clade A1means that they are not all the same species, andat least two were misidentified. As the CCAPlongicauda is morphologically markedly closer to

ARTICLE IN PRESS
Ta
ble
1.
18S
rDN
Aseq
uence
sig
natu
res
for
cla
des
of
Cerc
om
onas,
Eo
cerc
om
onas,
and
Para
cerc
om
on
as
as
ind
icate
do
nF
igure
s1
and
2.
Cla
de
co
de
1S
eq
uence
sig
natu
re(b
old
)2P
ositio
n3
No
tes
4
A1
CA
GC
TC
AT
TAA
AT
CA
GT
CA
TT
96/H
8,
~V
2a
A1
GA
GG
GA
CTA
TC
GG
TCG
AT
TTA
1402,
H44
bA
1T
CG
AG
CTT
TAC
AA
CC
TT
GG
TT
CT
1504,
H46
cA
1G
GA
CT
859
A2
TA
AT
T112/H
8,
~V
2d
A2
TC
GA
GC
TT
TAC
AA
CC
TT
GA
CT
1504,
H46
A2
CT
TCC
TG
TT
CTA
TT
TT
GT
TG
GT
TT
CTA
GG
AT
CG
G833,
E23_13/1
4,
V4
A1a
CA
C/T
TC
CA
TC
CT
TC
G700/H
E23
_1,
V4
A1b
CT
CTA
G498/H
17,
V3
A1b
1G
CT
TC
GG
666
eA
1b
1T
CT-
CC
CT
TC
TA
TCT
GG
GT
TG
GAG
CC
GG
A723,
E23
fA
1b
2G
AC
CA
TC
CA
702,
E23_1,
V4
gB
1T
CG
GC
CA
GA
GG
TG
AA
AT
TC
TT
TG
GA
TT
CG
A907,
H25
hB
1T
TA
TA
G112/H
8,
~V
2
1A
sin
dic
ate
do
nF
igure
1.
2B
old
nucle
otid
es¼
ancestr
al
synap
om
orp
hy
for
cla
de
(where
41
nt
mark
ed
,th
at
part
icula
rco
mb
inatio
nis
uniq
ue
toth
ecla
de:
within
Cerc
om
onad
idae,
unle
ss
oth
erw
ise
sta
ted
.S
ub
scrip
tnucle
otid
es¼
rare
seco
nd
ary
chang
es
toth
ep
reced
ing
nucle
otid
e.
3H
elix
(H,E
)and
variab
lere
gio
n(V
)lo
catio
nacco
rdin
gto
Wuyts
et
al.
(2000)/
po
sitio
no
fLH
-mo
st
nucle
otid
eo
fsig
natu
reseq
uence
rela
tive
toC
erc
om
onas
sp
.‘L
arg
e’
(AF
411266
AT
CC
PR
A-2
1).
4N
ote
s:
acf.
sig
natu
res
for
cla
des
A2
and
Bat
sam
ep
ositio
n.
b¼
sig
natu
reseq
uence
N1
inE
kelu
nd
et
al.
(2004).
Insert
ion
of
A(p
os.
1417)is
asynap
om
orp
hy
for
cla
de
A1,
once
chang
ed
toC
(in19-3
E).
c¼
sig
natu
reseq
uence
N2
inE
kelu
nd
et
al.
(2004)to
define
Neo
cerc
om
onas.
Po
ssib
leco
nfu
sio
n/c
onverg
ence
with
cla
de
B.
Thre
ep
ositio
ns
are
variab
leand
co
nverg
ew
ith
B,
but
this
sig
natu
recle
arly
dis
ting
uis
hes
cla
de
A1
fro
mA
2.
dcf.
sig
natu
res
for
cla
des
A1
and
Bat
sam
ep
ositio
n.
eU
niq
ue
within
Cerc
ozo
a.
f ‘—‘
ind
icate
sa
share
dd
ele
tio
nin
this
cla
de.
gU
niq
ue
within
Cerc
ozo
a.
hH
ighlig
hte
dnucle
otid
es
sho
wco
mp
ensato
ryb
ase
chang
esynap
om
orp
hy
for
cla
de
B1
seco
nd
arily
revers
ed
inC
s-9
.
129Cercomonad Kinetids and Phylogeny

ARTICLE IN PRESS
130 S.A. Karpov et al.
Dujardin’s (1841), as explained below, both theATCC and the SCCAP strains were misidentified,and we describe them below as new species in anew genus Paracercomonas.
Sequence Signature Analysis
Sequence signatures that are distinctive forparticular subclades of cercomonads were soughtby visual examination of a large alignment of over100 cercomonad sequences and a representativeselection of other Cercozoa; the signatures shownin Table 1 are not necessarily unique to cercomo-nads, as a complete eukaryote alignment was notstudied, but all unambiguously differentiate thespecified subclades from all other cercomonads inour database, and in some cases also from allother Cercozoa, as noted in Table 1. Sequencesignature analysis strongly supports the majorsplit between clades A1 and A2, with fourclear synapomorphies for each. However, N2 ofEkelund et al. (2004) does not discriminatebetween clade A1 and all clade B sequences inour database, as several clade B sequenceshave convergent substitutions. Thus only thefirst three A1 signatures are taxonomically diag-nostic for clade A1. All the sequence signaturesin Table 1 were checked for consistency withover 50 additional complete unpublished Cerco-monas sequences in our database (Bass and
Figure 2. Comparative alignment of the regions susignatures. Shaded regions indicate positions conservdiagnostic signatures and their position relative to the sePRA-21), which are also shown on Table 1; helix (H, E)et al. (2000) are also shown for each alignment regioboundaries between Clade A, Clade B, and other Cerc
Cavalier-Smith) — over 100 sequences altogether.Fig. 2 shows a comparative alignment of theregions surrounding some of the more importantsignatures.
None of the four sequences suggested byEkelund et al. (2004) to be diagnostic for theirtype 1 Cercomonas (our Paracercomonas, seebelow) is diagnostic for the whole of clade B. Thesecond A in their C1 is a synapomorphy only forclade B1, as our ‘Cercomonas’ Tempisque and‘Cercomonas’ sp. ‘anaerobic’ sequences (Cava-lier-Smith and Chao 2003) previously indicated.That mutation of a C to an A is invariablyaccompanied by a G—T substitution 28 nucleo-tides earlier in the sequence and is clearly aconserved and derived complementary base-pairsubstitution that uniquely defines clade B1, exceptfor two known reversions, e.g. in Cs-9; secondarystructure alignments show that these two nucleo-tides are indeed a derived U—A base pair (basepair 7 in helix 25) compared with the ancestralstate (G—C) for Filosa. The first G and last T intheir C4 (Ekelund et al. 2004) is a synapomorphynot for clade B, but for clade B1 only. Their C2 isnot diagnostic for clade B, as it is too variablewithin the clade; our database shows mutations atnine positions. The position of the third T could bediagnostic, as it is always a T, C or A in clade Band always a G in clade A, but its variability wouldmake that choice risky. Their C3 is also far too
rrounding some of the more important sequenceed across Cercozoa. Boxes indicate clade-specificquence of Cercomonas sp. ‘Large’ (AF411266 ATCCand variable region (V) location according to Wuytsn. The dashed and solid horizontal lines mark theozoa.
ARTICLE IN PRESS
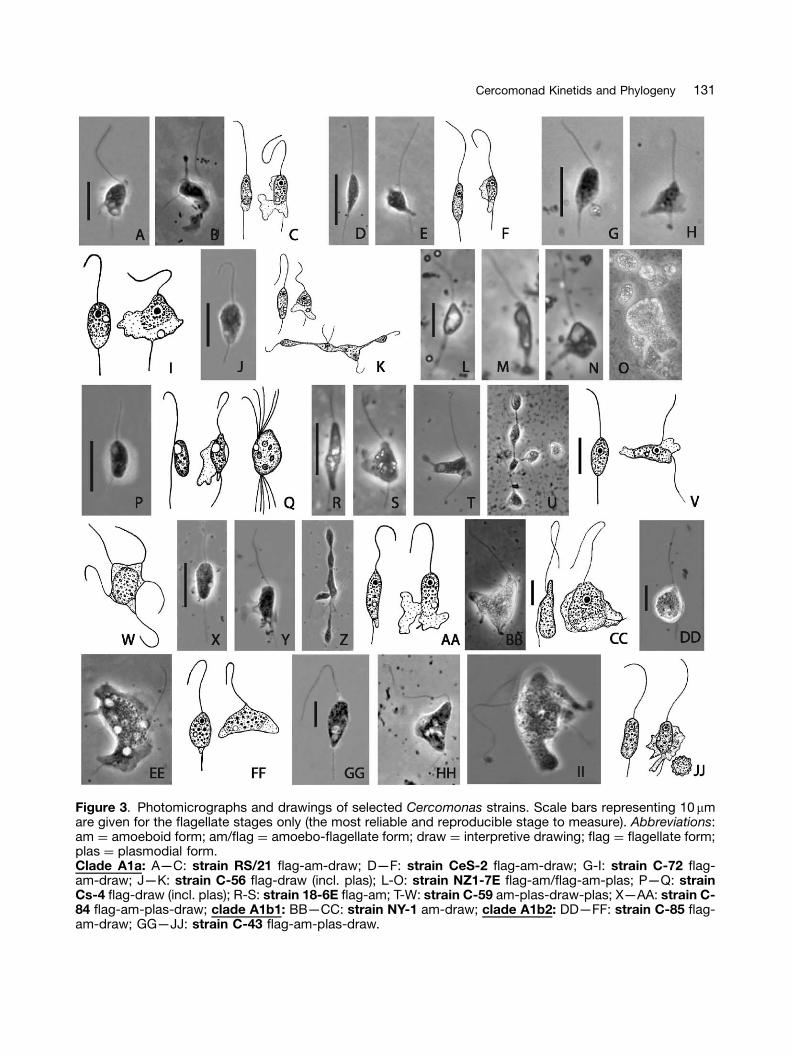
Figure 3. Photomicrographs and drawings of selected Cercomonas strains. Scale bars representing 10mmare given for the flagellate stages only (the most reliable and reproducible stage to measure). Abbreviations:am ¼ amoeboid form; am/flag ¼ amoebo-flagellate form; draw ¼ interpretive drawing; flag ¼ flagellate form;plas ¼ plasmodial form.Clade A1a: A—C: strain RS/21 flag-am-draw; D—F: strain CeS-2 flag-am-draw; G-I: strain C-72 flag-am-draw; J—K: strain C-56 flag-draw (incl. plas); L-O: strain NZ1-7E flag-am/flag-am-plas; P—Q: strainCs-4 flag-draw (incl. plas); R-S: strain 18-6E flag-am; T-W: strain C-59 am-plas-draw-plas; X—AA: strain C-84 flag-am-plas-draw; clade A1b1: BB—CC: strain NY-1 am-draw; clade A1b2: DD—FF: strain C-85 flag-am-draw; GG—JJ: strain C-43 flag-am-plas-draw.
131Cercomonad Kinetids and Phylogeny
ARTICLE IN PRESS
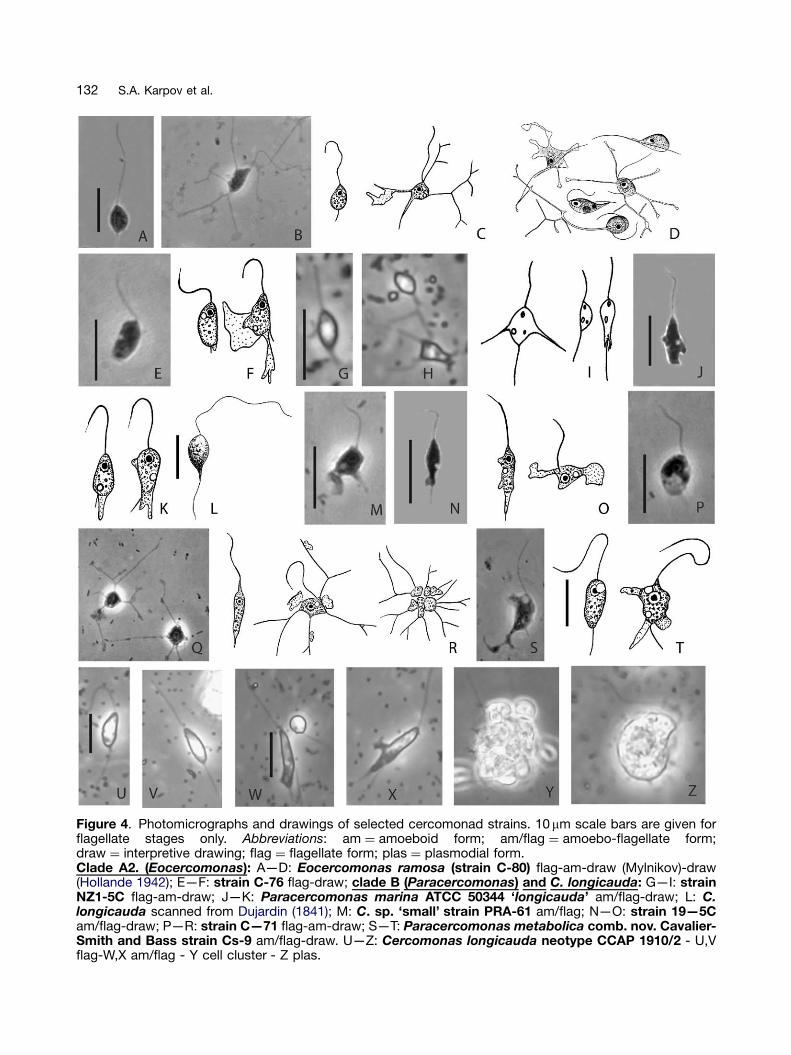
Figure 4. Photomicrographs and drawings of selected cercomonad strains. 10mm scale bars are given forflagellate stages only. Abbreviations: am ¼ amoeboid form; am/flag ¼ amoebo-flagellate form;draw ¼ interpretive drawing; flag ¼ flagellate form; plas ¼ plasmodial form.Clade A2. (Eocercomonas): A—D: Eocercomonas ramosa (strain C-80) flag-am-draw (Mylnikov)-draw(Hollande 1942); E—F: strain C-76 flag-draw; clade B (Paracercomonas) and C. longicauda: G—I: strainNZ1-5C flag-am-draw; J—K: Paracercomonas marina ATCC 50344 ‘longicauda’ am/flag-draw; L: C.longicauda scanned from Dujardin (1841); M: C. sp. ‘small’ strain PRA-61 am/flag; N—O: strain 19—5Cam/flag-draw; P—R: strain C—71 flag-am-draw; S—T: Paracercomonas metabolica comb. nov. Cavalier-Smith and Bass strain Cs-9 am/flag-draw. U—Z: Cercomonas longicauda neotype CCAP 1910/2 - U,Vflag-W,X am/flag - Y cell cluster - Z plas.
132 S.A. Karpov et al.

ARTICLE IN PRESS
133Cercomonad Kinetids and Phylogeny
variable, 16 of the nucleotide positions varyingwithin clade B, and under half our clade Bsequences sharing C3. Thus of their signaturesfor Cercomonas sensu Ekelund et al. (2004) woulddelimit clade B1, whereas the other two woulddefine a broader group; thus C1-C4 are notsuitable collectively for defining any genus. Thesignature sequence that Ekelund et al. (2004)used to define Cercomonas ‘longicauda’ (ourP. ekelundi) is not totally specific for theirstrain, but would identify six very closely relatedribotypes in our database (including Cercomonassp. small; its 4 unique single-nucleotide indelsand one in their strain sequence are probablysequencing errors) differing in very few nucleo-tides elsewhere in the molecule (sometimesjust one).
Light Microscope Morphology
Figure 3 shows phase contrast micrographs anddrawings of the strains at different stages of thelife cycle of a broad sample of 12 clade A1 strains(Cercomonas sensu stricto) arranged in the orderon the tree (Fig. 1). It is immediately obvious thatsubclade A1a cells are on average smaller thanA1b cells: mostly under 10mm long, whereasclade A1b cells are mostly over 10 mm and haveparticularly long flagella. Cercomonas jutlandica,the sole representative of clade A1b3, is also large(10—16mm) (Ekelund et al. 2004) and most closelyresembles C-85 from clade A1b2 (Figs 3 DD—FF).Of the four strains first studied phylogenetically(Cavalier-Smith and Chao 1996/7), sp. 23 (cladeA1b2) was markedly larger than species 21 and 22(clade A1a1) or 18 (clade B). Figure 4 similarlyillustrates two clade A2 and six clade B strains;most are similar in size to those of clade A1a. Thissurprising correlation between cell size and large-scale phylogeny needs further testing, but itappears that cercomonads were ancestrally smalland that cell size increased in the commonancestor of clade A1b; this increase may besignificant in relation to the marked ultrastructuraldifferences in cytoskeleton between clades A1and A2 reported below.
Inspection of Figure 3 also shows that pseudo-pods of all clade A1 strains are broad lamellipodia,never branched filopodia. However, althoughsome members of clades A2 and B producesimilar broad pseudopods, others have simpleor branched filopodia. C-80 in clade A2 andC-71 in clade B produce very similar highlybranched pseudopodia; initially both were mis-identified as Cercobodo cometa, but differences
in flagella dimensions indicate that neither is reallyC. cometa. Clearly their branched filopodiaevolved independently; there must be at leastthree different species of cercomonads withcometa-like morphology, so great care is neededin future identification. Both strains constitutingsubclade A1a2 (C-59 and C-84) are very prone toform linear chains of incompletely divided cells(Figs 3 U,Z). This behaviour is rarer in clade A1a1,being noted only in C-56 (Fig. 3 K). It is significantthat all A1a1 strains are very similar morphologi-cally; yet small differences can be seen, evenbetween the two strains with identical 18S rDNAsequences (C-72 and C-56), making it likely thatall 51 strains shown in Figure 1 are differentspecies.
The classical distinction between Cercobodo(flagella separate from cell, e.g. NY-1 of cladeA1b1 and Cs-4 of clade A1a1) and Cercomonas(flagella strongly adhering all along the length ofthe cell, e.g. C-43 of A1b2) (Krassilstschik 1886) isnot of deep phylogenetic significance, but can bea very reliable character for discriminating certainstrains. Contractile vacuoles are generally presentand occur at specific places in motile cells of eachstrain. Note that P. marina (ATCC ‘C. longicauda’)(Figs 4 J,K) is not markedly different in morphologyfrom the closely related, but ribotypically distinctP. ekelundi (‘C. longicauda’ of Ekelund et al. 2004);judging from their figs 5 and 12 (not clearly statedin their description), their strain may have a greatertendency for the posterior flagellum to adhere tothe trailing cytoplasm. Our Paracercomonas‘small’ is closely related to and morphologicallyvery similar to both. Descriptions are givenbelow of the five clade A strains selected forultrastructural study.
Cercomonas strain C-84 (Figs 3 X—AA)
Cells are ovoid or spindle shaped, 8.3—11.6mmlong. Both flagella are 1.5—2 times longer than thecell. Contractile vacuole is in the posterior part ofthe cell. Small plasmodia are about 25mm indiameter. Cysts not found.
Cercomonas strain C-59 (Figs 3 T—W)
Spindle-shaped body with pointed posterior end,9.9—13.2mm in length. Anterior flagellum 1.5—2times, and posterior 2—3 times longer than body.Contractile vacuole is in the posterior part of thecell. Pseudopodia flat and broad. May produceplasmodia. Cysts 5—6.6mm in diameter.

ARTICLE IN PRESS
134 S.A. Karpov et al.
Cercomonas strain RS/21 (ATCC 50317:Cercomonas sp. 21 of Cavalier-Smith andChao 1996/7: Figs 3 A—C)
Moving cells are oval, 8—14mm long, anteriorflagellum is 2—2.5 longer and the posterior is1.5—2 times longer than the cell body. One, rarelytwo, contractile vacuoles lie at the anterior part ofthe cell near the nucleus. Unbranched lamellatepseudopodia form at the posterior or middle partof the cell. Cyst diameter 5—8mm.
Figure 5. Kinetid orientation and flagellar roots ofCercomonas (clade A1). The direction of some of theroots is shown. A: View from the right side of the cell.B: View from the dorsal side of the cell. A — Anterior,P — Posterior ends; D — Dorsal, V — Ventral, L —Left, R — Right sides of the cell. As the discussionexplains, the anterior centriole is younger and theposterior one older; thus flagellum 1 is posterior andflagellum 2 anterior, according to recent convention(Moestrup 2000), and we label the ventral posteriorroots vp1 and vp2, according to their respectiveattachments to posterior centriole 1 and anteriorcentriole 2.Abbreviations for Figures 5—14: ab — anteriorfibrillar bridge, ac — centriole (basal body, kineto-some) of anterior flagellum, af — anterior flagellum,ag — Golgi apparatus, c — centriole (basal body,kinetosome), cf — concentric fibre (rings), cs —centrioles (basal bodies), cv — contractile vacuole,da — dorsal anterior root, di — diaphragm structure,dp — dorsal posterior root, dsm — dorsal band ofsecondary microtubules, er — endoplasmic reticu-lum, fb — fibrillar bridges connecting centrioles andcentrioles with roots, ff — fibrillar foot, fr—fibrillarroot, fsr — fibrillar striated root, lr — left anterior root,m — mitochondrion, mb — microbody, mc —microtubular cone, mr—microtubular root, mrb —
Cercomonas strain C-43 (Figs 3 GG—JJ)
The cell body has a broad-oval shape; markedlymetabolic. There are one or two tail pseudopodia,it can also produce short lateral ones. Cell lengthis normally 15—20mm; equal flagella a bit longerthan the body. Recurrent flagellum applies tothe cell, even if it turns or overcomes obstacles.One or two contractile vacuoles at the cellposterior. This strain was studied ultrastructurallyas C. crassicauda by Mylnikov (1989a), but as it ismuch larger than crassicauda of Dujardin (1841;6—10 mm) and has an obviously projecting poster-ior flagellum not usually obscured by a cytoplas-mic tail, it was probably misidentified. Note thatstrain RS18A, which is in clade B1a, unlike C-43,was labeled C. crassicauda by Cavalier-Smithand Chao (1986/7) following tentative identifica-tion by Humphrey Smith (Coventry), but thatmay not be correct either and the true identityof C. crassicauda requires clarification. Cysts9—15 mm; surface rough or wavy with bluntangular protuberances.
mushroom-like bodies, mtc—microtoxicysts, n —nucleus, nu — nucleolus, ob—osmiophilic bodies,og — osmiophilic granule, om — osmiophilicmaterial, pc—centriole of posterior flagellum, pf —posterior flagellum, pp — proximal partition of pc, pr— fibrillar projection from pc, ps — pseudopodium,sb — symbiotic bacteria, sm—secondary microtu-bules, sma—pair of microtubules from anteriorcentriole, sml — secondary microtubules from leftroot, vp — ventral posterior roots, vp1 — ventralposterior root of posterior flagellum, vp2 — ventralposterior root of anterior flagellum.
Eocercomonas ramosa sp. nov. strain C-80(Figs 4 A—D)
Moving cells are spindle shaped, 5—15mm long.Proximal part of anterior flagellum is surroundedby a cytoplasmic sheath. Posterior flagellum isalmost invisible. Contractile vacuole is in theanterior part of the cell. Trophic cells producehighly branched filopodia, and may group to-gether forming consortia. Cysts infrequent;smooth, spherical; c. 5 mm.
Almost identical to C-71, but has much longeranterior flagellum. Cysts not recorded in C-71. C-80 and C-71 are similar to Cercobodo cometaHollande (1942), but differ in both having relativelymuch shorter posterior flagella.
Electron Microscopy
Kinetid pattern and general ultrastructure of fivecercomonad strains were investigated: C-84, RS/21, C-43, C-59 from clade A1 (Cercomonas) and

ARTICLE IN PRESS
Figure 6. Reconstruction of kinetid structure ofCercomonas strain C-84 (clade A1). Viewed fromabove (dorsal side) but from the right (i.e. at an angleintermediate between Figs 5 A and B; for abbrevia-tions see Fig. 5 legend). To display the roots moreclearly in this perspective drawing and for ease ofcomparison with strain C-80 (Fig. 14) the apicalangle between the centrioles has been altered fromthe correct 90o to make it more obtuse than it is inthis strain.
135Cercomonad Kinetids and Phylogeny
C-80 from clade A2 (Eocercomonas ramosa).All strains studied differ in details of flagellarapparatus and general ultrastructure. Generalultrastructural features of C-43 (as Cercomonas‘crassicauda’) investigated earlier (Mylnikov 1989)were confirmed. Although four other strains (C-84,RS/21, C-59 and C-80) were investigated for thefirst time, we focus mainly on their kinetidstructure, which is complex and substantiallydifferent from that of all other protists. To orientthe reader we outline common features beforedescribing each strain in detail.
Both flagella insert at the cell apex and aremutually attached by their centrioles (shownschematically in Figs 5 A,B). Flagella are smooth,lacking paraxonemal structures. The anteriorflagellum points slightly to the left of the centralline and often beats asymmetrically to the left.Both centrioles lie almost in one plane. Thecentriole of the anterior flagellum typically pointsforward and to the left; the base of the posteriorcentriole is attached to the middle part of theventral side of the anterior centriole at an angle of901 or more, and its flagellum bends back to runalong the mid line of the cell. It usually lies in agroove or channel, depending on the species, but
in some may be less closely attached to the cell.At least three fibrillar bridges interconnect thecentrioles. The anterior fibrillar bridge is bestdeveloped (Fig. 5 A). The centriole of the posteriorflagellum has much more prominent fibrillar matrixaround it than the anterior one.
Two prominent ventral posterior roots of severalmicrotubules are always associated with thecentrioles, one (VP2) stemming from the anteriorand one (VP1) from the posterior centriole. Theyprobably provide mechanical support for theventral groove/channel when it is present (C-80,C-43). As the cells are too large for economicallyserially reconstructing the entire cytoskeleton, thispaper concentrates on flagellar transition zones,centrioles, and nearby cytoskeletal structures,which include many dense fibres and a variety ofmicrotubular roots (summarized in Fig. 6 for cladeA1). In all strains the nucleus is subtended by amicrotubular cone emanating from a nucleatingcentre associated with the centrioles, but itsnature and the geometry of association vary. Asexplained in the discussion, ultrastructural differ-ences among the strains correlate well withthe molecular tree. Those of the four clade A1strains (C-84, RS/21, C43, C-59) are broadlysimilar and described first, whereas C-80 (cladeA2) differs in important respects. We describe C-84 in detail; the other species are more brieflycompared with it.
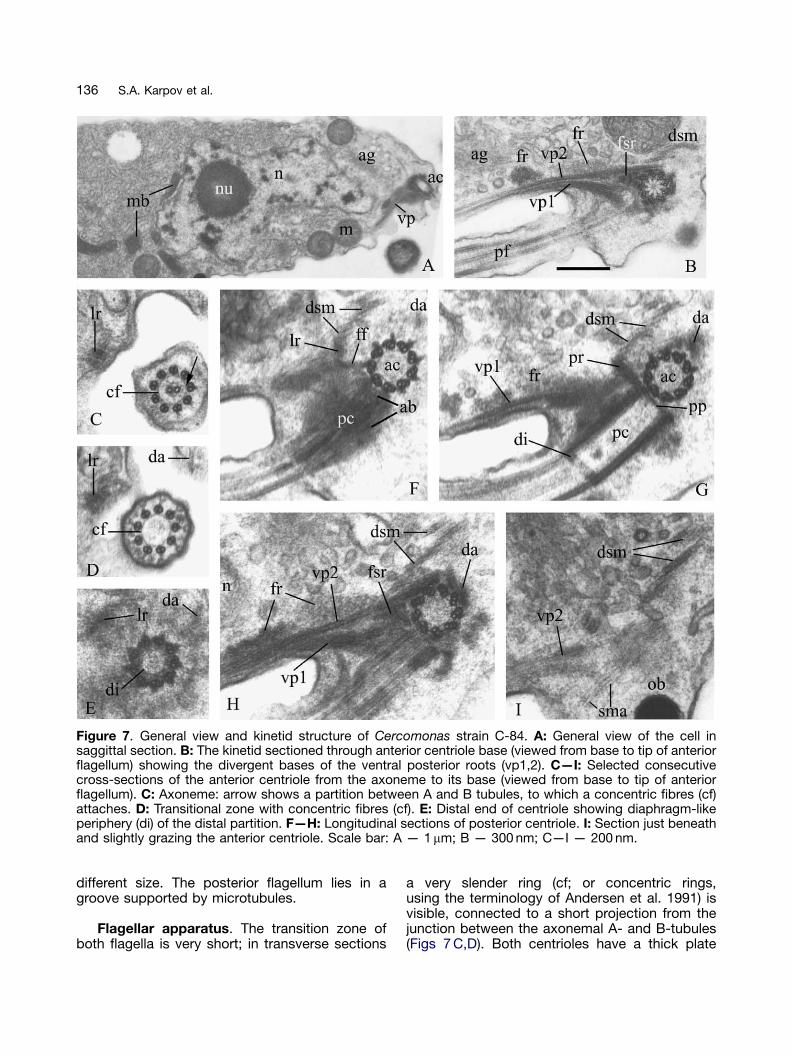
Cercomonas strain C-84 (Figs 6—8)
General ultrastructure. Both flagella emergefrom an apical tapering (Fig. 7 A). The vesicularnucleus has a large central nucleolus and promi-nent anterior projection directed towards thecentrioles. One or two Golgi dictyosomes attachto one or both sides of this nuclear projection.Mitochondria predominantly have slightly flat-tened tubular cristae, not vesicular or typicaltubular ones (Figs 7 A,B, 8 F). Microbodies arepartly applied to the nucleus and partly located inthe posterior cytoplasm. Extrusomes are rare,simple osmiophilic bodies, normally under theplasma membrane of the anterior half of the cell.There are many large membrane-bounded bodieswith central masses of dense material surroundedby numerous black dots or curved rods, whichresemble the mushroom-like bodies (MRBs) ofKatabia (Karpov et al. 2003a) but differ in lackingan internal stalk and crystal-like structure. Thecytoplasm is filled with rough endoplasmic reticu-lum (ER) cisternae, ribosomes, and vesicles of
ARTICLE IN PRESS
Figure 7. General view and kinetid structure of Cercomonas strain C-84. A: General view of the cell insaggittal section. B: The kinetid sectioned through anterior centriole base (viewed from base to tip of anteriorflagellum) showing the divergent bases of the ventral posterior roots (vp1,2). C—I: Selected consecutivecross-sections of the anterior centriole from the axoneme to its base (viewed from base to tip of anteriorflagellum). C: Axoneme: arrow shows a partition between A and B tubules, to which a concentric fibres (cf)attaches. D: Transitional zone with concentric fibres (cf). E: Distal end of centriole showing diaphragm-likeperiphery (di) of the distal partition. F—H: Longitudinal sections of posterior centriole. I: Section just beneathand slightly grazing the anterior centriole. Scale bar: A — 1mm; B — 300 nm; C—I — 200 nm.
136 S.A. Karpov et al.
different size. The posterior flagellum lies in agroove supported by microtubules.
Flagellar apparatus. The transition zone ofboth flagella is very short; in transverse sections
a very slender ring (cf; or concentric rings,using the terminology of Andersen et al. 1991) isvisible, connected to a short projection from thejunction between the axonemal A- and B-tubules(Figs 7 C,D). Both centrioles have a thick plate
ARTICLE IN PRESS
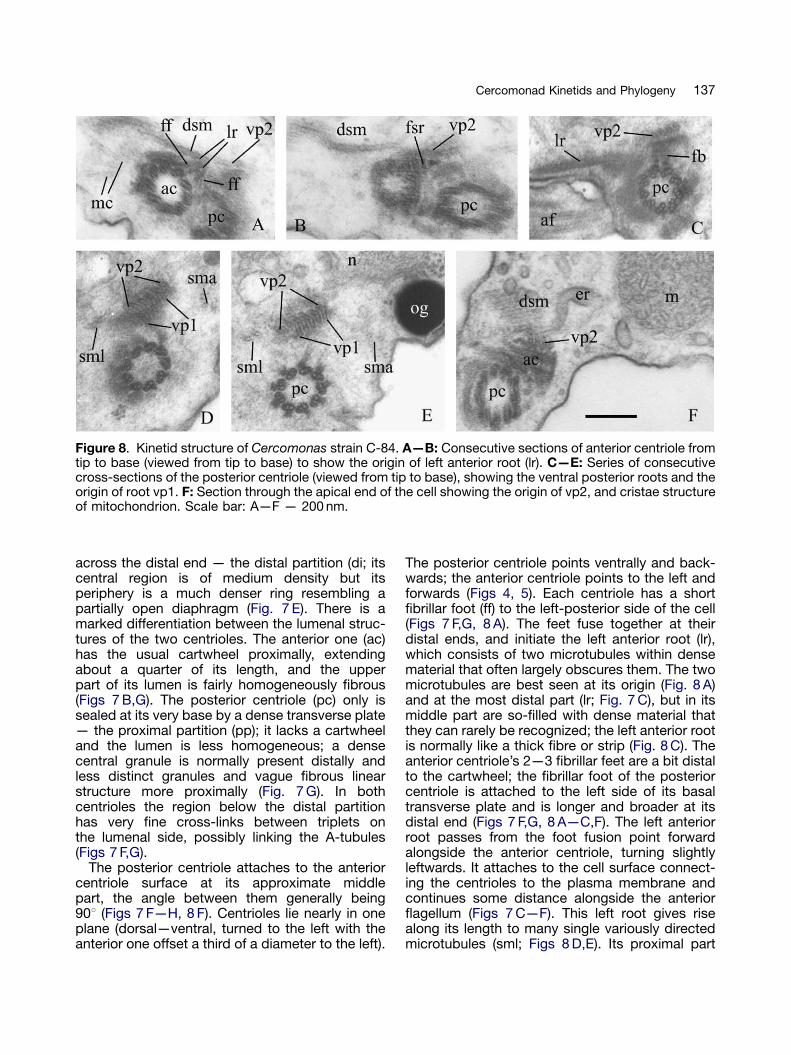
Figure 8. Kinetid structure of Cercomonas strain C-84. A—B: Consecutive sections of anterior centriole fromtip to base (viewed from tip to base) to show the origin of left anterior root (lr). C—E: Series of consecutivecross-sections of the posterior centriole (viewed from tip to base), showing the ventral posterior roots and theorigin of root vp1. F: Section through the apical end of the cell showing the origin of vp2, and cristae structureof mitochondrion. Scale bar: A—F — 200 nm.
137Cercomonad Kinetids and Phylogeny
across the distal end — the distal partition (di; itscentral region is of medium density but itsperiphery is a much denser ring resembling apartially open diaphragm (Fig. 7 E). There is amarked differentiation between the lumenal struc-tures of the two centrioles. The anterior one (ac)has the usual cartwheel proximally, extendingabout a quarter of its length, and the upperpart of its lumen is fairly homogeneously fibrous(Figs 7 B,G). The posterior centriole (pc) only issealed at its very base by a dense transverse plate— the proximal partition (pp); it lacks a cartwheeland the lumen is less homogeneous; a densecentral granule is normally present distally andless distinct granules and vague fibrous linearstructure more proximally (Fig. 7 G). In bothcentrioles the region below the distal partitionhas very fine cross-links between triplets onthe lumenal side, possibly linking the A-tubules(Figs 7 F,G).
The posterior centriole attaches to the anteriorcentriole surface at its approximate middlepart, the angle between them generally being901 (Figs 7 F—H, 8 F). Centrioles lie nearly in oneplane (dorsal—ventral, turned to the left with theanterior one offset a third of a diameter to the left).
The posterior centriole points ventrally and back-wards; the anterior centriole points to the left andforwards (Figs 4, 5). Each centriole has a shortfibrillar foot (ff) to the left-posterior side of the cell(Figs 7 F,G, 8 A). The feet fuse together at theirdistal ends, and initiate the left anterior root (lr),which consists of two microtubules within densematerial that often largely obscures them. The twomicrotubules are best seen at its origin (Fig. 8 A)and at the most distal part (lr; Fig. 7 C), but in itsmiddle part are so-filled with dense material thatthey can rarely be recognized; the left anterior rootis normally like a thick fibre or strip (Fig. 8 C). Theanterior centriole’s 2—3 fibrillar feet are a bit distalto the cartwheel; the fibrillar foot of the posteriorcentriole is attached to the left side of its basaltransverse plate and is longer and broader at itsdistal end (Figs 7 F,G, 8 A—C,F). The left anteriorroot passes from the foot fusion point forwardalongside the anterior centriole, turning slightlyleftwards. It attaches to the cell surface connect-ing the centrioles to the plasma membrane andcontinues some distance alongside the anteriorflagellum (Figs 7 C—F). This left root gives risealong its length to many single variously directedmicrotubules (sml; Figs 8 D,E). Its proximal part

ARTICLE IN PRESS
138 S.A. Karpov et al.
produces a band of 6—9 secondary microtubules(dsm) passing dorsal and backwards deep into thecell (Figs 7 F—I, 8 A,B,F). A short, striated,ventrally directed spur is near the proximal endof the anterior centriole, posterior to the left rootorigin (fsr; Figs 7 B,H, Fig 8B).
The short dorsal anterior root (da) of twomicrotubules nucleates from the right of anteriorcentriole (Figs 7 D—H), opposite the posteriorcentriole, and passes forward and slightly tothe left. It attaches to the plasma membrane,where it seems to end, and may producesecondary single microtubules passing back-wards into the cell — some pass from the baseof this root in the nuclear direction as a micro-tubular cone (mc, Fig. 8 A).
One major ventral root (VP2), initially of threemicrotubules, originates from the left dorsal sideof the anterior centriole, connecting to a thindense fibrillar sheet on the dorsal side of theanterior centriole (Figs 6, 7 B,H, 8). The micro-tubules in VP2 increase to six. An ER cisternausually applies to root VP2 (Fig. 8 F).
The second major ventral posterior root (VP1) offive microtubules starts from fibrillar materialaround the posterior left surface of the posteriorcentriole, passing backwards alongside it and rootVP2 (Figs 8 D,E). At its origin VP1 also connectswith a dense plate attached to the posteriorcentriole by fibrillar bridges (fb; Fig. 8 C). RootVP1 sharply turns from being perpendicular to theaxis of the anterior centriole and roughly 45degrees to the axis of the posterior centriole atits origin to parallel to it, then follows the posteriorcentriole and flagellum, between the plasmamembrane and VP2 (Figs 7 B,G—I, 8 D,E). VP1microtubules may increase to 7. At the level of theposterior flagellum/centriole junction both ventralroots split into two branches underlying theplasma membrane of the flagellar groove. RootVP2 is leftward of VP1. In some sections we foundthese VP microtubules at the middle and even inthe posterior part of the cell.
A more distinct posterior pair of secondarymicrotubules passes from the anterior centriolealong the posterior centriole in the same directionas, but well to the left of, the ventral posteriorroots (sml; Figs 8 D,E). A broad indistinct fibrillarroot of amorphous material (fr; Figs 7 B,G,H)passes from the left-posterior side of the anteriorcentriole and splits into two branches: a broadone directed to the nucleus and a narrow,dense branch alongside VP2. The nuclear projec-tion ends very near the VP1/VP2 association(Fig. 7 H), where the dense branch ends.
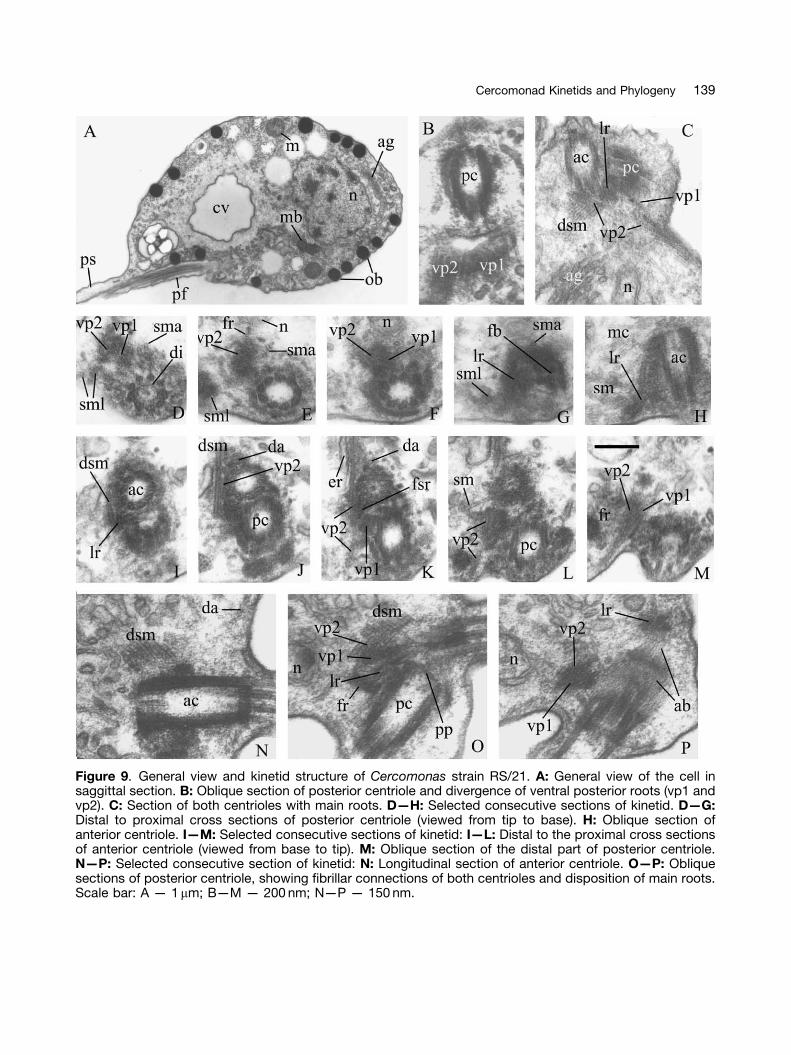
Cercomonas strain RS/21 (Fig. 9)
The structure and disposition of organelles,including the flagellar apparatus, are very similarto those of strain C-84 (Figs 9 A,O,P), but mush-room-like bodies were not found. Transition zonesof both flagella have concentric rings; eachcentriole has a thick distal partition with dia-phragm (Figs 9 D,N,P). A cartwheel is in theproximal quarter of the anterior centriole only(Figs 9 H,N,O). The proximal partition of theposterior centriole has a less prominent projectionthan in C-84 (Fig. 9 O). The posterior centrioleattaches to the anterior centriole middle; theangle between them is much more than 901(Figs 9 N—P), not 901 as in C-84. Their intercon-necting fibrillar bridges and origin and structure ofthe microtubular roots, short striated spur, and leftfibrillar feet producing the left anterior root are asin C-84 (Figs 9 B—P). The thick amorphous fibrillarroot of at least two branches passes from theanterior centriole very close to the nuclear projec-tion and VP2/VP1 association (Figs 9 E,F,L,M,O,P).The microtubular cone is associated with theproximal part of the anterior centriole and itsdorsal anterior root (Fig. 9 H). An ER cisternaapplies to root VP2 (Fig. 9 K).
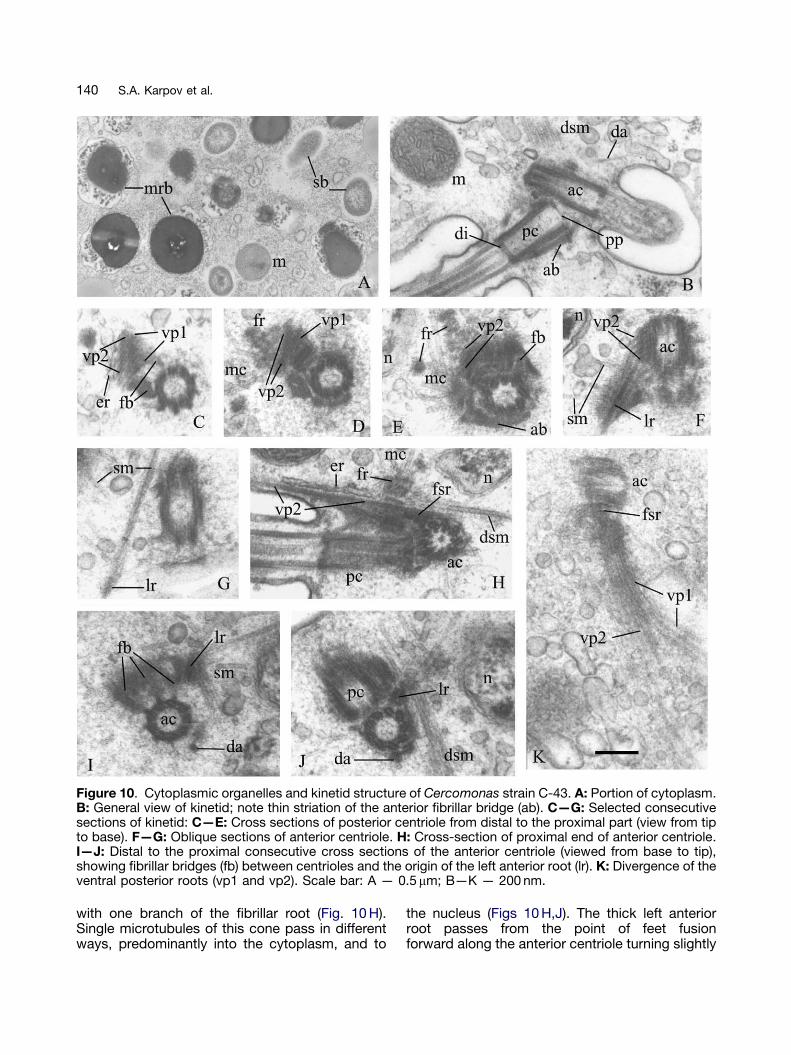
Cercomonas strain C-43 (Fig. 10)
General cell structure is as in the previous strains(Fig. 10 A,B), with MRBs as in C-84 (Fig. 10 A). Themain difference is the many symbiotic Gram-negative bacteria free in the cytoplasm withoutsurrounding membranes (Fig. 10 A). The posteriorflagellum is in a groove or channel supported bymicrotubules of roots VP2 and VP1 (Fig. 10 K).Flagella and centrioles are as in C-84 and RS/21.The angle between the centrioles is about 1201(Fig. 10 B). In some cells centriole orientationdiffers from that described above: the anteriorcentriole points to the left and the posteriorcentriole backwards, and the plane of bothcentrioles is parallel to the ventral side of the cell.The anterior connective between the centrioleshas thin striations (Fig. 10 B); conceivably it is acontractor to change the angle between thecentrioles. All roots are as in C-84 and RS/21(Figs 10 C—K), including the ER cisterna on VP2.The nuclear projection is much closer to thecentrioles and two fibrillar root branches directedto its surface are visible in Figures 10 E and 10 H.The microtubular cone is in different position(Figs 10 D,E,H): associated with fibrillar materialaround the posterior centriole (Figs 10 D,E), and
ARTICLE IN PRESS
Figure 9. General view and kinetid structure of Cercomonas strain RS/21. A: General view of the cell insaggittal section. B: Oblique section of posterior centriole and divergence of ventral posterior roots (vp1 andvp2). C: Section of both centrioles with main roots. D—H: Selected consecutive sections of kinetid. D—G:Distal to proximal cross sections of posterior centriole (viewed from tip to base). H: Oblique section ofanterior centriole. I—M: Selected consecutive sections of kinetid: I—L: Distal to the proximal cross sectionsof anterior centriole (viewed from base to tip). M: Oblique section of the distal part of posterior centriole.N—P: Selected consecutive section of kinetid: N: Longitudinal section of anterior centriole. O—P: Obliquesections of posterior centriole, showing fibrillar connections of both centrioles and disposition of main roots.Scale bar: A — 1mm; B—M — 200 nm; N—P — 150 nm.
139Cercomonad Kinetids and Phylogeny
ARTICLE IN PRESS
Figure 10. Cytoplasmic organelles and kinetid structure of Cercomonas strain C-43. A: Portion of cytoplasm.B: General view of kinetid; note thin striation of the anterior fibrillar bridge (ab). C—G: Selected consecutivesections of kinetid: C—E: Cross sections of posterior centriole from distal to the proximal part (view from tipto base). F—G: Oblique sections of anterior centriole. H: Cross-section of proximal end of anterior centriole.I—J: Distal to the proximal consecutive cross sections of the anterior centriole (viewed from base to tip),showing fibrillar bridges (fb) between centrioles and the origin of the left anterior root (lr). K: Divergence of theventral posterior roots (vp1 and vp2). Scale bar: A — 0.5 mm; B—K — 200 nm.
140 S.A. Karpov et al.
with one branch of the fibrillar root (Fig. 10 H).Single microtubules of this cone pass in differentways, predominantly into the cytoplasm, and to
the nucleus (Figs 10 H,J). The thick left anteriorroot passes from the point of feet fusionforward along the anterior centriole turning slightly

ARTICLE IN PRESS
Figure 11. Transverse section of the cell of Cerco-monas strain C-59. Scale bar: 1 mm.
141Cercomonad Kinetids and Phylogeny
leftwards (lr; Figs 10 F,G,I,J). It attaches to the cellsurface, connecting centrioles to the plasmamembrane (Fig. 10 G), and nucleates many var-iously oriented single microtubules (Figs 10 B,F,J)and a band of six secondary microtubules. Adorsal anterior root of 1—2 microtubules passesfrom the dorsal part of the sheet forward (da;Figs 10 B,I,J). Roots VP2 of three microtubulesand VP1 of 4-5 microtubules are as in otherclade A strains. They split into a left and rightpart, which pass alongside the posterior flagellum(Figs 10 C—E, H, K,).
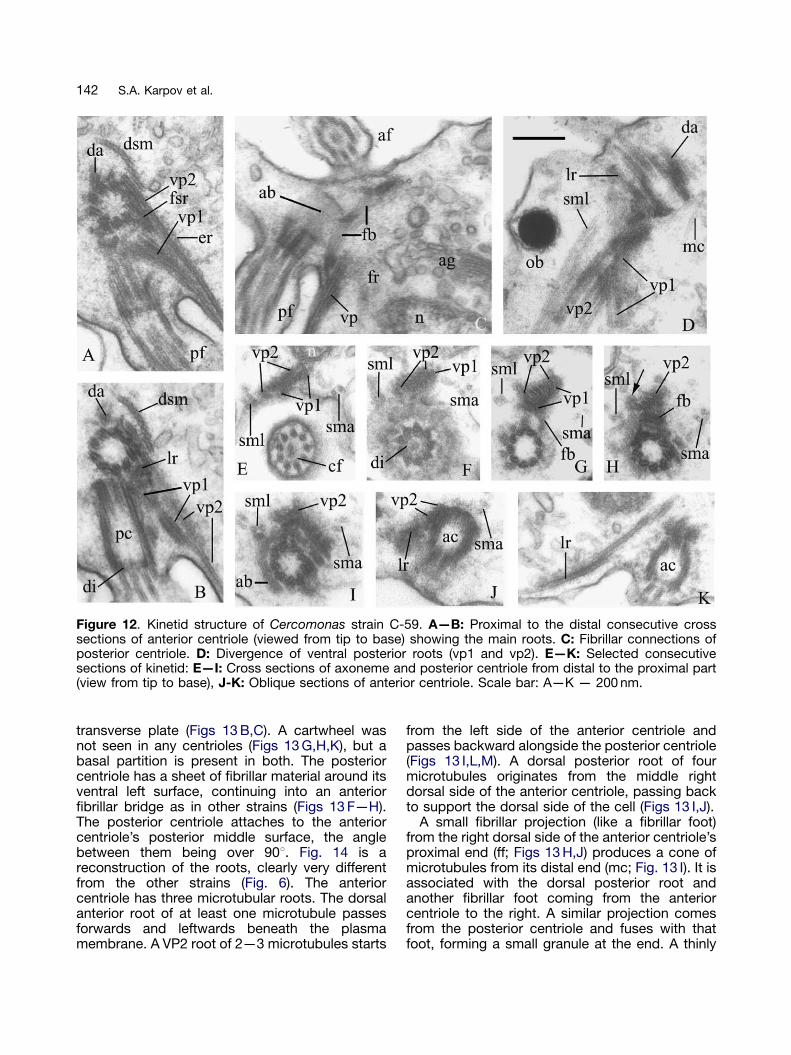
Cercomonas strain C-59 (Figs 11, 12)
Its general ultrastructure is most similar to C-84(Figs 11,12 C). The posterior flagellum lies in agroove. Mushroom-like bodies are rare, but verybig, sometimes half the nuclear dimension. Cen-trioles are joined at an obtuse angle. Transitionzone and centrioles of both flagella are as in C-84(Figs 12 A,B,E,F), as is the flagellar root system(Fig. 12) except that the foot of the posteriorcentriole is not as developed as in C-84—morelike that of C-43 and RS/21 (Figs 12 B—D). The
fibrillar sheet around the posterior centriole is clearin Figures 12 C and I, which show its structure andconnection with microtubular roots. The leftanterior and ventral roots VP2 and VP1 have thesame position and microtubule number as inpreviously described strains (Figs 12 A,B,D,J,K).Both ventral posterior roots are accompanied bytwo strands of two microtubules originating fromthe left anterior root and base of the anteriorcentriole respectively (Figs 12 A,E—J). After fol-lowing the posterior centriole and then its flagel-lum, VP2 and VP1 diverge to the left and right, andtogether with secondary microtubules from the leftanterior root and anterior centriole form twoseparate bands underlying the flagellar groovemembrane (Fig. 12 D). The number of microtu-bules may increase to six in VP2 and seven inVP1. Thus the complex posterior band mayhave 2+7+6+2 microtubules (Fig. 12 E). Theunbranched, inconspicuous, amorphous non-striated fibrillar root passes from the centriolestowards the nucleus (Fig. 12 C). Roots VP2/VP1associate with the nucleus as in C-84 (Fig. 12 E).An ER cisterna is on VP2 (Fig. 12 A). Themicrotubular cone is not well developed; itsmicrotubules associate with the anterior centriole(Fig. 12 D).
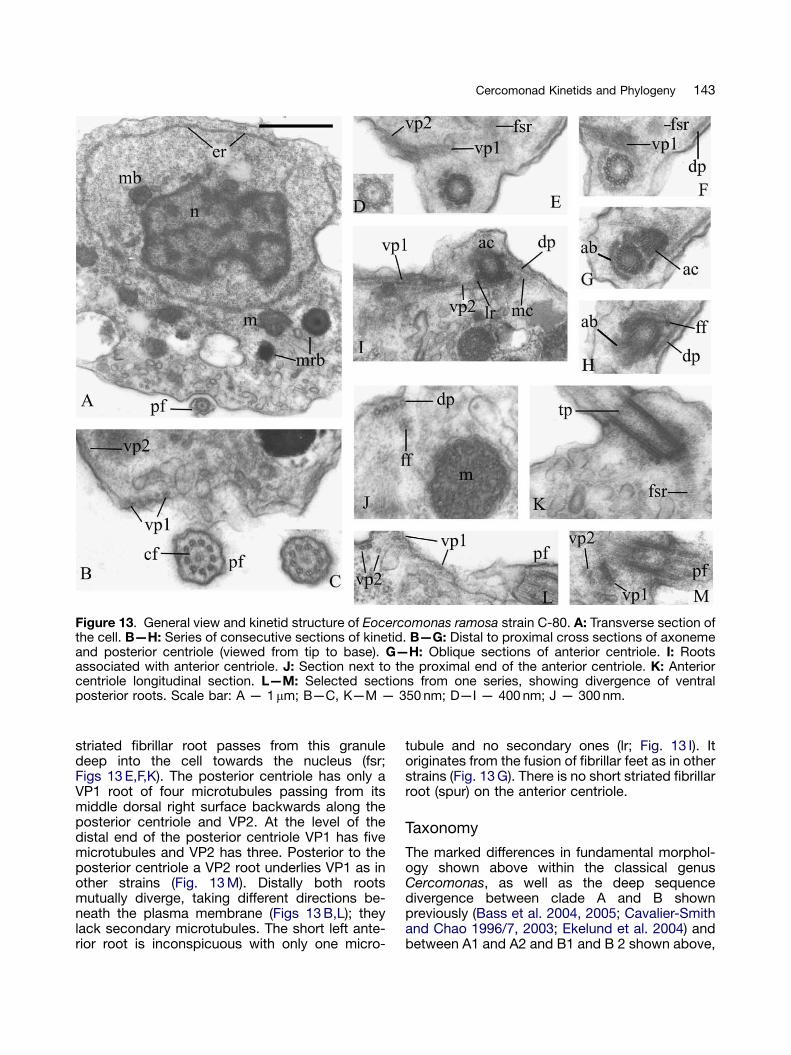
Eocercomonas ramosa: strain C-80(Figs 13, 14)
This strain differs from all the others in generalmorphology and flagellar apparatus structure. Thenucleus contains prominent peripheral and muchinterstitial heterochromatin (Fig. 13 A). Neither anucleolus nor a nuclear projection towards thecentrioles is evident. A large Golgi dictyosome isapplied to the anterior ventral part of the nucleus.Mitochondria predominantly have tubular cristae,some branched (Figs 13 A,J). Microbodies are notprominent, but near the nucleus. No extrusomeswere found. Mushroom-like bodies are scatteredthrough the cytoplasm. Its most peculiar feature isa prominent rough ER cisterna around the nucleusand part of the surrounding cytoplasm (Fig. 13 A).The posterior flagellum lies in a groove supportedby microtubules.
The transition zone of both flagella is short. Avery thin transverse plate is at the distal end of thecentriole, level with the transitional fibres con-necting it to the plasma membrane (Figs 13 K,M).In marked contrast to all clade A1 strains there isno thick distal partition with obvious peripheraldiaphragm. A concentric fibre or ring is above the
ARTICLE IN PRESS
Figure 12. Kinetid structure of Cercomonas strain C-59. A—B: Proximal to the distal consecutive crosssections of anterior centriole (viewed from tip to base) showing the main roots. C: Fibrillar connections ofposterior centriole. D: Divergence of ventral posterior roots (vp1 and vp2). E—K: Selected consecutivesections of kinetid: E—I: Cross sections of axoneme and posterior centriole from distal to the proximal part(view from tip to base), J-K: Oblique sections of anterior centriole. Scale bar: A—K — 200 nm.
142 S.A. Karpov et al.
transverse plate (Figs 13 B,C). A cartwheel wasnot seen in any centrioles (Figs 13 G,H,K), but abasal partition is present in both. The posteriorcentriole has a sheet of fibrillar material around itsventral left surface, continuing into an anteriorfibrillar bridge as in other strains (Figs 13 F—H).The posterior centriole attaches to the anteriorcentriole’s posterior middle surface, the anglebetween them being over 901. Fig. 14 is areconstruction of the roots, clearly very differentfrom the other strains (Fig. 6). The anteriorcentriole has three microtubular roots. The dorsalanterior root of at least one microtubule passesforwards and leftwards beneath the plasmamembrane. A VP2 root of 2—3 microtubules starts
from the left side of the anterior centriole andpasses backward alongside the posterior centriole(Figs 13 I,L,M). A dorsal posterior root of fourmicrotubules originates from the middle rightdorsal side of the anterior centriole, passing backto support the dorsal side of the cell (Figs 13 I,J).
A small fibrillar projection (like a fibrillar foot)from the right dorsal side of the anterior centriole’sproximal end (ff; Figs 13 H,J) produces a cone ofmicrotubules from its distal end (mc; Fig. 13 I). It isassociated with the dorsal posterior root andanother fibrillar foot coming from the anteriorcentriole to the right. A similar projection comesfrom the posterior centriole and fuses with thatfoot, forming a small granule at the end. A thinly
ARTICLE IN PRESS
Figure 13. General view and kinetid structure of Eocercomonas ramosa strain C-80. A: Transverse section ofthe cell. B—H: Series of consecutive sections of kinetid. B—G: Distal to proximal cross sections of axonemeand posterior centriole (viewed from tip to base). G—H: Oblique sections of anterior centriole. I: Rootsassociated with anterior centriole. J: Section next to the proximal end of the anterior centriole. K: Anteriorcentriole longitudinal section. L—M: Selected sections from one series, showing divergence of ventralposterior roots. Scale bar: A — 1mm; B—C, K—M — 350 nm; D—I — 400 nm; J — 300 nm.
143Cercomonad Kinetids and Phylogeny
striated fibrillar root passes from this granuledeep into the cell towards the nucleus (fsr;Figs 13 E,F,K). The posterior centriole has only aVP1 root of four microtubules passing from itsmiddle dorsal right surface backwards along theposterior centriole and VP2. At the level of thedistal end of the posterior centriole VP1 has fivemicrotubules and VP2 has three. Posterior to theposterior centriole a VP2 root underlies VP1 as inother strains (Fig. 13 M). Distally both rootsmutually diverge, taking different directions be-neath the plasma membrane (Figs 13 B,L); theylack secondary microtubules. The short left ante-rior root is inconspicuous with only one micro-
tubule and no secondary ones (lr; Fig. 13 I). Itoriginates from the fusion of fibrillar feet as in otherstrains (Fig. 13 G). There is no short striated fibrillarroot (spur) on the anterior centriole.
Taxonomy
The marked differences in fundamental morphol-ogy shown above within the classical genusCercomonas, as well as the deep sequencedivergence between clade A and B shownpreviously (Bass et al. 2004, 2005; Cavalier-Smithand Chao 1996/7, 2003; Ekelund et al. 2004) andbetween A1 and A2 and B1 and B 2 shown above,

ARTICLE IN PRESS
Figure 14. Reconstruction of kinetid structure ofEocercomonas ramosa strain C-80 (clade A2).Viewed from above (dorsal side) but from the right(i.e. at an angle intermediate between Figs 5 A and B;for abbreviations see Fig. 5 legend).
144 S.A. Karpov et al.
make it highly desirable to subdivide this undulybroad genus into several separate genera. But thiscannot be done without determining to whichclade the type species C. longicauda (thusdesignated by Fromentel (1874), according toEkelund et al. 2004) belongs, since only that cladecan retain the name Cercomonas. Thus noprogress can be made in revising the high levelclassification of Cercomonadidae without identify-ing a strain that is Cercomonas longicaudaDujardin. This is less easy than Ekelund et al.(2004) assumed, as almost every author sinceDujardin (1841) has had a different idea of whatC. longicauda looks like. For example C. long-icauda of Lemmermann (1914; fig. 59, p. 48, asCercobodo longicauda for which he gave theauthority (Klebs) Senn contrasting with (Stein)Senn in the text!) is so dramatically different fromDujardin’s description and figure that it cannot bethe same species. Fig. 4 L reproduces Dujardin’sfigure (scanned digitally and unchanged exceptfor contrast enhancement). The anterior flagellumis about 3.8� the body length and the clearlyvisible posterior flagellum projects a full bodylength behind the cell; hence the name long-icauda. Furthermore the flagellum adheres to thecell body right up to its extended posterior tip.None of these three characters, the only distin-guishing features of Dujardin’s longicauda apartfrom cell size, is true of Lemmermann’s figure norof either the SCCAP strain (see fig. 5 of Ekelundet al. 2004, where the elongated posterior of thecell and the flagellum are clearly distinct and the
posterior flagellum does not project beyond it) orthe ATCC strain (see Fig. 4 J—K). All three strainshave an anterior flagellum about equal to the body,not 3.8� its length as in Dujardin’s figure (or3—4� , implied by his description). Thus all threewere certainly misidentified as longicauda. Noneof them has the projecting posterior flagellum thatobviously caused Dujardin to confer that name inparticular. Translated into English, Dujardin’s com-plete diagnosis of C. longicauda is:
Flexible spindle-shaped body, terminated atthe rear by a very thin flexuous filament in theform of a tail. Body length 8—9mm. Tail [i.e.the projecting part of the posterior flagellum]15mm. Flagelliform filament [i.e. anteriorflagellum] very thin, 30—40mm.
Ekelund et al. (2004) did not explain why theyidentified their strain as longicauda, and made nocomparison with Dujardin’s original description,nor cited it. The only previous description theyreferred to was by Sandon (1927, p. 72), who gavea body length of 5—10mm, and 3—4� bodylength for the anterior flagellum (a marked dis-crepancy with the SCCAP strain; Ekelund et al.2004) and ‘slightly longer than body length’ for theposterior flagellum. Although the SCCAP, ATCC,and Lemmermann (1914) strains are mutuallysimilar in flagella/cell proportions and in theprotoplasmic tail often obscuring the no longerposterior flagellum, they all differ in size from eachother and Dujardin’s. Lemmermann’s strain is thelargest (18—36mm), SCCAP the smallest4—10mm, and ATCC intermediate (10—12mm).
Ekelund et al. (2004) designated their SCCAPC1 as ‘neotype’ for C. longicauda, even thoughit differed in every respect except size fromC. longicauda Dujardin, 1841. They also did notexpressly explain why neotypification was neces-sary or explicitly meet all seven of the conditionsthat the International Code of Zoological Nomen-clature (ICZN) requires for designation of aneotype. Thus SCCAP C1 is invalid as a neotypefor C. longicauda. As only a valid neotypificationhas authority, this still remains to be done.Because of the exceptional importance of stabiliz-ing the application of the name Cercomonasbefore the taxonomy of Cercomonadidae can berevised, it is necessary to designate a validneotype for C. longicauda here.
In recent decades four different clonalstrains have been contradictorily identified asC. longicauda. Both of those that belong in cladeB1 (SCCAP C1 and ATCC 50344) were definitely

ARTICLE IN PRESS
145Cercomonad Kinetids and Phylogeny
misidentified. Strain C-1 (now dead) of Mylnikov(1987), with ultrastructural features suggestive ofclade A1 not clade B — see discussion, wasprobably also misidentified, as C. longicaudasensu Mylnikov (2000) was 18—36mm — far largerthan Dujardin’s; its large size and ultrastructuretogether suggest that it belongs to clade A1b (seeabove), but it was distinct from all three A1bstrains in Figure 3. CCAP 1910/2 longicauda, lessobviously misidentified, is in clade A1a. The verysame excessive dimensions (18—36mm) weregiven by Klebs (1892), whose drawing differedgreatly from Dujardin’s, and by Zhukov (1993) withdrawings of longicauda (after Skuja, but essen-tially the same as Klebs) also nothing likeDujardin’s, but not unlike C. ekelundi; Kent’s(1880—2) drawings differ in showing a distinctposterior flagellum, but appear to have beencopied from those of Stein (1878), which also areprobably not of the same organism as Dujardin’s.The use of conflicting secondary sources foridentification has led to many different organismsbeing wrongly identified as longicauda and cras-sicauda (the only two species in Stein’s treatisethat can be accepted as Cercomonas). We heartilyconcur with the comments that ‘more than onespecies may be included’ among the numerousrecords of this ‘species’ (Al-Qassab et al. 2002),and that most (if not all) are ‘highly questionable’(Foissner 1991). The fact that four such differentcultures of cercomonads, belonging in threeradically different parts of the tree, have beenidentified as longicauda (three certainly incor-rectly), and the numerous discordant drawingsand descriptions in the literature, emphasize theurgent necessity to clarify and standardize thetaxonomic status of C. longicauda. This is bestdone by designating a neotype much closer toDujardin’s original description than is SCCAP C1,even though certainty that it is the same species isunattainable.
Neotypification of Cercomonaslongicauda Dujardin 1841
Conditions of article 75.3 of ICZN for validtypification are met thus:
1.
We designate the CCAP 1910/2 culture as thehapantotype and neotype for C. longicaudaDujardin 1841 for the express purpose ofclarifying the taxonomic status of that species.2.
Cercomonas longicauda can be distinguishedfrom all other species by its 18S rRNAsequence. Any strains differing by 3 nucleo-tides or more are to be regarded as differentspecies (except for any that might in future beshown to be sexual and able to interbreedfreely with it).Description of the neotype strain: Cell�8—10mm long when gliding, often amoeboid.Its posterior flagellum adheres strongly to itsbody and extends about a body length (i.e. itstotal length is 15—20mm) beyond its posteriortip and is thus always clearly visible duringgliding motion, when its body is spindle-shaped(Figs 4 U,V). Its anterior flagellum is about twiceits body length (usually 15—17 mm). Cysts arespherical and smooth walled. Contractilevacuole. Pseudopodia broad, flat and lobed(Figs 4 W, X). Nucleus anterior. Cells sometimesclump together (Fig. 4 Y). Plasmodial stagecompact and rounded, not a string of almostseparate cells (Fig. 4 Z).
3.
The neotype is strain CCAP 1910/2 isolated byZolffel in 1991, and could also be recognized byits 18S rRNA sequence together with a lightmicroscope appearance as described aboveand shown in Figures 4 U—Z, even if it weremislabelled in future. Thus recognition of theneotype designated is assured.4.
There is no reason to think that Dujardin evermade any type specimens of any of theprotozoa that he described. Thus no holotypeever existed. Designation of a neotype byEkelund et al. (2004) is invalid, as the desig-nated strain was misidentified and definitely notlongicauda as described by Dujardin (1841)(see above).5.
Half the 22 strains studied here (all in Fig. 3except plasmodialis, C-43 and C-85, plus long-icauda in Fig. 4 U—Z) are substantially more likeDujardin’s description than is SCCAP C1.Several are totally indistinguishable from Dujar-din’s in body form and size (e.g. CeS-2, C-72,NZ1, longicauda CCAP1910/2). However, all differin having somewhat shorter anterior flagella andsometimes also somewhat shorter posterior fla-gella. This makes it possible that none of them isreally precisely the same as Dujardin’s. However,this leaves out of account the probability ofvariation in flagellar lengths within a clone andthe fact that Dujardin’s description was not basedon clonal cultures anyway, and we cannot knowhow representative the individual cell that wasfigured was. We point out, for example, that theindividual cell of C-80 in Figure 4 A (photographedin the Borok lab) has an anterior flagellum over

ARTICLE IN PRESS
146 S.A. Karpov et al.
three times its body length, yet most gliding cellsof this strain growing in the Oxford laboratory haveanterior flagella less than twice the body length, asdo several several cells in the plasmodial phasesdepicted in Figure 4 D. Thus, even within a clonalCercomonas culture, there can be discrepanciesin anterior flagellum length as great as thedifferences seen between several clade A1 cul-tures and Dujardin’s. Thus, for most of thesecultures it could not be argued with confidencethat they are not the same species as Dujardin’s.Thus several of them could reasonably be chosenas a neotype. Conversely, however, the fact thatso many clade A1 cultures are almost indistin-guishable except for anterior flagellum length fromDujardin’s longicauda, yet have very differentrRNA sequences and must virtually all be differentspecies from each other and from his, makesselection of a specific one essentially arbitrary. Weshall never know what was the rRNA sequence ofDujardin’s strain, nor whether the strain heobserved had plasmodial phases and cysts ornot. Yet the need to chose a neotype and stabilisenomenclature is so great that one must bechosen, even though their morphological similaritynecessarily gives inadequate morphological basisfor doing so. We have over a hundred otherCercomonas cultures in the Oxford laboratory,nearly half of which would be almost as suitable,but none of which ordinarily have anterior flagella3—4 times body length.
We therefore designate CCAP 1910/2 as neo-type not only because (apart from its usuallysomewhat shorter anterior flagellum) it is indis-tinguishable from the original description, but alsobecause it is quite closely related to RS/21, whichwe have now well characterised ultrastructurally,and RS/22, both of which have been used forcharacterising protein genes as well as rRNA(Archibald et al. 2003; Bass et al. 2005; Cavalier-Smith and Chao 1996/7; Keeling 2001). However,CCAP 1910/2 is preferable to either RS/21 or 22as it is already available from CCAP and hassurvived there for 14 years of subculturing after itswas isolated and identified as longicauda by anindependent Cercomonas expert, Zolffel. Thelatter is important as ICZN also recommends(article 75B) that expert opinion be sought to verify‘that the proposed designation does not arouseserious objection from other specialists in thegroup in question’. Accordingly we also consultedK. Vickerman (Glasgow), who agrees that thisstrain would be an acceptable neotype and thatSCCAP C-1 from Danish agricultural soil, and theATCC strain (a marine strain, unlike Dujardin’s,
with distinctly non-adherent posterior flagellum)especially, were both misidentified.
The species C. longicauda was originally de-scribed from three infusions of licorice, gum orpotato in Rennes, France (Dujardin 1841; p. 290,Plate 4:15) and thus likely to have come from soilor freshwater; the neotype was isolated by Zolffelin 1991 in England from freshwater (http://www.ccap.ac.uk). Zooflagellate morphospeciesare widely thought to be cosmopolitan (Finlay2002; Finlay and Clarke 1999). Although this neednot be the case for species defined by morediscriminating molecular methods (Cavalier-Smithand Chao 2006; von der Heyden and Cavalier-Smith 2005), for at least a few Cercomonas strainswe have been able to recover precisely the sameCercomonas 18S rRNA sequence from sampleson different continents as far apart as England,Panama and New Zealand, so there is currently nosound reason to think that designating a neotypefrom a different locality matters in any way or thatignorance of the precise location of either theneotype or original is relevant to their identity.
The neotype culture is available from CCAP(http://www.ccap.ac.uk).
Cercomonas Dujardin 1841 emend.Karpov, Bass, Mylnikov, and Cavalier-Smith (non-Cercomonas emend. Ekelundet al. 2004), revised diagnosis
Gliding, typically spindle-shaped cercomonadswith the posterior flagellum trailing, usually ex-tending behind the cell body. Broad, flat lobedpseudopods or filopodia often formed, usuallyunbranched. A prominent nuclear extension direc-ted to the nearby centrioles. Tubular mitochondrialcristae usually slightly flattened. The anteriorcentriole usually has a proximal cartwheelbut the posterior one does not. Both centrioleshave a thick distal partition with a more prominentdiaphragm-like outer part. The anterior centriolehas a fibrillar striated spur. Posterior centrioleonly has a proximal transverse plate. Extru-somes are nearly isodiametric osmiophilicbodies. Some species have a complex life cycleincluding multinuclear plasmodia and sphericalcysts (smooth-walled or rugose). With threediagnostic signature sequences in 18S rRNA:CATT, GAGGGACTATCGGTCGATTTA (N1 of Eke-lund et al. 2004), and GGACT (see Table 1). N2 ofEkelund et al. (2004) is not diagnostic for clade A1on our rRNA trees. Type species C. longicauda

ARTICLE IN PRESS
147Cercomonad Kinetids and Phylogeny
Dujardin 1841, with neotype culture CCAP 1910/2and type sequence: DQ442884.
Cercomonas jutlandica (Ekelund et al.)comb. nov. Cavalier-Smith and Bass
Basionym Neocercomonas jutlandica Ekelund et al.(2004 p. 129). As the type species of Cercomonas(C. longicauda Dujardin) falls within the originaldefinition of Neocercomonas, Neocercomonas isnow a junior synonym of Cercomonas. But itremains available as a potential generic or sub-generic name for a differently defined restrictedsubset of clade A1 excluding longicauda butincluding jutlandica (e.g. clade A1b), should it beneeded.
Eocercomonas gen. nov. Karpov, Bass,Mylnikov, and Cavalier-Smith; Diagnosis
Gliding, typically spindle-shaped cercomonadswith the posterior flagellum trailing as in Cercomo-nas and Paracercomonas. Eocercomonas dif-fers from both these genera in having a dorsalposterior microtubule band and nuclear fibrillarstriated root. It differs from Cercomonas in lackinga nuclear extension to the centrioles, and inlacking cartwheels in both centrioles and havinga proximal transverse plate in the anterior as wellas the posterior centriole, thin distal centriolarpartition (without peripheral diaphragm), and along fibrillar striated root passing from the poster-ior centriole towards the nucleus, and mito-chondrial cristae are rounded tubules. Withthree diagnostic sequence signatures in 18SrRNA: TAATT, TCGAGCTTTACAACCTTGACT, andCTTCCTGTTCTATTTGTTGGTTTCTAGGATCGG (seeTable 1). Type species E. ramosa sp. nov. Corre-sponds with clade A2 on our rRNA trees. Etym. Eo-Gk. dawn, Cerco — Gk tail; monas Gk unit, becauseit is a tailed monad that is sister to Cercomonassensu stricto and diverged from it before anyCercomonas species separated from each other. Afeminine noun, like Cercomonas Dujardin.
Eocercomonas ramosa sp. nov. Karpov,Bass, Mylnikov, and Cavalier-Smith;Diagnosis
Gliding cells spindle-shaped, 5—15 mm long.Proximal part of anterior flagellum surrounded bya cytoplasmic sheath. Anterior flagellum usuallyover twice body length when actively gliding
(420mm). Posterior flagellum almost invisible,protruding only slightly behind cell. Contractilevacuole in the anterior part of the cell. Extrusomesabsent. Trophic cells produce highly branchedvery slender pseudopodia, and may group to-gether in consortia. Dorsal anterior root laterallyattached to anterior centriole. Cysts unknown.Etym. ramosa L. ¼ branching, because of itsbranching filopodia. Type strain C-80 isolatedfrom a freshwater lake in Antarctica by A.P.Mylnikov. Type illustrations Figures 4 A—D. Type18S rRNA sequence AY884327.
Cercomonas ( ¼ Cercobodo) cometa (Hollande)is the most similar named species but differs in itsrelatively shorter anterior flagellum and, notably,its posterior flagellum being equal to or longerthan the anterior one. We prefer to designate thewhole 18S rRNA sequence as the type becausethere is a danger that if one picks just one or twohighly variable segments that could inadvertentlyinclude several different related ribotypes.
Paracercomonas gen. nov. Cavalier-Smith and Bass; Diagnosis
Gliding, with flexible very metabolic body usuallyable to produce pseudopodia of different shapes,including branched filopodia, from any part of thecell. During movement the cell body attaches tothe substrate. Anterior flagellum makes slowrowing motion, posterior flagellum trails behindmore passively. Mitochondrial cristae roundedtubules. Some species have complex life cycleincluding multinuclear plasmodia and cysts. Dis-tinguished from Cercomonas and Eocercomonasby usually having cartwheels in both centriolesand by a diagnostic, highly conserved, andderived complementary base-pair substitutionU—A base pair at base pair 7 in helix 25, insteadof a G—C pair as in all other cercomonads (seeTable 1 and preceding analysis). Corresponds withclade B1 on our rRNA trees. Etym. Para- Gk.beside, contrary to; cerco — Gk tail; monas Gkunit, to emphasize that it is probably sister toCercomonas plus Eocercomonas, but more dra-matically different in sequence and to some extentmorphology than they are from each other. Afeminine noun, like Cercomonas Dujardin.
Paracercomonas marina sp. nov. Cavalier-Smith and Bass; Diagnosis
Cercomonad with irregular, sometimes lobed ortruncated, posterior, slightly longer than and often

ARTICLE IN PRESS
148 S.A. Karpov et al.
entirely obscuring its slightly shorter posteriorflagellum. Body length �10mm; anterior flagellum�10 mm. Unambiguously distinguishable frommorphologically similar species, e.g. the slightlylarger P. ekelundi by its 18S rRNA sequence.Plasmodial stage, contractile vacuole or cystsunrecorded. Marine. Type culture ATCC 50344;type 18S rRNA sequence AF411270; type figureFigure 4 J—K.
Paracercomonas ekelundi sp. nov. Cavalier-Smith and Bass; Diagnosis
Cercomonad with narrowly drawn out, sometimesforked, posterior, slightly longer than and oftenobscuring its similar length (or somewhat longer)posterior flagellum. Body slender, fusiform whengliding, length 10—16mm; anterior flagellum�10 mm. Unambiguously distinguishable frommorphologically similar species, e.g. the slightlysmaller P. marina by its 18S rRNA sequence.Contractile vacuole lateral; close to, usuallybehind nucleus. Soil habitat. Spherical 3—4mmcysts. Plasmodial phase unknown. Type cultureSCCAP C1; type 18S rRNA sequence AY496047;type figure figure 5 of Ekelund et al. (2004). BothP. marina and P. ekelundi are unambiguouslydistinguishable from C. longicauda by not havinga longer posterior flagellum that extends a bodylength behind the posterior tip of the cell and by itstypical non-adherence to that tip.
Paracercomonas metabolica (Mylnikov)Cavalier-Smith and Bass comb. nov.
Basionymn: Cercobodo metabolicus Mylnikov(1992). As we have shown that Mylnikov’sC. metabolicus strain Cs-9 falls robustly incercomonad clade B1b and has the B1 sequencesignature and the Paracercomonas sequencesignature defined above (Table 1, Fig. 1), it mustbe transferred from Cercomonas, where Karpovand Mylnikov (2004) placed it (as Cercomonas( ¼ Cercobodo) metabolicus; note that althoughsome later authors (Kent 1880—1882; Lee andPatterson 2000) have treated Cercomonas asmasculine, its originator Dujardin (1841) clearlytreated Cercomonas as feminine, so it shouldhave been Cercomonas metabolica; all othernames of ‘Cercomonas’ species originally de-scribed as Cercobodo (treated by Lemmermann,1914 as masculine and Hollande, 1932 asfeminine! Type Cercobodo laciniaegerens Kras-silstschik, 1886, gender unclear) but since trans-
ferred to ‘Cercomonas’ with species namesending in —us (Mylnikov and Karpov 2004) shouldsimilarly be corrected to �a.
Discussion
Ekelund et al. (2004) wrote that their ‘analysisstrongly suggest that Cercomonas is not amonophyletic group’ and gave that as the primereason for dividing Cercomonas into two genera.However, the failure of clades A and B to grouptogether on their trees and on some, but not all,previous trees (Cavalier-Smith and Chao 2003) isprobably a long-branch tree-reconstruction arti-fact caused by the more rapid evolution of cladeB. Their analysis did not strongly argue againstmonophyly. Only their parsimony trees gavemoderate support to the polyphyly or paraphylyof Cercomonas. There was no bootstrap supportfor this in their distance or maximum-likelihoodanalyses, which were technically superior byallowing for intersite variation. Holophyly of theclassical genus Cercomonas was found on amaximum-likelihood tree (Cavalier-Smith andChao 2003), on a gamma-corrected distance treewith massive taxonomic sampling (Bass andCavalier-Smith 2004), and on a taxonomicallyrather well-sampled Bayesian tree (Bass et al.2005). Although statistical support in each casewas weak, this consistency among very wellsampled trees using the theoretically best meth-ods means that there is no good reasonnot to continue to regard Cercomonadidae asholophyletic.
Division of Cercomonas into separate generamust therefore depend on their morphological andgenetic disparity, not on suggestions of polyphylyprobably based on computational artifacts. Ourphylogenetic, sequence signature, and ultrastruc-tural analyses all support treating clades A1, forwhich we reserve the name Cercomonas, andclade A2 (Eocercomonas), and B1 (Paracercomo-nas) as separate genera. Even though we unfortu-nately cannot accept the name Neocercomonas(Ekelund et al. 2004) for clade A1, as that cladeincludes the type species of Cercomonas, Neo-cercomonas could become a useful name forclade A1b, e.g. should it prove desirable in futureto establish separate subgenera (or even genera)for A1a and A1b. Our sequence and ultrastructuralstudies have thus resulted in Cercomonas beingdivided into three very distinct genera. Althoughthis reflects a large advance in evolutionaryunderstanding of the family Cercomonadidae,

ARTICLE IN PRESS
149Cercomonad Kinetids and Phylogeny
the division of Cercomonas into two genera byEkelund et al. (2004) and three by ourselves hasthe unfortunate practical consequence that cer-comonads can no longer be assigned to a genusby light microscopy alone, and we do not know towhich of the three genera most named speciesbelong — though we suspect that most will belongto Cercomonas and not require renaming. Wesuggest that the vernacular name cercomonad,corresponding to the family name Cercomonadi-dae, be used specifically to embrace all threegenera and not in a broader sense for a still largergroup of Cercozoa. We also suggest that names ofcercomonad species not yet assigned to generaare placed in inverted commas. Inability to assignnamed species to genera is not a serious scientificproblem, because one major conclusion from ourwork must be that no ecological or other observa-tions on Cercomonas can be very useful unlessthe strain being studied is placed on the rRNAtree. Sequences rather than light microscopiccharacters are the only way of adequatelyrepresenting the huge biological diversity ofthis family. New genera will also be neededafter further research for the anaerobic cercomo-nad and for the clade that includes the Tempisquestrain.
Our culturing, light microscopy and sequencinghave revealed a large number of new ‘Cercomo-nas’ strains. Except for C-56 and C-72, whichhave identical 18S rRNA sequences, the se-quences represented on Figure 1 are so differentthat there must be at least 50 separate species.Our light microscopy indicates that although someare morphologically very distinct, other super-ficially very similar strains, e.g. C-71 and C-80both resembling ‘C. cometa’, can be as geneti-cally distant as any ‘Cercomonas’. This is notsurprising since cercomonads can vary visibly inthe light microscope in fewer different ways thanthe 678 variable positions in their 18S rRNA. Apartfrom size and shape of the flagellates and cystsand the length of each flagellum, the majorvariables among strains of Cercomonadidae liein the form and position of pseudopods, thenumber, size and position of contractile vacuoles,the thickness and texture of cyst walls, and thepresence/absence of plasmodium and cysts in thelife cycle. Our demonstration that certain strains ofvery different sequence can have a similarcombination of light microscope charactersmeans that in future it will be essential to combinean rRNA sequence with light microscope mor-phology of cultures in describing new species.Very few (e.g. C. longicauda, C. alexeieffi, C-43,
and N. plasmodialis — Cs-4) of our strains can beidentified with established species; we shall nameadditional new species elsewhere. We wereagreeably surprised by the extent to which themolecular trees allow some patterns of phenoty-pic relationship of these remarkably pleomorphicamoeboflagellates to be discerned even in thelight microscope. With sufficiently extensive sam-pling it should be possible to define speciesrecognizable by a combination of morphologyand rRNA signatures.
Although we have argued that Cercomonadidaeare probably holophyletic, some recent treessuggest that this may not be true of the orderCercomonadida, currently comprising only Cerco-monadidae and Heteromitidae (Cavalier-Smithand Chao 2003). The best and most comprehen-sive trees consistently place Heteromitidae closerto Thecofilosea and/or Imbricatea than to Cerco-monadidae (Bass and Cavalier-Smith 2004; Basset al. 2005). Although this has only weak support,if it were verified by other molecular analyses thetwo families may need separate orders. Cerco-monadida were (weakly) holophyletic on only oneof four trees of Cavalier-Smith and Chao (2003)and on none of those of Ekelund et al. (2004).
Ultrastructural Contrasts betweenCercomonas (A1) and Eocercomonas (A2)
All 4 members of clade A1 (Cercomonas: C-84, C-59, RS/21 and C-43) have a general ultrastructurewith slightly flattened tubular mitochondrial cris-tae, nuclear projection to centrioles, well-devel-oped nucleolus, and extrusomes (osmiophilicbodies), whereas C-80 (clade A2: Eocercomonasramosa) has no nuclear projection, extrusomes ornucleolus, but has prominent heterochromatin,developed ER around the central part of the cellincluding the nucleus, and tubular branchingcristae. C-80 alone has branching filopodia in theamoeboid state. Centrioles also differ in these twoclades: A1 has a thick distal partition withperipheral diaphragm-like structure and a cart-wheel in the anterior centriole only; in A2 (C-80)both centrioles lack cartwheels, having a proximaltransverse plate instead, and the distal partition isvery thin with no peripheral diaphragm.
All four A1 strains have great similarity in rootstructure also (Table 2), but are rather differentfrom C-80. A1 ventral posterior microtubules areorganized as two distinct roots (VP2 and VP1), oneattached to the side of the anterior centriole andthe other embedded proximally in a dense sheet
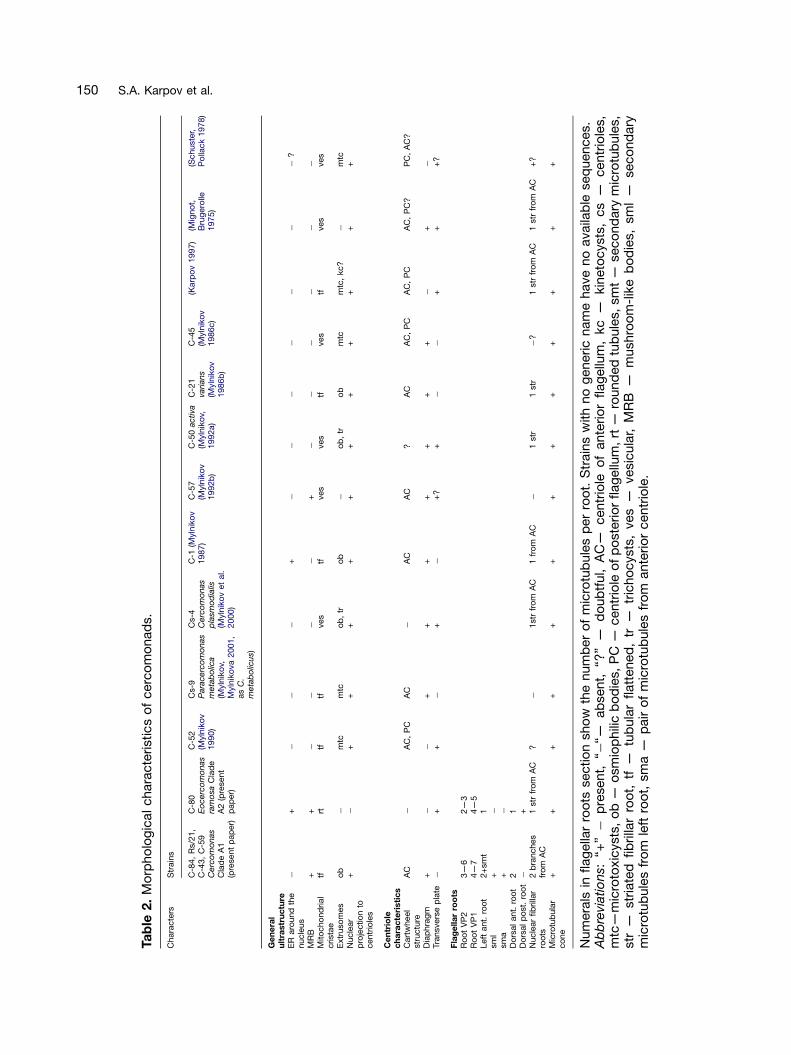
ARTICLE IN PRESS
Ta
ble
2.
Mo
rpho
log
icalchara
cte
ristics
of
cerc
om
onad
s.
Chara
cte
rsS
train
s
C-8
4,
Rs/2
1,
C-4
3,
C-5
9C
erc
om
onas
Cla
de
A1
(pre
sent
pap
er)
C-8
0E
ocerc
om
onas
ram
osa
Cla
de
A2
(pre
sent
pap
er)
C-5
2(M
yln
iko
v1990)
Cs-9
Para
cerc
om
onas
meta
bo
lica
(Myln
iko
v,M
yln
iko
va
2001,
as
C.
meta
bo
licus)
Cs-4
Cerc
om
onas
pla
smo
dia
lis(M
yln
iko
vet
al.
2000)
C-1
(Myln
iko
v1987)
C-5
7(M
yln
iko
v1992b
)
C-5
0activa
(Myln
iko
v,1992a)
C-2
1va
rians
(Myln
iko
v1986b
)
C-4
5(M
yln
iko
v1986c)
(Karp
ov
1997)
(Mig
no
t,B
rug
ero
lle1975)
(Schuste
r,P
olla
ck
1978)
Ge
ne
ral
ult
rastr
uc
ture
ER
aro
und
the
nucle
us
�+
��
�+
��
��
��
�?
MR
B+
+�
��
�+
��
��
��
Mito
cho
nd
rial
crista
etf
rttf
tfves
tfves
ves
tfves
tfves
ves
Extr
uso
mes
ob
�m
tcm
tco
b,
tro
b�
ob
,tr
ob
mtc
mtc
,kc?
�m
tcN
ucle
ar
pro
jectio
nto
centr
iole
s
+�
++
++
++
++
++
+
Ce
ntr
iole
ch
ara
cte
risti
cs
Cart
wheel
str
uctu
reA
C�
AC
,P
CA
C�
AC
AC
?A
CA
C,
PC
AC
,P
CA
C,
PC
?P
C,
AC
?
Dia
phra
gm
+�
�+
++
++
++
�+
�
Transvers
ep
late�
++
�+
�+
?+
��
++
+?
Fla
ge
lla
rro
ots
Ro
ot
VP
23—
62—
3R
oo
tV
P1
4—
74—
5Left
ant.
roo
t2+
sm
t1
sm
l+
�
sm
a+
�
Do
rsalant.
roo
t2
1D
ors
alp
ost.
roo
t�
+N
ucle
ar
fib
rilla
rro
ots
2b
ranches
fro
mA
C1
str
fro
mA
C?
�1str
fro
mA
C1
fro
mA
C�
1str
1str
�?
1str
fro
mA
C1
str
fro
mA
C+
?
Mic
rotu
bula
rco
ne
++
++
++
++
++
++
+
Num
era
lsin
flag
ella
rro
ots
sectio
nsho
wth
enum
ber
of
mic
rotu
bule
sp
er
roo
t.S
train
sw
ith
no
generic
nam
ehave
no
availa
ble
seq
uences.
Ab
bre
viatio
ns:
‘‘+’’�
pre
sent,
‘‘�‘‘—
ab
sent,
‘‘?’’
—d
oub
tful,
AC
—centr
iole
of
ante
rio
rflag
ellu
m,
kc
—kin
eto
cysts
,cs
—centr
iole
s,
mtc
—m
icro
toxic
ysts
,o
b—
osm
iop
hili
cb
od
ies,
PC
—centr
iole
of
po
ste
rio
rflag
ellu
m,
rt—
round
ed
tub
ule
s,
sm
t—
seco
nd
ary
mic
rotu
bule
s,
str
—str
iate
dfib
rilla
rro
ot,
tf—
tub
ula
rflatt
ened
,tr
—tr
icho
cysts
,ves
—vesic
ula
r,M
RB
—m
ushro
om
-lik
eb
od
ies,
sm
l—
seco
nd
ary
mic
rotu
bule
sfr
om
left
roo
t,sm
a—
pair
of
mic
rotu
bule
sfr
om
ante
rio
rcentr
iole
.
150 S.A. Karpov et al.

ARTICLE IN PRESS
151Cercomonad Kinetids and Phylogeny
adjacent to the posterior centriole. These roots arein all investigated strains, but the number ofmicrotubules is slightly different. VP2 normallystarts as a 3-microtubule root, and VP1 as a 4—5-microtubule root. In all strains additional cytoplas-mic microtubules broaden them to 5—7. Leftand right strands of two microtubules accompanyboth roots.
In C-80 the VP2 has at its origin 2—3 (in differentcells) and VP1 has 4—5 microtubules, both havingalmost no changes on their way backwards. A1cercomonads have a well-developed microtubule-nucleating left anterior root, while in A2 it is muchsimpler without secondary microtubules. At thesame time, it has an additional dorsal posteriorroot, absent in A1. C-80 has no striated spur fromthe anterior centriole, but its distinctive nuclearfibrillar root is striated, unlike the nuclear fibrillarroots of clade A1. The fewer microtubulesassociated with the posterior ventral roots (twodistinct pairs totally absent) in C-80 may be theprimitive state and its absence of secondarymicrotubules a derived state for Cercomonadidaeas the heteromitid Katabia lacks the former buthas the latter (Karpov et al. 2003a). Our prelimin-ary serial sectioning study of one clade B strainshows that it is even more different from A1 than isA2, and relatively simple, suggesting that severaldistinctive characters of A1 are synapomorphiesfor it and not primitive characters of Cercomona-didae as a whole. We defer discussion of this untilour clade B data are published.
Common Features of Clade A Flagella andKinetids
Centrioles attach to each other obtusely or nearlyorthogonally. In C-43, thin striation of the fibrillarmain and broadest anterior bridge, connectingboth centrioles, suggests its possible contractionand a flexibility of angle between the centrioles inlife. If so, the angle might not be important forkinetid description in ‘Cercomonas’. More essen-tial is the universal attachment of the posteriorcentriole to the surface of the anterior one at itsapproximate middle. Precisely this mode ofattachment and approximately orthogonal cen-triolar orientation was found in all investigated‘Cercomonas’ (Karpov 1997; Mignot and Bruger-olle 1975; Mylnikov 1986b, c 1987, 1989a, b,1990, 1992a, b, 1995, 2000, 2001; Mylnikov andMylnikova 2001; Mylnikov et al. 2000; Schusterand Pollack 1978): a very conservative character.
In all investigated strains five features of the rootsystem are conserved: an association of twoposterior ventral microtubular roots; left anteriorroot; dorsal anterior root; microtubular cone. Theanterior centriole has two microtubular roots inclade A1 and three in clade A2. The posteriorcentriole has one microtubular root in all fivestrains. The left anterior root is common forboth centrioles as it originates from their feetfusion, an important character found in Katabiaand Heteromita also (Karpov et al. 2003a). Mostroots pass to the left dorsal side of the cell, andthe nuclear projection of A1 is from the left dorsalside of the centrioles, indicating a conservedgeometry.
An ER cisterna was applied to root VP2 in allstrains. A similar association of ER with centriolesis common in animals, choanoflagellates (Karpovand Leadbeater 1998) and some bicoecids(Karpov, unpublished); it might be present butoverlooked in other Cercozoa and protists, andnot distinctive for cercomonads. All strains hadfibrillar roots to the nucleus, but their origin andcomposition are slightly different, and not con-servative.
Comparison of our Strains with OtherInvestigated ‘Cercomonas’ Species
Several strains were previously studied (Table 2),but none using serial sections, so detail was muchless, and the origin, direction and exact location ofthe roots were not known. The main characterpreviously noted in ‘Cercomonas’ root systemswas the microtubular cone — a set of singlemicrotubules originating usually from the foot ofthe anterior centriole and directed to the nucleus(Karpov 1997; Mylnikov 1986a—c, 1987, 1989a, b,1990, 1992a, b, 1995, 2000; Mylnikov andMylnikova 2001; Mylnikov et al. 2000). It wasaccepted as a character of family Cercomonadi-dae, absent in Heteromitidae (Mylnikov andKarpov 2004). We found a cone in all investigatedclade A strains, confirming its taxonomic signifi-cance. But the cone microtubules may originatefrom the anterior centriole (usually from its smallposterior foot: clade A1a), from the base of thedorsal anterior root as secondary microtubules(C-80), or from the posterior centriole andfibrillar root, as in C-43. Some secondary micro-tubules from the left anterior root also pass to thenucleus. This emphasizes the ability of ‘Cercomo-nas’ kinetids to extend microtubules towards thenucleus, possibly as spindle remnants afternuclear division.

ARTICLE IN PRESS
152 S.A. Karpov et al.
One strain shown here to belong to clade A1was studied previously: C. plasmodialis (Cs-4). Ithas a nuclear projection and obvious nucleolus,but has tubular non-flattened cristae and twokinds of extrusomes: trichocysts and osmiophilicbodies (Mylnikov et al. 2000, and our unpublisheddata). Our unpublished data show a distinctivenuclear striated root in Cs-4 even more prominentthan in C-80, indicating that in some respects itsmorphological characters fit those reported herefor clade A1, but it differs in a few, suggesting thatultrastructural diversity within Cercomonas (A1)may turn out to be even greater when more strainsare studied thoroughly. A1 characters werepresent in ‘C.’ activas (Mylnikov 1992a, Table 2).‘C’. sp. (Mylnikov 1992b), had a distal centriolardiaphragm and nuclear projection, and thus maybelong to clade A1.
Tubular flattened cristae were found in‘C.’ varians (Mylnikov 1986b), ‘C. longicauda’(Mylnikov 1987: there is no sequence evidencethat this C-1 strain is related to either misidentified‘longicauda’ of clade B), and P. metabolica(Mylnikov and Mylnikova 2001). They also have anuclear projection to the centrioles and thefirst two have osmiophilic bodies as extrusomes(Table 2). They could mostly belong to A1, butP. metabolica has microtoxicysts, not yet ob-served in A1. ‘C.’ varians (Mylnikov 1986b)resembles A1 in most respects, but its striatednuclear root is an A2 character. Unlike in allother A1 strains, the cartwheel was not found inthe anterior centriole of C. plasmodialis (Mylnikovet al. 2000); perhaps it was overlooked. A cartwheelwas found in both centrioles of ‘C.’ cometa (C-52:Mylnikov 1990) and ‘C.’ sp. (C-57) (Table 2),suggesting that they may belong in clade B.
Reviews of ‘Cercomonas’ transition zones em-brace at least six species (Mylnikov 1995; Karpovand Fokin 1995). Most belong to a short type withone partition (transverse plate level with theplasma membrane), but some may have twopartitions. ‘C.’ cometa (Mylnikov 1990, 1995) hasa distal centriolar diaphragm, unlike our C-80E. ramosa, which has a very thin transverse plate.This ultrastructural difference and the differentflagella lengths suggest that the original identifica-tion of C-80 as C. cometa was mistaken. By othercharacters ‘C.’ cometa (strain C-52) is moresimilar to clade A1 than to clade A2 (Table 2).‘C.’ activas has two distal partitions: one corre-sponds with the transverse plate and another tothe centriolar distal partition (Mylnikov 1992a,1995). Concentric rings present in transitionalregions of all members of clade A were found
earlier in many cercomonads (Karpov and Fokin1995; Mylnikov 1995; called a cylinder). As we donot know to which clade several of these speciesbelong, we cannot deduce their evolutionaryhistory. This emphasizes that all future ultrastruc-tural studies of cercomonads should be on strainscharacterized by rRNA sequencing.
Centriolar Transformation in Cercomonas
An important conclusion of our study is thatflagellar transformation occurs in Cercozoa andthat the direction of transformation is from ayounger anterior flagellum to an older posteriorflagellum, as in all well-studied bikonts (Cavalier-Smith 2002). This follows from the fact that in allfour clade A1 Cercomonas strains the posteriorcentriole is devoid of a cartwheel. Yet a cartwheelis present in both daughter centrioles, as in alleukaryotes. This means that later in developmentthe cartwheel must be removed from the posteriorcentriole alone. The simplest interpretation of ourevidence for the secondary removal of the cart-wheel is that this is associated with flagellartransformation. We suggest that the anteriorcentriole and flagellum is transformed into theposterior centriole and flagellum in the next cellcycle after it is first assembled, as has beendemonstrated to occur in many protists belongingto the plant and chromist kingdoms and to theprotozoan infrakingdoms Alveolata and Excavata(Moestrup 2000). Our observations are the firstevidence for centriole and flagellar transformationin any members of the protozoan infrakingdomRhizaria. The pattern of transformation is from ayoung anterior to older posterior, precisely aspredicted by Cavalier-Smith (2002) to be theancestral state for all bikonts, not from a youngerposterior to an older anterior as suggested byMoestrup (2000) by analogy with the mycetozoanPhysarum.
Cercomonad Kinetids Differ Radically fromthose of Mycetozoa
Preliminary studies of kinetids of Cercomonadidawere used to argue that Mycetozoa and Cerco-monadida are related (Karpov 1997). This idea isnot supported by molecular phylogeny andCavalier-Smith (2002) argued in detail that thesimilarities between flagellar roots of Mycetozoaand Cercomonas on which the idea of a directrelationship between them was based are super-ficial. Our present study shows that Cercomonas

ARTICLE IN PRESS
153Cercomonad Kinetids and Phylogeny
clade A does indeed have very different roots fromMycetozoa (compare with recent reconstructionsof three myxogastrids: Karpov et al. 2003b). Thisconfirms the overwhelming molecular evidence(Cavalier-Smith and Chao 2003) that Cercomona-dida do not belong within Mycetozoa as hassometimes been suggested (Karpov 2000b). Pre-viously flagellar transformation from a youngeranterior to an older posterior flagellum was clearlydemonstrated for many members of the plantkingdom, many chromalveolates and some ex-cavates. Our demonstration that precisely thesame pattern is present in Cercozoa, whichbelong to the only previously unstudied majorgroup of eukaryotes, Rhizaria (Cavalier-Smith2002), means that this pattern of flagellar trans-formation has now been found in all major bikontgroups, as predicted (Cavalier-Smith 2002), andthus is indeed a synapomorphy for bikonts as awhole, being never found in unikonts such asMycetozoa (see Cavalier-Smith et al. 2004). Thisstrongly supports the idea that bikonts are aclade, in agreement with evidence that the rootof the eukaryote tree lies between bikontsand unikonts (Cavalier-Smith 2002, 2003a, b;Richards and Cavalier-Smith 2005; Stechmannand Cavalier-Smith 2002, 2003a, b).
Comparisons with Katabia and otherBikonts
Reasonably convincing homologies can be drawnbetween the roots of Cercomonas and Eocerco-monas and those of the heteromitid Katabia(Karpov et al. 2003a), even though Katabia (likeHeteromita globosa) has no microtubular cone.The only Katabia root directly attached to theanterior centriole has two microtubules andappears positionally homologous to the 2-micro-tubule dorsal anterior root of clade A; it differs bybeing associated laterally with a fibrous band, andis thus a composite root (thus was labeled CR),not a simple root — we found a similar compositeroot in Cercomonas clade B (Karpov et al. inpreparation), so the associated fibrous band mayhave been lost by the ancestor of clade A. Likeclade A Katabia has a 2-microtubule left anteriorroot with secondary microtubules, starting alsofrom fibrillar material associated with the anteriorcentriole, near to the feet fusion; we suggest thattheir left anterior roots may be homologues; that ofKatabia was previously called the upper root (UR);the 2-microtubule posterior-pointing root calledLR in Katabia lacks an obvious homologue inclade A cercomonads.
Katabia has a ‘posterior band’ of 4—5 micro-tubules originating from the dorsal side of bothcentrioles that passes under the plasma mem-brane alone to the posterior; this is arguablypositionally homologous to cercomonad root VP2(with three microtubules in A2 and proximally withthree and distally with six microtubules in A1) as itinitiates in almost the same place — beside ratherthan directly on the side of the anterior centriole.Katabia has a ‘girdle root’ of 3 microtubulesstemming from the posterior centriole, whichinitially passes posteriorly beside the posteriorband, then turns around the centrioles to the rightside of the cell; the proximal association of thegirdle and posterior bands is similar to that ofroots VP1 and VP2 in C-80 (Fig. 13 M) and thegirdle band stems from the same point on theposterior centriole as VP1. Since Katabia’s girdleband is also proximally ventral to the posteriorband, as VP1 is to VP2, we suggest that these tworoots are probably homologues, even though thedistal path of the girdle band is much moredivergent than for VP1, and that the characteristicproximal association of VP1 and VP2 on the left ofthe posterior centriole may be a synapomorphy forSarcomonadea (heteromitids plus cercomonads).To test this and establish the ancestral state forFilosa, comparably detailed studies are neededfor more divergent lineages, e.g. Metopion andMetromonas. There appear to be no cercomonadhomologues of the 2-microtubule anterior band ofthe posterior centriole of Katabia. Thus fourmicrotubular roots may be homologous betweenheteromitids and cercomonads. Information onHeteromita globosa (Macdonald et al. 1977) andH. sp. (Karpov 1997) is too limited for usefulcomparison: two posterior bands and a leftanterior root were identified, but without morethorough study of Heteromita we cannot saywhether it really lacks other roots identified inKatabia.
Our conclusion of four distinct conservedmicrotubular roots for sarcomonads is importantfor eventually reconstructing the ancestral bikontkinetid. Root VP1 may be positionally homologouswith left root r1 of plants, chromalveolates, andexcavates (Cavalier-Smith 2003b; Moestrup 2000;Simpson and Roger 2004), but in marked contrastto most of these other bikonts there appears to beno right posterior microtubular root (r2) in sarco-monads. Conversely, no obvious homologue ofthe sarcomonad root VP2 is present in otherbikonts. Spongomonad cercozoans have only onemicrotubule band associated with each centriole,which might either be the ancestral state for all

ARTICLE IN PRESS
154 S.A. Karpov et al.
Cercozoa (Cavalier-Smith 2002) or a secondarysimplification. Kinetid structure must be deter-mined from several more cercozoan classesbefore the ancestral state of Cercozoa, and thenbikonts as a whole, can be reliably reconstructed.
Methods
Isolation and maintenance of cultures: TheMylnikov (Borok, Russia) cultures were isolatedfrom these sites: Cs-4, C-56 and Cs-9 from soil(Borok, Russia), CeS-2 from freshwater lake(Germany), C-43 from sewage water (Russia),C-72 Sphagnum bog (Russia), C-85 from smallpond (Borok, Russia), C-59 from freshwater lakeC-71 and C-84 from river (Russia), C-76 and C-80from a lake (Antarctica). P. marina was from ATCC(‘longicauda’ #50344). The Mylnikov strains wereall clonal cultures made by isolating single cells bya glass micropipette, and maintained in Petridishes on artificial Pratt medium (KNO3 0.1%,K2HPO4 � 3H2O 0.01%, MgSO4 �7H2O 0.01%,FeCl3 �6H2O 0.001%; pH 6.5—7.5) with the addi-tion of bacteria Klebsiella aerogenes or wheatgrains as the source of food at 20—22 1C. TheBass (Oxford) strains were isolated from crude soiland freshwater samples immediately after collec-tion by serial dilution into a 96-well cell cultureplate (Nunclon). Volvic mineral water (Danone) wasadded to soil samples immediately after collec-tion, without flooding them, to facilitate pipettingafter a period of incubation at room temperaturefor approximately 1 week. Serial dilution wasachieved by inoculating 250ml of medium (ca.150 ml Volvic water which had previously beenboiled in the microwave for 3 min with ca. 20grains of untreated wheat or barley; subsequentlymaintained at 4 1C) with 50—100ml of crudeculture. This was mixed by repeatedly pipettingup and down; then the same volume as theinocculant was transferred to a second wellcontaining 250ml fresh medium. This manoeuvrewas repeated 7 times for each strain. The 96-wellplate was sealed with parafilm and left at roomtemperature to incubate for 1—2 weeks. The wellswere then examined and cultures grown fromthose wells containing only ‘Cercomonas’ andbacteria. The serial dilution procedure was re-peated in cases where the final well containedother eukaryotes as well as ‘Cercomonas’. Stockcultures are maintained at 14 1C in the samemedium, including the cereal grain, and subcul-tured ideally at 3-monthly intervals. Cultures areavailable from the authors upon request.
Light microscopy: Light microscopy on livingmaterial in Russia used the Biolam-I (Russia)microscope equipped with a video camera MC-1009/S (Germany) and phase contrast waterimmersion 70�objective (KF-5 device, Russia).Development of the cultures was recorded by aPanasonic NV-HS850 (Japan) video recorder. Thevideotape recording was digitized by PC compu-ter and saved in formats such as avi, tiff. TheOxford strains were photographed using a � 40dry phase contrast lens on an invertedOlympus IX70 microscope equipped with a NikonCoolpix 995 digital camera. Digital (.jpeg) imageswere transferred to Photoshop v.5.5 (Adobe) forprocessing.
DNA purification, sequencing, phylogeneticanalysis: Cultures were harvested after reachingstationary growth phase by gently scraping thebottom of the culture dish with a sterile scraper,and concentrating the cells by one of twomethods: filtering under low pressure through aglass-fibre filter (Whatman GF/F) which was cutinto small pieces before the first (homogenization)step, or centrifuging the liquid culture at 2,000 rpmin a benchtop centrifuge for 10—15 min at 5 1C,then pipetting out the pellet of organic material.Total DNA was extracted using the UltraClean SoilDNA Isolation Kit (Mo Bio Laboratories), subject-ing either the finely chopped filter or 200ml ofmedium with organic pellet to the first (homo-genization) step or directly from 1 g soil samples.18S rRNA gene sequences were amplified by PCRwith one or two of three primer pairs: (1) the‘Cercomonas’ Clade A-specific 243F (50-CCA-ATGCACCCTCTGGGTGGTT-30) and the ‘Cerco-monas’ Clade A-biased 1733R (50-TGATCAA-GTTTGATTCAGTTCTCGGAT-3’); (2) 25F (50-CAT-ATGCTTGTCTCAAAGATTAAGCCA-30) and thecercozoan-specific 1256R (50-GCACCACCACC-CAYAGAATCAAGAAAGAWCTTC-30); (3) the Cer-cozoa-specific 1259F 50-GGT CCR GAC AYA GTRAGG ATT GAC AGA TTG AAG-30 and generaleukaryote 369R 50-TCG CAT TAC GTA TCG CATTTC GCT G-30 under cycling conditions: 5 min at95 1C, 30 cycles of (32 s at 95 1C, 36 s at 70 1C,and 3 min 30 s at 72 1C), then 8 min at 72 1C. Theproducts were run on a 1% agarose gel. Bands ofcorrect length were excised, cleaned using a GFXPCR DNA Gel Band Purification Kit (AmershamBiosciences), sequenced using dye terminatorsand separated on an automated ABI-377 sequen-cer. Sequencing primers were the initial PCRprimers and an internal primer Pre3NDf (50-CAGCAGGCGCGCAAATTACC-30). Complete se-quences were obtained from cultures but only

ARTICLE IN PRESS
155Cercomonad Kinetids and Phylogeny
partial ones from soil samples. Sequences wereedited in the trace editor TED and deposited inGenBank under accession numbers AY884312—AY884342 and DQ442884.
Sequences were aligned by eye with all existing‘Cercomonas’ sequences in our database usingGenetic Data Environment software. These se-quences included those from other cultures andfrom environmental libraries constructed usingCercozoa-specific primers (Bass and Cavalier-Smith 2004). Gaps and ambiguously alignedpositions were removed, leaving 1013 positionsacross 56 sequences, including the five hetero-mitid outgroups. ModelTest v.3.06 (Posada andCrandall 1998) selected the GTR model withgamma correction for intersite rate variation(G; a ¼ 0.524154) and allowance for invariant sites(i ¼ 0.330625). These parameters were used fordistance trees constructed using PAUP 4.0b10(Swofford 1999). Heuristic searches (1000 repli-cates with no time limit per replicate; randomaddition) using minimum evolution were made(with TBR branch swapping). Bootstrap supportvalues were estimated using 500 bootstrapreplicates for each of 50 addition sequencereplicates for a distance analysis using the sameGTR+G+I model. Sequence alignments are avail-able on request from DB.
The Bayesian tree (Fig. 1) was calculated usingMrBayes v3.0b4 (Huelsenbeck and Ronquist2001) based on 56 taxa and 1013 positions. Twoseparate MCMC runs with randomly generatedstarting trees were carried out for 3 milliongenerations each with one cold and three heatedchains. The evolutionary model applied includedGTR substitution matrix, gamma correction withshape parameter estimated from the data, thecovarion model, and autocorrelation. Trees weresampled every 100 generations. A total of 5500trees were discarded as burn-in (trees sampledbefore the likelihood plots reached a plateau). Aconsensus of the remaining trees was generatedto reveal posterior probabilities of the branchingpattern. Both independent runs resulted in basi-cally identical tree topologies and posteriorprobabilities, indicating that they lasted longenough to converge.
Electron microscopy: Cells were fixed with afresh mixture of 0.6% osmium tetroxide with 1.4%glutaraldehyde (2 ml fixative (1 ml 2% of osmiumtetroxide+1 ml of 4.25% glutaraldehyde)+1 ml ofcells in medium) in 0.05 M phosphate buffer pH7.3 (final concentrations) at 0 1C in the dark for30—40 min. After dehydration in an alcohol seriesand acetone, the pellet was embedded in Araldite.
Blocks were serially sectioned with a diamondknife on a Reichert ultramicrotome, mounted onformvar-coated slot grids, and post-stained withuranyl acetate and lead citrate. Sections wereviewed on a JEM 100CX electron microscopeoperating at 80 kV. For three-dimensional recon-struction of kinetid patterns, serial sections of theflagellar apparatus of at least 10 cells of eachspecies were examined. To exclude potentialmistakes with rootlet interpretation, we did notanalyse micrographs of predivision stages, whenadditional centrioles appear; just prior to mitosis,additional roots appear in conjunction with cen-triole duplication.
Note added in proof
The newly described cercozoan Aurigamonas alsoexhibits transformation from young anterior to oldposterior flagellum (Vickerman et al. 2005).
Acknowledgements
The work was supported by a NERC researchgrant and Professorial Fellowship to TC-S andstudentship to DB, by RFBR grants 02-04-49987and 02-04-48265 to APM and SAK, and a jointproject grant from the Royal Society. TC-S thanksthe Canadian Institute for Advanced ResearchEvolutionary Biology Programme for Fellowshipsupport and Ema Chao for assistance.
References
Al-Qassab S, Lee WJ, Murray S, Simpson AGB,Patterson DJ (2002) Flagellates from stromatolitesand surrounding sediments in Shark Bay, WesternAustralia. Acta Protozool 41: 91—144
Arndt H, Dietrich D, Auer B, Cleven E-J, Grafen-ham T, Weitere M, Mylnikov AP (2000) FunctionalDiversity in Aquatic Ecosystems. In Leadbeater BS,Green JC (eds) The Flagellates: Unity and Diversity.Taylor & Francis, London, pp 240—268
Archibald JM, Longet D, Pawlowski J, Keeling PJ(2003) A novel polyubiquitin structure in Cercozoaand foraminifera: evidence for a new eukaryoticsupergroup. Mol Biol Evol 20: 62—66
Bass D, Cavalier-Smith T (2004) Phylum-specificenvironmental DNA analysis reveals remarkably highglobal biodiversity of Cercozoa (Protozoa). Int J SystEvol Microbiol 54: 2393—2404

ARTICLE IN PRESS
156 S.A. Karpov et al.
Bass D, Moreira D, Lopez-Garcıa P, Polet S, ChaoEE, von der Heyden S, Pawlowski J, Cavalier-Smith T (2005) Polyubiquitin insertions and thephylogeny of Cercozoa and Rhizaria. Protist 156:149—161
Cavalier-Smith T (1974) Basal body and flagellardevelopment during the vegetative cell cycle and thesexual cycle of Chlamydomonas reinhardii. J Cell Sci16: 529—556
Cavalier-Smith T (1998) A revised six-kingdomsystem of life. Biol Rev Cambridge Philos Soc 73:203—266
Cavalier-Smith T (2002) The phagotrophic origin ofeukaryotes and phylogenetic classification of Proto-zoa. Int J Syst Evol Microbiol 52: 297—354
Cavalier-Smith T (2003a) The excavate protozoanphyla Metamonada Grasse emend. (Anaeromona-dea, Parabasalia, Carpediemonas, Eopharyngia) andLoukozoa emend. (Jakobea, Malawimonas): theirevolutionary affinities and new higher taxa. Int J SystEvol Microbiol 53: 1741—1758
Cavalier-Smith T (2003b) Protist phylogeny and thehigh-level classification of Protozoa. Europ J Protis-tol 39: 338—348
Cavalier-Smith T, Chao EE (1996/7) Sarcomonadribosomal RNA sequences, rhizopod phylogeny, andthe origin of euglyphid amoebae. Arch Protistenkd147: 227—236
Cavalier-Smith T, Chao EE (2003) Phylogeny andclassification of phylum Cercozoa (Protozoa). Protist154: 341—358
Cavalier-Smith T, Chao EE (2006) Phylogeny andmegasystematics of phagotrophic heterokonts(kingdom Chromista). J Mol Evol 62: 388—420
Cavalier-Smith T, Chao EE, Oates B (2004)Molecular phylogeny of Amoebozoa and the evolu-tionary significance of the unikont Phalansterium.Europ J Protistol 40: 21—48
Dujardin F (1841) Histoire Naturelle des ZoophytesInfusoires. Roret, Paris
Ekelund F, Patterson DJ (1997) Some heterotrophicflagellates from a cultivated soil in Australia. ArchProtistenkd 148: 461—478
Ekelund F, Daugbjerg N, Fredslund L (2004)Phylogeny of Heteromita, Cercomonas and Thau-matomonas based on SSU rDNA sequences, includ-ing the description of Neocercomonas jutlandica sp.nov., gen. nov. Europ J Protistol 40: 119—135
Finlay BJ (2002) Global dispersal of free-living micro-bial eukaryote species. Science 296: 1061—1063
Finlay BJ, Clarke KJ (1999) Apparent globalubiquity of species in the protist genus Paraphyso-monas. Protist 150: 419—430
Foissner W (1991) Diversity and Ecology of SoilFlagellates. In Patterson DJ, Larsen J (eds) TheBiology of Free-Living Heterotrophic Flagellates.Clarendon Press, Oxford, pp 93—112
Fromentel FDE (1874) Etudes sur les Microzoairesou Infusoires Proprement dit. Masson, Paris
Hollande A (1942) Etude Cytologique et Biologiquede Quelques Flagelles Libres. Arch Zool exp gen-erale 83: 1—268
Huelsenbeck JP, Ronquist F (2001) MRBAYES:Bayesian inference of phylogenetic trees. Bioinfor-matics 17: 754—755
Karpov SA (1997) Cercomonads and their relation-ship to myxomycetes. Arch Protistenkd 158:297—307
Karpov SA (2000a) Flagellate Phylogeny: an Ultra-structural Approach. In Leadbeater BSC, Green JC(eds) The Flagellates: Unity, Diversity and Evolution.Taylor & Francis, London, pp 336—360
Karpov SA (ed) (2000b) Protista: Handbook onZoology Pt 1. Nauka, St. Petersburg
Karpov SA, Fokin SI (1995) The structuraldiversity of flagellar transitional zone in heterotrophicflagellates and other protists. Tsitologia 37:1038—1052
Karpov SA, Ekelund F, Moestrup Ø (2003a) Katabiagromovi nov. gen., nov. sp. — a new soil flagellatewith affinities to Heteromita (Cercomonadida). Pro-tistology 3: 30—41
Karpov SA, Novozhilov YK, Christiakova LV(2003b) A comparative study of zoospore cytoske-leton in Symphytocarpus impexus, Arcyria cinereaand Lycogala epidendrum (Eumycetozoa). Protistol-ogy 3: 15—29
Keeling PJ (2001) Foraminifera and Cercozoa arerelated in actin phylogeny: two orphans find a home?Mol Biol Evol 18: 1551—1557
Kent WS (1880—1882) A Manual of the Infusoria.Bogue, London
Klebs G (1892) Flagellatenstudien. Zeit wiss Zool 55:265—445
Krassilstschik I (1886) The materials to the naturalhistory and taxonomy of flagellates. On Cercobodolaciniaegerens nov. gen. et nov. sp. ZapiskiNovosibirskogoobschestva estestvoispytatelei. 2, 1,211—245 (in Russian)

ARTICLE IN PRESS
157Cercomonad Kinetids and Phylogeny
Lee WJ, Patterson DJ (2000) Heterotrophic flagel-lates (Protista) from marine sediments of BotanyBay, Australia. J Nat History 34: 483—562
Lemmermann E (1914) Flagellatae 1. GustavFischer, Jena
Macdonald CM, Darbyshire JF, Ogden CG (1977)The morphology of a common soil zooflagellate,Heteromita globosa Stein (Mastigophorea: Protozoa).Bull Br Mus Nat Hist (Zool) 31: 255—264 + 6 plates
Mignot J-P, Brugerolle G (1975) Etude ultrastruc-turale de Cercomonas Dujardin ( ¼ CercobodoKrassilstchick) protiste flagelle. Protistologica 11:547—554
Moestrup Ø (2000) The Flagellate Cytoskeleton:Introduction of a General Terminology for Micro-tubular Roots in Protists. In Leadbeater BS, GreenJC (eds) The Flagellates: Unity, Diversity and Evolu-tion. Taylor & Francis, London, pp 69—94
Mylnikov AP (1986a) The biology and ultrastructureof amoeboid flagellates Cercomonadida ord. n.Zoologicheskii Zhurnal, Moscow 65: 683—692 (inRussian with English summary)
Mylnikov AP (1986b) The peculiarities of ultrastruc-ture Cercomonas varians. Tsitologiya 28: 414—417(in Russian with English summary)
Mylnikov AP (1986c) Ultrastructure of a colourlessamoeboid flagellate, Cercomonas sp. Arch Protis-tenkd 131: 239—247
Mylnikov AP (1987) The ultrastructure of flagellateCercomonas longicauda (Cercomonadida) Bot Zhurn72: 750—753 (in Russian with English summary)
Mylnikov AP (1989a) The cell structure of flagellateCercomonas crassicauda. Biologia vnutrenih vodInform bul 84: 22—26 (in Russian)
Mylnikov AP (1989b) The rootlet flagellar system ofcercomonads. Biologia vnutrenih vod Inform bul 84:52—54 (in Russian)
Mylnikov AP (1990) The cell structure of flagellateCercobodo cometa. Biologia vnutrenih vod Informbul 86: 33—38 (in Russian)
Mylnikov AP (1992a) The cell structure of theflagellate Cercomonas activus. Biologia vnutrenihvod Inform bul N 92: 14—19 (in Russian)
Mylnikov AP (1992b) The structure of amoeboidflagellate Cercomonas sp. Biologia vnutrenih vodInform bul N 94: 41—45 (in Russian)
Mylnikov AP (1992c) Some taxonomic features ofcercomonads. Biologia vnutrenih vod Inform bul 93:35—39 (in Russian)
Mylnikov AP (1995). The Free-Living HeterotrophicFlagellates (Ultrastructure, Systematics and Biol-ogy). DSc Thesis. St. Petersburg: St. PetersburgState University (in Russian)
Mylnikov AP (2000). Class Cercomonadea Mylnikov,1986 — Cercomonads. In Alimov AF, StepanjantzSD, Karpov SA (eds) Protista, I: Handbookon Zoology. Nauka, St. Petersburg. pp 411—417(in Russian with English summary)
Mylnikov AP (2002) New amoeboid flagellates of thegenus Cercomonas (Cercomonadida, Protozoa)from lakes of Spitzbergen island. ZoologicheskiZhurnal Moscow 81: 1187—1192 (in Russian withEnglish summary)
Mylnikov AP, Mylnikova ZM, Tsvetkov AI (2000)The fine structure of amoeboid flagellate Cercomo-nas plasmodialis Mylnikov. Biologia vnutrenih vod N4: 29—35 (in Russian with English summary)
Mylnikov AP, Mylnikova ZM (2001) The mainultrastructure of amoeboid flagellate Cercomonasmetabolicus Mylnikov, 1987. Biologiya Vnutrenih VodN 2: 65—69 (in Russian with English summary)
Mylnikov AP, Karpov SA (2004) Review of thediversity and taxonomy of cercomonads. Protistol-ogy 3: 201—217
Nikolaev SI, Berney C, Fahrni JF, Bolivar I, PoletS, Mylnikov AP, Aleshin VV, Petrov NB, PawlowskiJ (2004) The twilight of Heliozoa and rise of Rhizaria,an emerging supergroup of amoeboid eukaryotes.Proc Natl Acad Sci USA 101: 8066—8071
Polet S, Berney C, Fahrni J, Pawlowski J (2004)Small-subunit ribosomal RNA gene sequences ofPhaeodarea challenge the monophyly of Haeckel’sRadiolaria. Protist 155: 53—63
Posada D, Crandall KA (1998) MODELTEST: testingthe model of DNA substitution. Bioinformatics 14:817—818
Richards TA, Cavalier-Smith T (2005) Myosindomain evolution and the primary divergence ofeukaryotes. Nature 436: 1113—1118
Sandon H (1927) The Composition and Distributionof the Protozoan Fauna of the Soil. Oliver & Boyd,Edinburgh
Schuster FL, Pollack A (1978) Ultrastructure ofCercomonas sp., a free-living amoeboid flagellate.Arch Protistenkd 120: 206—212
Simpson AGB, Roger AJ (2004) Excavata and theOrigin of Amitochondriate Eukaryotes. In Hirt RP,Horner DS (eds) Organelles, Genomes, andEukaryote Phylogeny, The Systematics Association

ARTICLE IN PRESS
158 S.A. Karpov et al.
Special Volume Series 68. CRC Press, Boca Raton,Florida, pp 27—33
Skvortzov BV (1977) The flagellates of clear andpolluted waters. New taxa of the genus CercobodoKrassil: a colourless flagellate of Rhizomastigaceae,Pantostomatinae. Q J Taiwan Mus Taipei 30: 89—121
Stechmann A, Cavalier-Smith T (2002) Rooting theeukaryote tree by using a derived gene fusion.Science 297: 89—91
Stechmann A, Cavalier Smith T (2003a) The root ofthe eukaryote tree pinpointed. Curr Biol 13:R665—R666
Stechmann A, Cavalier-Smith T (2003b) Phyloge-netic analysis of eukaryotes using heat-shockprotein Hsp90. J Mol Evol 57: 408—419
Stein FR (1878) Der Organismus der Infusionsthiere,III: Der Organismus der Flagellaten I. Engelmann,Leipzig
Swofford DW (1999) PAUP* 4.0b10. Sinauer, Sun-derland, MS
Vickerman K, Appleton PL, Clarke KJ, Moreira D(2005) Aurigamonas solis n. gen., n. sp., a soil-dwelling predator with unusual helioflagellate orga-nisation and belonging to a novel clade within theCercozoa. Protist 156: 335—354
von der Heyden S, Cavalier-Smith T (2005) Cultur-ing and environmental DNA sequencing uncoverhidden kinetoplastid biodiversity and a majormarine clade within ancestrally freshwaterNeobodo designis. Int J Syst Evol Microbiol 55:2605—2621
Wuyts J, De Rijk P, Van de Peer Y, Pison G,Rousseeuw P, De Wachter R (2000) Comparativeanalysis of more than 3000 sequences reveals theexistence of two pseudoknots in area V4 ofeukaryotic small subunit ribosomal RNA. NucleicAcids Res 28: 4698—4708
Zhukov BF (1993) Atlas of Freshwater HeterotrophicFlagellates. Russian Academy of Sciences,Institute of the Biology of Inland Waters, Rubinsk(in Russian)



![Molecular Phylogeny and Evolution. Introduction to evolution and phylogeny Nomenclature of trees Five stages of molecular phylogeny: [1] selecting sequences.](https://static.fdocuments.us/doc/165x107/56649e265503460f94b155ae/molecular-phylogeny-and-evolution-introduction-to-evolution-and-phylogeny.jpg)














