
Molecular cloning and characterization of the RAD1 gene of Saccharomyces cerevisiae
8
119 Gene, 26 (1983) 119-126 Elsevier GENE 902 Molecular cloning and characterization of the RADI gene of Succharomyces cerevisiae (DNA repair; Escherichiu coli; yeasts; incision; excision; pyrimidine dimers; deletion mutants; UV sensitivity) David R. Higgins, Satya Prakash, Paul Reynolds * and Louise Prakash * Department of Biology, University of Rochester, Rochester, NY 14627 (U.S.A.) Tel. (716) 275-3847, and *Department of Radiation Biology and Biophysics, University of Rochester, School of Medicine, Rochester, NY 14642 (U.S.A.) Tel. (716) 275-2656 (Received July 8th, 1983) (Revision received and accepted September lst, 1983) SUMMARY We have cloned the RADI gene of Saccharomyces cerevisiae and physically mapped it to a 4.0-kb DNA fragment from chromosome XVI. The RADI gene determines a transcript of 3.1 kb, and the direction of transcription was found to be leftwards, from EcoRI towards BglII (Fig. 1). Deletions of the RADI gene were made and were found to have no effect on viability of vegetative cells or spores, or on sporulation. INTRODUCTION In Escherichia coli, the uvrA, uvrB and uvrC genes are required for the removal of ultraviolet light(UV)- induced pyrimidine dimers from DNA (Boyce and Howard-Flanders, 1964; Setlow and Carrier, 1964). In addition to being extremely UV-sensitive, uvrA, uvrB and uvrC mutants show enhanced sensitivity to other DNA damaging agents which introduce distor- tions in the DNA, such as nitroquinoline oxide (Ikenaga et al., 1975), mitomycin C (Kondo et al., 1970), and psoralen plus near UV light treatment (Cole, 1971). The uvrA, uvrB and uvrC genes encode proteins of J4, 114000 (Sancar et al., 1981a), 84000 (Sancar et al., 1981b) and 70000 (Sancar et al., 198 Ic), respectively. The uvrABC endonuclease Abbreviations: kb, kilobases or kilobase pairs; poly(A)+ , polyadenylated; tet, tetracycline; UV, ultraviolet light; [ 1, indi- cates plasmid carrier state. 0378-l 119/83/$03.00 0 1983 Elsevier Science Publishers activity has been reconstituted in vitro and shown to incise UV-irradiated DNA, producing two cuts in the damaged DNA strand, one on each side of the pyrimidine dimer (Sancar and Rupp, 1983). In eukaryotic organisms, excision repair seems to require a large number of genes and is probably a more complex process than in E. coli. In the yeast S. cerevisiae, radl, rad2, rad3, rad4, rad7, radl0, radl4, rad16, rad23 and mms19 mutants are defective in excision of UV-induced pyrimidine dimers (Unrau et al., 1971; Resnick and Setlow, 1972; Waters and Moustacchi, 1974; Prakash, 1975; 1977a,b; Reynolds, 1978; Prakash and P&ash, 1979; Miller et al., 1982a), as well as of psoralen plus 360 nm light-induced interstrand DNA crosslinks (Miller et al., 1982a,b). Of these mutants, rudl, rud2, rud3, rad4 and radl0 have been shown to be defective in the incision step of excision repair (Wilcox and Prakash, 198 1; Reynolds and Friedberg, 1981). Attempts to characterize pyrimidine dimer-incising
Transcript of Molecular cloning and characterization of the RAD1 gene of Saccharomyces cerevisiae

119 Gene, 26 (1983) 119-126
Elsevier
GENE 902
Molecular cloning and characterization of the RADI gene of Succharomyces cerevisiae
(DNA repair; Escherichiu coli; yeasts; incision; excision; pyrimidine dimers; deletion mutants; UV sensitivity)
David R. Higgins, Satya Prakash, Paul Reynolds * and Louise Prakash *
Department of Biology, University of Rochester, Rochester, NY 14627 (U.S.A.) Tel. (716) 275-3847, and *Department of
Radiation Biology and Biophysics, University of Rochester, School of Medicine, Rochester, NY 14642 (U.S.A.) Tel. (716) 275-2656
(Received July 8th, 1983)
(Revision received and accepted September lst, 1983)
SUMMARY
We have cloned the RADI gene of Saccharomyces cerevisiae and physically mapped it to a 4.0-kb DNA
fragment from chromosome XVI. The RADI gene determines a transcript of 3.1 kb, and the direction of
transcription was found to be leftwards, from EcoRI towards BglII (Fig. 1). Deletions of the RADI gene were
made and were found to have no effect on viability of vegetative cells or spores, or on sporulation.
INTRODUCTION
In Escherichia coli, the uvrA, uvrB and uvrC genes
are required for the removal of ultraviolet light(UV)-
induced pyrimidine dimers from DNA (Boyce and
Howard-Flanders, 1964; Setlow and Carrier, 1964).
In addition to being extremely UV-sensitive, uvrA,
uvrB and uvrC mutants show enhanced sensitivity to
other DNA damaging agents which introduce distor-
tions in the DNA, such as nitroquinoline oxide
(Ikenaga et al., 1975), mitomycin C (Kondo et al.,
1970), and psoralen plus near UV light treatment
(Cole, 1971). The uvrA, uvrB and uvrC genes encode
proteins of J4, 114000 (Sancar et al., 1981a), 84000
(Sancar et al., 1981b) and 70000 (Sancar et al.,
198 Ic), respectively. The uvrABC endonuclease
Abbreviations: kb, kilobases or kilobase pairs; poly(A)+ ,
polyadenylated; tet, tetracycline; UV, ultraviolet light; [ 1, indi-
cates plasmid carrier state.
0378-l 119/83/$03.00 0 1983 Elsevier Science Publishers
activity has been reconstituted in vitro and shown to
incise UV-irradiated DNA, producing two cuts in
the damaged DNA strand, one on each side of the
pyrimidine dimer (Sancar and Rupp, 1983).
In eukaryotic organisms, excision repair seems to
require a large number of genes and is probably a
more complex process than in E. coli. In the yeast
S. cerevisiae, radl, rad2, rad3, rad4, rad7, radl0,
radl4, rad16, rad23 and mms19 mutants are defective
in excision of UV-induced pyrimidine dimers (Unrau
et al., 1971; Resnick and Setlow, 1972; Waters
and Moustacchi, 1974; Prakash, 1975; 1977a,b;
Reynolds, 1978; Prakash and P&ash, 1979; Miller
et al., 1982a), as well as of psoralen plus 360 nm
light-induced interstrand DNA crosslinks (Miller
et al., 1982a,b). Of these mutants, rudl, rud2, rud3,
rad4 and radl0 have been shown to be defective in
the incision step of excision repair (Wilcox and
Prakash, 198 1; Reynolds and Friedberg, 1981).
Attempts to characterize pyrimidine dimer-incising

120
activity in vitro from S. cerevisiae have not been
successful (Reynolds and Friedberg, 1980; Prakash,
L., unpublished results), perhaps because the activity
exists as part of a labile complex.
To identify and characterize the protein products
of the RAD genes involved in the incision process
and to examine the regulation of these genes following
treatment with DNA-damaging agents, we have
cloned several of these RAD genes. In this paper, we
report the isolation of the RADl gene by complemen-
tation of the radl mutants of S. cerevisiae and show
that the RADI gene determines a transcript of
3.1 kb. We also demonstrate that radl deletions are
viable, in contrast to rad3 deletions, which are reces-
sive lethals (Higgins et al., 1983).
MATERIALS AND METHODS
(a) Yeast and bacterial strains
The following yeast strains were used: 7799-K,
MATa his4-17 ura3-52 RAD’ (obtained from G.
Fink); DH25-lB, MATa his3-Al Zeu2-3 leu2-112
1~~1-1 trpl-289 ura3-52 radl-2; DH50-2B, HO his-
ura3-52 RAD’ ; DH50-9B, HO lys- ura3-52 radl-2;
LP2693-6C, MATa leu2-3 leu2-112 trpl-289 ura3-52
RAD’ ; B-635, MATa ~~~1-115 hisl-1 lys2 trp2
RAD’.
E. coli strain JM103 was used to propagate Ml3
derivative phages. Yeast strains containing plasmids
(indicated by square brackets around plasmid) were
maintained and grown on selective media.
(b) Plasmids
The yeast recombinant library used, obtained from
D. Botstein, consisted of partial Sau3A digests of
yeast genomic DNA ligated into the BamHI site of
the 7.6-kb hybrid plasmid YEp24 (Botstein et al.,
1979; Carlson and Botstein, 1982). YEp24 consists
of pBR322, a 2.2-kb EcoRI fragment of yeast 2 p
circle DNA, form B, conferring the ability to replicate
autonomously in yeast, and a l.l-kb Hind111 frag-
ment containing the yeast URA3 gene, for selection
in yeast.
YIPS, the 5.5kb plasmid used for integrative
transformation, is also derived from pBR322 and
contains the yeast URA3 gene. However, it does not
replicate autonomously in yeast since it lacks a yeast
replication origin sequence and can transform only
by homologous integration into the yeast genome
(Scherer and Davis, 1979; Botstein et al., 1979).
YIp5 contains unique EcoRI, HindIII, BamHI and
Sal1 sites.
YEpl3 is a 10.7-kb plasmid, which is also derived
from pBR322 and which replicates autonomously in
yeast because of the 2 p origin of replication; it
contains the yeast LEU2 gene for selection in yeast
(Broach et al., 1979). All plasmids were maintained
in E. coli strain HBlOl.
(c) Transformation procedures and UV irradiation
The spheroplast method of Hinnen et al. (1978) or
the treatment of intact yeast cells with lithium acetate
to promote DNA uptake (Ito et al., 1983) were used
for transformation of yeast. Response to UV irradia-
tion of various yeast strains was determined by the
spot test or from survival curves following UV irradia-
tion as described by Prakash and Prakash (1977).
(d) Other methods
Nucleic acids were purified and electrophoresed
as described previously (Higgins et al., 1983).
Restriction fragments were transferred to nitrocellu-
lose paper by the method of Southern (1975) and
hybridizations were carried out according to the
method of Maniatis et al. (1982), except that a single-
stranded probe with the 1.6-kb BglII-EcoRI frag-
ment of pDH2 (Fig. l), prepared as described below,
was used in the hybridization mixture. The specific
activity ofthe probe was 4.3 x 10’ cpm/pg DNA and
a total of 4.3 x 10h cpm was used in the hybridiza-
tion mixture.
Poly(A)+ RNA was prepared by binding total
RNA to oligo-deoxythymidylate cellulose (type 3,
Collaborative Research) in 10 mM Tris . HCl,
pH 7.5-0.5 M NaCl and eluting the bound
poly(A)+RNA in 10 mM Tris. HCl, pH 7.5 (Aviv
and Leder, 1972). RNA sa.mples were fractionated
by electrophoresis in formaldehyde agarose gels,
transferred to “gene screen paper” (New England
Nuclear Corp.) as described by the manufacturer
and hybridized with “P-labeled DNA (Maniatis
et al., 1982). The 1.6-kb BglII-EcoRI restriction

fragment of pDH2 (Fig. 1) was cloned into BamHI-
EcoRI restricted M13mp8 and M13mp9 (Messing
and Vieira, 1982), and the method of Hu and
Messing (1982) was used to prepare the labeled
single-stranded probes. About 6 x lo6 cpm were
used per RNA-DNA hybridization. All autoradio-
graphs were exposed on Kodak XL-l (X-omat) film
at - 70°C with DuPont Lightning Plus intensifying
screens.
RESULTS
(a) Isolation of the RADZ gene
The pool of yeast genomic DNA in YEp24 was
used to transform the uracil requiring (Ursa),
UV-sensitive (Rad - ) yeast strain DH25-1B (~~3-52
r&l-2) to uracil prototrophy (Ura’ ). Approx.
20000 transformants were screened for UV resist-
ance by spot testing and six UV-resistant (Rad’)
colonies were obtained. Subcloning and retesting of
these six transformants revealed that the genetic
markers present in the original untransformed strain,
as well as UV resistance, were retained. Plasmid
DNA from each of the six UV-resistant transfor-
mants was prepared by a modification of the Hirt
procedure (1967) and used to transform E. coli strain
HB 101 to ampicillin resistance. Plasmid DNA from
these transformed E. coli strains was then used to
121
transform the uru3-52 rudl-2 yeast strain DH25- 1B
to Ura+ . 60 Ura+ transformants were tested from
each of the six transformations and in all six cases,
the Ura+ transformants were also UV-resistant,
indicating that UV resistance was carried on the
plasmid. The six plasmids were designated pDH 1 to
pDH6. Plasmids pDH 1, pDH2, pDH3 and pDH5,
each containing an 8.5-kb insert of yeast DNA, gave
identical patterns of digestion with Hind111 and other
restriction enzymes, whereas plasmids pDH4 and
pDH6 contained an additional 3 kb of yeast DNA.
A restriction map of the yeast DNA insert containing
the rudl complementing function in plasmid pDH2
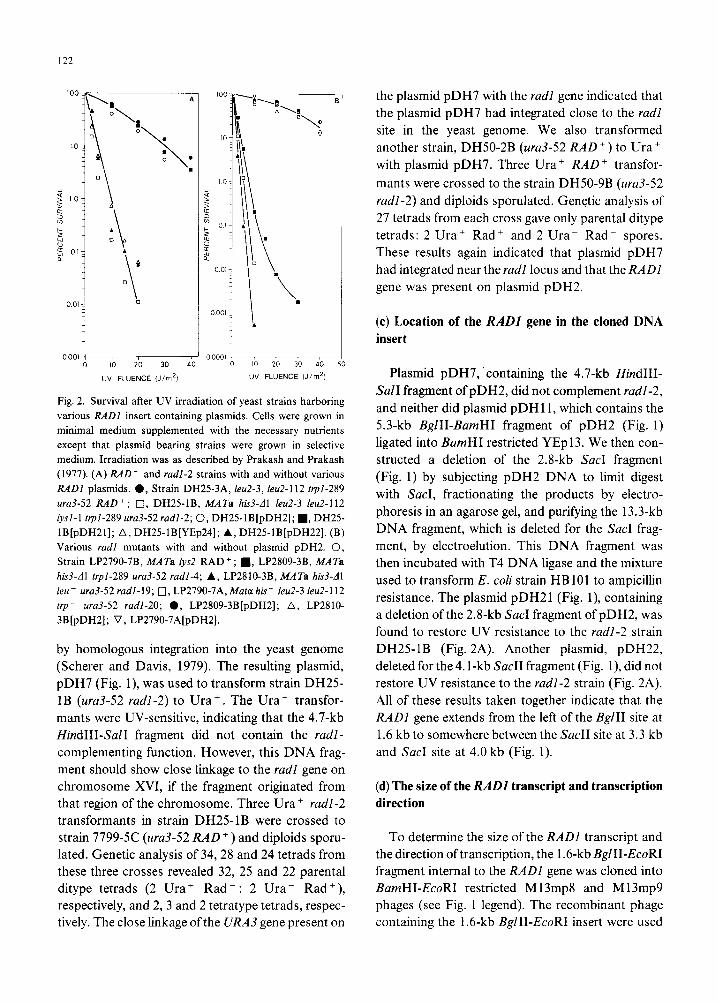
is given in Fig. 1. UV survival curves of various radl mutants, radl-2, radl-4, radl- 19 and radl-20, carry-
ing the plasmid pDH2 show normal levels of UV
resistance (Fig. 2).
(h) Genetic mapping of the cloned DNA segment
To verify that the yeast DNA insert in the pDH2
plasmid contained the yeast RADl gene and not a
suppressor of radl-2, or some other gene which
could complement radl-2 when present on a multi-
copy plasmid, we genetically mapped the 4.7-kb
HindIII-Sal1 fragment of pDH2 (Fig. 1). The 4.7-kb
HindIII-Sal1 fragment of plasmid pDH2 was ligated
into Hind111 + WI-restricted YIPS, a plasmid
containing the yeast URA3 gene but not the se-
quences required for autonomous replication in
yeast. Transformation with YIPS, therefore, occurs
Lb 6 i i j k j k ; i 1 Plarmtd Complementotion
E SII E SI H E SPSIB SII of rod!
__ __ pDH2 +
I pDH7
I I pDHII
__ 1 [+-- pDH2l +
__ I II-- pDH22 -
I I pDH23
Fig. 1. Restriction map ofthe yeast DNA insert in plasmid pDH2 and complementation pattern of pDH2 derivatives. Insert ofthe yeast
DNA segment in pDH2, given by the open bar, is in the BarnHI site of the ret gene of YEp24; the thin line represents YEp24 DNA.
* denotes the Sau3Al-BamHI junction of the fusion of insert with plasmid. Symbols for restriction enzymes are as follows: B, BarnHI;
Bg, BglII; E, EcoRI; H, HindIII; K, KpnI; SI, SacI; SII, SacII; Sa, SalI; Sp, SphI. The open bars below plasmid pDH2 represent DNA
segments of pDH2 cloned in various vectors, as indicated in the text. The 1.6-kb BglII-EcoRI fragment used to generate plasmid pDH23
was also inserted into M13mp8 and M13mp9 phages and used to generate single stranded probes for hybridization to yeast RNA to
determine the size and direction of the RADI transcript. The wavy line with the arrowhead below the DNA insert of plasmid pDH23
denotes the direction of the RADI transcript.
ooolM oooo’ UV FLUENCE (J/m’)
-0kFX-
UV FLUENCE (J/m*)
50
Fig. 2. Survival after UV irradiation of yeast strains harboring
various RADl insert containing plasmids. Cells were grown in
minimal medium supplemented with the necessary nutrients
except that plasmid bearing strains were grown in selective
medium. Irradiation was as described by Prakash and Prakash
(1977). (A) RAD’ and radl-2 strains with and without various
RADl plasmids. 0, Strain DH25-3A, leu2-3, leu2-112 ~I-289
ura3-52 RAD + ; 0, DH25-lB, MATa hb3-dl IeuZ-3 leu2-112
lysl-1 trpl-289 ~~3-52 radl-2; 0, DH25-lB[pDH2]; n , DH25-
lB[pDHZI]; A, DH25-lB[YEp24]; A, DH25_1B[pDH22]. (B)
Various rudl mutants with and without plasmid pDH2. 0,
Strain LP2790-7B, MATa rysZ RAD+; n , LP2809-3B, MATa his3-Al trpl-289 ura3-52 radl-4; A, LP2810-3B, MATa hW-Al
leu- ura3-52 radl-19; 0, LP2790-7A,Mata his- IeuZ-3 leu2-112
trp- ura3-52 radl-20; 0, LP2809-3B[pDH2]; A, LP2810-
3B[pDH2]; V, LP2790-7A[pDH2].
by homologous integration into the yeast genome
(Scherer and Davis, 1979). The resulting plasmid,
pDH7 (Fig. l), was used to transform strain DH25-
1B (ura3-52 r&l-2) to Ura+. The Ura+ transfor-
mants were UV-sensitive, indicating that the 4.7-kb
KndIII-Sal1 fragment did not contain the radl- complementing function. However, this DNA frag-
ment should show close linkage to the radl gene on
chromosome XVI, if the fragment originated from
that region of the chromosome. Three Ura+ radl-2 transformants in strain DH25-1B were crossed to
strain 7799-K (uru3-52 RAD + ) and diploids sporu-
lated. Genetic analysis of 34,28 and 24 tetrads from
these three crosses revealed 32, 25 and 22 parental
ditype tetrads (2 Ura+ Rad : 2 Ura- Rad +),
respectively, and 2,3 and 2 tetratype tetrads, respec-
tively. The close linkage ofthe URA3 gene present on
the plasmid pDH7 with the rudl gene indicated that
the plasmid pDH7 had integrated close to the radl site in the yeast genome. We also transformed
another strain, DH50-2B (ura3-52 RAD + ) to Ura +
with plasmid pDH7. Three Ura+ RAD’ transfor-
mants were crossed to the strain DH50-9B (ura3-52 radl-2) and diploids sporulated. Genetic analysis of
27 tetrads from each cross gave only parental ditype
tetrads: 2 Ura+ Rad’ and 2 Ura- Rad- spores.
These results again indicated that plasmid pDH7
had integrated near the radl locus and that the RADl gene was present on plasmid pDH2.
(c) Location of the RADl gene in the cloned DNA
insert
Plasmid pDH7, ‘containing the 4.7-kb HindIII-
Sal1 fragment of pDH2, did not complement radl-2, and neither did plasmid pDH 11, which contains the
5.3-kb BglII-BumHI fragment of pDH2 (Fig. 1)
ligated into BamHI restricted YEp13. We then con-
structed a deletion of the 2.8-kb Sac1 fragment
(Fig. 1) by subjecting pDH2 DNA to limit digest
with SacI, fractionating the products by electro-
phoresis in an agarose gel, and purifying the 13.3-kb
DNA fragment, which is deleted for the Sac1 frag-
ment, by electroelution. This DNA fragment was
then incubated with T4 DNA ligase and the mixture
used to transform E. coli strain HB 101 to ampicillin
resistance. The plasmid pDH21 (Fig. l), containing
a deletion of the 2.8-kb Sac1 fragment of pDH2, was
found to restore UV resistance to the radl-2 strain
DH25-1B (Fig. 2A). Another plasmid, pDH22,
deleted for the 4.1-kb Sac11 fragment (Fig. l), did not
restore UV resistance to the radl-2 strain (Fig. 2A).
All of these results taken together indicate that the
RADl gene extends from the left of the Bg111 site at
1.6 kb to somewhere between the Sac11 site at 3.3 kb
and Sac1 site at 4.0 kb (Fig. 1).
(d) The size of the RADZ transcript and transcription
direction
To determine the size of the RADl transcript and
the direction of transcription, the 1.6-kb BglII-EcoRI
fragment internal to the RADl gene was cloned into
BamHI-EcoRI restricted M 13mp8 and M 13mp9
phages (see Fig. 1 legend). The recombinant phage
containing the 1.6-kb BglII-EcoRI insert were used

123
as templates for DNA synthesis to generate radio-
actively labeled single-stranded probes, as described
by Hu and Messing (1982). A hybridization probe
primer (New England Biolabs), which anneals 45
nucleotides 5’ of the cloning site and therefore does
not prime DNA synthesis through the cloned insert,
was used to synthesize the complementary DNA
strand of either M 13mp8 or M13mp9. The
reaction, carried out in the presence of a radioactively
labeled deoxynucleoside triphosphate did not
proceed to completion under the conditions employ-
ed (Hu and Messing, 1982). Thus, the radioactively
labeled reaction product was single-stranded in the
region of the yeast insert, and the 5’ to 3’ orientation
of the RNA strand hybridizing to the single-stranded
DNA insert is opposite to the polarity of the DNA
strand with which it hybridizes. The labeled probes
were used in hybridizations to total yeast RNA from
the RAD’ strain B-635 and the rudl-2 strain
DH25- 1B with and without the pDH2 plasmid, and
to poly(A) + RNA from the RAD + strain B-635. The
RADl transcript is more abundant in the radl-2
strain containing the RADl gene on a multicopy
plasmid, pDH2 (Fig. 3A, lane 4) than in either the
A B kb
123 4
Fig. 3. Transcript ofthe RADl gene. Total RNA (20 fig per lane)
and 10 pg poly(A)‘RNA from yeast cells were fractionated on
formaldehyde-1.5%-agarose gels, transferred to gene screen
paper, and hybridized to the 1.6-kb BglII-EcoRI fragment of the
RADZ gene cloned into Ml3mp9. (A) Lane 1, total RNA from the
RAD+ strain B-635; lane 2, total RNA from the mdl-2 strain
DH25-1B; lane 3, total RNA from the radl-2 strain DH25-1B
containing the plasmid YEp24 without any additional yeast
DNA insert; lane 4, total RNA from the radl-2 strain DH25-IB
containing the radl complementing plasmid pDH2, the lower
narrow band being ribosomal RNA; (B) poly(A) + RNA from the
RAD + strain B-635.
RAD’ (Fig. 3A, lane 1) or radl-2 strain lacking the
pDH2 plasmid (Fig. 3A, lanes 2 and 3); the RADI
transcript is also more abundant in poly(A) + RNA
from the RAD+ strain (Fig. 3B) than in total RNA
from the RAD’ strain (Fig. 3, lane 1). The 3.1-kb
RADI transcript hybridizes to the 1.6-kb BglII-
EcoRI fragment of the RADI gene cloned in
M13mp9 (Fig. 3A, lane 4 and Fig. 3B), but not when
this fragment is cloned in M13mp8 (not shown).
Since the cloning sites of M13mp9 are arranged as
5’-BamHI-EcoRI-3’, and those of M13mp8 are
5’-EcoRI-BarnHI-3’, these results indicate that the
direction of transcription of the RADl gene is from
EcoRI to BglII (Fig. 1).
(e) Deletions of the RADl gene
Deletions of the RAD3 gene, which like the RADl
gene, is required for incising UV-damaged DNA, are
recessive lethals (Higgins et al., 1983). Deletions of
the RADl gene were generated by the method of
Shortle et al. (1982). The 1.6-kb BglII-EcoRI frag-
ment (see Fig. l), which is an internal fragment of the
RADl gene, was cloned into BumHI and EcoRI
restricted plasmid YIPS. The 6.7-kb plasmid thus
generated, pDH23 (Fig. l), was used to transform
the yeast strain 7799-5C (ura3-52 RAD + ) to Ura+ . Nine UV-sensitive Ura+ transformants were ob-
tained, which presumably resulted from integration
of the plasmid pDH23 at the RADI site in the yeast
genome, thereby creating deletions of the RADI
gene, as indicated in Fig. 4. Physical evidence for
integration of pDH23 at the RADl locus was
obtained by Southern analysis of yeast genomic
DNA from the recipient strain 7799-5C and the
Ura + transformants derived from it. Yeast genomic
DNA was restricted with BglII, fractionated by
electrophoresis in agarose gels, transferred to nitro-
cellulose by the method of Southern (1975), and then
hybridized to the nick translated 1.6-kb BgZII-EcoRI
fragment from pDH2. The size of the BglII fragment
from the recipient strain (Fig. 5, lane 1) hybridizing
to the RADI probe is 9.0 kb, and, as expected for
homologous integration at the RADI locus, the size
of the BglII fragment hybridizing to the RADI probe
from the 7799-5C[pDH23] transformants has in-
creased to 15.7 kb (Fig. 5, lanes 2 and 3).
In order to further ascertain that UV sensitivity in
these Ura + transformants was due to deletion at the

124
I- 16kb-
X
H HBg K E SII E SJ
CHROMOSOMAL RADl LOCUS . ..’ -f ’ a..
HOMOLOGOUS INTEGRATION AT RADl LOCUS
i H HBg K E E SII E SI
. . . . . . . .
Fig. 4. Generation of duplicated deletion of the RADI gene by homologous integration at the R.4DI locus of an internal segment of the
R&31 gene. The I&kb BglII-EcoRI fragment of pDH2 was ligated into BumHI-EcoRI restricted YIp5 to generate the plasmid pDH23.
As indicated diagramatically, integration by homologous recombination at the RAADI locus will generate a dupficated deletion at the
RADl locus flanking the YIp5 sequence. Symbols for restriction enzymes, as given in Fig. 1; thin line, pBR322 sequences; open segment,
RADl region DNA; hatched segment, yeast UK43 gene; heavy line, yeast chromosomal DNA.
1 2 3
kb
15.7
9.0
ML)1 locus, the Ura+ UV-sensitive tr~sform~ts were mated to the r&I-2 strain DH25-1B. The diploids were UV sensitive, as expected ifthe deletion mutation is allelic to the radl-2 mutation. Therefore, vegetatively growing haploid cells with a radl dele- tion are viable, unlike the rad3 deletion mutants. The radl deletion also had no effect on plating efficiency or growth rate. The rudl deletion mutants showed somewhat greater sensitivity to X-rays and methyl methanesulfonate than the radl-2 mutant, but similar levels of sensitivity as the other radl alleles examined, radl-4. radl-19 and radl-20. We then examined the viabiiity of radl deletion-bearing spores. Four Ura + UV-sensitive transformants in strain 7799-W were mated to the strain LP2693-6C (ura3-52 RAD + ), the diploids sporulated and 25-29 tetrads analyzed from each cross. In all four crosses, with the exception of
Fig. 5. Physical evidence for integration of plasmid pDH23
containing the I .6-kb BglII-EcoRI internal fragment ofthe RADl
gene at the RADl locus in the Yeast genome. . _ Plasmid pDH23 was used to transform the RAD’ ~~3-52
strain 7799-K to Ura +. Genomic DNA from the untransformed
strain (lane 1) and two W-sensitive transformallts (lanes 2 and
3) was purified and restricted with BglII. DNA fragments
separated by electrophoresis in agarose gels were transferred to
nitrocellulose and hybridized to 32P-labeled 1.6-kb BglII-EcoRI
fragment of pDH2.
one tetratype tetrad, only parental ditype tetrads showing 2 Ura+ Rad - and 2 Ural Rad + spores were obtained, demonstrating that Ura+ Rad - spores bearing the radl deletion are viable. The pre- dominance of parental ditype tetrads provides further genetic con~rmation for inte~ation of the plasmid pDH23 at the RAD1 site in strain 7799-5C. The radl Alradl A homozygotes were proficient in sporu- lation and produced viable spores.

DISCUSSION
We have cloned the RADl gene and used it to
determine the size of its transcript and the direction of transcription. The RADl gene encodes a 3.1-kb transcript and the direction of transcription is left- ward as shown in Fig. 1. Our subcloning results show that the RADI gene extends from the left of the BglII site at 1.6 kb to somewhere between the Sac11 site at 3.3 kb and Sac1 site at 4.0 kb (Fig. 1). The 3.1-kb RADl transcript could begin between the Sac11 and Sac1 sites on the right and end between the two Hind111 sites on the left (Fig. 1). The 3.1-kb transcript could code for a polypeptide of about 110 kDa1, similar in size to the uvrA protein ofE. coli.
We have also shown that deletions of the RADI
gene, created by integration of the internal 1.6-kb BglII-EcoRI fragment in the chromosomal RADI
gene, have no effect on viability or growth rate of vegetatively growing cells, or on sporulation or spore viability. Deletions of the RAD3 gene, which like the RADI gene is required for incising DNA containing pyrimidine dimers, are recessive lethals (Higgins et al., 1983). The RADl gene appears to be more specific to excision repair of damaged DNA, whereas the RAD3 gene is involved, in addition, in other vital cellular processes. Yeast cells containing the RADI
gene on a multicopy plasmid have much higher levels of RADI mRNA than cells with only the chromo- somal RADZ gene but show no alteration in growth rate or viability.
ACKNOWLEDGEMENTS
We thank Ravi Deshmukh for help with initial screening of transformants and Renata Polakowska for the Southern gel shown in Fig. 5. This investi- gation was supported by grants CA32514 and CA35035 to S.P. and GM19261 to L.P. from NIH.
REFERENCES
Aviv, H. and Leder, P.: Purification of biologically active globin
messenger RNA by chromatography on oligothymidylic
acid-cellulose. Proc. Natl. Acad. Sci. USA 69 (1972)
1408-1412.
125
Botstein, D., Falco, S.C., Stewart, S.E., Brennan, M., Scherer,
S., Stinchcomb, D.T., Struhl, K. and Davis, R.W.: Sterile
host yeasts (SHY): a eukaryotic system of biological contain-
ment for recombinant DNA experiments. Gene 8 (1979)
1 l-24.
Boyce, R. and Howard-Flanders, P.: Release of ultraviolet light-
induced thymine dimers from DNA in E. coli K-12.Proc.
Natl. Acad. Sci. USA 51 (1964) 293-300.
Broach, J.R., Strathern, J.N. and Hicks, J.B.: Transformation in
yeast: development of a hybrid cloning vector and isolation
of the CANI gene. Gene 8 (1979) 121-133.
Carlson, M. and Botstein, D.: Two differentially regulated
mRNAs with different 5’ ends encode secreted and intra-
cellular forms of yeast invertase. Cell 28 (1982) 145-154.
Cole, R.S.: Inactivation ofEscherichia cob, F’ episomes at trans-
fer, and bacteriophage lambda by psoralen plus 360-nm light:
significance of deoxyribonucleic acid cross-links. J. Bacterial.
107 (1971) 846-852.
Higgins, D.R., Prakash, S., Reynolds, P., Polakowska, R.,
Weber, S. and Prakash, L.: Isolation and characterization of
the RAD3 gene of Saccharomyces cerevisiae and inviability of
rad3 deletion mutants. Proc. Natl. Acad. Sci. USA 80 (1983)
5680-5684.
Hinnen, A., Hicks, J.B. and Fink, G.R.: Transformation ofyeast.
Proc. Natl. Acad. Sci. USA 75 (1978) 1929-1933.
Hirt, B.: Selective extraction on polyoma DNA from infected
mouse cell cultures. J. Mol. Biol. 26 (1967) 365-369.
Hu, N.-T. and Messing, J.: The making of strand-specific Ml3
probes. Gene 17 (1982) 271-277.
Ikenaga, M., Ichikawa-Ryo, H. and Kondo, S.: The major cause
of inactivation and mutation by 4-nitroquinoline l-oxide in
Escherichia cob: excisable 4 NQO-purine adducts. J. Mol.
Biol. 92 (1975) 341-356.
Ito, H., Fukuda, Y., Murata, K. and Kimura, A.: Transformation
of intact yeast cells treated with alkali cations. J. Bacterial.
153 (1983) 163-168.
Kondo, S., Ichikawa, H., Iwo, K. and Kato, T.: Base-change
mutagenesis and prophage induction in strains ofEscherichia
coli with different DNA repair capacities. Genetics 66 (1970)
187-217.
Maniatis, T., Fritsch, E.F. and Sambrook, J.: Molecular Cloning
- A Laboratory Manual. Cold Spring Harbor Laboratory,
Cold Spring Harbor, NY, 1982.
Messing, J. and Vieira, J.: A new pair of M 13 vectors for selecting
either DNA strand of double-digest restriction fragments.
Gene I9 (1982)269-276.
Miller, R.D., Prakash, L. and Prakash, S.: Defective excision of
pyrimidine dimers and interstrand DNA crosslinks in rad7
and rad23 mutants of Saccharomyces cerevisiae. Mol. Gen.
Genet. 188 (1982a) 235-239.
Miller, R.D., Prakash, L. and Prakash, S.: Genetic control of
excision of Saccharomyces cerevisiae interstrand DNA cross-
links induced by psoralen plus near-UV light. Mol. Cell. Biol.
2 (1982b) 939-948.
Prakash, L.: Repair of pyrimidine dimers in nuclear and mito-
chondrial DNA of yeast irradiated with low doses of ultra-
violet light. J. Mol. Biol. 98 (1975) 781-795.

126
Prakash, L.: Repair of pyrimidine dimers in radiation-sensitive mutants rad3, rad4, rad6, and rad9 of Saccharomyces cerevi-
Cue. Mutation Res. 45 (1977a) 13-20. Prakash, L.: Defective thymine dimer excision in radiation sensi-
tive mutants ran10 and rudlti of Saccharomyces cerevisiae.
Mol. Gen. Genet. 152 (1977b) 125-128.
Prakash, L. and Prakash, S.: Isolation and characterization of MMS sensitive mutants ofSaccharom.yce.7 cerevisiae. Genetics 86 (1977) 33-55.
Prakash, L. and P&cash, S.: Three additional genes involved in pyrimidine dimer removal in Succhar~mycescerev~~iae: ftAD7,
RADl4 and MMS19. Mol. Gen. Genet. 176 (1979) 351-359. Resnick, M.A. and Setlow, J.K.: Repair of pyrimidine dimer
damage induced in yeast by ultraviolet light. J. Bacterial. LO9 (1972) 979-986.
Reynolds, R.J.: Removal of pyrimidine dimers from Saccharu-
rnyce~ce~evj~~~~e nuclear DNA under nongrowth conditions as detected by a sensitive enzymatic assay. Mutation Res. 50 (1978) 43-56.
Reynolds, R.J. and Friedberg, E.C.: Molecular mechanisms of pyrimidine dimer excision in Succharomyces cerevisiae, I.
Studies with intact ceils and cell-free systems, in Generoso, W.M., Shelby, M.D. and de Serres, F.J. (Eds.), DNA Repair and Mutagenesis in Eukaryotes. Plenum, New York, 1980, pp. 121-139.
Reynolds, R.J. and Friedberg, EC.: Molecular mechanisms of pyrimidine dimer excision in Snccharomyces cerevisiae: in- cision of ultraviolet-irradiated deoxyribonucleic acid in vivo. J. Bacterial. 146 (1981) 692-704.
Sancar, A., Wharton, R.P., Seltzer, S., Kacinski, B.M., Clarke, N.D. and Rupp, W.D.: Identitication ofthe w-A gene product. J. Mol. Biol. 148 (1981a) 45-62.
Sancar, A., Clarke, N.D., Griswold, J., Kennedy, W.J. and Rupp, W.D.: Identification of the uvrB gene product. J. Mol. Biol. 148 (1981b)63-76.
Sancar, A., Kacinski, B.M., Mott, D.L. and Rupp, W.D.: Identifi- cation of the uvrcgene product. Proc. Natl. Acad. Sci. USA 78 (1981~) 5450-5454.
Sancar, A. and Rupp, W.D.: A novel repair enzyme: UVRABC excision nuclease of Escherichia cob cuts a DNA strand on both sides of the damaged region. Cell 33 (1983) 249-260.
Scherer, S. and Davis, R.W.: RepIacem~nt of chromosome seg- ments with altered DNA sequences constructed in vitro.
Proc. Natl. Acad. Sci. USA 76 (1979) 4951-4955. Setlow, R.B. and Carrier, W.L.: The disappearance of thymine
dimers from DNA: an error-correcting mechanism. Proc.
Natl. Acad. Sci. USA 51 (1964) 226-231. Shortle, D., Haber, J.E. and Botstein, D.: Lethal disruption ofthe
yeast actin gene by integrative DNA transformation. Science 217 (1982) 371-373.
Southern, E.M.: Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98 (1975) 503-517.
Unrau, P., Wheatcroft, R. and Cox, B.S.: The excision of pyrimidine dimers from DNA of ultraviolet irradiated yeast. Mol. Cen. Genet. 113 (1971) 359-362.
Waters, R. and Moustacchi, E.: The disappearance ofultraviolet- induced pyrimidine dimers from the nuclear DNA of expo- nential and stationary phase cells of Saccharomyces cerevisiae
following various post-irradiation treatments. Biochim. Biophys. Acta 353 (1974) 407-419.
Wilcox, D.R. and Prakash, L.: Incision and postincision steps of pyrimidine dimer removal in excision-defective mutants of Saccharomvces cerevisiae. J. Bacterial. 148 (1981) 618-623.
Communicated by S.R. Kushner


















