
Molecular and Genetic Basis for Strain-Dependent NK1 · James R. Carlyle,2* Aruz Mesci,* Belma...
15
of August 30, 2018. This information is current as Mouse NK Cells Strain-Dependent NK1.1 Alloreactivity of Molecular and Genetic Basis for Troke, Marie-France Proteau and Andrew P. Makrigiannis Belanger, Lee-Hwa Tai, Etienne Rousselle, Angela D. James R. Carlyle, Aruz Mesci, Belma Ljutic, Simon http://www.jimmunol.org/content/176/12/7511 doi: 10.4049/jimmunol.176.12.7511 2006; 176:7511-7524; ; J Immunol References http://www.jimmunol.org/content/176/12/7511.full#ref-list-1 , 22 of which you can access for free at: cites 39 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2006 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on August 30, 2018 http://www.jimmunol.org/ Downloaded from by guest on August 30, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of Molecular and Genetic Basis for Strain-Dependent NK1 · James R. Carlyle,2* Aruz Mesci,* Belma...

of August 30, 2018.This information is current as
Mouse NK CellsStrain-Dependent NK1.1 Alloreactivity of Molecular and Genetic Basis for
Troke, Marie-France Proteau and Andrew P. MakrigiannisBelanger, Lee-Hwa Tai, Etienne Rousselle, Angela D. James R. Carlyle, Aruz Mesci, Belma Ljutic, Simon
http://www.jimmunol.org/content/176/12/7511doi: 10.4049/jimmunol.176.12.7511
2006; 176:7511-7524; ;J Immunol
Referenceshttp://www.jimmunol.org/content/176/12/7511.full#ref-list-1
, 22 of which you can access for free at: cites 39 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2006 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Molecular and Genetic Basis for Strain-Dependent NK1.1Alloreactivity of Mouse NK Cells1
James R. Carlyle,2* Aruz Mesci,* Belma Ljutic,3* Simon Belanger,† Lee-Hwa Tai,†
Etienne Rousselle,† Angela D. Troke,† Marie-France Proteau,† and Andrew P. Makrigiannis2†
NK1.1 alloantigen expression can be used to define NK cells in certain mouse strains, such as B6 (NKR-P1C) and SJL (NKR-P1B).However, BALB/c NK cells do not react with the anti-NK1.1 mAb, PK136. To investigate the NK1.1� phenotype of BALB/c NKcells, we have undertaken NK1.1 epitope mapping and genomic analysis of the BALB/c Nkrp1 region. Bacterial artificial chro-mosome library analysis reveals that, unlike the Ly49 region, the Nkrp1-Ocil/Clr region displays limited genetic divergence betweenB6 and BALB/c mice. In fact, significant divergence is confined to the Nkrp1b and Nkrp1c genes. Strikingly, the B6 Nkrp1d geneappears to represent a divergent allele of the Nkrp1b gene in BALB/c mice and other strains. Importantly, BALB/c NK cellsexpress abundant and functional Nkrp1 transcripts, and the BALB/c NKR-P1B receptor functionally binds Ocil/Clr-b ligand.However, the BALB/c NKR-P1B/C sequences differ from those of the known NK1.1 alloantigens, and epitope mapping demon-strates that directed mutation of a single amino acid in the NKR-P1BBALB protein confers NK1.1 reactivity. Thus, PK136 mAbrecognizes, in part, a distal C-terminal epitope present in NKR-P1BSw/SJL and NKR-P1CB6, but absent in NKR-P1A/D/FB6 andNKR-P1B/CBALB. Allelic divergence of the Nkrp1b/c gene products and limited divergence of the BALB/c Nkrp1-Ocil/Clr regionexplain a longstanding confusion regarding the strain-specific NK1.1 alloantigen reactivity of mouse NK cells. The Journal ofImmunology, 2006, 176: 7511–7524.
N K cells are large granular lymphocytes capable of rec-ognizing and killing transformed, infected, Ab-coated,transplanted, and stressed cells (1, 2). Historically, NK
cells from selected mouse strains have been phenotypically definedusing NK-1 alloantigen-specific antisera (3) or the anti-NK1.1mAb, PK136 (4). The NK1.1 alloantigen is now known to identifyNK cells from CE, B6, NZB, C58, Ma/My, ST, SJL, FVB, andSwiss outbred mice, but not BALB/c, AKR, CBA, C3H, DBA, or129 mice (3–7). In fact, the NK1.1 alloantigen was originally iden-tified by immunizing BALB/c-background [(C3H � BALB)F1]host mice with donor CE splenocytes (3), and the same immuni-zation protocol was used to generate the PK136 mAb (4). Immu-nologically, this precludes the possibility that BALB/c (self) NKcells would react with anti-NK1.1 alloantibodies or PK136 mAb;thus, by definition, BALB/c NK cells are NK1.1� (3, 4, 8). Due toconventional use of the B6 mouse strain and initial molecular clon-ing of an NK1.1 Ag from B6 NK cells (9), anti-NK1.1 reactivity
has since become popularized as representing the product of theNkrp1c gene; however, the Nkrp1b gene product also reacts withPK136 mAb (5, 6).
Nonetheless, the underlying molecular basis for the lack ofNK1.1 reactivity of NK cells from BALB/c and other mousestrains remains an enigma. Previous studies have suggested thatBALB/c NK cells either lack or possess low-level expression ofthe Nkrp1 genes, as detected by Northern blotting (10); however,no genetic basis for a BALB/c defect in Nkrp1 expression has beenestablished. Thus, lack of NK1.1 reactivity could be due to dele-tion of Nkrp1 genes, defective gene expression, or allelic poly-morphism in BALB/c mice. Extreme variation in gene contentbetween the BALB/c and B6 haplotypes has been observed pre-viously for the related Ly49 gene family (11). This suggests thatother NK gene complex (NKC)4 regions, including the Nkrp1-Ocil/Clr region, also may be subject to rapid evolutionary diver-gence and/or polymorphism.
Importantly, since cognate NKR-P1 ligands have recently beenidentified (12–14), a BALB/c defect in NKR-P1 expression couldbe functionally significant for NK cell function and innate immu-nity. Moreover, while ligands for the stimulatory NKR-P1A/C re-ceptors remain elusive, ligands for the inhibitory NKR-P1B/D re-ceptors (12) and stimulatory NKR-P1F (13) receptors have beenidentified as products of the Ocil/Clr family of genes (15–17),which are intermingled with the Nkrp1 genes themselves in theNKC (18). Thus, determination of the gene content of the BALB/cNkrp1-Ocil/Clr region and the basis of the BALB/c defect inNK1.1 expression could have implications for the importance ofthe NKR-P1–Ocil/Clr system in self-nonself discrimination inmice and other species. Therefore, we have undertaken NK1.1
*Department of Immunology, University of Toronto, Sunnybrook and Women’s Re-search Institute, Toronto, ON, Canada; †Laboratory of Molecular Immunology, In-stitut de Recherches Cliniques de Montreal, Montreal, QC, Canada
Received for publication December 21, 2005. Accepted for publication March24, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants (to J.R.C. and A.P.M.) from the Canadian In-stitutes of Health Research (CIHR). J.R.C. is supported by a Career DevelopmentAward from the International Human Frontier Science Program Organization. A.M.was supported by a Life Sciences Award from the University of Toronto. A.P.M. issupported by a New Investigator Award from the CIHR.2 Address correspondence and reprint requests to Dr. James R. Carlyle, Department ofImmunology, University of Toronto, Sunnybrook and Women’s Research Institute,2075 Bayview Avenue (A-331), Toronto, ON, Canada M4N 3M5. E-mail address:[email protected] or Dr. Andrew P. Makrigiannis, Laboratory of MolecularImmunology, Institut de Recherches Cliniques de Montreal, 110 Avenue des PinsOuest, Montreal, QC, Canada H2W 1R7. E-mail address: [email protected] Current address: Immunology Research CA, Sanofi-Pasteur, 1755 Steeles AvenueWest, Toronto, ON, Canada M2R 3T4
4 Abbreviations used in this paper: NKC, NK gene complex; BAC, bacterial artificialchromosome; EC/UTR, extracellular/3�-untranslated region.
The Journal of Immunology
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$02.00
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

epitope mapping and genomic analysis of the BALB/c Nkrp1 re-gion to determine the functional significance of the NKR-P1 rec-ognition system in NK1.1� mouse strains.
In this study, we demonstrate that the BALB/c genome pos-sesses a full complement of Nkrp1 and Ocil/Clr genes, includingnovel family members present in both B6 and BALB/c mice. Fur-thermore, BALB/c NK cells possess normal Nkrp1 expression rel-ative to B6 NK cells, and the BALB/c NKR-P1B receptor func-tionally binds cognate Ocil/Clr-b ligand. Absent NK1.1 reactivityof BALB/c NK cells can be explained by Nkrp1 allelic divergence,specifically a single amino acid substitution (S191T) present in theBALB/c NKR-P1B/C receptors. In fact, divergence between theB6 and BALB/c Nkrp1-Ocil/Clr regions appears to be localized tothe Nkrp1b/c genes, and the B6 Nkrp1d gene appears to be anallele of the Nkrp1b gene found in BALB/c and other mousestrains. These results confirm the importance of the NKR-P1–Ocil/Clr recognition system across strain boundaries, and suggest thatthis mode of self-nonself discrimination is conserved in otherNK1.1� mouse strains, as well as other species.
Materials and MethodsMice
Mice were purchased from the Charles River Laboratories National CancerInstitute–Frederick Animal Production Area (Frederick, MD), and/or main-tained in our own animal facilities.
Cells
BWZ.36 cells (19) were obtained from Dr. N. Shastri (University of Cal-ifornia, Berkeley, CA). The 293T cells were obtained from Dr. D. Raulet(University of California, Berkeley, CA). Cells were grown in completeDMEM-HG (high glucose, 10% FCS). All cells were maintained at 37°Cin a humidified 5% CO2 atmosphere.
Expression constructs and NKR-P1 mutants
Mouse Nkrp1b (National Institutes of Health–Swiss–Sw) and Nkrp1a/c/d(C57BL/6–B6) constructs were generated previously (5). Mouse Nkrp1e/fcDNAs were amplified from NK libraries using the following primers:P1E-F, ATGGACACAGAAAGGATCTACCTC; P1E-R, TCAGGAGTCATGAAATATGGTTTC; and P1F-F, ATGGACACATCAAAGGTCCATG; P1F-R, TCAGACATGTATCAGGGTCTTTTG. Rat Nkrp1a/bcDNA clones (20) were generously provided by Dr. R. G. Miller (OntarioCancer Institute, Toronto, ON). Site-directed mutagenesis was performedby the method of gene splicing by overlap extension of DNA ends, asdescribed previously (12). Briefly, known Nkrp1 sequences were amplifiedby PCR using specific 5� or 3� outside primers and complementary internalprimers with specified point mutations; the resulting products were thenmixed and amplified using the outside primers alone to obtain the full-length mutant cDNA. All PCR products were directly cloned intopcDNA3.1/V5/HIS/TOPO (Invitrogen Life Technologies), and constructswere sequenced to confirm their identities. The resulting constructs werecotransfected into 293T cells using Effectene reagent (Qiagen) with apCMV-GFP-nuclear localization sequence reporter, or subcloned into thepMSCV2.2-CMV-IRES-GFP (pMCIG) retroviral vector (12) before trans-fection into 293T cells. Transfection efficiencies were monitored by GFPexpression, and transfection results are shown gated on GFP� cells for allcontstructs tested.
Flow cytometry and cell sorting
Cells were stained as described previously (21). Stained cells were ana-lyzed using a FACSCalibur flow cytometer (BD Biosciences) or weresorted on a FACSDiVa (BD Biosciences). Sorted cells were �99% pure,as determined by postsort analysis.
Receptor fusions, retroviral infections, and BWZ assay
A pMSCV2.2-CMV-IRES-GFP (pMCIG) retroviral vector (12) was mod-ified to include a CD3�/NKRP1B cassette (intracellular region of CD3�,membrane proximal and transmembrane regions of NKR-P1B). The CD3�/NKRP1B fusion cassette was generated by gene SOEing using the follow-ing primers: SALCD3Z-F, GTCGACATGAGAGCAAAATTCAGCAGGAGTG;CD3ZP1B-R,GCGAGGGCACCGACAGCGAGGGGCCAGGGTCTGC; CD3ZP1B-F, ACCCTGGCCCCTCGCTGTCGGTGCCCTCGCT
GGCATC; XHOP1BTM-R, CTCGAGTGATGATTTTTGTACTGATAG.The resulting product was TOPO TA-cloned (Invitrogen Life Technolo-gies) and cut with SalI and XhoI (New England Biolabs), then ligated intopMCIG that had been linearized with XhoI and treated with CIP. cDNAinserts were subcloned into the modified cassette vector, pMC3BIG, usingXhoI and NotI sites. BWZ.36 cells were infected with retroviral superna-tants (24–48 h) from transient triple-transfected 293T cells, as describedpreviously (12), then sorted at day 3–4 following infection. Stable BWZtransductants were analyzed using plate-bound mAb or cell mixtures, asdescribed previously (12).
cDNA libraries
BALB/c DX5� NK cell (22) and BALB/c-congenic C.B-17/SCID LAK(23) cDNA libraries were reported previously. The NKC of C.B-17/SCID(BALB/c.C57BL/Ka-Igh-1b scid/scid) mice has been confirmed to beBALB/c in gene content (Ref. 24 and data not shown). A (B6 � BALB/c)F1 LAK library was generously provided by Dr. R. G. Miller (OntarioCancer Institute, Toronto, ON). A B6 DX5� NK cell cDNA library wasprovided by Dr. H. Arase (Osaka University, Japan).
Evaluation of BAC gene content
BAC clones from the CHORI-28 BALB/c library (BACPAC Resources)containing Nkrp1 or Ocil/Clr genes were identified and size-estimated asdescribed previously (25). The gene content of BACs was determined byPCR and sequencing of products. Briefly, �10 ng of BAC DNA was sub-jected to PCR (94°C 30 s, 57°C 30 s, 72°C 1 min, 30 cycles) with primerscapable of amplifying known and predicted BALB/c fragments of Nkrp1and Ocil/Clr gene sequences (primers are shown in Table I). PCR productswere separated on 1% agarose gels and visualized with ethidium bromide.PCR products of each size were cloned by TOPO TA-cloning into pCR2.1(Invitrogen Life Technologies) and sequenced in-house using T7 and M13reverse primers. All BAC PCR amplifications were performed using Taqpolymerase on enriched, high-copy BAC DNA. Importantly, no mutationswere detected between distinct PCR (i.e., at least two clones of each productfrom separate PCR were sequenced). Partial exon sequence allowed the iden-tification of the Nkrp1 or Ocil/Clr gene that was amplified from each BACclone. The partial gene sequences from the BALB/c BACs were deposited inGenBank under the following accession numbers: Nkrp1a (exons 3–5,DQ143102); Nkrp1b (exons 3–5, DQ143103); Nkrp1c (exon 1, DQ143106);Nkrp1c (exons 3–5, DQ336140); Nkrp1e (exon 2, DQ143104); Nkrp1f (exons3–5, DQ143105); Nkrp1g (exons 3–5, DQ336141); Clr-a (exons 3–4,DQ143108); Clr-b (exons 3–4, DQ143111); Clr-c (3� end, DQ143114); Clr-d(exons 4–5, DQ143107); Clr-e (exons 4–5, DQ143109); Clr-f (exons 3–4,DQ143110); Clr-g (exons 3–4, DQ143112); and Clr-h (exons 3–4,DQ143113). Sequence-confirmed gene fragments were used in Southern blotanalyses of BAC DNA to confirm Nkrp1-Ocil/Clr gene content as describedpreviously (25).
RT-PCR and cDNA library PCR
Full-length cDNA clones were obtained using fresh cDNA isolated fromd6 BALB/c LAK cells (plastic-adherent IL-2 lymphokine-activated killercells). RNA was isolated using TRIzol (Invitrogen Life Technologies) andreverse-transcribed into cDNA with SuperScript first-strand cDNA synthe-sis kit (Invitrogen Life Technologies). PCR conditions, visualization, clon-ing, and sequencing were performed as outlined above. Library cDNAPCR was performed using primers specific for either the full-lengthNkrp1a/b/c coding regions (5) or the corresponding extracellular/3�-un-translated regions (21), under limiting cycle conditions (25 cycles PCRwith 50 ng cDNA library, representing �10–20 ng cDNA equivalents).For mammalian expression studies, PCR products were cloned directly intopcDNA3.1/V5/HIS/TOPO (Invitrogen Life Technologies) and sequencedto confirm identity and orientation. All cDNA PCR amplifications wereperformed using Expand High Fidelity enzyme (Roche Diagnostics), andsequences were determined from multiple clones from at least two separatePCR. Sequences were deposited in GenBank under the following accessionnumbers: Nkrp1aBALB, DQ237927; Nkrp1bBALB, DQ237928; Nkrp1cBALB,DQ237929; Nkrp1eBALB, DQ237930; and Nkrp1f BALB, DQ237931). Phy-logenetic analysis was performed as described previously (26).
BAC end-sequence characterization
BAC DNA (1 �g) was sequenced in-house with T7 and SP6 primers.Based on end-sequence results, primers were designed to PCR-amplify therespective BAC ends. Products were cloned using a TOPO-pCR2.1 kit(Invitrogen) and sequences were confirmed. Cloned end-sequence frag-ments were radioactively labeled and used as probes in Southern blots ofEcoRI-digested BAC DNA.
7512 THE BALB/c Nkrp1-Ocil/Clr GENE CLUSTER
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

ResultsDivergence of the BALB/c and B6 Nkrp1-Ocil/Clr regions
One simple explanation for the NK1.1� phenotype of BALB/c NKcells is that the two genes known to confer NK1.1 reactivity,Nkrp1b and Nkrp1c, like Ly49d and Ly49h, may not be present inthe BALB/c genome (11). BALB/c NK cells have been reported tobe deficient in Nkrp1 transcripts, as detected by Northern blotting(10). Thus, lack of NK1.1 reactivity of BALB/c NK cells could bedue to deletion of Nkrp1 genes or defective gene expression.
To investigate this possibility, we constructed a physical map ofthe BALB/c Nkrp1-Ocil/Clr region. To this end, a BALB/cgenomic library, CHORI-28, was probed with a mixture of knownNkrp1 and Ocil/Clr cDNAs. Positive BAC clones were isolatedand further screened by PCR using gene-specific primers for theknown Nkrp1 and Ocil/Clr genes (Table I). PCR products weresequenced to confirm gene identity, and Southern blots of EcoRI-digested BACs were probed to confirm the presence of specificgenes in each BAC clone (Table II). In some instances, the primerpairs cross-amplified closely related sequences (see Table I), re-sulting in the discovery of new gene fragments. One such novelsequence, cross-amplified using Clr-b primers, corresponds to anexon-3/intron-3/exon-4 (e3-i3-e4) genomic fragment resembling anear-identical (�99%) match to a sequence available from the lat-est B6 genome assembly (designated Clr-h; GenBank accessionno. XM487965). The matching region in the B6 genome does not
correspond to the location of any other known Clr gene. This sug-gests that the Clr-h sequence represents a new Clr family memberconserved in the B6 and BALB/c genomes. In addition, a new genewas detected in the region between Clr-f and Nkrp1c based onSouthern hybridization using an Nkrp1f probe. This sequence alsoresembles a near-identical (�99%) match in the B6 genome (des-ignated Nkrp1g; GenBank accession no. XM355818) that does notcorrespond to the location of any other known Nkrp1 gene. Thus,two new genes, Clr-h and Nkrp1g, are conserved in the B6 andBALB/c genomes.
With the exception of the Nkrp1c and Nkrp1d primers, allprimer pairs amplified the BALB/c allele of the intended gene. Asa positive control, all B6-derived primers amplified the intendedtarget sequence when B6 genomic DNA was used as a template.Sequencing of the BALB/c Nkrp1a/e/f and Clr-a/b/c/d/e/f/g genefragments showed that all were highly related to their B6 coun-terparts (�98% identity). Three different primer pairs specific forNkrp1cB6 gave variable results when BALB/c BACs were used astemplates (see Table I): one set failed to produce any product; asecond set cross-amplified Nkrp1e; and a third set cross-amplifiedboth Nkrp1e and an exon-1 sequence identical with a knownBALB/c Nkrp1 gene fragment, designated as gene-40 (GenBankaccession no. X64720) (10). Although this latter fragment resem-bles Nkrp1a exon 1 in coding sequence, it was distinct from theNkrp1aBALB sequence that we identified; therefore, because the
Table I. Primers used to amplify Nkrp1-Ocil/Clr-related genes from BAC clones and cDNA
Primer Specificity Sense Primer Antisense Primer
BALB/cGene(s)Detected
Nkrp1aB6 CGTTAATCTCTCTCTGTGTTTTCTCATTTAT(intron 2)
TGTCAGTGTTAGAGCCCTAC (intron 5) Nkrp1a
Nkrp1bSJL GTCAAGTCCCTCCATCTACA (exon 2) AATGGGGATGGGATTCACAG (exon 7) Nkrp1b(cDNA)
Nkrp1cB6 TTAGCGTTAATCCTCTCTG (intron 2) CAATTAGCCCCAAGGGGAAT (intron 5) aNPNkrp1cB6 AATCAGTGGGTGTGGGAACA (exon 1) TGCTCAAGTTTCACACTGAC (exon 1) Nkrp1e,
Nkrp1cNkrp1cB6 CAGCCTGCTTTATCAAAGCAAACAG
(intron 1)TCCAGGCCCAAGGGTCCCA (intron 2) Nkrp1e
Nkrp1cBALB TGCGAGTCCTAATACAAAAACCATCAAG(exon 3)
CCTTAGTCCAATCCAAAATGAATTGTAA(exon 5)
Nkrp1c(cDNA)
Nkrp1dB6 CTTCTGCCTGTGTTTTCTTATC(intron 2)
ACTATTAGCCCCAAGGGGAC (intron 5) Nkrp1b
Nkrp1fB6 CCTGAAGTGAGAACTAGAATATCC(intron 2)
TTCAGCTGAACCTTTCAAGA (intron 5) Nkrp1f
Nkrp1gB6/BALB CACTCCACCATTCATTTCAG (exon 3) CCCTCTCAAGAATTCATAAGG (exon 5) aNP (cDNA)Clr-aB6 CTGAAGTTTGGATGGCTAGA (intron 2) GCATCTCTTACAGATACAATGC
(intron 4)Clr-a
Clr-bB6 AAGACCTGCTTGCCATGATC (intron 2) TGACACTGCCTTTCATCTCC (intron 4) Clr-b, Clr-h,Clr-d
Clr-bB6 ACTCAGCTCCTCAGCTCTGA (exon 1) GGCTAAAAAGCGTCTCTTGG (exon 5) Clr-b(cDNA)
Clr-cB6 TGTAATGGGATGTGTTAACCTATTAGG(3� UTR)
GTACTTCATCAACCTTCATTTGTTATGC(3� UTR)
Clr-c
Clr-dB6 TCCTAATGAGATACAAGGCA (exon 4) CAGGAACAGGAGTTGGGCAG (exon 5) Clr-dClr-eB6 GGCCTGCACAGAGTCGTCAA (exon 4) GACAGGACACTAGCATTAGT (exon 5) Clr-eClr-fB6 GTGAGATTTGGATGGCTAGG (intron 2) GGCTTCACATGATATAGTCA (intron 4) Clr-fClr-gB6 TCAGGAGGGTGTGCCAAAAA (intron 2) CTCATAGCCACGACACACAT (intron 4) Clr-gClr-hB6 CTCCATCCTAGTGGGTACAA (exon 1) TGCTACAAATCCACTTCCTT (exon 5) aNP (cDNA)Cd69B6 ACAAATGTCCTTTCTGTCCAC
(intron 2)AAAGTGTCAGCCTAGAGAGG (intron 4) Cd69
BAC 358o7SP6-end
CACCTCTCTTGTTTAGAAGAGCCCT CTGAGGAGAACTTACATGATGAA aNA
BAC 387p19SP6-end
TACAGCCTCTGCTTTCCTTC CTGAAGCAGCGATCAGAAAG aNA
a No product detected for BALB/c BAC DNA cited in this study. A positive control (B6 genomic DNA) was included in every BAC PCR and always resulted in theamplification of the correct-sized product. In the case of Nkrp1c and Nkrp1d, the B6-derived product was confirmed by sequencing to verify the specificity of the primers. NP,No product; NA, not applicable.
7513The Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

gene-40 designation is synonymous with the Nkrp1c transcript inB6 mice, this sequence was designated Nkrp1cBALB. Primers spe-cific for Nkrp1d amplified a product with a coding sequence verysimilar to that of the known Nkrp1b gene from Sw and SJL mice.This was surprising, considering that the Nkrp1b gene product isresponsible for NK1.1 reactivity in Sw, SJL, and other mousestrains (5–7, 12).
In contrast with genomic PCR, PCR of cDNA libraries derivedfrom BALB/c NK cells revealed the existence of an Nkrp1c-liketranscript; however, only a partial extracellular domain fragmentcould be amplified, due to the location of the primers (data notshown). Southern analysis using this cDNA product, new frag-ments derived from the cDNA, and the previous genomic
Nkrp1cBALB exon-1 fragment as probes (Fig. 1 and data notshown) revealed that the Nkrp1c-like gene resided centromeric toNkrp1bBALB, approximating the location of Nkrp1c relative toNkrp1d in the B6 genome (27). Hybridization with Nkrp1bBALB
and Nkrp1cBALB probes showed that several BACs contained thesegenes (Fig. 1). Similarly, hybridization of the same BALB/c BACswith Nkrp1dB6 and Nkrp1cB6 probes revealed the same bands, al-beit with weaker intensity (Fig. 1).
A physical map for the BALB/c Nkrp1-Ocil/Clr gene cluster
A relative gene order of Nkrp1a, Clr-h, Clr-f, Nkrp1g, Nkrp1c,Nkrp1b, Clr-g, Clr-d, Clr-e, Clr-c, Nkrp1f, Clr-a, Nkrp1e, Clr-b,and Cd69 was revealed after assembling the BACs in order of gene
Table II. Gene content of BALB/c BAC clonesa
Gene/Marker Nkrp1a Clr-h Clr-f Nkrp1g 358o7-end 387p19-end Nkrp1c Nkrp1c Nkrp1b Clr-g Clr-d Clr-e Clr-c Nkrp1f Clr-a Nkrp1e Clr-b Cd69
Probe RegionExons3–5
Exons3–4
Exons3–4
Exons3–5 SP6 SP6
Exons3–5 Exon 1
Exons3–5
Exons3–4
Exons4–5
Exons4–5 3� End
Exons3–5
Exons3–4 cDNA
Exons3–4 Exon 2
Fragments(kb) 10.5 8 4.5 5.8 2.6 2.0
5.3/4.6 9.0 3.3 5.8 5.75 3.0 1.8
3.9/3.25 7.1
4/0.7/11.5 10.5 PCR
368i24 X299i20 X X X517h18 X X X331e2 X X X X X457k23 X X X X X387p19 X X X X X X296i7 X X X X358o7 X X X X X X493d24 X X X X521c8 X X X X501d7 X X X X X451l24 X X X X X X444p13 X X X X299f24 X X X X349l18 X X X X371k12 X X X X X510j5 X X X306n15 X X X334k17 X X X370a24 X X X445a21 X X445k14 X X
a BAC clones containing Nkrp1 and Ocil/Clr-related genes were initially identified from high-density spotted BAC library filters. Positive BACs were screened by PCR withgene-specific primers. One of each kind of product was sequenced to confirm gene identity. These PCR gene fragments then served as probes in Southern blots of EcoRI-digestedBAC DNA. In some cases, cDNAs were used to identify positive BACs.
FIGURE 1. Identification of BALB/c genomic clones containing Nkrp1b/c. BALB/c BAC clones previously found to contain Nkrp1 sequences (by PCR)were confirmed in their gene content by Southern blotting. BAC DNA was digested with EcoRI, gel-separated, and transferred. The blot was consecutivelyhybridized with the indicated probes. For simplicity, only BACs positive for Nkrp1b or Nkrp1c are shown. Probes Nkrp1cB6 exons 3–5, Nkrp1dB6 exons3–5, Nkrp1cBA exon 1, and Nkrp1bBA exons 3–5 are derived from BAC or genomic DNA. Probes Nkrp1cBA exons 3–5 and Nkrp1cBA exon 5–6 are derivedfrom cDNA. BA, BALB/c; ex, exon.
7514 THE BALB/c Nkrp1-Ocil/Clr GENE CLUSTER
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

content (Table II). There are some ambiguities in this assembly.Notably, the order of Clr-h and Clr-f in the BALB/c genome isunknown because of their identification on shared BACs; however,in the B6 genomic database, Clr-h lies centromeric to Clr-f and istranscribed in the opposite orientation. Similarly, Clr-c and Nkrp1falso were found on the same BACs, but their order is known in theB6 haplotype, with Clr-c lying centromeric to Nkrp1f.
To generate a physical map of the approximate relative locationof each gene on the chromosome, the size of each BAC clone wasascertained by pulsed-field gel electrophoresis (Fig. 2 and data notshown). Using the gene content and size of each BAC, the ap-proximate location of each gene on chromosome 6 of BALB/cmice was predicted (Fig. 2). The BALB/c Nkrp1-Ocil/Clr region ispredicted to be �640 kb in length (Nkrp1a exon 3 to Clr-b exon4), with a possible range from 575 to 700 kb (Fig. 2). In previousgene-mapping studies, the entire B6 Nkrp1-Ocil/Clr cluster waspredicted to be �700 kb in length (16, 27); however, the exactlength is unknown, as the B6 genome assembly for this regioncontains several gaps.
Determination of Nkrp1-Ocil/Clr haplotypes in different inbredmouse strains
Direct comparison of the gene content between BALB/c and B6mice shows that at least 13 genes are conserved, including
Nkrp1c and the novel, but shared, Clr-h and Nkrp1g genes (Fig.3). The only BALB/c-specific gene was Nkrp1b, but this genecorresponded to the approximate location of Nkrp1d in the B6genome (Fig. 2), and both the Nkrp1bBALB and Nkrp1dB6
probes hybridized to the same genes (Fig. 1). Thus, it is likelythat the two designations actually represent different alleles ofthe same gene, especially since their transmembrane and cyto-plasmic coding sequences are identical. Therefore, like the B6and BALB/c Ly49 gene clusters, the Nkrp1-Ocil/Clr regionsrepresented in these mice possess shared genes and have pre-sumably diverged from an ancestral haplotype. However, unlikethe Ly49 gene repertoire, that of the Nkrp1-Ocil/Clr genes ap-pears to be highly conserved, perhaps with the exception of aseemingly directed divergence of Nkrp1b and Nkrp1c. Thesesimilarities and differences are apparent in Southern analyses ofgenomic DNA (Fig. 4A). The RFLP patterns generated by theNkrp1f and Clr-f probes are similar in the two mouse strains,but the banding patterns resulting from the Nkrp1cB6 probe aredistinct (Fig. 4A). Hybridization of the Nkrp1cB6 and Clr-fprobes was conducted at lower stringency than that of Nkrp1fand confirms that the number of Nkrp1 and Ocil/Clr genes, asevidenced by cross-hybridizing genes, is similar in these twostrains.
FIGURE 2. BAC contig overlap and construction of a physical map for the BALB/c Nkrp1-Ocil/Clr gene cluster. The BAC gene content from TableII and BAC sizing data (pulsed-field gel electrophoresis; data not shown) were integrated to produce a map of the relative location of all known BALB/cNkrp1-Ocil/Clr genes. BACs are represented by horizontal lines, with the name and size of each given on the left side. F, the start of exon 4 of the indicatedgenes. The spacing between genes was based on the average of possible maximum and minimum sizes imposed by the BAC size and gene content. Thescale bar (top) is demarcated in kilobases. The markers 387p19-SP6� and 358o7-SP6�, represented by empty circles, are the end sequences of BACs 387p19and 358o7, respectively. After cloning, these BAC end sequence fragments were used as probes to show that BACs 387p19 and 358o7 overlap, despitenot sharing any Nkrp1-Ocil/Clr genes.
7515The Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The only other well-characterized Ly49 strain haplotype, repre-sented by the 129 group of inbred mice, is a third distinct haplo-type more complex than that of either B6 or BALB/c mice (26). Todetermine whether the 129 strain Nkrp1-Ocil/Clr cluster is similaror divergent, genomic Southern blot analyses were performed (Fig.4). Hybridization with Nkrp1bBALB and Nkrp1cB6 probes revealedthat the 129-derived genomic DNA gave an identical RFLP patternto that of BALB/c, suggesting the 129 and BALB/c strains share asimilar Nkrp1-Ocil/Clr haplotype, in contrast with their Ly49 re-gions (Fig. 4B). In a third experiment, hybridization with theNkrp1bBALB probe suggests that BALB/c, 129/Sv, 129S1, andFVB inbred mice have a similar Nkrp1 haplotype, one that is dis-tinct from that of B6 mice (Fig. 4C). Hybridization with a Clr-hBALB probe shows that this gene is present in all five mousestrains (Fig. 4C). Interestingly, the same probe cross-hybridized toone band at 5.8 kb in the BALB/c and 129-group mice (most likelyClr-g and Clr-d, see Table II), but in B6 and FVB mice, a doublet(5.9 and 6.0 kb) was detected (Fig. 4C). Collectively, these datasuggest that the commonly used inbred mouse strains have one ofat least three distinct Nkrp1-Ocil/Clr haplotypes, represented byB6 (group 1), BALB/c and the 129-group (group 2), and FVB(group 3). These findings extend earlier work showing that theBALB/c and 129-strain Nkrp1 haplotypes belong to one of fourRFLP groups, based on previous analysis of the Nkrp1a locus (28).In any case, BALB/c mice are not deficient in Nkrp1 gene content,so the lack of NK1.1 reactivity of BALB/c NK cells is not due todeletion of either the Nkrp1b or Nkrp1c genes.
BALB/c NK cells express abundant Nkrp1b and Nkrp1ctranscripts
To obtain cDNA clones of the Nkrp1 genes from BALB/c mice,RT-PCR was performed on fresh BALB/c A-LAK cDNA andBALB/c NK cell cDNA libraries using multiple primer sets. Thisapproach yielded full-length or near full-length cDNA clones forNkrp1aBALB, Nkrp1bBALB, Nkrp1cBALB, Nkrp1eBALB, andNkrp1fBALB (Fig. 5 and data not shown). This was unexpected,given that BALB/c NK cells were reported previously to lack de-tectable Nkrp1 expression in Northern blotting experiments (10).To determine whether Nkrp1b/c expression was specifically defi-cient in BALB/c mice, RT-PCR was applied to both BALB/c andB6 NK cell cDNA libraries using limited amounts of cDNA andamplification cycles. The Nkrp1 primer sets used have been pre-
viously reported, one set corresponding to the extracellular/3�-un-translated region (EC/UTR) (21), a second set corresponding to thefull-length coding sequence (FL/CDS) regions (5). As shown inFig. 5, Nkrp1 transcripts were easily detected in both strains. How-ever, while the EC/UTR primers yielded a significant signal forNkrp1cBALB, a FL/CDS Nkrp1cBALB signal could not be detected.This has been observed before for Sw strain NK cells (5, 21), andsuggests that the primer sets used may be divergent compared withthe Nkrp1cBALB sequence. Interestingly, note that the Nkrp1b FL/CDS primer set amplifies Nkrp1bSw (5), Nkrp1bSJL (6), andNkrp1bBALB (Fig. 5; see below), whereas it also cross-amplifiesNkrp1dB6 (5, 6). Therefore, to confirm the cDNA identities, anddetermine the discrepancy of the Nkrp1cBALB results, all PCRproducts were sequenced.
Sequencing results revealed that the EC/UTR Nkrp1cBALB PCRproduct was most highly related to the known Nkrp1cB6 sequencein overall BLAST score, followed by Nkrp1dB6, and Nkrp1bSw/SJL.Moreover, it shared 196 bp of nucleotide identity to the gene-40partial sequence derived from BALB/c LAK cells (GenBank ac-cession no. X64720) (10). As mentioned previously, the gene-40sequence is also identical over 89 bp to the genomic Nkrp1cBALB
exon-1 probe, confirming its Nkrp1cBALB designation. However,this sequence closely resembles Nkrp1a in its 5� region, whichcould confound attempts to amplify the full-length cDNA by PCRusing the Nkrp1c 5� primer. Therefore, to take advantage of thesequence similarity, we used PCR amplification of cDNA librariesusing a P1A-5�/P1C-3� combination of FL/CDS primers.
Interestingly, full-length transcripts for Nkrp1cBALB were de-tected equally in BALB/c, B6, and F1 LAK libraries by thismethod (Fig. 5B). Moreover, primers spanning exons 3–5 werecapable of specifically detecting Nkrp1cBALB transcripts in freshBALB/c but not B6 NK cells (Fig. 5C). Sequencing confirmed thatthe Nkrp1cBALB EC/UTR product, the P1A-5�/P1C-3� product, theNkrp1cBALB exon 3–5 product, and the gene-40 sequence (Gen-Bank accession no. X64720) (10) were all identical with one an-other. These results confirm the existence of a full-length codingNkrp1c cDNA in BALB/c NK cells. However, comparison of theNkrp1cBALB cDNA with the published Nkrp1cB6 cDNA revealsthat they differ by 20 nonsynonymous substitutions in the extra-cellular coding region alone (Fig. 6A). Such divergent sequenceswould be expected to confound attempts using Northern blotting tocompare expression of Nkrp1c transcripts between the two mouse
FIGURE 3. Comparison of gene content in the B6 and BALB Nkrp1-Ocil/Clr regions. The mouse NKC is depicted at the top, with major groups of NKcell receptor genes shown. Note that additional genes are present but are not shown for simplicity. Immediately below is an expanded view of the knowngenes in the B6 Nkrp1-Ocil/Clr region, and on the bottom, a diagram of the BALB/c region. Rectangles represent genes, and the arrows indicatetranscriptional orientation.
7516 THE BALB/c Nkrp1-Ocil/Clr GENE CLUSTER
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

strains (10). Notably, the similarity of the Nkrp1cBALB 5� region toNkrp1aB6 is reminiscent of Ly49o129/C57L, which resemblesLy49aB6 upstream and Ly49dB6 downstream (29, 30).
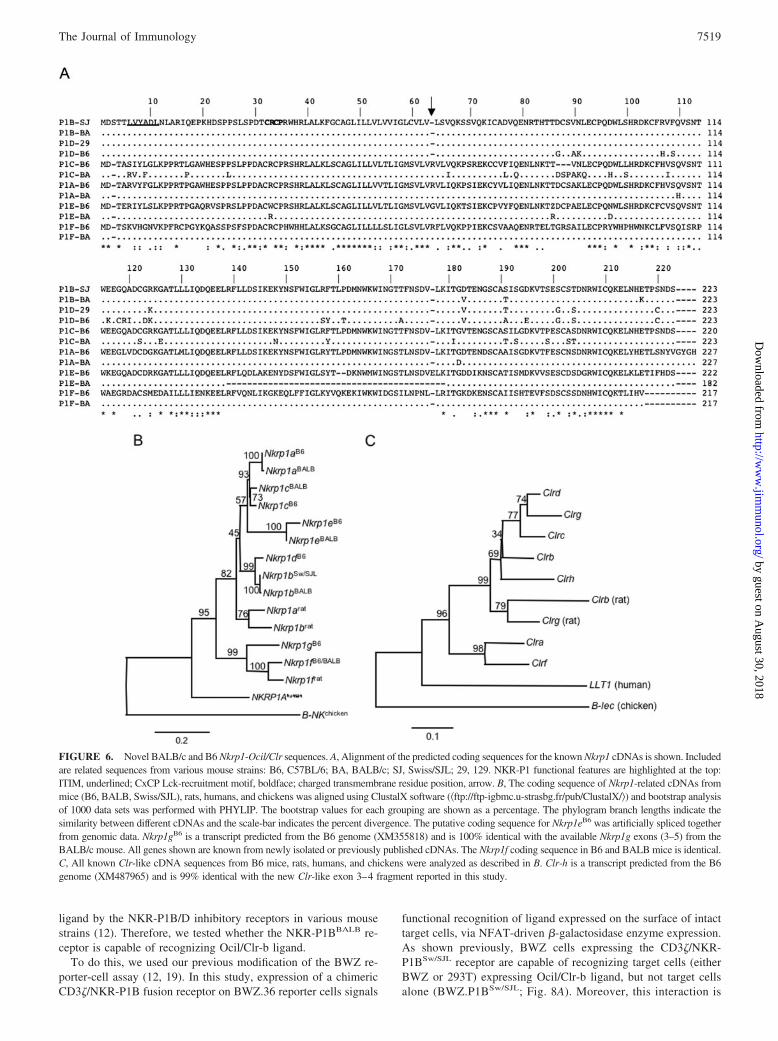
In contrast with Nkrp1cBALB, the Nkrp1aBALB sequence differsfrom its B6 counterpart by only two nonsynonymous substitutionsin its extracellular coding region, and the Nkrp1bBALB cDNA se-quence is �99% identical with Nkrp1bSw/SJL, differing by onlythree nonsynonymous substitutions in its extracellular coding re-gion (Fig. 6A). The ITIM motif in the Nkrp1bBALB coding se-quence is intact, and it lacks a charged residue in its transmem-brane region, so this cDNA is predicted to code for a functionalinhibitory transmembrane receptor. A near full-length (exon 5 de-leted) in-frame cDNA clone of Nkrp1eBALB was also isolated bythis method (Fig. 6A); in contrast, Nkrp1e is only known fromsingle genomic exons in B6 mice (27). The Nkrp1fBALB cDNAsequence is identical with the published Nkrp1f B6 sequence(Fig. 6A). Three independent primer sets failed to amplify anNkrp1gB6/BALB cDNA (Fig. 5C and data not shown), so this genemay represent a pseudogene, at least in the strains tested.
When these Nkrp1 sequences (and the B6 Ocil/Clr sequences)are compared over their coding regions, the Nkrp1bBALB andNkrp1bSw/SJL sequences are closely associated with each other and
with Nkrp1dB6 (Fig. 6B). The Nkrp1cBALB sequence is mostclosely related to the allelic sequence, Nkrp1cB6. All otherBALB/c Nkrp1 genes are closely related to their B6 alleles. Thenovel gene, Clr-h, groups with the Clr-b/c/d/g clade (Fig. 6C). AllClr genes are shown without allele designation, as the BALB/cfull-length coding sequences were not available for directcomparison.
Thus, while our results using RT-PCR differ from previousNorthern blot analysis of Nkrp1 transcript expression in BALB/cmice, our sequencing analysis confirms and extends previous find-ings, where our Nkrp1a/b/cBALB sequences correspond to theBALB/c partial sequences of gene 2 (GenBank accession no.X64723), gene 34 (GenBank accession no. X64719), and gene 40(GenBank accession no. X64720), respectively (10). Thus,BALB/c mice are not deficient in Nkrp1 expression, they simplypossess divergent Nkrp1 sequences compared with those ofB6 mice.
BALB/c NKR-P1B/C proteins lack NK1.1 reactivity due toallelic divergence
Our results demonstrate that BALB/c mice possess a full Nkrp1gene content and normal Nkrp1b/c transcript expression in NK/
FIGURE 4. Inference of Nkrp1-Ocil/Clr haplotypes in different inbred mouse strains by RFLP analysis. (A–C) Genomic DNA samples from theindicated mouse strains were digested with EcoRI, separated, and transferred to nylon membranes. The membranes were probed with the indicated probescontaining exons 3–5. In A, DNA samples from three different animals of each strain are shown. Hybridization with the Nkrp1cB6 and Clr-fBALB probeswas performed at less stringent conditions so that all Nkrp1- and Ocil/Clr-related fragments could be detected.
7517The Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

LAK cells when compared with NK1.1� mouse strains, suggestingthat the basis of NK1.1 alloreactivity between these mouse strainsmay be due to divergence in NKR-P1 protein sequences. As shownin Fig. 7, NK1.1 reactivity is specific to NKR-P1BSw/SJL andNKR-P1CB6 among known NKR-P1 proteins from both mice andrats. However, sequence alignment of these proteins does not di-rectly reveal an amino acid sequence unique to NK1.1� vsNK1.1� isoforms (Fig. 7A and data not shown). Therefore, welimited our analysis to the mouse NKR-P1 isoforms, and furtherfocused on differences between the mouse NKR-P1BSw/SJL andNKR-P1BBALB sequences, because the two differ in NK1.1 reac-tivity, but only differ in sequence by three amino acids (see Fig.7A). Of these three substitutions in the NKR-P1BSw/SJL sequence,two were expected to be nonconservative, resulting in charge al-terations in the NKR-P1BBALB isoform (D183V, E217K). How-ever, the D183V substitution also is present in the NK1.1� NKR-P1CB6 isoform, while the E217 amino acid is intact in the NK1.1�
NKR-P1CBALB isoform. This left a seemingly conservative S191Tsubstitution as a candidate determinant for NK1.1 epitope reactiv-ity (Fig. 7A). Interestingly, both NK1.1-reactive isoforms (NKR-P1BSw/SJL, NKR-P1CB6) possess an S residue at this position,whereas all other mouse NKR-P1 sequences have substitutions,including NKR-P1BBALB (T), NKR-P1CBALB (T), NKR-P1DB6
(A), NKR-P1D129 (T), NKR-P1AB6/BALB (A), NKR-P1EB6/BALB
(T), NKR-P1FB6/BALB (I), and the predicted NKR-P1GB6 (L).Therefore, to test the significance of this residue in conferringNK1.1 reactivity, the NKR-P1BBALB sequence was altered by site-directed mutagenesis.
As shown in Fig. 7C, the T191S mutation conferred NK1.1 re-activity to the NKR-P1BBALB protein, whereas a K217E controlmutation, found natively in the NKR-P1CBALB sequence, did notalter NK1.1 reactivity. This demonstrates that a single amino acidsubstitution present in both the BALB/c NKR-P1B and NKR-P1Csequences is sufficient to explain the lack of NK1.1 reactivity ofthe BALB/c mouse strain. Furthermore, these results suggestthat the basis of NK1.1 alloreactivity across mouse strains, includ-ing the original (C3H � BALB)F1 �-CE immunizations used togenerate both the polyclonal �-NK-1 (3) and monoclonal �-NK1.1(PK136; Ref. 4) specificities, is due to a single amino acidsubstitution in either NKR-P1B, NKR-P1C, or both receptors. Dueto a lack of information about NKR-P1 expression in the CEmouse strain, we cannot distinguish these possibilities at present.Nonetheless, these results demonstrate that BALB/c mice are notdeficient in NKR-P1 expression, rather they possess divergentNKR-P1 sequences sufficient to generate alloantibodies whenimmunized with NK cells from other mouse strains.
The BALB/c NKR-P1B receptor is functional and recognizescognate Ocil/Clr-b ligand
Although BALB/c mice express functional NKR-P1B/C tran-scripts, it remains formally possible that BALB/c NK cells may bedeficient in NKR-P1-mediated recognition, if substitutions in theirNKR-P1 coding sequences lead to loss of ligand recognition. Al-though cognate ligands for the NKR-P1A/C receptors remain un-known, NKR-P1-mediated missing-self recognition of tumor cellshas been shown to depend on functional recognition of Ocil/Clr-b
FIGURE 5. PCR analysis of Nkrp1transcript expression in BALB/c vs B6NK cDNA libraries and fresh tissues. A,Limited cycle PCR analysis with high-fidelity polymerase was applied to cloneBALB/c strain Nkr-p1 cDNA sequences.Shown for comparison are Nkr-p1 ampli-fications performed using a B6 NK cellcDNA library in parallel. Two sets ofprimers were used: EC/UTR primers cor-respond to the extracellular and 3�-un-translated region; FL/CDS primers cor-respond to the full-length codingsequence regions. The signals shown forNkr-p1b/d are combined, as the primersamplify both sequences. B, Amplificationof full-length Nkrp1c sequences fromB6, BALB/c, and (B6xBALB)F1 LAK li-braries using the upstream 5�-Nkrp1a and3�-Nkrp1c primer combination. All PCRproducts were sequenced to confirmidentity. C, RT-PCR analysis of Nkrp1-Ocil/Clr transcripts in fresh tissues.cDNA from B6 and BALB/c splenocytes(splen.), BALB/c GM-CSF-culturedbone marrow dendritic cells (DC), andBALB/c NK cells enriched from IL-2cultured splenocytes (ALAK) are shown.NT, no template. All PCR products werecloned and sequenced to confirm theiridentity. Primers (see Table I) were de-signed based on available sequence toamplify only the BALB/c allele(Nkrp1cBALB, exons 3–5), or both B6 andBALB alleles (Nkrp1g, exons 3–5; Clr-b,full-length coding region; Clr-h, exons1–5).
7518 THE BALB/c Nkrp1-Ocil/Clr GENE CLUSTER
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
ligand by the NKR-P1B/D inhibitory receptors in various mousestrains (12). Therefore, we tested whether the NKR-P1BBALB re-ceptor is capable of recognizing Ocil/Clr-b ligand.
To do this, we used our previous modification of the BWZ re-porter-cell assay (12, 19). In this study, expression of a chimericCD3�/NKR-P1B fusion receptor on BWZ.36 reporter cells signals
functional recognition of ligand expressed on the surface of intacttarget cells, via NFAT-driven �-galactosidase enzyme expression.As shown previously, BWZ cells expressing the CD3�/NKR-P1BSw/SJL receptor are capable of recognizing target cells (eitherBWZ or 293T) expressing Ocil/Clr-b ligand, but not target cellsalone (BWZ.P1BSw/SJL; Fig. 8A). Moreover, this interaction is
FIGURE 6. Novel BALB/c and B6 Nkrp1-Ocil/Clr sequences. A, Alignment of the predicted coding sequences for the known Nkrp1 cDNAs is shown. Includedare related sequences from various mouse strains: B6, C57BL/6; BA, BALB/c; SJ, Swiss/SJL; 29, 129. NKR-P1 functional features are highlighted at the top:ITIM, underlined; CxCP Lck-recruitment motif, boldface; charged transmembrane residue position, arrow. B, The coding sequence of Nkrp1-related cDNAs frommice (B6, BALB, Swiss/SJL), rats, humans, and chickens was aligned using ClustalX software (�ftp://ftp-igbmc.u-strasbg.fr/pub/ClustalX/�) and bootstrap analysisof 1000 data sets was performed with PHYLIP. The bootstrap values for each grouping are shown as a percentage. The phylogram branch lengths indicate thesimilarity between different cDNAs and the scale-bar indicates the percent divergence. The putative coding sequence for Nkrp1eB6 was artificially spliced togetherfrom genomic data. Nkrp1gB6 is a transcript predicted from the B6 genome (XM355818) and is 100% identical with the available Nkrp1g exons (3–5) from theBALB/c mouse. All genes shown are known from newly isolated or previously published cDNAs. The Nkrp1f coding sequence in B6 and BALB mice is identical.C, All known Clr-like cDNA sequences from B6 mice, rats, humans, and chickens were analyzed as described in B. Clr-h is a transcript predicted from the B6genome (XM487965) and is 99% identical with the new Clr-like exon 3–4 fragment reported in this study.
7519The Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

specific for Ocil/Clr-b ligand, as it can be blocked using the�-Ocil/Clr-b mAb, 4A6 (Fig. 8A). Importantly, BWZ cells ex-pressing the NKR-P1BBALB receptor are also capable of specifi-cally recognizing Ocil/Clr-b ligand (BWZ.P1BBALB; Fig. 8A),while BWZ cells expressing a control CD69 fusion receptor do notrecognize Ocil/Clr-b (BWZ.CD69; Fig. 8A). Furthermore,BWZ.P1BBALB reporter cells also recognize native Ocil/Clr-b li-gand on the surface of various hemopoietic cells ex vivo, includingbone marrow (BM), lymph node (LN), spleen (SP), and thymus(TH) (Fig. 8B). These results demonstrate that the BALB/c NKR-P1B receptor is functional and recognizes cognate Ocil/Clr-b li-gand on the surface of normal cells. Thus, the NKR-P1B–Ocil/Clr-b missing-self recognition system is intact in BALB/c micedespite a single amino acid substitution that is responsible for thelack of NK1.1 reactivity of BALB/c NK cells.
DiscussionIt is widely believed that the lack of NK1.1 reactivity of BALB/cNK cells is due to a strain-specific deficiency in the expression ofone or more members of the Nkrp1 family (10). We show here thatBALB/c mice are normal with respect to Nkrp1 gene content andgenomic organization (Figs. 1–6), Nkrp1 transcript expression(Figs. 5–6), and NKR-P1B receptor function (Figs. 7–8). Further-more, we show that the lack of NK1.1 reactivity of BALB/c NK
cells is due to a single amino acid substitution (S191T) found inthe extracellular regions of both the BALB/c NKR-P1B and NKR-P1C receptors (the known NK1.1 Ags (5, 6, 9)), which abolishesthe epitope recognized by the �-NK1.1 mAb, PK136 (4) (Fig. 7).Despite allelic differences, the BALB/c NKR-P1B receptor is fullyfunctional and recognizes Ocil/Clr-b ligand on the surface of trans-fected and normal cells (Fig. 8). Our finding that the BALB/c and129 strains appear to share a related Nkrp1 haplotype (Fig. 4)suggests that other NK1.1� mouse strains possess similar allelicdifferences. These results indicate that the NKR-P1–Ocil/Clr re-ceptor-ligand system plays a broader and more important role intarget cell recognition than previously thought.
Insight from the BALB/c Nkrp1 region: Nkrp1b and Nkrp1ddelineate divergent murine NKC haplotypes
A significant finding of this study is that the mouse Nkrp1b andNkrp1d genes appear to represent alleles of the same genetic locus(Fig. 3). It has long been suspected that this might be the case, fora number of reasons. Although the original Nkrp1b cDNA se-quence (gene 34; Ref. 31) was reportedly derived from B6 mice,along with Nkrp1a (gene 2; Ref. 31); mNKR-P1.7 (9, 28) andNkrp1c (gene 40; Ref. 31); mNKR-P1.9 (9), we and others havebeen unable to identify an Nkrp1b genomic or cDNA sequence, or
FIGURE 7. NK1.1 epitope mapping of mouse NKR-P1 proteins. A, Amino acid alignment of the mouse NKR-P1 distal C-terminal sequences regionsis shown. Residues that differ between NKR-P1BBALB and NKR-P1BSw/SJL are shown in boldface. †Predicted coding sequence from the B6 genomicdatabase (XM355818) (B) NK1.1 reactivity of known mouse and rat NKR-P1 proteins. 293T cells were transfected with vectors encoding the indicatedcDNA’s (plus GFP reporter vector; see Materials and Methods), then cells were analyzed by flow cytometry using the PK136 �-NK1.1 mAb. Alltransfections are shown gated on GFP� transfected cells. C, NK1.1 reactivity of NKR-P1BBALB variants generated by site-directed mutagenesis. Shownare control NKR-P1BSw/SJL and NKR-P1BBALB transfectants of 293T cells (as in B), as well as NKR-P1BBALB (K217E) and NKR-P1BBALB (T191S)mutants (gated on GFP� transfected cells).
7520 THE BALB/c Nkrp1-Ocil/Clr GENE CLUSTER
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

an NKR-P1B protein, from the B6 background (5, 6, 27). How-ever, our group and Kung et al. (6) were able to isolate an identicalNkrp1b cDNA sequence derived from the Sw (5) and SJL strains(6), respectively. Thus, it is possible, due to the large numbers ofNK cells required for cDNA library preparation and Northern blot-ting, that a mixed strain may have been included in the originalidentification of the mouse Nkrp1 cDNA family. In any case, weand others were successful in cloning an Nkrp1b-related cDNAfrom B6 NK cells by RT-PCR (5, 6). Due to significant codingsequence divergence in the extracellular region (21 nonsynony-mous substitutions, a difference similar to that observed betweenother Nkrp1 genes), we gave this cDNA a novel designation,Nkrp1d (GenBank accession no. AF338321; Ref. 5); however,Kung et al. (6) only detected 15-aa substitutions and designatedtheir cDNA sequence, Nkrp1bB6 (GenBank accession no.AF354260). Notwithstanding these differences, our collectivecDNA findings were later confirmed at the genomic level throughrandom sequencing of the B6 Nkrp1 region: no evidence for theexistence of a B6 Nkrp1b sequence could be found, resulting in thedesignation of the B6 gene, Nkrp1d (GenBank accession no.AF324825; Ref. 27). In the absence of information about theNkrp1 genomic region from other strains, including Sw or SJLmice, this issue has remained unresolved until now.
Our new data from the BALB/c genome, however, sheds lighton the allelic nature of Nkrp1b and Nkrp1d: (1) only a single in-hibitory-class cDNA and/or gene could be identified in BALB/c,similar to B6 and other strains analyzed to date; (2) the genomiclocation of the BALB/c Nkrp1b gene is similar to that of the B6Nkrp1d gene (3); the Nkrp1bBALB and Nkrp1dB6 probes both hy-bridize to the same bands on genomic Southern blots; and (4) theNkrp1bBALB gene shares a highly conserved e3-i3-e4 organizationand sequence with the Nkrp1dB6 locus (Fig. 9). If Nkrp1b and
Nkrp1d are indeed allelic, they show much greater allelic diver-gence than other Nkrp1 genes among B6, BALB/c, and othermouse strains (with exception of the Nkrp1c locus). The reason forthis specific and directed divergence is unclear, but it is even moreintriguing considering that the NKR-P1B/D receptors share a com-mon ligand, the product of the Ocil/Clr-b locus, which is identicalamong the B6, BALB/c, and 129 strains (12, 13). Differential bind-ing of NKR-P1B and NKR-P1D to a single allele of Ocil/Clr-b hasbeen observed before in functional assays (12); however, thiscould reflect a differential affinity of each gene product, as a con-sequence of their divergent sequences, for a common ligand. Inturn, such differential binding of NKR-P1 receptors to their Ocil/Clr ligands could have implications for the role of this system inmissing self recognition and transplantation biology (12). Confir-mation of such a role will have to await the elucidation of Ocil/Clralleles and haplotypes, as well as functional analyses of the role ofOcil/Clr proteins in allotransplantation.
Another interesting finding of the genomic analyses is that theBALB/c and 129 strains appear to share a related Nkrp1 haplotype(Fig. 4 and Ref. 28). In fact, an Nkrp1b-like sequence has beenidentified in 129-strain mice, designated Nkrp1d129, that resemblesNkrp1bBALB more than Nkrp1dB6 (13); indeed, the NKR-P1D129
protein differs from the NKR-P1BBALB and NKR-P1BSw/SJL pro-teins by only 5- or 6-aa, respectively, and possesses the S191Tsubstitution that abrogates NK1.1 reactivity (Fig. 7). As bothBALB/c and 129 are well-known NK1.1� mouse strains, this sug-gests that other NK1.1� strains possess similar allelic differences.Interestingly, our classification of Nkrp1 haplotypes thus far ap-pears to delineate functionally distinct NK1.1 alloantigen expres-sion patterns: B6 strain (group 1) NK cells are known express anNK1.1 alloantigen (Nkrp1c) (9); BALB/c and 129 strain (group 2)NK cells do not express NK1.1 (4); while FVB strain (group 3) NK
FIGURE 8. Functional analysis of Ocil/Clr-b li-gand binding by the BALB/c NKR-P1B receptor. A,BWZ reporter cell assay analysis of NKR-P1 trans-ductants vs a panel of target cells. Parental and Ocil/Clr-b-expressing BWZ variants and 293T transfec-tants with and without 4A6 �-Ocil/Clr-b blockingmAb were tested. B, BWZ reporter cell assay anal-ysis of NKR-P1 transductants vs cells from normalmouse tissues ex vivo. Shown are parental BWZcells, transductants bearing the NKR-P1BSw/SJL andNKR-P1BBALB receptors, as well as transductantsexpressing mouse CD69, as a control.
7521The Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

cells are also known to express NK1.1 (Nkrp1b) (7). Notably, FVBmice represent an inbred Swiss-related mouse strain (7), perhapsexplaining the similar basis for NK1.1 reactivity (Nkrp1b) of FVB
(7), Sw (5), SJL (6), and CD-1 (12) NK cells; thus, it is likely thatthese strains all share a related Nkrp1 haplotype (group 3). Becausethere are currently no data on the relationship of any of these
FIGURE 9. Comparison of Nkrp1 gene structure. A, A scale diagram of the organization of exons 3–5 of various mouse Nkrp1 genes is shown.Differentially shaded boxes indicate exons and various types of repetitive elements in introns as highlighted at the bottom. Numbers indicate the percentagenucleotide identity of the exon 3–5 region between B6 and available BALB/c alleles. The identity score shown for Nkrp1cB6 and Nkrp1cBALB ignores theLINE element in the Nkrp1cBALB gene. B, Individual exon 3–5 sequences of Nkrp1 genes from B6 and BALB/c mice were aligned using ClustalX software,and bootstrap analysis of 1000 data sets was performed with Phylip software (�http://evolution.genetics.washington.edu/phylip.html�). The bootstrap valuesfor each grouping are shown as a percentage. Phylogram branch lengths indicate the exon similarity among different Nkrp1 genes, and the scale bar indicatesthe percent divergence. When different strains are grouped together in one gene, this indicates exon identity.
7522 THE BALB/c Nkrp1-Ocil/Clr GENE CLUSTER
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

strains to the donor CE strain used for immunizations to generatethe original polyclonal (3) and monoclonal (4) anti-NK1.1 alloan-tibodies, the identity of the original NK1.1 Ag remains unknown.
Nevertheless, before any final conclusions can be made con-cerning Nkrp1 (and Ocil/Clr) haplotype variation, more haplotypeswill have to be characterized. The genomic organization and contentof the BALB/c and B6 Nkrp1-Ocil/Clr regions are more similarthan the Ly49 regions between these strains. If one assumes thatthe BALB/c and B6 Nkrp1-Ocil/Clr and Ly49 haplotype associa-tion is a natural one and not a consequence of the original deri-vation of inbred mice, then it would appear that the Nkrp1-Ocil/Clrregion is evolving less rapidly than the Ly49 region. Although thereason for this remains speculative at present, a striking differencebetween the two NK cell receptor systems is that the Nkrp1 genesare genetically intermingled in the NKC with the Ocil/Clr genesthat encode their ligands (12, 13); thus, both are coinherited. Incontrast, the Ly49 genes segregate independently of the genes en-coding their ligands, because the MHC is located on a differentchromosome than the NKC (18). Coinheritance of ligand-encodinggenes may negatively affect the rate of accumulated receptor genemutations (i.e., receptor evolution), and vice-versa, because aminoacid changes that decrease receptor-ligand binding affinities mightbe selected against. In contrast, the observed level of allelic diver-gence between Nkrp1b and Nkrp1d (and within the Nkrp1c locus)suggests that this may not always be the case. Further character-ization of Ocil/Clr haplotypes is necessary to clarify the relation-ship between allelic divergence of the Nkrp1 and Ocil/Clr genesand how that influences their function. Intriguingly, the underlyingbasis for the evolution of numerous Nkrp1 genes in rodents andonly a single gene (NKRP1A; Ref. 32) in humans is unknown. Yetlike the Ly49 receptor system, this dichotomy is probably a func-tional consequence of differences in receptor usage by rodent vshuman NK cells. Only a single Ocil/Clr-b-like gene exists in hu-mans, designated LLT1 (18, 33). Indeed, the recent finding thathuman NKR-P1A–LLT1 interact and inhibit NK cell functiondemonstrates that the NKR-P1 missing-self recognition system isintact in humans (34, 35).
Identification of novel Nkrp1 and Ocil/Clr genes
Southern cross-hybridization and PCR cross-amplification of BACclones and cDNA libraries identified sequences in the BALB/cgenome that appear to represent novel Nkrp1 and Ocil/Clr genes.The first of these, designated Clr-h, was identified from BAC anal-ysis using primers intended for Clr-b (cross-reactive to Clr-d).This new gene is not unique to the BALB/c genome; rather, thesequence is readily identifiable in the latest assembly of the publicB6 genome database (99% identical across e3-i3-e4; GenBank ac-cession no. XM487965). Moreover, the corresponding region inthe B6 genome does not fit the location of any other known Clrgene (18). Such a high degree of sequence conservation suggeststhat Clr-h either represents a ligand for a highly conserved recep-tor, or that it is not subject to selection pressure and thus mayrepresent a nonfunctional gene/pseudogene (at least in the strainstested; Fig. 5C), or encode a ligand for a nonfunctional receptor. Inthis light, it is interesting to note that the Nkrp1a sequence ishighly conserved, the Nkrp1f sequence is identical between B6(27) and BALB/c (this study), and the Nkrp1e sequence appears tobe nonfunctional in both strains (this study, and Ref. 27). As theNKR-P1F receptor binds to Ocilrp2/Clr-g (13, 16, 17, 36, 37), it islikely that at least some of the remaining NKR-P1 receptors andOcil/Clr ligands bind to one another. Although the Clr-c/d/e genesare not novel, this study elucidates their relative order within theNKC (i.e., Clr-g, Clr-d, Clr-e, Clr-c, and Nkrp1f)
A second new gene, Nkrp1g, is predicted to exist in the intervalbetween Clr-f and Nkrp1c, based upon Southern cross-hybridiza-tion with an Nkrp1f cDNA probe. Although it is not knownwhether the sequence encodes a functional receptor, a sequence iscurrently available for this putative new gene in the B6 genomedatabase (GenBank accession no. XM355818). The coding se-quence bears some similarity to the NKR-P1F receptor in the ex-tracellular region (16), but the predicted protein appears to lackknown signaling motifs, including a cytoplasmic ITIM or Y resi-due, a putative CxCP recruitment motif for the Lck tyrosine ki-nase, and a charged transmembrane R residue (5, 38). Thus, fullBAC sequencing may be required to assess the significance of theBALB/c cross-hybridization results, if no related cDNA can beisolated. This approach may also provide a clue to the origin andrelationship of this sequence to the other Nkrp1 genes and/or pseu-dogenes. Although attempts to isolate an Nkrp1g cDNA were un-successful (at least in the strains tested; Fig. 5C), the Nkrp1g se-quence appears to be conserved in the rat genome (Ensembl IDENSRNOT00000035766; �www.ensembl.org�), along with Nkrp1a(GenBank accession no. M62891), Nkrp1b (GenBank accessionno. U56936), and Nkrp1f (GenBank accession no. X97477).
Conformational nature of the NK1.1 epitope
As mentioned previously, the NK1.1 epitope is specific to NKR-P1BSw/SJL and NKR-P1CB6 among known NKR-P1 proteins fromboth mice and rats (Fig. 7). However, sequence alignment of theseNKR-P1 proteins does not provide a clue as to the context of theNK1.1-reactive sequence. Our identification of a substitution in theNKR-P1BBALB vs NKR-P1BSw/SJL sequence that confers NK1.1reactivity by no means indicates that this amino acid alone is suf-ficient to generate the NK1.1 epitope. In contrast, both the ratNKR-P1A and NKR-P1B sequences contain the NK1.1-reactive Sresidue at the correct position, yet neither receptor bears the NK1.1epitope. This is not surprising considering that NKR-P1B/C can beimmunoprecipitated but not Western blotted using PK136 mAb(J.R.C., unpublished observations). This suggests that the NK1.1epitope is conformational in nature, relying on additional context-dependent amino acid residues, and that denaturation of the three-dimensional structure of the epitope destroys its reactivity.
It was surprising that the seemingly conservative S191T substi-tution found in both NKR-P1BBALB and NKR-P1CBALB couldabolish NK1.1 reactivity, whereas reversing this single amino acidsubstitution could confer NK1.1 reactivity (Fig. 7). The chemicalnature of S and T residues is quite similar: both residues contain ahydroxymethyl group, yet the T residue has one additional methylgroup. The implications of this are not entirely clear, but since bothS and T residues can be posttranslationally modified by O-linkedpolysaccharides, it is possible that a specific O-linked glycosyl-transferase may be partially responsible for either conferring ordestroying the NK1.1 epitope on these receptors. Carbohydratemodification of the NKR-P1 receptors is not functionally requiredfor ligand binding, because tetramers of NKR-P1 proteins pro-duced in bacteria bind to their cognate ligands (12, 13); however,such modifications could alter the affinity of the receptor–ligandinteraction, because bacterially produced NKR-P1 tetramers onlybind their ligands weakly (12, 13). This is evident in the findingsthat NKR-P1D tetramers fail to visualize native Ocil/Clr-b ligandon normal cells ex vivo (13), and NKR-P1B tetramers bind moreweakly to Ocil/Clr-b ligand than the �-Ocil/Clr-b mAb, 4A6(which is a low-affinity IgM) (12). Nonetheless, our present find-ings using the NKR-P1BBALB receptor in the BWZ reporter sys-tem recapitulate our in vitro and ex vivo results using an NKR-P1BSw/SJL receptor reporter cell (12). These results confirm that
7523The Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

the cognate Ocil/Clr-b ligand recognized by these inhibitory re-ceptors is expressed in a broad manner reminiscent of MHC classI (12), in contrast with the more restricted expression pattern sug-gested by other reports (13). The correlation between Ocil/Clr-btranscript expression (15, 16) and surface protein staining using�-Ocil/Clr-b 4A6 mAb (12) seems to be in agreement with ourcurrent findings.
Thus, the NKR-P1B–Ocil/Clr-b missing self recognition systemis intact in BALB/c mice, despite their well-known NK1.1� straindesignation. The generation of mAbs to the BALB/c NKR-P1 pro-teins should facilitate direct analyses of receptor expression andfunction. In addition, further analysis of the known and novelNKR-P1–Ocil/Clr interactions in various mouse strains should elu-cidate the complex nature of this system of at least 6 potentialreceptor and 8 potential ligand genes linked to one another in theNKC; heterodimerization of these proteins could offer as yet un-appreciated additional complexity to NK cell recognition of targetcells.
Note added in proof. The genomic sequences and intron–exonstructure of the Nkrp1 genes in this study have been modified fromLjutic et. al. (39) to conform to the Ly49 gene nomenclature (i.e.,exon 2, cytoplasmic; exon 3, transmembrane, exon 4, stalk; exons5–7, C-type lectin-like domain).
AcknowledgmentsWe thank Dr. J.C. Zuniga-Pflucker for support; Drs. David Raulet, LewisLanier, Richard Miller, and Stephen Anderson for suggestions; Renee De-Pooter for critical reading of the manuscript; and Gisele Knowles for cellsorting.
DisclosuresThe authors have no financial conflict of interest.
References1. Trinchieri, G. 1989. Biology of natural killer cells. Adv. Immunol. 47: 187–376.2. Lanier, L. L. 2005. NK cell recognition. Annu. Rev. Immunol. 23: 225–274.3. Glimcher, L., F. W. Shen, and H. Cantor. 1977. Identification of a cell-surface
antigen selectively expressed on the natural killer cell. J. Exp. Med. 145: 1–9.4. Koo, G. C., and J. R. Peppard. 1984. Establishment of monoclonal anti-NK-1.1
antibody. Hybridoma 3: 301–303.5. Carlyle, J. R., A. Martin, A. Mehra, L. Attisano, F. W. Tsui, and
J. C. Zuniga-Pflucker. 1999. Mouse NKR-P1B, a novel NK1.1 antigen with in-hibitory function. J. Immunol. 162: 5917–5923.
6. Kung, S. K., R. C. Su, J. Shannon, and R. G. Miller. 1999. The NKR-P1B geneproduct is an inhibitory receptor on SJL/J NK cells. J. Immunol. 162: 5876–5887.
7. Liu, J., M. A. Morris, P. Nguyen, T. C. George, E. Koulich, W. C. Lai,J. D. Schatzle, V. Kumar, and M. Bennett. 2000. Ly49I NK cell receptor trans-gene inhibition of rejection of H2b mouse bone marrow transplants. J. Immunol.164: 1793–1799.
8. Hackett, J., Jr., M. Tutt, M. Lipscomb, M. Bennett, G. Koo, and V. Kumar. 1986.Origin and differentiation of natural killer cells. II. Functional and morphologicstudies of purified NK-1.1� cells. J. Immunol. 136: 3124–3131.
9. Ryan, J. C., J. Turck, E. C. Niemi, W. M. Yokoyama, and W. E. Seaman. 1992.Molecular cloning of the NK1.1 antigen, a member of the NKR-P1 family ofnatural killer cell activation molecules. J. Immunol. 149: 1631–1635.
10. Giorda, R., E. P. Weisberg, K. I. Tze, and M. Trucco. 1992. Genomic structureand strain-specific expression of the natural killer cell receptor NKR-P1. J. Im-munol. 149: 1957–1963.
11. Anderson, S. K., K. Dewar, M. L. Goulet, G. Leveque, and A. P. Makrigiannis.2005. Complete elucidation of a minimal class I MHC natural killer cell receptorhaplotype. Genes Immun. 6: 481–492.
12. Carlyle, J. R., A. M. Jamieson, S. Gasser, C. S. Clingan, H. Arase, andD. H. Raulet. 2004. Missing self recognition of Ocil/Clr-b by inhibitory NKR-P1natural killer cell receptors. Proc. Natl. Acad. Sci. USA 101: 3527–3532.
13. Iizuka, K., O. Naidenko, B. Plougastel, D. Fremont, and W. M. Yokoyama. 2003.Genetically linked C-type lectin-related ligands for the NKR-P1 NK cell recep-tors. Nat. Immunol. 4: 801–807.
14. Kumar, V., and M. E. McNerney. 2005. A new self: MHC-class-I-independentnatural-killer-cell self-tolerance. Nat. Rev. Immunol. 5: 363–374.
15. Zhou, H., V. Kartsogiannis, Y. S. Hu, J. Elliott, J. M. Quinn, W. J. McKinstry,M. T. Gillespie, and K. W. Ng. 2001. A novel osteoblast-derived C-type lectinthat inhibits osteoclast formation. J. Biol. Chem. 276: 14916–14923.
16. Plougastel, B., C. Dubbelde, and W. M. Yokoyama. 2001. Cloning of Clr, a newfamily of lectin-like genes localized between mouse Nkrp1a and Cd69. Immu-nogenetics 53: 209–214.
17. Zhou, H., V. Kartsogiannis, J. M. Quinn, C. Ly, C. Gange, J. Elliott, K. W. Ng,and M. T. Gillespie. 2002. Osteoclast inhibitory lectin, a family of new osteoclastinhibitors. J. Biol. Chem. 277: 48808–48815.
18. Yokoyama, W. M., and B. F. Plougastel. 2003. Immune functions encoded by thenatural killer gene complex. Nat. Rev. Immunol. 3: 304–316.
19. Sanderson, S., and N. Shastri. 1994. Lacz inducible, antigen/MHC-specific T-cellhybrids. Int. Immunol. 6: 369–376.
20. Li, J., B. A. Rabinovich, R. Hurren, J. Shannon, and R. G. Miller. 2003. Expres-sion cloning and function of the rat NK activating and inhibitory receptors NKR-P1A and -P1B. Int. Immunol. 15: 411–416.
21. Carlyle, J. R., A. M. Michie, S. K. Cho, and J. C. Zuniga-Pflucker. 1998. Naturalkiller cell development and function precede �� T cell differentiation in mousefetal thymic ontogeny. J. Immunol. 160: 744–753.
22. Arase, H., T. Saito, J. H. Phillips, and L. L. Lanier. 2001. Cutting edge: the mouseNK cell-associated antigen recognized by DX5 monoclonal antibody is CD49b(�2 integrin, very late antigen-2). J. Immunol. 167: 1141–1144.
23. Stoneman, E. R., M. Bennett, J. An, K. A. Chesnut, E. K. Wakeland,J. B. Scheerer, M. J. Siciliano, V. Kumar, and P. A. Mathew. 1995. Cloning andcharacterization of 5E6 (Ly49C), a receptor molecule expressed on a subset ofmurine natural killer cells. J. Exp. Med. 182: 305–313.
24. Dorshkind, K., S. B. Pollack, M. J. Bosma, and R. A. Phillips. 1985. Naturalkiller (NK) cells are present in mice with severe combined immunodeficiency(scid). J. Immunol. 134: 3798–3801.
25. Proteau, M. F., E. Rousselle, and A. P. Makrigiannis. 2004. Mapping of theBALB/c Ly49 cluster defines a minimal natural killer cell receptor gene reper-toire. Genomics 84: 669–677.
26. Makrigiannis, A. P., D. Patel, M. L. Goulet, K. Dewar, and S. K. Anderson. 2005.Direct sequence comparison of two divergent class I MHC natural killer cellreceptor haplotypes. Genes. Immun. 6: 71–83.
27. Plougastel, B., K. Matsumoto, C. Dubbelde, and W. M. Yokoyama. 2001. Anal-ysis of a 1-Mb BAC contig overlapping the mouse Nkrp1 cluster of genes: clon-ing of three new Nkrp1 members, Nkrp1d, Nkrp1e, and Nkrp1f. Immunogenetics53: 592–598.
28. Yokoyama, W. M., J. C. Ryan, J. J. Hunter, H. R. Smith, M. Stark, andW. E. Seaman. 1991. cDNA cloning of mouse NKR-P1 and genetic linkage withLy-49. Identification of a natural killer cell gene complex on mouse chromosome6. J. Immunol. 147: 3229–3236.
29. Makrigiannis, A. P., P. Gosselin, L. H. Mason, L. S. Taylor, D. W. McVicar,J. R. Ortaldo, and S. K. Anderson. 1999. Cloning and characterization of a novelactivating Ly49 closely related to Ly49A. J. Immunol. 163: 4931–4938.
30. Mehta, I. K., H. R. Smith, J. Wang, D. H. Margulies, and W. M. Yokoyama.2001. A “chimeric” C57l-derived Ly49 inhibitory receptor resembling the Ly49Dactivation receptor. Cell Immunol. 209: 29–41.
31. Giorda, R., and M. Trucco. 1991. Mouse NKR-P1. A family of genes selectivelycoexpressed in adherent lymphokine-activated killer cells. J. Immunol. 147:1701–1708.
32. Lanier, L., C. Chang, and J. Phillips. 1994. Human NKR-P1A. A disulfide-linkedhomodimer of the C-type lectin superfamily expressed by a subset of NK and Tlymphocytes. J. Immunol. 153: 2417–2428.
33. Boles, K. S., R. Barten, P. R. Kumaresan, J. Trowsdale, and P. A. Mathew. 1999.Cloning of a new lectin-like receptor expressed on human NK cells. Immunoge-netics 50: 1–7.
34. Aldemir, H., V. Prod’homme, M. J. Dumaurier, C. Retiere, G. Poupon,J. Cazareth, F. Bihl, and V. M. Braud. 2005. Cutting edge: Lectin-like transcript1 is a ligand for the CD161 receptor. J. Immunol. 175: 7791–7795.
35. Rosen, D. B., J. Bettadapura, M. Alsharifi, P. A. Mathew, H. S. Warren, andL. L. Lanier. 2005. Cutting edge: Lectin-like transcript-1 is a ligand for theinhibitory human NKR-P1A receptor. J. Immunol. 175: 7796–7799.
36. Tian, W., R. Nunez, S. Cheng, Y. Ding, J. Tumang, C. Lyddane, C. Roman, andH. C. Liou. 2005. C-type lectin OCILRP2/Clr-g and its ligand NKRP1f costimu-late T cell proliferation and IL-2 production. Cell Immunol. 234: 39–53.
37. Tian, W., B. Feng, and H. C. Liou. 2005. Silencing OCILRP2 leads to intrinsicdefects in T cells in response to antigenic stimulation. Cell Immunol. 235: 72–84.
38. Ljutic, B., J. R. Carlyle, D. Filipp, R. Nakagawa, M. Julius, andJ. C. Zuniga-Pflucker. 2005. Functional requirements for signaling through thestimulatory and inhibitory mouse NKR-P1 (CD161) NK cell receptors. J. Immu-nol. 174: 4789–4796.
39. Ljutic, B., J. R. Carlyle, and J. C. Zuniga-Pflucker. 2003. Identification of up-stream cis-acting regulatory elements controlling lineage-specific expression ofthe mouse NK cell activation receptor, NKR-P1C. J. Biol. Chem. 278:31909–31917. Vol. 176, No. 12
7524 THE BALB/c Nkrp1-Ocil/Clr GENE CLUSTER
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from



![tAymnielt-7, · 2019. 11. 28. · Amir Misirlic 3000 Anida Kapo 1000 Asim 'åHOLORYLþ 6000 Asim 0XMNLþ 1000 Belma Tvico 10000 Benjamin %DMUDPRYLþ 1500 Berin 7X]OLþ 3000 Damir](https://static.fdocuments.us/doc/165x107/60fe8b77205eb35ca86b5c21/taymnielt-7-2019-11-28-amir-misirlic-3000-anida-kapo-1000-asim-holoryl.jpg)














