
Modulatory effect of quercetin on azathioprine induced ...
5
Indian Journal of Experimental Biology Vol. 39, February 2001, pp. 132-136 Modulatory effect of quercetin on azathioprine induced membrane changes in the mouse spleen S Chitra, S Devipriya & C S Shyamala Devi* Department of Biochemistry and Molecular Biology, University of Madras, Guindy Campus, Chennai 600 025, India Received 12 November 1999; revised I November 2000 Modulatory effect of quercetin on azathioprine induced toxic changes was studied in spleen of experimental animals. Azathioprine trea tment caused an increase in serum albumin/globin ratio and a decrease in total protein in spleen tissue. An increase in a membrane bound ATPases was also noted. Supplementation of quercetin with azathioprine increased the protein content and lowe red the ac ti vities of membrane ATPase in spleen. There was a decrease in serum albumin globulin ratio. It was concluded that quercetin modulated the prote in and membrane bound ATPase activities and prOtected the spleen from azathioprine induced membrane damage. Azathioprine (6, 1-methyl-4-nitroimidazole-5-thio puine) is the main cytotoxic agent used for immunosuppres- sion and is widely used to control tissue rejection in transplant surgery. After oral or intravenous admini- stration, it disappears rapidly from the circulation and is extensively metabolised to mercaptopurine', purine analogue that inhibits DNA synthesis. Spleen is the most affected organ in the immunosuppressive condition 2 • Other side effects associated with azathioprine include drug fever, liver damage 3 and pancreatitis 4 . Azathioprine exacerbates the condition and also causes joint and skin lesions. Other adverse effects are nausea, vomiting and pyrexia 5 . Flavonoids have been recognised as antioxidative a nd free radical scavenging agents 6 and as membrane ATPase inhibitors 7 . Quercetin [(3,3',4'5,7) pentahydroxy flavone)] is one of the most common phenolic compounds in vascular plants. It occurs in conjugated or free form in many plant products used for food, including many fruits, vegetables and tea 8 . In mice quercetin increases the hepatic and pulmonary activities. Increase in lung and liver activities may be either due to detoxification of the chemicals or by controlling the free radical mediated bioreactions 9 . This confirms the antioxidant potential of quercetin. The present study was undertaken to study the modulatory changes in the spleen on co-administration of quercetin with azathioprine in an experimental mice. Materials and Methods Drugs and chemicals-Quercetin (commercially *Correspondent author available powdered form was used) was obtained from M/s. Research Organics Chemical Company, Chennai, India. Azathioprine, bovine serum albumin were obtained from Sigma USA. Other chemicals used were of analytical grade. Animals -Male albino mice (25 g) were maintained in the following standard environmental conditions. They were fed with commercial pelleted diet obtained from Hindustan Lever Limited anc given food and water ad libi tu m. Animals wen housed six per cage at 27°±2°C with constan humidity under (55%) 12 hr light /dark cycle. Quercetin in the powdered form was dissolved it dimethyl sulfoxide (0.8 mL) reagent and administerec intraperitoneally (ip). Azathioprine was dissolved it water and was given orally (3mg/kg body w( Animals were divided into four groups consisting of · mice in each group, azathioprine tablets wer administered orally, but experienced difficuity i administering DMSO by oral route. Group I animals (control) received no treatmeri group II animals received azathioprine (3 mg/kg bod wt) for 28 days; group III animals received quercet (25 mg/kg body wt) 10 every week for 28 days (da: 1,7,14,21 and 28); and group IV animals receivt simultaneously a treatment of azathioprine (3mg!l body wt) and quercetin (25 mglkg body wt) for : days as given in groups II and III. At the end of the treatment, the animals were fast• for 24 hr to avoid metabolic disturbances a sacrificed by cervical decapitation. Serum w collected by centrifugation for estimation of album globulin ratio 11 • Agarose slide gel electrophoresis
Transcript of Modulatory effect of quercetin on azathioprine induced ...

Indian Journal of Experimental Biology Vol. 39, February 2001, pp. 132-136
Modulatory effect of quercetin on azathioprine induced membrane changes in the mouse spleen
S Chitra, S Devipriya & C S Shyamala Devi*
Department of Biochemistry and Molecular Biology, University of Madras, Guindy Campus, Chennai 600 025, India
Received 12 November 1999; revised I November 2000
Modulatory effect of quercetin on azathioprine induced toxic changes was studied in spleen of experimental animals. Azathioprine treatment caused an increase in serum albumin/globin ratio and a decrease in total protein in spleen tissue. An increase in a membrane bound ATPases was also noted. Supplementation of quercetin with azathioprine increased the protein content and lowered the acti vities of membrane ATPase in spleen. There was a decrease in serum albumin globulin ratio. It was concluded that quercetin modulated the protein and membrane bound ATPase activities and prOtected the spleen from azathioprine induced membrane damage.
Azathioprine (6, 1-methyl-4-nitroimidazole-5-thio puine) is the main cytotoxic agent used for immunosuppression and is widely used to control tissue rejection in transplant surgery. After oral or intravenous administration, it disappears rapidly from the circulation and is extensively metabolised to mercaptopurine' , purine analogue that inhibits DNA synthesis. Spleen is the most affected organ in the immunosuppressive condition2
• Other side effects associated with azathioprine include drug fever, liver damage3 and pancreatitis4
. Azathioprine exacerbates the condition and also causes joint and skin lesions. Other adverse effects are nausea, vomiting and pyrexia5
.
Flavonoids have been recognised as antioxidative and free radical scavenging agents6 and as membrane ATPase inhibitors7
. Quercetin [(3,3',4'5,7) pentahydroxy flavone)] is one of the most common phenolic compounds in vascular plants. It occurs in conjugated or free form in many plant products used for food, including many fruits, vegetables and tea8
. In mice quercetin increases the hepatic and pulmonary activities. Increase in lung and liver activities may be either due to detoxification of the chemicals or by controlling the free radical mediated bioreactions9
.
This confirms the antioxidant potential of quercetin. The present study was undertaken to study the modulatory changes in the spleen on co-administration of quercetin with azathioprine in an experimental mice.
Materials and Methods Drugs and chemicals-Quercetin (commercially
*Correspondent author
available powdered form was used) was obtained from M/s . Research Organics Chemical Company, Chennai, India. Azathioprine, bovine serum albumin were obtained from Sigma USA. Other chemicals used were of analytical grade.
Animals -Male albino mice (25 g) were maintained in the following standard environmental conditions. They were fed with commercial pelleted diet obtained from Hindustan Lever Limited anc given food and water ad libitum. Animals wen housed six per cage at 27°±2°C with constan humidity under (55%) 12 hr light /dark cycle.
Quercetin in the powdered form was dissolved it dimethyl sulfoxide (0.8 mL) reagent and administerec intraperitoneally (ip). Azathioprine was dissolved it water and was given orally (3mg/kg body w( Animals were divided into four groups consisting of · mice in each group, azathioprine tablets wer administered orally, but experienced difficuity i administering DMSO by oral route.
Group I animals (control) received no treatmeri group II animals received azathioprine (3 mg/kg bod wt) for 28 days; group III animals received quercet (25 mg/kg body wt) 10 every week for 28 days (da: 1,7,14,21 and 28); and group IV animals receivt simultaneously a treatment of azathioprine (3 mg!l body wt) and quercetin (25 mglkg body wt) for : days as given in groups II and III.
At the end of the treatment, the animals were fast• for 24 hr to avoid metabolic disturbances a sacrificed by cervical decapitation. Serum w collected by centrifugation for estimation of album globulin ratio 11
• Agarose slide gel electrophoresis

CHITRA eta/.: MODULATORY EFFECT OF QUERCETIN ON AZATHIOPRINE INDUCED MEMBRANE CHANGES 133
serum protein in control and experimental animals was carried out. Spleen was dissected out, washed with chilled physiological saline, weighed, homogenised in 0.1 M Tris- HCl buffer (pH 7.4) at 4°C in Potter-Elvejem homogenizer and then used for evaluation of protein content' 2 and membrane bound ATPase activities 13
-16
• Activities of xanthine oxidase 17
, adenosine deaminase18 and 5' nucleotidase19 were estimated in serum and spleen tissue.
Statistical evaluatioll- Student's t test and two way analysis of variance (Anova) were used for statistical analysis.
Results and Discussion In azathioprine treated animals the albumin/
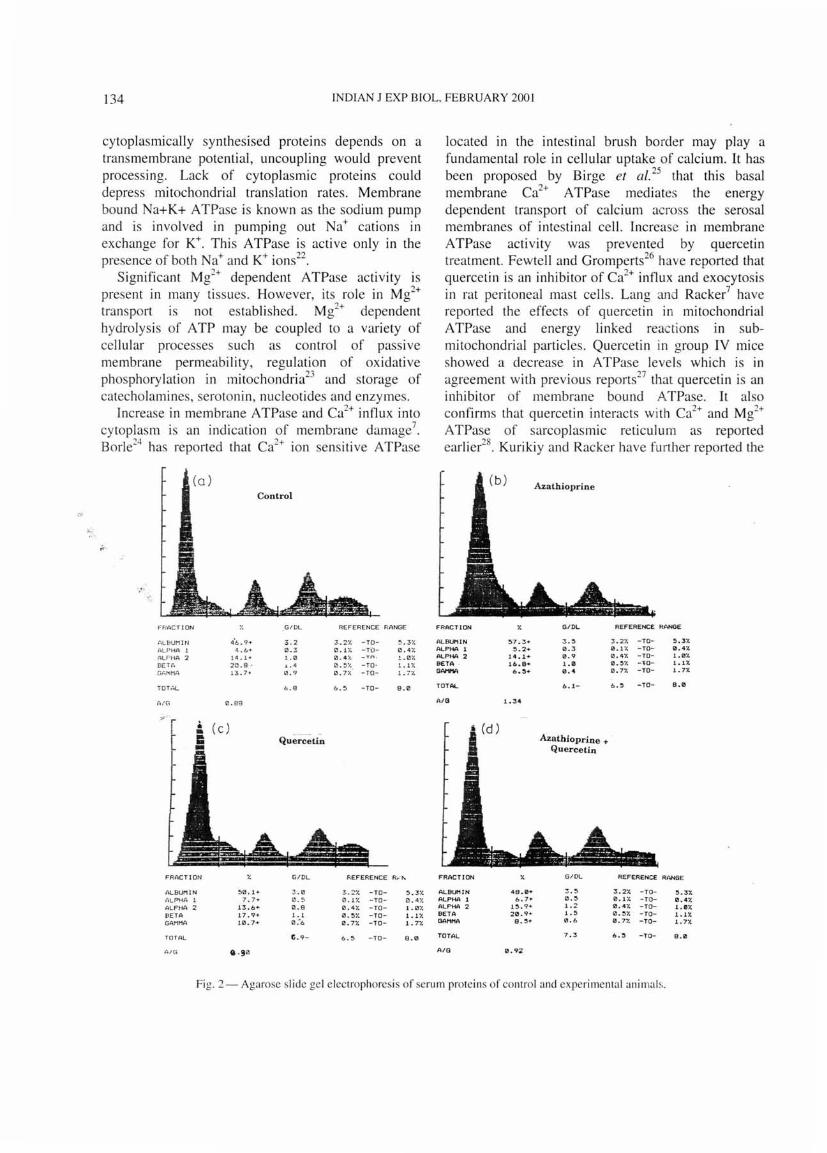
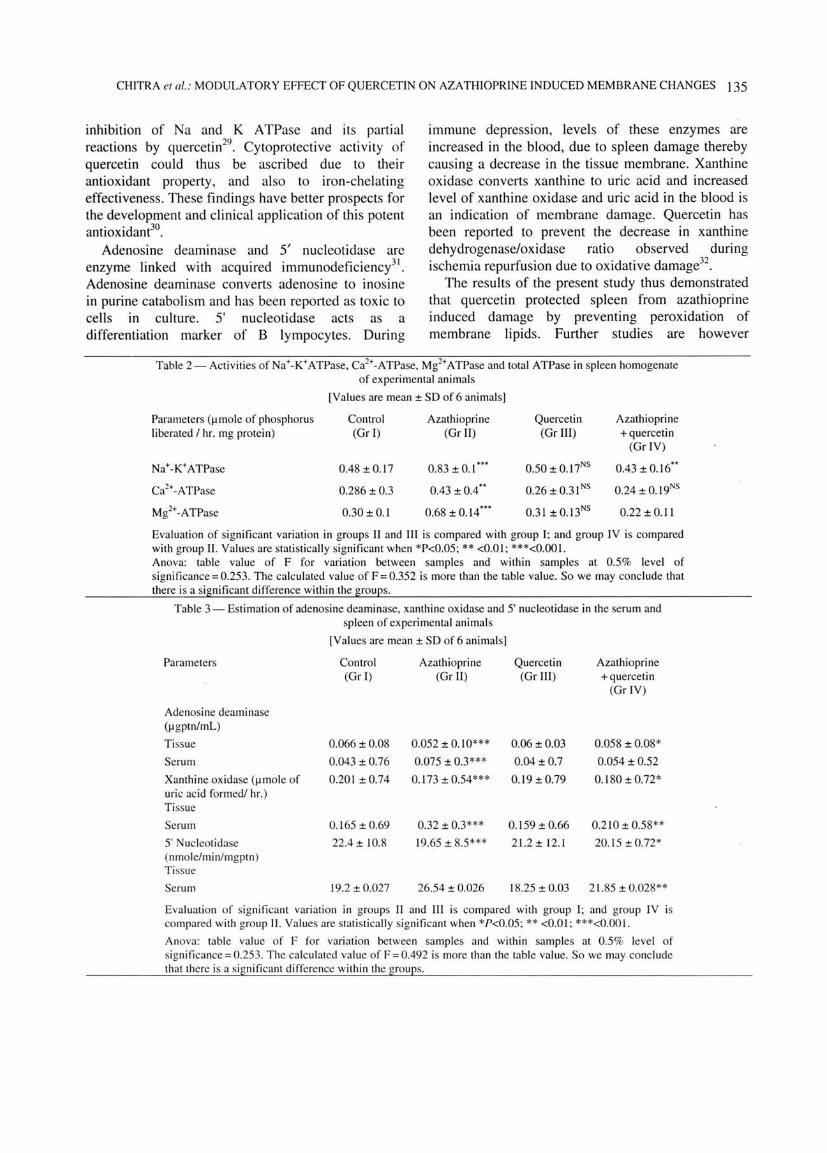
globulin ratio increased as compared to control (Table 1). No significant changes were noted in quercetin treated animals as compared to control animals. In group IV there was a decrease in albumin/globulin ratio as compared to group II animals. After azathioprine treatment, there was a depletion in yglobulin levels as compared to control (Figs 1,2). In group IV animals, globulin level increased with a relative decrease in albumin/globulin ratio as compared to animals treated with azathioprine alone. In azathioprine treated animals, total protein also decreased as compared to control animals. However, in group IV mice there was an increase in total protein levels as compared to group II animals.
In group II animals membrane ATPase activities in spleen increased as compared to normal control (Table 2). Quercetin supplementation caused a decrease of membrane A TPases as compared to animals treated with azathioprine alone.
Group II animals showed an increased level of
marker enzymes in serum with concomittant decrease in spleen tissue (Table 3). Quercetin treatment caused a decrease in serum enzyme levels and slight increase in tissue as compared to animals treated with azathioprine alone.
The results obtained indicated azathioprine induced toxicity in terms of increase in membrane A TPase activity, a known marker for membrane damage by azathioprine. Azathioprine has been reported to affect the functional activity of the spleen cells in vivo20
.
Decrease in protein in azathioprine treated mice may be due to decrease in intra-mitochondrial A TP concentration21
• Mitochondrial membrane carrier for external A TP requires a transmembrane potential gradient which could be abolished by increase in mitochondrial Ca2
+. Therefore sufficient A TP may not be available to drive mitochondria protein synthesis. Protein synthesis may be further inhibited in uncoupled mitochondria because of failure to import proteins. Since the processing of some
c B T H
Fig. I- Agarose slide gel electrophoresis of serum proteins of control and experimental animals [C- control ; B- azathioprine treated; T-quercetin treated; and H- azathioprine and quercetin treated]
Table I- Estimation of protein content and albumin and globulin ratio in the spleen and serum of experimental animals
[Values are mean± SD of 6 animals in each experiment]
Parameters Control Azathioprine (Gr II) Quercetin Azathioprine + (Gr I) (Gr lll) quercetin
(Gr IV)
Total protein 6.8 6.1·· 6.9NS 7.3 ...
(mg/g tissue)
Albumin globu lin ratio 0.88 1.34 .. 0.90 0.92·· (percentage)
Evaluation of significant variation in groups II and lii are compared with group I; and group IV is compared with group II. Values are statistically significant when *?<0.05: ** <0.01: ***<0.001. Anova: table value of F for variation between samples and within samples at 0.5% level of significance= 0.253. The calculated value ofF= 2.96 is more than the table value. So we may conclude that there is a significant difference within the rou s.
134 INDIAN J EXP BIOL, FEBRUARY 2001
cytoplasmically synthesised proteins depends on a transmembrane potential, uncoupling would prevent processing. Lack of cytoplasmic proteins could depress mitochondrial translation rates. Membrane bound Na+K+ ATPase is known as the sod ium pump and is involved in pumping out Na+ cations in exchange for K+. This ATPase is active on ly in the presence of both Na+ and K+ ions22
.
Significant Mg2+ dependent ATPase actiVIty is present in many tissues. However, its role in Mg2+ transport is not established. Mg2+ dependent hydrolysis of A TP may be coupled to a variety of cellular processes such as control of passive membrane permeability, regulation of oxidative phosphorylation in mitochondria23 and storage of catecholamines, serotonin, nucleotides and enzymes.
Increase in membrane ATPase and Ca2+ influx into cytoplasm is an indication of membrane damage7
•
Borle24 has reported that Ca2+ ion sensitive ATPase
(a) Control
FRACT ION G/OL REFERENCE RANGE
ALBUMIN 4·6 . 9 + 3 . 2 3.2'l. - TO - ~.31.
ALPHA I 4 . 6+ 0.3 0.1 % -TO- 0 . 4 Z 11LPHA 2 14.1• 1.0 0 . 4 ~- !.01. BETA 20.8 · . 0. 0. 5'l. - TO - 1.11. GAI>IMA 13.7 .. 0.9 0. 71. -TO- 1. 71.
TOTHL 6.8 6.5 -TO- 8 . 0
A.'G 0 . 88
(c)
FRACTION " G/OL REFERENCE R,: /'1..
ALBUMIN 50. 1+ 3 .0 3 . :Z 'l. -TO- :) .31. ALPHA l 7 . 7• 0.5 0 .1% - TO - e! . 4Y. ALPHA 2 13.b+ 0.8 0.41. -TO- 1.07. ~ETA 17. 9+ 1.1 0.~%. -TO- 1.11. GAMMA 10. 7 + 0.~6 0. 71. -TO- 1. 71.
TOTAL 6.9 - 6,, -TO- 8.0
A/G o . g 0
located in the intestinal brush border may play a fundamental role in cellular uptake of calcium. It has been proposed by Birge et al. 25 that this basal membrane Ca2+ A TPase mediates the energy dependent transport of calcium across the serosal membranes of intestinal cell. Increase in membrane ATPase act1v1ty was prevented by quercetin treatment. Fewtell and Gromperts26 have reported that quercetin is an inhibitor of Ca2+ influx and exocytosis in rat peritoneal mast cells. Lang and Racker7 have reported the effects of quercetin in mitochondrial ATPase and energy linked reactions in submitochondrial particles. Quercetin in group IV mice showed a decrease in ATPase levels which is in agreement with previous reports27 that quercetin is an inhibitor of membrane bound ATPase. It also confirms that quercetin interacts with Ca2+ and Mg2+ ATPase of sarcoplasmic reticulum as reported earlier28
. Kurikiy and Racker have further reported the
(b)
FRACTION " ALBUHIN :)7.3+ ALPHA 1 ::..2+ ALPHA 2 1 4.1+ BETA 16.8+ GAMIIA 6.::.+
TOTAL
A/0 1.34
(d)
FRACTION " ALBUMIN 48.0+ ALPHA 1 b. 7-+-ALPHA 2 1~.9· BETA 20.9+ GAMMA B . ~+
TOTAL
A/G 0 . 94:
Azathioprine
G/DL
3 . > 0.J 0.9 1.0 0.4
6.1-
Azathioprine + Quercetin
G/OL
3. ~ 0., 1.2
"' 0.6
7 .3
REFERENCE RANGE
3.27. -TO- ~ . :n
0,1 1. -TO- 0 , 41. 0.41. -TO- 1.07. 0.:):1. -lO- l.lX 0. 71. -TO- 1. 7X
6. > -TO- 8.0
REFERENCE RANGE
3.21.. - TO- :..3X 0 . 1 1. -TO- 0 . 47, 0.41. -TO- 1.0X 0.~1. -TO- 1.17. 0.n:. -TO- 1..71.
6 .> -TO- 8.0
Fig. 2- Agarose slide gel electrophoresis of serum proteins of control and experimental animal s.
CHITRA era/.: MODULATORY EFFECT OF QUERCETIN ON AZATHIOPRINE INDUCED MEMBRANE CHANGES 135
inhibition of Na and K ATPase and its partial reactions by quercetin29
. Cytoprotective activity of quercetin could thus be ascribed due to their antioxidant property, and also to iron-chelating effectiveness. These findings have better prospects for the development and clinical application of this potent antioxidant30
.
Adenosine deaminase and 5' nucleotidase are enzyme linked with acquired immunodeficiency31
.
Adenosine deaminase converts adenosine to inosine in purine catabolism and has been reported as toxic to cells in culture. 5' nucleotidase acts as a differentiation marker of B lympocytes. During
immune depression, levels of these enzymes are increased in the blood, due to spleen damage thereby causing a decrease in the tissue membrane. Xanthine oxidase converts xanthine to uric acid and increased level of xanthine oxidase and uric acid in the blood is an indication of membrane damage. Quercetin has been reported to prevent the decrease in xanthine dehydrogenase/oxidase ratio observed during ischemia repurfusion due to oxidative damage32
.
The results of the present study thus demonstrated that quercetin protected spleen from azathioprine induced damage by preventing peroxidation of membrane lipids. Further studies are however
Table 2- Activities of Na+-K+ ATPase, Ca2+-ATPase, Mg2+ATPase and total ATPase in spleen homogenate of experimental animals
[Values are mean± SD of 6 animals]
Parameters (}.I mole of phosphorus Control Azathioprine Quercetin Azathioprine liberated I hr. mg protein) (Gr I) (Gr II) (Gr III) +quercetin
(Gr IV)
Na+-K+ ATPase 0.48 ± 0.17 0.83 ± 0.1··· 0.50 ± 0.17NS 0.43 ±0.16 ..
Ca2+-ATPase 0.286 ± 0.3 0.43 ± 0.4 .. 0.26 ± 0.3 INS 0.24 ± 0. I 9NS
Mg2+ -ATPase 0.30 ± 0.1 0.68 ± 0.14··· 0.31 ±0.13NS 0.22 ± 0.11
Evaluation of significant variation in groups II and III is compared with group I; and group IV is compared with group II. Values are statistically significant when *P<0.05; ** <0.01; ***<0.001. Anova: table value of F for variation between samples and within samples at 0.5% level of significance= 0.253. The calculated value ofF= 0.352 is more than the table value. So we may conclude that there is a significant difference within the groups.
Table 3- Estimation of adenosine deaminase, xanthine oxidase and 5' nucleotidase in the serum and spleen of experimental animals
[Values are mean± SD of 6 animals]
Parameters Control Azathioprine Quercetin Azathioprine (Gr I) (Gr II) (Gr III) +quercetin
(Gr IV)
Adenosine deaminase (}Jgptn/mL)
Tissue 0.066 ±0.08 0.052 ± 0.1 0*** 0.06 ±0.03 0.058 ± 0.08*
Serum 0.043 ± 0.76 0.075 ± 0.3*** 0.04 ± 0.7 0.054 ± 0.52
Xanthine ox idase (}.I mole of 0.201 ± 0.74 0.173 ± 0.54*** 0.19 ± 0.79 0.180 ± 0.72* uric acid formed/ hr.) Ti ssue
Serum 0. 165 ± 0.69 0.32 ± 0.3*** 0.159 ± 0.66 0.210 ± 0.58**
5' Nucleotidase 22.4 ± 10.8 19.65 ± 8.5*** 21.2± 12.1 20.15 ± 0.72* (nmole/min/mgptn) Ti ssue
Serum 19.2 ± 0.027 26.54 ± 0.026 18.25 ± 0.03 21.85 ± 0.028**
Evaluation of significant variation in groups II and Ill is compared with group I; and group IV is compared with group II. Values are stati stically significant when *P<0.05: ** <0.01: ***<0.001.
Anova: table value of F for variation between samples and within samples at 0.5% level of significance= 0.253. The calculated value ofF = 0.492 is more than the table val ue. So we may conclude that there is a significant difference within the groups.

136 INDIAN J EXP BIOL. FEBRUARY 2001
necessary to understand the mechanism of quercetin modulation azathioprine effect.
References I Bach J F, Metabolism of azathioprine, J Am Med, 222 ( 1972)
1458. 2 Woods J E, Dewerd J H, Johnson W J & Anderson C F,
Splenectomy in renal transpl antation. J Am Med Assoc, 21 8 ( 1971 ) 1430.
3 Ireland P, Abdur Rashid, Franz Yon Lichtenberg, Tito Cavallo, John P. Merill , Liver di sease in kidney transplant patients receiving aza thioprine, Arcl1s lllf Med, 132 (1 973) 29.
4 Nogueira J R & Freedman M A, Acute pancreatiti s as a complication of imuran therapy in regional enti riti s, Gastroellferology, 62 ( 1972) I 040.
5 Brooke B N, Hoffmann D C & Swarbrick E T , Azathioprine fo r Crohn's disease, Lancet, 2 ( 1969) 6 12.
6 Ba rs, Nakayama, Galvez, Antiox idati ve acti vity of fl avono ids in various systems of lipid perox idation, J ?harm Phannacol, 50 (5) (1 994) 56 1.
7 Lang D N & Raiker E, Effects of quercetin and F1 inhibited on mitochondrial ATPase and energy linked reactions in submitochondrial part ic les, Biochim Biophys Acta, 333 ( 174) 180.
8 Bramley P M & Pridham J B, The re lative antioxidant act ivities of pl ant deri ved polypheno lic fl avonoids, Free Radic Res, 22 (4) (1 995) 375.
9 Gandhi R K, Quercetin is a scavenger of superox ide anions, /nt J Toxicol Occup Environ Health , VI ( I) (1991) 190.
10 Johann Hofmann , He iner H, Fiebig, Bernd R, Winterhalter, Dietmar P, Berger & Hans Grunicke, Enhancement of the anti proli fe rative acti vity o f c isdiammine dichloropl atinum (ii ) by quercetin , lnt J Cancer, 45 ( 1990) 536.
II Harold Varley, Pract Clin Biochem, IV (1993) 967. 12 Lowry 0 H, Roserbrough N J, Farr A L & Randall R S,
protein determination with the fo lin's reagent, J Bioi Chem, 193 ( 195 1) 265 .
13 Ba nting S L in membrane and ion transport fro m biobehavioural bases of coronary heart di sease, Vo l I, edited by T M Dembroski , T H Schmidt & G Blumcher (Wiley lnterscience, London) 1970, 257.
14 Ohni shi T, Scizuki T , Suzuki Y & Ozawa K A, presence of enzy me system in mammalian ti ssues, clin exp pharm physio/, 9(1982)2 13.
15 Hjerten S & Pan H, Purification and characteri sation o f two forms of a low affinity Ca2+ ATPase from erythrocyte membranes, Biochim Biophys Acta, 728 ( 1983) 28 1.
16 Hoki n Le, Dhal T L,Deukprea J D, Dikon J E, Hackney J F & Perdue J E, Stud ies on the characterization o f the sodiumpotass ium transport adenosine triphosphatase, J Bioi Chem,
248 ( 1973) 2593. I 7 Homodlarehe F & Bach J F Dardenem, in vitro inhibition of
prostaglandin produc tio n by azathioprine and 6 mercaptopurine, Prostagland, 35 (4) (1 988) 479 .
18 Kalckar H M, Differential spectrophotometry of purine compounds by means of specific enzymes, J Bioi Chem, 167, (1947b) 46 1.
19 Georges Van De n Berghe, Chri sti an Van Pottelsberghe & He nri-Gery Hers, A kinetic study of the soluble 5' nucleotidase of rat liver, Biochem J, 162 (1 977) 611.
20 Dianzani M U, The content o f adenosine po lyphosphates in fatty livers. Biochem J , 65 ( 1957) 11 6.
2 I Schatz G & Butow R A, How are proteins imported into mitochondri a, Cell , 32 (1 983) 3 16.
22 Smith E K M & Welt L G, Acti vation o f human erythrocyte membrane ATPase by low concentrations of sodium and potass ium, Am J Physiol, 220 ( I 971 ) 119.
23 Chao D L S & Dav is E J, Studies on the roJe of Mg2+ and the Mg2+ stimulated adenosine triphosphatase in oxidative phosphorylation, Biochemis11y, II (1 972) 1943 .
24 Borle A B, Ro le o f Ca2+ ATPase in Ca2+ uptake in intestinal brush border epithelium, Anm1 Rev Physiol. 36 ( 1974) 36 1.
25 Birge J R, Gilbert H R & Avio li L V, Intestinal calcium transport. The ro le o f sodium, Science. 176 ( 1972) 168.
26 Fewte ll C M S & Gro mperts B D, Quercetin : A novel inhibitor o f Ca2+ influx and exocytosis in rat peritoneal mast cells, Biochim Biophys Acta , 469 (1977) 52.
27 Suo llina E M, Buchsbaum M & Racker E, The effec t of fl avonoids on aerobic g lycolys is and gro wth of tumor ce ll s, Cancer Res, 35 ( 1975) 1865.
28 Shoshan V & Maclennan D H, Quercetin interaction with the (Ca2+ + Mg2+) ATPase o f sarcoplasmic reticulum, J Bioi Chem, 256 (198 1) 887.
29 Kuriki Y & Racker E, Inhibition o f (Na+, K+) adenosine triphosphatase and its parti al reac tion by quercetin. Biochim Biophys Acta , 15 ( 1976) 4951.
30 Morell , Lescoat G, Cognet P, Sergent 0 . Pasdelout N. Bri ssot P, Cillard P & Cillard J , Antiox idant and iron che lating acti vities of fl avonoids catechin, quercetin and diosmetin on iron loaded rat hepatocyte c ultures, Biochem Plw m wco/, 45( 1) (1 993) 13.
3 I Hitzig W H, in Immuno logic di sorders in infants and children, edited by E.R. Stiehm & Y. Fulginit i (Saunders) 1973, pp 300.
32 Sanhueza J, Valdes J, Ca mpos R, Garido A & Valenzuela A. Changes in the xanthine dehydrogenase xanthine ox idase ratic in the rat kidney subjected to ische mi a - repurfusion .stress · pre venti ve effec t o f some fl avonoids, Res Commun Chen Pat hoi Pharmacol, 78 (2) ( I 992) 2 11.


















