
Model peptide studies of sequence repeats derived from the intracrystalline biomineralization...
12
Model Peptide Studies of Sequence Repeats Derived from the Intracrystalline Biomineralization Protein, SM50. II. Pro,Asn-Rich Tandem Repeats Bo Zhang Guangzhao Xu John Spencer Evans* Laboratory for Chemical Physics, New York University, 345 E. 24th Street, New York, NY 10010 Received 31 January 2000; accepted 2 May 2000 Abstract: In the biomineralization process, a number of Pro-rich proteins participate in the formation of three-dimensional supramolecular structures. One such protein superfamily, the Pro,Gly-rich sea urchin intracrystalline spicule matrix proteins, form protein–protein supramo- lecular assemblies that modify the microstructure of the inorganic mineral phase (calcite) within embryonic sea urchin spicules and adult sea urchin spines. These proteins represent a useful model for understanding Pro sequence usage and the resulting generation of extended or “open” structures for protein–protein and/or protein– crystal recognition. In the sea urchin spicule matrix protein, SM50 (Strongylocentrotus purpuratus), there exists an unusual 20-residue Pro,Asn-con- taining repeat, OPNNPNNPNPNNPNNPNNPNPO, which links the upstream 15-residue C-termi- nal domain and the downstream 211-residue b-spiral repeat domain. To define the structural preferences of this 20-residue repeat, we created a 20-residue N- and C-terminal “capped” peptidomimetic of this sequence. Using far-uv CD dichroism, CH a and a- 15 N conformational shifts, 3 J NH-CHa coupling constants, sequential d NN(i, i 1 1) rotating frame nuclear Overhauser effect connectivities, d aN(i, i 1 1) /d NN(i, i 1 1) intensity ratios, amide temperature shift coefficients, amide solvent exchange, and simulated annealing refinement protocols, we have determined that this 20-residue repeat motif adopts an extended “twist” structure consisting of turn- and coil-like regions. These findings are consistent with previous studies, which have shown that Pro-rich tandem repeats adopt extended, flexible structures in solution. We hypothesize that this 20-residue repeat may fulfill the role of a mineral-binding domain, a protein–protein docking domain, or as an internal “molecular spacer” for the SM50 protein during spicule biocomposite formation. © 2000 John Wiley & Sons, Inc. Biopoly 54: 464 – 475, 2000 Keywords: nmr; sea urchin embryo; sea urchine spicule; biomineralization; proline repeats; self-assembly INTRODUCTION Proline-rich sequence regions occur in a wide variety of proteins. 1–4 The most interesting of these are the Pro-containing repeat sequences, which facilitate in- termolecular protein–protein association. These repet- itive sequences can be classified into three groupings: Pro–Pro pairs, short Pro-containing repeats, or Pro- Correspondence to: John Spencer Evans; jse@dave-edmunds. dental.nyu.edu Contract grant sponsor: National Science Foundation Contract grant number: CAREER MCB 95-13250, MCB 98- 16703, and DMR 99-01356 Biopolymers, Vol. 54, 464 – 475 (2000) © 2000 John Wiley & Sons, Inc. 464
Transcript of Model peptide studies of sequence repeats derived from the intracrystalline biomineralization...

Model Peptide Studies ofSequence Repeats Derivedfrom the IntracrystallineBiomineralization Protein,SM50. II. Pro,Asn-RichTandem Repeats
Bo ZhangGuangzhao Xu
John Spencer Evans*Laboratory for Chemical
Physics,New York University,
345 E. 24th Street,New York, NY 10010
Received 31 January 2000;accepted 2 May 2000
Abstract: In the biomineralization process, a number of Pro-rich proteins participate in theformation of three-dimensional supramolecular structures. One such protein superfamily, thePro,Gly-rich sea urchin intracrystalline spicule matrix proteins, form protein–protein supramo-lecular assemblies that modify the microstructure of the inorganic mineral phase (calcite) withinembryonic sea urchin spicules and adult sea urchin spines. These proteins represent a useful modelfor understanding Pro sequence usage and the resulting generation of extended or “open”structures for protein–protein and/or protein–crystal recognition. In the sea urchin spicule matrixprotein, SM50 (Strongylocentrotus purpuratus), there exists an unusual 20-residue Pro,Asn-con-taining repeat,OPNNPNNPNPNNPNNPNNPNPO, which links the upstream 15-residue C-termi-nal domain and the downstream 211-residueb-spiral repeat domain. To define the structuralpreferences of this 20-residue repeat, we created a 20-residue N- and C-terminal “capped”peptidomimetic of this sequence. Using far-uv CD dichroism, CHa anda-15N conformational shifts,3JNH-CHa coupling constants, sequentialdNN(i, i 1 1) rotating frame nuclear Overhauser effectconnectivities,daN(i, i 1 1)/dNN(i, i 1 1) intensity ratios, amide temperature shift coefficients, amidesolvent exchange, and simulated annealing refinement protocols, we have determined that this20-residue repeat motif adopts an extended “twist” structure consisting of turn- and coil-likeregions. These findings are consistent with previous studies, which have shown that Pro-rich tandemrepeats adopt extended, flexible structures in solution. We hypothesize that this 20-residue repeatmay fulfill the role of a mineral-binding domain, a protein–protein docking domain, or as aninternal “molecular spacer” for the SM50 protein during spicule biocomposite formation.© 2000 John Wiley & Sons, Inc. Biopoly 54: 464–475, 2000
Keywords: nmr; sea urchin embryo; sea urchine spicule; biomineralization; proline repeats;self-assembly
INTRODUCTION
Proline-rich sequence regions occur in a wide varietyof proteins.1–4 The most interesting of these are the
Pro-containing repeat sequences, which facilitate in-termolecular protein–protein association. These repet-itive sequences can be classified into three groupings:Pro–Pro pairs, short Pro-containing repeats, or Pro-
Correspondence to:John Spencer Evans; [email protected]
Contract grant sponsor: National Science FoundationContract grant number: CAREER MCB 95-13250, MCB 98-
16703, and DMR 99-01356Biopolymers, Vol. 54, 464–475 (2000)© 2000 John Wiley & Sons, Inc.
464

containing multiple tandem repeats.1–4 The short re-petitive Pro-rich sequences have the general form(XP)n or (XPY)n,3 and are believed to adopt apolyproline type II structure.1–4 Protein domainscomprised of short repetitive Pro-rich sequences tendto form extended, rigid structures or “sticky arms,”1–4
which permit rapid and reversible protein–protein as-sociations.3 Conversely, the tandem repeats containlonger Pro-rich sequences (5–8 residues),1–4 and arebelieved to form extended, flexible structures thatpermit functional protein–protein assemblies.3 In gen-eral, the strength of the intermolecular associationsthat occur between Pro-containing repeats and otherprotein domains can be correlated to the molecularrigidity, pyrrolidine ring hydrophobicity, and the highpotential for Pro carbonyl oxygen hydrogen bondingwithin these repeat domains.3
Given that Pro-containing repeat regions may actas potential protein interaction domains, it is not sur-prising that some biomineralization-specific proteinsalso contain Pro-rich domains. In many organisms,the biomineralization process utilizes a number ofPro-rich polypeptides to form three-dimensional su-pramolecular structures in the extracellular ma-trix.5–11 One such protein superfamily, the Pro,Gly-rich sea urchin intracrystalline spicule matrix proteinsform protein–protein supramolecular assemblies thatmodify the microstructure of the inorganic mineralphase (calcite) within embryonic sea urchin spi-cules8–12and adult sea urchin spines.2 These proteinsassemble into a fibrillar array of concentric polymer“sleeves” that are embedded within the calcite mineralphase.8–12 What is so intriguing about this class ofproteins is that they convey fracture-resistant proper-ties to the inorganic mineral phase.13–15Hence, theseproteins represent a useful model for understandingPro sequence usage and the resulting generation ofextended or “open” structures for protein–protein rec-ognition. In addition, the spicule matrix proteins couldalso serve as useful mimetic models for developingbiologically based self-assembling materials.
An analysis of the available amino acid sequencesfor the sea urchin spicule matrix proteins reveals thepresence of unique Pro-rich motifs.8–12 Of these, themost unusual is the 20-residue Pro,Asn repeat foundin the intracrystalline spicule matrix protein, SM50.9
This repeat links the upstream 15-residue C-terminaldomain, ORFNRPRMLQEADALAOCOOH, andthe downstream 211-residue consensusb-spiral repeatelastic domain (Figure 1). This 20-residue repeat(hereafter referred to as the PNNP repeat) is intriguingfor two reasons. First, only two types of amino acidsare utilized: Asn represents a putative site for side-chain hydrogen bonding,16 and Pro represents a puta-tive site for hydrophobic interaction.3 Second, theSM50 PNNP repeat is homologous to theONNO-rich repeats found in the aragonite-specific proteinnacrein,17 and to theOPXPO andOPXXPO repeatsof the aragonite-specific protein, Lustrin A.18 It hasbeen postulated that these aragonite-specific proteinsutilize these repeats for mineral interaction17 or as aninternal molecular spacer domain.18 Hence, the struc-ture of the PNNP repeat may play an important role inSM50 function within the context of spicule mineral-ization.
Due to sequence repetition and absence of amidehydrogen atoms, it is difficult to deduce the secondarystructure of Pro-rich domains using nmr spectroscopy.Moreover, many of the proteins that contain repetitivePro-rich domains often aggregate in solution,3,19mak-ing the task of structure determination even moreproblematic. Hence, a special strategy is required: theuse of native-sequence derived peptide models (pep-tidomimetics) to obtain Pro-rich repeat sequences sec-ondary structure preferences. The use of peptidomi-metics has already advanced our understanding ofprotein folding and the relationship between primarysequence and secondary structure.20–28Recently, thisstrategy was utilized to establish secondary structurepreferences for the b-turn OGVGGRO andOGMGGQO repeats in the SM50 intracrystallineprotein.19
FIGURE 1 Domain map of theStrongylocentrotus purpuratusspicule matrix protein, SM50. Thismap represents the mature isoform (prepro leader sequence deleted).
Model Peptide Studies of Sequence Repeats465

In this report, we detail a multidisciplinary ap-proach for determining the secondary structure pref-erences for Pro-rich sequence repeats. The strategyinvolves (a) the use of CD and1H-nmr spectroscopyto determine the global conformation of a repeat se-quence peptidomimetic derived from the parent pro-tein, (b) site-selective15N isotope-filtered nmr meth-ods19,29–31to establish conformational preferences atspecific locations within the peptidomimetic, and (c)simulated annealing (SA) molecular dynamics struc-ture refinement32 to generate conformational ensem-bles based upon available nmr restraints. We utilizethis combined strategy to determine the conforma-tional preferences for the following SM50 peptidomi-metic:
Na-acetyl-PNNPNNPNPNNPNNPNNPNP-Ca-
amide (sequence position 411–430)
which represents the PNNP repeat in toto. As part ofthe site-selective nmr strategy, we incorporateda-15N-Pro at positions 7, 9, 12, 15 (underlined), and,a,g-15N2 Asn at positions 8, 10, 11 (boldfaced) duringthe syntheses of separate peptides. The placement ofa-15N-Asn anda-15N-Pro within the peptidomimeticpermits the unambiguous determination of (a) long-range heteronuclearJ couplings (3JNH-CHa)19,30,31
that reflect Asnf torsion angle preferences, (b) Asnrotating frame nuclear Overhauser effect1H–1H re-laxation transfer,19 and (c) sequence-specific Asn andPro 1H,15N chemical shifts. As described in this re-port, our experimental and simulated annealing dataindicate that the PNNP repeat motif adopts an ex-tended twist structure consisting of turn- and coil-likeregions. These findings are consistent with the ex-tended, flexible structures adopted by Pro-rich tandemrepeats.1–4
MATERIALS AND METHODS
Peptide Synthesis, Purification, andSample Preparation
The Na-acetyl, Ca amide SM50 20-mer and Na-acetylL-(PPPPPPPPPP)-Ca-amide 10-mer peptides were synthe-sized using an Applied Biosystems 431A Peptide Synthe-sizer and Fmoc MBHA resin (Fmoc: 9-flourenylmethyoxy-carbonyl; Novabiochem/Calbiochem). Typical peptide syn-thesis runs were carried out at the 0.2 mmole level using theApplied Biosystems “FastMoc 0.25” HBTU/HOBt/NMPprotocol [HBTU: 2-(1H-benzotriazole-1-yl)-1,1,3,3-tetra-methyluronium hexafluorophosphate; HOBt: N-hydroxy-benzotriazole; NMP:n-methylpyrrolidone; Applied biosys-
tems Technical Notes, November 1993], double couplingprocedure, and acetic anhydride capping at each couplingstep. To avoid possible side-chain modification during syn-thesis, Ng-Trityl-Na-Fmoc-Asn (abbreviated as Fmoc-Asn-Trt) was utilized in 20-mer synthesis runs. In individuallysynthesized peptides, Fmoca-15N Asn (98% 15N, Cam-bridge Isotope Labs) was incorporated at positions 8, 10,and 11 in the 20-mer. Similarly, Fmoca-15N Pro (98%15N,Cambridge Isotope Labs) was incorporated at positions 7, 9,12, and 15 in the 20-mer, and at position 5 in the 10-mer.The completed peptides were treated with acetic anhydridein NMP/DIEA to create the Na-acetyl derivative, thendeprotected and cleaved (reaction time5 3 h, at 25°C) fromthe resin using a cleavage cocktail (15 mL/g resin) contain-ing 90% v/v triflouroacetic acid, (TFA), 2.5% v/v water,2.5% v/v ethanedithiol, 2.5% v/v phenol, and 2.5% v/vthioanisole.19 The thiol scavengers were utilized to preventoxidation and enhance trityl removal. The reaction mixturewas filtered under reduced pressure. The crude peptideswere separately dissolved in deionized distilled water, ex-tracted three times with diethylether, and then concentratedand lyophilized. Peptides were analyzed using a Perkin-Elmer API III LC/MS Electrospray Mass Spectrometer; theexperimental molecular weight of each 20-mer was deter-mined to be 2.207 kDa, in agreement with the theoreticalmolecular weight of 2.207 kDa. Similarly, the 10-mer ex-hibited a molecular weight of 1.032 kDa, which is in agree-ment with the theoretical molecular weight of 1.0312 kDa.Peptide purification involved the use of a Waters DeltaPakC18 reverse-phase high performance liquid chromatography(HPLC) column, using 0.1% TFA/water mobile phase andeluting with a 80% acetonitrile/0.1% TFA/water linear gra-dient. Peptide elution was monitored at 230 nm. IndividualHPLC fractions were analyzed using electrospray massspectrometry; the 20-mer was found to be.96% in purityand free of sidechain protection. The 10-mer was found tobe.94% in purity. For nmr studies, purified peptides werethen dissolved in 1 mM Na2HPO4 in deionized distilledwater, pH 7.4, containing 10% v/v deuterium oxide (99.9%atom D, Cambridge Isotope Labs) and 10 micromolard4-TSP. Parallel nmr samples were also dissolved in 1 mMNaH2PO4, pH 4.0. Both peptides were soluble in water ateach pH value; we believe this is due to the presence of Asnin the 20-mer, and the solvent accessibility of the polar Proa-carbonyl anda-amide groups in both the 10- and 20-merpeptides. For each15N-labeled peptide, the final concentra-tion was 4.0 mM in a volume of 450mL. CD studies utilized1 mM concentrations in the same buffer for unlabeled20-mer and 10-mer samples. At these concentrations, tur-bidity measurements at 380 nm revealed no evidence ofpeptide aggregation for all samples; this was also confirmedby analyses of nmr proton chemical shifts and line widths,and the absence of long-rangedSC-SCrotating frame nuclearOverhauser effect connectivities.
CD and NMR ExperimentsCD spectra of the 20-mer and 10-mer peptides were ob-tained using a AVIV 60 CD Spectrometer, running 60DS
466 Zhang, Xu, and Evans

software version 4.1t. The 20-mer and 10-mer samples werescanned from 180 to 240 nm at 5°C, using 1 nm bandwidthand a scan rate of 1 nm/s. The spectrometer was previouslycalibrated withd-10-camphorsulfphonic acid. A total of 4scans were acquired for each peptide. Mean residue ellip-ticity [uM] is expressed in deg cm2 dmol21 per mole pep-tide.33–35
All nmr experiments were performed on a VarianUNITY 500 spectrometer as detailed elsewhere.19 With theexception of the proton amide temperature shift experi-ments, all reported nmr experiments were conducted at 278K. Proton assignments were obtained using “excitationsculpting” versions of the two-dimensional 2D z-PFG-“clean” total correlation spectroscopy (TOCSY), compen-sated spin–lock rotating frame nuclear Overhauser effectspectroscopy (ROESY) and transverse ROESY (T-ROESY)experiments, and15N v1-filtered selective excitationROESY experiments.19,31 The proton chemical shifts and3JNH-CHa coupling constants for15N-labeled Asn residueswere obtained using the “excitation sculpting” PFG15N-filtered “in-phase/antiphase” TOCSY experiments.29,30 Noobservable cross-relaxation crosstalk artifacts were detectedin theJ-coupling spectra.30 Direct observe15N experiments,involving gated1H WALTZ-16 composite pulse decoupling(1 kHz field strength) during acquisition, were utilized toobtain 15N chemical shifts for Asn and Pro residues. Nu-clear magnetic resonance data were processed using FE-LIX2.30 software (MSI/Biosym Technologies, Inc). Rele-vant nmr acquisition and processing parameters are pro-vided in the figure legends or in the legend to Table II.Using TOCSY or T-ROESY experiments at 278, 283, 288,293, and 298 K, 20-mer amide proton temperature coeffi-cients were determined from the slope of the temperature vsamide proton chemical shift curves for each Asn group-ing.36 Residue-specific amide temperature shift coefficientswere obtained for Asn8, Asn10, and Asn11 using PFG15Nv1-filtered selective excitation TOCSY or ROESY experi-ments. Temperature gradients are expressed in units ofppb/K with a negative sign indicating an upfield shift uponwarming. Temperature calibration of the VT unit was de-termined prior to experimentation using neat methanol overa temperature range of 273–320 K.
Structure Refinement
Three-dimensional structures of the 20-mer were obtainedusing the Crystallography and NMR System (CNS) soft-ware package (version 0.9a; Yale University)32 followingthe reported protocol.37–39 As demonstrated below in theResults section, Asn8, Asn10, and Asn11 are conformation-ally representative of theOPNPO, OPNNO, andONNPO residue clusters, respectively, within the 20-mer.Moreover, the Pro residues within the 20-mer represent aconformationally equivalent ensemble (see Results, below).Hence, we approached the issue of assigning experimentalrestraints in the following manner. Backbonef torsionangle,a-CH, and Asna-NH chemical shift restraints ob-tained for Asn8, Asn10, and Asn11 were applied to other
corresponding Asn residues in the 20-mer (i.e., Asn8
5 Asn19; Asn10 5 Asn2, Asn5, Asn10, Asn13, and Asn16;Asn11 5 Asn3, Asn6, Asn14, and Asn17). Thea-CH chem-ical shift restraints for Pro7, Pro9, and Pro12 were applied toother corresponding Pro residues in the 20-mer (i.e., Pro7
5 Pro18; Pro12 5 Pro4, Pro15). Pro1 and Pro20 were ex-cluded from the structure calculations. A total of 44 re-straints (12J-coupling and 32 chemical shift) were utilizedfor structure refinement as described in the next paragraph.
The generated structures (total5 30) were obtainedusing the simulated annealing molecular dynamics codeprovided in the CNS software package.32 The refinementprotocol involved a high temperature dynamics stage (start-ing at 2000 K with heating to 50,000 K in 1000 steps),followed by primary (2000 steps) and secondary (3000steps) quenching stages, with termination at a final simula-tion temperature of 300 K. The simulated annealing runconcluded with conjugate-gradient minimization to conver-gence. The final accepted structures (total5 2) were thenobtained using the “accept” algorithm and the experimentalnmr restraints. These accepted structures were then aver-aged into a single structure using a least-squares fittingroutine, and finally refined with conjugate-gradient energyminimization to convergence.
RESULTS
Circular Dichroism
The far-uv CD spectra of the SM50 Na-acetyl PNNPCa-amide peptidomimetic at 278 K in 1 mMNa2HPO4, pH 7.4, are presented in Figure 2. Theabsorption curve for the 20-mer exhibits a negativeband at 204 nm. This feature has been equated withthe presence of some secondary structure in equilib-rium with extended or coil structures34,35,40; specifi-cally, this feature is found in CD spectra obtained forpeptides that containb-turn and/orb-hairpin struc-tures.34,35,40,41For comparison, we synthesized Na-
FIGURE 2 CD spectra of SM50 Na-acetyl PNNP Ca-amide (grey) and Na-acetyl-(P)10-C
a-amide (black) pep-tidomimetics at 278 K in 1 mM Na2HPO4, pH 7.4.
Model Peptide Studies of Sequence Repeats467
acetyl-(P)10-Ca-amide and performed parallel CD ex-
periments on this polyproline type II peptidomimetic.As shown in Figure 2, Na-acetyl-(P)10-C
a-amide ex-hibits a single intense negative band at 216 nm; notethat this minimum is red shifted from the 195 nmminimum obtained for short Poly-L-Pro type II helicesfound in globular proteins.33 Clearly, on the basis ofthe CD spectra, the 20-mer peptidomimetic does notshare the same conformational characteristics as thePoly-L-Pro peptide, despite the fact that nearly 40% ofthe 20-mer sequence contains proline. Hence, we con-clude that the 20-mer sea urchin PNNP sequence doesnot adopt any detectable Poly-L-proline type II struc-ture under the conditions utilized in this study.
NMR Spectroscopy
Table I lists the amino acid assignments for the 20-mer SM50 peptidomimetic at 278 K in 90% water/10% deuterium oxide, pH 7.4. For comparison, wealso present nmr data obtained for Na-acetyl-(P)10-Ca-amide. Since both polypeptides are capped, thenmr findings (e.g.,1H chemical shifts, ROESY crosspeaks,3JNH-CHa couplings) obtained for each peptideat pH 7.4 were found to be identical to those obtainedat pH 2.0. Not surprisingly, we observe spectral over-lap for Pro and Asn NHa and CHa resonances that
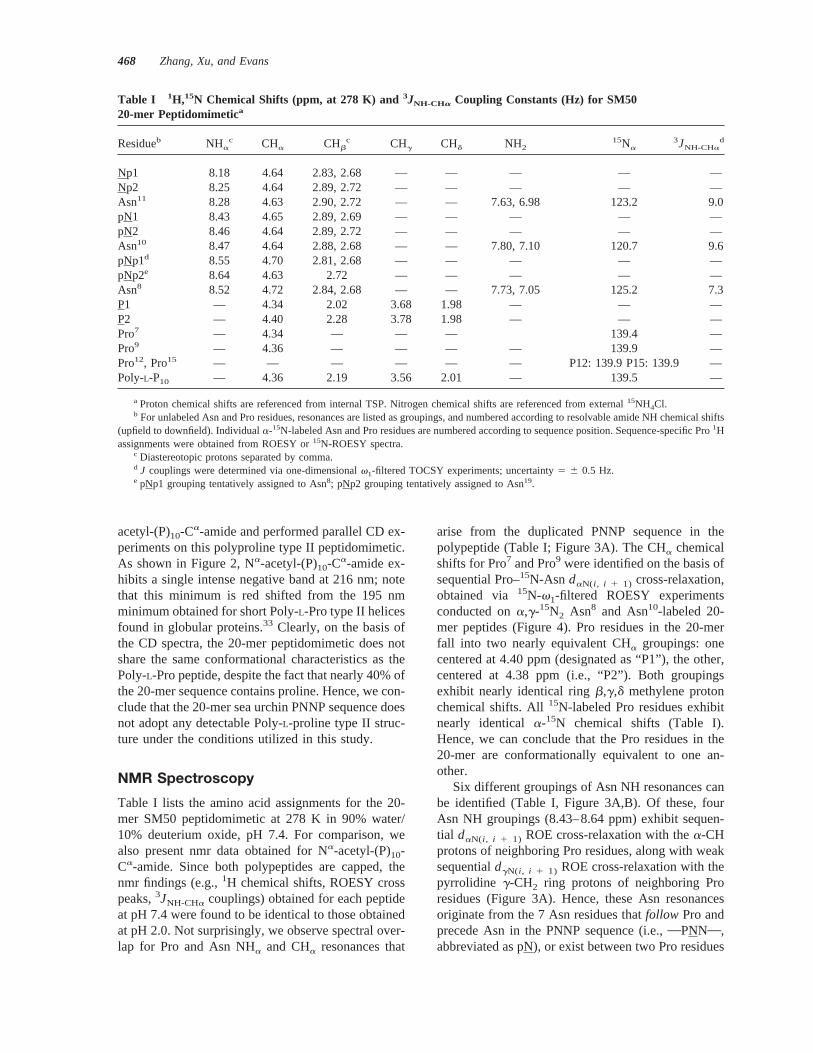
arise from the duplicated PNNP sequence in thepolypeptide (Table I; Figure 3A). The CHa chemicalshifts for Pro7 and Pro9 were identified on the basis ofsequential Pro–15N-Asn daN(i , i 1 1) cross-relaxation,obtained via 15N-v1-filtered ROESY experimentsconducted ona,g-15N2 Asn8 and Asn10-labeled 20-mer peptides (Figure 4). Pro residues in the 20-merfall into two nearly equivalent CHa groupings: onecentered at 4.40 ppm (designated as “P1”), the other,centered at 4.38 ppm (i.e., “P2”). Both groupingsexhibit nearly identical ringb,g,d methylene protonchemical shifts. All15N-labeled Pro residues exhibitnearly identical a-15N chemical shifts (Table I).Hence, we can conclude that the Pro residues in the20-mer are conformationally equivalent to one an-other.
Six different groupings of Asn NH resonances canbe identified (Table I, Figure 3A,B). Of these, fourAsn NH groupings (8.43–8.64 ppm) exhibit sequen-tial daN(i , i 1 1) ROE cross-relaxation with thea-CHprotons of neighboring Pro residues, along with weaksequentialdgN(i , i 1 1) ROE cross-relaxation with thepyrrolidine g-CH2 ring protons of neighboring Proresidues (Figure 3A). Hence, these Asn resonancesoriginate from the 7 Asn residues thatfollow Pro andprecede Asn in the PNNP sequence (i.e.,OPNNO,abbreviated as pN), or exist between two Pro residues
Table I 1H,15N Chemical Shifts (ppm, at 278 K) and3JNH-CHa Coupling Constants (Hz) for SM5020-mer Peptidomimetica
Residueb NHac CHa CHb
c CHg CHd NH215Na
3JNH-CHad
Np1 8.18 4.64 2.83, 2.68 — — — — —Np2 8.25 4.64 2.89, 2.72 — — — — —Asn11 8.28 4.63 2.90, 2.72 — — 7.63, 6.98 123.2 9.0pN1 8.43 4.65 2.89, 2.69 — — — — —pN2 8.46 4.64 2.89, 2.72 — — — — —Asn10 8.47 4.64 2.88, 2.68 — — 7.80, 7.10 120.7 9.6pNp1d 8.55 4.70 2.81, 2.68 — — — — —pNp2e 8.64 4.63 2.72 — — — — —Asn8 8.52 4.72 2.84, 2.68 — — 7.73, 7.05 125.2 7.3P1 — 4.34 2.02 3.68 1.98 — — —P2 — 4.40 2.28 3.78 1.98 — — —Pro7 — 4.34 — — — 139.4 —Pro9 — 4.36 — — — — 139.9 —Pro12, Pro15 — — — — — — P12: 139.9 P15: 139.9 —Poly-L-P10 — 4.36 2.19 3.56 2.01 — 139.5 —
a Proton chemical shifts are referenced from internal TSP. Nitrogen chemical shifts are referenced from external15NH4Cl.b For unlabeled Asn and Pro residues, resonances are listed as groupings, and numbered according to resolvable amide NH chemical shifts
(upfield to downfield). Individuala-15N-labeled Asn and Pro residues are numbered according to sequence position. Sequence-specific Pro1Hassignments were obtained from ROESY or15N-ROESY spectra.
c Diastereotopic protons separated by comma.d J couplings were determined via one-dimensionalv1-filtered TOCSY experiments; uncertainty5 6 0.5 Hz.e pNp1 grouping tentatively assigned to Asn8; pNp2 grouping tentatively assigned to Asn19.
468 Zhang, Xu, and Evans
(OPNPO, abbreviated as pNp). The other two AsnNH resonance groupings (8.18–8.30 ppm), which donot exhibit any short-range or long-range rOe cross-relaxation transfer to any Pro ring protons (Figure3A), originate from the remaining Asn residues thatpreceded Pro in the 20-mer sequence and follow Asn(i.e.,ONNPO). This Asn grouping will be referred toasNp. To identify the Asn groupings more precisely,we obtained the15N-filtered proton chemical shifts fora,g-15N2-labeled Asn8, Asn10, and Asn11 residues(Table I, Figure 4), using15N-filteredv1-TOCSY andROESY experiments. Technically speaking, Asn8,Asn10, and Asn11 are representative of pNp, pN, and
Np Asn groupings, respectively. As shown in Table I,we assigned the groupings as follows. Asn11 NHa,CHa, and b-CH2 proton chemical shifts correspondwith those obtained for two Np Asn groupings (sub-categorized as Np1 and Np2). The Asn10 NHa, CHa,and b-CH2 proton chemical shifts correspond withtwo pN groupings (subcategorized as pN1, pN2), andthe Asn8 NHa, CHa, and b-CH2 proton chemicalshifts correspond to the remaining Asn groupings(i.e., pNp1, pNp2). Note that the chemical shifts ob-tained for Asn8 closely correspond with those ob-tained for pNp1; hence, this grouping is tentativelyassigned to Asn8, and by extension, pNp2 is assignedto Asn19 near the C-terminus. These findings confirmthat Asn8, Asn10, and Asn11 are conformationallyrepresentative of similar Asn residues (i.e., pNp, pN,Np) located throughout the 20-mer.
CHa and a-15N Conformational Shifts. Proton CHa
chemical shifts are sensitive to peptide backbone con-formation, and the deviation of these chemical shiftsfrom “random coil” values42,43 (referred to as CHaconformational shifts orDdHa, where DdHa 5dHa, observed2 dHa “random coil”) have been used toqualitatively deduce secondary structure in peptidesand proteins.19,26,27,36,40,42,43It is generally observedthat b-turn anda-helical residues are upfield shiftedrelative to the random coil state; betab-strand resi-dues are downfield shifted.19,26,27,36,40,42,43Using theWishart et al. CHa chemical shift index (database of70 proteins),42 and implementing corrections for Proand Asn residues preceding Pro in the sequence,44 wecalculated theDdHa values for Asn,Pro groupings andfor a-15N-Asn residues in the 20-mer peptidomimetic(Figure 5A). Here, it can be seen that Np1, Np2,Asn11, and pNp1/Asn8 all exhibit upfield shifted CHaconformational shifts that exceed the random coilthreshold of 0.1 ppm.19,26,27,36,40,42,43The presence ofupfield shifted CHa values indicates that these Asnresidues are conformationally similar to either thea-helix orb-turn secondary structure classification. Incontrast, the pN1, pN2, pNp2/Asn19, Asn10, and allPro residues exhibit downfield CHa conformationalshifts that do not exceed the random coil threshold of0.1 ppm. Clearly, the conformational preferences ofpN, Asn19, and Pro residues are most likely that of anextended or coil conformational state, as opposed to atrue b-strand conformational state.
In addition to the CHa conformational shifts, wecalculated the a-15N conformational shifts42–44
(DdNa) at residues Asn8, Asn10, Asn11, Pro7, Pro9,Pro12, and Pro15 within the PNNP 20-mer, and Pro5
within poly-L-(P)10 (Figure 5B) using the Wishart etal. a-15N chemical shift index42 and corrections for
FIGURE 3 (A) NHa—CHa and (B) NHa fingerprint re-gions of 1H–1H “excitation sculpting” z-PFG-T-ROESYspectrum of SM50 20-mer peptidomimetic, pH 7.4. Thecarrier is centered on the water resonance. Acquisition pa-rameters included a relaxation delay of 1 s, spectral window5 5200 Hz, “hard” 90°5 10.5 ms, “hard” 180°5 21 ms;“soft” selective 180°5 3.2 ms; 128 transients/experimentwere acquired; 2048 and 256 complex points were collectedin t1 andt2, respectively. The spin–lock period (180° pulsetrain) was 250 ms.Z-gradient parameters:G1 5 14 G/cm,G2 5 6 G/cm, each with a duration of 1 ms with 500msstabilization time. The hypercomplex phase-sensitivemethod was utilized for processing both 2D spectra, withzero-filling in the F2 dimension. NSEQ5 sequential ROEs.“D” 5 diagonal peaks. Proton chemical shifts are refer-enced from internald4-TSP.
Model Peptide Studies of Sequence Repeats469

Pro and Asn residues that precede Pro in the se-quence.43 Because there is considerable overlap be-tween a-helix and b-strand a-15N conformationalshift categories,42,43 a-15N conformational shifts canonly be used to qualitatively estimate the presence ofnonrandom coil conformational states within a pep-tide or protein. As shown in Figure 5B, the confor-mational shifts Asn8, Asn11, and all Proa-15N spins inthe 20-mer and P5 in the 10-mer are downfield shifted$4 ppm relative to the random coil state.42,43 Notethat Asn10 exhibits aa-15N conformational shift ofapproximately 2 ppm. Note thata-15N conformationalshifts .3 ppm have been observed in structured do-mains of globular proteins.44 Collectively, the1H and15N conformational shift data qualitatively confirmthe presence of a heterogenous structure consisting ofnonrandom coil and extended conformational states.
3JNH-CHa Coupling Constants.Coupling constantshave been widely utilized for the estimation of local
conformational preferences in peptides. The3JNH-CHa
coupling constants are linear averages over all con-formers; they are independent of the particular chem-ical environment and measure only thef angle pop-ulation at each position in the main chain.19,45Due tochemical shift overlap, we were unable to obtainresolvable Asn3JNH-CHa couplings from1H–1H ex-clusive correlated spectroscopy (COSY) or doublequantum filtered (DQF)-COSY experiments. How-ever, by taking advantage of the placement ofa,g-15N2-labeled Asn residues, we were able to determineAsn3JNH-CHa coupling constants for Asn8, Asn10, andAsn11 in the peptidomimetic (Table I).29,30 3JNH-CHa-coupling values obtained for Asn8, Asn10, and Asn11
would be presumed to be representative of pNp, pN,and Np groupings, respectively, in the repetitive 20-mer. The deviation of3JNH-CHa coupling constantsfrom random coil values (DJ) was also determined(Figure 5C). In general,J couplings larger than ran-dom coil values (1 DJ) are indicative ofb-strand
FIGURE 4 Representative one-dimensional z-PFG15N v1-filtered selective excitation ROESYspectra of the SM50 20-mer peptidomimetic (a,g-15N labeled Asn10), pH 7.4. Transmitter offsetwas applied on-resonance with the solvent signal. The one-dimensional spectrum utilized 8 Kcomplex data points, 1024 scans, spectral window5 5200 Hz, and a Lorentzian apodization windowfunction of 2 Hz was applied. Proton chemical shifts are referenced from internald4-TSP.Sequential cross-relaxation transfer gives rise to in-phase (i.e., positive) peaks for Asn10 magneti-zation, and out-of-phase (i.e., inverted) peaks for Pro9 and Asn11 magnetization.
470 Zhang, Xu, and Evans

conformation, whereasJ couplings smaller than ran-dom coil values (2 DJ) representa-helix conforma-tion.21,45 Beta turn values can be either1 DJ or 2DJ.21,45 As shown in Figure 5C, Asn10 and Asn11
have1 DJ values. 1 Hz; however, theDJ value forAsn8 is , 1 Hz and is negative. These qualitativeestimates suggest that the Asn10 and Asn11 exist in aconformational state that reflect the presence ofb-strand,b-turn, or “coil” conformations. However,
Asn8, which is flanked on either side by Pro residues,possesses a smallDJ value and may exist in anextended state.
rOe Connectivities.SequentialdNN(i , i 1 1) ROEmagnetization transfer was observed betweenNp1,Np2 Asn amide proton groupings (8.18–8.25ppm) and pN1,pN2 Asn amide proton groupings(8.43–8.50 ppm) in the 20-mer (Figure 3B). However,we did not observe any sequential ROE magnetizationtransfer involving either the pNp1 or pNp2 groupings.Using 15N-v1-filtered PFG-ROESY experiments, wealso detected sequentialdNN(i , i 1 1) ROE magnetiza-tion transfer between Asn10 and Asn11 (Figure 4).
Earlier nmr studies of peptides have establishedthat sequentialdNN(i , i 1 1) Overhauser magnetizationtransfer is a diagnostic indicator ofb-turn folding inpeptides.19–27 However, other studies have shownthat the short-range sequentialdNN(i , i 1 1) connec-tivities can also arise from conformations available toflexible random coil peptides,26,46 we can utilize thed
aN(i , i 1 1)/dNN(i , i 1 1) intensity ratio to qualitatively
distinguish between random coil and structured do-mains.19,26,46 In general,daN(i , i 1 1)/dNN(i , i 1 1)
intensity ratios# 1.4 are indicative of random coilregions.26,46 Using the Np and pN grouping ROEcross-peak volumes obtained from our PFG-T-ROESY spectra (Figure 3A,B), we find that thedaN(i ,
i 1 1)/dNN(i , i 1 1) intensity ratio for N3 N amidegroupings within the 20-mer is 4.55, a value that isthree times larger than the random coil threshold of1.4.
Medium- and long-rangeda-a(i , j ), dSC-SC(i , j ) ROEcross peaks were not observed in our T-ROESY spec-tra. In addition, we did not observe long-rangedNN(i ,
j ) ROE cross peaks between pN and Np amide group-ings (Figures 3B and 4). These observations rule outthe presence ofa-helix and b-hairpin structures.Hence, from the Overhauser data, we conclude thatNp1,Np2 and pN1,pN2 Asn residues exist in a con-formational state that may consist of shortb-turn-likeconformations in the vicinity of eachONNO region.Theseb-turn segments would be assumed to be inconformational equilibria with a random coil state.
Amide Temperature Coefficients and Solvent Ex-change. It has been shown that “random coil” orextended peptides exhibit amide temperature shift co-efficients in the range of26.6 to 29.0 ppb/K inaqueous solution.36 For the PNNP 20-mer, we foundthat the coefficients obtained for Asn8, Asn10, andAsn11 were 28.5, 29.0, and28.8 ppb/K, respec-tively. In addition, amide temperature shift coeffi-cients were also estimated for Asn amide groupings
FIGURE 5 (A) DdHa values for the SM50 20-mer pep-tidomimetic in 90% water, 10% D2O, pH 7.4, 278 K.Negative values represent upfield shifts, positive valuesrepresent downfield shifts. No corrections for terminal res-idues effects have been made.43 Corrected CHa chemicalshift values were utilized for all Pro and Xaa–Pro nearestneighbors (where Xaa5 Asn).43 Proton chemical shiftswere referenced from internald4-TSP. (B)DdNa values forthe SM50 20-mer peptidomimetic in 90% water, 10% D2O,pH 7.4, 278 K. Negative values represent upfield shifts,positive values represent downfield shifts. Corrected15Na
chemical shift values were utilized for all Pro and Xaa–Pronearest neighbors (where Xaa5 Asn).43 No corrections forterminal residues effects have been made.43 Nitrogen-15chemical shifts were referenced from external15NH4Cl. (C)Calculated difference (DJ) between experimental and ran-dom coil3JNH-CHa values for the SM50 20-mer peptidomi-metic in 90% water, 10% D2O at 278 K, pH 7.4. Note: forA, B, C please refer to the Results section for definitions ofshorthand abbreviations used to identify1H and 15N spingroupings.
Model Peptide Studies of Sequence Repeats471
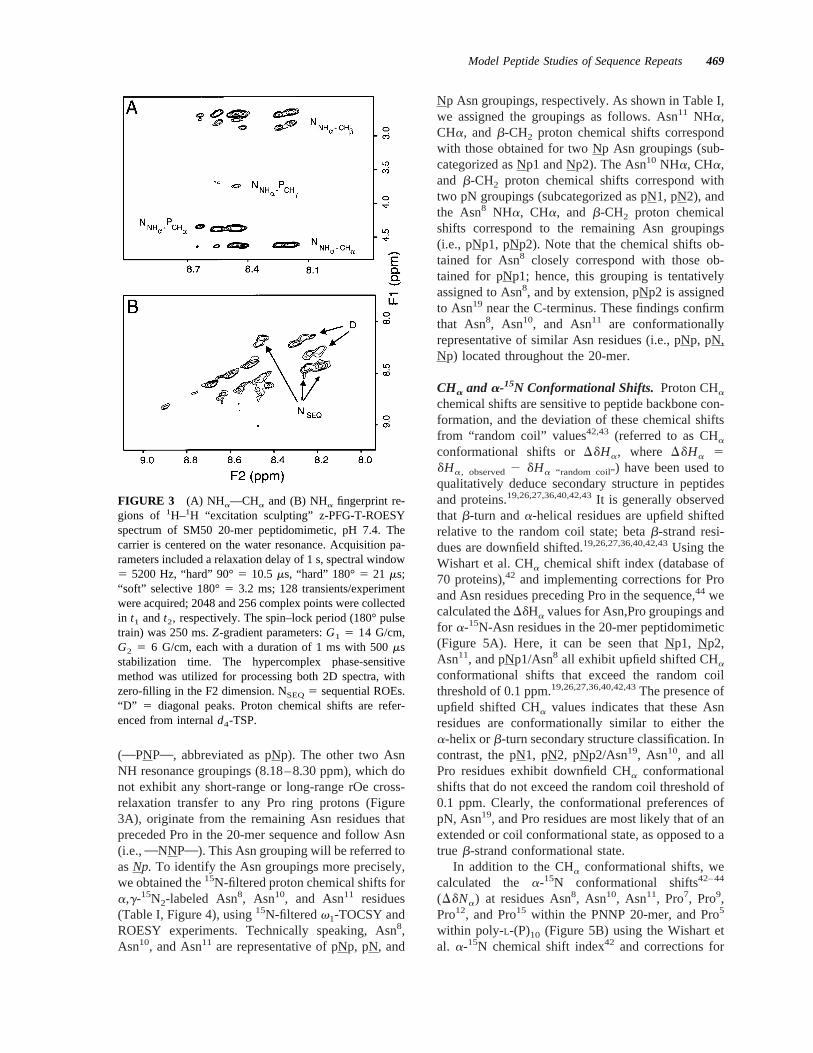
(Figure 3A,B). These values were found to be in therange of 28.0 to 29.0. These values suggest thatnone of the Asn amide hydrogens are participating inweak intramolecular hydrogen bonding with carbonyloxygens elsewhere along the polypeptide backbone.To confirm these findings, we estimated the presenceor absence of solvent exchange between H2O and thebackbone amide protons in the PNNP 20-mer, usingPFG-T-ROESY experiments.19 The exchange regimethat we can measure is on the nmr timescale (milli-seconds to seconds); any exchange processes thatoccur on a slower timescale will not be observed. Asshown in Figure 6, the pNp2 (i.e., Asn19) amidegroupings exhibit shielding from solvent chemicalexchange. Given that the amide temperature shift co-efficients for these groupings are large, we concludethat the observed amide proton solvent shielding doesnot arise from intrastrand hydrogen bonding. It isplausible that the amide shielding arises fromthe presence of flanking hydrophobic pyrrolidinerings.
Structure Refinement
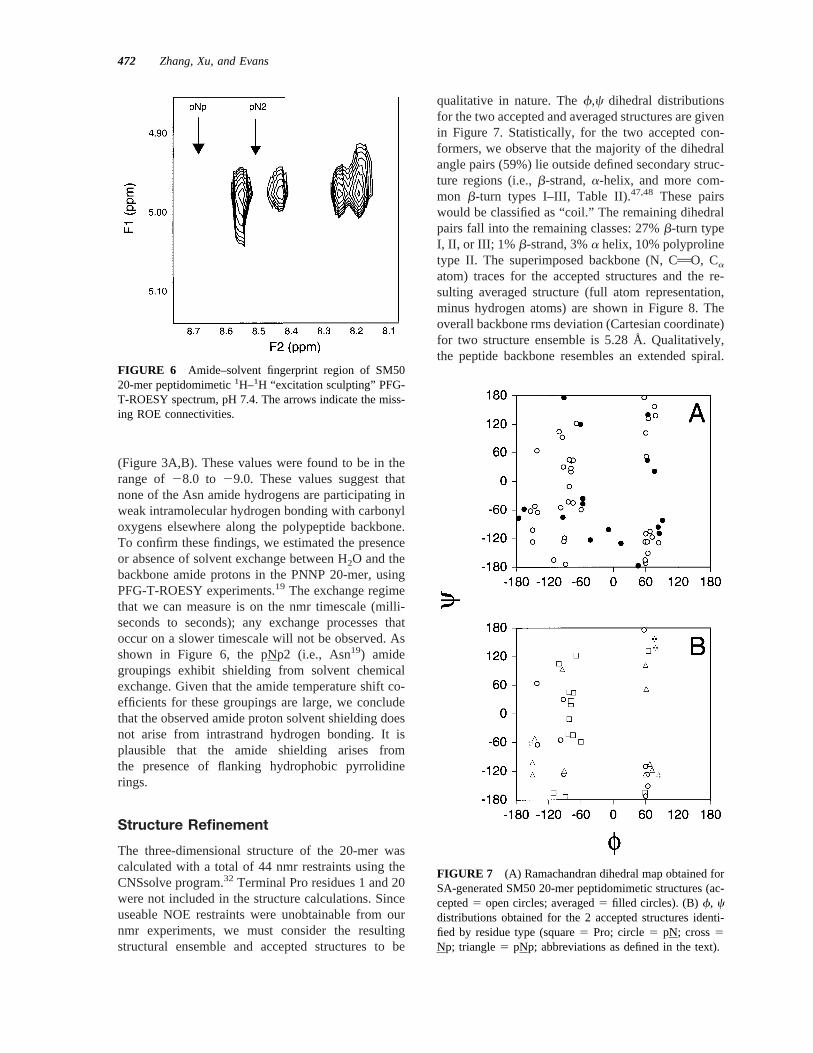
The three-dimensional structure of the 20-mer wascalculated with a total of 44 nmr restraints using theCNSsolve program.32 Terminal Pro residues 1 and 20were not included in the structure calculations. Sinceuseable NOE restraints were unobtainable from ournmr experiments, we must consider the resultingstructural ensemble and accepted structures to be
qualitative in nature. Thef,c dihedral distributionsfor the two accepted and averaged structures are givenin Figure 7. Statistically, for the two accepted con-formers, we observe that the majority of the dihedralangle pairs (59%) lie outside defined secondary struc-ture regions (i.e.,b-strand,a-helix, and more com-mon b-turn types I–III, Table II).47,48 These pairswould be classified as “coil.” The remaining dihedralpairs fall into the remaining classes: 27%b-turn typeI, II, or III; 1% b-strand, 3%a helix, 10% polyprolinetype II. The superimposed backbone (N, CAO, Ca
atom) traces for the accepted structures and the re-sulting averaged structure (full atom representation,minus hydrogen atoms) are shown in Figure 8. Theoverall backbone rms deviation (Cartesian coordinate)for two structure ensemble is 5.28 Å. Qualitatively,the peptide backbone resembles an extended spiral.
FIGURE 7 (A) Ramachandran dihedral map obtained forSA-generated SM50 20-mer peptidomimetic structures (ac-cepted5 open circles; averaged5 filled circles). (B)f, cdistributions obtained for the 2 accepted structures identi-fied by residue type (square5 Pro; circle5 pN; cross5Np; triangle5 pNp; abbreviations as defined in the text).
FIGURE 6 Amide–solvent fingerprint region of SM5020-mer peptidomimetic1H–1H “excitation sculpting” PFG-T-ROESY spectrum, pH 7.4. The arrows indicate the miss-ing ROE connectivities.
472 Zhang, Xu, and Evans

This is consistent with our understanding that Pro-richtandem repeats form extended, flexible structures insolution.3 The averaged structure features Asn residuepairs that orient outward from the peptide backbone.Pro residues are known to induce kinks or bendswithin a sequence stretch,1–4 and these features areobserved in the vicinity of the Pro residues of thestructural ensemble (Figure 8); Overall, we find thatthese refined structures are qualitatively in agreementwith our nmr observations,viz: (a) A majority of theAsn–Asn sequence pairs exhibit sequential
dNN(i , i 1 1) ROEs anddaN(i , i 1 1)/dNN(i , i 1 1)
intensity ratios that are not consistent with a randomcoil structure, but are characteristic of a turn confor-mation.19–27 Note that turn-like structures are ob-served in Figure 8. (b) The 20-mer does not exhibitobservable intrastrand hydrogen bonding, as evi-denced by the absence of shielded Asn NHa protonsthat correlate with sequential Asn–AsndNN(i , i 1 1)
ROEs (Figures 3A,B and 6). We did not observe anyevidence of intrastrand hydrogen bonding within therefined structures obtained via simulated annealing.
Table II Major f,c Regionsa
Type f(i ) (deg) f(i 1 1) (deg) f(i 1 2) (deg) c(i ) (deg) c(i 1 1) (deg) c(i 1 2) (deg)
b-Strand 21206 45 N/A N/A 1306 30 N/A N/Aa-Helix 2556 20 N/A N/A 2556 20 N/A N/Ab-Turn type I NR N/A 2606 20 2906 20 N/A 2306 40 06 30b-Turn type II NR N/A 2606 20 806 30 N/A 1206 20 06 30b-Turn type III
NR N/A 2606 40 2606 40 N/A 2306 40 2306 20Polyproline type II 275 N/A N/A 145 N/A N/A
a Dihedral values and secondary structure classifications were obtained from Refs. 47 and 48. NR: nonreverse.
FIGURE 8 Conformational ensemble (left) and averaged structure (right) obtained from CNS-solve SA simulations of Na-acetyl-PNNPNNPNPNNPNNPNNPNP-Ca-amide. The conformationalensemble (3 structures, consisting of the 2 accepted structures plus the averaged structure) isrepresented in ribbon form; the averaged structure on the right is shown in ribbon representation forthe backbone and heteroatom representation for the side chains.
Model Peptide Studies of Sequence Repeats473

(c) The DJ values obtained for Asn8, 10, 11are con-sistent with conformational states that consist of mix-tures ofb-turn or “coil” structures.21,45Again, as seenin Figure 8, turn- and coil-like structures are presentin the accepted structures.
DISCUSSION
The SM50 20-mer PNNP repeat motif represents achallenging peptide for structure determination. How-ever, by using CD spectrometry, and selective isoto-pic incorporation and nmr filtering techniques, we areable to obtain qualitative and quantitative estimates ofthe conformational preferences adopted by the 20-merpeptidomimetic in aqueous solution. Based upon theavailable CHa and NHa conformational shifts, ROEmagnetization transfer connectivities,J-coupling con-stants, solvent exchange data, and simulated anneal-ing runs, we find that the PNNP sequence adopts anextended, twisted conformation consisting of a mix-ture of turn-like and coil-like regions (Figure 8), mostlikely interconverting with the random coil state. Thisproposed conformation is consistent with the confor-mational behavior of tandem repeat Pro-rich se-quences, and is inconsistent with the polyproline typeII conformation that is typical of short Pro repeats1–4
(Figure 2). Interestingly, protein database studies haverevealed that turn regions can experience conforma-tional interconversions, which in turn affects back-bone dynamics at the turn region.48 Given that turn-like regions (i.e.,ONNO) have been identifiedwithin the PNNP 20-mer, it is likely that our nmr dataset (ROEs,J couplings, chemical shifts) reflects thisconformational averaging. One would expect the 20-residue sequence to exhibit different backbone dy-namics within the 46 kDa SM50 protein, and as aresult, the conformational sampling experienced bythe PNNP domain within the SM50 protein will bedifferent, compared to the 20 residue peptide. Thus,due to the limited number of nmr restraints availableto us, and differences in backbone conformationalaveraging, we would advise the reader that our struc-tural findings regarding this 20-residue sequenceshould be considered qualitative at this point. Futureconformational studies conducted on the native SM50protein or protein fragments will either corroborate orrefute our peptidomimetic findings.
Although the function of the intracrystalline SM50protein has not been fully established, recent experi-mental evidence has provided more insight into therole of this protein during the spicule mineralizationprocess. Recent studies indicate that the spicule min-eralization process is complex: using electron micros-copy and Fourier transform infrared, it has been dem-
onstrated that amorphous calcium carbonate is ini-tially deposited within the spicule and then slowlytransforms into calcite.49,50During the mineralizationprocess, SM50 has been identified at two locations inthe spicule: the surface of the mineral and occludedwithin the mineral phase.51 Clearly, SM50 is involvedin the process of matrix assembly, and may also haveinteractions with the developing calcium carbonatemineral phase. It is possible that the SM50 protein,like other multifunctional mineral-specific proteinssuch as Lustrin A,18 amelogenin,6,7 and nacrein,17 ispartitioned into multifunctional domains, each ofwhich performs one or more specific molecular tasks.At least 4 different SM50 domains have been delin-eated: (a) The N-terminal domain, a portion of whichhas homology to the C-lectins51; (b) the;200 residueconsensus repeat region, a portion of which adopts arepeatingb-turn conformation similar to elastin andthe spider silk dragline protein19; (c) the PNNP re-peat; and (d) the charged C-terminal motif. Given thatthe proposed structure for the PNNP repeat featuresan extended twisted backbone from which Asn sidechains are accessible to the environment, we wouldlike to hypothesize some possible roles for the PNNPrepeat that are in accord with SM50 function: (a)Mineral recognition domain.—The Asn amide side-chain groups could possible form hydrogen bondswith carbonate groups at exposed interfaces of theamorphous calcium carbonate and/or calcite mineralphases in the developing spicule, thus allowing theSM50 protein to adsorb to the calcium carbonatephase(s). A similar role has been proposed for theONNGO repeat in the aragonite-specific nacreinprotein17; (b) Self-assembly domain—Given that Asnside chains are known to form hydrogen-bondingnetworks in globular proteins,16 and that the Pro pyr-rolidine ring is hydrophobic, it is conceivable that thePNNP repeat could act as a docking site for SM50–SM50 self-association, or SM50 association with an-other spicule matrix protein; (c)Molecular spacer—Itis possible that the 15-residue C-terminal domain andthe 200 amino acid consensus repeat domains ofSM50 require spatial separation from one another inorder to perform their particular roles in spicule ma-trix development. Thus, the extended PNNP sequencemay position each of these domains away from oneanother. Clearly, further experimental studies will berequired to determine the correct function of thePNNP repeat within the context of SM50 function.
This work was supported by the National Science Founda-tion (CAREER MCB 95-13250, MCB 98-16703, DMR99-01356). This paper represents contribution number 12from the Laboratory for Chemical Physics.
474 Zhang, Xu, and Evans

REFERENCES
1. Adzhubei, A. A.; Sternberg, M. J. E. J Mol Biol 1993,229, 472–493.
2. MacArthur, M. W.; Thornton, J. M. J Mol Biol 1991,218, 397–412.
3. Williamson, M. P. Biochem J 1994, 297, 249–260.4. Creamer, T. P. Proteins Struct Func Gene 1998, 33,
218–226.5. Lowenstam, H. A.; Weiner, S. On Biomineralization;
Oxford University Press, New York, 1989; pp 20–49.6. Moradian-Oldak, J.; Tan, J.; Fincham, A. G. Biopoly-
mers 1998, 46, 225–238.7. Moradian-Oldak, J.; Leung, W.; Fincham, A. G. J
Struct Biol 1998, 122, 320–327.8. Killian, C. E.; Wilt, F. C. J Biol Chem 1996, 271,
9150–9159.9. Katoh-Fukui, Y.; Noce, T.; Ueda, T.; Fujiwara, Y.;
Hashimoto, N.; Higashinakagawa, T.; Killian, C. E.;Livingston, B. T.; Wilt, F. H.; Benson, S. C.; Sucov,H. M.; Davidson, E. H. Devel Biol 1991, 145, 201.
10. Livingston, B. T.; Shaw, R.; Bailey, A.; Wilt, F. H.Devel Biol 1991, 148, 473–480.
11. Harkey, M. A.; Klueg, K.; Sheppard, P.; Raff, R. A.Devel Biol 1995, 168, 549–566.
12. Benson, S. C.; Wilt, F. H. In Calcification in BiologicalSystems; Bonucci, E., Ed.; CRC Press: Boca Raton, FL,1992; pp 157–178.
13. Berman, A.; Addadi, L.; Weiner, S. Nature 1993, 331,546–548.
14. Berman, A.; Hanson, J.; Leiserowitz, L.; Koetzle, T. F.;Weiner, S.; Addadi, L. Science 1993, 259, 776–779.
15. Berman, A.; Hanson, J.; Leiserowitz, L.; Koetzle, T. F.;Weiner, S.; Addadi, L. J Phys Chem 1993, 97, 5162–5170.
16. Le Questel, J.-Y., Morris, D. G., Maccallum, P. H.,Poet, R.; Milner-White, E. J. J Mol Biol 1993, 231,888–896.
17. Miyamoto, H.; Miyashita, T.; Okushima, M.; Nakano,S.; Morita, T.; Matsushiro, A. Proc Natl Acad Sci USA1996, 93, 9657–9660.
18. Shen, X.; Belcher, A. M.; Hansma, P. K.; Stucky,G. D.; Morse, D. E. J Biol Chem 1997, 272, 32472–32481.
19. Xu, G.; Evans, J. S. Biopolymers 1999, 49, 303–312.20. de Alba, E.; Jimenez, M. A.; Rico, M. J Am Chem Soc
1997, 119, 175–183.21. Liang, G.-B.; Rito, C. J.; Gellman, S. H. J Am Chem
Soc 1992, 114, 4440–4442.22. Dyson, H. J.; Rance, M.; Houghten, R. A.; Lerner,
R. A.; Wright, P. E. J Mol Biol 1988, 201, 161–200.23. Johnson, W. C.; Pagano, T. G.; Basson, T.; Madri,
J. A.; Gooley, P.; Armitage, I. M. Biochemistry 1993,32, 268–273.
24. Ramirez-Alvarado, M.; Blanco, F. J.; Niemann, H.;Serrano, L. J Mol Biol 1997, 273, 898–912.
25. Imperiali, B.; Spencer, J. R.; Struthers, M. D. J AmChem Soc 1994, 116, 1516–1517.
26. Maynard, A. J.; Sharman, G. J.; Searle, M. S. J AmChem Soc 1998, 120, 1996–2007.
27. Haque, T. S.; Gellman, S. H. J Am Chem Soc 1997,119, 2303–2304.
28. Evans, J. S.; Chiu, T.; Chan, S. I. Biopolymers 1994,34, 1359–1375.
29. Xu, G.; Evans, J. S. J Magn Reson Series A 1996, 123,105–110.
30. Xu, G.; Evans, J. S. J Magn Reson 1999, 138, 127–134.31. Xu, G.; Evans, J. S. J Magn Reson Series B 1996, 111,
183–185.32. Brunger, A. T.; Adams, P. D.; Clore, G. M.; Delano,
W. L.; Gros, P.; Grosse-Kunstleve, R. W.; Jiang, J.-S.;Kuszewski, J.; Nilges, M.; Pannu, N. S.; Read, R. J.;Rice, L. M.; Simonson, T.; Warren, G. L. Acta Cryst1998, D54, 905–921.
33. Sreerama, N.; Woody, R. W. Biochemistry 1994, 33,10022–10025.
34. Manning, M. C.; Illangasekare, M.; Woody, R. W.Biophys Chem 1998, 31, 77–86.
35. Perczel, A.; Hollosi, M.; Sandor, P.; Fasman, G. D. IntJ Pept Protein Res 1993, 41, 223–236.
36. Andersen, N. H.; Neidigh, J. W.; Harris, S. M.; Lee,G. M.; Liu, Z.; Tong, H. J Am Chem Soc 1997, 119,8547–8561.
37. Garrett, D. S.; Kuszewski, J.; Hancock, T. J.; Lodi,P. J.; Vuister, G. W.; Gronenborn, A. M.; Clore, G. M.J Magn Reson Ser B 1994, 104, 99–103.
38. Kuszewski, J.; Gronenborn, A. M.; Clore, G. M. JMagn Reson Ser B 1995, 107, 293–297.
39. Stein, E. G.; Rice, L. M.; Brunger, A. T. J Magn ResSer B 1997, 124, 154–164.
40. Blanco, F. J.; Rivas, G.; Serrano, L. Nature Struct Biol1994, 1, 584–590.
41. Gibbs, A. C.; Kondejewski, L. H.; Gronwald, W.; Nip,A. M.; Hodges, R. S.; Sykes, B. D.; Wishart, D. S.Nature Struct Biol 1998, 5, 284–288.
42. Wishart, D. S.; Sykes, B. D.; Richards, F. M. J Mol Biol1991, 222, 311–333.
43. Wishart, D. S.; Bigam, C. G.; Holm, A.; Hodges, R. S.;Sykes, B. D. J Biomol NMR 1995, 5, 67–81.
44. Braun, D.; Wider, G.; Wuthrich, K. J Am Chem Soc1994, 116, 8466–8469.
45. Smith, L. J.; Bolin, K. A.; Schwalbe, H.; MacArthur,M. W.; Thornton, J. M.; Dobson, C. M. J Mol Biol1996, 255, 494–506.
46. Fiebig, K. M.; Schwalbe, H.; Buck, M.; Smith, L. J.;Dobson, C. M. J Phys Chem 1996, 100, 2661–2666.
47. Ball, J. B.; Andrews, P. R.; Alewood, P. F.; Hughes,R. A. FEBS Lett 1990, 273, 15–18.
48. Gunasekaran, K.; Gomathi, L.; Ramakrishnan, C.;Chandrasekhar, J.; Balaram, P. J Mol Biol 1998, 259,1505–1516.
49. Beniash, E.; Aisenberg, J.; Addadi, L.; Weiner, S. ProcRoy Soc Lond B 1997, 264, 461–465.
50. Beniash, E.; Addadi, L.; Weiner, S. J Struct Biol 1999,125, 50–62.
51. Wilt, F. H. J Struct Biol 1999, 126, 216–226.
Model Peptide Studies of Sequence Repeats475


















