
Mineralogical Biosignatures and the Search for Life on Mars · Mineralogical Biosignatures and the...
20
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/11020286 Mineralogical Biosignatures and the Search for Life on Mars Article in Astrobiology · February 2001 DOI: 10.1089/153110701753593856 · Source: PubMed CITATIONS 84 READS 250 5 authors, including: Some of the authors of this publication are also working on these related projects: Genome enabled Investigation of S(0) Cycling in a Subterranean Microbial Ecosystem View project John W Moreau University of Melbourne 76 PUBLICATIONS 987 CITATIONS SEE PROFILE Clara S. Chan University of Delaware 56 PUBLICATIONS 1,583 CITATIONS SEE PROFILE All content following this page was uploaded by John W Moreau on 16 May 2014. The user has requested enhancement of the downloaded file.
Transcript of Mineralogical Biosignatures and the Search for Life on Mars · Mineralogical Biosignatures and the...

See discussions stats and author profiles for this publication at httpswwwresearchgatenetpublication11020286
Mineralogical Biosignatures and the Search for Life on Mars
Article in Astrobiology middot February 2001
DOI 101089153110701753593856 middot Source PubMed
CITATIONS
84
READS
250
5 authors including
Some of the authors of this publication are also working on these related projects
Genome enabled Investigation of S(0) Cycling in a Subterranean Microbial Ecosystem View project
John W Moreau
University of Melbourne
76 PUBLICATIONS 987 CITATIONS
SEE PROFILE
Clara S Chan
University of Delaware
56 PUBLICATIONS 1583 CITATIONS
SEE PROFILE
All content following this page was uploaded by John W Moreau on 16 May 2014
The user has requested enhancement of the downloaded file
ASTROBIOLOGYVolume 1 Number 4 2001copy Mary Ann Liebert Inc
Research Paper
Mineralogical Biosignatures and the Search for Life on Mars
JILLIAN F BANFIELD1 JOHN W MOREAU1 CLARA S CHAN1 SUSAN A WELCH1
and BRENDA LITTLE2
ABSTRACT
If life ever existed or still exists on Mars its record is likely to be found in minerals formedby or in association with microorganisms An important concept regarding interpretation ofthe mineralogical record for evidence of life is that broadly defined life perturbs disequi-libria that arise due to kinetic barriers and can impart unexpected structure to an abiotic sys-tem Many features of minerals and mineral assemblages may serve as biosignatures even iflife does not have a familiar terrestrial chemical basis Biological impacts on minerals andmineral assemblages may be direct or indirect Crystalline or amorphous biominerals an im-portant category of mineralogical biosignatures precipitate under direct cellular control aspart of the life cycle of the organism (shells tests phytoliths) or indirectly when cell surfacelayers provide sites for heterogeneous nucleation Biominerals also form indirectly as by-products of metabolism due to changing mineral solubility Mineralogical biosignatures in-clude distinctive mineral surface structures or chemistry that arise when dissolution andorcrystal growth kinetics are influenced by metabolic by-products Mineral assemblages them-selves may be diagnostic of the prior activity of organisms where barriers to precipitation ordissolution of specific phases have been overcome Critical to resolving the question ofwhether life exists or existed on Mars is knowing how to distinguish biologically inducedstructure and organization patterns from inorganic phenomena and inorganic self-organiza-tion This task assumes special significance when it is acknowledged that the majority of andperhaps the only material to be returned from Mars will be mineralogical Key Words MarsmdashMicroorganismsmdashBiosignaturesmdashMinerals Astrobiology 1 447ndash465
447
INTRODUCTION
THE FEW UNAMBIGUOUS MINERALOGICAL biosig-natures available today are mostly restricted
to minerals produced under direct cellular con-
trol (eg coccoliths and diatoms) or within com-plex mineralndashpolymer intergrowths (eg shells)Although highly diagnostic of life in terrestrialsamples their relevance to detection of life onMars may be minimal In the absence of clearly
1Department of Geology and Geophysics University of Wisconsin Madison WI2Naval Research Laboratory Stennis Space Center MSPresent address Departments of Earth and Planetary Science and of Environmental Science Policy and Man-
agement University of California at Berkeley Berkeley CA
identifiable remains of organisms a variety ofmineralogical biosignatures may serve to estab-lish that life existed in an environment These in-clude minerals formed as a direct or indirect con-sequence of microbial metabolism mineralassemblages templated by organic polymers andmodifications to mineral surfaces or mineral dis-tribution patterns
A subset of terrestrial microorganisms derivesmetabolic energy by catalysis of inorganic chem-ical reactions Microbes use inorganic and organicsubstrates to generate energy by coupling theproduction of ATP to enzymatic transport of elec-trons Numerous electron donor and acceptorpairs are available in the form of organic mole-cules minerals dissolved ions and metals andgases especially in environments characterizedby redox transitions (eg in sediments or at theinterface between basaltic rocks and oxidizingsurface-derived fluids) Microbial utilization ofinorganic substrates in energy generation cancause minerals to dissolve or to precipitate Thecharacteristics of biologically induced mineralproducts may bear testament to their biologicalorigin Minerals formed as metabolic by-productsare typically nanocrystalline owing to their rapidprecipitation from solutions that locally arehighly supersaturated At this time insufficientinformation is available to determine whether thecharacteristics of these minerals can be used asrigorous biosignatures
In addition to modifying the redox state ofaqueous ions microbial metabolism can impactthe composition of surrounding solutionsthrough a change in pH for example via the pro-duction of organic or inorganic complexingagents This phenomenon may induce precipita-tion of minerals with crystal structures crystalsizes defect structures and morphologies thatwould not be produced in the absence of biolog-ical activity Similarly complexation of metals byorganic ligands may promote dissolution of oth-erwise extremely insoluble phases Dissolutionand precipitation may be accompanied by bio-logically induced fractionation of trace and ma-jor elements andor isotopes
Cell surfaces and cell-derived polymers canpromote heterogeneous nucleation of mineralswith unusual characteristics In some cases theseprecipitates may preserve the morphologicalcharacteristics of the cell (microfossils) Althoughsuch morphological microfossils are difficult todistinguish from mineral aggregates produced by
inorganic processes trace element isotopic andmicrostructural features may provide clues thatin combination with morphological characteris-tics could indicate a biological origin
In general life on Earth has evolved to use com-pounds readily available from the environmentespecially from solutions However as the geo-chemical characteristics of an environment changesome metabolically required elements may be-come scarce and organisms may be driven to in-crease their proximity to mineral surfaces In ad-dition organisms attach to solid substrates forprotection against environmental extremes (egelectromagnetic radiation mechanical abrasiontemperature stress hazardous chemicals tem-perature shock desiccation etc) Close mi-crobendashmineral association can promote mineraldissolution and the enhanced release of biologi-cally useful constituents to solution Distinctivesurface compositional characteristics and etchingpatterns may result Although detailed surfacefeatures may preserve evidence of life consider-able research is needed to develop reliable crite-ria for recognizing such features in the rockrecord
EXAMPLES OF TERRESTRIAL MINERALBIOSIGNATURES WITH POSSIBLE
RELEVANCE TO MARS
On Earth a variety of mineral characteristicsrecord evidence of biological activity
Minerals whose phase indicates a biogenic origin
The formation of aragonite the high-pressureCaCO3 polymorph rather than calcite by algae(de Vrind de Jong and de Vrind 1997) and inshells (Falini et al 1996) is an example of biolog-ical activity leading to precipitation of a structurenot predicted to occur given ambient physicaland chemical conditions However the crystal-lization of aragonite at low temperature can oc-cur under some specific conditions via abioticpathways For example Renault and Jones (1997)observed a cyclic precipitation of calcite and ara-gonite in a hot spring travertine which they at-tributed to rapid degassing of CO2
Algae octocorallia and echinoderm tests areoften composed of high-Mg calcite and the shellsof molluscs are typically composed of lower-Mgcalcites (eg Dauphin 1999) In the case of echi-
BANFIELD ET AL448
noderms it has been suggested that an organicmatrix with calcium-binding ability is framed ina periodic three-dimensional structure made ofconcentric layers bridged by radial threads (Am-eye et al 1998) Yet both high- and low-Mg cal-cites are found in beach cements and are some-times inferred to form inorganically (Kneale andViles 2000)
Unexpected minerals also can form if biologi-cal activity leads to the creation of redox condi-tions not anticipated based on bulk solution pa-rameters Mackinawite (a tetragonal iron sulfide)is considered to be a biosignature for microbio-logically influenced corrosion of iron by sulfate-reducing bacteria Experimental databases indi-cate mackinawite should only form underextremely reducing conditions If the ferrous ionconcentration in the electrolyte is low macki-nawite alters to greigite (Rickard 1969 King etal 1976) The alteration is not observed in non-biological systems containing tetragonal FeS An-other example is ferrimagnetic greigite whichsome bacteria form from nonmagnetic macki-nawite (McNeil and Little 1990) Interpretationof these minerals as possible biosignatures re-quires understanding of ldquobulkrdquo and microenvi-ronmental redox conditions
Minerals with major element compositionsdiagnostic of a biogenic origin
Certain microorganisms such as those found inlichens make oxalate minerals (Freytet and Ver-recchia 1995 Allen et al 1996 Edwards et al 1997Russell et al 1998) For example calcium oxalateand other divalent metal oxalates are found in soils(Graustein et al 1977) Since oxalate is not com-monly produced abiotically its existence may in-dicate biological activity Dolomite CaMg(CO3)2with structurally ordered Mg and Ca can form viabiological pathways yet it is difficult to precipitateinorganically in most modern environments (Vas-concelos et al 1995 Warthmann et al 2000) How-ever because dolomite may form during diagen-esis its existence cannot be construed as evidenceof biological activity unless alternative origins canbe ruled out
Organic functional groups (eg carboxylgroups) strongly select for some elements overothers (eg U Mn Zn etc) Consequentlynucleation of phases on cell surfaces or at othercell-related sites could lead to distinctive mineralchemistries
Minerals whose trace element compositionindicates a biogenic origin
Some minerals contain unusual concentrationsof trace elements This is probably largely due tothe kinetics of biologically induced precipitationExamples include some high-Sr calcites and somephosphates high in specific trace elements Mor-timer et al (1997) reported that the Mg and Ca con-centrations in siderite were inversely proportionalto the rate of activity of microorganisms associ-ated with their formation Derry et al (2001) re-ported that phytoliths had low GeSi ratios com-pared with the source fluids This was attributedto selective uptake of uncharged Si rather thancharged Ge species and because Ge in soil solu-tions was complexed by organic matter
Minerals whose isotopic composition indicates abiogenic origin
Distinctive isotope fractionation patterns canindicate the role of organisms in mineral precip-itation For example the sulfur isotopic compo-sition of sulfides reflects fractionation due to se-lective uptake of isotopically light sulfur into thecell for use as an electron acceptor in respiration(eg of organics) Methanogens create methanethat is highly enriched in 12C and leave behindisotopically heavy carbon In contrast methan-otrophs can convert light carbon in methane toisotopically light carbonates These examples il-lustrate that the interpretation of isotopic biosig-natures requires some understanding of the geo-chemical system in which they formed
The existence of anomalously high or low iso-tope signatures or small-scale isotopic hetero-geneities may also be evidence of biological ac-tivity and serve as mineralogical biosignaturesLarge numbers of stable isotopes are fractionatedin biominerals In addition to S O C and N Feis garnering specific attention Experimentalwork has demonstrated a d56Fe fractionation of213permil between the Fe31 substrate and biologi-cally reduced Fe21 (Beard et al 1999) Oxygenisotopes in phosphate have recently been pro-posed as an inorganic indicator of enzymatic ac-tivity and P metabolism (Blake et al 2001)
Minerals whose surface composition suggests thepresence of microorganisms
Organisms can alter the trace or major elementchemistry of mineral surface layers or impact the
MINERALOGICAL BIOSIGNATURES 449
abundance and distribution of surface alterationproducts For example sulfur-oxidizing bacteriarapidly remove elemental sulfur from the sur-faces of dissolving metal sulfide minerals(McGuire et al 2001) In the absence of sulfur-ox-idizing microbes elemental sulfur would accu-mulate owing to its slow dissolution kinetics inmost aqueous solutions
The stoichiometry of dissolution of silicateminerals is variable depending on the pH andwhether organic ligands are present In acidic so-lutions regardless of whether organic ligands arepresent the net release of AlSi from the feldsparframework can be either approximately stoichio-metric with respect to the dissolving phase or Alcan be preferentially released to solution leavingSi-enriched surface layers At near neutral pH inthe absence of organic compounds Si is prefer-entially released to solution compared with Alleaving Al-rich residual layers However if or-ganic ligands are present the AlSi release is stoi-chiometric (Welch and Ullman 1993 Stillings etal 1996) It is not certain that such experimentalresults translate to the identification of usefulbiosignatures First observed phenomena cannotbe interpreted uniquely without additional con-straints and second clay mineral assemblagesare the likely surface products of silicate dissolu-tion under most conditions given sufficient time
Minerals whose crystal morphology indicates abiogenic origin
Most magnetotactic organisms arrange theirmagnetic crystals into chains Some species pro-duce biogenic magnetite with unique character-istics elongated nonequilibrium prismatic mor-phology chemically pure few defects includingtwins restricted to single domainsuperpara-magnetic size range restricted widthlength ra-tio and restricted anisotrophic size distribution(nonndashlog-normal distribution or truncated) Ithas been argued that on Earth magnetites withthese characteristics form only as the result of mi-crobial activity (Thomas-Keprta et al 2000 2001)Collapsed chains of magnetite crystals have beenused as biosignatures for magnetotactic bacteriain terrestrial soil samples (Fassbinder et al 1990Kirshvink and Vali 1999)
More generally crystal growth in the presenceof organic ligands can modify the crystal habit(crystal form) of any phase through alteration ofthe relative stability of crystallographically dis-
tinct surfaces (eg Mann et al 1993 Albeck et al1996)
Minerals whose surface morphology indicatesbiological impact
Organisms can affect the way in which the sur-face topography of a mineral evolves by induc-ing localized pitting Specific surface step mor-phologies and larger-scale surface topographieshave been shown to arise when minerals dissolvein the presence of organic compounds [egamino acids (Teng and Dove 1997 see Fig 1Welch et al 2002 see Fig 2)] ldquoCell-sized and cell-shapedrdquo etch pits observed on the surfaces ofmany minerals have been attributed to enhanceddissolution under attached cells [eg silicates(Fisk et al 1998 also see Thorseth et al 1995)] Insome cases it has been confirmed experimentallythat the cells are associated with pit formationand thus the pits are of microbial origin (Fig 3)However a variety of nonbiological processescan lead to formation of micron-scale pits and itmay be very difficult to develop criteria to dis-tinguish these from pits formed by microorgan-isms (eg Edwards et al 2001)
Mineral accumulations with unusual particle size
Typically when microbial redox transforma-tions induce mineral precipitation external to thecell the particle size of the products is excep-tionally small often 2ndash5 nm (Fig 4) For exam-ple enzymatic catalysis of iron oxidation by neu-trophilic iron-oxidizing bacteria leads to theformation of few-nanometer-diameter iron oxy-hydroxide particles (eg Banfield et al 2000)
BANFIELD ET AL450
FIG 1 Atomic force microscope image of calcite re-acted inorganically (left) and in the presence of anamino acid (Asp as inset right) The field of view of eachimage is 10 mm Note the very different surface step mor-phologies (photomicrograph courtesy of Patricia Dovefrom Dove and Grantham manuscript in preparation)
Small particle size does not arise uniquely owingto biological activity However nanoparticle for-mation is favored when enzymatic activity causesrapid supersaturation (leading to many simulta-neous nucleation events) under conditions wherecrystal growth would be inhibited or occur at avery slow rate In addition large crystal size orunusual crystal length in one or two dimensionsmay result from polymer-based nucleation(Calvert and Rieke 1996 MacLachlan et al 2000)
Minerals whose spatial arrangement suggests thepresence of cells
Mineral aggregates can assume unusual diag-nostically biological shapes For example ironand manganese oxides precipitate onto polymerstrands or sheaths of bacteria and preserve theshapes of these polymers Classic examples in-clude the mineralized twisted ribbons generatedby Gallionella spp and the drinking straw-likestructures formed by Leptothrix spp (Fig 5) Theproducts of these organisms have been used toinfer a biogenic origin for Fe-rich deposits in thegeological record (eg Alt 1988) Examples in-clude Cambrian sea-floor silicandashiron oxide de-posits described by Duhig et al (1992) and a Juras-sic hydrothermal vent community described byLittle et al (1999) (also see review by Hofmannand Farmer 2000)
Minerals often precipitate on or adhere to cellsurfaces For example crystallization of sulfatesand carbonates onto cell surface layers (eg pe-riodic S-layers) may lead to products with dis-tinctive size microstructure and aggregate char-acteristics (Schultze-Lam et al 1996 Fortin et al1997) In some cases mineralization may preservethe morphology of the microorganisms even af-ter the cell materials have degraded (eg mor-phological microfossils) However identificationof the former presence of life based on cell-likemorphologies is fraught with problems Manymineralization processes can generate small cell-shaped and approximately cell-sized objectsFurthermore some minerals commonly encoun-tered in soil and aqueous environments grow asmicron- and submicron-sized spherical or tubu-lar particles In some cases such morphologicaldata has been used to argue for the existence oflife forms smaller than the most commonly ac-cepted ldquosmallestrdquo bacteria [eg ldquonannobacteriardquo(Folk and Lynch 1997 Folk 1999)] generatingconsiderable controversy (also see Kirkland et al1999) Figures 6ndash8 are scanning electron micro-scope (SEM) images of micron- and submicron-scale objects resembling rounded rod- and spir-rilum-shaped cells respectively The objectsdisplay few characteristics typical of minerals(eg crystal faces) and they are fairly uniform insize These characteristics could lead the observerto conclude that the objects are cells or mineral-ized cells
MINERALOGICAL BIOSIGNATURES 451
FIG 2 Examples of the response of the apatite surfaceto experimental dissolution in (a) inorganic acid and (b)the presence of actively metabolizing cells Note the ar-row in (b) indicates the location of a cell Scale bars 5 2mm In this case the surface texture may provide evidencefor the presence of organic acids at the surface during dis-solution (Welch et al 2002)
FIG 3 SEM image of a cell-shaped pit formed under-neath a surface-attached microorganism that was dehy-drated under vacuum During the experiment the roundor disc-shaped cells of the iron-oxidizer Ferroplasma aci-darmanus colonized highly polished arsenopyrite (FeAsS)surfaces Pitting of the surface surrounding the cell is dueto the release of ferric iron which oxidizes surface sulfidegroups (see Edwards et al 2001)
a
b
The SEM is a powerful tool for detection andcharacterization of cells (eg Firtel and Bev-eridge 1995) However to verify that an objectthat looks like a cell is (or was) a cell it is neces-sary first to establish what cells should look likevia the method of observation Hydrated cellsand nonconducting material can be viewed withan environmental SEM (Collins et al 1993) withlimited resolution (10 nm) Dehydrated cells canbe examined with scanning electron microscopyat low accelerating voltages (a few kilovolts) butare difficult to visualize (Fig 9) in the absence ofmineralization Better resolution requires fixa-tion either freezing (as for Fig 10) or chemicaltreatment followed by dehydration and criticalpoint drying Cells can be ion-sputter-coated withgold palladium iridium chromium or otherheavy metals and examined at higher voltages(15ndash20 kV) Fixation dehydration and sputter-coating increase the possibly of better imagingbut also increase the possibility for introducingartifacts that may complicate interpretation ofshapes (Little et al 1991) In general additional
evidence such as a DNA or RNA stain (usuallyobserved via epifluorescence optical microscopy)is highly desirable If possible additional charac-terization methods should be employed
Transmission electron microscope (TEM)analysis of the cell-like objects shown in Figure 6revealed that they are composed of aggregates ofa fine-grain particulate lanthanide phosphatemineral TEM cross sections of the material illus-trated in Fig 7 coupled with energy-dispersivemicroanalysis demonstrated conclusively thatthe objects are spherical and tubular halloysite(see TEM image in the lower part of Fig 7) TEMcharacterization of objects in Fig 8 confirmed thatthe filaments consist of aggregated iron oxyhy-droxides An interesting phenomenon revealedby the TEM is the presence of a core of semi-aligned crystals of FeOOH within these aggre-gates (Welch et al 2001 Chan et al manuscriptin preparation) While organisms may have con-tributed to the formation of these structures theaggregates cannot be identified as cells or miner-alized cells
BANFIELD ET AL452
FIG 4 TEM image of ZnS pro-duced as a result of the activity ofsulfate-reducing bacteria in solu-tions containing 1 ppm Zn (fordetails see Labrenz et al 2000)Note that the particle size is excep-tionally small often 2 nm(smaller black arrows) The largerblack arrow indicates an aggregateof slightly misoriented particlesThe black and white arrow marksa particle exhibiting a stacking dis-order of intergrown sphalerite andwurtzite structures
Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

ASTROBIOLOGYVolume 1 Number 4 2001copy Mary Ann Liebert Inc
Research Paper
Mineralogical Biosignatures and the Search for Life on Mars
JILLIAN F BANFIELD1 JOHN W MOREAU1 CLARA S CHAN1 SUSAN A WELCH1
and BRENDA LITTLE2
ABSTRACT
If life ever existed or still exists on Mars its record is likely to be found in minerals formedby or in association with microorganisms An important concept regarding interpretation ofthe mineralogical record for evidence of life is that broadly defined life perturbs disequi-libria that arise due to kinetic barriers and can impart unexpected structure to an abiotic sys-tem Many features of minerals and mineral assemblages may serve as biosignatures even iflife does not have a familiar terrestrial chemical basis Biological impacts on minerals andmineral assemblages may be direct or indirect Crystalline or amorphous biominerals an im-portant category of mineralogical biosignatures precipitate under direct cellular control aspart of the life cycle of the organism (shells tests phytoliths) or indirectly when cell surfacelayers provide sites for heterogeneous nucleation Biominerals also form indirectly as by-products of metabolism due to changing mineral solubility Mineralogical biosignatures in-clude distinctive mineral surface structures or chemistry that arise when dissolution andorcrystal growth kinetics are influenced by metabolic by-products Mineral assemblages them-selves may be diagnostic of the prior activity of organisms where barriers to precipitation ordissolution of specific phases have been overcome Critical to resolving the question ofwhether life exists or existed on Mars is knowing how to distinguish biologically inducedstructure and organization patterns from inorganic phenomena and inorganic self-organiza-tion This task assumes special significance when it is acknowledged that the majority of andperhaps the only material to be returned from Mars will be mineralogical Key Words MarsmdashMicroorganismsmdashBiosignaturesmdashMinerals Astrobiology 1 447ndash465
447
INTRODUCTION
THE FEW UNAMBIGUOUS MINERALOGICAL biosig-natures available today are mostly restricted
to minerals produced under direct cellular con-
trol (eg coccoliths and diatoms) or within com-plex mineralndashpolymer intergrowths (eg shells)Although highly diagnostic of life in terrestrialsamples their relevance to detection of life onMars may be minimal In the absence of clearly
1Department of Geology and Geophysics University of Wisconsin Madison WI2Naval Research Laboratory Stennis Space Center MSPresent address Departments of Earth and Planetary Science and of Environmental Science Policy and Man-
agement University of California at Berkeley Berkeley CA
identifiable remains of organisms a variety ofmineralogical biosignatures may serve to estab-lish that life existed in an environment These in-clude minerals formed as a direct or indirect con-sequence of microbial metabolism mineralassemblages templated by organic polymers andmodifications to mineral surfaces or mineral dis-tribution patterns
A subset of terrestrial microorganisms derivesmetabolic energy by catalysis of inorganic chem-ical reactions Microbes use inorganic and organicsubstrates to generate energy by coupling theproduction of ATP to enzymatic transport of elec-trons Numerous electron donor and acceptorpairs are available in the form of organic mole-cules minerals dissolved ions and metals andgases especially in environments characterizedby redox transitions (eg in sediments or at theinterface between basaltic rocks and oxidizingsurface-derived fluids) Microbial utilization ofinorganic substrates in energy generation cancause minerals to dissolve or to precipitate Thecharacteristics of biologically induced mineralproducts may bear testament to their biologicalorigin Minerals formed as metabolic by-productsare typically nanocrystalline owing to their rapidprecipitation from solutions that locally arehighly supersaturated At this time insufficientinformation is available to determine whether thecharacteristics of these minerals can be used asrigorous biosignatures
In addition to modifying the redox state ofaqueous ions microbial metabolism can impactthe composition of surrounding solutionsthrough a change in pH for example via the pro-duction of organic or inorganic complexingagents This phenomenon may induce precipita-tion of minerals with crystal structures crystalsizes defect structures and morphologies thatwould not be produced in the absence of biolog-ical activity Similarly complexation of metals byorganic ligands may promote dissolution of oth-erwise extremely insoluble phases Dissolutionand precipitation may be accompanied by bio-logically induced fractionation of trace and ma-jor elements andor isotopes
Cell surfaces and cell-derived polymers canpromote heterogeneous nucleation of mineralswith unusual characteristics In some cases theseprecipitates may preserve the morphologicalcharacteristics of the cell (microfossils) Althoughsuch morphological microfossils are difficult todistinguish from mineral aggregates produced by
inorganic processes trace element isotopic andmicrostructural features may provide clues thatin combination with morphological characteris-tics could indicate a biological origin
In general life on Earth has evolved to use com-pounds readily available from the environmentespecially from solutions However as the geo-chemical characteristics of an environment changesome metabolically required elements may be-come scarce and organisms may be driven to in-crease their proximity to mineral surfaces In ad-dition organisms attach to solid substrates forprotection against environmental extremes (egelectromagnetic radiation mechanical abrasiontemperature stress hazardous chemicals tem-perature shock desiccation etc) Close mi-crobendashmineral association can promote mineraldissolution and the enhanced release of biologi-cally useful constituents to solution Distinctivesurface compositional characteristics and etchingpatterns may result Although detailed surfacefeatures may preserve evidence of life consider-able research is needed to develop reliable crite-ria for recognizing such features in the rockrecord
EXAMPLES OF TERRESTRIAL MINERALBIOSIGNATURES WITH POSSIBLE
RELEVANCE TO MARS
On Earth a variety of mineral characteristicsrecord evidence of biological activity
Minerals whose phase indicates a biogenic origin
The formation of aragonite the high-pressureCaCO3 polymorph rather than calcite by algae(de Vrind de Jong and de Vrind 1997) and inshells (Falini et al 1996) is an example of biolog-ical activity leading to precipitation of a structurenot predicted to occur given ambient physicaland chemical conditions However the crystal-lization of aragonite at low temperature can oc-cur under some specific conditions via abioticpathways For example Renault and Jones (1997)observed a cyclic precipitation of calcite and ara-gonite in a hot spring travertine which they at-tributed to rapid degassing of CO2
Algae octocorallia and echinoderm tests areoften composed of high-Mg calcite and the shellsof molluscs are typically composed of lower-Mgcalcites (eg Dauphin 1999) In the case of echi-
BANFIELD ET AL448
noderms it has been suggested that an organicmatrix with calcium-binding ability is framed ina periodic three-dimensional structure made ofconcentric layers bridged by radial threads (Am-eye et al 1998) Yet both high- and low-Mg cal-cites are found in beach cements and are some-times inferred to form inorganically (Kneale andViles 2000)
Unexpected minerals also can form if biologi-cal activity leads to the creation of redox condi-tions not anticipated based on bulk solution pa-rameters Mackinawite (a tetragonal iron sulfide)is considered to be a biosignature for microbio-logically influenced corrosion of iron by sulfate-reducing bacteria Experimental databases indi-cate mackinawite should only form underextremely reducing conditions If the ferrous ionconcentration in the electrolyte is low macki-nawite alters to greigite (Rickard 1969 King etal 1976) The alteration is not observed in non-biological systems containing tetragonal FeS An-other example is ferrimagnetic greigite whichsome bacteria form from nonmagnetic macki-nawite (McNeil and Little 1990) Interpretationof these minerals as possible biosignatures re-quires understanding of ldquobulkrdquo and microenvi-ronmental redox conditions
Minerals with major element compositionsdiagnostic of a biogenic origin
Certain microorganisms such as those found inlichens make oxalate minerals (Freytet and Ver-recchia 1995 Allen et al 1996 Edwards et al 1997Russell et al 1998) For example calcium oxalateand other divalent metal oxalates are found in soils(Graustein et al 1977) Since oxalate is not com-monly produced abiotically its existence may in-dicate biological activity Dolomite CaMg(CO3)2with structurally ordered Mg and Ca can form viabiological pathways yet it is difficult to precipitateinorganically in most modern environments (Vas-concelos et al 1995 Warthmann et al 2000) How-ever because dolomite may form during diagen-esis its existence cannot be construed as evidenceof biological activity unless alternative origins canbe ruled out
Organic functional groups (eg carboxylgroups) strongly select for some elements overothers (eg U Mn Zn etc) Consequentlynucleation of phases on cell surfaces or at othercell-related sites could lead to distinctive mineralchemistries
Minerals whose trace element compositionindicates a biogenic origin
Some minerals contain unusual concentrationsof trace elements This is probably largely due tothe kinetics of biologically induced precipitationExamples include some high-Sr calcites and somephosphates high in specific trace elements Mor-timer et al (1997) reported that the Mg and Ca con-centrations in siderite were inversely proportionalto the rate of activity of microorganisms associ-ated with their formation Derry et al (2001) re-ported that phytoliths had low GeSi ratios com-pared with the source fluids This was attributedto selective uptake of uncharged Si rather thancharged Ge species and because Ge in soil solu-tions was complexed by organic matter
Minerals whose isotopic composition indicates abiogenic origin
Distinctive isotope fractionation patterns canindicate the role of organisms in mineral precip-itation For example the sulfur isotopic compo-sition of sulfides reflects fractionation due to se-lective uptake of isotopically light sulfur into thecell for use as an electron acceptor in respiration(eg of organics) Methanogens create methanethat is highly enriched in 12C and leave behindisotopically heavy carbon In contrast methan-otrophs can convert light carbon in methane toisotopically light carbonates These examples il-lustrate that the interpretation of isotopic biosig-natures requires some understanding of the geo-chemical system in which they formed
The existence of anomalously high or low iso-tope signatures or small-scale isotopic hetero-geneities may also be evidence of biological ac-tivity and serve as mineralogical biosignaturesLarge numbers of stable isotopes are fractionatedin biominerals In addition to S O C and N Feis garnering specific attention Experimentalwork has demonstrated a d56Fe fractionation of213permil between the Fe31 substrate and biologi-cally reduced Fe21 (Beard et al 1999) Oxygenisotopes in phosphate have recently been pro-posed as an inorganic indicator of enzymatic ac-tivity and P metabolism (Blake et al 2001)
Minerals whose surface composition suggests thepresence of microorganisms
Organisms can alter the trace or major elementchemistry of mineral surface layers or impact the
MINERALOGICAL BIOSIGNATURES 449
abundance and distribution of surface alterationproducts For example sulfur-oxidizing bacteriarapidly remove elemental sulfur from the sur-faces of dissolving metal sulfide minerals(McGuire et al 2001) In the absence of sulfur-ox-idizing microbes elemental sulfur would accu-mulate owing to its slow dissolution kinetics inmost aqueous solutions
The stoichiometry of dissolution of silicateminerals is variable depending on the pH andwhether organic ligands are present In acidic so-lutions regardless of whether organic ligands arepresent the net release of AlSi from the feldsparframework can be either approximately stoichio-metric with respect to the dissolving phase or Alcan be preferentially released to solution leavingSi-enriched surface layers At near neutral pH inthe absence of organic compounds Si is prefer-entially released to solution compared with Alleaving Al-rich residual layers However if or-ganic ligands are present the AlSi release is stoi-chiometric (Welch and Ullman 1993 Stillings etal 1996) It is not certain that such experimentalresults translate to the identification of usefulbiosignatures First observed phenomena cannotbe interpreted uniquely without additional con-straints and second clay mineral assemblagesare the likely surface products of silicate dissolu-tion under most conditions given sufficient time
Minerals whose crystal morphology indicates abiogenic origin
Most magnetotactic organisms arrange theirmagnetic crystals into chains Some species pro-duce biogenic magnetite with unique character-istics elongated nonequilibrium prismatic mor-phology chemically pure few defects includingtwins restricted to single domainsuperpara-magnetic size range restricted widthlength ra-tio and restricted anisotrophic size distribution(nonndashlog-normal distribution or truncated) Ithas been argued that on Earth magnetites withthese characteristics form only as the result of mi-crobial activity (Thomas-Keprta et al 2000 2001)Collapsed chains of magnetite crystals have beenused as biosignatures for magnetotactic bacteriain terrestrial soil samples (Fassbinder et al 1990Kirshvink and Vali 1999)
More generally crystal growth in the presenceof organic ligands can modify the crystal habit(crystal form) of any phase through alteration ofthe relative stability of crystallographically dis-
tinct surfaces (eg Mann et al 1993 Albeck et al1996)
Minerals whose surface morphology indicatesbiological impact
Organisms can affect the way in which the sur-face topography of a mineral evolves by induc-ing localized pitting Specific surface step mor-phologies and larger-scale surface topographieshave been shown to arise when minerals dissolvein the presence of organic compounds [egamino acids (Teng and Dove 1997 see Fig 1Welch et al 2002 see Fig 2)] ldquoCell-sized and cell-shapedrdquo etch pits observed on the surfaces ofmany minerals have been attributed to enhanceddissolution under attached cells [eg silicates(Fisk et al 1998 also see Thorseth et al 1995)] Insome cases it has been confirmed experimentallythat the cells are associated with pit formationand thus the pits are of microbial origin (Fig 3)However a variety of nonbiological processescan lead to formation of micron-scale pits and itmay be very difficult to develop criteria to dis-tinguish these from pits formed by microorgan-isms (eg Edwards et al 2001)
Mineral accumulations with unusual particle size
Typically when microbial redox transforma-tions induce mineral precipitation external to thecell the particle size of the products is excep-tionally small often 2ndash5 nm (Fig 4) For exam-ple enzymatic catalysis of iron oxidation by neu-trophilic iron-oxidizing bacteria leads to theformation of few-nanometer-diameter iron oxy-hydroxide particles (eg Banfield et al 2000)
BANFIELD ET AL450
FIG 1 Atomic force microscope image of calcite re-acted inorganically (left) and in the presence of anamino acid (Asp as inset right) The field of view of eachimage is 10 mm Note the very different surface step mor-phologies (photomicrograph courtesy of Patricia Dovefrom Dove and Grantham manuscript in preparation)
Small particle size does not arise uniquely owingto biological activity However nanoparticle for-mation is favored when enzymatic activity causesrapid supersaturation (leading to many simulta-neous nucleation events) under conditions wherecrystal growth would be inhibited or occur at avery slow rate In addition large crystal size orunusual crystal length in one or two dimensionsmay result from polymer-based nucleation(Calvert and Rieke 1996 MacLachlan et al 2000)
Minerals whose spatial arrangement suggests thepresence of cells
Mineral aggregates can assume unusual diag-nostically biological shapes For example ironand manganese oxides precipitate onto polymerstrands or sheaths of bacteria and preserve theshapes of these polymers Classic examples in-clude the mineralized twisted ribbons generatedby Gallionella spp and the drinking straw-likestructures formed by Leptothrix spp (Fig 5) Theproducts of these organisms have been used toinfer a biogenic origin for Fe-rich deposits in thegeological record (eg Alt 1988) Examples in-clude Cambrian sea-floor silicandashiron oxide de-posits described by Duhig et al (1992) and a Juras-sic hydrothermal vent community described byLittle et al (1999) (also see review by Hofmannand Farmer 2000)
Minerals often precipitate on or adhere to cellsurfaces For example crystallization of sulfatesand carbonates onto cell surface layers (eg pe-riodic S-layers) may lead to products with dis-tinctive size microstructure and aggregate char-acteristics (Schultze-Lam et al 1996 Fortin et al1997) In some cases mineralization may preservethe morphology of the microorganisms even af-ter the cell materials have degraded (eg mor-phological microfossils) However identificationof the former presence of life based on cell-likemorphologies is fraught with problems Manymineralization processes can generate small cell-shaped and approximately cell-sized objectsFurthermore some minerals commonly encoun-tered in soil and aqueous environments grow asmicron- and submicron-sized spherical or tubu-lar particles In some cases such morphologicaldata has been used to argue for the existence oflife forms smaller than the most commonly ac-cepted ldquosmallestrdquo bacteria [eg ldquonannobacteriardquo(Folk and Lynch 1997 Folk 1999)] generatingconsiderable controversy (also see Kirkland et al1999) Figures 6ndash8 are scanning electron micro-scope (SEM) images of micron- and submicron-scale objects resembling rounded rod- and spir-rilum-shaped cells respectively The objectsdisplay few characteristics typical of minerals(eg crystal faces) and they are fairly uniform insize These characteristics could lead the observerto conclude that the objects are cells or mineral-ized cells
MINERALOGICAL BIOSIGNATURES 451
FIG 2 Examples of the response of the apatite surfaceto experimental dissolution in (a) inorganic acid and (b)the presence of actively metabolizing cells Note the ar-row in (b) indicates the location of a cell Scale bars 5 2mm In this case the surface texture may provide evidencefor the presence of organic acids at the surface during dis-solution (Welch et al 2002)
FIG 3 SEM image of a cell-shaped pit formed under-neath a surface-attached microorganism that was dehy-drated under vacuum During the experiment the roundor disc-shaped cells of the iron-oxidizer Ferroplasma aci-darmanus colonized highly polished arsenopyrite (FeAsS)surfaces Pitting of the surface surrounding the cell is dueto the release of ferric iron which oxidizes surface sulfidegroups (see Edwards et al 2001)
a
b
The SEM is a powerful tool for detection andcharacterization of cells (eg Firtel and Bev-eridge 1995) However to verify that an objectthat looks like a cell is (or was) a cell it is neces-sary first to establish what cells should look likevia the method of observation Hydrated cellsand nonconducting material can be viewed withan environmental SEM (Collins et al 1993) withlimited resolution (10 nm) Dehydrated cells canbe examined with scanning electron microscopyat low accelerating voltages (a few kilovolts) butare difficult to visualize (Fig 9) in the absence ofmineralization Better resolution requires fixa-tion either freezing (as for Fig 10) or chemicaltreatment followed by dehydration and criticalpoint drying Cells can be ion-sputter-coated withgold palladium iridium chromium or otherheavy metals and examined at higher voltages(15ndash20 kV) Fixation dehydration and sputter-coating increase the possibly of better imagingbut also increase the possibility for introducingartifacts that may complicate interpretation ofshapes (Little et al 1991) In general additional
evidence such as a DNA or RNA stain (usuallyobserved via epifluorescence optical microscopy)is highly desirable If possible additional charac-terization methods should be employed
Transmission electron microscope (TEM)analysis of the cell-like objects shown in Figure 6revealed that they are composed of aggregates ofa fine-grain particulate lanthanide phosphatemineral TEM cross sections of the material illus-trated in Fig 7 coupled with energy-dispersivemicroanalysis demonstrated conclusively thatthe objects are spherical and tubular halloysite(see TEM image in the lower part of Fig 7) TEMcharacterization of objects in Fig 8 confirmed thatthe filaments consist of aggregated iron oxyhy-droxides An interesting phenomenon revealedby the TEM is the presence of a core of semi-aligned crystals of FeOOH within these aggre-gates (Welch et al 2001 Chan et al manuscriptin preparation) While organisms may have con-tributed to the formation of these structures theaggregates cannot be identified as cells or miner-alized cells
BANFIELD ET AL452
FIG 4 TEM image of ZnS pro-duced as a result of the activity ofsulfate-reducing bacteria in solu-tions containing 1 ppm Zn (fordetails see Labrenz et al 2000)Note that the particle size is excep-tionally small often 2 nm(smaller black arrows) The largerblack arrow indicates an aggregateof slightly misoriented particlesThe black and white arrow marksa particle exhibiting a stacking dis-order of intergrown sphalerite andwurtzite structures
Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

identifiable remains of organisms a variety ofmineralogical biosignatures may serve to estab-lish that life existed in an environment These in-clude minerals formed as a direct or indirect con-sequence of microbial metabolism mineralassemblages templated by organic polymers andmodifications to mineral surfaces or mineral dis-tribution patterns
A subset of terrestrial microorganisms derivesmetabolic energy by catalysis of inorganic chem-ical reactions Microbes use inorganic and organicsubstrates to generate energy by coupling theproduction of ATP to enzymatic transport of elec-trons Numerous electron donor and acceptorpairs are available in the form of organic mole-cules minerals dissolved ions and metals andgases especially in environments characterizedby redox transitions (eg in sediments or at theinterface between basaltic rocks and oxidizingsurface-derived fluids) Microbial utilization ofinorganic substrates in energy generation cancause minerals to dissolve or to precipitate Thecharacteristics of biologically induced mineralproducts may bear testament to their biologicalorigin Minerals formed as metabolic by-productsare typically nanocrystalline owing to their rapidprecipitation from solutions that locally arehighly supersaturated At this time insufficientinformation is available to determine whether thecharacteristics of these minerals can be used asrigorous biosignatures
In addition to modifying the redox state ofaqueous ions microbial metabolism can impactthe composition of surrounding solutionsthrough a change in pH for example via the pro-duction of organic or inorganic complexingagents This phenomenon may induce precipita-tion of minerals with crystal structures crystalsizes defect structures and morphologies thatwould not be produced in the absence of biolog-ical activity Similarly complexation of metals byorganic ligands may promote dissolution of oth-erwise extremely insoluble phases Dissolutionand precipitation may be accompanied by bio-logically induced fractionation of trace and ma-jor elements andor isotopes
Cell surfaces and cell-derived polymers canpromote heterogeneous nucleation of mineralswith unusual characteristics In some cases theseprecipitates may preserve the morphologicalcharacteristics of the cell (microfossils) Althoughsuch morphological microfossils are difficult todistinguish from mineral aggregates produced by
inorganic processes trace element isotopic andmicrostructural features may provide clues thatin combination with morphological characteris-tics could indicate a biological origin
In general life on Earth has evolved to use com-pounds readily available from the environmentespecially from solutions However as the geo-chemical characteristics of an environment changesome metabolically required elements may be-come scarce and organisms may be driven to in-crease their proximity to mineral surfaces In ad-dition organisms attach to solid substrates forprotection against environmental extremes (egelectromagnetic radiation mechanical abrasiontemperature stress hazardous chemicals tem-perature shock desiccation etc) Close mi-crobendashmineral association can promote mineraldissolution and the enhanced release of biologi-cally useful constituents to solution Distinctivesurface compositional characteristics and etchingpatterns may result Although detailed surfacefeatures may preserve evidence of life consider-able research is needed to develop reliable crite-ria for recognizing such features in the rockrecord
EXAMPLES OF TERRESTRIAL MINERALBIOSIGNATURES WITH POSSIBLE
RELEVANCE TO MARS
On Earth a variety of mineral characteristicsrecord evidence of biological activity
Minerals whose phase indicates a biogenic origin
The formation of aragonite the high-pressureCaCO3 polymorph rather than calcite by algae(de Vrind de Jong and de Vrind 1997) and inshells (Falini et al 1996) is an example of biolog-ical activity leading to precipitation of a structurenot predicted to occur given ambient physicaland chemical conditions However the crystal-lization of aragonite at low temperature can oc-cur under some specific conditions via abioticpathways For example Renault and Jones (1997)observed a cyclic precipitation of calcite and ara-gonite in a hot spring travertine which they at-tributed to rapid degassing of CO2
Algae octocorallia and echinoderm tests areoften composed of high-Mg calcite and the shellsof molluscs are typically composed of lower-Mgcalcites (eg Dauphin 1999) In the case of echi-
BANFIELD ET AL448
noderms it has been suggested that an organicmatrix with calcium-binding ability is framed ina periodic three-dimensional structure made ofconcentric layers bridged by radial threads (Am-eye et al 1998) Yet both high- and low-Mg cal-cites are found in beach cements and are some-times inferred to form inorganically (Kneale andViles 2000)
Unexpected minerals also can form if biologi-cal activity leads to the creation of redox condi-tions not anticipated based on bulk solution pa-rameters Mackinawite (a tetragonal iron sulfide)is considered to be a biosignature for microbio-logically influenced corrosion of iron by sulfate-reducing bacteria Experimental databases indi-cate mackinawite should only form underextremely reducing conditions If the ferrous ionconcentration in the electrolyte is low macki-nawite alters to greigite (Rickard 1969 King etal 1976) The alteration is not observed in non-biological systems containing tetragonal FeS An-other example is ferrimagnetic greigite whichsome bacteria form from nonmagnetic macki-nawite (McNeil and Little 1990) Interpretationof these minerals as possible biosignatures re-quires understanding of ldquobulkrdquo and microenvi-ronmental redox conditions
Minerals with major element compositionsdiagnostic of a biogenic origin
Certain microorganisms such as those found inlichens make oxalate minerals (Freytet and Ver-recchia 1995 Allen et al 1996 Edwards et al 1997Russell et al 1998) For example calcium oxalateand other divalent metal oxalates are found in soils(Graustein et al 1977) Since oxalate is not com-monly produced abiotically its existence may in-dicate biological activity Dolomite CaMg(CO3)2with structurally ordered Mg and Ca can form viabiological pathways yet it is difficult to precipitateinorganically in most modern environments (Vas-concelos et al 1995 Warthmann et al 2000) How-ever because dolomite may form during diagen-esis its existence cannot be construed as evidenceof biological activity unless alternative origins canbe ruled out
Organic functional groups (eg carboxylgroups) strongly select for some elements overothers (eg U Mn Zn etc) Consequentlynucleation of phases on cell surfaces or at othercell-related sites could lead to distinctive mineralchemistries
Minerals whose trace element compositionindicates a biogenic origin
Some minerals contain unusual concentrationsof trace elements This is probably largely due tothe kinetics of biologically induced precipitationExamples include some high-Sr calcites and somephosphates high in specific trace elements Mor-timer et al (1997) reported that the Mg and Ca con-centrations in siderite were inversely proportionalto the rate of activity of microorganisms associ-ated with their formation Derry et al (2001) re-ported that phytoliths had low GeSi ratios com-pared with the source fluids This was attributedto selective uptake of uncharged Si rather thancharged Ge species and because Ge in soil solu-tions was complexed by organic matter
Minerals whose isotopic composition indicates abiogenic origin
Distinctive isotope fractionation patterns canindicate the role of organisms in mineral precip-itation For example the sulfur isotopic compo-sition of sulfides reflects fractionation due to se-lective uptake of isotopically light sulfur into thecell for use as an electron acceptor in respiration(eg of organics) Methanogens create methanethat is highly enriched in 12C and leave behindisotopically heavy carbon In contrast methan-otrophs can convert light carbon in methane toisotopically light carbonates These examples il-lustrate that the interpretation of isotopic biosig-natures requires some understanding of the geo-chemical system in which they formed
The existence of anomalously high or low iso-tope signatures or small-scale isotopic hetero-geneities may also be evidence of biological ac-tivity and serve as mineralogical biosignaturesLarge numbers of stable isotopes are fractionatedin biominerals In addition to S O C and N Feis garnering specific attention Experimentalwork has demonstrated a d56Fe fractionation of213permil between the Fe31 substrate and biologi-cally reduced Fe21 (Beard et al 1999) Oxygenisotopes in phosphate have recently been pro-posed as an inorganic indicator of enzymatic ac-tivity and P metabolism (Blake et al 2001)
Minerals whose surface composition suggests thepresence of microorganisms
Organisms can alter the trace or major elementchemistry of mineral surface layers or impact the
MINERALOGICAL BIOSIGNATURES 449
abundance and distribution of surface alterationproducts For example sulfur-oxidizing bacteriarapidly remove elemental sulfur from the sur-faces of dissolving metal sulfide minerals(McGuire et al 2001) In the absence of sulfur-ox-idizing microbes elemental sulfur would accu-mulate owing to its slow dissolution kinetics inmost aqueous solutions
The stoichiometry of dissolution of silicateminerals is variable depending on the pH andwhether organic ligands are present In acidic so-lutions regardless of whether organic ligands arepresent the net release of AlSi from the feldsparframework can be either approximately stoichio-metric with respect to the dissolving phase or Alcan be preferentially released to solution leavingSi-enriched surface layers At near neutral pH inthe absence of organic compounds Si is prefer-entially released to solution compared with Alleaving Al-rich residual layers However if or-ganic ligands are present the AlSi release is stoi-chiometric (Welch and Ullman 1993 Stillings etal 1996) It is not certain that such experimentalresults translate to the identification of usefulbiosignatures First observed phenomena cannotbe interpreted uniquely without additional con-straints and second clay mineral assemblagesare the likely surface products of silicate dissolu-tion under most conditions given sufficient time
Minerals whose crystal morphology indicates abiogenic origin
Most magnetotactic organisms arrange theirmagnetic crystals into chains Some species pro-duce biogenic magnetite with unique character-istics elongated nonequilibrium prismatic mor-phology chemically pure few defects includingtwins restricted to single domainsuperpara-magnetic size range restricted widthlength ra-tio and restricted anisotrophic size distribution(nonndashlog-normal distribution or truncated) Ithas been argued that on Earth magnetites withthese characteristics form only as the result of mi-crobial activity (Thomas-Keprta et al 2000 2001)Collapsed chains of magnetite crystals have beenused as biosignatures for magnetotactic bacteriain terrestrial soil samples (Fassbinder et al 1990Kirshvink and Vali 1999)
More generally crystal growth in the presenceof organic ligands can modify the crystal habit(crystal form) of any phase through alteration ofthe relative stability of crystallographically dis-
tinct surfaces (eg Mann et al 1993 Albeck et al1996)
Minerals whose surface morphology indicatesbiological impact
Organisms can affect the way in which the sur-face topography of a mineral evolves by induc-ing localized pitting Specific surface step mor-phologies and larger-scale surface topographieshave been shown to arise when minerals dissolvein the presence of organic compounds [egamino acids (Teng and Dove 1997 see Fig 1Welch et al 2002 see Fig 2)] ldquoCell-sized and cell-shapedrdquo etch pits observed on the surfaces ofmany minerals have been attributed to enhanceddissolution under attached cells [eg silicates(Fisk et al 1998 also see Thorseth et al 1995)] Insome cases it has been confirmed experimentallythat the cells are associated with pit formationand thus the pits are of microbial origin (Fig 3)However a variety of nonbiological processescan lead to formation of micron-scale pits and itmay be very difficult to develop criteria to dis-tinguish these from pits formed by microorgan-isms (eg Edwards et al 2001)
Mineral accumulations with unusual particle size
Typically when microbial redox transforma-tions induce mineral precipitation external to thecell the particle size of the products is excep-tionally small often 2ndash5 nm (Fig 4) For exam-ple enzymatic catalysis of iron oxidation by neu-trophilic iron-oxidizing bacteria leads to theformation of few-nanometer-diameter iron oxy-hydroxide particles (eg Banfield et al 2000)
BANFIELD ET AL450
FIG 1 Atomic force microscope image of calcite re-acted inorganically (left) and in the presence of anamino acid (Asp as inset right) The field of view of eachimage is 10 mm Note the very different surface step mor-phologies (photomicrograph courtesy of Patricia Dovefrom Dove and Grantham manuscript in preparation)
Small particle size does not arise uniquely owingto biological activity However nanoparticle for-mation is favored when enzymatic activity causesrapid supersaturation (leading to many simulta-neous nucleation events) under conditions wherecrystal growth would be inhibited or occur at avery slow rate In addition large crystal size orunusual crystal length in one or two dimensionsmay result from polymer-based nucleation(Calvert and Rieke 1996 MacLachlan et al 2000)
Minerals whose spatial arrangement suggests thepresence of cells
Mineral aggregates can assume unusual diag-nostically biological shapes For example ironand manganese oxides precipitate onto polymerstrands or sheaths of bacteria and preserve theshapes of these polymers Classic examples in-clude the mineralized twisted ribbons generatedby Gallionella spp and the drinking straw-likestructures formed by Leptothrix spp (Fig 5) Theproducts of these organisms have been used toinfer a biogenic origin for Fe-rich deposits in thegeological record (eg Alt 1988) Examples in-clude Cambrian sea-floor silicandashiron oxide de-posits described by Duhig et al (1992) and a Juras-sic hydrothermal vent community described byLittle et al (1999) (also see review by Hofmannand Farmer 2000)
Minerals often precipitate on or adhere to cellsurfaces For example crystallization of sulfatesand carbonates onto cell surface layers (eg pe-riodic S-layers) may lead to products with dis-tinctive size microstructure and aggregate char-acteristics (Schultze-Lam et al 1996 Fortin et al1997) In some cases mineralization may preservethe morphology of the microorganisms even af-ter the cell materials have degraded (eg mor-phological microfossils) However identificationof the former presence of life based on cell-likemorphologies is fraught with problems Manymineralization processes can generate small cell-shaped and approximately cell-sized objectsFurthermore some minerals commonly encoun-tered in soil and aqueous environments grow asmicron- and submicron-sized spherical or tubu-lar particles In some cases such morphologicaldata has been used to argue for the existence oflife forms smaller than the most commonly ac-cepted ldquosmallestrdquo bacteria [eg ldquonannobacteriardquo(Folk and Lynch 1997 Folk 1999)] generatingconsiderable controversy (also see Kirkland et al1999) Figures 6ndash8 are scanning electron micro-scope (SEM) images of micron- and submicron-scale objects resembling rounded rod- and spir-rilum-shaped cells respectively The objectsdisplay few characteristics typical of minerals(eg crystal faces) and they are fairly uniform insize These characteristics could lead the observerto conclude that the objects are cells or mineral-ized cells
MINERALOGICAL BIOSIGNATURES 451
FIG 2 Examples of the response of the apatite surfaceto experimental dissolution in (a) inorganic acid and (b)the presence of actively metabolizing cells Note the ar-row in (b) indicates the location of a cell Scale bars 5 2mm In this case the surface texture may provide evidencefor the presence of organic acids at the surface during dis-solution (Welch et al 2002)
FIG 3 SEM image of a cell-shaped pit formed under-neath a surface-attached microorganism that was dehy-drated under vacuum During the experiment the roundor disc-shaped cells of the iron-oxidizer Ferroplasma aci-darmanus colonized highly polished arsenopyrite (FeAsS)surfaces Pitting of the surface surrounding the cell is dueto the release of ferric iron which oxidizes surface sulfidegroups (see Edwards et al 2001)
a
b
The SEM is a powerful tool for detection andcharacterization of cells (eg Firtel and Bev-eridge 1995) However to verify that an objectthat looks like a cell is (or was) a cell it is neces-sary first to establish what cells should look likevia the method of observation Hydrated cellsand nonconducting material can be viewed withan environmental SEM (Collins et al 1993) withlimited resolution (10 nm) Dehydrated cells canbe examined with scanning electron microscopyat low accelerating voltages (a few kilovolts) butare difficult to visualize (Fig 9) in the absence ofmineralization Better resolution requires fixa-tion either freezing (as for Fig 10) or chemicaltreatment followed by dehydration and criticalpoint drying Cells can be ion-sputter-coated withgold palladium iridium chromium or otherheavy metals and examined at higher voltages(15ndash20 kV) Fixation dehydration and sputter-coating increase the possibly of better imagingbut also increase the possibility for introducingartifacts that may complicate interpretation ofshapes (Little et al 1991) In general additional
evidence such as a DNA or RNA stain (usuallyobserved via epifluorescence optical microscopy)is highly desirable If possible additional charac-terization methods should be employed
Transmission electron microscope (TEM)analysis of the cell-like objects shown in Figure 6revealed that they are composed of aggregates ofa fine-grain particulate lanthanide phosphatemineral TEM cross sections of the material illus-trated in Fig 7 coupled with energy-dispersivemicroanalysis demonstrated conclusively thatthe objects are spherical and tubular halloysite(see TEM image in the lower part of Fig 7) TEMcharacterization of objects in Fig 8 confirmed thatthe filaments consist of aggregated iron oxyhy-droxides An interesting phenomenon revealedby the TEM is the presence of a core of semi-aligned crystals of FeOOH within these aggre-gates (Welch et al 2001 Chan et al manuscriptin preparation) While organisms may have con-tributed to the formation of these structures theaggregates cannot be identified as cells or miner-alized cells
BANFIELD ET AL452
FIG 4 TEM image of ZnS pro-duced as a result of the activity ofsulfate-reducing bacteria in solu-tions containing 1 ppm Zn (fordetails see Labrenz et al 2000)Note that the particle size is excep-tionally small often 2 nm(smaller black arrows) The largerblack arrow indicates an aggregateof slightly misoriented particlesThe black and white arrow marksa particle exhibiting a stacking dis-order of intergrown sphalerite andwurtzite structures
Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

noderms it has been suggested that an organicmatrix with calcium-binding ability is framed ina periodic three-dimensional structure made ofconcentric layers bridged by radial threads (Am-eye et al 1998) Yet both high- and low-Mg cal-cites are found in beach cements and are some-times inferred to form inorganically (Kneale andViles 2000)
Unexpected minerals also can form if biologi-cal activity leads to the creation of redox condi-tions not anticipated based on bulk solution pa-rameters Mackinawite (a tetragonal iron sulfide)is considered to be a biosignature for microbio-logically influenced corrosion of iron by sulfate-reducing bacteria Experimental databases indi-cate mackinawite should only form underextremely reducing conditions If the ferrous ionconcentration in the electrolyte is low macki-nawite alters to greigite (Rickard 1969 King etal 1976) The alteration is not observed in non-biological systems containing tetragonal FeS An-other example is ferrimagnetic greigite whichsome bacteria form from nonmagnetic macki-nawite (McNeil and Little 1990) Interpretationof these minerals as possible biosignatures re-quires understanding of ldquobulkrdquo and microenvi-ronmental redox conditions
Minerals with major element compositionsdiagnostic of a biogenic origin
Certain microorganisms such as those found inlichens make oxalate minerals (Freytet and Ver-recchia 1995 Allen et al 1996 Edwards et al 1997Russell et al 1998) For example calcium oxalateand other divalent metal oxalates are found in soils(Graustein et al 1977) Since oxalate is not com-monly produced abiotically its existence may in-dicate biological activity Dolomite CaMg(CO3)2with structurally ordered Mg and Ca can form viabiological pathways yet it is difficult to precipitateinorganically in most modern environments (Vas-concelos et al 1995 Warthmann et al 2000) How-ever because dolomite may form during diagen-esis its existence cannot be construed as evidenceof biological activity unless alternative origins canbe ruled out
Organic functional groups (eg carboxylgroups) strongly select for some elements overothers (eg U Mn Zn etc) Consequentlynucleation of phases on cell surfaces or at othercell-related sites could lead to distinctive mineralchemistries
Minerals whose trace element compositionindicates a biogenic origin
Some minerals contain unusual concentrationsof trace elements This is probably largely due tothe kinetics of biologically induced precipitationExamples include some high-Sr calcites and somephosphates high in specific trace elements Mor-timer et al (1997) reported that the Mg and Ca con-centrations in siderite were inversely proportionalto the rate of activity of microorganisms associ-ated with their formation Derry et al (2001) re-ported that phytoliths had low GeSi ratios com-pared with the source fluids This was attributedto selective uptake of uncharged Si rather thancharged Ge species and because Ge in soil solu-tions was complexed by organic matter
Minerals whose isotopic composition indicates abiogenic origin
Distinctive isotope fractionation patterns canindicate the role of organisms in mineral precip-itation For example the sulfur isotopic compo-sition of sulfides reflects fractionation due to se-lective uptake of isotopically light sulfur into thecell for use as an electron acceptor in respiration(eg of organics) Methanogens create methanethat is highly enriched in 12C and leave behindisotopically heavy carbon In contrast methan-otrophs can convert light carbon in methane toisotopically light carbonates These examples il-lustrate that the interpretation of isotopic biosig-natures requires some understanding of the geo-chemical system in which they formed
The existence of anomalously high or low iso-tope signatures or small-scale isotopic hetero-geneities may also be evidence of biological ac-tivity and serve as mineralogical biosignaturesLarge numbers of stable isotopes are fractionatedin biominerals In addition to S O C and N Feis garnering specific attention Experimentalwork has demonstrated a d56Fe fractionation of213permil between the Fe31 substrate and biologi-cally reduced Fe21 (Beard et al 1999) Oxygenisotopes in phosphate have recently been pro-posed as an inorganic indicator of enzymatic ac-tivity and P metabolism (Blake et al 2001)
Minerals whose surface composition suggests thepresence of microorganisms
Organisms can alter the trace or major elementchemistry of mineral surface layers or impact the
MINERALOGICAL BIOSIGNATURES 449
abundance and distribution of surface alterationproducts For example sulfur-oxidizing bacteriarapidly remove elemental sulfur from the sur-faces of dissolving metal sulfide minerals(McGuire et al 2001) In the absence of sulfur-ox-idizing microbes elemental sulfur would accu-mulate owing to its slow dissolution kinetics inmost aqueous solutions
The stoichiometry of dissolution of silicateminerals is variable depending on the pH andwhether organic ligands are present In acidic so-lutions regardless of whether organic ligands arepresent the net release of AlSi from the feldsparframework can be either approximately stoichio-metric with respect to the dissolving phase or Alcan be preferentially released to solution leavingSi-enriched surface layers At near neutral pH inthe absence of organic compounds Si is prefer-entially released to solution compared with Alleaving Al-rich residual layers However if or-ganic ligands are present the AlSi release is stoi-chiometric (Welch and Ullman 1993 Stillings etal 1996) It is not certain that such experimentalresults translate to the identification of usefulbiosignatures First observed phenomena cannotbe interpreted uniquely without additional con-straints and second clay mineral assemblagesare the likely surface products of silicate dissolu-tion under most conditions given sufficient time
Minerals whose crystal morphology indicates abiogenic origin
Most magnetotactic organisms arrange theirmagnetic crystals into chains Some species pro-duce biogenic magnetite with unique character-istics elongated nonequilibrium prismatic mor-phology chemically pure few defects includingtwins restricted to single domainsuperpara-magnetic size range restricted widthlength ra-tio and restricted anisotrophic size distribution(nonndashlog-normal distribution or truncated) Ithas been argued that on Earth magnetites withthese characteristics form only as the result of mi-crobial activity (Thomas-Keprta et al 2000 2001)Collapsed chains of magnetite crystals have beenused as biosignatures for magnetotactic bacteriain terrestrial soil samples (Fassbinder et al 1990Kirshvink and Vali 1999)
More generally crystal growth in the presenceof organic ligands can modify the crystal habit(crystal form) of any phase through alteration ofthe relative stability of crystallographically dis-
tinct surfaces (eg Mann et al 1993 Albeck et al1996)
Minerals whose surface morphology indicatesbiological impact
Organisms can affect the way in which the sur-face topography of a mineral evolves by induc-ing localized pitting Specific surface step mor-phologies and larger-scale surface topographieshave been shown to arise when minerals dissolvein the presence of organic compounds [egamino acids (Teng and Dove 1997 see Fig 1Welch et al 2002 see Fig 2)] ldquoCell-sized and cell-shapedrdquo etch pits observed on the surfaces ofmany minerals have been attributed to enhanceddissolution under attached cells [eg silicates(Fisk et al 1998 also see Thorseth et al 1995)] Insome cases it has been confirmed experimentallythat the cells are associated with pit formationand thus the pits are of microbial origin (Fig 3)However a variety of nonbiological processescan lead to formation of micron-scale pits and itmay be very difficult to develop criteria to dis-tinguish these from pits formed by microorgan-isms (eg Edwards et al 2001)
Mineral accumulations with unusual particle size
Typically when microbial redox transforma-tions induce mineral precipitation external to thecell the particle size of the products is excep-tionally small often 2ndash5 nm (Fig 4) For exam-ple enzymatic catalysis of iron oxidation by neu-trophilic iron-oxidizing bacteria leads to theformation of few-nanometer-diameter iron oxy-hydroxide particles (eg Banfield et al 2000)
BANFIELD ET AL450
FIG 1 Atomic force microscope image of calcite re-acted inorganically (left) and in the presence of anamino acid (Asp as inset right) The field of view of eachimage is 10 mm Note the very different surface step mor-phologies (photomicrograph courtesy of Patricia Dovefrom Dove and Grantham manuscript in preparation)
Small particle size does not arise uniquely owingto biological activity However nanoparticle for-mation is favored when enzymatic activity causesrapid supersaturation (leading to many simulta-neous nucleation events) under conditions wherecrystal growth would be inhibited or occur at avery slow rate In addition large crystal size orunusual crystal length in one or two dimensionsmay result from polymer-based nucleation(Calvert and Rieke 1996 MacLachlan et al 2000)
Minerals whose spatial arrangement suggests thepresence of cells
Mineral aggregates can assume unusual diag-nostically biological shapes For example ironand manganese oxides precipitate onto polymerstrands or sheaths of bacteria and preserve theshapes of these polymers Classic examples in-clude the mineralized twisted ribbons generatedby Gallionella spp and the drinking straw-likestructures formed by Leptothrix spp (Fig 5) Theproducts of these organisms have been used toinfer a biogenic origin for Fe-rich deposits in thegeological record (eg Alt 1988) Examples in-clude Cambrian sea-floor silicandashiron oxide de-posits described by Duhig et al (1992) and a Juras-sic hydrothermal vent community described byLittle et al (1999) (also see review by Hofmannand Farmer 2000)
Minerals often precipitate on or adhere to cellsurfaces For example crystallization of sulfatesand carbonates onto cell surface layers (eg pe-riodic S-layers) may lead to products with dis-tinctive size microstructure and aggregate char-acteristics (Schultze-Lam et al 1996 Fortin et al1997) In some cases mineralization may preservethe morphology of the microorganisms even af-ter the cell materials have degraded (eg mor-phological microfossils) However identificationof the former presence of life based on cell-likemorphologies is fraught with problems Manymineralization processes can generate small cell-shaped and approximately cell-sized objectsFurthermore some minerals commonly encoun-tered in soil and aqueous environments grow asmicron- and submicron-sized spherical or tubu-lar particles In some cases such morphologicaldata has been used to argue for the existence oflife forms smaller than the most commonly ac-cepted ldquosmallestrdquo bacteria [eg ldquonannobacteriardquo(Folk and Lynch 1997 Folk 1999)] generatingconsiderable controversy (also see Kirkland et al1999) Figures 6ndash8 are scanning electron micro-scope (SEM) images of micron- and submicron-scale objects resembling rounded rod- and spir-rilum-shaped cells respectively The objectsdisplay few characteristics typical of minerals(eg crystal faces) and they are fairly uniform insize These characteristics could lead the observerto conclude that the objects are cells or mineral-ized cells
MINERALOGICAL BIOSIGNATURES 451
FIG 2 Examples of the response of the apatite surfaceto experimental dissolution in (a) inorganic acid and (b)the presence of actively metabolizing cells Note the ar-row in (b) indicates the location of a cell Scale bars 5 2mm In this case the surface texture may provide evidencefor the presence of organic acids at the surface during dis-solution (Welch et al 2002)
FIG 3 SEM image of a cell-shaped pit formed under-neath a surface-attached microorganism that was dehy-drated under vacuum During the experiment the roundor disc-shaped cells of the iron-oxidizer Ferroplasma aci-darmanus colonized highly polished arsenopyrite (FeAsS)surfaces Pitting of the surface surrounding the cell is dueto the release of ferric iron which oxidizes surface sulfidegroups (see Edwards et al 2001)
a
b
The SEM is a powerful tool for detection andcharacterization of cells (eg Firtel and Bev-eridge 1995) However to verify that an objectthat looks like a cell is (or was) a cell it is neces-sary first to establish what cells should look likevia the method of observation Hydrated cellsand nonconducting material can be viewed withan environmental SEM (Collins et al 1993) withlimited resolution (10 nm) Dehydrated cells canbe examined with scanning electron microscopyat low accelerating voltages (a few kilovolts) butare difficult to visualize (Fig 9) in the absence ofmineralization Better resolution requires fixa-tion either freezing (as for Fig 10) or chemicaltreatment followed by dehydration and criticalpoint drying Cells can be ion-sputter-coated withgold palladium iridium chromium or otherheavy metals and examined at higher voltages(15ndash20 kV) Fixation dehydration and sputter-coating increase the possibly of better imagingbut also increase the possibility for introducingartifacts that may complicate interpretation ofshapes (Little et al 1991) In general additional
evidence such as a DNA or RNA stain (usuallyobserved via epifluorescence optical microscopy)is highly desirable If possible additional charac-terization methods should be employed
Transmission electron microscope (TEM)analysis of the cell-like objects shown in Figure 6revealed that they are composed of aggregates ofa fine-grain particulate lanthanide phosphatemineral TEM cross sections of the material illus-trated in Fig 7 coupled with energy-dispersivemicroanalysis demonstrated conclusively thatthe objects are spherical and tubular halloysite(see TEM image in the lower part of Fig 7) TEMcharacterization of objects in Fig 8 confirmed thatthe filaments consist of aggregated iron oxyhy-droxides An interesting phenomenon revealedby the TEM is the presence of a core of semi-aligned crystals of FeOOH within these aggre-gates (Welch et al 2001 Chan et al manuscriptin preparation) While organisms may have con-tributed to the formation of these structures theaggregates cannot be identified as cells or miner-alized cells
BANFIELD ET AL452
FIG 4 TEM image of ZnS pro-duced as a result of the activity ofsulfate-reducing bacteria in solu-tions containing 1 ppm Zn (fordetails see Labrenz et al 2000)Note that the particle size is excep-tionally small often 2 nm(smaller black arrows) The largerblack arrow indicates an aggregateof slightly misoriented particlesThe black and white arrow marksa particle exhibiting a stacking dis-order of intergrown sphalerite andwurtzite structures
Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

abundance and distribution of surface alterationproducts For example sulfur-oxidizing bacteriarapidly remove elemental sulfur from the sur-faces of dissolving metal sulfide minerals(McGuire et al 2001) In the absence of sulfur-ox-idizing microbes elemental sulfur would accu-mulate owing to its slow dissolution kinetics inmost aqueous solutions
The stoichiometry of dissolution of silicateminerals is variable depending on the pH andwhether organic ligands are present In acidic so-lutions regardless of whether organic ligands arepresent the net release of AlSi from the feldsparframework can be either approximately stoichio-metric with respect to the dissolving phase or Alcan be preferentially released to solution leavingSi-enriched surface layers At near neutral pH inthe absence of organic compounds Si is prefer-entially released to solution compared with Alleaving Al-rich residual layers However if or-ganic ligands are present the AlSi release is stoi-chiometric (Welch and Ullman 1993 Stillings etal 1996) It is not certain that such experimentalresults translate to the identification of usefulbiosignatures First observed phenomena cannotbe interpreted uniquely without additional con-straints and second clay mineral assemblagesare the likely surface products of silicate dissolu-tion under most conditions given sufficient time
Minerals whose crystal morphology indicates abiogenic origin
Most magnetotactic organisms arrange theirmagnetic crystals into chains Some species pro-duce biogenic magnetite with unique character-istics elongated nonequilibrium prismatic mor-phology chemically pure few defects includingtwins restricted to single domainsuperpara-magnetic size range restricted widthlength ra-tio and restricted anisotrophic size distribution(nonndashlog-normal distribution or truncated) Ithas been argued that on Earth magnetites withthese characteristics form only as the result of mi-crobial activity (Thomas-Keprta et al 2000 2001)Collapsed chains of magnetite crystals have beenused as biosignatures for magnetotactic bacteriain terrestrial soil samples (Fassbinder et al 1990Kirshvink and Vali 1999)
More generally crystal growth in the presenceof organic ligands can modify the crystal habit(crystal form) of any phase through alteration ofthe relative stability of crystallographically dis-
tinct surfaces (eg Mann et al 1993 Albeck et al1996)
Minerals whose surface morphology indicatesbiological impact
Organisms can affect the way in which the sur-face topography of a mineral evolves by induc-ing localized pitting Specific surface step mor-phologies and larger-scale surface topographieshave been shown to arise when minerals dissolvein the presence of organic compounds [egamino acids (Teng and Dove 1997 see Fig 1Welch et al 2002 see Fig 2)] ldquoCell-sized and cell-shapedrdquo etch pits observed on the surfaces ofmany minerals have been attributed to enhanceddissolution under attached cells [eg silicates(Fisk et al 1998 also see Thorseth et al 1995)] Insome cases it has been confirmed experimentallythat the cells are associated with pit formationand thus the pits are of microbial origin (Fig 3)However a variety of nonbiological processescan lead to formation of micron-scale pits and itmay be very difficult to develop criteria to dis-tinguish these from pits formed by microorgan-isms (eg Edwards et al 2001)
Mineral accumulations with unusual particle size
Typically when microbial redox transforma-tions induce mineral precipitation external to thecell the particle size of the products is excep-tionally small often 2ndash5 nm (Fig 4) For exam-ple enzymatic catalysis of iron oxidation by neu-trophilic iron-oxidizing bacteria leads to theformation of few-nanometer-diameter iron oxy-hydroxide particles (eg Banfield et al 2000)
BANFIELD ET AL450
FIG 1 Atomic force microscope image of calcite re-acted inorganically (left) and in the presence of anamino acid (Asp as inset right) The field of view of eachimage is 10 mm Note the very different surface step mor-phologies (photomicrograph courtesy of Patricia Dovefrom Dove and Grantham manuscript in preparation)
Small particle size does not arise uniquely owingto biological activity However nanoparticle for-mation is favored when enzymatic activity causesrapid supersaturation (leading to many simulta-neous nucleation events) under conditions wherecrystal growth would be inhibited or occur at avery slow rate In addition large crystal size orunusual crystal length in one or two dimensionsmay result from polymer-based nucleation(Calvert and Rieke 1996 MacLachlan et al 2000)
Minerals whose spatial arrangement suggests thepresence of cells
Mineral aggregates can assume unusual diag-nostically biological shapes For example ironand manganese oxides precipitate onto polymerstrands or sheaths of bacteria and preserve theshapes of these polymers Classic examples in-clude the mineralized twisted ribbons generatedby Gallionella spp and the drinking straw-likestructures formed by Leptothrix spp (Fig 5) Theproducts of these organisms have been used toinfer a biogenic origin for Fe-rich deposits in thegeological record (eg Alt 1988) Examples in-clude Cambrian sea-floor silicandashiron oxide de-posits described by Duhig et al (1992) and a Juras-sic hydrothermal vent community described byLittle et al (1999) (also see review by Hofmannand Farmer 2000)
Minerals often precipitate on or adhere to cellsurfaces For example crystallization of sulfatesand carbonates onto cell surface layers (eg pe-riodic S-layers) may lead to products with dis-tinctive size microstructure and aggregate char-acteristics (Schultze-Lam et al 1996 Fortin et al1997) In some cases mineralization may preservethe morphology of the microorganisms even af-ter the cell materials have degraded (eg mor-phological microfossils) However identificationof the former presence of life based on cell-likemorphologies is fraught with problems Manymineralization processes can generate small cell-shaped and approximately cell-sized objectsFurthermore some minerals commonly encoun-tered in soil and aqueous environments grow asmicron- and submicron-sized spherical or tubu-lar particles In some cases such morphologicaldata has been used to argue for the existence oflife forms smaller than the most commonly ac-cepted ldquosmallestrdquo bacteria [eg ldquonannobacteriardquo(Folk and Lynch 1997 Folk 1999)] generatingconsiderable controversy (also see Kirkland et al1999) Figures 6ndash8 are scanning electron micro-scope (SEM) images of micron- and submicron-scale objects resembling rounded rod- and spir-rilum-shaped cells respectively The objectsdisplay few characteristics typical of minerals(eg crystal faces) and they are fairly uniform insize These characteristics could lead the observerto conclude that the objects are cells or mineral-ized cells
MINERALOGICAL BIOSIGNATURES 451
FIG 2 Examples of the response of the apatite surfaceto experimental dissolution in (a) inorganic acid and (b)the presence of actively metabolizing cells Note the ar-row in (b) indicates the location of a cell Scale bars 5 2mm In this case the surface texture may provide evidencefor the presence of organic acids at the surface during dis-solution (Welch et al 2002)
FIG 3 SEM image of a cell-shaped pit formed under-neath a surface-attached microorganism that was dehy-drated under vacuum During the experiment the roundor disc-shaped cells of the iron-oxidizer Ferroplasma aci-darmanus colonized highly polished arsenopyrite (FeAsS)surfaces Pitting of the surface surrounding the cell is dueto the release of ferric iron which oxidizes surface sulfidegroups (see Edwards et al 2001)
a
b
The SEM is a powerful tool for detection andcharacterization of cells (eg Firtel and Bev-eridge 1995) However to verify that an objectthat looks like a cell is (or was) a cell it is neces-sary first to establish what cells should look likevia the method of observation Hydrated cellsand nonconducting material can be viewed withan environmental SEM (Collins et al 1993) withlimited resolution (10 nm) Dehydrated cells canbe examined with scanning electron microscopyat low accelerating voltages (a few kilovolts) butare difficult to visualize (Fig 9) in the absence ofmineralization Better resolution requires fixa-tion either freezing (as for Fig 10) or chemicaltreatment followed by dehydration and criticalpoint drying Cells can be ion-sputter-coated withgold palladium iridium chromium or otherheavy metals and examined at higher voltages(15ndash20 kV) Fixation dehydration and sputter-coating increase the possibly of better imagingbut also increase the possibility for introducingartifacts that may complicate interpretation ofshapes (Little et al 1991) In general additional
evidence such as a DNA or RNA stain (usuallyobserved via epifluorescence optical microscopy)is highly desirable If possible additional charac-terization methods should be employed
Transmission electron microscope (TEM)analysis of the cell-like objects shown in Figure 6revealed that they are composed of aggregates ofa fine-grain particulate lanthanide phosphatemineral TEM cross sections of the material illus-trated in Fig 7 coupled with energy-dispersivemicroanalysis demonstrated conclusively thatthe objects are spherical and tubular halloysite(see TEM image in the lower part of Fig 7) TEMcharacterization of objects in Fig 8 confirmed thatthe filaments consist of aggregated iron oxyhy-droxides An interesting phenomenon revealedby the TEM is the presence of a core of semi-aligned crystals of FeOOH within these aggre-gates (Welch et al 2001 Chan et al manuscriptin preparation) While organisms may have con-tributed to the formation of these structures theaggregates cannot be identified as cells or miner-alized cells
BANFIELD ET AL452
FIG 4 TEM image of ZnS pro-duced as a result of the activity ofsulfate-reducing bacteria in solu-tions containing 1 ppm Zn (fordetails see Labrenz et al 2000)Note that the particle size is excep-tionally small often 2 nm(smaller black arrows) The largerblack arrow indicates an aggregateof slightly misoriented particlesThe black and white arrow marksa particle exhibiting a stacking dis-order of intergrown sphalerite andwurtzite structures
Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats
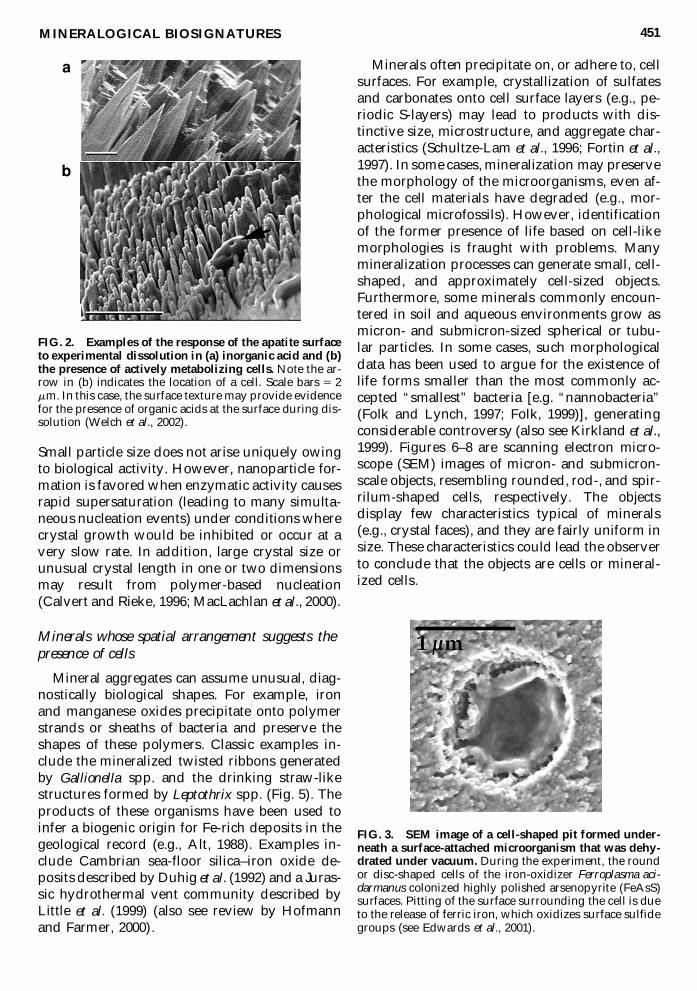
Small particle size does not arise uniquely owingto biological activity However nanoparticle for-mation is favored when enzymatic activity causesrapid supersaturation (leading to many simulta-neous nucleation events) under conditions wherecrystal growth would be inhibited or occur at avery slow rate In addition large crystal size orunusual crystal length in one or two dimensionsmay result from polymer-based nucleation(Calvert and Rieke 1996 MacLachlan et al 2000)
Minerals whose spatial arrangement suggests thepresence of cells
Mineral aggregates can assume unusual diag-nostically biological shapes For example ironand manganese oxides precipitate onto polymerstrands or sheaths of bacteria and preserve theshapes of these polymers Classic examples in-clude the mineralized twisted ribbons generatedby Gallionella spp and the drinking straw-likestructures formed by Leptothrix spp (Fig 5) Theproducts of these organisms have been used toinfer a biogenic origin for Fe-rich deposits in thegeological record (eg Alt 1988) Examples in-clude Cambrian sea-floor silicandashiron oxide de-posits described by Duhig et al (1992) and a Juras-sic hydrothermal vent community described byLittle et al (1999) (also see review by Hofmannand Farmer 2000)
Minerals often precipitate on or adhere to cellsurfaces For example crystallization of sulfatesand carbonates onto cell surface layers (eg pe-riodic S-layers) may lead to products with dis-tinctive size microstructure and aggregate char-acteristics (Schultze-Lam et al 1996 Fortin et al1997) In some cases mineralization may preservethe morphology of the microorganisms even af-ter the cell materials have degraded (eg mor-phological microfossils) However identificationof the former presence of life based on cell-likemorphologies is fraught with problems Manymineralization processes can generate small cell-shaped and approximately cell-sized objectsFurthermore some minerals commonly encoun-tered in soil and aqueous environments grow asmicron- and submicron-sized spherical or tubu-lar particles In some cases such morphologicaldata has been used to argue for the existence oflife forms smaller than the most commonly ac-cepted ldquosmallestrdquo bacteria [eg ldquonannobacteriardquo(Folk and Lynch 1997 Folk 1999)] generatingconsiderable controversy (also see Kirkland et al1999) Figures 6ndash8 are scanning electron micro-scope (SEM) images of micron- and submicron-scale objects resembling rounded rod- and spir-rilum-shaped cells respectively The objectsdisplay few characteristics typical of minerals(eg crystal faces) and they are fairly uniform insize These characteristics could lead the observerto conclude that the objects are cells or mineral-ized cells
MINERALOGICAL BIOSIGNATURES 451
FIG 2 Examples of the response of the apatite surfaceto experimental dissolution in (a) inorganic acid and (b)the presence of actively metabolizing cells Note the ar-row in (b) indicates the location of a cell Scale bars 5 2mm In this case the surface texture may provide evidencefor the presence of organic acids at the surface during dis-solution (Welch et al 2002)
FIG 3 SEM image of a cell-shaped pit formed under-neath a surface-attached microorganism that was dehy-drated under vacuum During the experiment the roundor disc-shaped cells of the iron-oxidizer Ferroplasma aci-darmanus colonized highly polished arsenopyrite (FeAsS)surfaces Pitting of the surface surrounding the cell is dueto the release of ferric iron which oxidizes surface sulfidegroups (see Edwards et al 2001)
a
b
The SEM is a powerful tool for detection andcharacterization of cells (eg Firtel and Bev-eridge 1995) However to verify that an objectthat looks like a cell is (or was) a cell it is neces-sary first to establish what cells should look likevia the method of observation Hydrated cellsand nonconducting material can be viewed withan environmental SEM (Collins et al 1993) withlimited resolution (10 nm) Dehydrated cells canbe examined with scanning electron microscopyat low accelerating voltages (a few kilovolts) butare difficult to visualize (Fig 9) in the absence ofmineralization Better resolution requires fixa-tion either freezing (as for Fig 10) or chemicaltreatment followed by dehydration and criticalpoint drying Cells can be ion-sputter-coated withgold palladium iridium chromium or otherheavy metals and examined at higher voltages(15ndash20 kV) Fixation dehydration and sputter-coating increase the possibly of better imagingbut also increase the possibility for introducingartifacts that may complicate interpretation ofshapes (Little et al 1991) In general additional
evidence such as a DNA or RNA stain (usuallyobserved via epifluorescence optical microscopy)is highly desirable If possible additional charac-terization methods should be employed
Transmission electron microscope (TEM)analysis of the cell-like objects shown in Figure 6revealed that they are composed of aggregates ofa fine-grain particulate lanthanide phosphatemineral TEM cross sections of the material illus-trated in Fig 7 coupled with energy-dispersivemicroanalysis demonstrated conclusively thatthe objects are spherical and tubular halloysite(see TEM image in the lower part of Fig 7) TEMcharacterization of objects in Fig 8 confirmed thatthe filaments consist of aggregated iron oxyhy-droxides An interesting phenomenon revealedby the TEM is the presence of a core of semi-aligned crystals of FeOOH within these aggre-gates (Welch et al 2001 Chan et al manuscriptin preparation) While organisms may have con-tributed to the formation of these structures theaggregates cannot be identified as cells or miner-alized cells
BANFIELD ET AL452
FIG 4 TEM image of ZnS pro-duced as a result of the activity ofsulfate-reducing bacteria in solu-tions containing 1 ppm Zn (fordetails see Labrenz et al 2000)Note that the particle size is excep-tionally small often 2 nm(smaller black arrows) The largerblack arrow indicates an aggregateof slightly misoriented particlesThe black and white arrow marksa particle exhibiting a stacking dis-order of intergrown sphalerite andwurtzite structures
Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

The SEM is a powerful tool for detection andcharacterization of cells (eg Firtel and Bev-eridge 1995) However to verify that an objectthat looks like a cell is (or was) a cell it is neces-sary first to establish what cells should look likevia the method of observation Hydrated cellsand nonconducting material can be viewed withan environmental SEM (Collins et al 1993) withlimited resolution (10 nm) Dehydrated cells canbe examined with scanning electron microscopyat low accelerating voltages (a few kilovolts) butare difficult to visualize (Fig 9) in the absence ofmineralization Better resolution requires fixa-tion either freezing (as for Fig 10) or chemicaltreatment followed by dehydration and criticalpoint drying Cells can be ion-sputter-coated withgold palladium iridium chromium or otherheavy metals and examined at higher voltages(15ndash20 kV) Fixation dehydration and sputter-coating increase the possibly of better imagingbut also increase the possibility for introducingartifacts that may complicate interpretation ofshapes (Little et al 1991) In general additional
evidence such as a DNA or RNA stain (usuallyobserved via epifluorescence optical microscopy)is highly desirable If possible additional charac-terization methods should be employed
Transmission electron microscope (TEM)analysis of the cell-like objects shown in Figure 6revealed that they are composed of aggregates ofa fine-grain particulate lanthanide phosphatemineral TEM cross sections of the material illus-trated in Fig 7 coupled with energy-dispersivemicroanalysis demonstrated conclusively thatthe objects are spherical and tubular halloysite(see TEM image in the lower part of Fig 7) TEMcharacterization of objects in Fig 8 confirmed thatthe filaments consist of aggregated iron oxyhy-droxides An interesting phenomenon revealedby the TEM is the presence of a core of semi-aligned crystals of FeOOH within these aggre-gates (Welch et al 2001 Chan et al manuscriptin preparation) While organisms may have con-tributed to the formation of these structures theaggregates cannot be identified as cells or miner-alized cells
BANFIELD ET AL452
FIG 4 TEM image of ZnS pro-duced as a result of the activity ofsulfate-reducing bacteria in solu-tions containing 1 ppm Zn (fordetails see Labrenz et al 2000)Note that the particle size is excep-tionally small often 2 nm(smaller black arrows) The largerblack arrow indicates an aggregateof slightly misoriented particlesThe black and white arrow marksa particle exhibiting a stacking dis-order of intergrown sphalerite andwurtzite structures
Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

Spatial patterns of minerals can provide veryconvincing evidence that life existed in an envi-ronment (eg trace fossils) Bioturbation con-tributes significantly to the development of soiland sediment structure and can impart distinc-tive textural and mineralogical organization [egworm tubes (Aller 1984)] Soil aggregates are alsobiologicalndashmineralogical entities with potentialutility as biosignatures
Minerals whose aggregation state indicates a rolefor low-molecular-weight by-products
Distinctive solution chemistry particularly thepresence of small organic molecules may lead toparticle aggregation patterns that would not beanticipated in an inorganic system For exampleaggregates may arise when organic ligands bindto nanoparticle surfaces and bridge adjacent par-ticles to form three-dimensional arrays The ex-ternal shape of the resulting aggregates is oftenspherical (Fig 6) However within the array thenanoparticles may be highly oriented
Presence of organic molecules in crystalsindicating biomineralization
Organic polymers are often incorporated intobiominerals as they form (eg shells pearlsbone and coccoliths formed by calcareous algae)Microbial cells can be permeated by silica fixingorganic molecules in varying stages of degrada-tion During mineral replacement the organicstructure acts as a template Minerals replacingmicroorganisms and potentially intergrown withorganic polymers include silica clays oxides sul-fides carbonates and phosphates
Unusual mineral assemblages indicating abiological process
Certain mineral assemblages may be impossi-ble to form without biological activity For ex-ample associations of finely particulate metalsulfides with low-temperature phases (eg clayminerals) in very low-grade carbon-rich sedi-ments are likely to have a biological origin ther-mochemical sulfate reduction is strongly inhib-ited below 175degC over reasonable time scalesdue to large kinetic barriers (eg Ohmoto andLasaga 1982) Though other processes can gen-erate very small sulfide particles (eg impactabrasion and condensation from the solar nebu-lar) these products have compositional charac-
MINERALOGICAL BIOSIGNATURES 453
FIG 5 Biomineralization products of iron-oxidizingbacteria Ferric iron oxyhydroxides coat the cell or cell-produced polymers giving rise to highly diagnosticshapes a Field emission SEM image of Leptothrix sheathcoated with ferrihydrite b Field emission SEM image ofa Gallionella stalk coated with ferrihydrite c TEM imageof mineralized polymer produced by a new isolate of aniron-oxidizing bacteria (photomicrograph courtesy ofEmerson unpublished) Larger crystals coating the poly-mer are the FeOOH phase akaganeite Although Martianiron-oxidizing cells may have different forms their mor-phologies should be preserved by a similar process (egChan et al manuscript in preparation)
a
b
c
teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

teristics (eg elevated Ni concentration) and as-sociations with other minerals that indicate theirnonbiological origin [eg metal sulfide nanopar-ticles in carbonaceous chondrites (Lauretta et al1997)]
Specific major element distribution patternsindicating biological effects
Abundance patterns for biologically essentialmajor elements may indicate biological activityFor example phosphate depletion in biologicallyaltered weathering profiles may be strikingTaunton et al (2000ab) showed that lanthanideand phosphorus concentrations varied dramati-cally between biologically altered and predomi-nantly inorganically chemically weathered zonesThey attributed such chemical variations to mi-crobial phosphate scavenging in low-phosphateenvironments (eg P uptake in combination withcomplexation of the metals by an organic ligandleading to dissolution of otherwise extremely in-soluble phosphate minerals) The end result ofthis process was a change in mineralogy Afterphosphate removal Ce31 was oxidized to Ce41
and precipitated as insoluble oxides in biologi-cally altered zones Zones little impacted by mi-
croorganisms contained abundant lanthanidephosphate phases
Some organisms deploy siderophores to scav-enge iron Iron oxides may also be solubilized byreductive dissolution facilitated by soluble or-ganic electron shuttle molecules [eg 26-AQDS(see review by Hernandez and Newman 2002)]Both of these processes may also lead to major el-ement redistribution patterns that could consti-tute biosignatures
Terrestrial soils contain numerous potentialbiosignatures In addition to high concentrationsof organic compounds the patterns of elementdistribution may strongly indicate the operationof biological pathways For example soils con-
BANFIELD ET AL454
FIG 6 SEM image of rounded objects with shapes andsizes resembling microbial cells Compositional analy-sis and TEM characterization demonstrated that these ob-jects are precipitates of lanthanide aluminum phosphateand weathering products of apatite (reproduced with per-mission from Banfield and Eggleton 1989)
FIG 7 SEM image of rod-shaped and round objectson the surface of a mineral in soil Compositional analy-sis and TEM characterization show these to be sphericalhalloysite and halloysite tubes (lower image see Banfieldand Eggleton 1990 for details)
taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

taining abundant organic acids tend to be de-pleted in Al and Fe because these elements arecomplexed and solubilized by organic ligands Incontrast Al and Fe are concentrated in organic-poor environments For example Weed and Nor-ton (1991) and Johnston and Vestal (1993) showedthat organic acids such as oxalate caused reloca-tion of Al and Fe to deeper portions of Antarcticweathering profiles compared with sites withoutsignificant biological activity In Australian gran-ite weathering profiles geochemical data sug-gested that K and Mg are depleted in saproliteimmediately below the soil profile which is con-sistent with faster biologically induced weather-ing of biotite compared with feldspar (Welch etal 1999)
Unusual trace element accumulations mayindicate biological impact
Microorganisms require small quantities ofspecific elements as enzyme cofactors For exam-ple Archaea require Ni for certain enzymes in-cluding most hydrogenases carbon monoxidedehydrogenase and coenzyme F430mdashpart of thekey enzyme involved in methanogenesis (Ermleret al 1997) Also Cu is required by some bacte-ria for cytochrome c oxidase and Mn is presentin certain superoxide dismutases and P680chlorophyll a (the water-splitting enzyme in mostoxygenic phototrophs) (eg Bratton et al 2000Nugent et al 2001)
If a biologically essential ion such as Co NiCu Mn Mo Se and V which are usually foundin low abundance is scavenged from the envi-ronment concentrations of these elements couldconstitute a mineralogical biosignature Further-more toxic ions are sequestered into granules bysome microorganisms For example Se is se-questered into S granules by sulfide-oxidizingbacteria (Nelson et al 1996) Recent studies showuranium-rich phosphate granules inside sulfate-reducing bacteria in the aerobic zone of U-con-taminated sediments (Suzuki and Banfield 2001)Consequently biological activity may be indi-cated both by occurrence of unusual trace ele-ment concentrations and by unexpected elemen-tal ratios Other examples of potential importanceinclude fractionation of ions with geochemicallysimilar distribution patterns due to differentialcomplexation by organic molecules [eg Ge vsSi Al vs Ga (Pokrovski and Schott 1998)]
MINERALOGICAL BIOSIGNATURES 455
FIG 8 SEM image of high-contrast curled roundedobjects Although these filamentous forms resemble verysmall cells TEM data below indicate that they consist ofaggregates of fine particulate iron oxyhydroxide miner-als As shown in the bottom image the filaments containa very thin FeOOH core consisting of aligned nanocrys-tals Although these objects are clearly not cells the roleof cells in their formation cannot be ruled out
MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

MARTIAN GEOCHEMISTRYCONSTRAINTS FOR POSSIBLE BIOMES
It is likely that a Martian biosphere will be lo-calized in interfacial reaction zones It is reason-able to propose that for most of Marsrsquo history theprocess of planetary evolution in combinationwith atmospheric photochemical processes andcatastrophic events such as meteorite impactshas maintained some level of disequilibriumamong the atmosphere hydrosphere and litho-sphere Thus in addition to sunlight there aresources of energy that could be harvested by lifeforms
Soil samples taken at the Pathfinder site closelyresemble those collected several thousand kilo-meters away at the two Viking sites (Economouet al 1998 Larsen et al 2000) Data have been in-terpreted to indicate that the aeolian dust is aphysical mixture of basalt and andesite-derivedmaterials in approximately equal proportions(Larsen et al 2000) In addition a close chemicalrelationship has been established for Martian me-teorites and Pathfinder rocks (Driebus 1998)Pathfinder data indicate the presence of high-sil-ica rocks with SiO2 compositions ranging from525 to 612 wt The concentration of SiO2 is in-versely correlated with sulfur extrapolating to anS-free rock at an SiO2 concentration of 62 wt(Britt et al 1998) Rocks composing the Martian
surface are inferred to be Mg-Fe-basalts and lowMg- low S-ldquoandesitesrdquo (also see Hamilton et al2001) Analysis of SNC meteorites suggests thatMars rocks are reduced and contain primarily fer-rous iron with minor amounts of reduced sulfurPrimary minerals in SNC rocks include olivinespyroxenes chromite magnetite apatite pyriteand pyhrrotite Alteration products reportedfrom SNC meteorites include hematite goethitemagnetite smectite siderite nanophase ferric ox-ideshydroxides halite and gypsum (McSweenand Treiman 1998)
Martian soil is highly oxidized (Fe21Fe31 5073 in rock Fe21Fe31 5 320 in soils) and sul-fate is a major component When compared withrock compositions soils are depleted in silicaaluminum and potassium while enriched in ti-tanium chromium iron magnesium and sulfur(Britt et al 1998 Economou et al 1998 Bell et al2000) The soil is inferred to be acidic Iron is inthe form of composites of nanophase ferric iron-rich minerals and more coarsely crystallinehematite Other constituents inferred to be pres-ent include sulfates probably calcium sulfates(gypsum) Ferrous silicates occur in the rocks andsoil but do not appear to be present in the dustOver a significant period of Martian surficial his-
BANFIELD ET AL456
FIG 9 SEM image of a living or recently dead cellNote that the cell which is barely visible in the image isdehydrated (white arrows mark cell margins) The imagewas recorded using the SE2 detector of an LEO 1530 fieldemission SEM with accelerating voltage of 3 kV and 4 mmworking distance If cells are imaged at higher accelerat-ing voltage they are virtually invisible unless they aremineralized or fixed via cryogenic or chemical meansprior to imaging
FIG 10 SEM image of the ZnS biofilm described byLabrenz et al (2000) The sample was impregnated andprepared for microscopy via a freeze-etch method (RWebb personal communication 2000) The SEM imageshows microbial cells and polymers associated withspherical aggregates The aggregates consist of 108nanoparticles of ZnS (see Fig 4) produced by sulfate-re-ducing bacteria The spherical shape has been reproducedin experiments where ZnS is precipitated in the presenceof organic molecules (Huang et al manuscript in prepa-ration)
tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

tory redox reactions many of which probably in-volved Fe and S occurred at the planet surface(Bell et al 2000 Edgett and Malin 2000 Morriset al 2000)
BIOSIGNATURES IN THE MARTIAN CONTEXT
Models for the Martian hydrological cycle in-clude episodic ocean formation glaciation andflooding and persistence of ocean regions earlyin the history of Mars (Baker et al 1991 Malinand Edgett 2000) Ground ice in the crust and soilmay be significant modern reservoirs of water onMars (eg Clifford 1993 Mustard et al 2001)Evidence includes geomorphologic features in-dicative of viscous creep at higher latitudes Mid-latitude soils have also been identified as reser-voirs for water (Fanale et al 1986) Mustard et al(2001) report evidence for a 1ndash10-m-thick reser-voir of near-surface water as ice within the porespace of soils in the 30ndash60deg latitude range
Oceans or lakes represent potential locationsfor microbial communities These could be simi-lar to environments from the early Earth in whichmicrofossils are found (eg shallow-marine or la-custrine basins) Organisms that derive energyfrom inorganic compounds or light might be an-ticipated The last open water on Mars was mostlikely found in alkaline hypersaline lakes Thesebodies of water could have been stratified lead-ing to a range of redox environments withinwhich various types of metabolisms could haveoperated Furthermore a fluviallacustrineocean system could be expected to produce sed-iments and thus a range of sedimentary habitatssimilar to those encountered in terrestrial set-tings Clearly this scenario gives rise to a widediversity of geochemical cycles that may producemineralogical biosignatures for which terrestrialanalogs exist
Martian environments in which the formationof terrestrial mineral biosignatures could occurinclude subsurface volcanoes hydrothermalvent systems metal sulfide-rich deposits andsurface water gradients Though microorgan-isms in extraterrestrial habitats may haveevolved different metabolic activities than ter-restrial organisms they would have had to in-teract with similar rock surfaces and may havedeveloped similar strategies to cope with envi-ronmental challenges Such interactions may
have led to comparable patterns of elemental dis-tribution and mineral formation Furthermoreeven if metabolic differences existed life is likelyto rely on some form of electron transfer reac-tions Thus focus on available redox-active Mar-tian surface mineral constituents may be appro-priate when developing model systems formineralogical biosignature analysis A few ex-amples of possible environments and their po-tential to produce mineralogical biosignatures arepresented below
Iron
Redox disequilibrium is a key requirement forchemoautotrophic biogeochemical systems Ironis one of the most abundant redox-active com-ponents of the Martian lithosphere It occurs inreduced forms in rocks and in oxidized forms indust and sediments One approach in the searchfor Martian mineral biosignatures would be to fo-cus on the chemistry of Fe and other electrondonors such as sulfur compounds or H2 Thuswe consider the potential environments where re-dox reactions involving Fe may sustain life onMars and the possible mineralogical biosigna-tures associated with these
Even if the biochemical design of extraterres-trial life is not comparable to that on Earth thecharacteristics of mineral precipitates resultingfrom organism-based catalysis of iron redox re-actions may be similar In terrestrial microbialmetabolisms operating under anoxic conditionsFe31 can be used as an electron acceptor Con-versely under oxidizing conditions Fe21 can beused as an electron donor A number of elementscan serve as the related electron donors and ac-ceptors
The formation of very small intracellular mag-netite (Fe3O4) or greigite (Fe3S4) crystals in somecases with unusual morphologies is associatedwith microorganisms that populate relativelylow oxygen environments (eg Posfai et al1998) In addition to magnetosome magnetite (seeBazylinski et al 1995) extracellular oxides andsulfides are produced in large quantities underanoxic conditions by iron- and sulfate-reducingbacteria [eg dissimilatory iron-reducing bacte-ria (Fredrickson et al 1998)] Moskowitz et al(1989) discussed the differences between theproducts of extracellular and intracellular ironmineralization reactions The use of magnetitesize morphology and microstructure as a biosig-
MINERALOGICAL BIOSIGNATURES 457
nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

nature has been a matter of considerable recentdebate with regard to the biogenicity of magnetitein the Martian meteorite ALH84001 (eg Goldenet al 2001 Thomas-Keprta et al 2001)
Martian organisms that utilize ferric iron as ispresent in Martian dust as an electron acceptormight be encountered in anoxic aqueous or sed-imentary habitats Other Martian organismscould provide reduced organic carbon com-pounds to serve as electron donors Ferric iron re-duction could also be coupled to the oxidation ofhydrogen possibly supplied from the planetaryinterior (eg geothermal systems)
A number of metabolic options that rely uponthe use oxidation of Fe21 are possible on MarsThough organisms tend not to use iron oxidationas an energy source in well-oxidized solutionsowing to rapid inorganic oxidation kinetics theycan do so when the kinetics of inorganic oxida-tion reactions are inhibited by high acidity or lowoxygen concentrations Thus on Earth iron oxi-dation forms the basis of ecosystems located inacid rockmine drainage environments or wheresteep gradients occur between oxic and anoxic re-gions in solutions or sediments (eg water wellsflooded underground mines)
A potential Martian biome could rely on dis-equilibrium between ferrous iron-rich volcanics(mafic rocks) and the atmosphere or hydro-sphere In this scenario organisms might use ironoxidation for metabolic energy generation in thegradient between the oxic surface and reducedrock reservoirs or in subsurface hydrous envi-ronments in proximity to basalts and basaltic an-desites Subsurface lava tubes as suggested bychains of collapsed structures imaged by the MarsGlobal Surveyor could provide open space insubsurface aqueous environments
On Earth ferrous iron utilized in energy gen-eration is usually supplied by mineral dissolu-tion either through reductive dissolution of fer-ric iron minerals in adjacent anoxic zones or bydissolution of ferrous iron sulfides or silicates Ifiron sulfides are the source of iron sulfuric acidis generated owing to sulfide oxidation (see be-low) This can lead to low pH environments thatcan sustain significant biomass owing to the slowrate of inorganic iron oxidation in acid solutions(eg Bond et al 2000) If the pH buffering ca-pacity of the surrounding rocks is small the pHof weathering solutions may remain low andlimit precipitation of ferric iron minerals and the
formation of mineralogical biosignatures Proba-ble mineralogical biosignatures formed underthese conditions may be restricted to etching fea-tures on sulfide mineral surfaces such as the dis-tinctive pits created by direct contact betweencells and the surface Since pits can form inor-ganically however the interpretation of suchetch features as biological in origin is often diffi-cult if not impossible (Edwards et al 2001)
Microbiological communities supported by theoxidation of ferrous iron released by silicate min-eral dissolution at near-neutral pH are relativelyunstudied though some effort has been devotedto analysis of iron-oxidizing bacteria in ground-water soils and sediments (eg Emerson andMoyer 1997) Neutrophilic iron-oxidizing mi-crobes are difficult to cultivate and are typicallystudied some distance from the dissolving min-erals However an effective metabolism couldharvest iron released from the surface of fayalite(an Fe-silicate) by dissolution as follows
Fe2SiO4 1 4H1 THORN 2Fe21 1 H4SiO4
Oxidation of aqueous ferrous iron in slightly ox-idized solutions could be coupled to the reduc-tion of oxygen nitrate or other electron accep-tors
Thermodynamic calculations indicate ampleenergy is available for microbiological metabo-lism from iron silicate redox reactions (Jakoskyand Shock 1998) Recent kinetic studies suggestthat in aqueous solutions Fe21 is released by dis-solution of phases such as biotite pyroxene andolivine at rates sufficient to sustain significant mi-crobial populations especially under acidic con-ditions (Santelli et al 2001)
Mineral deposits associated with iron-oxidizingbacteria living at neutral pH have very high po-tential as mineral biosignatures Both the mor-phology and mineralogy of the products may bequite distinctive In some cases biogenicity may beindicated by filmy mineralized sheets similar tothose produced by Mn-oxidizing bacteria (Man-dernack et al 1995) In other cases Fe accumulateson extracellular polymeric compounds leading tothe formation of assemblages such as those shownin Fig 5 for Gallionella and Leptothrix In the case ofsome isolates of iron-oxidizing bacteria the min-eral produced is either ferrihydrite or feroxyhite(Banfield et al 2000) In other examples the pre-cipitate may be akaganeite (Chan et al manuscript
BANFIELD ET AL458
in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

in preparation) sometimes displaying extremelyunusual morphologies (Welch et al 2001) Mineralbiosignatures such as these have been used to in-fer a biogenic origin for ancient iron deposits asreferenced above In terrestrial examples identifi-cation of morphological biosignatures relies uponthe existence of life forms similar to those found onthe Earth Although life forms identical to those onEarth may not have emerged on Mars rapid pre-cipitation of nanophase iron oxyhydroxides with astrong tendency to flocculate or adhere to organiccompounds may give rise to distinctly bioinorganicmaterials
During the production of the large-scalebanded iron formations (BIF) on Earth biologicprocesses may have played a major role in pre-cipitating iron minerals Candidate microbialgroups include purple bacteria (Widdel et al1993 Ehrenreich and Widdel 1994 Glasby 1998)neutrophilic iron oxidizers (eg Emerson andMoyer 1997) and microaerophilic organisms thatcouple iron oxidation to nitrate reduction (egStraub et al 2001) However the critical role ofmicrobes in BIF formation is far from widely ac-cepted Large (350 3 350ndash750 km) hematite de-posits in the Sinus Meridiani region of Mars de-tected by the Thermal Emission Spectrometerinstrument on the Mars Global Surveyor (Chris-tensen et al 2000) have been interpreted as pos-sible BIF analogues Christensen et al (2000) fa-vor models for the formation of the Mars hematitedeposits that involve chemical precipitation fromFe-rich water This is due in part to the appar-ent association with sedimentary materials Thefeatures of this region have led to its considera-tion as a potential sample return locality for exo-biology (eg Allen et al 1996)
One possibility for creation of the hematite-richareas on Mars involves precipitation of nano-crystalline iron oxyhydroxides from solutions via a biologically-influenced pathway Subsequentburial and heating may have converted theseproducts to coarser hematite Documentation ofall types of biosignatures in modern iron oxide-depositing environments and in ancient terres-trial BIF samples is needed to generate a com-parison database for analysis of samples from re-gions such as Sinus Meridiani This work willrequire detailed analyses of terrestrial biosigna-tures (mineralogical isotopic textural) producedby iron-oxidizing prokaryotes Since mineralsmay have been metamorphosed and altered by
subsequence heat pressure and chemical reac-tions the fate of iron mineral biosignatures dur-ing diagenesis should be investigated
The sulfur cycle
Like iron sulfur is likely to exist in multipleoxidation states at and near the Martian surfaceand could participate in energy generation reac-tions under oxic and anoxic conditions Given theextensive amount of volcanism that has occurredon Mars sulfur oxidation during the alteration ofvolcanogenic sulfide deposits is likely Other lo-cations for potential S-based Martian biomes in-clude subsurface environments where geother-mal heat could permit liquid water and at siteswhere prior hydrothermal activity may have oc-curred (eg Fisk and Giovannoni 1999)
Under oxidizing conditions microorganismson Earth can use sulfide polysulfide sulfur andsulfoxy compounds as electron donors These re-actions are frequently responsible for acidifi-cation of the environment Oxidation of metal sulfide in air or solution typically leads to accu-mulation of elemental sulfur on metal sulfidemineral surfaces These deposits are rapidly re-moved by sulfur-oxidizing bacteria Althoughperhaps difficult to interpret the absence of ele-mental sulfur accumulations and the associatedacidification may constitute a mineralogicalbiosignature In other environments where sul-fide released by communities of sulfate-reducingbacteria is oxidized by microorganisms the re-sulting elemental sulfur can be stored as granulesinside cells These granules may also constitute amineralogical biosignature Selenium follows sul-fur owing to its geochemical similarity Its bio-logical reduction reoxidation and sequestrationmay lead to a fractionation in relative abundanceof Se and S Further research is needed to evalu-ate the value of such patterns as mineralogicalbiosignatures
Under anaerobic conditions bacteria catalyze atlow temperatures the kinetically slow sulfate re-duction reaction In the presence of even smallquantities (sub ppm-level concentrations) of met-als such as Cu Zn Pb Cd and Fe this leads to pre-cipitation of nanocrystals of metal sulfide phases(eg Labrenz et al 2000) These particles tend toflocculate to form distinctive micron-scale spherespossibly owing to the presence of surface-boundorganic molecules (Fig 3) These aggregates have
MINERALOGICAL BIOSIGNATURES 459
considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

considerable potential as mineralogical biosigna-tures especially if combined with evidence of theirorigin in a low-temperature environment and forbiologic fractionation of 3432S For example someterrestrial sulfide deposits have been interpreted asformed by bacterial sulfate reduction on the basisof S isotope data (eg Ohmoto et al 1993Kakegawa et al 2000) and petrographic texturessuggestive of mineralized microbial mats (Mc-Goldrick 1999) These deposits await further de-tailed investigation and comparison with nanocrys-talline sulfides formed by sulfate-reducing bacteriain modern geologic settings and in vitro
Other targets
Sedimentary deposits have considerable poten-tial as sites of mineral biosignature formation Earlydiagenetic infusion of silica or phosphate or rapidburial of organic materials in fine-grained clayscould preserve cell textures and thus produce a va-riety of biosignatures including microfossils
Novel mechanisms for harvesting chemical orlight energy unlike those identified on Earthmay have emerged on Mars For example silicatemineral dissolution and clay precipitation reac-tions are thermodynamically favored yet kineti-cally inhibited Proton motive force may be gen-erated through precipitation of minerals on thecell surface (eg calcite and clays)
Ca(H2O)621 1 HCO32 THORN CaCO3 1 6H2O 1 H1
Biosignatures for such a process can only be in-ferred from other mineralndashorganism phenomena
OPPORTUNITIES
Our review of mineralogical phenomena aris-ing from biological activity suggests somepromising potential mineral biosignatures thatmay be developed into rigorous criteria for lifedetection However much remains to be learnedTo advance the utility of minerals as recorders ofrecent or ancient biological activity on Earth andMars and to interpret reliably this record we sug-gest two categories of further research
Building a database of mineralogicalbiosignatures
A database of mineralogical biosignatures is re-quired As discussed above there are many ways
in which organisms can impact their surround-ings Additional work is needed to explore thepotential of each of the groups of phenomena asmineral biosignatures Minerals that are pro-duced directly by organisms are of particular in-terest The forms (morphologies) and composi-tions of submicrometer magnetites sulfidesphosphates oxides and other phases from bothbiogenic and inorganic systems must be cata-logued As criteria to establish additional phe-nomena as mineral biosignatures are developedit is essential to distinguish them from the resultsof inorganic and organic processes Both charac-terization of natural systems and careful experi-mental studies are needed to achieve this goalExplicit consideration of probable differences be-tween Martian and terrestrial geological systemsis required to constrain appropriately the exper-imental conditions for inorganic controls
Given the significant number of ecosystems inwhich life and thus evidence of life may be en-countered on Earth the search for Martian min-eral biosignatures should focus initially on a sub-set of terrestrial environments selected as analogsfor targeted Martian environments Extensivedocumentation of the mineralogy geochemistryand microbiology of target terrestrial biomesmust be conducted through the study of naturalhabitats and the characterization and analysis ofsimplified systems in the laboratory The formcomposition and morphology of minerals pro-duced by cells should be analyzed in detail withenough spatial resolution to distinguish the com-position and morphology of biomineral productsand the topography of reacted surfaces at thescale at which the features vary The bulk andtrace elemental composition of minerals and thesolutions from which they form should be ana-lyzed This approach is essential in order to re-veal the extent to which microorganisms modifyelement partitioning and other aspects of thechemistry and structure of their environmentsSolution chemistry can be interpreted in the con-text of thermodynamic stability diagrams and thesimplicity principle for precipitation reactionsThis will allow identification of minerals whoseformation could not be predicted based on ther-modynamic consideration of the bulk system
Before mineralogical biosignatures can be con-fidently identified and interpreted in extremelysmall quantities of Martian materials researchersmust expand the database of unambiguous biosig-natures from the ancient Earth record For exam-
BANFIELD ET AL460
ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

ple it is critical that terrestrial BIF be analyzed inas much detail as possible and criteria be estab-lished to determine unequivocally whether suchdeposits preserve life signatures This workshould be complemented by parallel studies ofmodern sites where iron oxidendashrich deposits areaccumulating
Detection and calibration of mineral biosigna-tures in natural environments can be greatly as-sisted by laboratory experiments in which envi-ronmental factors and time can be carefullyregulated Environments should differ in pH Ehtemperature water activity solution composi-tion and pressure Within each of the target habi-tats representative groups of microbes should beidentified and analyzed in terms of the environ-mentmineral interactions Such experimentsshould be used to establish which of multiple en-vironmental variables determines the nature andextent of the resultant mineral biosignatures Theuse of sterile controls will provide valuable in-formation on the extent rate and products ofstrictly abiotic reactions In addition systematiclaboratory studies should be undertaken using avariety of microbes and their mineral products toevaluate the effects of cells on mineral surfacesunder a diversity of geochemical conditions
Understanding the fate of biosignatures
It is critical to consider how long-term aging al-teration and thermal treatment of the cellndashmineralassemblage will modify potential mineralogicalbiosignatures Most biominerals are extremelyfinely crystalline Since crystal growth is driven bysurface energy minimization it will occur at alltemperatures but over shorter times at highertemperatures Even in the absence of heating dueto burial volcanism or impacts the texture andmineralogy of assemblages will be altered espe-cially for Martian surface deposits formed billionsof years ago Specifically the crystal growth of pri-mary biominerals morphology development im-purity redistribution surface topographic changeisotope redistribution and transformation to otherstable and possibly anhydrous phases should beanalyzed Whether studies of appropriate envi-ronments will reveal that primary mineral biosig-natures are converted to recognizable ancient min-eral biosignatures by burial desiccation heatingor other processes remains to be determined
The interpretation of textures and patterns ofelemental and isotope distribution in ancient de-
posits will benefit greatly from understandinghow primary biosignatures may have been mod-ified by crystal growth phase transformationsdegradation of organic matter silicification etcBecause the size of minerals produced as the re-sult of enzymatic redox reactions is extremelysmall questions related to particle evolution overtime are directly linked to questions of nanopar-ticle properties stability and reactivity
Studies of the formation of mineralogicalbiosignatures should not be conducted withoutconsideration of their biological context Theemerging approaches that utilize genomic andgene expression data have the potential to revo-lutionize our understanding of how microorgan-isms interact with and impact their environ-ments These data could be employed to advancegreatly our understanding of mineralogical bio-signature formation
CONCLUSIONS
The search for mineralogical biosignatures thatmight establish the biogenicity of features in aMartian sample could be advanced by further de-tailed high-resolution multitechnique character-ization of both modern terrestrial ecosystems andancient deposits of presumed biogenic originMultiple methods of analysis are needed to con-strain the characteristics of the system sufficientlyand to rule out nonbiological explanations forseemingly biological phenomena In addition ex-perimental biomineralization studies and inves-tigations of the ways in which time and temper-ature modify textural and chemical biosignaturesare required These data will be critical to inter-pretation of the record of the ancient Earth as wellas of Martian samples The sites and samples cho-sen for detailed study should be the best possi-ble analogs for possible Mars biomes selected onthe basis of what is known about the Martian sur-face and subsurface regions
ACKNOWLEDGMENTS
This paper was developed following a NASA-sponsored workshop on biosignatures held inSan Diego CA in 2000 and organized by JohnKerridge Patricia Dove is thanked for providingFig 1 and John Kerridge and David McKay arethanked for their contributions to discussions
MINERALOGICAL BIOSIGNATURES 461
Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

Funding to support the collection of original dataincluded in this article was provided by theNASA Astrobiology Institute (JPL) the Depart-ment of Energy Basic Energy Sciences Programand the National Science Foundation (EAR)
ABBREVIATIONS
BIF banded iron formations SEM scanningelectron microscope TEM transmission electronmicroscope
REFERENCES
Albeck S Addadi L and Weiner S (1996) Regulationof calcite crystal morphology by intracrystalline acidicproteins and glycoproteins Connect Tissue Res 35 419ndash424
Allen MF Figueroa C Weinbaum BS Barlow SBand Allen EB (1996) Differential production of oxalateby mycorrhizal fungi in arid ecosystems Biol FertilSoils 22 287ndash292
Aller RC (1984) The importance of relict burrow struc-tures and burrow irrigation in controlling sedimentarysolute distributions Geochim Cosmochim Acta 48 1929ndash1934
Alt JC (1988) Hydrothermal oxide and nontronite de-posits on seamounts in the Eastern Pacific Marine Geol81 227ndash239
Ameye L Compere P Dille J and Dubois P (1998)Ultrastructure and cytochemistry of the early calcifica-tion site and of its mineralization organic matrix inParacentrotus lividus (Echinodermata Echinoidea) His-tochem Cell Biol 110 285ndash294
Baker VR Strom RG Gulick VC Kargel JS Ko-matsu G and Kale VS (1991) Ancient oceans icesheets and the hydrological cycle on Mars Nature 352589ndash594
Banfield JF and Eggleton RA (1989) Apatite replace-ment and rare earth mobilization fractionation and fix-ation during weathering Clay Clay Min 37 113ndash127
Banfield JF and Eggleton RA (1990) Analytical trans-mission electron microscope studies of plagioclasemuscovite and K-feldspar weathering Clay Clay Min38 77ndash89
Banfield JF Welch SA Zhang H Ebert TT andPenn RL (2000) The role of aggregation in crystalgrowth and transformations in nanophase FeOOH bio-mineralization products Science 289 751ndash754
Bazylinski DA Frankel RB Heywood BR Mann SKing JW Donaghay PL and Hanson AK (1995)Controlled biomineralization of magnetite (Fe3O4) andgreigite (Fe3S4) in a magnetotactic bacterium Appl En-viron Microbiol 61 3232ndash3239
Beard BL Johnson CM Cox L Sun H NealsonKH and Aguilar C (1999) Iron isotope biosignaturesScience 285 1889ndash1892
Bell JF III McSween HY Jr Crisp JA Morris RVMurchie SL Bridges NT Johnson JR Britt DTGolombek MP Moore HJ Ghosh A Bishop JLAnderson RC Bruumlckner J Economou T Green-wood JP Gunnlaugsson HP Hargraves RMHviid S Knudsen JM Madsen MB Reid RRieder R and Soderblom L (2000) Mineralogic andcompositional properties of Martian soil and dust re-sults from Mars Pathfinder J Geophys Res 105 1721ndash1755
Blake RE Alt JC and Martini AM (2001) Oxygen iso-tope ratios of PO4 an inorganic indicator of enzymaticactivity and P metabolism and a new biomarker in thesearch for life Proc Natl Acad Sci USA 95 2148ndash2153
Bond PL Druschel GK and Banfield JF (2000) Com-parison of acid mine drainage microbial communitiesin physically and geochemically distinct ecosystemsAppl Environ Microbiol 66 4962ndash4973
Bratton MR Hiser L Antholine WE Hoganson Cand Hosler JP (2000) Identification of the structuralsubunits required for formation of the metal centers insubunit I of cytochrome c oxidase of Rhodobactersphaeroides Biochemistry 39 12989ndash12995
Britt DT Anderson R Bell JF III Crisp J EconomouT Herkenhoff KE Madsen MB McSween HYMurchie S Reid R Rieder R Singer RB andSoderblom L (1998) The mineralogy of the MarsPathfinder landing site 29th Lunar and Planetary Sci-ence Conference Abstracts Available at wwwlpiusraedumeetingsLPSC98pdf1776pdf
Calvert P and Rieke P (1996) Biomimetic mineralizationin and on polymers Chem Material 8 1715
Christensen PR Bandfield JL Clark RN Edgett KSHamilton VE Hoefen T Kieffer HH Kuzmin ROLane MD Malin MC Morris RV Pearl JC Pear-son R Roush TL Ruff SW and Smith MD (2000)Detection of crystalline hematite mineralization onMars by the Thermal Emission Spectrometer evidencefor near-surface water J Geophys Res 105E 9623ndash9642
Clifford SM (1993) A model for the hydrologic and cli-matic behavior of water on Mars J Geophys Res Plan-ets 98 10973ndash11016
Collins SP Pope RK Scheetz RW Ray RI WagnerPA and Little BJ (1993) Advantages of environmen-tal scanning electron microscopy in studies of mi-croorganisms Microsc Res Tech 25 398ndash405
Dauphin Y (1999) Infrared spectra and elemental com-position in recent biogenic calcites relationships be-tween the nu(4) band wavelength and Sr and Mg con-centrations Appl Spectrosc 53 184ndash190
de Vrind de Jong EW and de Vrind JPM (1997) Algaldeposition of carbonates and silicates In Reviews inMineralogy Vol 35 Geomicrobiology Interactions BetweenMicrobes and Minerals edited by JF Banfield and KHNealson Mineralogical Society of America Washing-ton DC pp 267ndash307
Derry LA Kurtz AC Ziegler K Chadwick OA andKelley EF (2001) Plant phytolith source of dissolvedsilica in Hawaiian streams from GeSi ratios 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3805pdf
BANFIELD ET AL462
Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

Driebus G Ryabchikov I Rieder R Economou TBruumlckner J McSween MY Jr and Wanke H (1998)Relationship between rocks and soil at the Pathfinderlanding site and the Martian meteorites 29th Lunar andPlanetary Science Conference Abstracts Available atwwwlpiusraedumeetingsLPSC98pdf1348pdf
Duhig NC Davidson GJ and Stolz J (1992) Microbialinvolvement in the formation of Cambrian sea-floor sil-ica-iron oxide deposits Australia Geology 20 511ndash514
Economou TE Rieder R Waumlnke H Turkevich ABruumlckner J Dreibus G Crisp J and McSween H Jr(1998) The chemical composition of Martian rocks andsoil preliminary analyses 29th Lunar and PlanetaryScience Conference Available at wwwlpiusraedumeetingsLPSC98pdf1711pdf
Edgett KS and Malin MC (2000) New views of Marseolian activity materials and surface properties threevignettes from the Mars Global Surveyor Mars OrbiterCamera J Geophys Res 105 1623ndash1650
Edwards HGM Russell NC and Wynn-WilliamsDD (1997) Fourier transform Raman spectroscopic andscanning electron microscopic study of cryptoen-dolithic lichens from Antarctica J Raman Spectrosc 28685ndash690
Edwards KJ Hu B Hamers RJ and Banfield JF(2001) A new look at microbial leaching patterns on sul-fide minerals FEMS Microb Ecol 34 197ndash206
Ehrenreich A and Widdel F (1994) Anaerobic oxidationof ferrous iron by purple bacteria a new type of pho-totrophic metabolism Appl Environ Microbiol 604517ndash4526
Emerson D and Moyer C (1997) Isolation and charac-terization of novel iron-oxidizing bacteria that grow atcircumneutral pH Appl Environ Microbiol 63 4784ndash4792
Ermler U Grabarse W Shima S Goubeaud M andThauer RK (1997) Crystal structure of methyl-coen-zyme M reductase the key enzyme of biologicalmethane formation Science 278 1457ndash1462
Falini G Albeck S Weiner S and Addadi L (1996)Control of aragonite or calcite polymorphism by mol-lusk shell macromolecules Science 271 67ndash69
Fanale FP Salvail JR Zent AP and Postawko SE(1986) Global distribution and migration of subsurfaceice on Mars Icarus 67 1ndash18
Fassbinder JWE Stanjek H and Vali H (1990) Oc-currence of magnetic bacteria in soil Nature 343 161ndash163
Firtel M and Beveridge TJ (1995) Scanning probe mi-croscopy in microbiology Micron 26 347ndash362
Fisk MR and Giovannoni SJ (1999) Source of nutrientsand energy for a deep biosphere on Mars J GeophysRes Planets 104E 11805ndash11815
Fisk MR Giovannoni SJ and Thorseth IH (1998) Al-teration of oceanic volcanic glass textural evidence ofmicrobial activity Science 281 978ndash980
Folk RL (1999) Nannobacteria and the precipitation ofcarbonate in unusual environments Sediment Geol 12647ndash55
Folk RL and Lynch FL (1997) The possible role of nan-nobacteria (dwarf bacteria) in clay-mineral diagenesis
and the importance of careful sample preparation inhigh-magnification SEM study J Sediment Res 67583ndash589
Fortin D Ferris FG and Beveridge TJ (1997) Surface-mediated mineral development by bacteria In Reviewsin Mineralogy Vol 35 Geomicrobiology Interactions Be-tween Microbes and Minerals edited by JF Banfield andKH Nealson Mineralogical Society of America Wash-ington DC pp 162ndash180
Fredrickson JK Zachara JM Kennedy DW DongHL Onstott TC Hinman NW and Li SM (1998)Biogenic iron mineralization accompanying the dis-similatory reduction of hydrous ferric oxide by agroundwater bacterium Geochim Cosmochim Acta 623239ndash3257
Freytet P and Verrecchia E (1995) Discovery of Ca ox-alate crystals associated with fungi in moss travertines(bryoherms fresh-water heterogeneous stromatolites)Geomicrobiol J 13 117ndash127
Glasby GP (1998) Earliest life in the Archean rapid dis-persal of CO2-utilizing bacteria from submarine hy-drothermal vents Episodes 21 252ndash256
Golden DC Ming DW Schwandt CS Lauer HVSocki RA Morris RV Lofgren GE and McKayGA (2001) A simple inorganic process for formationof carbonates magnetite and sulfides in Martian me-teorite ALH84001 Am Mineral 86 370ndash375
Graustein WC Cromack K Jr and Sollins P (1977)Calcium oxalate occurrence in soils and effect on nu-trient and geochemical cycles Science 198 1252ndash1254
Hamilton VE Wyatt MB McSween HY and Chris-tensen PR (2001) Analysis of terrestrial and Martianvolcanic compositions using thermal emission spec-troscopymdash2 Application to Martian surface spectrafrom the Mars Global Surveyor Thermal Emission Spec-trometer J Geophys Res Planets 106 14733ndash14746
Hernandez ME and Newman DK (2001) Extracellularelectron transfer Cell Mol Life Sci 58 1562ndash1571
Hofmann BA and Farmer JD (2000) Filamentous fab-rics in low-temperature mineral assemblages are theyfossil biomarkers Implications for the search for a sub-surface fossil record on the early Earth and Mars PlanetSpace Sci 48 1077ndash1086
Jakosky BM and Shock EL (1998) The biological po-tential of Mars the early Earth and Europa J GeophysRes 103 19359ndash19364
Johnston CG and Vestal JR (1993) Biogeochemistry ofoxalate in Antarctic cryptoendolithic lichen-dominatedcommunity Microb Ecol 25 305ndash319
Kakegawa T Kasahara Y Hayashi K Ohmoto H(2000) Sulfur and carbon isotope analyses of the 27 GaJeerinah Formation Fortescue Group AustraliaGeochem J 34 121ndash133
King RA Dittmer CK and Miller JDA (1976) Effectof ferrous iron concentration on the corrosion of ironin semi-batch cultures of sulphate-reducing bacteriaBr Corros J 11 105
Kirkland BL Lynch FL Rahnis MA Folk RL Mo-lineux IJ and McLean RJC (1999) Alternative ori-gins for nannobacteria-like objects in calcite Geology 27347ndash350
MINERALOGICAL BIOSIGNATURES 463
Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

Kirshvink JL and Vali H (1999) Criteria for the identi-fication of bacterial magnetofossils on Earth or Mars30th Lunar and Planetary Science Conference Ab-stracts Available at wwwlpiusraedumeetingsLPSC99pdf1681pdf
Kneale D and Viles HA (2000) Beach cement incipientCaCO3-cemented beachrock development in the upperintertidal zone North Uist Scotland Sediment Geol132 165ndash170
Labrenz M Druschel GK Thomsen-Ebert T GilbertB Welch SA Kemner K Logan GA Summons RDe Stasio G Bond PL Lai B Kelley SD and Ban-field JF (2000) Natural formation of sphalerite (ZnS)by sulfate-reducing bacteria Science 290 1744ndash1747
Larsen KW Arvidson RE Joliff BL and Clark BC(2000) Correspondence and least squares analyses ofsoil and rock compositions for the Viking Lander 1 andPathfinder sites 31st Lunar and Planetary Science Con-ference Abstracts Available at wwwlpiusraedumeetingsLPSC2000pdf
Lauretta DS Lodders K and Fegley B Jr (1997) Ex-perimental simulations of sulfide formation in the So-lar Nebula Science 277 358ndash360
Little B Wagner P Ray R Pope R and Scheetz R(1991) Biofilms artifacts introduced during SEM prepa-ration evaluated by ESEM J Industrial Microbiol 8213ndash222
Little CTS Herrington RJ Haymon RM andDanelian T (1999) Early Jurassic hydrothermal ventcommunity from the Franciscan Complex San RafaelMountains California Geology 27 167ndash170
Malin M C and Edgett KS (2000) Sedimentary rocks ofearly Mars Science 290 1927ndash1939
MacLachlan MJ Manners I and Ozin GA (2000) New(inter)faces polymers and inorganic materials AdvMaterials 12 675
Mandernack KW Post J and Tebo BM (1995) Man-ganese mineral formation by bacterial spores of the ma-rine bacillus strain SG-1mdashevidence for the direct oxi-dation of Mn (II) to Mn (IV) Geochim Cosmochim Acta59 4393ndash4408
Mann S Archibald DD Didymus JM Douglas THeywood BR Meldrum FC and Reeves NJ (1993)Crystallization at inorganic-organic interfacesmdashbio-minerals and biomimetic synthesis Science 261 1286ndash1292
McGoldrick P (1999) Northern Australian ldquoSedexrdquo Zn-Pb deposits microbial oases in Proterozoic seas In Min-eral Deposits Processes to Processing (Proceedings of theFifth Biennial SGA Meeting and the Tenth QuadrennialIAGOD Symposium) Vol 2 edited by CJ Stanley AHRankin RJ Bodnar J Naden BWD Yardley AJCriddle RD Hagni AP Gize J Paseva AJ Fleet RSeltmann C Halls M Stemprok B Williamson RJHerrington RET Hill HM Prichard F Wall CTWilliams I McDonald JJ Wilkinson D Cooke NJCook BJ Marshall P Spry K Zhaw L Meinart KSundblad PW Scott SHB Clark E Valsami-JonesNJ Beukes HJ Stein JL Hannah F Neubauer DJ
Blundell DHM Alderton MP Smith S Mulshawand RA Ixer AA Balkema Rotterdam pp 885ndash888
McGuire MM Edwards KJ Banfield JF and HamersRJ (2001) Kinetics surface chemistry and structuralevolution of microbially mediated sulfide mineral dis-solution Geochem Cosmochim Acta 65 57ndash72
McNeil MB and Little B (1990) Mackinawite formationduring microbial corrosion J Corros 46 599ndash600
McSween HY Jr and Treiman AH (1998) Martian me-teorites in planetary materials Rev Mineral 36 61ndash653
Morris RV Golden DC Bell JF III Tad SDScheinost AC Hinman NW Furniss G MertzmanSA Bishop JL Ming DW Allen CC and BrittDT (2000) Mineralogy composition and alteration ofMars Pathfinder rocks and soils evidence from multi-spectral elemental and magnetic data on terrestrialanalogue SNC meteorite and Pathfinder samples JGeophys Res 105 1757ndash1817
Mortimer RJG Coleman ML and Rae JE (1997) Ef-fect of bacteria on the elemental composition of earlydiagenetic siderite implications for palaeoenviron-mental interpretations Sedimentology 44 759ndash765
Moskowitz BM Frankel RB Bazylinski DA Jan-nasch HW and Lovley DR (1989) A comparison ofmagnetite particles produced anaerobically by magne-totactic and dissimilatory iron-reducing bacteria Geo-phys Res Lett 16 665ndash668
Mustard JF Cooper CD and Rifkin MK (2001) Evi-dence for recent climate change on Mars from the iden-tification of youthful near-surface ground ice Nature412 411ndash414
Nelson DC Casey WH Sison JD Mack EE AhmadA and Pollack JS (1996) Selenium uptake by sulfur-accumulating bacteria Geochim Cosmochim Acta 603531ndash3539
Nugent JHA Rich AM and Evans MCW (2001)Photosynthetic water oxidation towards a mechanismBiochim Biophys Acta 1503 138ndash146
Ohmoto H and Lasaga AC (1982) Kinetics of reactionsbetween aqueous sulfates and sulfides in hydrothermalsystems Geochim Cosmochim Acta 46 1727ndash1745
Ohmoto H Kakegawa T and Lowe D (1993) 34 Bil-lion-year-old biogenic pyrites from Barberton SouthAfrica sulfur isotope evidence Science 262 555ndash557
Pokrovski GS and Schott J (1998) Experimental studyof the complexation of silicon and germanium withaqueous organic species implications for germaniumand silicon transport and GeSi ratio in natural watersGeochim Cosmochim Acta 62 3413ndash3428
Posfai M Buseck P Bazylinski D and Frankel R(1998) Reaction sequence of iron sulfide minerals in bac-teria and their use as biomarkers Science 280 880ndash883
Renault RW and Jones B (1997) Controls on aragoniteand calcite precipitation in hot spring travertines atChemurkeu Lake Bogoria Kenya Can J Earth Sci 34801ndash818
Richard DT (1969) The microbiological formation of ironsulphies Stockholm Contrib Geol 20 49ndash66
BANFIELD ET AL464
Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats

Russell NC Edwards HGM and Wynn-WilliamsDD (1998) FT-Raman spectroscopic analysis of en-dolithic microbial communities from Beacon sandstonein Victoria Land Antarctica Antarctic Sci 10 63ndash74
Santelli CM Welch SA Westrich HR and BanfieldJF (2001) The effect of Fe-oxidizing bacteria on Fe-sil-icate mineral dissolution rates Chem Geol 180 99ndash115
Schultze-Lam S Fortin D Davis BS and BeveridgeTJ (1996) Mineralization of bacterial surfaces ChemGeol 132 171ndash181
Stillings LL Drever JI Brantley SL Sun Y andOxburgh R (1996) Rates of feldspar dissolution at pH3ndash7 with 0ndash8 mM oxalic acid Chem Geol 132 79ndash89
Straub KL Benz M and Schink B (2001) Iron metab-olism in anoxic environments at near neutral pH FEMSMicrobiol Ecol 34 181ndash186
Suzuki Y and Banfield JF (2001) Microbial formationof uranium-bearing minerals In Proceedings of the ClayMinerals Society Annual Meeting Clay Minerals SocietyMadison WI p 36
Taunton AE Welch SA and Banfield JF (2000a) Geo-microbiological controls on light rare earth element Yand Ba distributions during granite weathering and soilformation J Alloys Compounds 303 30ndash36
Taunton AE Welch SA and Banfield JF (2000b) Mi-crobial controls on phosphate and lanthanide distribu-tions during granite weathering Chem Geol 169 371ndash382
Teng HH and Dove PM (1997) Surface site-specific in-teractions of aspartate with calcite during dissolution im-plications for biomineralization Am Mineral 82 878ndash887
Thomas-Keprta KL Bazylinski DA Kirschvink JLClemett SJ McKay DS Wentworth SJ Vali HGibson EK and Romanek CS (2000) Elongated pris-matic magnetite crystals in ALH84001 carbonate glob-ules potential Martian magnetofossils Geochim Cos-mochim Acta 64 4049ndash4081
Thomas-Keprta KL Clemett SJ Bazylinski DAKirschvink JL McKay DS Wentworth SJ Vali HGibson EK McKay MF and Romanek CS (2001)Truncated hexa-octahedral magnetite crystals inALH84001 presumptive biosignatures Proc Natl AcadSci USA 98 2164ndash2169
Thorseth IH Torsvik T Furnes H and MuehlenbachsK (1995) Microbes play an important role in the alter-ation of oceanic crust Chem Geol 126 137ndash146
Vasconcelos C McKenzie JA Bernasconi S Grujic Dand Tien AJ (1995) Microbial mediation as a possiblemechanism for natural dolomite formation at low-tem-peratures Nature 377 220ndash222
Warthmann R van Lith Y Vasconcelos C McKenzieJA and Karpoff AM (2000) Bacterially induceddolomite precipitation in anoxic culture experimentsGeology 28 1091ndash1094
Weed R and Norton SA (1991) Siliceous crusts quartzrinds and biotic weathering of sandstones in the colddesert of Antarctica In Developments in GeochemistryVol 6 Diversity of Environmental Biogeochemistry editedby J Berthelin Elsevier Amsterdam pp 327ndash339
Welch SA and Ullman WJ (1993) The effect of organicacids on plagioclase dissolution rates and stoichiome-try Geochim Cosmochim Acta 57 2725ndash2736
Welch SA Taunton AE Santelli CM and BanfieldJF (1999) Role of microorganisms in the weathering ofgranite and formation of soil [abstract] In Proceedingsof the American Society for Microbiology Annual MeetingAmerican Society for Microbiology Washington DC
Welch SA Nesterova M Chan CS Skatvold AMand Banfield JF (2001) Biologically induced formationof novel nanocrystalline iron oxyhydroxides 11th An-nual VM Goldschmidt Conference Available atwwwlpiusraedumeetingsgold2001pdf3901pdf
Welch SA Taunton AE and Banfield JF (2002) Effectof microorganisms and microbial metabolites on apatitedissolution Geomicrobiol J (in press)
Widdel F Schnell S Heising S Ehrenreich A Ass-mus B and Schink B (1993) Ferrous iron oxidation byanoxygenic phototrophic bacteria Nature 362 834ndash836
Address reprint requests toDr Jillian A Banfield
Departments of Earth and Planetary Sciences andEnvironmental Science Policy and Management
369 McCone HallUniversity of California Berkeley
Berkeley CA 94720-4767
E-mail jillseismoberkeleyedu
MINERALOGICAL BIOSIGNATURES 465
View publication statsView publication stats


















