
Mimicking the plant-cell interior under water stress by ... · Jean-Marie Mouillon1#, Sylvia K...
42
Mimicking the plant-cell interior under water stress by macromolecular crowding: disordered dehydrin proteins are highly resistant to structural collapse. Corresponding author: Pia Harryson Department of Biochemistry and Biophysics Arrhenius Laboratories for Natural Sciences Stockholm University S-106 91 Stockholm, Sweden Tel: + 46 8 164238 Fax: + 46 8 153679 [email protected] Research area: Biochemistry and/or Environmental stress and adaptation 1 Plant Physiology Preview. Published on October 10, 2008, as DOI:10.1104/pp.108.124099 Copyright 2008 by the American Society of Plant Biologists www.plantphysiol.org on July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.
Transcript of Mimicking the plant-cell interior under water stress by ... · Jean-Marie Mouillon1#, Sylvia K...

Mimicking the plant-cell interior under water stress by
macromolecular crowding: disordered dehydrin proteins are highly
resistant to structural collapse.
Corresponding author:
Pia Harryson
Department of Biochemistry and Biophysics
Arrhenius Laboratories for Natural Sciences
Stockholm University
S-106 91 Stockholm, Sweden
Tel: + 46 8 164238 Fax: + 46 8 153679
Research area:
Biochemistry and/or Environmental stress and adaptation
1
Plant Physiology Preview. Published on October 10, 2008, as DOI:10.1104/pp.108.124099
Copyright 2008 by the American Society of Plant Biologists
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Jean-Marie Mouillon1#, Sylvia K Eriksson* and Pia Harryson*
1 Umeå Plant Science Centre
Department of Plant Physiology
Umeå University
S-901 87 UMEÅ
SWEDEN
# Present address:
Fluxome Sciences A/S
Diplomvej 378
DK-2800 Lyngby
Denmark
* Department of Biochemistry and Biophysics
Arrhenius Laboratories for Natural Sciences
Stockholm University
S-106 91 Stockholm, Sweden
Tel: + 46 8 164238 Fax: + 46 8 153679
2 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Financial source:
This project has been funded by the Swedish Research Council for the Environment,
Agricultural Sciences and Spatial Planning (P.H.) and the Carl Tryggers Foundation (P.H.)
and the Lawski Foundation (S.E.).
Corresponding author:
[email protected] Fax: + 46 8 153679
3 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

ABSTRACT
The dehydrins are a class of drought-induced proteins in plants that lack a fixed three-
dimensional structure. Their specific molecular action, as well as the reason for their
disordered character, is as yet poorly understood. It has been speculated, however, that the
dehydrins are tuned to acquire a biologically active structure only under the conditions at
which they normally function, i.e. upon dehydration. To test this hypothesis, we here
investigate the effect of reduced water content and macromolecular crowding on three
dehydrins from Arabidopsis thaliana. As a simplistic model for mimicking cellular
dehydration we used polyethylene glycol (PEG), glycerol, and sugars which plants naturally
employ as compatible solutes, i.e. sucrose and glucose. Macromolecular crowding was
induced by the large polysaccharides ficoll and dextran. The results show that the dehydrins
are remarkably stable in their disordered state and are only modestly affected by the solvent
alterations. A notable exception is the dehydrin Cor47 which shows a small, intrinsic increase
in helical structure at high concentrations of osmolytes. We also examined the effect of
phosphorylation but found no evidence that such post-translational modifications of the
dehydrin sequences modulate their structural response to osmolytes and crowding agents.
The results suggest that the dehydrins are highly specialised proteins that have evolved to
maintain their disordered character under conditions where unfolded states of several globular
proteins would tend to collapse.
Key words: dehydrin, intrinsically disordered protein, macromolecular crowding, circular
dichroism (CD)
4 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

INTRODUCTION
The loss of water from plant cells during drought and freezing triggers a series of adaptation
processes. One of these is the production of osmolytes such as sucrose, proline and betaine
(Hasegawa et al., 2000; Wang et al., 2003). As a consequence, the already crowded interior
of the cell gets even denser and the physiological processes need to adjust to elevated
backgrounds of non-reacting macromolecules and steric confinement. In some cases the
increase in macromolecular density can be radical, going from 300–400 g/L under ideal
growth conditions to >900 g/L upon severe desiccation (Ellis, 2001; Bryant et al., 2005).
Such cellular crowding does not only present a challenge in terms of modulating diffusion-
based chemical reactions (Ellis, 2001; Minton, 2005; 2006), but will also affect structural
integrity. Crowding promotes protein assembly by favouring compact conformations over
extended ones, and the reduction in bulk water will directly influence membrane topology and
dynamics. The molecular conditions in water-starved plant cells are therefore very different
from those normally used to characterise biochemical processes in vitro. It has been put
forward that in such situations the biological function of proteins could be regulated through
crowding-induced conformational changes (Hall, 2006; Hall and Dobson, 2006). Under these
extreme conditions the plant cells respond by expressing a characteristic class of stress
proteins, the dehydrins (Close, 1997; Garay-Arroyo et al., 2000) (Fig. 1).
The function of dehydrins is as yet unknown. A complicating factor, from a biochemical
perspective, is that they seem to lack ordered structure (Lisse et al., 1996; Soulages et al.,
2003; Mouillon et al., 2006). Even so, all dehydrins contain, by definition, at least one copy
of a highly conserved sequence segment, the K-seg, and may or may not include other
conserved sequences called the S-seg, the Y-seg (Close, 1996) and the ChP-1 (Mouillon et al.,
2006). Since the dehydrins accumulate at very high concentrations inside cells (Bartels and
Salamini, 2001) they are unlikely to be signal molecules or conventional enzymes. The role
of the dehydrins is probably of a more general nature. For example, they may stabilise
membranes, act as chaperones or by other means buffer the altered solvent properties inside
water-stressed cells (Close, 1996; Garay-Arroyo et al., 2000; Boudet et al., 2006). As a clue
to how the stress response is orchestrated molecularly, it has been suggested that the
functional structure of the dehydrins is induced by their hydration status (Boudet et al., 2006).
Accordingly, the disordered appearance of the dehydrins may be converted into active, three-
dimensional structures provided that the conditions inside a drought-stressed plant cell are
sufficiently well reproduced. The disordered conformations seen under dilute conditions
5 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

would then represent the denatured state and, consequently, tell very little about the
dehydrin’s actual biological function. This idea is supported by the observations that water
can modulate ionisation potentials, pKA values and protein-binding potentials (Kornblatt and
Kornblatt, 2002). Other classes of disordered proteins have been found to participate in
molecular recognition, regulation and cell signalling (Dunker et al., 2002; Uversky et al.,
2005). Function is, then, induced by folding of the disordered protein into a structured state
where the functional groups are placed at the right locations for interactions. Folding can be
initiated (or templated) by binding to a biological partner such as other proteins, DNA, RNA,
or metals (Dunker et al., 2002; Tompa and Csermely, 2004).
Of particular interest with respect to the dehydrins is that the conformational changes are
sometimes conditional, i.e. the protein structure responds to changes in the environment such
as temperature (McNulty et al., 2006), pH (Tornroth-Horsefield et al., 2006), availability of
water (Luo and Baldwin, 1997) or macromolecular crowding (Minton, 2005a; 2005b; Hall,
2006). Disorder is used to allow the function to be switched on and off, e.g. a 10% change in
the concentration of intracellular proteins can lead to changes up to a factor 10 in the activity
of molecular regulatory species (Al-Habori, 2001). Computational studies have recently
demonstrated that the effect of crowding, despite its underlying complexity, can be simply
mimicked by encapsulation of the proteins in spherical pores (Klimov et al., 2002; Cheung
and Thirumalai, 2007). This freedom in how the crowding pressure can be applied suggests
that simplified experiments where crowding effects are tested by addition of large branched
polymers, such as ficoll and dextran, could produce valid approximations of the in vivo
condition. In fact, it has recently been verified by nuclear magnetic resonance (NMR) that the
effect of such crowding agents in vitro reproduces the intracellular conditions fairly well
(Selenko et al., 2006).
In this study, we examine whether the addition of macromolecular crowding agents in vitro
can force the dehydrins to fold. The progressive substitution of water with various polymers
is performed as a simplistic model of how the cellular interior changes upon
drought/desiccation. As representative crowding agents we chose a series of natural and
synthetic osmolytes, the effects of which are well characterised in other systems. To
distinguish the response of the full-length dehydrins from that of their constituent K- and
ChP-1 segments, the latter have been analysed separately in the form of peptides. The results
show that both the full-length dehydrins, and the isolated K- and ChP-1 segments, retain their
6 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

disordered character also at extremely high levels of osmolytes. Moreover, the propensity of
the dehydrins to undergo unspecific collapse, c.f. amorphous glass, appears even lower than
that of denatured states of globular proteins. Taken together, these observations suggest that
the dehydrin sequence is highly evolved and adapted to remain disordered also under
conditions of severe dehydration. In this respect, the dehydrins are different from the class of
disordered proteins that rely on folding to become functional. The function of the dehydrins
is therefore likely to lie in the interactions of the conserved segments with their specific
biological targets.
7 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

RESULTS
Analysis of structural transitions, using circular dichroism. Circular dichroism is a
common spectroscopic method for detecting secondary structure in proteins, where α-helices,
β-sheets and random coils display characteristic spectra at wavelengths 190–250 nm (Fig. 2).
Accordingly, the CD spectrum of a typical folded protein is a characteristic structural
fingerprint made up by the sum of its constituent secondary-structure elements. In
physiological buffer the CD spectra of the dehydrins, however, resemble those of unfolded
global proteins (Mouillon et al., 2006) (Figs. 2 and 3). This signal differs somewhat from that
of the pure random coil since it contains contributions from dynamic, residual secondary-
structure and polyproline helices II (Mouillon et al., 2006). In this study we used this coil-
like signal as a reference to test whether a more ordered structure is induced upon addition of
solutes that mimic the solvent conditions experienced inside a living plant cell. For example,
the induction of helical structure in the dehydrins upon addition of trifluoroethylene (TFE)
(Mouillon et al., 2006) is seen as a characteristic decrease in the CD signal around 222 nm in
combination with an increase at around 200 nm (cf. Fig. 2).
The natural stress solutes have little impact on the dehydrin structure. Under conditions
of stress, plants are known to produce a variety of compatible solutes such as betaine (Chen
and Murata, 2002), proline (Yoshiba et al., 1997) and different sugars (Ingram and Bartels,
1996; Hoekstra et al., 2001). Of particular interest for this study are sucrose and glucose
which are naturally produced in response to low temperature and drought (Hoekstra et al.,
2001). To investigate whether increased levels of these sugars contribute to modulating the
conformational properties of the dehydrins, we undertook titration experiments (followed by
CD) in which the sucrose and glucose content was varied between 0% and 80%.
Interestingly, the results suggest that sucrose and glucose have only a marginal effect on the
disordered character of Cor47, Lti29 and Lti30 (Fig. 3). Even at very high concentrations of
sugar, the CD signal at 222 nm remains largely the same as in physiological buffer, apart from
a small but significant decrease in ellipticity at between 210 nm and 230 nm for Cor47. At
shorter wavelengths the characteristic minimum of the random-coil signal tends to be
progressively weakened. Under the extreme solvent conditions used here, however, these
spectral changes should be considered relatively modest. The increased CD signal at 200 nm
possibly reflects a small increase in helical structure. Such increased helical propensity is
expected upon weakening of the hydrogen bonding between water and the protein backbone
8 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

at high concentrations of solute. Even so, the strength of this signal is too small to imply that
the observed effect has any bearing on the physiological function of the dehydrin proteins.
The effect of glycerol follows the intrinsic α-helical propensities of the polypeptide
chains. In comparison with the small sugars sucrose and glucose, the structural effect of
adding glycerol is much more pronounced. Above 50% glycerol, the CD spectra of all
proteins in this study indicate a significant increase in the occupancy of α-helical structure
(Fig. 3). Moreover, this effect seems to be larger for Cor47 than for Lti29 and Lti30. The
result is in good accord with the intrinsic helical propensities of the three proteins. The
relative response of the different dehydrins to additions of glycerol follows precisely that of
the helical inducer TFE (Mouillon et al., 2006). In other words, the magnitude of the glycerol
response can be correlated to the protein’s intrinsic α-helical propensity, i.e. Cor47 > Lti30 =
Lti29. To illustrate this correspondence more clearly, in Figure 4 we compare the effect of
adding 50% glycerol with that of adding 50% of the strong helical inducer TFE. Although the
magnitude of the glycerol response is in general much smaller than that of the TFE response,
the features of the difference spectra appear very much the same. Analysis of the different
CD spectra using the structural prediction program Dichroweb (Whitmore and Wallace, 2004)
shows that Cor47 is 15% α-helical in 50% glycerol while for corresponding value of Lti29
and Lti30, is 7.5%. Upon comparison with other proteins, it is apparent that this low α-helical
propensity matches the background levels of most polypeptide sequences and supports the
notion that the extent of residual α-helical structure observed in this study is unlikely to
constitute the key to the molecular action of the dehydrins in vivo.
The reason why the α-helical response is higher with glycerol than with sucrose and glucose
could simply be that the former is a stronger helical inducer (Table 1). Even so, we observed
that Cor47, which is most sensitive to glycerol, displays a CD change that deviates from an
ideal two-state transition into α-helical structure. Upon titration with glycerol the CD
spectrum of Cor47 does not display a single iso-elliptic point, as is typical for TFE titrations,
but acquires the α-helical signal in two steps, i.e. A ⇔ B ⇔ C (Fig. 5). First, the CD
difference spectra indicate a mixed α-helical structure at between 0% and 75% glycerol.
Then, the spectrum of Cor47 undergoes a second transition to a more characteristic α-helical
signal at >80% glycerol. Apparently, Cor47 first responds to glycerol by adopting low levels
of a mixed α -helical structure. Finally, at very high glycerol concentrations, this mixed
9 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

structure is converted to more purely α-helical structures (Fig. 5). These α-helical structures
could first form locally and then extend to cover a larger part of the sequence. Analysis using
Dichroweb (Whitmore and Wallace, 2004) shows 22% helical structures at 75% glycerol and
48% helical structure at 95% glycerol. Thus, by quantitative comparison, the results suggest
that the degree of α-helical structure induced by 95% glycerol is similar to that in 25% TFE
(Mouillon et al., 2006). It should also be noted that at 25% TFE the level of α-helical
structure of Cor47, as analysed using CDPro has levelled out at a maximum value of 47%
(Mouillon et al., 2006). Therefore, Cor47 is likely to acquire a significant content of local
structure in 95% glycerol.
The corresponding signal for Lti29 and Lti30 is much smaller. This indicates that the
sensitivity to glycerol is a specific feature of the Cor47 sequence and not a common,
functional property of the dehydrin proteins as a group.
Structural collapse in the presence of polyethylene glycol: comparison with the unfolded
states of globular proteins. The tendency of the polypeptide chain to collapse into compact
states at high concentrations of osmolytes such as glycerol is not unique to Cor47 but has
generally been observed for small peptides (Venkatesu et al., 2007) and denatured states of
globular proteins (Fig. 2) (Foord and Leatherbarrow, 1998; Silow and Oliveberg, 2003). If
the pressure to minimise the surface area is sufficiently high, any unfolded polypeptide is
expected to collapse into compact forms that need not always resemble the native structures
(Bryngelson and Thirumalai, 1996). The ability to resist such unspecific coil collapse under
physiological conditions is a key feature that distinguishes natural proteins from randomly
assembled polypeptides (Bryngelson and Thirumalai, 1996). This property renders the
denatured states of natural proteins flexible and facilitates their folding into unique native
structures. In an attempt to benchmark the collapse propensity of the dehydrin proteins along
such a general scale, we have monitored the effect of adding PEG, following the conditions
used in protein folding studies. Again, the structural response was highest for Cor47,
followed by Lti30 (Fig. 6). Least affected was Lti29. As the effect of PEG 4,000 and PEG
6,000 resembles that of glycerol, the mode of action of these osmolytes is most likely similar.
Analysis of the different PEG spectra using Dichroweb reveals that the induced structure is a
mix of β-sheets and α-helices (14% helical in 50% PEG 6,000). It is interesting to note,
however, that the susceptibility of the dehydrins to change their structure in the presence of
osmolytes generally seems lower than for denatured states of small globular proteins (c.f.
10 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Figs. 2 and 6). Although the globular proteins typically adopt their folded states in highly
concerted two-state transitions, i.e. without populating intermediates, their denatured states
have been observed to become trapped in compact, sterically restricted conformations already
at 30% PEG or ethylene glycol (EG) (Qu et al., 1998; Silow and Oliveberg, 2003). In the
case of the archetypical two-state protein CI2, the acid-denatured state shows striking changes
in the CD spectrum at 70% PEG (Fig. 2) (Silow and Oliveberg, 2003). Comparable effects
are only seen with Cor47, while the impact on the CD spectra of Lti29 and Lti30 is less
pronounced. Consistently, the dehydrins also show very limited response to sulphate, which
is another well-known misfolding and aggregation agent for globular proteins (Otzen and
Oliveberg, 1999). Therefore, when it comes to resisting unspecific collapse, it is apparent that
the dehydrins are equally, if not more, robust than the unfolded states of globular proteins.
The response of the dehydrins to crowding agents ficoll and dextran. The preferred way
to examine the effect of macromolecular crowding would be to subject the dehydrins to high
concentrations of other proteins, RNA and colloidal particles. In vivo, the total concentration
of proteins and RNA is in excess of 300 mg/mL (Zimmerman and Trach, 1991). An intrinsic
problem with such experiments, however, is that the spectroscopic handle of the dehydrin
structure, in this case the CD signal, would disappear against the much larger background of
the co-solutes. As an alternative, we used the large polysaccharides ficoll (70 kDa) and
dextran (40 kDa and 70 kDa). Interestingly, the presence of these crowding agents showed no
appreciable effect on the dehydrin proteins. At 50% ficoll or dextran the CD spectra of Lti29
and Lti30 were virtually identical to those in pure buffer while the spectrum of Cor47 showed
an increase in the 200 nm region similar to that in glucose and sucrose (Fig. 6). Although the
changes in the lower wavelengths of the Cor47 spectrum are difficult to rationalise
structurally (Mouillon et al., 2006), the results on the whole underline the dehydrins’
resistance to altering their disordered character in response to co-solute exposure. Therefore,
the disordered character of these proteins could be a key requirement for their function and
this property seems to be maintained in the crowded interior in the dehydrated plant cell.
Combinations of sugars and osmolytes. Although tests of the pure, individual osmolytes
and crowding agents are required for experimental stringency, these experiments need not
represent the complex conditions inside stressed cells where a multitude of co-solutes act in
concert. To examine whether there are any synergistic effects of mixing sugars, osmolytes
and crowding agents, we also screened the response of the dehydrins to a series of mixtures.
11 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

As a base, we used 25% glycerol, and added to this an additional 25% of the other
compounds. Notably, these systematic mixtures did not contribute to enhancing or changing
the effect on the dehydrin CD spectra: in all cases the observed effect was the combination of
the signals from the isolated compounds. To further rule out the possibility of a delayed
response to the solvent perturbations, the samples were re-run after 24 hours but without any
detectable difference in the CD spectra (data not shown). These findings are in line with the
recent conclusions by Holthauzen and Bolen (2007).
The conserved segments behave approximately like the full-length proteins. Even though
the response of the full-length proteins to solvent additives is generally small, it is possible
that this response stems from a strong structural effect in a small part of the sequence. The
prime candidates for such local structure formation would then be the sequence regions
containing the conserved segments. These are the K-segment (K-seg:
EKKGIMDKIKEKLPG) and the charged-peptide 1 (ChP-1: EEGEDGEKKKKEKKKKI), in
other words, the charged part following the serine-rich segment found in Cor47 and Lti29, i.e.
in the SKn type of dehydrins (c.f. Fig. 1). The function of these characteristic and repeatedly
occurring sequence stretches is still unknown. In partial contrast to the full-length dehydrins,
peptides of the K-seg and ChP-1 show the most pronounced structural response to PEG 6,000
and not to glycerol (Fig. 7). At 50% PEG 6,000, structural prediction analysis using
Dichroweb (Whitmore and Wallace, 2004) suggests an α-helical content of 9% for K-seg and
11% for ChP-1. By comparison, the effect of PEG 4,000 is substantially smaller than that of
PEG 6,000 and similar to the structural response induced by glycerol.
The different effects of PEG 6,000 and PEG 4,000 are interesting since the PEGs are
substantially larger than the peptide fragments and the full-length dehydrins and would,
accordingly, be expected to affect these in an equivalent way. Ficoll and dextran are also
considerably larger than either peptide but give as low a response as with the full-length
dehydrins. Upon titration with glycerol, the K-segment and ChP-1 both show an
approximately linear decrease of the CD signal at 222 nm. Therefore, K-seg and ChP-1
behave very much like the full-length proteins Lti29 and Lti30 and lack the characteristic late
transition of Cor47 at 95% glycerol (data not shown).
Phosphorylation has only a marginal effect on the dehydrin structures. Recombinantly
expressed dehydrins are not phosphorylated when purified from E. coli (data not shown).
12 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

However, dehydrins are known to undergo phosphorylation both in vivo and in vitro (Jiang
and Wang, 2004; Alsheikh et al., 2005; Brini et al., 2007; Rohrig et al., 2006). To investigate
the structural impact of phosphorylation, we treated the dehydrins with casein kinase II. Both
Cor47 and Lti29, but not Lti30, became phosphorylated. Such selective phosphorylation of
the different dehydrins is in good agreement with previous reports (Alsheikh et al., 2005).
The results show that phosphorylation of Lti29 has no detectable effect on the CD spectra
under any of the solvent conditions tested in this study (data not shown). In the presence of
50% glycerol, however, phosphorylation of Cor47 produces a small decrease in the coil
content, indicated by the CD spectrum below 210 nm (Fig. 8). Notably, this signal cannot be
seen in the presence of glucose or PEG 4,000, suggesting that the structural relevance is after
all very small (data not shown). The effect of phosphorylation on the dehydrin structure is
therefore very small and does not significantly enhance the response to osmolytes or crowding
agents in vitro.
13 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

DISCUSSION
Crowding has no observable role in modulating the dehydrin structures. How the
molecular function of proteins and membranes is maintained in organisms during
environmental stress is a poorly understood area of biology that relates to the detailed
interplay between solvent and solute molecules. The latter include supra-structures of lipids,
proteins and carbohydrates. In plants, drought can reduce the water content to only a fraction
of the normal levels, often accompanied by compensating production of osmolytes and
changes in protein composition. The benefit of such stress-induced adjustments of the intra-
cellular conditions is to safeguard biological viability. At a molecular level, however, not
only is it challenging to reproduce this complex response in vitro but the response is also
challenging to rationalise mechanistically: the interior of cells can as yet not be treated
exactly as in vivo and simplifications are needed. In the present study of the dehydrin
proteins, one such simplification is to parameterise the experimental conditions in terms of
macromolecular crowding or excluded volume effects. As the water content of the cell is
decreased, the background of other molecules is enriched, leading to a geometric restriction of
the available space for polypeptide chains. Typically, proteins function under conditions
where the solute concentration is around 400 g/L (Zimmerman and Minton, 1993) and in
severely dehydrated plant cells this number can be substantially higher (Bartels and Salamini,
2001). Such increased background of crowding molecules, i.e. the osmolytes and all other
solvent components taken together, is expected to have an important impact on the cellular
processes (Ellis, 2001a; 2001b). By simply competing for volume, macromolecular crowding
favours compact states of the polypeptide chain and increases the stability of native proteins,
provided that no competing misfolded conformations are produced. Accompanying this
general stability effect, crowding increases also the occupancy of compact denatured species,
which, eventually, leads to a retardation of the protein folding rates due to increased
reconfiguration times. As the normally flexible and dynamic polypeptide chain gets
entangled in compact conformations its internal movements become energetically more
cumbersome (Bryngelson and Thirumalai, 1996; Oliveberg and Wolynes, 2005). It has been
put forward that such collapse/expansion of certain inert biopolymers could actually be used
as a means to regulate function of proteins in biological environments via crowding (Hall,
2006; Hall and Dobson, 2006).
14 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

The effect of macromolecular crowding on the protein structure is therefore expected to
resemble that of small stabilising agents such as sodium sulphate, Na2SO4. However, the
efficiency of macromolecular crowding in promoting protein stability is relatively modest and
is substantially smaller than that of Na2SO4. The equilibrium constant for RNAse T1 was
only shifted 7.5-fold towards the native state in 400 mg/mL dextran 70 (Qu and Bolen, 2002),
while the corresponding effect of Na2SO4 on the protein L23 is several times larger (Hedberg
and Oliveberg, 2004). Consistent with the earlier observation that Na2SO4 has no appreciable
impact on the dehydrin structures (Mouillon et al., 2006), we could not discern any
compelling effect upon crowding by the large branched sugars ficoll and dextran (Fig. 6).
The only outcome is a small change in the low wavelength region of the CD spectra. This
change could possibly reflect a relaxation of PII helices towards more flexible coil
conformations. By comparison, induction of a classic secondary structure, such as α-helices
or β-sheets, would lead to a reduction in the 200 nm signal with a simultaneous increase in the
minimum around 220 nm (c.f. Fig. 2). However, it is necessary to consider also the size of
the tested crowding agent in relation to that of the dehydrins. At a first approximation, a
small crowding agent is expected to get closer to the protein and, hence, occupy volume more
efficiently compared with a large crowding agent. Even so, the dehydrin structures are
equally indifferent to the smaller sugars glucose and sucrose (Figs. 3 and 4).
We could not distinguish any correspondence between changes in CD spectra and the size of
any of the added solutes in this study. The disordered nature of the dehydrin polypeptide
seems to persist also when the available space is geometrically restricted. A plausible
explanation for this result is that the dehydrins do not easily adapt compact conformations,
either in the form of uniquely defined folded states, as has been observed for several other
disordered proteins (Dawson et al., 2003; Love et al., 2004; Fink, 2005), or in the form of
unspecifically collapsed conformations that are sometimes observed in the denatured states of
globular proteins (Lattman et al., 1994; Otzen and Oliveberg, 2002; Ittah et al., 2004). The
behaviour is in good agreement with earlier reports on the disordered c-Fos and p27Kip1
proteins, where crowding with differently sized dextrans (9 kDa, 37.5 kDa or 77 kDa) and
ficoll 70 at concentrations of up to 250 g/L was not sufficient to induce ordered structure
(Flaugh and Lumb, 2001). The dehydrins resemble also the Parkinson-related α-synuclein
that remains disordered in the crowded interior of E. coli cells and at high concentrations of
BSA (McNulty et al., 2006). In some contrast, the transcription factor regulator FlgM partly
15 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

gains structure under the corresponding conditions (Dedmon et al., 2002), and FlgM also
gains structure upon binding to its biological target (Daughdrill et al., 1997). These
differences have been taken to exemplify two classes of disordered proteins: those that are
functionally tuned to remain disordered in the crowded interior of the cell (Flaugh and Lumb,
2001), and those that are not. According to the data in Figures 3–8, the dehydrins belong to
the former category. Taken together, the effect of macromolecular crowding on disordered
protein conformations therefore seems to be variable and needs to be tested in each separate
case. A brief summary of the crowding effects on other disordered proteins and denatured
globular proteins is given in Table 1. Another effect of water loss is increased ionic strength.
We have earlier demonstrated, however, that increased ionic strength or elevated levels of
metal ions has no appreciable impact on the dehydrin structure in vitro (Mouillon et al.,
2006).
Preferential protein-solute exclusion increases the sampling of the local α-helical
structure. The conformational transitions underlying the altered CD signals of Cor47 in the
presence of glycerol and PEGs are therefore likely to have a molecular origin different from
pure crowding. From a functional perspective it is reasonable to examine whether these
molecular factors are somehow coupled to changes in the accessibility of water. For example,
they may be manifested in altered structure of the protein hydration shells. However, such
water-mediated effects are intrinsically difficult to mechanistically rationalise because of the
complexity of aqueous solutions. A simplified view is offered by weak interaction models.
These models assume that the solute molecules, typically osmolytes, are either accumulated
or excluded from the protein-solvent interface. In the event that the osmolyte is preferentially
attracted to the protein surface, this will favour extended conformations and promote
destabilisation according to mass action (Schellman, 2002). Examples of such destabilising
osmolytes are the denaturants urea and guanidium chloride (GdmCl). Conversely, if the
osmolyte is preferentially excluded from the protein surface, this will lead to minimisation of
the protein-solvent interface area and protein stabilisation (Bolen and Baskakov, 2001). Such
stabilising, or protective, osmolytes are sugars, free amino acids and glycerol. The reason for
the “affinity differences” between the osmolytes seems partly related to the degree by which
they can form favourable polar interactions with the polypeptide backbone (Street et al.,
2006). Computational (Street et al., 2006) and experimental (Qu and Bolen, 2002) analysis
suggests that the order of stabilising capacity of the osmolytes used in this study is sucrose >
sorbitol > proline > glycerol. From this perspective, it is interesting to note that the osmolyte
16 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

that most efficiently perturbed the CD spectra of the dehydrin Cor47 is glycerol (Fig. 5). The
effect of 80% sucrose, on the other hand, is still relatively modest. An explanation for this
reversed order could lie in the somewhat unusual amino-acid composition of the dehydrin
sequences.
The computational ranking of the osmolyte effect is obtained from backbone interactions
alone (Street et al., 2006), which is in good accord with the stability effect on RNAse T1 (Qu
and Bolen, 2002) with a hydrophobic side-chain content typical of globular proteins (cf. pdf
structure 1bu4). In contrast, the dehydrin sequences contain a relatively high proportion of
charged and polar side chains, presenting a higher degree of hydrogen bonding opportunities
with the surrounding solution molecules. This peculiarity could also explain why α-
synuclein, in contrast to the dehydrins, collapses upon addition of glucose (Morar et al.,
2001): α-synuclein contains approximately 40% hydrophobic residues while the dehydrins
contain just 15% (Lti30) to 30% (Cor47 and Lti29).
Even so, and regardless of the mechanistic origin of the accentuated glycerol effect of the
dehydrins, it is apparent from the CD data that the structural impact of adding osmolytes is
only small at physiologically relevant concentrations (Figs. 3 and 5). The same is true for the
fragments of their conserved segments (Fig. 7). The glycerol effect is not clearly manifested
unless the glycerol content is increased above 30%, a level that is likely to greatly exceed the
physiological concentrations. By comparison, the highest reported level of sucrose in vivo is
40% of the dry weight and this in a resurrection plant that more or less tolerates complete
desiccation (Ingram and Bartels, 1996). Moreover, the structural transition observed at
extreme concentrations of glycerol corresponds mainly to an increased sampling of the α-
helical structure, which is in good accordance with the intrinsic α-helical propensity of the
different dehydrin proteins (Fig. 1). The largest effect was observed for Cor47, followed by
Lti29 and Lti30, which precisely matched the gain of the α-helical structure in TFE titrations
(Mouillon et al., 2006). Consistently, a similar structural response was observed upon
addition of PEG 4,000 and PEG 6,000. These polymers are well-known protein stabilisers
(Kornblatt and Kornblatt, 2002; Kozer and Schreiber, 2004; Ren et al., 2006) that have also
been observed to unspecifically collapse the denatured states of globular proteins (Silow and
Oliveberg, 2003). On this basis, we conclude that the exertion of pressure to minimise the
solvent-accessible surface area of the dehydrin chains has no appreciable effects on their
17 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

tertiary structures. They generally remain disordered. The effect on the local α-helical
content merely follows the intrinsic α-helical propensity of the polypeptide chains. The latter,
however, is more likely to reflect the generic properties of the polypeptide chain than specific
function traits.
Possible interactions between dehydrins and lipid membranes. A frequently proposed
function of dehydrins and other LEA proteins is as membrane stabilisers. In essence, the
dehydrins would interact with membranes to modulate their topology and phase transitions
under stress, and in this process the dehydrins could also gain ordered structure. The idea is
supported by several findings of interactions between lipid vesicles and dehydrins and other
LEA proteins (Soulages et al., 2002; Koag et al., 2003; Soulages et al., 2003; Tolleter et al.,
2007; Kovacs et al., 2008). In some cases, as with the maize dehydrin DHN1, the membrane
interaction leads to conformational changes in the protein’s secondary structure, i.e. an
increase in the α –helical content (Koag et al., 2003). However, it is not clear to what extent
these effects can be transferred to other members of the protein family as, in contrast, the
interaction between Lti29 and negative charged lipids did not result in any conformational
changes (Kovacs et al., 2008). Also, the presence of lipids fail to induce any structural effect
on the soybean LEA protein GmD-19 and dehydrin GmDHN1 (Soulages et al., 2002;
Soulages et al., 2003). Although the involvement of the dehydrins in safeguarding membrane
structure during stress is indeed an attractive possibility, its generality remains to be
established.
Phosphorylation does not generate any significant structural changes of dehydrins.
Phosphorylation is a post-translational modification that is essential for regulation of cellular
functions. It was recently shown that most phosphorylation sites, 86%, are within disordered
regions of proteins (Collins et al., 2008). Even so, there are few studies on the effect of
phosphorylation on the structure of disordered proteins. Of particular interest for this study is
the fact that phosphorylation of the dehydrins occurs in vivo and has been reported to
modulate the coordination of calcium ions (Jiang and Wang, 2004; Alsheikh et al., 2005;
Brini et al., 2007; Rohrig et al., 2006). In this study only Cor47 and Lti29 could be
phosphorylated by casein kinase II. The result of this phosphorylation, however, had no
significant bearing on the structural behaviour of Cor47 and Lti29. Recent NMR studies of α-
synuclein have shown that phosphorylation at Ser 129 by casein kinase II increases the
18 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

disordered character of the protein. It was further shown that this structural alteration
increased the proteins’ tendency to self-associate (Sasakawa et al., 2007). As seen in Figure
8, the structure of Cor47 is left unaltered upon phosphorylation and also shows no tendency to
aggregate. Crowding by glycerol makes no difference. Even though Cor47 and Lti29 seem to
be structurally unchanged, it is important to note that phosphorylation can still play a key role
as regulatory factor for the interaction with other cellular targets.
Conclusions. From a physiological standpoint, the separate tests of crowding agents and
osmolytes constitute a fairly crude representation of the intracellular conditions where spatial
confinement and the presence of complex mixtures of background molecules act in concert on
the dehydrin structures. However, we failed to observe any synergistic structural effects in
mixtures of glycerol and the other solutes used in this study, lending further support to the
idea that the dehydrin structures are generally insensitive to changing solvent conditions.
Based on previous results, the structural insensitivity includes also changes of ionic strength
and levels of metal ions (Mouillon et al., 2006). This suggests that the dehydrin sequences
are evolved to remain flexible also under severe stress conditions, and that this property is an
integral requirement of their physiological function. Possibly, the high content of charged
and polar residues of the dehydrins effectively prevents structural collapse due to the
accompanying desolvation penalties of ionic moieties and hydrogen-bond partners. Such
transitions into compact glass-like species are otherwise characteristic of random polymers
(Bryngelson and Thirumalai, 1996) and to some extent also of denatured states of globular
proteins (Silow and Oliveberg, 2003). In both instances, the affected sequences generally
contain higher fractions of hydrophobic side chains than observed for the dehydrins.
Additional indications that the dehydrins are tuned to remain flexible under in vivo conditions
come from their high solubility and resistance to precipitation (Mouillon et al., 2006). As a
contrasting example, α-synuclein readily assembles in ordered fibrillar aggregates at elevated
protein concentrations or upon addition of PEG/osmolytes (Goers et al., 2003; Fink, 2006). In
discussions about the biological function of the dehydrins, it has been pointed out that their
unfolded nature would effectively maximise the coordination of water. According to NMR
experiments, this water is more tightly bound than in other disordered proteins (Bokor et al.,
2005). Some of the water seems tied up in bridging interactions within PII helices (Mouillon
et al., 2006). Upon removal of the coordinated water by evaporation, other proteins in the
LEA family have been observed to undergo structural rearrangements (Wolkers et al., 2001;
19 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Goyal et al., 2003; Goyal et al., 2005). For example, the LEA protein AavLea1 increases its
content of α-helical structure (Goyal et al., 2003). Could then the dehydrins act as water
reservoirs that expel coordinated water to the surrounding medium (Boudet et al., 2006)?
Considering the finding that the in vivo concentration of dehydrins does not seem to exceed
2–4% of cytoplasmic proteins, however, the benefit of such water expulsion is likely to be
marginal (Roberts et al., 1993; Ceccardi et al., 1994; Close, 1996), at least in moderate
drought. More likely, the molecular action of the dehydrins is more specific. One possibility
is that the dehydrins provide structural support, either spatially by cross-linking with other
macromolecular constituents (Abu-Abied et al., 2006) or laterally by adhering to membrane
surfaces (Tolleter et al., 2007). Desiccation of the cell will not only alter the molecular
composition of the aqueous compartments of the cell but will also distort its physical
dimensions, including the shape and proximity of membrane structures.
It is easy to envisage that such global alterations could have a pronounced effect on the
colloidal processes, for example by promoting phase transitions, topological frustration and
altered dynamics of membrane fusion and budding. The role of the dehydrins may be to
intervene with these processes by filling out, and perhaps topologically organising, crevices in
bulging membranes and thereby to safeguard their structural integrity. Alternatively, the
dehydrins could support macromolecular structures of proteins, carbohydrates or nucleotides.
In any case, the elements of recognition with the cellular target are likely to be the conserved
segments. The remarkable capacity of the dehydrins to remain coil-like suggests that the high
flexibility of the sequence regions connecting these segments is an important part of their
function (Fig. 1): the conserved segments are organised as “hooks on a string” (Mouillon et
al., 2006). Notably, this arrangement not only restricts the spatial location of the conserved
elements but also increases their local concentration.
20 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

MATERIALS AND METHODS
Expression and heat fractionation. Expression, purification and identification of the
recombinant Arabidopsis thaliana dehydrin proteins were as described previously (Svensson
et al., 2000). Glycerol stocks of the different Escherichia coli strains were made and 200 μL
was spread on Luria agar (LA) plates with 150 μg ampicillin. The plates were kept at 37°C
and grown overnight. The cells were suspended and added to 1 litre (L) of LB medium
containing 50 μg/mL ampicillin. Expression was induced at an optical density (OD) of 0.3 by
adding isopropyl ß-D-thiogalactopyranoside (IPTG) to a final concentration of 1 mM. The
suspensions were kept at 37°C until grown to an OD of 0.8–1.0 (i.e. 2–4 hours). Cells were
harvested by centrifugation at 4,000 g for 45 min and the pellet stored at –20°C. The thawed
cells from 1 L cultures were resuspended in 25 mL of 20 mM Na2HPO4, pH 7.2, 150 mM
NaCl. To this was added 1 mM (final concentration) phenylmethylsulfonyl fluoride (PMSF)
and 0.1 mg/mL lysosyme and the suspension was left for 30 min on ice. Lysated cells were
sonicated 6 x 15 s and centrifuged at 9,000 g for 30 min. The supernatants were placed in a
water bath at 70°C for 20 min. To precipitate heat-denatured proteins the sample was
centrifuged for 30 min at 9,000 g and the supernatant stored at –80°C.
Purification of dehydrins. Purification of dehydrins on immobilised metal ion affinity
chromatography (IMAC) and the following step on ion exchange chromatography was
according to Svensson et al. (2000). The supernatants from heat precipitation were diluted
1:2 with 20 mM NaHPO4, pH 7.2, 1.85 M NaCl and 1 mM PMSF. The samples were loaded
on a 5 mL HiTrap IDA-Sepharose column (Pharmacia) connected to a FPLC system
(Pharmacia) and charged with 2 mL of 3 mg/mL CuSO4. Absorbance was read at 280 nm.
Before applying the sample, five volumes of 20 mM Na2HPO4, pH 7.2, and 1.0 M NaCl were
used to equilibrate the column. The same buffer (40 volumes) was used to wash off unbound
sample from the column. Fractions of 5 mL were collected for analysis during the whole run.
Elution was performed with 2 M NH4Cl in 20 mM Na2HPO4, pH 7.2, 1.0 M NaCl, in one
step. The column was then equilibrated with ten volumes of 20 mM Na2HPO4, pH 7.2,
followed by elution of the Cu with 10mM EDTA in 20 mM Na2HPO4, pH 7.2. Precipitation
of protein was done with 80% (NH4)2SO4 and proteins were collected by centrifugation at
18,000 g for 30 min. The different dehydrins were resuspended in 2 mL of the following
21 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

buffers: Lti29 and Cor47 in 20 mM Bistris, pH 6.0, Rab18 in 20 mM Tris-HCl, pH 8.0, and
Lti30 in 50 mM glycine, pH 9.0. The dehydrins were desalted on a PD-10 column
(Pharmacia) by the resuspension buffers. The achieved 3 mL fractions were put on an anionic
exchange column (1-mL Mono Q HR 5/5) (Pharmacia) in the case of Cor47 and Lti29, and on
a cationic exchange column (1-ml Mono S HR 5/5, Pharmacia) connected to an FPLC in the
case of Lti30, and absorbance was read at 280 nm. The columns were equilibrated with the
resuspension buffers and elution by a NaCl gradient from 0 to 0.5 M in respective buffer over
30 volumes. Fractions of 0.5 mL were collected during the runs for analysis. The purity was
tested by SDS-PAGE gel electrophoresis, and without exception the purified material was run
as untreated dehydrins from crude cell extracts (data not shown). An extensive identification
of the recombinant dehydrins has previously been published by Svensson et al. (2000).
Analysis by circular dichroism. Circular dichroism (CD) measurements were carried out
using a JASCO J-810 spectropolarimeter (JASCO, Japan) and presented as mean ellipticity
per residue. The scan rate was 20 nm/min at a band pass of 0.2 nm, and 20 mdeg sensitivity.
All samples were mixed 1 hour prior to the CD analysis and centrifuged at 12,000 g for 2 min
before filling the cuvettes. Protein concentration was 1 mg/mL in 20 mM MES (0.2 mm
cuvette). Unless stated otherwise, all runs were performed at 25°C.
Preparation of samples. Unless otherwise stated, concentrations are given as % referring to
g/L. Dextrans (average molecular weight of 40 kDa or 70 kDa) (Sigma), ficolls (average
molecular weight of 70 kDa) (Sigma) and polyethylene glycol (PEG) (average molecular
weights of 4,000 or 6,000 Da) (Sigma) were used in concentrations of up to 500–800 g/L. All
osmolytes, sugars and crowding agents were titrated in steps of 10–25%. Each titration series
was repeated at least twice. The exception was Lti30 where the titration steps were 15–25%
and titration ended at 50% due to low amount of high-purity Lti30 protein.
Phosphorylation of dehydrins. The dehydrins were phosphorylated by 5.4 units/mL of
casein kinase II (CKII) (Sigma) in 1 mM ATP, 20 mM Tris-HCl, 50 mM KCl and 10 mM
MgCl2 at pH 7.5 for 10 hours. The phosphorylation status of the dehydrins was tested by Pro-
Q Diamond Phosphoprotein Gel stain (Invitrogen) on 15% Tris-Hcl gels (Biorad) and the
background levels of protein were measured by SYPRO Ruby gel stain (Invitrogen). Stained
gels were visualised by an FLA-3000G (Fuji Photo Film Co. Ltd) and an ultraviolet (UV)
transilluminator. The phosphorylation status was measured on three different batches of
22 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

dehydrins, with identical results. For the structural analysis, the CKII was separated from the
dehydrins by precipitation at 70°C, followed by centrifugation. The effect of solvent
additives on the phosphorylated dehydrins was measured as described above.
23 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

LEGENDS TO FIGURES
Figure 1. Distribution of the conserved segments in the sequences of the dehydrins Cor47,
Lti29 and Lti30 from Arabidopsis. The dehydrins constitute group 2 of the large family of the
hydrophilic LEA proteins (Close, 1996; Bartels and Salamini, 2001).
The amino acid sequences are as follows:
COR47: MAEEYKNNVPEHETPTVATEESPATTTEVTDRGLFDFLGKKEEEVKPQETTTLESEFDH
KAGISEPELAAEHEEVKENKITLLEELGEKTEEDEENKPSVIEKLHRSNSSSSSSSDEEGE
EKKEKKKKIVEGEEDKKGLVEKIKEKLPGHHDKTAEDDVPVSTTIPVPVSESVVEHDHP
E IKEKLPGYHAKTTEEEVKKEKESDD
LTI29: MAEEYKNTYPEGETPKVATEESSAPEIKERGMFDFLKKKEEVKPQETTTLASEFEH
KTGISEPESFVAKHEEEEHKPTLLEGLHQKHEEEEENKPSLLDKLHRSNSSSSSSSDEE
GEDGEKPEEEEKKGFMDKIKEKLPGHSKKPEDSGVVNTTPLVETATPIADIPEEKKGF
MDKIKEKLPGYHAKTTGEEEKKEKVSD
LTI30: MNSHQNQTGVQKKGITEKIMEKLPGHHGPTNTGVVHHEKKGMTEKVMEQLPGHHGA
TGTGGVHHEKKGMTEKVMEQLPGHHGSHQTGTNTTYGTTNTGGVHHEKKSVTEKVM
EKLPGHHGSHQTGTNTAYGTNTNVVHHEKKGIAEKIKEGLPGHHGTHKTGTTTSYGNT
GVVHHENKST
Figure 2. Reference circular dichroism (CD) spectra of the common types of secondary
structure and random coil conformations. A. Pure α-helical structure (dotted line), pure β-
sheet structure (striped line) and random coil (solid line). B. The acid-denatured state of the
globular protein CI2 in pure buffer and in 70% of EG and PEG. Notably the structural
induction of CI2 by EG and PEG is not along the normal folding pathway but seems to
involve non-native interactions (Silow and Oliveberg, 2003).
Figure 3. Circular dichroism (CD) signal of Cor47, Lti29 in the presence of 80% sucrose,
glucose and glycerol, and Lti30 in the presence of 50% sucrose and glycerol, illustrating a
difference in response to the different sugars.
Figure 4. (Top panel): Difference spectra of Cor47, Lti29 and Lti30 in 50% glycerol.
(Bottom panel): Difference spectra of Cor47, Lti29 and Lti30 in 50% of the strong helical
inducer TFE. The results indicate that the effects of both glycerol and TFE reflect the
intrinsic helical propensity of the dehydrin sequences.
24 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Figure 5. (Top panel): Circular dichroism (CD) spectra of the glycerol titration of Cor47.
The spectra show from top at 220 nm, 0%, 10%, 25%, 50%, 60%, 75% and 95% glycerol.
(Middle panel): Circular dichroism difference spectra showing that the structural induction of
Cor47 with glycerol occurs in two steps. Between 0% and 75%, glycerol Cor47 displays a
mix of β-sheet and α-helices while above 75% it displays mainly α-helices. (Bottom panel):
Changes in the CD signal at 200 nm and 222 nm upon titration of Cor47 with glycerol,
illustrating the transitions to mixed and pure α-helical structures, respectively.
Figure 6. Crowding of Cor47, Lti29 and Lti30 with polymers. The concentration of
polymers is 50%.
Figure 7. Circular dichroism (CD) difference spectra of the conserved segments of the
dehydrins, showing the effect of adding 50% polymers. (Top panel): The K-segment
(EKKGIMDKIKEKLPG) is present in all dehydrins. (Bottom panel): The ChP-1 segment
(EEGEDGEKKKKEKKKKI) is present in Cor47 and Lti29.
Figure 8. Circular dichroism (CD) spectra of phosphorylated Cor47 in pure buffer and in
50% glycerol.
25 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Acknowledgements
We are most grateful to Professor Mikael Oliveberg, Stockholm University for the data
regarding CI2 and for helpful discussions.
26 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Table 1. Effects of various crowding agents on intrinsically disordered proteins and denatured
globular proteins.
27 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Co-solute Effect on disordered proteins
Effect on denatured globular proteins
Effect on other systems
Dextrans 1. No effects on p27KIP1 or c-FOS 3. 2. Folding of TCAM RNAse 4.
1. Increased refolding (kf) of lysosyme 1. 2. Change to molten globular state of cyt C 2.
1. Decreased unfolding (ku) of GAPDH 5.
Ficolls 1. No effects on p27KIP1 or c-FOS 3. 2. No effects on TCAM RNAse stability (KD-N) 4.
1. Minor effect on FKB12 stability 6. 2. Minor effect on refolding (kf) of lysosymes 7.
EG/PEGs 1. Accelerated fibrillation of α- synuclein 10.
1. Collapse of the coil state (decreased kf) of CI2 at high PEG 8. 2. Increased folding rate (kf) of CI2 at low PEG 8. 3. Decreased folding rate (kf) of CspB (EG) 9.
1. Decreased unfolding (ku) of GAPDH 5.
Glycerol 1. Accelerated fibrillation of α- synuclein 10.
1. Stabilised native state of RNAseA 11.
2. Increased refolding(kf) of GS with GroEL12. 3. Increased refolding (kf) of MHD 13.
Glucose 2. Induced collapse of α-synuclein 14.
1. Folding of Fe cyt C 14.
1. Stabilised dimeric chymotrypsin 15.
Sucrose 1. Contracted RCAM RNAse 17.
1. Increased refolding (kf) of GS 12. 2. Increased refolding (kf) of CI2 16. 3. Increased refolding (kf) of MHD 13.
2. Stabilised dimeric chymotrypsin 15.
28 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

1. Zhou BR, Liang Y, Du F, Zhou Z, & Chen J. (2004) J Biol Chem 279, 55109–16. 2. Sasahara K, McPhie P, & Minton, AP. (2003) J Mol Biol 326, 1227–37. 3. Flaugh SL, & Lumb KJ. (2001) Biomacromolecules 2, 538–40. 4. Qu Y, & Bolen DW. (2002) Biophys Chem 101–102, 155–65. 5. Ren G, Lin Z, Tsou CL, & Wang CC. (2003) J Protein Chem 22, 431–9. 6. Spencer DS, Xu K, Logan TM, & Zhou HX. (2005) J Mol Biol 351, 219–32. 7. Van den Berg B, Wain R, Dobson CM, & Ellis RJ. (2000) Embo J 19, 3870–5. 8. Silow M, & Oliveberg M. (2003) J Mol Biol 326, 263–71. 9. Jacob M, Geeves M, Holtermann G, & Schmid FX. (1999) Nat Struct Biol 6, 923–6. 10. Munishkina LA, Cooper EM, Uversky VN, & Fink AL. (2004) J Mol Recognit 17,
456–64. 11. Gekko K, & Ito H. (1990) J Biochem (Tokyo) 107, 572–7. 12. Voziyan PA, & Fisher MT. (2002) Arch Biochem Biophys 397, 293–7. 13. Tieman BC, Johnston MF, & Fisher MT. (2001) J Biol Chem 276, 44541–50. 14. Morar AS, Olteanu A, Young GB, & Pielak GJ. (2001) Protein Sci 10, 2195–9. 15. Patel CN, Noble SM, Weatherly GT, Tripathy A, Winzor DJ, & Pielak GJ. (2002)
Protein Sci 11, 997–1003. 16. Ladurner AG, & Fersht AR. (1999) Nat Struct Biol 6, 28–31. 17. Qu Y, Bolen CL, & Bolen DW. (1998) Proc Natl Acad Sci U S A 95, 9268–73.
29 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

References
Abu-Abied M, Golomb L, Belausov E, Huang S, Geiger B, Kam Z, Staiger CJ, Sadot E (2006) Identification of plant cytoskeleton-interacting proteins by screening for actin stress fiber association in mammalian fibroblasts. Plant J 48: 367-379
Al-Habori M (2001) Macromolecular crowding and its role as intracellular signalling of cell volume regulation. Int J Biochem Cell Biol 33: 844-864
Alsheikh MK, Svensson JT, Randall SK (2005) Phosphorylation regulated ion-binding is a property shared by the acidic subclass dehydrins. Plant, Cell and Environment 28: 1114-1122
Bartels D, Salamini F (2001) Desiccation tolerance in the resurrection plant Craterostigma plantagineum. A contribution to the study of drought tolerance at the molecular level. Plant Physiol 127: 1346-1353
Bokor M, Csizmok V, Kovacs D, Banki P, Friedrich P, Tompa P, Tompa K (2005) NMR relaxation studies on the hydrate layer of intrinsically unstructured proteins. Biophys J 88: 2030-2037
Bolen DW, Baskakov IV (2001) The osmophobic effect: natural selection of a thermodynamic force in protein folding. J Mol Biol 310: 955-963
Boudet J, Buitink J, Hoekstra FA, Rogniaux H, Larre C, Satour P, Leprince O (2006) Comparative analysis of the heat stable proteome of radicles of Medicago truncatula seeds during germination identifies late embryogenesis abundant proteins associated with desiccation tolerance. Plant Physiol: 1481-1436
Brini F, Hanin M, Lumbreras V, Irar S, Pages M, Masmoudi K (2006) Functional Charaterization of DHN-%, a dehydrin showing a differential phopshorylation pattern in two Tunisian durum wheat (Triticum durum Desf.) varieties with marked difference in salt and drought tolerance. Plant Science 172: 20-28
Bryant JE, Lecomte JT, Lee AL, Young GB, Pielak GJ (2005) Protein dynamics in living cells. Biochemistry 44: 9275-9279
Bryngelson JD, Thirumalai D (1996) Internal constraints induce localization in an isolated polymer molecule. Phys Rev Lett 76: 542-545
Ceccardi TL, Meyer NC, Close TJ (1994) Purification of a maize dehydrin. Protein Expr Purif 5: 266-269
Chen TH, Murata N (2002) Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr Opin Plant Biol 5: 250-257
Cheung MS, Thirumalai D (2007) Effects of crowding and confinement on the structures of the transition state ensemble in proteins. J Phys Chem B 111: 8250-8257
Close TJ (1996) Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiol Plant 97: 795-803
Close TJ (1997) Dehydrins: a commonality in the response of plants to dehydration and low temperature. Physiol Plant 100: 291-296
Collins MO, Yu L, Campuzano I, Grant SG, Choudhary JS (2008) Phosphoproteomic analysis of the mouse brain cytosol reveals a predominance of protein phosphorylation in regions of intrinsic sequence disorder. Mol Cell Proteomics In press
Daughdrill GW, Chadsey MS, Karlinsey JE, Hughes KT, Dahlquist FW (1997) The C-terminal half of the anti-sigma factor, FlgM, becomes structured when bound to its target, sigma 28. Nat Struct Biol 4: 285-291
Dawson R, Muller L, Dehner A, Klein C, Kessler H, Buchner J (2003) The N-terminal domain of p53 is natively unfolded. J Mol Biol 332: 1131-1141
Dedmon MM, Patel CN, Young GB, Pielak GJ (2002) FlgM gains structure in living cells. Proc Natl Acad Sci U S A 99: 12681-12684
30 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Dunker AK, Brown CJ, Lawson JD, Iakoucheva LM, Obradovic Z (2002) Intrinsic disorder and protein function. Biochemistry 41: 6573-6582
Ellis RJ (2001a) Macromolecular crowding: an important but neglected aspect of the intracellular environment. Curr Opin Struct Biol 11: 114-119
Ellis RJ (2001b) Macromolecular crowding: obvious but underappreciated. Trends Biochem Sci 26: 597-604
Fink AL (2005) Natively unfolded proteins. Curr Opin Struct Biol 15: 35-41 Fink AL (2006) The aggregation and fibrillation of alpha-synuclein. Acc Chem Res 39: 628-
634 Flaugh SL, Lumb KJ (2001) Effects of macromolecular crowding on the intrinsically
disordered proteins c-Fos and p27(Kip1). Biomacromolecules 2: 538-540 Foord RL, Leatherbarrow RJ (1998) Effect of osmolytes on the exchange rates of
backbone amide protons in proteins. Biochemistry 37: 2969-2978 Garay-Arroyo A, Colmenero-Flores JM, Garciarrubio A, Covarrubias AA (2000) Highly
hydrophilic proteins in prokaryotes and eukaryotes are common during conditions of water deficit. J Biol Chem 275: 5668-5674
Goers J, Uversky VN, Fink AL (2003) Polycation-induced oligomerization and accelerated fibrillation of human alpha-synuclein in vitro. Protein Sci 12: 702-707
Goyal K, Tisi L, Basran A, Browne J, Burnell A, Zurdo J, Tunnacliffe A.(2003) Transition from natively unfolded to folded state induced by desiccation in an anhydrobiotic nematode protein. J Biol Chem. 278(15):12977-84
Goyal K, Pinelli C, Maslen SL, Rastogi RK, Stephens E, Tunnacliffe A (2005) Dehydration-regulated processing of late embryogenesis abundant protein in a desiccation-tolerant nematode. FEBS Lett 579: 4093-4098
Hall D (2006) Protein self-association in the cell: a mechanism for fine tuning the level of macromolecular crowding? Eur Biophys J 35: 276-280
Hall D, Dobson CM (2006) Expanding to fill the gap: A possible role for inert biopolymers in regulating the extent of the 'macromolecular crowding' effect. FEBS Lett 580: 2584-2590
Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ (2000) Plant Cellular and Molecular Responses to High Salinity. Annu Rev Plant Physiol Plant Mol Biol 51: 463-499
Hedberg L, Oliveberg M (2004) Scattered Hammond plots reveal second level of site-specific information in protein folding: phi' (beta++). Proc Natl Acad Sci U S A 101: 7606-7611
Hoekstra FA, Golovina EA, Buitink J (2001) Mechanisms of plant desiccation tolerance. Trends Plant Sci 6: 431-438
Hoekstra FA, Golovina EA, Tetteroo FA, Wolkers WF (2001) Induction of desiccation tolerance in plant somatic embryos: how exclusive is the protective role of sugars? Cryobiology 43: 140-150
Holthauzen LM, Bolen DW (2007) Mixed osmolytes: the degree to which one osmolyte affects the protein stabilizing ability of another. Protein Sci 16: 293-298
Ingram J, Bartels D (1996) The Molecular Basis of Dehydration Tolerance in Plants. Annu Rev Plant Physiol Plant Mol Biol 47: 377-403
Ittah V, Kahana E, Amir D, Haas E (2004) Applications of time-resolved resonance energy transfer measurements in studies of the molecular crowding effect. J Mol Recognit 17: 448-455
Jiang X, Wang Y (2004) Beta-elimination coupled with tandem mass spectrometry for the identification of in vivo and in vitro phosphorylation sites in maize dehydrin DHN1 protein. Biochemistry 43: 15567-15576
31 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Klimov DK, Newfield D, Thirumalai D (2002) Simulations of beta-hairpin folding confined to spherical pores using distributed computing. Proc Natl Acad Sci U S A 99: 8019-8024
Koag MC, Fenton RD, Wilkens S, Close TJ (2003) The binding of maize DHN1 to lipid vesicles. Gain of structure and lipid specificity. Plant Physiol 131: 309-316
Kornblatt JA, Kornblatt MJ (2002) The effects of osmotic and hydrostatic pressures on macromolecular systems. Biochim Biophys Acta 1595: 30-47
Kovacs D, Kalmar E, Torok Z, Tompa P (2008) Chaperone activity of ERD10 and ERD14, two disordered stress-related plant proteins. Plant Physiol 147: 381-390
Kozer N, Schreiber G (2004) Effect of crowding on protein-protein association rates: fundamental differences between low and high mass crowding agents. J Mol Biol 336: 763-774
Lattman EE, Fiebig KM, Dill KA (1994) Modeling compact denatured states of proteins. Biochemistry 33: 6158-6166
Lisse T, Bartels D, Kalbitzer HR, Jaenicke R (1996) The recombinant dehydrin-like desiccation stress protein from the resurrection plant Craterostigma plantagineum displays no defined three-dimensional structure in its native state. Biol Chem 377: 555-561
Love JJ, Li X, Chung J, Dyson HJ, Wright PE (2004) The LEF-1 high-mobility group domain undergoes a disorder-to-order transition upon formation of a complex with cognate DNA. Biochemistry 43: 8725-8734
Luo P, Baldwin RL (1997) Mechanism of helix induction by trifluoroethanol: a framework for extrapolating the helix-forming properties of peptides from trifluoroethanol/water mixtures back to water. Biochemistry 36: 8413-8421
McNulty BC, Tripathy A, Young GB, Charlton LM, Orans J, Pielak GJ (2006) Temperature-induced reversible conformational change in the first 100 residues of alpha-synuclein. Protein Sci 15: 602-608
McNulty BC, Young GB, Pielak GJ (2006) Macromolecular crowding in the Escherichia coli periplasm maintains alpha-synuclein disorder. J Mol Biol 355: 893-897
Minton AP (2005a) Influence of macromolecular crowding upon the stability and state of association of proteins: predictions and observations. J Pharm Sci 94: 1668-1675
Minton AP (2005b) Models for excluded volume interaction between an unfolded protein and rigid macromolecular cosolutes: macromolecular crowding and protein stability revisited. Biophys J 88: 971-985
Minton AP (2006) Macromolecular crowding. Curr Biol 16: R269-271 Morar AS, Olteanu A, Young GB, Pielak GJ (2001) Solvent-induced collapse of alpha-
synuclein and acid-denatured cytochrome c. Protein Sci 10: 2195-2199 Mouillon JM, Gustafsson P, Harryson P (2006) Structural investigation of disordered stress proteins: comparison of full-length Dehydrins with isolated peptides of their conserved segments. Plant Physiol 141: 638-650 Oliveberg M, Wolynes PG (2005) The experimental survey of protein-folding energy
landscapes. Q Rev Biophys 38: 245-288 Otzen DE, Oliveberg M (1999) Salt-induced detour through compact regions of the protein
folding landscape. Proc Natl Acad Sci U S A 96: 11746-11751 Otzen DE, Oliveberg M (2002) Conformational plasticity in folding of the split beta-alpha-
beta protein S6: evidence for burst-phase disruption of the native state. J Mol Biol 317: 613-627
Qu Y, Bolen CL, Bolen DW (1998) Osmolyte-driven contraction of a random coil protein. Proc Natl Acad Sci U S A 95: 9268-9273
32 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Qu Y, Bolen DW (2002) Efficacy of macromolecular crowding in forcing proteins to fold. Biophys Chem 101-102: 155-165
Ren X, Yang Z, Kuang T (2006) Solvent-induced changes in photochemical activity and conformation of photosystem I particles by glycerol. Biol Chem 387: 23-29
Roberts JK, DeSimone NA, Lingle WL, Dure L, 3rd (1993) Cellular Concentrations and Uniformity of Cell-Type Accumulation of Two Lea Proteins in Cotton Embryos. Plant Cell 5: 769-780
Rohrig H, Schmidt J, Colby T, Brautigam A, Hufnagel P, Bartels D (2006) Desiccation of the resurrection plant Craterostigma plantagineum induces dynamic changes in protein phosphorylation. Plant Cell Environ 29: 1606-1617
Sasakawa H, Sakata E, Yamaguchi Y, Masuda M, Mori T, Kurimoto E, Iguchi T, Hisanaga S, Iwatsubo T, Hasegawa M, Kato K (2007) Ultra-high field NMR studies of antibody binding and site-specific phosphorylation of alpha-synuclein. Biochem Biophys Res Commun 363: 795-799
Schellman JA (2002) Fifty years of solvent denaturation. Biophys Chem 96: 91-101 Selenko P, Serber Z, Gadea B, Ruderman J, Wagner G (2006) Quantitative NMR analysis
of the protein G B1 domain in Xenopus laevis egg extracts and intact oocytes. Proc Natl Acad Sci U S A 103: 11904-11909
Silow M, Oliveberg M (2003) High concentrations of viscogens decrease the protein folding rate constant by prematurely collapsing the coil. J Mol Biol 326: 263-271
Soulages JL, Kim K, Arrese EL, Walters C, Cushman JC (2003) Conformation of a group 2 late embryogenesis abundant protein from soybean. Evidence of poly (L-proline)-type II structure. Plant Physiol 131: 963-975
Soulages JL, Kim K, Walters C, Cushman JC (2002) Temperature-induced extended helix/random coil transitions in a group 1 late embryogenesis-abundant protein from soybean. Plant Physiol 128: 822-832
Street TO, Bolen DW, Rose GD (2006) A molecular mechanism for osmolyte-induced protein stability. Proc Natl Acad Sci U S A 103: 13997-14002
Svensson J, Palva ET, Welin B (2000) Purification of recombinant Arabidopsis thaliana dehydrins by metal ion affinity chromatography. Protein Expr Purif 20: 169-178
Tolleter D, Jaquinod M, Mangavel C, Passirani C, Saulnier P, Manon S, Teyssier E, Payet N, Avelange-Macherel MH, Macherel D (2007) Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell 19: 1580-1589
Tompa P, Csermely P (2004) The role of structural disorder in the function of RNA and protein chaperones. Faseb J 18: 1169-1175
Tornroth-Horsefield S, Wang Y, Hedfalk K, Johanson U, Karlsson M, Tajkhorshid E, Neutze R, Kjellbom P (2006) Structural mechanism of plant aquaporin gating. Nature 439: 688-694
Uversky VN, Oldfield CJ, Dunker AK (2005) Showing your ID: intrinsic disorder as an ID for recognition, regulation and cell signaling. J Mol Recognit 18: 343-384
Venkatesu P, Lee MJ, Lin HM (2007) Thermodynamic characterization of the osmolyte effect on protein stability and the effect of GdnHCl on the protein denatured state. J Phys Chem B 111: 9045-9056
Wang W, Vinocur B, Altman A (2003) Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta 218: 1-14
Whitmore L, Wallace BA (2004) DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res 32: W668-673
33 www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

34
Wolkers WF, McCready S, Brandt WF, Lindsey GG, Hoekstra FA (2001) Isolation and characterization of a D-7 LEA protein from pollen that stabilizes glasses in vitro. Biochim Biophys Acta 1544: 196-206
Yoshiba Y, Kiyosue T, Nakashima K, Yamaguchi-Shinozaki K, Shinozaki K (1997) Regulation of levels of proline as an osmolyte in plants under water stress. Plant Cell Physiol 38: 1095-1102
Zimmerman SB, Minton AP (1993) Macromolecular crowding: biochemical, biophysical, and physiological consequences. Annu Rev Biophys Biomol Struct 22: 27-65
Zimmerman SB, Trach SO (1991) Estimation of macromolecule concentrations and excluded volume effects for the cytoplasm of Escherichia coli. J Mol Biol 222: 599-620
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

CHP-1K-seg S-seg
Cor47 SK3 (265 aa)Lti29 SK3 (260 aa)Lti30 K6 (193 aa)
Figure 1.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

230 250210
wavelength (nm)
acid-denatured CI2
+70% EG
Figure 2.
A. B.
+70% PEG
random coil
α-helix
β-sheet
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.
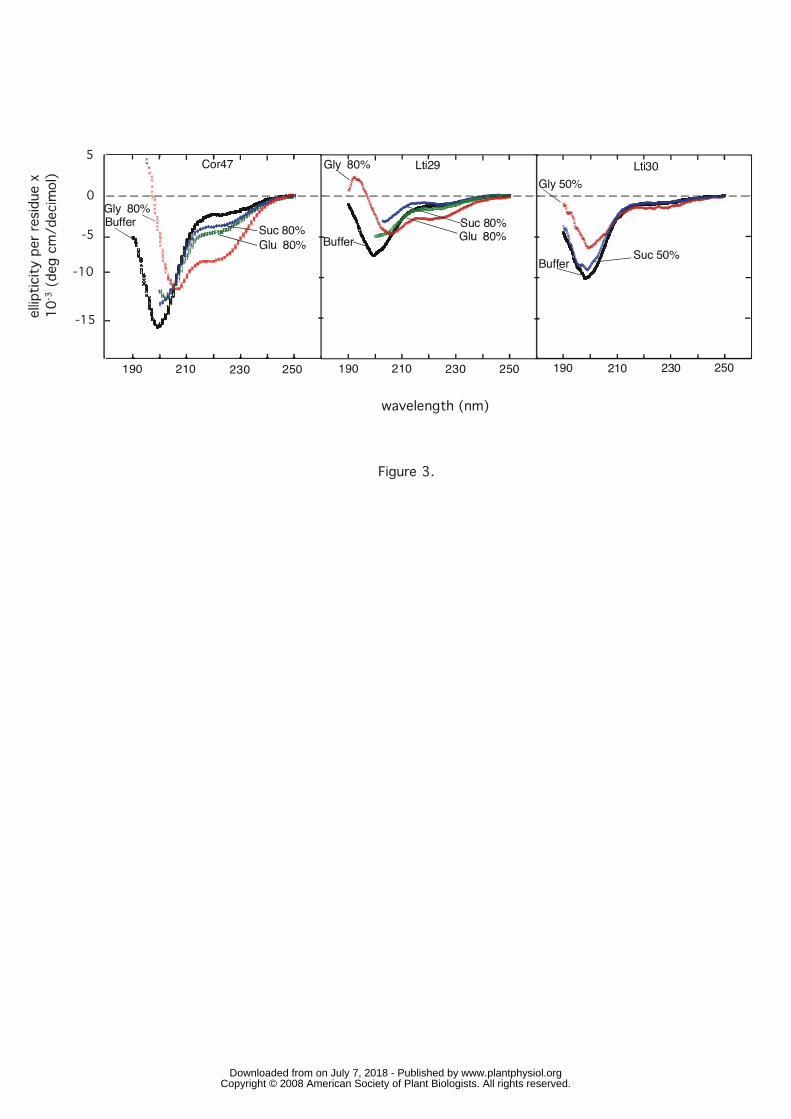
190 210 230 250 190 210 230 250 190 210 230 250
Buffer Suc 80%Gly 80%
Cor47
BufferSuc 80%
Gly 80% Lti29 Lti30
BufferSuc 50%
Gly 50%
-10
5
-5
-15
0
Figure 3.
wavelength (nm)
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Cor47
Lti29
Lti30
Cor47Lti29
Lti30
Difference spectra Glycerol (50%)
Difference spectra TFE (50%)
0
8
6
4
2
-2
10
0
-10
-20
Figure 4.
190 210 230 250
wavelength (nm)
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

222nm
200nm
Cor47
Glycerol (%)
-4
-8
-12
-16
0 20 40 60 80 100
Cor47 + Glycerol 0%-95%
Difference spectraCor47-Glycerol
75% - 0%95% - 75%
5
0
-5
-10
-15
10
5
0
-5
190 210 230 250
Figure 5.
wavelength (nm)
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

BufferDex7000Fic7000PEG4000PEG6000
Buffer
Dex7000Fic7000PEG4000PEG6000
Dex4000 BufferDex7000Fic7000PEG4000
Cor47 Lti29 Lti305
0
-5
-10
-15
Figure 6.
190 210 230 250 190 210 230 250 190 230210 250wavelength (nm)
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Dex7000Fic7000GlyPeg4000Peg6000
Dex4000
Difference spectra K-seg,Polymers 50%
Difference spectra,ChP-1Polymers 50%
Dex7000Fic7000GlyPeg4000Peg6000
3
2
1
0
-1
-2
3
2
1
0
-1
-2
Figure 7.
190 210 230 250wavelength (nm)
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Cor47 phosphorylated + glycerol
0
-5
-10
-15
190 210 230 250
Cor47+Gly 50%
Cor47 phosphorylatedCor47Cor47 Phosphorylated + Gly50%
wavelength (nm)
Figure 8
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.


















