
microbial ecology
15
A Radioisotope Assay for the Quantification of Hydrocarbon Biodegradation Potential in Environmental Samples D. M. CAPARELLO AND P. A. LAROCK Department of Oceanography, Florida State University, Tallahassee, bTorida 32306 Abstract An enrichment culture method is described for quantifying the activity of hydro- carbon oxidizing bacteria in water and sediments. Application of the procedure indicated that the hydrocarbon oxidizing potential of environmental samples reflects the hydro- carbon burden of the area, the ability of the microflora to utilize hydrocarbons, and that lakes with large aquatic plant communities contain populations of hydrocarbon bacteria comparable to those found in oil-polluted harbors. Introduction During the past 50 years at least five million tons of petroleum have been spilled into the sea (Pilpel, 1968). Currently, maritime spillage is estimated to be five to ten million tons per year while the volume of hydrocarbons derived from terrestrial sources such as road runoffs, and industrial effluent is approaching similar magnitudes (Blumer, 1968). This input, however, has not resulted in any great accumulation of oil. The disappearance is the result primarily of natural degradation measures initiated first by sunlight and followed by microbial bio- degradation [11 ]. Microbial hydrocarbon oxidizers are widely distributed on land and coastal waters, particularly in areas subject to frequent oil pollution (ZoBell [20]). Measurements of microbial hydrocarbon oxidation have been made using a variety of procedures including bacterial enumeration techniques (Davis [8]; Gunkel [12]), respirometric methods (Davis [8]; Jones [14, 15]; Davies and Hughes [7] ), labeled hydrocarbons (Davis and Raymond [9]), infrared spectro- photometry (Davis [8]), and oxygen electrodes (Buswell and Jurtshuk [5]). Each of these methods has its limitations. Infrared methods seem to be consis- tent for oils high in aliphatics but less suitable for lower members of the aro- matic series (Floodgate [10]). Manometric techniques are subject to vapor pres- 28 MICROBIAL ECOLOGY, Vol. 2, 28-42 (1975) 1975 by Springer-Verlag New York Inc.
-
Upload
nhanhnafi5 -
Category
Documents
-
view
216 -
download
0
Transcript of microbial ecology

A Radioisotope Assay for the Quantification of Hydrocarbon Biodegradation Potential
in Environmental Samples
D. M. CAPARELLO AND P. A. LAROCK
Department of Oceanography, Florida State University, Tallahassee, bTorida 32306
Abstract
An enrichment culture method is described for quantifying the activity of hydro- carbon oxidizing bacteria in water and sediments. Application of the procedure indicated that the hydrocarbon oxidizing potential of environmental samples reflects the hydro- carbon burden of the area, the ability of the microflora to utilize hydrocarbons, and that lakes with large aquatic plant communities contain populations of hydrocarbon bacteria comparable to those found in oil-polluted harbors.
Introduction
During the past 50 years at least five million tons of petroleum have been spilled into the sea (Pilpel, 1968). Currently, maritime spillage is estimated to be five to ten million tons per year while the volume of hydrocarbons derived from terrestrial sources such as road runoffs, and industrial effluent is approaching similar magnitudes (Blumer, 1968). This input, however, has not resulted in any great accumulation of oil. The disappearance is the result primarily of natural degradation measures initiated first by sunlight and followed by microbial bio- degradation [11 ].
Microbial hydrocarbon oxidizers are widely distributed on land and coastal waters, particularly in areas subject to frequent oil pollution (ZoBell [20]). Measurements of microbial hydrocarbon oxidation have been made using a variety of procedures including bacterial enumeration techniques (Davis [8]; Gunkel [12]), respirometric methods (Davis [8]; Jones [14, 15]; Davies and Hughes [7] ), labeled hydrocarbons (Davis and Raymond [9]), infrared spectro- photometry (Davis [8]), and oxygen electrodes (Buswell and Jurtshuk [5]). Each of these methods has its limitations. Infrared methods seem to be consis- tent for oils high in aliphatics but less suitable for lower members of the aro- matic series (Floodgate [10]). Manometric techniques are subject to vapor pres-
28
MICROBIAL ECOLOGY, Vol. 2, 28-42 (1975) �9 1975 by Springer-Verlag New York Inc.

Radioisotope Assay for Quantification of Hydrocarbon Biodegeneration Potential 29
sure limitations when used with some hydrocarbons (Floodgate [10]). Davis [8] found manometric procedures to be complicated systems often requiring 5 to 30 days to obtain measurements for ethane oxidizing bacteria. Bacterial enumeration procedures have been described by several investigators. Gunkel [12] used a high speed homogenizer with an emulsifier to prepare serial dilu- tions for the Most Probable Number estimates. Davis [8] cites several advantages for the plate count technique, but Jones and Edington [16] found plating to be unwieldy owing to the precautions necessary to insure that the agar be free of organic impurities. Plate counts may even result in the emergence of cultures that in the natural habitat are of minor significance in oxidizing hydrocarbons. Jones [15] used a buried slide technique to enumerate hydrocarbon oxidizers in soils. He concluded, however, that population studies in microbial oil degrada- tion were less useful than the measurement of oxidizing activity since bacterial
numbers yield no information on the rate of microbial oil degradation. Gunkel
[12] had arrived at the same conclusion.
The purpose of this paper is to describe a simple enrichment assay pro- cedure that permits a rapid quantification of bacterial hydrocarbon oxidizing
activity in natural environments.
Material and Methods
Cultural Procedures
Marine hydrocarbon oxidizing bacteria were grown on enriched seawater (ENSW) composed of (NH 4) 2SO4, l0 ppt; K2HPO4, 50 ppm; and yeast extract, 0.1% in artificial seawater ("Instant Ocean," Aquarium Systems Inc., Wickliffe, Ohio). Freshwater bacteria were grown on Bushnell and Haas mineral salts medium (BH), supplemented with 0.1% yeast extract (Bushnell and Haas [4]). Estuarine bacteria were grown on BH medium to which 1.0% NaC1 and 0.05% yeast extract were added. When a solid medium was desired for plate counting, 1.5% agar was added to the appropriate basal salts medium. The hydro- carbon source was Fisher heavy paraffin oil and Fisher kerosene (PK) mixed 1:1 v/v and was added to the lids of inverted Petri dishes. Cultures for laboratory experiments were isolated from marine and freshwater environments or~ the appropriate medium except the yeast extract concentration was lowered to 0.01%. After isolation, cultures were main- tained on slants of the appropriate agar medium with 1 ml of sterile PK pipetted on the agar surface.
The two hydrocarbon sources uscd were 14Ci-hexadecane (14C-16) and a synthetic mixture of commercially available n-alkanes varying in chain length from C-t0 to C-25 (HC MIX). HC MIX was prepared so that the lower molecular weight alkanes (C-10 to C-17) were present in twice the concentration of C-18 to C-25 to maintain the complete mixture as a liquid at room temperature.
Sample Collection
Water and slick samples were collected by skimming the surface with a sterile "Whirl-pak." Sediment samples were collected by inserting sterile 8 mm diam. glass core tubes into the sediments and then plugging the ends with sterile corks. All samples were refrigerated until assayed, which was usually within 12 hr after collection.

30 D . M . Caparello and P. A. LaRock
Determination of biomass by adenosine triphosphate (ATP) was made according to the method of Quammen et aL[ l 9].
Measurement o f Microbial Hydrocarbon Oxidation
1. Radioisotope Assay. Bacterial hydrocarbon oxidizing activity was measured by collecting tile 14CO 2 evolved from the oxidation of 14C1-16. The procedure used was patterned after the technique of Harrison et al. [13] with minor modifications to permit the use of hydrocarbons. In place of serum bottles, 50 ml Erlenmeyer flasks were used, each having 4 glass vanes indented around the base to assure mixing and dispersal of the hydrocarbon in the basal medium. The Erlenmeyer flasks also minimized splashing of the medium onto the filter paper CO 2 trap at the high shaking rates used.
In application, a series of 50 ml Erlenmeyer flasks containing 10 ml of either BH or ENSW media were sealed with aluminum foil and autoclaved for 20 rain at 15 psi. Next, 1 ml of the sample to be assayed, either a suspension of a pure culture or environmental sample, was added to each test flask. Sediment inocula were prepared by first withdrawing the overlying water in the corer to within l cm of the sediment. The sediment was extruded and discarded until only the top 2 cm of sediment remained in the corer. This surface sedi- ment was emptied into a reaction flask with a separate core of each sample used to inocu-
late the desired number of flasks.
Twenty-five microliters of 14C-16 with a specific activity of 2 )< 106 dpm/ml was then added to the water surface of each flask using a 50 ~1 syringe (Hamilton Corp.). Repeti- tive testing indicated that this method of adding hydrocarbons was highly reproducible for a given operator. The flasks were sealed with a rubber stopper (Priezer Scientific, Philadelphia, Pa.) fitted with a center well and Filter paper assembly (Harrison et al. [13]) and incubated at 35~ on a gyrotory shaker at 240 rpm. Uninoculated flasks served as controls.
After the desired incubation time, duplicate growth flasks and controls were sacri- riced by injecting 0.2 ml of &phenethyla_mine on the filter paper, followed by 2.0 ml of 1 N H2SO 4 into the medium to stop the reaction and release the CO 2. After acidification, the flasks were shaken for an additional hour to permit absorption of the I4CO 2. The filter paper was removed, placed in 15 ml of Aquasot (New England Nuclear, Boston, Mass.), and counted on a Nuclear Chicago Model 4534 liquid scintillation counter. Quench was corrected for by the channels ratio method. The activity of duplicate flasks was averaged and the 14CO 2 evolved expressed as a percentage of the activity initially added as 14C-16
according to the following equation:
Percent 14CO2 evolved - dpm (experimental filter) - dpm (control filter) • 100
dpm (14C-16 added)
The 14CO 2 evolved was expressed as a percentage of the activity added to compen- sate for variations in specific activity of different batcfies of the hexadecane.
2. Gravimetric Assay for the Determination o f n-Alkane Oxidation. In batch culture experiments using HC MIX, hydrocarbon loss was determined gravimetrically. Duplicate 125 ml Erlenmeyer flasks, each containing 20 ml of ENSW for marine bacteria, or 20 ml of BH for freshwater organisms, were inoculated with 1 ml of a suspension of the desired culttue to give a final density of 5 • 106 cells]ml. Next, 50 pl of HC MIX were added with a syringe, the flasks plugged with cotton and sealed with aluminum foil, The flasks were incu- bated at 35~ for 24 hr on a gyrotory shaker operating at 160 rpm. After incubation, 1 pl

Radioisotope Assay for Quantificafioz~ of Hydrocarbon Biodegeneration Potential 31
of 14C_16 in a hexane solvent was added to the reaction flasks followed by 20 ml of hexane for the extraction of the residual hydrocarbons. The radioisotope was used to determine the efficiency of the extraction process and had an activity of approximately 10,000 dpm/td, The/]asks were shaken for t hr and the phases separated. The hexane containing the remain- ing HC MIX was concentrated under a stream of nitrogen to 2.5 ml in calibrated glass- stoppered centrifuge tubes. Next, 0.5 ml of this solution was withdrawn and counted using 15 ml of Aquasol and the efficiency of the extraction determined. The remaining solution was transferred to a tared aluminmn weighing dish, the hexane evaporated under nitrogen, and the dishes reweighed on a Mettler HI6 balance. The percentage of the hydrocarbons oxidized was calculated from the following equation:
Percent hydrocarbon oxidized 0 . 8 N W - WJ x 10 2 0.8 NW
where
N = extraction efficiency W = weight of 50 #1 of HC MIX
14[/ = weight of residual HC MIX.
Generation o f Growth Curves
The effect of inoculmn density on culture outgrowth under conditions used in the radioassay was studied for several pure cultures and enviromnental samples. Environmental samples were used directly to inoculate the growth flasks. Pure cultures were pregrown on the appropriate agar and incubated for 24 hours at 35~ The cells were harvested with BH or ENSW medium and the suspension brought to an optical density of 0.3 at 610 rim. This yielded a suspension containing between 2 X 108 to 2 X 109 cells/ml depending on the organisms used. The desired inoculum density was obtained by serial dilutions to cover a range of 2.2 x 103 to 2.2 • 106 cells/flask, using ml inoculum size of 1 ml. Preparation of the growth flasks and their analysis was carried out as described in the radioisotope assay except that duplicate flasks of each sample were sacrificed at the first indication of turbidity and at 2 - 4 hr intervals thereafter. The experiment was terminated after 30 hr.
Results
In the radioassay, we a s sumed n - h e x a d e c a n e cou ld replace a m i x t u r e o f
n-a lkanes as a h y d r o c a r b o n subs t r a t e w i t h o u t a f fec t ing the relat ive q u a n t i t y o f
h y d r o c a r b o n oxid ized . We f u r t h e r a s sumed tha t the ~ 4CO2 evolved would serve
as an accura te measure o f the h y d r o c a r b o n ut i l ized. To test this ra t ionale , a
series o f tes t cu l tu res hav ing vary ing abi l i ty to ut i l ize h y d r o c a r b o n s (Table 1)
were g rown us ing 14C-16 u n d e r the c o n d i t i o n s o f the radioassay and w i t h a
m i x t u r e o f n-a lkanes u n d e r the c o n d i t i o n s o f the g rav imet r ic assay. All flasks
were i n c u b a t e d 24 hr .
In Fig. 1, t he results o f the t w o assay m e t h o d s have been p l o t t e d against
one a n o t h e r . Bo th m e t h o d s r anked the test cu l tu res in the s~une o rde r accord ing
to thei r abi l i ty to use the respect ive h y d r o c a r b o n subs t ra te . The l inear i ty o f the
po in t s ind ica tes tha t , numer ica l ly , the m a g n i t u d e o f the response each m e t h o d

32 D. M. Caparello and P. A. LaRock
Table 1 Test Cultures and Location o f Isolation
Culture Area of isolation
FHR-2 Surface water from the Fenholloway River, Perry, Florida
LM-1 Surface water from Lake Munson, Tallahassee, Florida
LM-2 Surface water from Lake Munson
HR-2 Surface water from Hillsborough River, Tampa, Florida
C-5 Beach sand from Coal Oil Point, California
L-2 Surface oil slick in Barataria Bay, Louisiana
C-22 Water from Boston Harbor, Boston, Massachusetts
12
10
>. 8-
~ -
2"
HR-2
LM-2
6 1'0 1'4 1'8 22 26 3"0 3'4 Percent HC MIX degraded - gravimetrie determination
Fig. 1. Comparison of the radioisotope assay and gravimetric technique as mea- sures of hydrocarbon oxidation by pure cultures, lnoculum size for each culture was 10 8 cells.

Radioisotope Assay for Quantification of Hydrocarbon Biodegeneration Potential 33
measures is directly proportional. Hexadecane may therefore substitute for a mixture of n-alkanes, and the 14C02 evolved is a measure of the ability of the test culture to oxidize n-alkanes.
The growth characteristics and rates of 14CO 2 evolution of active and non- active hydrocarbon oxidizers were examined using the radioassay (Fig. 2). The 14CO2 evolved for each culture indicated that the active oxidizer, HR-2, had a much higher stationary phase 14CO2 evolution than either C-22 or C-5. The length of the lag and slope of the exponential phases of each culture varied. The more active oxidizer had the greatest log phase slope and the shortest lag time. The weakest oxidizer, C-5, had the smallest log phase slope and the longest lag
�9 time. The inoculum density for C-5 was 3.3 X 108 cells/flask, for C-22 it was 4.1 • 108 cells/flask, and for HR-2 it was 2.2 X 104 cells/flask.
A family of growth curves was generated using the radioassay for the active oxidizer HR-2 over an inoculum range of 2.2 X 103 to 2.2 • 106 cells/flask (Fig. 3). All inoculated flasks eventually reached the same stationary phase cell density and 14CO 2 evolution. The length of the lag time for each curve was in- versely proportional to the logarithm of the inoculum density (Fig. 4). The dis- placement of the growth curve, or more precisely the length of the lag period, is therefore a measure of the size of the bacterial population in the original sample. Varying the inoculum density for a given culture did not affect the slope of the
12 A
10
O~ 6 HR-2
~, C-22 n
2
0 ;- I I I I I I I I | I
10 12 14 16 18 20 22 24 26 28 30
Time, hr
Fig. 2. Growth characteristics and rates of 1 4 C O 2 evolution for three pure cul- tures of varying hydrocarbon oxidizing activity. The cultures were HR-2, C-22, and C-5 with inoculum densities of 2.2 X 10 4, 4.1 • 108, and 3.3 X 108 cells/ flask, respectively.

34 D . M . Caparello and P. A. LaRock
exponential phase or the quantity of 14CO2 evolved when the culture reached the stationary phase.
The growth characteristics of environmental samples were investigated by generating a set of growth curves similar to those of the pure culture experiment. Water samples (Table 2, sample 4) were collected from the Sparkman ship canal in Tampa Bay, Florida and used as an inoculum full strength, and with 5-, 10-, and 102-fold dilutions. The resultant growth curves (Fig. 5) had the same basic features of the pure culture experiment. The length of the lag phase was inversely proportional to the logarithm of the inoculum density, each curve had a similar slope for the exponential phase, and the same quantity of 14CO2 was evolved when the cultures reached the stationary phase.
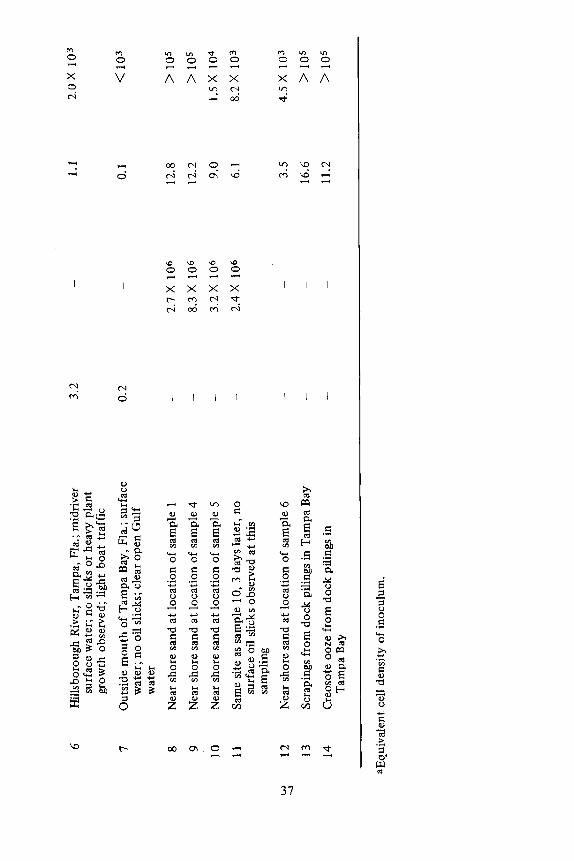
The hydrocarbon oxidizing activity of the microflora in a number of en- vironmental samples was measured using the radioassay. In these experiments, 1 ml of aqueous sample or a single core (1 cc total volume) was added to a series of reaction flasks and incubated for 24 hours. The 14CO2 evolved in this incuba- tion period is presented in Table 2 and suggests the oxidizing potential of an en- vironmental sample may reflect the degree of hydrocarbon addition to which the area is subjected. Sample 1 had the greatest ~ 4CO 2 evolved by the water samples but the activity of the freshwater samples surpassed most of the estuarine and marine samples. Comparison of the 14CO2 evolved and ATP biomass of samples 1, 4, 5, 6, and 7 indicated that samples from areas of relatively little hydro- carbon burden (samples 6 and 7) had less 14CO 2 evolved than polluted areas did (samples 1, 4, and 5). Estimates of biomass using ATP will indicate only the total microbial population irrespective of what elements compose the microflora.
10
2
E 4
2
A E F G ~ ,8 6, .4 . ,2,1..5 ,,8,,6,4, , 2 ! .~ .86 4 . , 2 ! ~
12 106 105 104 103
L I I I L 1 I I 1 I t I I - ~ [ 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Time, hr
Fig. 3. The ef fec t of vary ing the i n o c u l u m dens i ty on the pe rcen t J 4CO2 evolved by culture HR-2. The inoculum density for the family of curves is: A, 2.2 X 106; B, 1.7X 106; C, 1.1 X 106;D, 5.5 X 10s; E, 2.2X IOS;F, 2.2X 104;G, 2.2X 103 cells/flask. The log scale insert is used to interpolate between curves.

Radioisotope Assay for Quantification of Hydrocarbon Biodegeneration Potential 35
Samples 1 and 6 had nearly equivalent biomass but the 14C0 2 evolved by sample 6 was one-tenth that of sample 1.
Analysis of sand (samples 8 to 12) showed the same trend as was found with the corresponding water samples. However, a greater amount of 14CO2 was evolved than was found with the water samples. This is seen by comparing samples 1, 4, 5, and 6 to samples 8, 9, 10, and 12, respectively. The most active samples were cresote scrapings collected from dock pilings (samples 13 and 14).
7
-8 E
o
_J
3 I 1 1 1 1
0 5 10 15 20 25 3 0
Time, hr, for 5% 14C02 evolved
Fig. 4. Duration of the lag phase for culture HR-2 as a function of the log of inoculum density.

Tab
le 2
L
ocat
ion
and
Des
crip
tion
o
/En
viro
nm
enta
l Sa
mpl
es T
este
d w
ith
the
Rad
ioas
say
Sam
ple
Loc
atio
n an
d de
scri
ptio
n
Pla
te
AT
P C
once
ntra
tion
co
unt
(/ag
lit
er-
1 )
(cel
ls m
l- 1
) i
4C
O 2
Ev
olv
ed
(per
cent
)
Rad
ioas
say
esti
mat
ed
popu
lati
on
(cel
ls m
l- 1
)a
0%
4
Spa
rkm
an s
hip
chan
nel,
Tam
pa B
ay,
Fla
.;
near
sho
re s
urfa
ce w
ater
; pr
omin
ent
surf
ace
oil
slic
ks; a
ctiv
e sh
ippi
ng
acti
vity
in
chan
nel
Lak
e M
unso
n, T
alla
hass
ee,
Fla.
; ne
ar
shor
e su
rfac
e w
ater
, su
rfac
e sl
icks
pr
esen
t; r
ecei
ves
city
tre
ated
sew
age;
la
rge
alga
l an
d pl
ant
popu
lati
on
Lak
e Ja
ckso
n, T
alla
hass
ee,
Fla
.; s
hore
su
rfac
e w
ater
; no
sli
cks
obse
rved
; la
rge
plan
t po
pula
tion
Cov
e in
Spa
rkm
an c
hann
el;
near
sho
re
surf
ace
wat
er;
litt
le o
r no
sli
cks
ob-
serv
ed; r
ecei
ves
sew
age
effl
uent
Cov
e ad
jace
nt t
o lo
cati
on 4
; ne
ar s
hore
su
rfac
e w
ater
; re
ceiv
es s
ewag
e ef
flue
nt;
no o
il s
lick
s ob
serv
ed
5.0
16.0
3
.6 X
10
6
17.0
2.
9 X 1
0 6
11.7
9.9
4.4
2.5
2.0
> 10
s
1.5
X 1
0 4
5.5
X 1
0 3
3.6
X 1
0 a
3.2
X 1
0 3
6 8 9 10
11
12
13
14
Hil
lsb
oro
ug
h R
iver
, T
amp
a, F
la.;
mid
riv
er
surf
ace
wat
er;
no
sli
cks
or h
eav
y p
lant
g
row
th o
bse
rved
; li
ght
bo
at t
raff
ic
Ou
tsid
e m
ou
th o
f T
amp
a B
ay,
Fla
.; s
urfa
ce
wat
er;
no
oil
sli
cks;
cle
ar o
pen
Gu
lf
wat
er
Nea
r sh
ore
san
d at
lo
cati
on
of
sam
ple
1
Nea
r sh
ore
san
d at
lo
cati
on
of
sam
ple
4
Nea
r sh
ore
san
d at
lo
cati
on
of
sam
ple
5
Sam
e si
te a
s sa
mp
le 1
0, 3
day
s la
ter,
no
su
rfac
e oi
l sl
icks
ob
serv
ed a
t th
is
sam
plin
g
Nea
r sh
ore
san
d at
lo
cati
on
of
sam
ple
6
Scr
apin
gs f
rom
do
ck p
ilin
gs i
n T
amp
a B
ay
Cre
oso
te o
oze
fro
m d
ock
pil
ings
in
Tam
pa
Bay
3.2
0.2
F
2.7
X 1
0 6
8.3
X 1
0 6
3.2
X 1
0 6
2.4
X 1
0 6
1.1
0.1
aEq
uiv
alen
t ce
ll d
ensi
ty o
f in
ocu
lum
.
12.8
12.2
9.0
6.1
3.5
16.6
11.2
2.0
X 1
0 3
<
10 3
lO s
lO s
1.5
X 1
0 4
8.2
X 1
0 3
4.5
X 1
0 3
10 s
l0 s

38 D.M. Caparello and P. A. LaRock
0 2
8 1
o .
I I I I I I I I I I I i 0 2 4 6 8 10 12 14 16 18 20 22 24
Time, hr
Fig. 5. The effect of varying the inoculum density on the percent 14CO 2 evolved by an environmental surface water sample from Sparkman Channel, Tampa Bay, Florida. The inoculum density for the family of curves is: A, 3.6 X 106; B, 2X 105;C, 3.6 X 105;D, 3.6 • 104 cells/ml.
In comparing the radioassay estimations (for a 24 hr incubation period) to ATP biomass and plate counts (Table 2), it is seen that the biomass for sample 1 was less than for samples 4 and 5, but the 14CO z evolved was five times greater. Similarly, plate counts of samples 8, 10, and l 1 were equivalent to samples 4 and 5 but their 14CO 2 evolved was three to five times greater. This suggests that the hydrocarbon oxidizing potential of a sample is a function of the oxidiz- ing activity of the microflora as well as its cell density.
Discussion
In the enrichment culture technique described, the important parameters of bacterial development are (1) the quantity of 14CO 2 evolved is a measure of the amount of hydrocarbon oxidized (Fig. 1); (2) the quantity of 14CO2 evolved is a measure of the culture's ability to utilize hydrocarbons (Fig. 2); (3) the length of the lag phase of growth is a function of the inoculum density. The more cells added to the growth flasks, the shorter the lag period (Fig. 3); and (4) the quantity of CO2 evolved in the stationary phase for a given culture is independent of the inoculum density, but for smaller inocula a longer incuba- tion period is required to attain this final 14CO 2 concentration (Figs. 3 and 4).
These features permit the radioassay to be used to quantify hydrocarbon oxidizing activity in two ways. The first method merely measures the 14CO2 released after a fixed incubation period and thus indicates the relative ability of the sample to degrade hydrocarbons. For example, if the cultures shown in Fig. 2

Radioisotope Assay for Quantification of Hydrocarbon Biodegeneration Potential 39
were assayed at 24 hr, the ability of each to oxidize hydrocarbons would be proportional to the amount of 14CO2 evolved during the fixed incubation period. On this basis, culture HR-2 would be found to be the strongest oxidizer and the C-5 the weakest. When used with environmental samples, the amount of 14CO 2 released in a fixed incubation period is thus a relative measure of the ability of the specified sample to assimilate hydrocarbon materials.
The second method for using the radioassay introduces time as an addi- tional variable, thereby extending the range of the tectmique. In general, the duration of the lag phase of growth in a batch culture is inversely proportional to the logarithm of tile cell density in the inoculum (Lamanna and Mallette [17]). Using a hydrocarbon oxidizing bacterium (Culture HR-2, Table 1), a series of growth curves were generated for inocula of different sizes (Fig. 3) and" the duration of the lag was found to be proportional to the logarithm of the inoculum density under the conditions of the radioassay (Fig. 4). The duration of the lag phase may thus be used to arrive at a numerical estimate of the cell density in the inoculating material. Under this condition the growth curves of Fig. 3 are used to translate differences in lag into population estimates. Using the incubation time and 14CO2 evolved by an environmental sample or test cul- ture, a point is located within the family of standard curves (Fig. 3), and a numerical estimate of the bacterial population is read off the curves. The cell density so obtained is, in reality, the inoculum density of the standardizing cul- ture that will produce an equivalent amount of 14CO 2 in the same incubation period, and not the actual population density contained in the sample. This technique permits comparison of the hydrocarbon oxidizing activity of different environments made up of different populations. The laboratory culture used to generate the standard curves serves only to provide a base for comparing environ- mental samples. The actual activity of the standardizing culture is not important.
In using Fig. 3 to estimate the number of hydrocarbon bacteria in a sample, one must consider that the radioassay indicates only that the quantity of 14CO2 evolved by a sample can be produced in the same period of time by a laboratory culture of known inoculum density. Thus, a large population of less active oxidizers and another smaller, but more active population could produce similar amounts of 14CO 2 and therefore be interpreted as equal populations. Numerically, the populations are not equivalent, but their ability to assimilate hydrocarbons is.
Application of the radioassay to environmental samples collected in and around Tampa Bay and in two North Florida lakes indicated that hydrocarbon oxidizing bacteria are ubiquitous. They were found in water and sediment in both unpolluted and oil polluted areas, as had previously been reported (ZoBell [20, 21]). The abundance of oil degrading bacteria, however, was greatest in areas receiving waste hydrocarbon materials. The water sample taken from a ship canal in Tampa Bay, where there were visible oil slicks, (Table 2, sample 1), had

40 D .M. Caparello and P. A. LaRock
greater hydrocarbon utilizing populations than did the Hillsborough River which feeds into the bay (sample 6) or the Gulf of Mexico at the mouth of the bay (sample 7). The total microbial biomass, as estimated by ATP, for samples 1,4, 5, and 6 varied by a factor of 5. The hydrocarbon utilizing population, however, varied by a factor of at least 50 (Table 2). The enhanced hydrocarbon oxidation observed for sample 1 indicates that for a given microbial biomass, exposure of the sample to oil will either increase the hydrocarbon utilizing component or replace the existing bacterial component by another population that is more adapted to oxidize oil. The low hydrocarbon oxidizing activity and high ATP content of samples 4 and 5, relative to sample 1, may be the result of the addi- tion of the effluent of a sewage treatment plant. Under similar conditions in Raritan Bay, N.J., Atlas and Bartha [2] reported a substantial increase in the total number of microorganisms at a sewage outfall, but no change was noted in the numbers of oil oxidizing bacteria.
Atlas and Bartha [1 ] further indicated that low phosphate and nitrate con- centrations are limiting factors in the microbial degradation of hydrocarbons, particularly in seawater. They found that no observable microbial oil degradation occurred within 18 days in offshore water with 0.98 rag/liter nitrogen and .07 mg]titer phosphorous. The nitrate-nitrogen and phosphate-phosphorous concen- trations in sample 7 were 0.158 mg/liter and 0.067 mg/liter, respectively. The low oxidizing activity at this location may be the result of either low nutrient concentrations or a microbial population that was relatively inactive for hydro- carbon oxidation. In Tampa Bay proper neither nitrates or phosphates are limit- ing as a result of the discharge of sewage and the wastes from phosphate mining.
The analysis of sediment or sand samples (Table 2, samples 8 - 12) indi- cates that areas with higher hydrocarbon burdens have greater hydrocarbon oxidizing activity and that sediments have larger hydrocarbon utilizing popula- tions than the overlying water. Similar results have been reported by Colwell et al. [6] in Chesapeake Bay where sediments from oil polluted areas contained as much as 100 times more oil utilizing bacteria than did the overlying water.
Of great interest was the finding that freshwater lakes contain hydrocar- bon utilizing populations of comparable magnitude to those found in oil pol- luted estuarine environments. Lake Munson receives the effluent from a sewage treatment plant and the nutrients added to the lake have resulted in the de- velopment of water plants and frequent heavy algal blooms. At times visible oil slicks are present which could originate either from grease and oils in the sewage or from plant hydrocarbons. Lake Jackson is located in a relatively undeveloped area and as such receives runoff from predominantly wooded land. At present we can only speculate that the hydrocarbon oxidizing activity found in these lakes may be in response to material contributed primarily as plant hydrocarbons, al- though a more exhaustive study is necessary to resolve this question.

Radioisotope Assay for Quantification of Hydrocarbon Biodegeneration Potential 41
A c k n o w l e d g m e n t s
Dominic M. Caparcllo was supported by NSF Grant GZ-2337 to Florida State Uni- versity. Samples were collected in Tampa Bay aboard the R. V. Tursiops through funds pro- vided by the NSF under Contract GC-28174.
Re fe rences
1. Atlas, R. M., and Bartha, R. 1972. Degradation and mineralization of petroleum in sea water: Limitation by nitrogen and phosphorous. Bioteeh. Bioeng. 14: 309 - 318.
2. Atlas, R. M., and Bartha, R. 1973. Abundance, distribution and oil biodegradation potential of microorganisms in Raritan Bay. Environ. Pollut. 4:291 - 300.
3. Blumer, M. 1968. Oil pollution of the ocean. In." Oil on the Sea. D. Hould, editor. Plenum Press, New York. pp. 5 - 13.
4. Bushnell, L. D., and Haas, H. F. 1941. The utilization of certain hydrocarbons by microorganisms. J. Bacteriol. 41:653 - 673.
5. Buswell, J. A., and Jurtshuk, P. 1969. Microbial oxidation of hydrocarbons measured by oxygraphy. Arch. Microbiol. 64:215 - 222.
6. Colwell, R. R., Walker, J. D., and Nelson, J. D. 1973. Microbial ecology and the problem of petroleum degradation in Chesapeake Bay. ln: The Microbial Degradation o f Oil Pollutants. D. G. Ahearn and S. P. Meyers, editors. Center for Wetland Resources, Louisiana State Univ. Publ. No. LSU-SG-73-01. pp. 185 -197.
7. Davies, J. A., and Hughes, D. E. 1968. The biochemistry and microbiology of crude oil degradation, ln: Field Studies Council Symposium. The Biological Effects o f Oil Pollution on Littoral Communities. Carthy and Arthur, editors. Field Studies Council, London, pp. 139 - 144.
8. Davis, J. B. 1967. Petroleum Microbiology. Elsevier, New York, 604 pp.
9. Davis, J. B., and Raymond, R. L. 1961. Oxidation of alkyl substituted cyclic hydro- carbons by a Noeardia during growth on n-alkanes. Appl. Microbiol. 9: 383 - 388.
10. Floodgate, G. D. 1972. Biodcgradation of hydrocarbons in the sea. In: Water Pollu- tion Microbiology. R. Mitchell, editor. Wiley-lnterscience, New York. pp. 153 - 171.
l l . Friede, J., Quire, P., Gholson, R., Gaudy, E., and Gaudy, A. F. 1972. Assessment of biodegradation potential for controlling oil spills on the high seas. Project 4110.1/3.1, Dept. of Transportation. 130 pp.
12. Gunkel, W. 1968. Bacteriological investigations of oil polluted sediments from the Cornish coast following the Torrey Canyon disaster, ln: Field Studies Council Symposium. The Biological E lfeets o]" Oil Pollution on Littoral Communities. Carthy and Arthur, editors. Field Studies Council, London, pp. 115 - 158.
13. ttarrison, M. J., Wright, R. T., and Morita, R. Y. 1971. Method for measuring mineralization in lake sediments. Appl. Microbiol. 21:698 - 702.
14. Jones, J. G. 1968. Methods of studying the effect and metabolism of hydrocarbons in natural environments, hT: Field Studies Council Symposium. The Biological

42 D . M . CapareUo and P. A. LaRock
Effects o f Oil Pollution on Littoral Communities. Carthy and Arthur , editors. Field Studies Council, London, pp. 145 - 150.
15. Jones, J. G. 1969. The determinat ion of microbial hydrocarbon metabol ism in natural environments. Arch. Microbiol. 6 7 : 3 9 7 - 407.
16. Jones, J. G., and Edington, M. A. 1968. An ecolo~ca~ survey of hydrocarbon- oxidizing micro-organisms. J. Gen. MicrobioL 5 2 : 3 8 1 - 390.
17. Lamanna, C., and Mallette, M. F. 1965. Basic Bacteriology, 3rd ed. The Williams and Wilkins Co., Baltimore.
18. Pilpel, N. 1968. The natural fate o f oil on the sea. Endeavors' 2 7 : 1 1 - 13.
19. Quammen, M., LaRock, P. A., and Calder, J. A. 1973. Environmenta l effects o f pulp mill wastes. In. Proceedings o f the Belle Baruch Symposium on Estuarine Microbial Ecology. H. L. Stevenson and R. R. Colwell, editors. Univ. o f South Carolina, Columbia, South Carolina. In press.
20. ZoBell, C. E. 1969. Microbial modif icat ion of crude oil in the sea. In: Proceedings o]" Joint Conference on Prevention and Control o f Oil Spills, 1969. American Petroleum Insti tute, New York. pp. 317 - 326.
21. ZoBell, C. E. 1972. Microbial degradation o f oil: Present status, problems and per- spectives. In: The Microbial Degradation o f Oil Pollutants. D. G. Ahearn and S. P. Meyers, editors. Center for Wetland Resources, Louisiana State Univ. Publ. No. LSU-SG-73-01. pp. 3 - 16.


















