
MHCI promotes developmental synapse elimination and aging...
12
Full-length Article MHCI promotes developmental synapse elimination and aging-related synapse loss at the vertebrate neuromuscular junction Mazell M. Tetruashvily a,b,1 , Marin A. McDonald c,d,1 , Karla K. Frietze a , Lisa M. Boulanger a,c,e,⇑ a Department of Molecular Biology, Princeton University, Princeton, NJ 08544, United States b Rutgers Robert Wood Johnson Medical School, Piscataway, NJ 08854, United States c Department of Neurosciences, University of California, San Diego 92093, United States d Medical Scientist Training Program, University of California, San Diego 92093, United States e Princeton Neuroscience Institute, Princeton University, Princeton, NJ 08544, United States article info Article history: Received 14 September 2015 Received in revised form 12 January 2016 Accepted 12 January 2016 Available online xxxx Keywords: MHCI Major histocompatibility complex class I Neuromuscular junction NMJ Synapse elimination Aging Development Denervation abstract Synapse elimination at the developing neuromuscular junction (NMJ) sculpts motor circuits, and synapse loss at the aging NMJ drives motor impairments that are a major cause of loss of independence in the elderly. Here we provide evidence that at the NMJ, both developmental synapse elimination and aging-related synapse loss are promoted by specific immune proteins, members of the major histocom- patibility complex class I (MHCI). MHCI is expressed at the developing NMJ, and three different methods of reducing MHCI function all disrupt synapse elimination during the second postnatal week, leaving some muscle fibers multiply-innervated, despite otherwise outwardly normal synapse formation and maturation. Conversely, overexpressing MHCI modestly accelerates developmental synapse elimination. MHCI levels at the NMJ rise with aging, and reducing MHCI levels ameliorates muscle denervation in aged mice. These findings identify an unexpected role for MHCI in the elimination of neuromuscular synapses during development, and indicate that reducing MHCI levels can preserve youthful innervation of aging muscle. Ó 2016 Elsevier Inc. All rights reserved. 1. Introduction Synapse elimination sculpts mature patterns of connectivity throughout the vertebrate nervous system. During prenatal and early postnatal development, axons retract, refining initially dif- fuse projections. Consistent with a critical role in circuit matura- tion, defects in synapse elimination have been implicated in neurodevelopmental disorders (e.g., (Hayashi-Takagi et al., 2014; Krey et al., 2013; Noutel et al., 2011; Pfeiffer et al., 2010; Tang et al., 2014; Tsai et al., 2012)). Synapse loss also occurs with aging, when it is associated with significant motor and cognitive impair- ments (reviewed in (Hedden and Gabrieli, 2004; Rudolf et al., 2014)). However, the molecular drivers of developmental synapse elimination remain largely unknown. Furthermore, it is unclear if developmental synapse elimination and aging-related denervation occur through related, or distinct, molecular mechanisms. The canonical model for studying developmental synapse elim- ination is the vertebrate neuromuscular junction (NMJ) (Brown et al., 1976; Thompson, 1985). Before birth, each muscle fiber receives inputs from many motor neurons (MNs) (Nguyen and Lichtman, 1996; Sanes and Lichtman, 1999; Tapia et al., 2012). MN axon branches are extensively pruned, and by the end of the second postnatal week in rodents, each muscle fiber receives input from only a single MN (Brown et al., 1976; Busetto et al., 2000; Keller-Peck et al., 2001; Personius and Balice-Gordon, 2001; Redfern, 1970). Previous studies have significantly advanced our understanding of the factors that control the rate of synapse elimination. Several manipulations either delay (Je et al., 2013; Lee et al., 2011; Li et al., 2004; Misgeld et al., 2002; Nguyen et al., 1998; Personius et al., 2008; Rafuse et al., 2000; Roche et al., 2014; Sleigh et al., 2014; Thompson et al., 1979) or accelerate (Bogdanik et al., 2012; O’Brien et al., 1978; Personius et al., 2007; Thompson, 1983) synapse elimination at the developing NMJ. However, in most cases where synapse elimination is delayed, uniform monoinner- vation is still established within the first few postnatal weeks. Thus the endogenous factors that ensure elimination of every excess MN input remain largely unknown. http://dx.doi.org/10.1016/j.bbi.2016.01.008 0889-1591/Ó 2016 Elsevier Inc. All rights reserved. ⇑ Corresponding author at: Princeton Neuroscience Institute, Princeton Univer- sity, Princeton, NJ 08544, United States. E-mail address: [email protected] (L.M. Boulanger). 1 Authors contributed equally. Brain, Behavior, and Immunity xxx (2016) xxx–xxx Contents lists available at ScienceDirect Brain, Behavior, and Immunity journal homepage: www.elsevier.com/locate/ybrbi Please cite this article in press as: Tetruashvily, M.M., et al. MHCI promotes developmental synapse elimination and aging-related synapse loss at the ver- tebrate neuromuscular junction. Brain Behav. Immun. (2016), http://dx.doi.org/10.1016/j.bbi.2016.01.008
Transcript of MHCI promotes developmental synapse elimination and aging...

Brain Behavior and Immunity xxx (2016) xxxndashxxx
Contents lists available at ScienceDirect
Brain Behavior and Immunity
journal homepage wwwelsevier comlocate ybrbi
Full-length Article
MHCI promotes developmental synapse elimination and aging-relatedsynapse loss at the vertebrate neuromuscular junction
httpdxdoiorg101016jbbi2016010080889-1591 2016 Elsevier Inc All rights reserved
uArr Corresponding author at Princeton Neuroscience Institute Princeton Univer-sity Princeton NJ 08544 United States
E-mail address lboulangprincetonedu (LM Boulanger)1 Authors contributed equally
Please cite this article in press as Tetruashvily MM et al MHCI promotes developmental synapse elimination and aging-related synapse loss at ttebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg101016jbbi201601008
Mazell M Tetruashvily ab1 Marin A McDonald cd1 Karla K Frietze a Lisa M Boulanger aceuArraDepartment of Molecular Biology Princeton University Princeton NJ 08544 United StatesbRutgers Robert Wood Johnson Medical School Piscataway NJ 08854 United StatescDepartment of Neurosciences University of California San Diego 92093 United StatesdMedical Scientist Training Program University of California San Diego 92093 United Statese Princeton Neuroscience Institute Princeton University Princeton NJ 08544 United States
a r t i c l e i n f o
Article historyReceived 14 September 2015Received in revised form 12 January 2016Accepted 12 January 2016Available online xxxx
KeywordsMHCIMajor histocompatibility complex class INeuromuscular junctionNMJSynapse eliminationAgingDevelopmentDenervation
a b s t r a c t
Synapse elimination at the developing neuromuscular junction (NMJ) sculpts motor circuits and synapseloss at the aging NMJ drives motor impairments that are a major cause of loss of independence in theelderly Here we provide evidence that at the NMJ both developmental synapse elimination andaging-related synapse loss are promoted by specific immune proteins members of the major histocom-patibility complex class I (MHCI) MHCI is expressed at the developing NMJ and three different methodsof reducing MHCI function all disrupt synapse elimination during the second postnatal week leavingsome muscle fibers multiply-innervated despite otherwise outwardly normal synapse formation andmaturation Conversely overexpressing MHCI modestly accelerates developmental synapse eliminationMHCI levels at the NMJ rise with aging and reducing MHCI levels ameliorates muscle denervation in agedmice These findings identify an unexpected role for MHCI in the elimination of neuromuscular synapsesduring development and indicate that reducing MHCI levels can preserve youthful innervation of agingmuscle
2016 Elsevier Inc All rights reserved
1 Introduction
Synapse elimination sculpts mature patterns of connectivitythroughout the vertebrate nervous system During prenatal andearly postnatal development axons retract refining initially dif-fuse projections Consistent with a critical role in circuit matura-tion defects in synapse elimination have been implicated inneurodevelopmental disorders (eg (Hayashi-Takagi et al 2014Krey et al 2013 Noutel et al 2011 Pfeiffer et al 2010 Tanget al 2014 Tsai et al 2012)) Synapse loss also occurs with agingwhen it is associated with significant motor and cognitive impair-ments (reviewed in (Hedden and Gabrieli 2004 Rudolf et al2014)) However the molecular drivers of developmental synapseelimination remain largely unknown Furthermore it is unclear ifdevelopmental synapse elimination and aging-related denervationoccur through related or distinct molecular mechanisms
The canonical model for studying developmental synapse elim-ination is the vertebrate neuromuscular junction (NMJ) (Brownet al 1976 Thompson 1985) Before birth each muscle fiberreceives inputs from many motor neurons (MNs) (Nguyen andLichtman 1996 Sanes and Lichtman 1999 Tapia et al 2012)MN axon branches are extensively pruned and by the end of thesecond postnatal week in rodents each muscle fiber receives inputfrom only a single MN (Brown et al 1976 Busetto et al 2000Keller-Peck et al 2001 Personius and Balice-Gordon 2001Redfern 1970)
Previous studies have significantly advanced our understandingof the factors that control the rate of synapse elimination Severalmanipulations either delay (Je et al 2013 Lee et al 2011 Li et al2004 Misgeld et al 2002 Nguyen et al 1998 Personius et al2008 Rafuse et al 2000 Roche et al 2014 Sleigh et al 2014Thompson et al 1979) or accelerate (Bogdanik et al 2012OrsquoBrien et al 1978 Personius et al 2007 Thompson 1983)synapse elimination at the developing NMJ However in mostcases where synapse elimination is delayed uniform monoinner-vation is still established within the first few postnatal weeks Thusthe endogenous factors that ensure elimination of every excess MNinput remain largely unknown
he ver-
2 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
In adults responses to peripheral motor neuron injury are reg-ulated by specific immune proteins members of the major histo-compatibility complex class I (MHCI) MHCI has been detected inadult motor neurons (Edstrom et al 2004 Linda et al 1999Thams et al 2009) but adult muscle cells are thought to repressesMHCI genes except after injury or in disease states (Karpati et al1988) Consequently studies of the role of MHCI in the peripheralnervous system have focused almost exclusively on responses toinjury In injury models reducing MHCI expression exacerbatesloss of synapses made onto axotomized motor neuron cell bodiesin the spinal cord and impairs functional recovery (Berg et al2013 Oliveira et al 2004 Thams et al 2009) These and otherstudies have reinforced the view that at the NMJ MHCI is primarilyinvolved in responses to injury In the central nervous systemMHCI has been implicated in pruning of developing retinal projec-tions (Datwani et al 2009 Huh et al 2000 Lee et al 2014) whileelimination of developing cerebellar climbing fibers is MHCI-independent (Letellier et al 2008 McConnell et al 2009) Herewe explored if MHCI plays a role in developmental synapse elimi-nation or aging-related synapse loss at the vertebrate NMJ
2 Results
21 Impaired synapse elimination in mice deficient for cell-surfaceMHCI
MHCI genes are expressed at the NMJ as synapse elimination isoccurring (Tetruashvily et al in press) If MHCI promotes synapseelimination at the developing NMJ then reducing MHCI expressionshould increase the number of multiply-innervated muscle fibersTo test this prediction we visualized innervation of postsynapticsites in diaphragm muscle fibers from mice lacking stable cell-surface expression of most MHCI proteins (b2mTAP micesee Section 4) The diaphragm is a thin planar muscle with awell-organized end plate band that is particularly amenable toimaging in whole-mount preparations Postsynaptic nicotinicacetylcholine receptors (nAChRs) were labeled using tetramethylrhodamine-conjugated a-bungarotoxin (a-btx) and presynapticmotor neuron axons were visualized using antibodies against theaxon cytoskeletal element neurofilament and the presynaptic vesi-cle protein SV2 Because individual nAChR clusters may lay aboveone another in whole mount muscle 3D reconstructions of confo-cal Z-stacks were created for all suspected multiply-innervatedpostsynaptic sites and reconstructed stacks were viewed frommultiple depths and angles to determine if motor neuron brancheswere distinct and if they innervated a single nAChR cluster or twoclusters in different focal planes (see Section 4)
In WT mice nearly all postsynaptic sites are contacted by a sin-gle MN axon by postnatal day 15 (P15) as expected (Kopp et al2000 Redfern 1970) In contrast significantly more postsynapticsites remain multiply-innervated in mice lacking cell-surface MHCI(Fig 1AndashC Multiple innervation at P15 b2mTAP 164 plusmn 22WT 35 plusmn 11 p = 00011) Multiply-innervated postsynaptic sitesdetected in P15 MHCI-deficient mice all received two inputsAlthough multiple innervation persists b2mTAP mice are ofnormal size and weight and eye opening fur growth weaningand reproduction all occur at appropriate ages Development ofthe NMJ is also otherwise outwardly normal Every muscle fibercontains only a single postsynaptic nAChR cluster at P15 and thearea perimeter and intensity of nAChR labeling and morphologyof the end plate band are all normal in b2mTAP mice(Fig S2) Furthermore the distribution of nAChR labeling changesas expected from plaque-like to pretzel-like as the animalsmatured Despite this maturation multiple innervation persistsin MHCI-deficient adults (Fig 1AndashC b2mTAP 186 plusmn 43WT 23 plusmn 17 p = 00116) and remarkably is still present at one
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
year of age (Fig 1D b2mTAP 95 plusmn 13 WT 25 plusmn 05p = 00069) Thus loss of MHCI prevents multiple innervation fromfully resolving at the NMJ
Loss of MHCI could cause multiply-innervated muscle fibers topersist because it directly or indirectly disrupts synapse elimina-tion or because it increases synapse formation via changes in neu-rogenesis programmed cell death or motor neuron outgrowth orbranching However several lines of evidence suggest that changesin synapse formation do not underlie the persistent multiple inner-vation in MHCI-deficient animals At birth (P0) and at P7 when MNprogrammed cell death and synapse formation are largely com-plete (Tapia et al 2012 Xu et al 2006 Yamamoto andHenderson 1999) a normal proportion of postsynaptic sites aremultiply-innervated in MHCI-deficient mice (Fig 1AndashC) Further-more at P7 each muscle fiber is contacted by the same averagenumber of axons in WT and MHCI-deficient mice (Fig S2E andF) Thus measures that are sensitive to changes in motor neuronand muscle fiber number and synapse formation are normal inMHCI-deficient mice These results suggest that in the absence ofMHCI patterns of innervation are established normally but inner-vation fails to decline over the second postnatal week consistentwith a disruption of developmental pruning
However it is still possible that loss of MHCI prevents theestablishment of monoinnervation without changes in synapseelimination eg by prolonging neurogenesis programmed celldeath andor synapse formation well past their normaldevelopmental window If synapse elimination is not impaired inMHCI-deficient mice then ongoing synapse formation would benecessary to explain the stable synapse density from P7 onwardsHowever we did not see evidence of ongoing axon sprouting out-growth or synapse formation over the second postnatal weekTogether these results suggest that changes in synapse formationare unlikely to contribute significantly to persistent multiple inner-vation in MHCI-deficient animals Instead they are more consis-tent with a role for MHCI in allowing synapse elimination toreach completion
Extensive synapse elimination occurs at the NMJ prior to theclose of the first postnatal week (Tapia et al 2012) This initialsynapse elimination does not require MHCI because at P7 the per-centage of postsynaptic sites that remain multiply-innervated(Fig 1) and the extent of MN convergence onto individual postsy-naptic sites (Fig S2E and F) are both normal in MHCI-deficientmice Consistent with the fact that early synapse elimination hap-pens normally MHCI-deficient mice do not display gross motordeficits (eg weakness inactivity or tremor) (McConnell et al2009 Nelson et al 2013) Over the second postnatal week how-ever far fewer postsynaptic sites achieve mono-innervation inMHCI-deficient animals a failure of circuit maturation that coulddisrupt the emergence of fine motor control Remarkably there isno further decline in multiply-innervated postsynaptic sitesbetween P15 and adulthood (Fig 1B and C)
22 Loss of MHCI impairs synapse elimination specifically during thesecond postnatal week
If MHCI is specifically involved in synapse elimination during thesecond postnatal week then acutely disrupting MHCI function dur-ing the second postnatal week should causemultiple innervation topersist To test this hypothesiswe inhibitedMHCI function fromP7-P15 using an anti-MHCI antibody (OX18) that is function-blockinginmultiple assays Incubating hepatocytes or tumor cells in OX18 issufficient to trigger lysis by NK cells (Blom et al 1999 Smits et al1994) a sensitive and specific bioassay for loss of MHCI functionOX18 recognizes MHCI in Western blots and immunohistochem-istry from mouse nervous system samples (eg (Adelson et al2012 Corriveau et al 1998 Datwani et al 2009 Dixon-Salazar
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fig 1 Persistent multiple innervation at the MHCI-deficient NMJ (A) Confocal images of intact muscle stained with a-btx (red) and anti-neurofilament and anti-SV2 (green)Scales 10 lm Arrows mark distinct MN axons in P15 and adult (B) Multiple innervation in confocal images Multiple innervation at P7 631 plusmn 17 in b2mTAP (n = 6) vs585 plusmn 33 in WT (n = 8) p = 028 P15 b2mTAP n = 9 WT n = 8 adult (P29ndash50) n = 4 per genotype (values see text) (C) Time course of synapse elimination Multipleinnervation at P0 833 plusmn 31 in b2mTAP vs 902 plusmn 46 in WT p = 013 n = 4 animals per genotype (B) and (C) p = 00011 p = 00116 (D) Multiple innervation in oneyear old diaphragm b2mTAP n = 5 WT n = 3 p = 00069 (E) Multiple innervation in mice lacking classical MHCIs Kb n = 3 Db n = 6 Gray bars multipleinnervation (mean plusmn SEM) at the same age in b2mTAP (top) or WT (bottom) diaphragm from (B) p = 0002 p 6 00001
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 3
et al 2014 Goddard et al 2007 Huh et al 2000 Needleman et al2010)) Remarkably injected antibodies cause a nearly threefoldincrease in the retention of multiple inputs relative to IgG1-injected littermate controls (Fig 2) The quantitatively smallereffect of antibody injection compared to genetic MHCI deficiencyis not surprising for three reasons (1) antibody penetration maynot be complete resulting in only partial block of MHCI functionat the NMJ (2) antibody is delivered transiently and may be grad-ually cleared resulting in only a temporary block of MHCI functionover the week-long experiment and (3) antibody may not com-pletely block the function of all of the MHCI proteins whose cell-surface expression is reduced in genetically MHCI-deficient mice(see Section 4) The results of these antibody injections show thatinterfering with MHCI function over the second postnatal week sig-nificantly impairs establishment of monoinnervation in WT ani-mals Since MN cell death and synapse formation were complete
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
before MHCI function was blocked (Tapia et al 2012 Xu et al2006 Yamamoto and Henderson 1999) this finding further sup-ports the idea that increased multiple innervation in MHCI-deficient mice is not due to changes in synapse formation butrather reflects a disruption of the final stages of synapseelimination
23 Classical MHCIs promote synapse elimination at the developingNMJ
If MHCI promotes synapse elimination then overexpressingMHCI might accelerate synapse elimination To test this predictionwe compared multiple innervation at P7 when synapse elimina-tion is not yet complete in WT mice and in mice that overexpressthe mouse MHCI H-2Db under the control of the neuron-specificenolase promoter (NSE-Db mice (Rall et al 1995)) At P7 just over
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
A
controlantibody
0
12
6
3
9
B
m
ultip
ly in
nerv
ated
cont
rol
antib
ody
MH
CI
antib
ody
P15
MHCI antibody
C P7
WT NSE-Db
70
35
0
Fig 2 Anti-MHCI antibodies impair synapse elimination at the NMJ in WT mice while overexpressing MHCI in neurons accelerates synapse elimination (A) RepresentativeNMJs from OX18- and IgG1-injected WT animals at P15 nAChRs labeled with a-btx (red) presynaptic axons and terminals labeled with anti-neurofilament and anti-SV2(green) Arrows individual MN axons Scales 10 lm (B) Quantification of multiple innervation in IgG1- vs OX18-injected animals at P15 OX18 88 plusmn 17 IgG 30 plusmn 08p = 002 OX18 injection n = 9 animals from 4 litters IgG1 injection n = 6 animals from 3 litters (C) At P7 when synapse elimination is not yet complete there is a modestbut significant decrease in the percentage of muscle fibers that remain multiply-innervated in NSE-Db mice reflecting the fact that postsynaptic sites more rapidly achievemonoinnervation in MHCI-overexpressing animals WT n = 3 NSE-Db n = 5 p = 0012
4 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
half of postsynaptic sites remain multiply-innervated in WT ani-mals However a modestly but significantly lower percentage ofpostsynaptic sites remained multiply-innervated in MHCI-overexpressing animals (Fig 2C) The small magnitude of the accel-eration may reflect to fact that H-2Db expression is only modestlyincreased in these transgenics (Lee et al 2014 Rall et al 1995)Together these results demonstrate that developmental synapseelimination at the NMJ can be bi-directionally regulated bychanges in MHCI levels increasing MHCI accelerates synapse elim-ination while reducing MHCI disrupts synapse elimination
These results show that overexpressing just one member of thelarge MHCI gene family the classical MHCI H-2Db is sufficient toaccelerate synapse elimination at the NMJ To determine if classicalMHCIs are involved in synapse elimination at the developing NMJwe examined innervation in KbDb double mutant mice whichlack both of the classical MHCI genes found in C57Bl6 mice(Vugmeyster et al 1998) Both H-2Db and H-2Kb genes areexpressed at the developing NMJ (Tetruashvily et al in press) AtP15 the percentage of multiply-innervated postsynaptic siteswas three times WT levels in KbDb mice (multiple innerva-tion 106 plusmn 11 n = 5 p = 0001 relative to WT) This was not sig-nificantly different from b2mTAP values (p = 009)suggesting that loss of H-2K and H-2D could fully explain the initialsynapse elimination defects in b2mTAP mice In contrast lossof either H-2K or H-2D alone did not affect synapse elimination(Fig 1E) suggesting that only a single classical MHCI is necessaryfor normal synapse elimination to occur Thus three different mod-els that disrupt MHCI function (b2mTAP and KbDb miceand anti-MHCI function-blocking antibodies injected into WTmice) all cause multiple innervation to persist together stronglysuggesting that MHCI promotes synapse elimination at the devel-oping NMJ Furthermore results in NSE-Db and KbDb micesuggest that synapse elimination at the NMJ is promoted by classi-cal MHCI proteins
24 MHCI is expressed at the developing NMJ
MHCI protein is not thought to be expressed in uninjured mus-cle cells but has been detected in adult MNs at the NMJ (Thamset al 2009) mRNAs encoding several MHCI proteins have beendetected at the NMJ at multiple ages (Tetruashvily et al inpress) To determine if MHCI protein is present at the NMJ assynapse elimination is occurring we immunostained intact muscle
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
using a mouse monoclonal antibody (OX-18) that recognizes amonomorphic epitope of rat MHCI (RT-1A (Fukumoto et al1982)) and reacts with MHCI from rat mouse and primate brainin Western blots and immunohistochemistry (Corriveau et al1998 Datwani et al 2009 Dixon-Salazar et al 2014 Goddardet al 2007 Huh et al 2000 Needleman et al 2010 Rollekeet al 2006) Several lines of evidence support OX-18rsquos specificityfor MHCI in the mouse nervous system (see Section 4) UsingOX18 we detect specific MHCI labeling at the NMJ at P7 assynapse elimination is occurring (Fig S1A) This labeling is abol-ished by genetic destabilization of MHCI proteins (in b2mTAP
mice) or by replacing OX-18 with an isotype-control antibody(Fig S1) Whole-mount muscles were triple-labeled with OX18 tolabel MHCI a-btx to label postsynaptic nAChRs and anti-synaptophysin to visualize presynaptic MN terminals Becausepre- and post-synaptic structures are closely overlaid at the NMJthree-dimensional reconstruction of staining in confocal Z-stacksis necessary to unambiguously discriminate co-localization withpre- vs post-synaptic markers In P7 diaphragm MHCI co-localizes more extensively in individual optical sections with post-synaptic nAChRs than with presynaptic synaptophysin (Fig S1A)Specific MHCI labeling does not appear throughout the musclefiber but is restricted to the postsynaptic site Thus MHCI proteinis expressed at the NMJ at the time when developmental synapseelimination is occurring
25 Excess MN inputs can be recruited by graded stimulation in MHCI-deficient mice
Surplus MN inputs are functionally silenced before they arephysically removed (Colman et al 1997 Kopp et al 2000) Ourresults show that MHCI promotes synapse elimination but it isunclear if MHCI is involved in only the final physical removal ofsurplus axons or is involved in the earlier functional silencingTo discriminate between these possibilities we performed sharpelectrode recordings in P15 WT and MHCI-deficient animals andapplied graded stimulation to the phrenic nerve to sequentiallyrecruit MN inputs (Redfern 1970) Graded stimulation producedsingle-amplitude end plate potentials (EPPs) in 100 of recordingsfrom P15 WT animals as expected reflecting functional monoin-nervation of all postsynaptic sites at this age In MHCI-deficientanimals in contrast graded stimulation evoked a large stepwiseincrease in the EPP amplitude in 21 of recordings reflecting
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 5
recruitment of multiple MN inputs Multiple inputs could still berecruited at 13 of junctions in MHCI-deficient adults (Fig 3)These functional measures independently corroborate the anatom-ical evidence that synapse elimination is impaired in MHCI-deficient mice and show that some postsynaptic sites continueto receive inputs from MN axons that are distinct all the way tothe phrenic nerve typical of younger animals before synapse elim-ination is complete (Redfern 1970 Tapia et al 2012) Thus persis-tent multiple innervation in MHCI-deficient animals may resultfrom a failure to silence competing terminal branches rather thana failure to physically remove silent inputs
26 Increased synaptic strength at MHCI-deficient NMJs
Competing axons develop a disparity in synaptic strength andweaker inputs are eventually eliminated (Colman et al 1997Kopp et al 2000) Our anatomical and functional data supportthe idea that MHCI promotes silencing and elimination of MNsynapses In the adult hippocampus MHCI is required for long-term depression a form of persistent synaptic weakening (Huhet al 2000 Nelson et al 2013) To determine if MHCI is involvedin synapse weakening at the developing NMJ we measured synap-tic strength Quantal size is a measure of synaptic strength thatreflects the postsynaptic response to individual quanta of neuro-transmitter (Pennefather and Quastel 1981) At the NMJ miniatureend plate potential (MEPP) amplitude provides a reliable estimateof quantal size (Del Castillo and Katz 1954) MEPP amplitude issignificantly larger than WT in MHCI-deficient animals at P15(b2mTAP 178 plusmn 014 mV WT 115 plusmn 016 mV p = 0022)Quantal size decreases with age as previously observed(Wareham et al 1994) but remains larger in b2mTAP adults(P29-P45 b2mTAP 10 plusmn 008 mV WT 066 plusmn 006 mVp = 0004 Fig S3) Increased quantal size in MHCI-deficient NMJsis not associated with changes in input resistance EPP decay timeMEPP frequency paired-pulse facilitation (Fig S4) postsynapticnAChR density (Fig S2) or the number of quanta released perpresynaptic nerve impulse (quantal content) (Fig S4E) Togetherthese results show that loss of MHCI is associated with an earlyand persistent increase in synaptic strength Thus endogenousMHCI promotes synaptic weakening at the developing NMJ andmay contribute to the decline in synaptic strength that precedesfunctional and physical elimination of competing inputs duringnormal circuit development (Buffelli et al 2003 Colman et al1997 Kopp et al 2000)
27 MHCI levels rise in aged muscle
MHCI mRNA is up-regulated in aged MNs as they become sus-ceptible to denervation (Edstrom et al 2004) but is relativelylow in MNs that resist aging-related synapse loss (Edstrom et al2004 Linda et al 1999 Valdez et al 2012) MHCI promotessynapse elimination at the developing NMJ raising the possibilitythat higher levels of MHCI could be involved in aging-related mus-cle denervation However elevated mRNA levels do not guaranteeincreased transcribed protein levels and to our knowledge nostudies have assessed MHCI protein levels at the aging NMJ Todetermine if MHCI protein levels rise at the aging NMJ as denerva-tion is occurring we compared MHCI expression in extensor digi-torum longus (EDL) from P15 and 2-year-old mice using westernblotting The EDL was chosen because it shows significant denerva-tion at this age (Chai et al 2011 Valdez et al 2012) Becauseeither H2-K or H2-D are required for developmental synapse elim-ination (Fig 1E) we used an antibody (p8) that is specific for H2-KValues were normalized to a loading control (see Section 4) Com-paring MHCI expression in EDL from P15 vs 2-year-old mice wefound that H2-K levels rise significantly in aging EDL relative to
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
younger ages (Fig 4A) Thus MHCI protein is expressed at theNMJ during development and MHCI levels rise significantly atthe aging NMJ
28 MHCI promotes aging-related synapse loss
With aging synapses are eliminated frommature muscle fibersleaving some fibers fully denervated (Rudolf et al 2014 Valdezet al 2012) We find that during development MHCI promotessynapse elimination and MHCI levels rise in aging EDL raisingthe possibility that the increase in MHCI expression with age drivesaberrant synapse elimination in mature singly-innervated musclefibers To directly test if MHCI contributes to pathological synapseloss in aging muscle denervation of EDL muscle fibers wasassessed in aged (2-year-old) WT and b2mTAP mice Full orpartial denervation are evident when postsynaptic nAChRs(labeled with a-btx) are not completely apposed by the nerve ter-minal represented by the presynaptic marker SV2 (Valdez et al2010) As expected a subset of muscle fibers were completely den-ervated in aged EDL (122 plusmn 08 fully denervated n = 4) In con-trast both full and partial denervation was dramatically reducedin MHCI-deficient EDL (Fig 4 full denervation 45 plusmn 10p = 00009 relative to WT) While developmental synapse elimina-tion in the EDL was modestly impaired in MHCI-deficient animals(percent multiply-innervated in P15 EDL WT 10 plusmn 05 b2m-TAP 52 plusmn 11 n = 3 animals per age p = 00045) this differenceis not sufficient to fully explain the reduced denervation in MHCI-deficient EDL Thus the relative lack of denervation in aged MHCI-deficient animals cannot be fully explained by a different startingpoint for the aging process but instead reflects a degree of protec-tion from aging-related denervation Together these results showthat MHCI levels rise with aging at the NMJ and that reducingMHCI expression can ameliorate synapse loss in aging muscle
3 Discussion
Synapse elimination is essential for the establishment of matureneural circuits while synapse loss later in life is a significant causeof aging-associated pathology Here we show that MHCI promotesboth developmental synapse elimination and aging-relatedsynapse loss at the vertebrate NMJ MHCI is expressed at the devel-oping NMJ and three different manipulations that reduce MHCIfunction-genetic destabilization or deletion of MHCI proteins orinjection of function-blocking anti-MHCI antibodies-cause multi-ple innervation to persist at otherwise normally-developingsynapses Reducing MHCI expression does not affect patterns ofinnervation prior to the close of the first postnatal week suggest-ing it does not alter earlier synapse formation or elimination How-ever synapse elimination during the second week is significantlydisrupted preventing establishment of uniform mono-innervation Multiply-innervated postsynaptic sites are still pre-sent in MHCI-deficient animals at one year of age are evident bothanatomically and functionally and are associated with increasedquantal size Thus MHCI is required to complete the transition touniform mono-innervation in developing muscle and promotessynaptic weakening one of the earliest hallmarks of competitionamong multiple inputs to a single muscle fiber With aging MHCIlevels rise in motor neurons (Edstrom et al 2004) and in MHCI-deficient mice aging-related muscle denervation is markedlyreduced
Several experimental conditions delay synapse elimination atthe NMJ including changes in synaptic activity (Misgeld et al2002 Thompson et al 1979) NMDAR activation (Personiuset al 2008) GDNF (Nguyen et al 1998) BDNF or its receptor TrkB(Je et al 2013) NCAM (Rafuse et al 2000) glial neurofascin
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Adult
ββ2m--TAP--WT
A P15
15
0 5
10
2520
WT β2m--
TAP--
WT β2m--
TAP--
P15
m
ultip
ly in
nerv
ated
β2m--TAP--
0
15
5
10Adul
t
B
WT
Fig 3 Multiple functional inputs persist at MHCI-deficient NMJs (A) Representative EPPs recorded from P15 or adult (P29-45) muscle Scales P15 1 mV 15 ms adult10 mV 10 ms Stimulus artifacts blanked for clarity (B) Summed electrophysiological data shows increased multiple innervation in MHCI-deficient mice P15 WT 16 cellsfrom 8 animals b2mTAP 19 cells from 6 animals adult WT 25 cells from 5 animals b2mTAP 38 cells from 8 animals
6 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
(Roche et al 2014) or PKC theta (Li et al 2004) Synapse elimina-tion is also disrupted in mice which bear a dominant-negativemutation in the GARS gene a mouse model of Charcot-Marie-Tooth type 2D which includes apparent developmental arrest ofNMJ maturation (Sleigh et al 2014) Other manipulations includ-ing increasing activity reducing gap junctional coupling or loss ofthe presynaptic glycine transporter GlyT2 accelerate synapseelimination (Bogdanik et al 2012 OrsquoBrien et al 1978 Personiuset al 2007) These and other studies have provided fundamentalinsights into the molecular and cellular control of the rate ofsynapse elimination and suggest factors that may allow synapseelimination to occur normally prior to P7 in MHCI-deficient miceIn some of these models development of the NMJ is grossly dis-rupted which could contribute to their failure to undergo synapseelimination on a normal schedule While similar gross develop-mental disruption is not present at MHCI-deficient NMJs we can-not rule out the possibility that loss of MHCI perturbs synapseelimination indirectly through changes in aspects of developmentthat were not assessed With most manipulations however uni-form monoinnervation is still established within the first few post-natal weeks while multiple innervation is still evident at one yearof age in MHCI-deficient mice suggesting that synapse eliminationis persistently disrupted
We detect MHCI protein at the developing NMJ where it colo-calizes with nAChR labeling suggesting that it is expressed in themuscle fiber Importantly MHCI may also be expressed in themotor neuron andor Schwann cell during development andor inaging a possibility we cannot exclude because there are currentlyno antibodies to detect most members of the large MHCI familyMultiple MHCI mRNAs are expressed at the NMJ at different ages(Tetruashvily et al in press) MHCI protein has been detected inpresynaptic motor neurons in adult mice (Thams et al 2009)and in human and rat Schwann cells in vitro (Armati et al 1990Samuel et al 1987) Furthermore we find that modest overexpres-sion of the MHCI H2-D in neurons (Rall et al 1995) is sufficient tomodestly accelerate synapse elimination (Fig 2C) Ultimatelyimmuno-electron or super-resolution microscopy coupled withdevelopment of dozens of new antibodies will be necessary tofully characterize the pre- vs post-synaptic expression of the many
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
MHCI proteins Nevertheless the current results show that MHCI ispresent at the developing NMJ when synapse elimination isoccurring
MHCI is thought to be repressed by healthy muscle cells andupregulated only after injury or in disease states (Karpati et al1988) The current results demonstrate an unexpected role forMHCI at the healthy developing NMJ Remarkably MHCIrsquos functionat the developing NMJ is strikingly different to its role after injuryReducing MHCI expression exacerbates loss of synapses made ontoaxotomized motor neurons (Berg et al 2013 Oliveira et al 2004)while we find that both developmental and aging-related loss ofmotor neuron synapses onto muscles are inhibited
The bulk of excess MN synapses are pruned prior to P7 (Tapiaet al 2012) but mono-innervation is largely established betweenP7 and P15 as MNs begin to fire asynchronously (Buffelli et al2002 Personius et al 2001) In MHCI-deficient mice 15ndash20of postsynaptic sites retain two inputs at P15 No postsynaptic sitesreceived three or more inputs at P15 in MHCI-deficient animalsconsistent with the fact that very few postsynaptic sites stillreceive 3 or more inputs at P7 in WTs (Fig S2F) the age whenMHCIrsquos effects on synapse elimination begin to emerge The failureof circuit maturation in MHCI-deficient animals is not associatedwith gross motor defects in mice but could prevent the emergenceof fine-motor control that is behaviorally relevant in humansStrikingly the extent of the synapse elimination deficit at theMHCI-deficient NMJ is similar to the extent of axon pruning defi-cits in the lateral geniculate nucleus of mice lacking MHCI(Datwani et al 2009 Huh et al 2000 Lee et al 2014) or lackingneuron pentraxin (Bjartmar et al 2006) or components of thecomplement cascade (Stevens et al 2007) The reason for the par-tial nature of the pruning failure in the visual system in these mod-els remains unknown At the NMJ however we find that MHCI isspecifically required for the final stages of synapse eliminationduring the second postnatal week (Fig 1) In adult motor neuronslevels of the classical MHCI H-2D appear to be heterogeneous(Thams et al 2009) raising the possibility that different levels ofMHCI during development may also render some inputs more sus-ceptible to elimination Alternatively other factors including fibertype and associated activity patterns may influence the role that
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fullydenervated
15
10
5
0 β2m--
TAP--WT
3
2
1
0 β2m--
TAP--WT
E
Partially denervated
AChR
s (α
-btx
)
B C
SV2
5
4
D
mer
ge
A
MH
CI
P1540 80 160μg
MH
CI
2 Y O40 80 160μg
p
artia
lly d
ener
vate
d
fu
lly d
ener
vate
d
P15 2 Y OMH
CI l
abel
ing
in E
DL
(nor
mal
ized
to G
APD
H)
0
05025
07510
Fig 4 MHCI protein levels climb in aging EDL and reducing MHCI expression prevents aging-related denervation (A) Western blots for MHCI in lysates of EDL muscle fromP15 and 2-year-old mice Three different total protein concentrations are shown Blots were stripped and probed for GAPDH as a loading control Bottom MHCI densitometrynormalized to GAPDH for six technical replicates (two per concentration) at each age p = 00048 (B C) EDL muscles from 24ndash26 month-old mice were stained with a-btx(red) to detect nAChRs and SV2 (green) to detect presynaptic nerve terminals Representative images show examples of partially (arrows B) and fully (oval C) denervatedAChR clusters in WT EDL (red with no associated green in merge) Scales 7 lm (D E) Mean partial (D) and full (E) denervation in 2 year old EDL muscle from WT and MHCI-deficient b2mTAP mice (n = 4 per genotype) p = 003 p = 00009
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 7
MHCI plays in elimination of specific axon branches This is consis-tent with the finding that MHCI is required for synapse eliminationto reach completion in the diaphragm which consists of a mixtureof fast and slow fiber types but has a smaller influence on synapseelimination in the EDL which is almost exclusively composed offast-twitch fibers (Eddinger et al 1985) Together the currentresults suggest that synapse elimination at the NMJ may be dividedinto stages with distinct molecular requirements and that MHCI isessential as the last postsynaptic sites transition from dual- tomono-innervation
During synapse elimination at the NMJ competing axonsdevelop a disparity in synaptic strengthwhich precedeswithdrawalof the weaker input (Colman et al 1997 Kopp et al 2000) While itis unknown if larger synaptic strength at the MHCI-deficient NMJ isrelated to deficits in synapse elimination stronger inputs may beless vulnerable to competition-induced synaptic weakening In thisway loss of MHCI might prevent the development of sufficient dis-parity in synaptic strength to trigger functional silencing and phys-ical elimination in the most closely-matched competitions
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Interestingly paired increases in synapse strength and density likethose we observe at the MHCI-deficient NMJ are also seen in thecortex (Glynn et al 2011) and lateral geniculate nucleus (LGN)(Lee et al 2014) of MHCI-deficient mice Additional studies inMHCI-deficient mice found increases in synapse density in cortex(Adelson et al 2014) and increased synapse density (Dixon-Salazar et al 2014) and strength (Fourgeaud et al 2010) in hip-pocampus While the present study identifies a role for MHCI insynapse elimination at the NMJ it is as yet unknown if MHCI inhi-bits synapse formation andor promotes synapse elimination incortex or hippocampus
It will be important to determine if MHCI expression is regu-lated by activity at the developing NMJ as it is in the visual system(Corriveau et al 1998) to determine if it is specifically responsiveto the levels patterns andor synchronization of activity and tofully define the cellular and molecular networks linking MHCI tochanges in the strength and connectivity of synapses Animmunoreceptor for MHCI PirB has been detected in the CNS(Syken et al 2006) and at the adult NMJ (Thams et al 2009)
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

2 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
In adults responses to peripheral motor neuron injury are reg-ulated by specific immune proteins members of the major histo-compatibility complex class I (MHCI) MHCI has been detected inadult motor neurons (Edstrom et al 2004 Linda et al 1999Thams et al 2009) but adult muscle cells are thought to repressesMHCI genes except after injury or in disease states (Karpati et al1988) Consequently studies of the role of MHCI in the peripheralnervous system have focused almost exclusively on responses toinjury In injury models reducing MHCI expression exacerbatesloss of synapses made onto axotomized motor neuron cell bodiesin the spinal cord and impairs functional recovery (Berg et al2013 Oliveira et al 2004 Thams et al 2009) These and otherstudies have reinforced the view that at the NMJ MHCI is primarilyinvolved in responses to injury In the central nervous systemMHCI has been implicated in pruning of developing retinal projec-tions (Datwani et al 2009 Huh et al 2000 Lee et al 2014) whileelimination of developing cerebellar climbing fibers is MHCI-independent (Letellier et al 2008 McConnell et al 2009) Herewe explored if MHCI plays a role in developmental synapse elimi-nation or aging-related synapse loss at the vertebrate NMJ
2 Results
21 Impaired synapse elimination in mice deficient for cell-surfaceMHCI
MHCI genes are expressed at the NMJ as synapse elimination isoccurring (Tetruashvily et al in press) If MHCI promotes synapseelimination at the developing NMJ then reducing MHCI expressionshould increase the number of multiply-innervated muscle fibersTo test this prediction we visualized innervation of postsynapticsites in diaphragm muscle fibers from mice lacking stable cell-surface expression of most MHCI proteins (b2mTAP micesee Section 4) The diaphragm is a thin planar muscle with awell-organized end plate band that is particularly amenable toimaging in whole-mount preparations Postsynaptic nicotinicacetylcholine receptors (nAChRs) were labeled using tetramethylrhodamine-conjugated a-bungarotoxin (a-btx) and presynapticmotor neuron axons were visualized using antibodies against theaxon cytoskeletal element neurofilament and the presynaptic vesi-cle protein SV2 Because individual nAChR clusters may lay aboveone another in whole mount muscle 3D reconstructions of confo-cal Z-stacks were created for all suspected multiply-innervatedpostsynaptic sites and reconstructed stacks were viewed frommultiple depths and angles to determine if motor neuron brancheswere distinct and if they innervated a single nAChR cluster or twoclusters in different focal planes (see Section 4)
In WT mice nearly all postsynaptic sites are contacted by a sin-gle MN axon by postnatal day 15 (P15) as expected (Kopp et al2000 Redfern 1970) In contrast significantly more postsynapticsites remain multiply-innervated in mice lacking cell-surface MHCI(Fig 1AndashC Multiple innervation at P15 b2mTAP 164 plusmn 22WT 35 plusmn 11 p = 00011) Multiply-innervated postsynaptic sitesdetected in P15 MHCI-deficient mice all received two inputsAlthough multiple innervation persists b2mTAP mice are ofnormal size and weight and eye opening fur growth weaningand reproduction all occur at appropriate ages Development ofthe NMJ is also otherwise outwardly normal Every muscle fibercontains only a single postsynaptic nAChR cluster at P15 and thearea perimeter and intensity of nAChR labeling and morphologyof the end plate band are all normal in b2mTAP mice(Fig S2) Furthermore the distribution of nAChR labeling changesas expected from plaque-like to pretzel-like as the animalsmatured Despite this maturation multiple innervation persistsin MHCI-deficient adults (Fig 1AndashC b2mTAP 186 plusmn 43WT 23 plusmn 17 p = 00116) and remarkably is still present at one
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
year of age (Fig 1D b2mTAP 95 plusmn 13 WT 25 plusmn 05p = 00069) Thus loss of MHCI prevents multiple innervation fromfully resolving at the NMJ
Loss of MHCI could cause multiply-innervated muscle fibers topersist because it directly or indirectly disrupts synapse elimina-tion or because it increases synapse formation via changes in neu-rogenesis programmed cell death or motor neuron outgrowth orbranching However several lines of evidence suggest that changesin synapse formation do not underlie the persistent multiple inner-vation in MHCI-deficient animals At birth (P0) and at P7 when MNprogrammed cell death and synapse formation are largely com-plete (Tapia et al 2012 Xu et al 2006 Yamamoto andHenderson 1999) a normal proportion of postsynaptic sites aremultiply-innervated in MHCI-deficient mice (Fig 1AndashC) Further-more at P7 each muscle fiber is contacted by the same averagenumber of axons in WT and MHCI-deficient mice (Fig S2E andF) Thus measures that are sensitive to changes in motor neuronand muscle fiber number and synapse formation are normal inMHCI-deficient mice These results suggest that in the absence ofMHCI patterns of innervation are established normally but inner-vation fails to decline over the second postnatal week consistentwith a disruption of developmental pruning
However it is still possible that loss of MHCI prevents theestablishment of monoinnervation without changes in synapseelimination eg by prolonging neurogenesis programmed celldeath andor synapse formation well past their normaldevelopmental window If synapse elimination is not impaired inMHCI-deficient mice then ongoing synapse formation would benecessary to explain the stable synapse density from P7 onwardsHowever we did not see evidence of ongoing axon sprouting out-growth or synapse formation over the second postnatal weekTogether these results suggest that changes in synapse formationare unlikely to contribute significantly to persistent multiple inner-vation in MHCI-deficient animals Instead they are more consis-tent with a role for MHCI in allowing synapse elimination toreach completion
Extensive synapse elimination occurs at the NMJ prior to theclose of the first postnatal week (Tapia et al 2012) This initialsynapse elimination does not require MHCI because at P7 the per-centage of postsynaptic sites that remain multiply-innervated(Fig 1) and the extent of MN convergence onto individual postsy-naptic sites (Fig S2E and F) are both normal in MHCI-deficientmice Consistent with the fact that early synapse elimination hap-pens normally MHCI-deficient mice do not display gross motordeficits (eg weakness inactivity or tremor) (McConnell et al2009 Nelson et al 2013) Over the second postnatal week how-ever far fewer postsynaptic sites achieve mono-innervation inMHCI-deficient animals a failure of circuit maturation that coulddisrupt the emergence of fine motor control Remarkably there isno further decline in multiply-innervated postsynaptic sitesbetween P15 and adulthood (Fig 1B and C)
22 Loss of MHCI impairs synapse elimination specifically during thesecond postnatal week
If MHCI is specifically involved in synapse elimination during thesecond postnatal week then acutely disrupting MHCI function dur-ing the second postnatal week should causemultiple innervation topersist To test this hypothesiswe inhibitedMHCI function fromP7-P15 using an anti-MHCI antibody (OX18) that is function-blockinginmultiple assays Incubating hepatocytes or tumor cells in OX18 issufficient to trigger lysis by NK cells (Blom et al 1999 Smits et al1994) a sensitive and specific bioassay for loss of MHCI functionOX18 recognizes MHCI in Western blots and immunohistochem-istry from mouse nervous system samples (eg (Adelson et al2012 Corriveau et al 1998 Datwani et al 2009 Dixon-Salazar
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fig 1 Persistent multiple innervation at the MHCI-deficient NMJ (A) Confocal images of intact muscle stained with a-btx (red) and anti-neurofilament and anti-SV2 (green)Scales 10 lm Arrows mark distinct MN axons in P15 and adult (B) Multiple innervation in confocal images Multiple innervation at P7 631 plusmn 17 in b2mTAP (n = 6) vs585 plusmn 33 in WT (n = 8) p = 028 P15 b2mTAP n = 9 WT n = 8 adult (P29ndash50) n = 4 per genotype (values see text) (C) Time course of synapse elimination Multipleinnervation at P0 833 plusmn 31 in b2mTAP vs 902 plusmn 46 in WT p = 013 n = 4 animals per genotype (B) and (C) p = 00011 p = 00116 (D) Multiple innervation in oneyear old diaphragm b2mTAP n = 5 WT n = 3 p = 00069 (E) Multiple innervation in mice lacking classical MHCIs Kb n = 3 Db n = 6 Gray bars multipleinnervation (mean plusmn SEM) at the same age in b2mTAP (top) or WT (bottom) diaphragm from (B) p = 0002 p 6 00001
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 3
et al 2014 Goddard et al 2007 Huh et al 2000 Needleman et al2010)) Remarkably injected antibodies cause a nearly threefoldincrease in the retention of multiple inputs relative to IgG1-injected littermate controls (Fig 2) The quantitatively smallereffect of antibody injection compared to genetic MHCI deficiencyis not surprising for three reasons (1) antibody penetration maynot be complete resulting in only partial block of MHCI functionat the NMJ (2) antibody is delivered transiently and may be grad-ually cleared resulting in only a temporary block of MHCI functionover the week-long experiment and (3) antibody may not com-pletely block the function of all of the MHCI proteins whose cell-surface expression is reduced in genetically MHCI-deficient mice(see Section 4) The results of these antibody injections show thatinterfering with MHCI function over the second postnatal week sig-nificantly impairs establishment of monoinnervation in WT ani-mals Since MN cell death and synapse formation were complete
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
before MHCI function was blocked (Tapia et al 2012 Xu et al2006 Yamamoto and Henderson 1999) this finding further sup-ports the idea that increased multiple innervation in MHCI-deficient mice is not due to changes in synapse formation butrather reflects a disruption of the final stages of synapseelimination
23 Classical MHCIs promote synapse elimination at the developingNMJ
If MHCI promotes synapse elimination then overexpressingMHCI might accelerate synapse elimination To test this predictionwe compared multiple innervation at P7 when synapse elimina-tion is not yet complete in WT mice and in mice that overexpressthe mouse MHCI H-2Db under the control of the neuron-specificenolase promoter (NSE-Db mice (Rall et al 1995)) At P7 just over
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
A
controlantibody
0
12
6
3
9
B
m
ultip
ly in
nerv
ated
cont
rol
antib
ody
MH
CI
antib
ody
P15
MHCI antibody
C P7
WT NSE-Db
70
35
0
Fig 2 Anti-MHCI antibodies impair synapse elimination at the NMJ in WT mice while overexpressing MHCI in neurons accelerates synapse elimination (A) RepresentativeNMJs from OX18- and IgG1-injected WT animals at P15 nAChRs labeled with a-btx (red) presynaptic axons and terminals labeled with anti-neurofilament and anti-SV2(green) Arrows individual MN axons Scales 10 lm (B) Quantification of multiple innervation in IgG1- vs OX18-injected animals at P15 OX18 88 plusmn 17 IgG 30 plusmn 08p = 002 OX18 injection n = 9 animals from 4 litters IgG1 injection n = 6 animals from 3 litters (C) At P7 when synapse elimination is not yet complete there is a modestbut significant decrease in the percentage of muscle fibers that remain multiply-innervated in NSE-Db mice reflecting the fact that postsynaptic sites more rapidly achievemonoinnervation in MHCI-overexpressing animals WT n = 3 NSE-Db n = 5 p = 0012
4 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
half of postsynaptic sites remain multiply-innervated in WT ani-mals However a modestly but significantly lower percentage ofpostsynaptic sites remained multiply-innervated in MHCI-overexpressing animals (Fig 2C) The small magnitude of the accel-eration may reflect to fact that H-2Db expression is only modestlyincreased in these transgenics (Lee et al 2014 Rall et al 1995)Together these results demonstrate that developmental synapseelimination at the NMJ can be bi-directionally regulated bychanges in MHCI levels increasing MHCI accelerates synapse elim-ination while reducing MHCI disrupts synapse elimination
These results show that overexpressing just one member of thelarge MHCI gene family the classical MHCI H-2Db is sufficient toaccelerate synapse elimination at the NMJ To determine if classicalMHCIs are involved in synapse elimination at the developing NMJwe examined innervation in KbDb double mutant mice whichlack both of the classical MHCI genes found in C57Bl6 mice(Vugmeyster et al 1998) Both H-2Db and H-2Kb genes areexpressed at the developing NMJ (Tetruashvily et al in press) AtP15 the percentage of multiply-innervated postsynaptic siteswas three times WT levels in KbDb mice (multiple innerva-tion 106 plusmn 11 n = 5 p = 0001 relative to WT) This was not sig-nificantly different from b2mTAP values (p = 009)suggesting that loss of H-2K and H-2D could fully explain the initialsynapse elimination defects in b2mTAP mice In contrast lossof either H-2K or H-2D alone did not affect synapse elimination(Fig 1E) suggesting that only a single classical MHCI is necessaryfor normal synapse elimination to occur Thus three different mod-els that disrupt MHCI function (b2mTAP and KbDb miceand anti-MHCI function-blocking antibodies injected into WTmice) all cause multiple innervation to persist together stronglysuggesting that MHCI promotes synapse elimination at the devel-oping NMJ Furthermore results in NSE-Db and KbDb micesuggest that synapse elimination at the NMJ is promoted by classi-cal MHCI proteins
24 MHCI is expressed at the developing NMJ
MHCI protein is not thought to be expressed in uninjured mus-cle cells but has been detected in adult MNs at the NMJ (Thamset al 2009) mRNAs encoding several MHCI proteins have beendetected at the NMJ at multiple ages (Tetruashvily et al inpress) To determine if MHCI protein is present at the NMJ assynapse elimination is occurring we immunostained intact muscle
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
using a mouse monoclonal antibody (OX-18) that recognizes amonomorphic epitope of rat MHCI (RT-1A (Fukumoto et al1982)) and reacts with MHCI from rat mouse and primate brainin Western blots and immunohistochemistry (Corriveau et al1998 Datwani et al 2009 Dixon-Salazar et al 2014 Goddardet al 2007 Huh et al 2000 Needleman et al 2010 Rollekeet al 2006) Several lines of evidence support OX-18rsquos specificityfor MHCI in the mouse nervous system (see Section 4) UsingOX18 we detect specific MHCI labeling at the NMJ at P7 assynapse elimination is occurring (Fig S1A) This labeling is abol-ished by genetic destabilization of MHCI proteins (in b2mTAP
mice) or by replacing OX-18 with an isotype-control antibody(Fig S1) Whole-mount muscles were triple-labeled with OX18 tolabel MHCI a-btx to label postsynaptic nAChRs and anti-synaptophysin to visualize presynaptic MN terminals Becausepre- and post-synaptic structures are closely overlaid at the NMJthree-dimensional reconstruction of staining in confocal Z-stacksis necessary to unambiguously discriminate co-localization withpre- vs post-synaptic markers In P7 diaphragm MHCI co-localizes more extensively in individual optical sections with post-synaptic nAChRs than with presynaptic synaptophysin (Fig S1A)Specific MHCI labeling does not appear throughout the musclefiber but is restricted to the postsynaptic site Thus MHCI proteinis expressed at the NMJ at the time when developmental synapseelimination is occurring
25 Excess MN inputs can be recruited by graded stimulation in MHCI-deficient mice
Surplus MN inputs are functionally silenced before they arephysically removed (Colman et al 1997 Kopp et al 2000) Ourresults show that MHCI promotes synapse elimination but it isunclear if MHCI is involved in only the final physical removal ofsurplus axons or is involved in the earlier functional silencingTo discriminate between these possibilities we performed sharpelectrode recordings in P15 WT and MHCI-deficient animals andapplied graded stimulation to the phrenic nerve to sequentiallyrecruit MN inputs (Redfern 1970) Graded stimulation producedsingle-amplitude end plate potentials (EPPs) in 100 of recordingsfrom P15 WT animals as expected reflecting functional monoin-nervation of all postsynaptic sites at this age In MHCI-deficientanimals in contrast graded stimulation evoked a large stepwiseincrease in the EPP amplitude in 21 of recordings reflecting
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 5
recruitment of multiple MN inputs Multiple inputs could still berecruited at 13 of junctions in MHCI-deficient adults (Fig 3)These functional measures independently corroborate the anatom-ical evidence that synapse elimination is impaired in MHCI-deficient mice and show that some postsynaptic sites continueto receive inputs from MN axons that are distinct all the way tothe phrenic nerve typical of younger animals before synapse elim-ination is complete (Redfern 1970 Tapia et al 2012) Thus persis-tent multiple innervation in MHCI-deficient animals may resultfrom a failure to silence competing terminal branches rather thana failure to physically remove silent inputs
26 Increased synaptic strength at MHCI-deficient NMJs
Competing axons develop a disparity in synaptic strength andweaker inputs are eventually eliminated (Colman et al 1997Kopp et al 2000) Our anatomical and functional data supportthe idea that MHCI promotes silencing and elimination of MNsynapses In the adult hippocampus MHCI is required for long-term depression a form of persistent synaptic weakening (Huhet al 2000 Nelson et al 2013) To determine if MHCI is involvedin synapse weakening at the developing NMJ we measured synap-tic strength Quantal size is a measure of synaptic strength thatreflects the postsynaptic response to individual quanta of neuro-transmitter (Pennefather and Quastel 1981) At the NMJ miniatureend plate potential (MEPP) amplitude provides a reliable estimateof quantal size (Del Castillo and Katz 1954) MEPP amplitude issignificantly larger than WT in MHCI-deficient animals at P15(b2mTAP 178 plusmn 014 mV WT 115 plusmn 016 mV p = 0022)Quantal size decreases with age as previously observed(Wareham et al 1994) but remains larger in b2mTAP adults(P29-P45 b2mTAP 10 plusmn 008 mV WT 066 plusmn 006 mVp = 0004 Fig S3) Increased quantal size in MHCI-deficient NMJsis not associated with changes in input resistance EPP decay timeMEPP frequency paired-pulse facilitation (Fig S4) postsynapticnAChR density (Fig S2) or the number of quanta released perpresynaptic nerve impulse (quantal content) (Fig S4E) Togetherthese results show that loss of MHCI is associated with an earlyand persistent increase in synaptic strength Thus endogenousMHCI promotes synaptic weakening at the developing NMJ andmay contribute to the decline in synaptic strength that precedesfunctional and physical elimination of competing inputs duringnormal circuit development (Buffelli et al 2003 Colman et al1997 Kopp et al 2000)
27 MHCI levels rise in aged muscle
MHCI mRNA is up-regulated in aged MNs as they become sus-ceptible to denervation (Edstrom et al 2004) but is relativelylow in MNs that resist aging-related synapse loss (Edstrom et al2004 Linda et al 1999 Valdez et al 2012) MHCI promotessynapse elimination at the developing NMJ raising the possibilitythat higher levels of MHCI could be involved in aging-related mus-cle denervation However elevated mRNA levels do not guaranteeincreased transcribed protein levels and to our knowledge nostudies have assessed MHCI protein levels at the aging NMJ Todetermine if MHCI protein levels rise at the aging NMJ as denerva-tion is occurring we compared MHCI expression in extensor digi-torum longus (EDL) from P15 and 2-year-old mice using westernblotting The EDL was chosen because it shows significant denerva-tion at this age (Chai et al 2011 Valdez et al 2012) Becauseeither H2-K or H2-D are required for developmental synapse elim-ination (Fig 1E) we used an antibody (p8) that is specific for H2-KValues were normalized to a loading control (see Section 4) Com-paring MHCI expression in EDL from P15 vs 2-year-old mice wefound that H2-K levels rise significantly in aging EDL relative to
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
younger ages (Fig 4A) Thus MHCI protein is expressed at theNMJ during development and MHCI levels rise significantly atthe aging NMJ
28 MHCI promotes aging-related synapse loss
With aging synapses are eliminated frommature muscle fibersleaving some fibers fully denervated (Rudolf et al 2014 Valdezet al 2012) We find that during development MHCI promotessynapse elimination and MHCI levels rise in aging EDL raisingthe possibility that the increase in MHCI expression with age drivesaberrant synapse elimination in mature singly-innervated musclefibers To directly test if MHCI contributes to pathological synapseloss in aging muscle denervation of EDL muscle fibers wasassessed in aged (2-year-old) WT and b2mTAP mice Full orpartial denervation are evident when postsynaptic nAChRs(labeled with a-btx) are not completely apposed by the nerve ter-minal represented by the presynaptic marker SV2 (Valdez et al2010) As expected a subset of muscle fibers were completely den-ervated in aged EDL (122 plusmn 08 fully denervated n = 4) In con-trast both full and partial denervation was dramatically reducedin MHCI-deficient EDL (Fig 4 full denervation 45 plusmn 10p = 00009 relative to WT) While developmental synapse elimina-tion in the EDL was modestly impaired in MHCI-deficient animals(percent multiply-innervated in P15 EDL WT 10 plusmn 05 b2m-TAP 52 plusmn 11 n = 3 animals per age p = 00045) this differenceis not sufficient to fully explain the reduced denervation in MHCI-deficient EDL Thus the relative lack of denervation in aged MHCI-deficient animals cannot be fully explained by a different startingpoint for the aging process but instead reflects a degree of protec-tion from aging-related denervation Together these results showthat MHCI levels rise with aging at the NMJ and that reducingMHCI expression can ameliorate synapse loss in aging muscle
3 Discussion
Synapse elimination is essential for the establishment of matureneural circuits while synapse loss later in life is a significant causeof aging-associated pathology Here we show that MHCI promotesboth developmental synapse elimination and aging-relatedsynapse loss at the vertebrate NMJ MHCI is expressed at the devel-oping NMJ and three different manipulations that reduce MHCIfunction-genetic destabilization or deletion of MHCI proteins orinjection of function-blocking anti-MHCI antibodies-cause multi-ple innervation to persist at otherwise normally-developingsynapses Reducing MHCI expression does not affect patterns ofinnervation prior to the close of the first postnatal week suggest-ing it does not alter earlier synapse formation or elimination How-ever synapse elimination during the second week is significantlydisrupted preventing establishment of uniform mono-innervation Multiply-innervated postsynaptic sites are still pre-sent in MHCI-deficient animals at one year of age are evident bothanatomically and functionally and are associated with increasedquantal size Thus MHCI is required to complete the transition touniform mono-innervation in developing muscle and promotessynaptic weakening one of the earliest hallmarks of competitionamong multiple inputs to a single muscle fiber With aging MHCIlevels rise in motor neurons (Edstrom et al 2004) and in MHCI-deficient mice aging-related muscle denervation is markedlyreduced
Several experimental conditions delay synapse elimination atthe NMJ including changes in synaptic activity (Misgeld et al2002 Thompson et al 1979) NMDAR activation (Personiuset al 2008) GDNF (Nguyen et al 1998) BDNF or its receptor TrkB(Je et al 2013) NCAM (Rafuse et al 2000) glial neurofascin
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Adult
ββ2m--TAP--WT
A P15
15
0 5
10
2520
WT β2m--
TAP--
WT β2m--
TAP--
P15
m
ultip
ly in
nerv
ated
β2m--TAP--
0
15
5
10Adul
t
B
WT
Fig 3 Multiple functional inputs persist at MHCI-deficient NMJs (A) Representative EPPs recorded from P15 or adult (P29-45) muscle Scales P15 1 mV 15 ms adult10 mV 10 ms Stimulus artifacts blanked for clarity (B) Summed electrophysiological data shows increased multiple innervation in MHCI-deficient mice P15 WT 16 cellsfrom 8 animals b2mTAP 19 cells from 6 animals adult WT 25 cells from 5 animals b2mTAP 38 cells from 8 animals
6 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
(Roche et al 2014) or PKC theta (Li et al 2004) Synapse elimina-tion is also disrupted in mice which bear a dominant-negativemutation in the GARS gene a mouse model of Charcot-Marie-Tooth type 2D which includes apparent developmental arrest ofNMJ maturation (Sleigh et al 2014) Other manipulations includ-ing increasing activity reducing gap junctional coupling or loss ofthe presynaptic glycine transporter GlyT2 accelerate synapseelimination (Bogdanik et al 2012 OrsquoBrien et al 1978 Personiuset al 2007) These and other studies have provided fundamentalinsights into the molecular and cellular control of the rate ofsynapse elimination and suggest factors that may allow synapseelimination to occur normally prior to P7 in MHCI-deficient miceIn some of these models development of the NMJ is grossly dis-rupted which could contribute to their failure to undergo synapseelimination on a normal schedule While similar gross develop-mental disruption is not present at MHCI-deficient NMJs we can-not rule out the possibility that loss of MHCI perturbs synapseelimination indirectly through changes in aspects of developmentthat were not assessed With most manipulations however uni-form monoinnervation is still established within the first few post-natal weeks while multiple innervation is still evident at one yearof age in MHCI-deficient mice suggesting that synapse eliminationis persistently disrupted
We detect MHCI protein at the developing NMJ where it colo-calizes with nAChR labeling suggesting that it is expressed in themuscle fiber Importantly MHCI may also be expressed in themotor neuron andor Schwann cell during development andor inaging a possibility we cannot exclude because there are currentlyno antibodies to detect most members of the large MHCI familyMultiple MHCI mRNAs are expressed at the NMJ at different ages(Tetruashvily et al in press) MHCI protein has been detected inpresynaptic motor neurons in adult mice (Thams et al 2009)and in human and rat Schwann cells in vitro (Armati et al 1990Samuel et al 1987) Furthermore we find that modest overexpres-sion of the MHCI H2-D in neurons (Rall et al 1995) is sufficient tomodestly accelerate synapse elimination (Fig 2C) Ultimatelyimmuno-electron or super-resolution microscopy coupled withdevelopment of dozens of new antibodies will be necessary tofully characterize the pre- vs post-synaptic expression of the many
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
MHCI proteins Nevertheless the current results show that MHCI ispresent at the developing NMJ when synapse elimination isoccurring
MHCI is thought to be repressed by healthy muscle cells andupregulated only after injury or in disease states (Karpati et al1988) The current results demonstrate an unexpected role forMHCI at the healthy developing NMJ Remarkably MHCIrsquos functionat the developing NMJ is strikingly different to its role after injuryReducing MHCI expression exacerbates loss of synapses made ontoaxotomized motor neurons (Berg et al 2013 Oliveira et al 2004)while we find that both developmental and aging-related loss ofmotor neuron synapses onto muscles are inhibited
The bulk of excess MN synapses are pruned prior to P7 (Tapiaet al 2012) but mono-innervation is largely established betweenP7 and P15 as MNs begin to fire asynchronously (Buffelli et al2002 Personius et al 2001) In MHCI-deficient mice 15ndash20of postsynaptic sites retain two inputs at P15 No postsynaptic sitesreceived three or more inputs at P15 in MHCI-deficient animalsconsistent with the fact that very few postsynaptic sites stillreceive 3 or more inputs at P7 in WTs (Fig S2F) the age whenMHCIrsquos effects on synapse elimination begin to emerge The failureof circuit maturation in MHCI-deficient animals is not associatedwith gross motor defects in mice but could prevent the emergenceof fine-motor control that is behaviorally relevant in humansStrikingly the extent of the synapse elimination deficit at theMHCI-deficient NMJ is similar to the extent of axon pruning defi-cits in the lateral geniculate nucleus of mice lacking MHCI(Datwani et al 2009 Huh et al 2000 Lee et al 2014) or lackingneuron pentraxin (Bjartmar et al 2006) or components of thecomplement cascade (Stevens et al 2007) The reason for the par-tial nature of the pruning failure in the visual system in these mod-els remains unknown At the NMJ however we find that MHCI isspecifically required for the final stages of synapse eliminationduring the second postnatal week (Fig 1) In adult motor neuronslevels of the classical MHCI H-2D appear to be heterogeneous(Thams et al 2009) raising the possibility that different levels ofMHCI during development may also render some inputs more sus-ceptible to elimination Alternatively other factors including fibertype and associated activity patterns may influence the role that
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fullydenervated
15
10
5
0 β2m--
TAP--WT
3
2
1
0 β2m--
TAP--WT
E
Partially denervated
AChR
s (α
-btx
)
B C
SV2
5
4
D
mer
ge
A
MH
CI
P1540 80 160μg
MH
CI
2 Y O40 80 160μg
p
artia
lly d
ener
vate
d
fu
lly d
ener
vate
d
P15 2 Y OMH
CI l
abel
ing
in E
DL
(nor
mal
ized
to G
APD
H)
0
05025
07510
Fig 4 MHCI protein levels climb in aging EDL and reducing MHCI expression prevents aging-related denervation (A) Western blots for MHCI in lysates of EDL muscle fromP15 and 2-year-old mice Three different total protein concentrations are shown Blots were stripped and probed for GAPDH as a loading control Bottom MHCI densitometrynormalized to GAPDH for six technical replicates (two per concentration) at each age p = 00048 (B C) EDL muscles from 24ndash26 month-old mice were stained with a-btx(red) to detect nAChRs and SV2 (green) to detect presynaptic nerve terminals Representative images show examples of partially (arrows B) and fully (oval C) denervatedAChR clusters in WT EDL (red with no associated green in merge) Scales 7 lm (D E) Mean partial (D) and full (E) denervation in 2 year old EDL muscle from WT and MHCI-deficient b2mTAP mice (n = 4 per genotype) p = 003 p = 00009
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 7
MHCI plays in elimination of specific axon branches This is consis-tent with the finding that MHCI is required for synapse eliminationto reach completion in the diaphragm which consists of a mixtureof fast and slow fiber types but has a smaller influence on synapseelimination in the EDL which is almost exclusively composed offast-twitch fibers (Eddinger et al 1985) Together the currentresults suggest that synapse elimination at the NMJ may be dividedinto stages with distinct molecular requirements and that MHCI isessential as the last postsynaptic sites transition from dual- tomono-innervation
During synapse elimination at the NMJ competing axonsdevelop a disparity in synaptic strengthwhich precedeswithdrawalof the weaker input (Colman et al 1997 Kopp et al 2000) While itis unknown if larger synaptic strength at the MHCI-deficient NMJ isrelated to deficits in synapse elimination stronger inputs may beless vulnerable to competition-induced synaptic weakening In thisway loss of MHCI might prevent the development of sufficient dis-parity in synaptic strength to trigger functional silencing and phys-ical elimination in the most closely-matched competitions
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Interestingly paired increases in synapse strength and density likethose we observe at the MHCI-deficient NMJ are also seen in thecortex (Glynn et al 2011) and lateral geniculate nucleus (LGN)(Lee et al 2014) of MHCI-deficient mice Additional studies inMHCI-deficient mice found increases in synapse density in cortex(Adelson et al 2014) and increased synapse density (Dixon-Salazar et al 2014) and strength (Fourgeaud et al 2010) in hip-pocampus While the present study identifies a role for MHCI insynapse elimination at the NMJ it is as yet unknown if MHCI inhi-bits synapse formation andor promotes synapse elimination incortex or hippocampus
It will be important to determine if MHCI expression is regu-lated by activity at the developing NMJ as it is in the visual system(Corriveau et al 1998) to determine if it is specifically responsiveto the levels patterns andor synchronization of activity and tofully define the cellular and molecular networks linking MHCI tochanges in the strength and connectivity of synapses Animmunoreceptor for MHCI PirB has been detected in the CNS(Syken et al 2006) and at the adult NMJ (Thams et al 2009)
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

Fig 1 Persistent multiple innervation at the MHCI-deficient NMJ (A) Confocal images of intact muscle stained with a-btx (red) and anti-neurofilament and anti-SV2 (green)Scales 10 lm Arrows mark distinct MN axons in P15 and adult (B) Multiple innervation in confocal images Multiple innervation at P7 631 plusmn 17 in b2mTAP (n = 6) vs585 plusmn 33 in WT (n = 8) p = 028 P15 b2mTAP n = 9 WT n = 8 adult (P29ndash50) n = 4 per genotype (values see text) (C) Time course of synapse elimination Multipleinnervation at P0 833 plusmn 31 in b2mTAP vs 902 plusmn 46 in WT p = 013 n = 4 animals per genotype (B) and (C) p = 00011 p = 00116 (D) Multiple innervation in oneyear old diaphragm b2mTAP n = 5 WT n = 3 p = 00069 (E) Multiple innervation in mice lacking classical MHCIs Kb n = 3 Db n = 6 Gray bars multipleinnervation (mean plusmn SEM) at the same age in b2mTAP (top) or WT (bottom) diaphragm from (B) p = 0002 p 6 00001
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 3
et al 2014 Goddard et al 2007 Huh et al 2000 Needleman et al2010)) Remarkably injected antibodies cause a nearly threefoldincrease in the retention of multiple inputs relative to IgG1-injected littermate controls (Fig 2) The quantitatively smallereffect of antibody injection compared to genetic MHCI deficiencyis not surprising for three reasons (1) antibody penetration maynot be complete resulting in only partial block of MHCI functionat the NMJ (2) antibody is delivered transiently and may be grad-ually cleared resulting in only a temporary block of MHCI functionover the week-long experiment and (3) antibody may not com-pletely block the function of all of the MHCI proteins whose cell-surface expression is reduced in genetically MHCI-deficient mice(see Section 4) The results of these antibody injections show thatinterfering with MHCI function over the second postnatal week sig-nificantly impairs establishment of monoinnervation in WT ani-mals Since MN cell death and synapse formation were complete
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
before MHCI function was blocked (Tapia et al 2012 Xu et al2006 Yamamoto and Henderson 1999) this finding further sup-ports the idea that increased multiple innervation in MHCI-deficient mice is not due to changes in synapse formation butrather reflects a disruption of the final stages of synapseelimination
23 Classical MHCIs promote synapse elimination at the developingNMJ
If MHCI promotes synapse elimination then overexpressingMHCI might accelerate synapse elimination To test this predictionwe compared multiple innervation at P7 when synapse elimina-tion is not yet complete in WT mice and in mice that overexpressthe mouse MHCI H-2Db under the control of the neuron-specificenolase promoter (NSE-Db mice (Rall et al 1995)) At P7 just over
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
A
controlantibody
0
12
6
3
9
B
m
ultip
ly in
nerv
ated
cont
rol
antib
ody
MH
CI
antib
ody
P15
MHCI antibody
C P7
WT NSE-Db
70
35
0
Fig 2 Anti-MHCI antibodies impair synapse elimination at the NMJ in WT mice while overexpressing MHCI in neurons accelerates synapse elimination (A) RepresentativeNMJs from OX18- and IgG1-injected WT animals at P15 nAChRs labeled with a-btx (red) presynaptic axons and terminals labeled with anti-neurofilament and anti-SV2(green) Arrows individual MN axons Scales 10 lm (B) Quantification of multiple innervation in IgG1- vs OX18-injected animals at P15 OX18 88 plusmn 17 IgG 30 plusmn 08p = 002 OX18 injection n = 9 animals from 4 litters IgG1 injection n = 6 animals from 3 litters (C) At P7 when synapse elimination is not yet complete there is a modestbut significant decrease in the percentage of muscle fibers that remain multiply-innervated in NSE-Db mice reflecting the fact that postsynaptic sites more rapidly achievemonoinnervation in MHCI-overexpressing animals WT n = 3 NSE-Db n = 5 p = 0012
4 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
half of postsynaptic sites remain multiply-innervated in WT ani-mals However a modestly but significantly lower percentage ofpostsynaptic sites remained multiply-innervated in MHCI-overexpressing animals (Fig 2C) The small magnitude of the accel-eration may reflect to fact that H-2Db expression is only modestlyincreased in these transgenics (Lee et al 2014 Rall et al 1995)Together these results demonstrate that developmental synapseelimination at the NMJ can be bi-directionally regulated bychanges in MHCI levels increasing MHCI accelerates synapse elim-ination while reducing MHCI disrupts synapse elimination
These results show that overexpressing just one member of thelarge MHCI gene family the classical MHCI H-2Db is sufficient toaccelerate synapse elimination at the NMJ To determine if classicalMHCIs are involved in synapse elimination at the developing NMJwe examined innervation in KbDb double mutant mice whichlack both of the classical MHCI genes found in C57Bl6 mice(Vugmeyster et al 1998) Both H-2Db and H-2Kb genes areexpressed at the developing NMJ (Tetruashvily et al in press) AtP15 the percentage of multiply-innervated postsynaptic siteswas three times WT levels in KbDb mice (multiple innerva-tion 106 plusmn 11 n = 5 p = 0001 relative to WT) This was not sig-nificantly different from b2mTAP values (p = 009)suggesting that loss of H-2K and H-2D could fully explain the initialsynapse elimination defects in b2mTAP mice In contrast lossof either H-2K or H-2D alone did not affect synapse elimination(Fig 1E) suggesting that only a single classical MHCI is necessaryfor normal synapse elimination to occur Thus three different mod-els that disrupt MHCI function (b2mTAP and KbDb miceand anti-MHCI function-blocking antibodies injected into WTmice) all cause multiple innervation to persist together stronglysuggesting that MHCI promotes synapse elimination at the devel-oping NMJ Furthermore results in NSE-Db and KbDb micesuggest that synapse elimination at the NMJ is promoted by classi-cal MHCI proteins
24 MHCI is expressed at the developing NMJ
MHCI protein is not thought to be expressed in uninjured mus-cle cells but has been detected in adult MNs at the NMJ (Thamset al 2009) mRNAs encoding several MHCI proteins have beendetected at the NMJ at multiple ages (Tetruashvily et al inpress) To determine if MHCI protein is present at the NMJ assynapse elimination is occurring we immunostained intact muscle
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
using a mouse monoclonal antibody (OX-18) that recognizes amonomorphic epitope of rat MHCI (RT-1A (Fukumoto et al1982)) and reacts with MHCI from rat mouse and primate brainin Western blots and immunohistochemistry (Corriveau et al1998 Datwani et al 2009 Dixon-Salazar et al 2014 Goddardet al 2007 Huh et al 2000 Needleman et al 2010 Rollekeet al 2006) Several lines of evidence support OX-18rsquos specificityfor MHCI in the mouse nervous system (see Section 4) UsingOX18 we detect specific MHCI labeling at the NMJ at P7 assynapse elimination is occurring (Fig S1A) This labeling is abol-ished by genetic destabilization of MHCI proteins (in b2mTAP
mice) or by replacing OX-18 with an isotype-control antibody(Fig S1) Whole-mount muscles were triple-labeled with OX18 tolabel MHCI a-btx to label postsynaptic nAChRs and anti-synaptophysin to visualize presynaptic MN terminals Becausepre- and post-synaptic structures are closely overlaid at the NMJthree-dimensional reconstruction of staining in confocal Z-stacksis necessary to unambiguously discriminate co-localization withpre- vs post-synaptic markers In P7 diaphragm MHCI co-localizes more extensively in individual optical sections with post-synaptic nAChRs than with presynaptic synaptophysin (Fig S1A)Specific MHCI labeling does not appear throughout the musclefiber but is restricted to the postsynaptic site Thus MHCI proteinis expressed at the NMJ at the time when developmental synapseelimination is occurring
25 Excess MN inputs can be recruited by graded stimulation in MHCI-deficient mice
Surplus MN inputs are functionally silenced before they arephysically removed (Colman et al 1997 Kopp et al 2000) Ourresults show that MHCI promotes synapse elimination but it isunclear if MHCI is involved in only the final physical removal ofsurplus axons or is involved in the earlier functional silencingTo discriminate between these possibilities we performed sharpelectrode recordings in P15 WT and MHCI-deficient animals andapplied graded stimulation to the phrenic nerve to sequentiallyrecruit MN inputs (Redfern 1970) Graded stimulation producedsingle-amplitude end plate potentials (EPPs) in 100 of recordingsfrom P15 WT animals as expected reflecting functional monoin-nervation of all postsynaptic sites at this age In MHCI-deficientanimals in contrast graded stimulation evoked a large stepwiseincrease in the EPP amplitude in 21 of recordings reflecting
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 5
recruitment of multiple MN inputs Multiple inputs could still berecruited at 13 of junctions in MHCI-deficient adults (Fig 3)These functional measures independently corroborate the anatom-ical evidence that synapse elimination is impaired in MHCI-deficient mice and show that some postsynaptic sites continueto receive inputs from MN axons that are distinct all the way tothe phrenic nerve typical of younger animals before synapse elim-ination is complete (Redfern 1970 Tapia et al 2012) Thus persis-tent multiple innervation in MHCI-deficient animals may resultfrom a failure to silence competing terminal branches rather thana failure to physically remove silent inputs
26 Increased synaptic strength at MHCI-deficient NMJs
Competing axons develop a disparity in synaptic strength andweaker inputs are eventually eliminated (Colman et al 1997Kopp et al 2000) Our anatomical and functional data supportthe idea that MHCI promotes silencing and elimination of MNsynapses In the adult hippocampus MHCI is required for long-term depression a form of persistent synaptic weakening (Huhet al 2000 Nelson et al 2013) To determine if MHCI is involvedin synapse weakening at the developing NMJ we measured synap-tic strength Quantal size is a measure of synaptic strength thatreflects the postsynaptic response to individual quanta of neuro-transmitter (Pennefather and Quastel 1981) At the NMJ miniatureend plate potential (MEPP) amplitude provides a reliable estimateof quantal size (Del Castillo and Katz 1954) MEPP amplitude issignificantly larger than WT in MHCI-deficient animals at P15(b2mTAP 178 plusmn 014 mV WT 115 plusmn 016 mV p = 0022)Quantal size decreases with age as previously observed(Wareham et al 1994) but remains larger in b2mTAP adults(P29-P45 b2mTAP 10 plusmn 008 mV WT 066 plusmn 006 mVp = 0004 Fig S3) Increased quantal size in MHCI-deficient NMJsis not associated with changes in input resistance EPP decay timeMEPP frequency paired-pulse facilitation (Fig S4) postsynapticnAChR density (Fig S2) or the number of quanta released perpresynaptic nerve impulse (quantal content) (Fig S4E) Togetherthese results show that loss of MHCI is associated with an earlyand persistent increase in synaptic strength Thus endogenousMHCI promotes synaptic weakening at the developing NMJ andmay contribute to the decline in synaptic strength that precedesfunctional and physical elimination of competing inputs duringnormal circuit development (Buffelli et al 2003 Colman et al1997 Kopp et al 2000)
27 MHCI levels rise in aged muscle
MHCI mRNA is up-regulated in aged MNs as they become sus-ceptible to denervation (Edstrom et al 2004) but is relativelylow in MNs that resist aging-related synapse loss (Edstrom et al2004 Linda et al 1999 Valdez et al 2012) MHCI promotessynapse elimination at the developing NMJ raising the possibilitythat higher levels of MHCI could be involved in aging-related mus-cle denervation However elevated mRNA levels do not guaranteeincreased transcribed protein levels and to our knowledge nostudies have assessed MHCI protein levels at the aging NMJ Todetermine if MHCI protein levels rise at the aging NMJ as denerva-tion is occurring we compared MHCI expression in extensor digi-torum longus (EDL) from P15 and 2-year-old mice using westernblotting The EDL was chosen because it shows significant denerva-tion at this age (Chai et al 2011 Valdez et al 2012) Becauseeither H2-K or H2-D are required for developmental synapse elim-ination (Fig 1E) we used an antibody (p8) that is specific for H2-KValues were normalized to a loading control (see Section 4) Com-paring MHCI expression in EDL from P15 vs 2-year-old mice wefound that H2-K levels rise significantly in aging EDL relative to
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
younger ages (Fig 4A) Thus MHCI protein is expressed at theNMJ during development and MHCI levels rise significantly atthe aging NMJ
28 MHCI promotes aging-related synapse loss
With aging synapses are eliminated frommature muscle fibersleaving some fibers fully denervated (Rudolf et al 2014 Valdezet al 2012) We find that during development MHCI promotessynapse elimination and MHCI levels rise in aging EDL raisingthe possibility that the increase in MHCI expression with age drivesaberrant synapse elimination in mature singly-innervated musclefibers To directly test if MHCI contributes to pathological synapseloss in aging muscle denervation of EDL muscle fibers wasassessed in aged (2-year-old) WT and b2mTAP mice Full orpartial denervation are evident when postsynaptic nAChRs(labeled with a-btx) are not completely apposed by the nerve ter-minal represented by the presynaptic marker SV2 (Valdez et al2010) As expected a subset of muscle fibers were completely den-ervated in aged EDL (122 plusmn 08 fully denervated n = 4) In con-trast both full and partial denervation was dramatically reducedin MHCI-deficient EDL (Fig 4 full denervation 45 plusmn 10p = 00009 relative to WT) While developmental synapse elimina-tion in the EDL was modestly impaired in MHCI-deficient animals(percent multiply-innervated in P15 EDL WT 10 plusmn 05 b2m-TAP 52 plusmn 11 n = 3 animals per age p = 00045) this differenceis not sufficient to fully explain the reduced denervation in MHCI-deficient EDL Thus the relative lack of denervation in aged MHCI-deficient animals cannot be fully explained by a different startingpoint for the aging process but instead reflects a degree of protec-tion from aging-related denervation Together these results showthat MHCI levels rise with aging at the NMJ and that reducingMHCI expression can ameliorate synapse loss in aging muscle
3 Discussion
Synapse elimination is essential for the establishment of matureneural circuits while synapse loss later in life is a significant causeof aging-associated pathology Here we show that MHCI promotesboth developmental synapse elimination and aging-relatedsynapse loss at the vertebrate NMJ MHCI is expressed at the devel-oping NMJ and three different manipulations that reduce MHCIfunction-genetic destabilization or deletion of MHCI proteins orinjection of function-blocking anti-MHCI antibodies-cause multi-ple innervation to persist at otherwise normally-developingsynapses Reducing MHCI expression does not affect patterns ofinnervation prior to the close of the first postnatal week suggest-ing it does not alter earlier synapse formation or elimination How-ever synapse elimination during the second week is significantlydisrupted preventing establishment of uniform mono-innervation Multiply-innervated postsynaptic sites are still pre-sent in MHCI-deficient animals at one year of age are evident bothanatomically and functionally and are associated with increasedquantal size Thus MHCI is required to complete the transition touniform mono-innervation in developing muscle and promotessynaptic weakening one of the earliest hallmarks of competitionamong multiple inputs to a single muscle fiber With aging MHCIlevels rise in motor neurons (Edstrom et al 2004) and in MHCI-deficient mice aging-related muscle denervation is markedlyreduced
Several experimental conditions delay synapse elimination atthe NMJ including changes in synaptic activity (Misgeld et al2002 Thompson et al 1979) NMDAR activation (Personiuset al 2008) GDNF (Nguyen et al 1998) BDNF or its receptor TrkB(Je et al 2013) NCAM (Rafuse et al 2000) glial neurofascin
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Adult
ββ2m--TAP--WT
A P15
15
0 5
10
2520
WT β2m--
TAP--
WT β2m--
TAP--
P15
m
ultip
ly in
nerv
ated
β2m--TAP--
0
15
5
10Adul
t
B
WT
Fig 3 Multiple functional inputs persist at MHCI-deficient NMJs (A) Representative EPPs recorded from P15 or adult (P29-45) muscle Scales P15 1 mV 15 ms adult10 mV 10 ms Stimulus artifacts blanked for clarity (B) Summed electrophysiological data shows increased multiple innervation in MHCI-deficient mice P15 WT 16 cellsfrom 8 animals b2mTAP 19 cells from 6 animals adult WT 25 cells from 5 animals b2mTAP 38 cells from 8 animals
6 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
(Roche et al 2014) or PKC theta (Li et al 2004) Synapse elimina-tion is also disrupted in mice which bear a dominant-negativemutation in the GARS gene a mouse model of Charcot-Marie-Tooth type 2D which includes apparent developmental arrest ofNMJ maturation (Sleigh et al 2014) Other manipulations includ-ing increasing activity reducing gap junctional coupling or loss ofthe presynaptic glycine transporter GlyT2 accelerate synapseelimination (Bogdanik et al 2012 OrsquoBrien et al 1978 Personiuset al 2007) These and other studies have provided fundamentalinsights into the molecular and cellular control of the rate ofsynapse elimination and suggest factors that may allow synapseelimination to occur normally prior to P7 in MHCI-deficient miceIn some of these models development of the NMJ is grossly dis-rupted which could contribute to their failure to undergo synapseelimination on a normal schedule While similar gross develop-mental disruption is not present at MHCI-deficient NMJs we can-not rule out the possibility that loss of MHCI perturbs synapseelimination indirectly through changes in aspects of developmentthat were not assessed With most manipulations however uni-form monoinnervation is still established within the first few post-natal weeks while multiple innervation is still evident at one yearof age in MHCI-deficient mice suggesting that synapse eliminationis persistently disrupted
We detect MHCI protein at the developing NMJ where it colo-calizes with nAChR labeling suggesting that it is expressed in themuscle fiber Importantly MHCI may also be expressed in themotor neuron andor Schwann cell during development andor inaging a possibility we cannot exclude because there are currentlyno antibodies to detect most members of the large MHCI familyMultiple MHCI mRNAs are expressed at the NMJ at different ages(Tetruashvily et al in press) MHCI protein has been detected inpresynaptic motor neurons in adult mice (Thams et al 2009)and in human and rat Schwann cells in vitro (Armati et al 1990Samuel et al 1987) Furthermore we find that modest overexpres-sion of the MHCI H2-D in neurons (Rall et al 1995) is sufficient tomodestly accelerate synapse elimination (Fig 2C) Ultimatelyimmuno-electron or super-resolution microscopy coupled withdevelopment of dozens of new antibodies will be necessary tofully characterize the pre- vs post-synaptic expression of the many
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
MHCI proteins Nevertheless the current results show that MHCI ispresent at the developing NMJ when synapse elimination isoccurring
MHCI is thought to be repressed by healthy muscle cells andupregulated only after injury or in disease states (Karpati et al1988) The current results demonstrate an unexpected role forMHCI at the healthy developing NMJ Remarkably MHCIrsquos functionat the developing NMJ is strikingly different to its role after injuryReducing MHCI expression exacerbates loss of synapses made ontoaxotomized motor neurons (Berg et al 2013 Oliveira et al 2004)while we find that both developmental and aging-related loss ofmotor neuron synapses onto muscles are inhibited
The bulk of excess MN synapses are pruned prior to P7 (Tapiaet al 2012) but mono-innervation is largely established betweenP7 and P15 as MNs begin to fire asynchronously (Buffelli et al2002 Personius et al 2001) In MHCI-deficient mice 15ndash20of postsynaptic sites retain two inputs at P15 No postsynaptic sitesreceived three or more inputs at P15 in MHCI-deficient animalsconsistent with the fact that very few postsynaptic sites stillreceive 3 or more inputs at P7 in WTs (Fig S2F) the age whenMHCIrsquos effects on synapse elimination begin to emerge The failureof circuit maturation in MHCI-deficient animals is not associatedwith gross motor defects in mice but could prevent the emergenceof fine-motor control that is behaviorally relevant in humansStrikingly the extent of the synapse elimination deficit at theMHCI-deficient NMJ is similar to the extent of axon pruning defi-cits in the lateral geniculate nucleus of mice lacking MHCI(Datwani et al 2009 Huh et al 2000 Lee et al 2014) or lackingneuron pentraxin (Bjartmar et al 2006) or components of thecomplement cascade (Stevens et al 2007) The reason for the par-tial nature of the pruning failure in the visual system in these mod-els remains unknown At the NMJ however we find that MHCI isspecifically required for the final stages of synapse eliminationduring the second postnatal week (Fig 1) In adult motor neuronslevels of the classical MHCI H-2D appear to be heterogeneous(Thams et al 2009) raising the possibility that different levels ofMHCI during development may also render some inputs more sus-ceptible to elimination Alternatively other factors including fibertype and associated activity patterns may influence the role that
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fullydenervated
15
10
5
0 β2m--
TAP--WT
3
2
1
0 β2m--
TAP--WT
E
Partially denervated
AChR
s (α
-btx
)
B C
SV2
5
4
D
mer
ge
A
MH
CI
P1540 80 160μg
MH
CI
2 Y O40 80 160μg
p
artia
lly d
ener
vate
d
fu
lly d
ener
vate
d
P15 2 Y OMH
CI l
abel
ing
in E
DL
(nor
mal
ized
to G
APD
H)
0
05025
07510
Fig 4 MHCI protein levels climb in aging EDL and reducing MHCI expression prevents aging-related denervation (A) Western blots for MHCI in lysates of EDL muscle fromP15 and 2-year-old mice Three different total protein concentrations are shown Blots were stripped and probed for GAPDH as a loading control Bottom MHCI densitometrynormalized to GAPDH for six technical replicates (two per concentration) at each age p = 00048 (B C) EDL muscles from 24ndash26 month-old mice were stained with a-btx(red) to detect nAChRs and SV2 (green) to detect presynaptic nerve terminals Representative images show examples of partially (arrows B) and fully (oval C) denervatedAChR clusters in WT EDL (red with no associated green in merge) Scales 7 lm (D E) Mean partial (D) and full (E) denervation in 2 year old EDL muscle from WT and MHCI-deficient b2mTAP mice (n = 4 per genotype) p = 003 p = 00009
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 7
MHCI plays in elimination of specific axon branches This is consis-tent with the finding that MHCI is required for synapse eliminationto reach completion in the diaphragm which consists of a mixtureof fast and slow fiber types but has a smaller influence on synapseelimination in the EDL which is almost exclusively composed offast-twitch fibers (Eddinger et al 1985) Together the currentresults suggest that synapse elimination at the NMJ may be dividedinto stages with distinct molecular requirements and that MHCI isessential as the last postsynaptic sites transition from dual- tomono-innervation
During synapse elimination at the NMJ competing axonsdevelop a disparity in synaptic strengthwhich precedeswithdrawalof the weaker input (Colman et al 1997 Kopp et al 2000) While itis unknown if larger synaptic strength at the MHCI-deficient NMJ isrelated to deficits in synapse elimination stronger inputs may beless vulnerable to competition-induced synaptic weakening In thisway loss of MHCI might prevent the development of sufficient dis-parity in synaptic strength to trigger functional silencing and phys-ical elimination in the most closely-matched competitions
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Interestingly paired increases in synapse strength and density likethose we observe at the MHCI-deficient NMJ are also seen in thecortex (Glynn et al 2011) and lateral geniculate nucleus (LGN)(Lee et al 2014) of MHCI-deficient mice Additional studies inMHCI-deficient mice found increases in synapse density in cortex(Adelson et al 2014) and increased synapse density (Dixon-Salazar et al 2014) and strength (Fourgeaud et al 2010) in hip-pocampus While the present study identifies a role for MHCI insynapse elimination at the NMJ it is as yet unknown if MHCI inhi-bits synapse formation andor promotes synapse elimination incortex or hippocampus
It will be important to determine if MHCI expression is regu-lated by activity at the developing NMJ as it is in the visual system(Corriveau et al 1998) to determine if it is specifically responsiveto the levels patterns andor synchronization of activity and tofully define the cellular and molecular networks linking MHCI tochanges in the strength and connectivity of synapses Animmunoreceptor for MHCI PirB has been detected in the CNS(Syken et al 2006) and at the adult NMJ (Thams et al 2009)
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

A
controlantibody
0
12
6
3
9
B
m
ultip
ly in
nerv
ated
cont
rol
antib
ody
MH
CI
antib
ody
P15
MHCI antibody
C P7
WT NSE-Db
70
35
0
Fig 2 Anti-MHCI antibodies impair synapse elimination at the NMJ in WT mice while overexpressing MHCI in neurons accelerates synapse elimination (A) RepresentativeNMJs from OX18- and IgG1-injected WT animals at P15 nAChRs labeled with a-btx (red) presynaptic axons and terminals labeled with anti-neurofilament and anti-SV2(green) Arrows individual MN axons Scales 10 lm (B) Quantification of multiple innervation in IgG1- vs OX18-injected animals at P15 OX18 88 plusmn 17 IgG 30 plusmn 08p = 002 OX18 injection n = 9 animals from 4 litters IgG1 injection n = 6 animals from 3 litters (C) At P7 when synapse elimination is not yet complete there is a modestbut significant decrease in the percentage of muscle fibers that remain multiply-innervated in NSE-Db mice reflecting the fact that postsynaptic sites more rapidly achievemonoinnervation in MHCI-overexpressing animals WT n = 3 NSE-Db n = 5 p = 0012
4 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
half of postsynaptic sites remain multiply-innervated in WT ani-mals However a modestly but significantly lower percentage ofpostsynaptic sites remained multiply-innervated in MHCI-overexpressing animals (Fig 2C) The small magnitude of the accel-eration may reflect to fact that H-2Db expression is only modestlyincreased in these transgenics (Lee et al 2014 Rall et al 1995)Together these results demonstrate that developmental synapseelimination at the NMJ can be bi-directionally regulated bychanges in MHCI levels increasing MHCI accelerates synapse elim-ination while reducing MHCI disrupts synapse elimination
These results show that overexpressing just one member of thelarge MHCI gene family the classical MHCI H-2Db is sufficient toaccelerate synapse elimination at the NMJ To determine if classicalMHCIs are involved in synapse elimination at the developing NMJwe examined innervation in KbDb double mutant mice whichlack both of the classical MHCI genes found in C57Bl6 mice(Vugmeyster et al 1998) Both H-2Db and H-2Kb genes areexpressed at the developing NMJ (Tetruashvily et al in press) AtP15 the percentage of multiply-innervated postsynaptic siteswas three times WT levels in KbDb mice (multiple innerva-tion 106 plusmn 11 n = 5 p = 0001 relative to WT) This was not sig-nificantly different from b2mTAP values (p = 009)suggesting that loss of H-2K and H-2D could fully explain the initialsynapse elimination defects in b2mTAP mice In contrast lossof either H-2K or H-2D alone did not affect synapse elimination(Fig 1E) suggesting that only a single classical MHCI is necessaryfor normal synapse elimination to occur Thus three different mod-els that disrupt MHCI function (b2mTAP and KbDb miceand anti-MHCI function-blocking antibodies injected into WTmice) all cause multiple innervation to persist together stronglysuggesting that MHCI promotes synapse elimination at the devel-oping NMJ Furthermore results in NSE-Db and KbDb micesuggest that synapse elimination at the NMJ is promoted by classi-cal MHCI proteins
24 MHCI is expressed at the developing NMJ
MHCI protein is not thought to be expressed in uninjured mus-cle cells but has been detected in adult MNs at the NMJ (Thamset al 2009) mRNAs encoding several MHCI proteins have beendetected at the NMJ at multiple ages (Tetruashvily et al inpress) To determine if MHCI protein is present at the NMJ assynapse elimination is occurring we immunostained intact muscle
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
using a mouse monoclonal antibody (OX-18) that recognizes amonomorphic epitope of rat MHCI (RT-1A (Fukumoto et al1982)) and reacts with MHCI from rat mouse and primate brainin Western blots and immunohistochemistry (Corriveau et al1998 Datwani et al 2009 Dixon-Salazar et al 2014 Goddardet al 2007 Huh et al 2000 Needleman et al 2010 Rollekeet al 2006) Several lines of evidence support OX-18rsquos specificityfor MHCI in the mouse nervous system (see Section 4) UsingOX18 we detect specific MHCI labeling at the NMJ at P7 assynapse elimination is occurring (Fig S1A) This labeling is abol-ished by genetic destabilization of MHCI proteins (in b2mTAP
mice) or by replacing OX-18 with an isotype-control antibody(Fig S1) Whole-mount muscles were triple-labeled with OX18 tolabel MHCI a-btx to label postsynaptic nAChRs and anti-synaptophysin to visualize presynaptic MN terminals Becausepre- and post-synaptic structures are closely overlaid at the NMJthree-dimensional reconstruction of staining in confocal Z-stacksis necessary to unambiguously discriminate co-localization withpre- vs post-synaptic markers In P7 diaphragm MHCI co-localizes more extensively in individual optical sections with post-synaptic nAChRs than with presynaptic synaptophysin (Fig S1A)Specific MHCI labeling does not appear throughout the musclefiber but is restricted to the postsynaptic site Thus MHCI proteinis expressed at the NMJ at the time when developmental synapseelimination is occurring
25 Excess MN inputs can be recruited by graded stimulation in MHCI-deficient mice
Surplus MN inputs are functionally silenced before they arephysically removed (Colman et al 1997 Kopp et al 2000) Ourresults show that MHCI promotes synapse elimination but it isunclear if MHCI is involved in only the final physical removal ofsurplus axons or is involved in the earlier functional silencingTo discriminate between these possibilities we performed sharpelectrode recordings in P15 WT and MHCI-deficient animals andapplied graded stimulation to the phrenic nerve to sequentiallyrecruit MN inputs (Redfern 1970) Graded stimulation producedsingle-amplitude end plate potentials (EPPs) in 100 of recordingsfrom P15 WT animals as expected reflecting functional monoin-nervation of all postsynaptic sites at this age In MHCI-deficientanimals in contrast graded stimulation evoked a large stepwiseincrease in the EPP amplitude in 21 of recordings reflecting
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 5
recruitment of multiple MN inputs Multiple inputs could still berecruited at 13 of junctions in MHCI-deficient adults (Fig 3)These functional measures independently corroborate the anatom-ical evidence that synapse elimination is impaired in MHCI-deficient mice and show that some postsynaptic sites continueto receive inputs from MN axons that are distinct all the way tothe phrenic nerve typical of younger animals before synapse elim-ination is complete (Redfern 1970 Tapia et al 2012) Thus persis-tent multiple innervation in MHCI-deficient animals may resultfrom a failure to silence competing terminal branches rather thana failure to physically remove silent inputs
26 Increased synaptic strength at MHCI-deficient NMJs
Competing axons develop a disparity in synaptic strength andweaker inputs are eventually eliminated (Colman et al 1997Kopp et al 2000) Our anatomical and functional data supportthe idea that MHCI promotes silencing and elimination of MNsynapses In the adult hippocampus MHCI is required for long-term depression a form of persistent synaptic weakening (Huhet al 2000 Nelson et al 2013) To determine if MHCI is involvedin synapse weakening at the developing NMJ we measured synap-tic strength Quantal size is a measure of synaptic strength thatreflects the postsynaptic response to individual quanta of neuro-transmitter (Pennefather and Quastel 1981) At the NMJ miniatureend plate potential (MEPP) amplitude provides a reliable estimateof quantal size (Del Castillo and Katz 1954) MEPP amplitude issignificantly larger than WT in MHCI-deficient animals at P15(b2mTAP 178 plusmn 014 mV WT 115 plusmn 016 mV p = 0022)Quantal size decreases with age as previously observed(Wareham et al 1994) but remains larger in b2mTAP adults(P29-P45 b2mTAP 10 plusmn 008 mV WT 066 plusmn 006 mVp = 0004 Fig S3) Increased quantal size in MHCI-deficient NMJsis not associated with changes in input resistance EPP decay timeMEPP frequency paired-pulse facilitation (Fig S4) postsynapticnAChR density (Fig S2) or the number of quanta released perpresynaptic nerve impulse (quantal content) (Fig S4E) Togetherthese results show that loss of MHCI is associated with an earlyand persistent increase in synaptic strength Thus endogenousMHCI promotes synaptic weakening at the developing NMJ andmay contribute to the decline in synaptic strength that precedesfunctional and physical elimination of competing inputs duringnormal circuit development (Buffelli et al 2003 Colman et al1997 Kopp et al 2000)
27 MHCI levels rise in aged muscle
MHCI mRNA is up-regulated in aged MNs as they become sus-ceptible to denervation (Edstrom et al 2004) but is relativelylow in MNs that resist aging-related synapse loss (Edstrom et al2004 Linda et al 1999 Valdez et al 2012) MHCI promotessynapse elimination at the developing NMJ raising the possibilitythat higher levels of MHCI could be involved in aging-related mus-cle denervation However elevated mRNA levels do not guaranteeincreased transcribed protein levels and to our knowledge nostudies have assessed MHCI protein levels at the aging NMJ Todetermine if MHCI protein levels rise at the aging NMJ as denerva-tion is occurring we compared MHCI expression in extensor digi-torum longus (EDL) from P15 and 2-year-old mice using westernblotting The EDL was chosen because it shows significant denerva-tion at this age (Chai et al 2011 Valdez et al 2012) Becauseeither H2-K or H2-D are required for developmental synapse elim-ination (Fig 1E) we used an antibody (p8) that is specific for H2-KValues were normalized to a loading control (see Section 4) Com-paring MHCI expression in EDL from P15 vs 2-year-old mice wefound that H2-K levels rise significantly in aging EDL relative to
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
younger ages (Fig 4A) Thus MHCI protein is expressed at theNMJ during development and MHCI levels rise significantly atthe aging NMJ
28 MHCI promotes aging-related synapse loss
With aging synapses are eliminated frommature muscle fibersleaving some fibers fully denervated (Rudolf et al 2014 Valdezet al 2012) We find that during development MHCI promotessynapse elimination and MHCI levels rise in aging EDL raisingthe possibility that the increase in MHCI expression with age drivesaberrant synapse elimination in mature singly-innervated musclefibers To directly test if MHCI contributes to pathological synapseloss in aging muscle denervation of EDL muscle fibers wasassessed in aged (2-year-old) WT and b2mTAP mice Full orpartial denervation are evident when postsynaptic nAChRs(labeled with a-btx) are not completely apposed by the nerve ter-minal represented by the presynaptic marker SV2 (Valdez et al2010) As expected a subset of muscle fibers were completely den-ervated in aged EDL (122 plusmn 08 fully denervated n = 4) In con-trast both full and partial denervation was dramatically reducedin MHCI-deficient EDL (Fig 4 full denervation 45 plusmn 10p = 00009 relative to WT) While developmental synapse elimina-tion in the EDL was modestly impaired in MHCI-deficient animals(percent multiply-innervated in P15 EDL WT 10 plusmn 05 b2m-TAP 52 plusmn 11 n = 3 animals per age p = 00045) this differenceis not sufficient to fully explain the reduced denervation in MHCI-deficient EDL Thus the relative lack of denervation in aged MHCI-deficient animals cannot be fully explained by a different startingpoint for the aging process but instead reflects a degree of protec-tion from aging-related denervation Together these results showthat MHCI levels rise with aging at the NMJ and that reducingMHCI expression can ameliorate synapse loss in aging muscle
3 Discussion
Synapse elimination is essential for the establishment of matureneural circuits while synapse loss later in life is a significant causeof aging-associated pathology Here we show that MHCI promotesboth developmental synapse elimination and aging-relatedsynapse loss at the vertebrate NMJ MHCI is expressed at the devel-oping NMJ and three different manipulations that reduce MHCIfunction-genetic destabilization or deletion of MHCI proteins orinjection of function-blocking anti-MHCI antibodies-cause multi-ple innervation to persist at otherwise normally-developingsynapses Reducing MHCI expression does not affect patterns ofinnervation prior to the close of the first postnatal week suggest-ing it does not alter earlier synapse formation or elimination How-ever synapse elimination during the second week is significantlydisrupted preventing establishment of uniform mono-innervation Multiply-innervated postsynaptic sites are still pre-sent in MHCI-deficient animals at one year of age are evident bothanatomically and functionally and are associated with increasedquantal size Thus MHCI is required to complete the transition touniform mono-innervation in developing muscle and promotessynaptic weakening one of the earliest hallmarks of competitionamong multiple inputs to a single muscle fiber With aging MHCIlevels rise in motor neurons (Edstrom et al 2004) and in MHCI-deficient mice aging-related muscle denervation is markedlyreduced
Several experimental conditions delay synapse elimination atthe NMJ including changes in synaptic activity (Misgeld et al2002 Thompson et al 1979) NMDAR activation (Personiuset al 2008) GDNF (Nguyen et al 1998) BDNF or its receptor TrkB(Je et al 2013) NCAM (Rafuse et al 2000) glial neurofascin
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Adult
ββ2m--TAP--WT
A P15
15
0 5
10
2520
WT β2m--
TAP--
WT β2m--
TAP--
P15
m
ultip
ly in
nerv
ated
β2m--TAP--
0
15
5
10Adul
t
B
WT
Fig 3 Multiple functional inputs persist at MHCI-deficient NMJs (A) Representative EPPs recorded from P15 or adult (P29-45) muscle Scales P15 1 mV 15 ms adult10 mV 10 ms Stimulus artifacts blanked for clarity (B) Summed electrophysiological data shows increased multiple innervation in MHCI-deficient mice P15 WT 16 cellsfrom 8 animals b2mTAP 19 cells from 6 animals adult WT 25 cells from 5 animals b2mTAP 38 cells from 8 animals
6 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
(Roche et al 2014) or PKC theta (Li et al 2004) Synapse elimina-tion is also disrupted in mice which bear a dominant-negativemutation in the GARS gene a mouse model of Charcot-Marie-Tooth type 2D which includes apparent developmental arrest ofNMJ maturation (Sleigh et al 2014) Other manipulations includ-ing increasing activity reducing gap junctional coupling or loss ofthe presynaptic glycine transporter GlyT2 accelerate synapseelimination (Bogdanik et al 2012 OrsquoBrien et al 1978 Personiuset al 2007) These and other studies have provided fundamentalinsights into the molecular and cellular control of the rate ofsynapse elimination and suggest factors that may allow synapseelimination to occur normally prior to P7 in MHCI-deficient miceIn some of these models development of the NMJ is grossly dis-rupted which could contribute to their failure to undergo synapseelimination on a normal schedule While similar gross develop-mental disruption is not present at MHCI-deficient NMJs we can-not rule out the possibility that loss of MHCI perturbs synapseelimination indirectly through changes in aspects of developmentthat were not assessed With most manipulations however uni-form monoinnervation is still established within the first few post-natal weeks while multiple innervation is still evident at one yearof age in MHCI-deficient mice suggesting that synapse eliminationis persistently disrupted
We detect MHCI protein at the developing NMJ where it colo-calizes with nAChR labeling suggesting that it is expressed in themuscle fiber Importantly MHCI may also be expressed in themotor neuron andor Schwann cell during development andor inaging a possibility we cannot exclude because there are currentlyno antibodies to detect most members of the large MHCI familyMultiple MHCI mRNAs are expressed at the NMJ at different ages(Tetruashvily et al in press) MHCI protein has been detected inpresynaptic motor neurons in adult mice (Thams et al 2009)and in human and rat Schwann cells in vitro (Armati et al 1990Samuel et al 1987) Furthermore we find that modest overexpres-sion of the MHCI H2-D in neurons (Rall et al 1995) is sufficient tomodestly accelerate synapse elimination (Fig 2C) Ultimatelyimmuno-electron or super-resolution microscopy coupled withdevelopment of dozens of new antibodies will be necessary tofully characterize the pre- vs post-synaptic expression of the many
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
MHCI proteins Nevertheless the current results show that MHCI ispresent at the developing NMJ when synapse elimination isoccurring
MHCI is thought to be repressed by healthy muscle cells andupregulated only after injury or in disease states (Karpati et al1988) The current results demonstrate an unexpected role forMHCI at the healthy developing NMJ Remarkably MHCIrsquos functionat the developing NMJ is strikingly different to its role after injuryReducing MHCI expression exacerbates loss of synapses made ontoaxotomized motor neurons (Berg et al 2013 Oliveira et al 2004)while we find that both developmental and aging-related loss ofmotor neuron synapses onto muscles are inhibited
The bulk of excess MN synapses are pruned prior to P7 (Tapiaet al 2012) but mono-innervation is largely established betweenP7 and P15 as MNs begin to fire asynchronously (Buffelli et al2002 Personius et al 2001) In MHCI-deficient mice 15ndash20of postsynaptic sites retain two inputs at P15 No postsynaptic sitesreceived three or more inputs at P15 in MHCI-deficient animalsconsistent with the fact that very few postsynaptic sites stillreceive 3 or more inputs at P7 in WTs (Fig S2F) the age whenMHCIrsquos effects on synapse elimination begin to emerge The failureof circuit maturation in MHCI-deficient animals is not associatedwith gross motor defects in mice but could prevent the emergenceof fine-motor control that is behaviorally relevant in humansStrikingly the extent of the synapse elimination deficit at theMHCI-deficient NMJ is similar to the extent of axon pruning defi-cits in the lateral geniculate nucleus of mice lacking MHCI(Datwani et al 2009 Huh et al 2000 Lee et al 2014) or lackingneuron pentraxin (Bjartmar et al 2006) or components of thecomplement cascade (Stevens et al 2007) The reason for the par-tial nature of the pruning failure in the visual system in these mod-els remains unknown At the NMJ however we find that MHCI isspecifically required for the final stages of synapse eliminationduring the second postnatal week (Fig 1) In adult motor neuronslevels of the classical MHCI H-2D appear to be heterogeneous(Thams et al 2009) raising the possibility that different levels ofMHCI during development may also render some inputs more sus-ceptible to elimination Alternatively other factors including fibertype and associated activity patterns may influence the role that
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fullydenervated
15
10
5
0 β2m--
TAP--WT
3
2
1
0 β2m--
TAP--WT
E
Partially denervated
AChR
s (α
-btx
)
B C
SV2
5
4
D
mer
ge
A
MH
CI
P1540 80 160μg
MH
CI
2 Y O40 80 160μg
p
artia
lly d
ener
vate
d
fu
lly d
ener
vate
d
P15 2 Y OMH
CI l
abel
ing
in E
DL
(nor
mal
ized
to G
APD
H)
0
05025
07510
Fig 4 MHCI protein levels climb in aging EDL and reducing MHCI expression prevents aging-related denervation (A) Western blots for MHCI in lysates of EDL muscle fromP15 and 2-year-old mice Three different total protein concentrations are shown Blots were stripped and probed for GAPDH as a loading control Bottom MHCI densitometrynormalized to GAPDH for six technical replicates (two per concentration) at each age p = 00048 (B C) EDL muscles from 24ndash26 month-old mice were stained with a-btx(red) to detect nAChRs and SV2 (green) to detect presynaptic nerve terminals Representative images show examples of partially (arrows B) and fully (oval C) denervatedAChR clusters in WT EDL (red with no associated green in merge) Scales 7 lm (D E) Mean partial (D) and full (E) denervation in 2 year old EDL muscle from WT and MHCI-deficient b2mTAP mice (n = 4 per genotype) p = 003 p = 00009
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 7
MHCI plays in elimination of specific axon branches This is consis-tent with the finding that MHCI is required for synapse eliminationto reach completion in the diaphragm which consists of a mixtureof fast and slow fiber types but has a smaller influence on synapseelimination in the EDL which is almost exclusively composed offast-twitch fibers (Eddinger et al 1985) Together the currentresults suggest that synapse elimination at the NMJ may be dividedinto stages with distinct molecular requirements and that MHCI isessential as the last postsynaptic sites transition from dual- tomono-innervation
During synapse elimination at the NMJ competing axonsdevelop a disparity in synaptic strengthwhich precedeswithdrawalof the weaker input (Colman et al 1997 Kopp et al 2000) While itis unknown if larger synaptic strength at the MHCI-deficient NMJ isrelated to deficits in synapse elimination stronger inputs may beless vulnerable to competition-induced synaptic weakening In thisway loss of MHCI might prevent the development of sufficient dis-parity in synaptic strength to trigger functional silencing and phys-ical elimination in the most closely-matched competitions
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Interestingly paired increases in synapse strength and density likethose we observe at the MHCI-deficient NMJ are also seen in thecortex (Glynn et al 2011) and lateral geniculate nucleus (LGN)(Lee et al 2014) of MHCI-deficient mice Additional studies inMHCI-deficient mice found increases in synapse density in cortex(Adelson et al 2014) and increased synapse density (Dixon-Salazar et al 2014) and strength (Fourgeaud et al 2010) in hip-pocampus While the present study identifies a role for MHCI insynapse elimination at the NMJ it is as yet unknown if MHCI inhi-bits synapse formation andor promotes synapse elimination incortex or hippocampus
It will be important to determine if MHCI expression is regu-lated by activity at the developing NMJ as it is in the visual system(Corriveau et al 1998) to determine if it is specifically responsiveto the levels patterns andor synchronization of activity and tofully define the cellular and molecular networks linking MHCI tochanges in the strength and connectivity of synapses Animmunoreceptor for MHCI PirB has been detected in the CNS(Syken et al 2006) and at the adult NMJ (Thams et al 2009)
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 5
recruitment of multiple MN inputs Multiple inputs could still berecruited at 13 of junctions in MHCI-deficient adults (Fig 3)These functional measures independently corroborate the anatom-ical evidence that synapse elimination is impaired in MHCI-deficient mice and show that some postsynaptic sites continueto receive inputs from MN axons that are distinct all the way tothe phrenic nerve typical of younger animals before synapse elim-ination is complete (Redfern 1970 Tapia et al 2012) Thus persis-tent multiple innervation in MHCI-deficient animals may resultfrom a failure to silence competing terminal branches rather thana failure to physically remove silent inputs
26 Increased synaptic strength at MHCI-deficient NMJs
Competing axons develop a disparity in synaptic strength andweaker inputs are eventually eliminated (Colman et al 1997Kopp et al 2000) Our anatomical and functional data supportthe idea that MHCI promotes silencing and elimination of MNsynapses In the adult hippocampus MHCI is required for long-term depression a form of persistent synaptic weakening (Huhet al 2000 Nelson et al 2013) To determine if MHCI is involvedin synapse weakening at the developing NMJ we measured synap-tic strength Quantal size is a measure of synaptic strength thatreflects the postsynaptic response to individual quanta of neuro-transmitter (Pennefather and Quastel 1981) At the NMJ miniatureend plate potential (MEPP) amplitude provides a reliable estimateof quantal size (Del Castillo and Katz 1954) MEPP amplitude issignificantly larger than WT in MHCI-deficient animals at P15(b2mTAP 178 plusmn 014 mV WT 115 plusmn 016 mV p = 0022)Quantal size decreases with age as previously observed(Wareham et al 1994) but remains larger in b2mTAP adults(P29-P45 b2mTAP 10 plusmn 008 mV WT 066 plusmn 006 mVp = 0004 Fig S3) Increased quantal size in MHCI-deficient NMJsis not associated with changes in input resistance EPP decay timeMEPP frequency paired-pulse facilitation (Fig S4) postsynapticnAChR density (Fig S2) or the number of quanta released perpresynaptic nerve impulse (quantal content) (Fig S4E) Togetherthese results show that loss of MHCI is associated with an earlyand persistent increase in synaptic strength Thus endogenousMHCI promotes synaptic weakening at the developing NMJ andmay contribute to the decline in synaptic strength that precedesfunctional and physical elimination of competing inputs duringnormal circuit development (Buffelli et al 2003 Colman et al1997 Kopp et al 2000)
27 MHCI levels rise in aged muscle
MHCI mRNA is up-regulated in aged MNs as they become sus-ceptible to denervation (Edstrom et al 2004) but is relativelylow in MNs that resist aging-related synapse loss (Edstrom et al2004 Linda et al 1999 Valdez et al 2012) MHCI promotessynapse elimination at the developing NMJ raising the possibilitythat higher levels of MHCI could be involved in aging-related mus-cle denervation However elevated mRNA levels do not guaranteeincreased transcribed protein levels and to our knowledge nostudies have assessed MHCI protein levels at the aging NMJ Todetermine if MHCI protein levels rise at the aging NMJ as denerva-tion is occurring we compared MHCI expression in extensor digi-torum longus (EDL) from P15 and 2-year-old mice using westernblotting The EDL was chosen because it shows significant denerva-tion at this age (Chai et al 2011 Valdez et al 2012) Becauseeither H2-K or H2-D are required for developmental synapse elim-ination (Fig 1E) we used an antibody (p8) that is specific for H2-KValues were normalized to a loading control (see Section 4) Com-paring MHCI expression in EDL from P15 vs 2-year-old mice wefound that H2-K levels rise significantly in aging EDL relative to
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
younger ages (Fig 4A) Thus MHCI protein is expressed at theNMJ during development and MHCI levels rise significantly atthe aging NMJ
28 MHCI promotes aging-related synapse loss
With aging synapses are eliminated frommature muscle fibersleaving some fibers fully denervated (Rudolf et al 2014 Valdezet al 2012) We find that during development MHCI promotessynapse elimination and MHCI levels rise in aging EDL raisingthe possibility that the increase in MHCI expression with age drivesaberrant synapse elimination in mature singly-innervated musclefibers To directly test if MHCI contributes to pathological synapseloss in aging muscle denervation of EDL muscle fibers wasassessed in aged (2-year-old) WT and b2mTAP mice Full orpartial denervation are evident when postsynaptic nAChRs(labeled with a-btx) are not completely apposed by the nerve ter-minal represented by the presynaptic marker SV2 (Valdez et al2010) As expected a subset of muscle fibers were completely den-ervated in aged EDL (122 plusmn 08 fully denervated n = 4) In con-trast both full and partial denervation was dramatically reducedin MHCI-deficient EDL (Fig 4 full denervation 45 plusmn 10p = 00009 relative to WT) While developmental synapse elimina-tion in the EDL was modestly impaired in MHCI-deficient animals(percent multiply-innervated in P15 EDL WT 10 plusmn 05 b2m-TAP 52 plusmn 11 n = 3 animals per age p = 00045) this differenceis not sufficient to fully explain the reduced denervation in MHCI-deficient EDL Thus the relative lack of denervation in aged MHCI-deficient animals cannot be fully explained by a different startingpoint for the aging process but instead reflects a degree of protec-tion from aging-related denervation Together these results showthat MHCI levels rise with aging at the NMJ and that reducingMHCI expression can ameliorate synapse loss in aging muscle
3 Discussion
Synapse elimination is essential for the establishment of matureneural circuits while synapse loss later in life is a significant causeof aging-associated pathology Here we show that MHCI promotesboth developmental synapse elimination and aging-relatedsynapse loss at the vertebrate NMJ MHCI is expressed at the devel-oping NMJ and three different manipulations that reduce MHCIfunction-genetic destabilization or deletion of MHCI proteins orinjection of function-blocking anti-MHCI antibodies-cause multi-ple innervation to persist at otherwise normally-developingsynapses Reducing MHCI expression does not affect patterns ofinnervation prior to the close of the first postnatal week suggest-ing it does not alter earlier synapse formation or elimination How-ever synapse elimination during the second week is significantlydisrupted preventing establishment of uniform mono-innervation Multiply-innervated postsynaptic sites are still pre-sent in MHCI-deficient animals at one year of age are evident bothanatomically and functionally and are associated with increasedquantal size Thus MHCI is required to complete the transition touniform mono-innervation in developing muscle and promotessynaptic weakening one of the earliest hallmarks of competitionamong multiple inputs to a single muscle fiber With aging MHCIlevels rise in motor neurons (Edstrom et al 2004) and in MHCI-deficient mice aging-related muscle denervation is markedlyreduced
Several experimental conditions delay synapse elimination atthe NMJ including changes in synaptic activity (Misgeld et al2002 Thompson et al 1979) NMDAR activation (Personiuset al 2008) GDNF (Nguyen et al 1998) BDNF or its receptor TrkB(Je et al 2013) NCAM (Rafuse et al 2000) glial neurofascin
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Adult
ββ2m--TAP--WT
A P15
15
0 5
10
2520
WT β2m--
TAP--
WT β2m--
TAP--
P15
m
ultip
ly in
nerv
ated
β2m--TAP--
0
15
5
10Adul
t
B
WT
Fig 3 Multiple functional inputs persist at MHCI-deficient NMJs (A) Representative EPPs recorded from P15 or adult (P29-45) muscle Scales P15 1 mV 15 ms adult10 mV 10 ms Stimulus artifacts blanked for clarity (B) Summed electrophysiological data shows increased multiple innervation in MHCI-deficient mice P15 WT 16 cellsfrom 8 animals b2mTAP 19 cells from 6 animals adult WT 25 cells from 5 animals b2mTAP 38 cells from 8 animals
6 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
(Roche et al 2014) or PKC theta (Li et al 2004) Synapse elimina-tion is also disrupted in mice which bear a dominant-negativemutation in the GARS gene a mouse model of Charcot-Marie-Tooth type 2D which includes apparent developmental arrest ofNMJ maturation (Sleigh et al 2014) Other manipulations includ-ing increasing activity reducing gap junctional coupling or loss ofthe presynaptic glycine transporter GlyT2 accelerate synapseelimination (Bogdanik et al 2012 OrsquoBrien et al 1978 Personiuset al 2007) These and other studies have provided fundamentalinsights into the molecular and cellular control of the rate ofsynapse elimination and suggest factors that may allow synapseelimination to occur normally prior to P7 in MHCI-deficient miceIn some of these models development of the NMJ is grossly dis-rupted which could contribute to their failure to undergo synapseelimination on a normal schedule While similar gross develop-mental disruption is not present at MHCI-deficient NMJs we can-not rule out the possibility that loss of MHCI perturbs synapseelimination indirectly through changes in aspects of developmentthat were not assessed With most manipulations however uni-form monoinnervation is still established within the first few post-natal weeks while multiple innervation is still evident at one yearof age in MHCI-deficient mice suggesting that synapse eliminationis persistently disrupted
We detect MHCI protein at the developing NMJ where it colo-calizes with nAChR labeling suggesting that it is expressed in themuscle fiber Importantly MHCI may also be expressed in themotor neuron andor Schwann cell during development andor inaging a possibility we cannot exclude because there are currentlyno antibodies to detect most members of the large MHCI familyMultiple MHCI mRNAs are expressed at the NMJ at different ages(Tetruashvily et al in press) MHCI protein has been detected inpresynaptic motor neurons in adult mice (Thams et al 2009)and in human and rat Schwann cells in vitro (Armati et al 1990Samuel et al 1987) Furthermore we find that modest overexpres-sion of the MHCI H2-D in neurons (Rall et al 1995) is sufficient tomodestly accelerate synapse elimination (Fig 2C) Ultimatelyimmuno-electron or super-resolution microscopy coupled withdevelopment of dozens of new antibodies will be necessary tofully characterize the pre- vs post-synaptic expression of the many
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
MHCI proteins Nevertheless the current results show that MHCI ispresent at the developing NMJ when synapse elimination isoccurring
MHCI is thought to be repressed by healthy muscle cells andupregulated only after injury or in disease states (Karpati et al1988) The current results demonstrate an unexpected role forMHCI at the healthy developing NMJ Remarkably MHCIrsquos functionat the developing NMJ is strikingly different to its role after injuryReducing MHCI expression exacerbates loss of synapses made ontoaxotomized motor neurons (Berg et al 2013 Oliveira et al 2004)while we find that both developmental and aging-related loss ofmotor neuron synapses onto muscles are inhibited
The bulk of excess MN synapses are pruned prior to P7 (Tapiaet al 2012) but mono-innervation is largely established betweenP7 and P15 as MNs begin to fire asynchronously (Buffelli et al2002 Personius et al 2001) In MHCI-deficient mice 15ndash20of postsynaptic sites retain two inputs at P15 No postsynaptic sitesreceived three or more inputs at P15 in MHCI-deficient animalsconsistent with the fact that very few postsynaptic sites stillreceive 3 or more inputs at P7 in WTs (Fig S2F) the age whenMHCIrsquos effects on synapse elimination begin to emerge The failureof circuit maturation in MHCI-deficient animals is not associatedwith gross motor defects in mice but could prevent the emergenceof fine-motor control that is behaviorally relevant in humansStrikingly the extent of the synapse elimination deficit at theMHCI-deficient NMJ is similar to the extent of axon pruning defi-cits in the lateral geniculate nucleus of mice lacking MHCI(Datwani et al 2009 Huh et al 2000 Lee et al 2014) or lackingneuron pentraxin (Bjartmar et al 2006) or components of thecomplement cascade (Stevens et al 2007) The reason for the par-tial nature of the pruning failure in the visual system in these mod-els remains unknown At the NMJ however we find that MHCI isspecifically required for the final stages of synapse eliminationduring the second postnatal week (Fig 1) In adult motor neuronslevels of the classical MHCI H-2D appear to be heterogeneous(Thams et al 2009) raising the possibility that different levels ofMHCI during development may also render some inputs more sus-ceptible to elimination Alternatively other factors including fibertype and associated activity patterns may influence the role that
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fullydenervated
15
10
5
0 β2m--
TAP--WT
3
2
1
0 β2m--
TAP--WT
E
Partially denervated
AChR
s (α
-btx
)
B C
SV2
5
4
D
mer
ge
A
MH
CI
P1540 80 160μg
MH
CI
2 Y O40 80 160μg
p
artia
lly d
ener
vate
d
fu
lly d
ener
vate
d
P15 2 Y OMH
CI l
abel
ing
in E
DL
(nor
mal
ized
to G
APD
H)
0
05025
07510
Fig 4 MHCI protein levels climb in aging EDL and reducing MHCI expression prevents aging-related denervation (A) Western blots for MHCI in lysates of EDL muscle fromP15 and 2-year-old mice Three different total protein concentrations are shown Blots were stripped and probed for GAPDH as a loading control Bottom MHCI densitometrynormalized to GAPDH for six technical replicates (two per concentration) at each age p = 00048 (B C) EDL muscles from 24ndash26 month-old mice were stained with a-btx(red) to detect nAChRs and SV2 (green) to detect presynaptic nerve terminals Representative images show examples of partially (arrows B) and fully (oval C) denervatedAChR clusters in WT EDL (red with no associated green in merge) Scales 7 lm (D E) Mean partial (D) and full (E) denervation in 2 year old EDL muscle from WT and MHCI-deficient b2mTAP mice (n = 4 per genotype) p = 003 p = 00009
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 7
MHCI plays in elimination of specific axon branches This is consis-tent with the finding that MHCI is required for synapse eliminationto reach completion in the diaphragm which consists of a mixtureof fast and slow fiber types but has a smaller influence on synapseelimination in the EDL which is almost exclusively composed offast-twitch fibers (Eddinger et al 1985) Together the currentresults suggest that synapse elimination at the NMJ may be dividedinto stages with distinct molecular requirements and that MHCI isessential as the last postsynaptic sites transition from dual- tomono-innervation
During synapse elimination at the NMJ competing axonsdevelop a disparity in synaptic strengthwhich precedeswithdrawalof the weaker input (Colman et al 1997 Kopp et al 2000) While itis unknown if larger synaptic strength at the MHCI-deficient NMJ isrelated to deficits in synapse elimination stronger inputs may beless vulnerable to competition-induced synaptic weakening In thisway loss of MHCI might prevent the development of sufficient dis-parity in synaptic strength to trigger functional silencing and phys-ical elimination in the most closely-matched competitions
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Interestingly paired increases in synapse strength and density likethose we observe at the MHCI-deficient NMJ are also seen in thecortex (Glynn et al 2011) and lateral geniculate nucleus (LGN)(Lee et al 2014) of MHCI-deficient mice Additional studies inMHCI-deficient mice found increases in synapse density in cortex(Adelson et al 2014) and increased synapse density (Dixon-Salazar et al 2014) and strength (Fourgeaud et al 2010) in hip-pocampus While the present study identifies a role for MHCI insynapse elimination at the NMJ it is as yet unknown if MHCI inhi-bits synapse formation andor promotes synapse elimination incortex or hippocampus
It will be important to determine if MHCI expression is regu-lated by activity at the developing NMJ as it is in the visual system(Corriveau et al 1998) to determine if it is specifically responsiveto the levels patterns andor synchronization of activity and tofully define the cellular and molecular networks linking MHCI tochanges in the strength and connectivity of synapses Animmunoreceptor for MHCI PirB has been detected in the CNS(Syken et al 2006) and at the adult NMJ (Thams et al 2009)
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
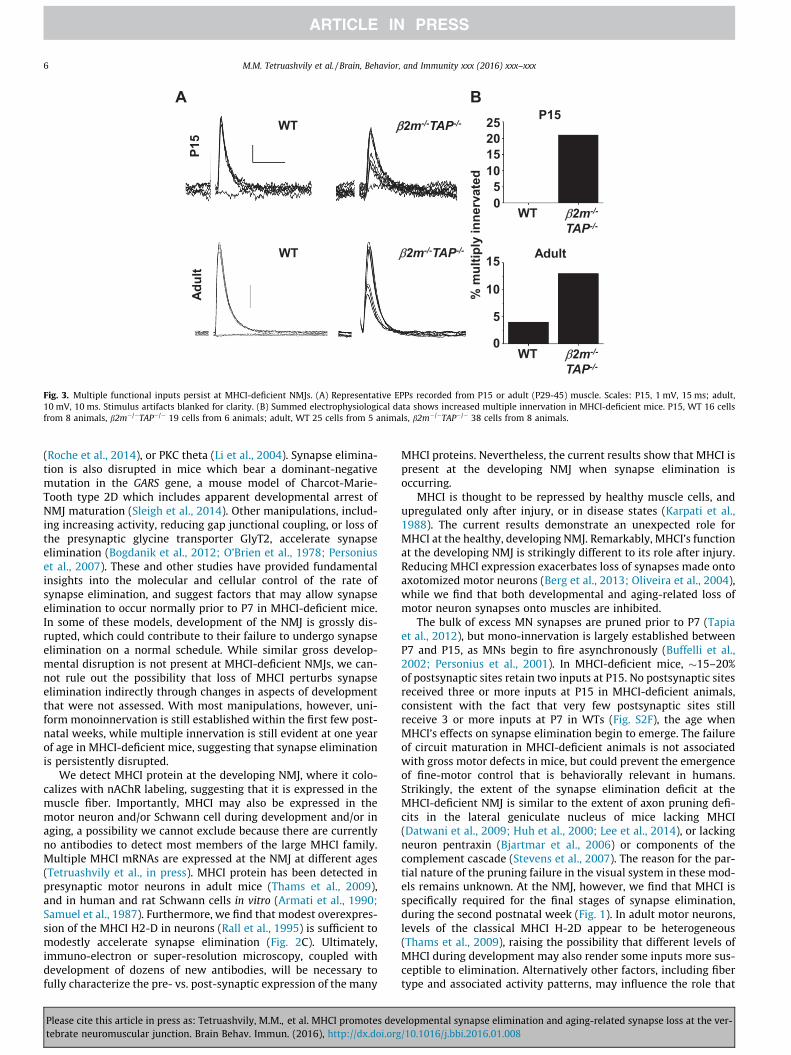
Adult
ββ2m--TAP--WT
A P15
15
0 5
10
2520
WT β2m--
TAP--
WT β2m--
TAP--
P15
m
ultip
ly in
nerv
ated
β2m--TAP--
0
15
5
10Adul
t
B
WT
Fig 3 Multiple functional inputs persist at MHCI-deficient NMJs (A) Representative EPPs recorded from P15 or adult (P29-45) muscle Scales P15 1 mV 15 ms adult10 mV 10 ms Stimulus artifacts blanked for clarity (B) Summed electrophysiological data shows increased multiple innervation in MHCI-deficient mice P15 WT 16 cellsfrom 8 animals b2mTAP 19 cells from 6 animals adult WT 25 cells from 5 animals b2mTAP 38 cells from 8 animals
6 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
(Roche et al 2014) or PKC theta (Li et al 2004) Synapse elimina-tion is also disrupted in mice which bear a dominant-negativemutation in the GARS gene a mouse model of Charcot-Marie-Tooth type 2D which includes apparent developmental arrest ofNMJ maturation (Sleigh et al 2014) Other manipulations includ-ing increasing activity reducing gap junctional coupling or loss ofthe presynaptic glycine transporter GlyT2 accelerate synapseelimination (Bogdanik et al 2012 OrsquoBrien et al 1978 Personiuset al 2007) These and other studies have provided fundamentalinsights into the molecular and cellular control of the rate ofsynapse elimination and suggest factors that may allow synapseelimination to occur normally prior to P7 in MHCI-deficient miceIn some of these models development of the NMJ is grossly dis-rupted which could contribute to their failure to undergo synapseelimination on a normal schedule While similar gross develop-mental disruption is not present at MHCI-deficient NMJs we can-not rule out the possibility that loss of MHCI perturbs synapseelimination indirectly through changes in aspects of developmentthat were not assessed With most manipulations however uni-form monoinnervation is still established within the first few post-natal weeks while multiple innervation is still evident at one yearof age in MHCI-deficient mice suggesting that synapse eliminationis persistently disrupted
We detect MHCI protein at the developing NMJ where it colo-calizes with nAChR labeling suggesting that it is expressed in themuscle fiber Importantly MHCI may also be expressed in themotor neuron andor Schwann cell during development andor inaging a possibility we cannot exclude because there are currentlyno antibodies to detect most members of the large MHCI familyMultiple MHCI mRNAs are expressed at the NMJ at different ages(Tetruashvily et al in press) MHCI protein has been detected inpresynaptic motor neurons in adult mice (Thams et al 2009)and in human and rat Schwann cells in vitro (Armati et al 1990Samuel et al 1987) Furthermore we find that modest overexpres-sion of the MHCI H2-D in neurons (Rall et al 1995) is sufficient tomodestly accelerate synapse elimination (Fig 2C) Ultimatelyimmuno-electron or super-resolution microscopy coupled withdevelopment of dozens of new antibodies will be necessary tofully characterize the pre- vs post-synaptic expression of the many
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
MHCI proteins Nevertheless the current results show that MHCI ispresent at the developing NMJ when synapse elimination isoccurring
MHCI is thought to be repressed by healthy muscle cells andupregulated only after injury or in disease states (Karpati et al1988) The current results demonstrate an unexpected role forMHCI at the healthy developing NMJ Remarkably MHCIrsquos functionat the developing NMJ is strikingly different to its role after injuryReducing MHCI expression exacerbates loss of synapses made ontoaxotomized motor neurons (Berg et al 2013 Oliveira et al 2004)while we find that both developmental and aging-related loss ofmotor neuron synapses onto muscles are inhibited
The bulk of excess MN synapses are pruned prior to P7 (Tapiaet al 2012) but mono-innervation is largely established betweenP7 and P15 as MNs begin to fire asynchronously (Buffelli et al2002 Personius et al 2001) In MHCI-deficient mice 15ndash20of postsynaptic sites retain two inputs at P15 No postsynaptic sitesreceived three or more inputs at P15 in MHCI-deficient animalsconsistent with the fact that very few postsynaptic sites stillreceive 3 or more inputs at P7 in WTs (Fig S2F) the age whenMHCIrsquos effects on synapse elimination begin to emerge The failureof circuit maturation in MHCI-deficient animals is not associatedwith gross motor defects in mice but could prevent the emergenceof fine-motor control that is behaviorally relevant in humansStrikingly the extent of the synapse elimination deficit at theMHCI-deficient NMJ is similar to the extent of axon pruning defi-cits in the lateral geniculate nucleus of mice lacking MHCI(Datwani et al 2009 Huh et al 2000 Lee et al 2014) or lackingneuron pentraxin (Bjartmar et al 2006) or components of thecomplement cascade (Stevens et al 2007) The reason for the par-tial nature of the pruning failure in the visual system in these mod-els remains unknown At the NMJ however we find that MHCI isspecifically required for the final stages of synapse eliminationduring the second postnatal week (Fig 1) In adult motor neuronslevels of the classical MHCI H-2D appear to be heterogeneous(Thams et al 2009) raising the possibility that different levels ofMHCI during development may also render some inputs more sus-ceptible to elimination Alternatively other factors including fibertype and associated activity patterns may influence the role that
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
Fullydenervated
15
10
5
0 β2m--
TAP--WT
3
2
1
0 β2m--
TAP--WT
E
Partially denervated
AChR
s (α
-btx
)
B C
SV2
5
4
D
mer
ge
A
MH
CI
P1540 80 160μg
MH
CI
2 Y O40 80 160μg
p
artia
lly d
ener
vate
d
fu
lly d
ener
vate
d
P15 2 Y OMH
CI l
abel
ing
in E
DL
(nor
mal
ized
to G
APD
H)
0
05025
07510
Fig 4 MHCI protein levels climb in aging EDL and reducing MHCI expression prevents aging-related denervation (A) Western blots for MHCI in lysates of EDL muscle fromP15 and 2-year-old mice Three different total protein concentrations are shown Blots were stripped and probed for GAPDH as a loading control Bottom MHCI densitometrynormalized to GAPDH for six technical replicates (two per concentration) at each age p = 00048 (B C) EDL muscles from 24ndash26 month-old mice were stained with a-btx(red) to detect nAChRs and SV2 (green) to detect presynaptic nerve terminals Representative images show examples of partially (arrows B) and fully (oval C) denervatedAChR clusters in WT EDL (red with no associated green in merge) Scales 7 lm (D E) Mean partial (D) and full (E) denervation in 2 year old EDL muscle from WT and MHCI-deficient b2mTAP mice (n = 4 per genotype) p = 003 p = 00009
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 7
MHCI plays in elimination of specific axon branches This is consis-tent with the finding that MHCI is required for synapse eliminationto reach completion in the diaphragm which consists of a mixtureof fast and slow fiber types but has a smaller influence on synapseelimination in the EDL which is almost exclusively composed offast-twitch fibers (Eddinger et al 1985) Together the currentresults suggest that synapse elimination at the NMJ may be dividedinto stages with distinct molecular requirements and that MHCI isessential as the last postsynaptic sites transition from dual- tomono-innervation
During synapse elimination at the NMJ competing axonsdevelop a disparity in synaptic strengthwhich precedeswithdrawalof the weaker input (Colman et al 1997 Kopp et al 2000) While itis unknown if larger synaptic strength at the MHCI-deficient NMJ isrelated to deficits in synapse elimination stronger inputs may beless vulnerable to competition-induced synaptic weakening In thisway loss of MHCI might prevent the development of sufficient dis-parity in synaptic strength to trigger functional silencing and phys-ical elimination in the most closely-matched competitions
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Interestingly paired increases in synapse strength and density likethose we observe at the MHCI-deficient NMJ are also seen in thecortex (Glynn et al 2011) and lateral geniculate nucleus (LGN)(Lee et al 2014) of MHCI-deficient mice Additional studies inMHCI-deficient mice found increases in synapse density in cortex(Adelson et al 2014) and increased synapse density (Dixon-Salazar et al 2014) and strength (Fourgeaud et al 2010) in hip-pocampus While the present study identifies a role for MHCI insynapse elimination at the NMJ it is as yet unknown if MHCI inhi-bits synapse formation andor promotes synapse elimination incortex or hippocampus
It will be important to determine if MHCI expression is regu-lated by activity at the developing NMJ as it is in the visual system(Corriveau et al 1998) to determine if it is specifically responsiveto the levels patterns andor synchronization of activity and tofully define the cellular and molecular networks linking MHCI tochanges in the strength and connectivity of synapses Animmunoreceptor for MHCI PirB has been detected in the CNS(Syken et al 2006) and at the adult NMJ (Thams et al 2009)
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

Fullydenervated
15
10
5
0 β2m--
TAP--WT
3
2
1
0 β2m--
TAP--WT
E
Partially denervated
AChR
s (α
-btx
)
B C
SV2
5
4
D
mer
ge
A
MH
CI
P1540 80 160μg
MH
CI
2 Y O40 80 160μg
p
artia
lly d
ener
vate
d
fu
lly d
ener
vate
d
P15 2 Y OMH
CI l
abel
ing
in E
DL
(nor
mal
ized
to G
APD
H)
0
05025
07510
Fig 4 MHCI protein levels climb in aging EDL and reducing MHCI expression prevents aging-related denervation (A) Western blots for MHCI in lysates of EDL muscle fromP15 and 2-year-old mice Three different total protein concentrations are shown Blots were stripped and probed for GAPDH as a loading control Bottom MHCI densitometrynormalized to GAPDH for six technical replicates (two per concentration) at each age p = 00048 (B C) EDL muscles from 24ndash26 month-old mice were stained with a-btx(red) to detect nAChRs and SV2 (green) to detect presynaptic nerve terminals Representative images show examples of partially (arrows B) and fully (oval C) denervatedAChR clusters in WT EDL (red with no associated green in merge) Scales 7 lm (D E) Mean partial (D) and full (E) denervation in 2 year old EDL muscle from WT and MHCI-deficient b2mTAP mice (n = 4 per genotype) p = 003 p = 00009
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 7
MHCI plays in elimination of specific axon branches This is consis-tent with the finding that MHCI is required for synapse eliminationto reach completion in the diaphragm which consists of a mixtureof fast and slow fiber types but has a smaller influence on synapseelimination in the EDL which is almost exclusively composed offast-twitch fibers (Eddinger et al 1985) Together the currentresults suggest that synapse elimination at the NMJ may be dividedinto stages with distinct molecular requirements and that MHCI isessential as the last postsynaptic sites transition from dual- tomono-innervation
During synapse elimination at the NMJ competing axonsdevelop a disparity in synaptic strengthwhich precedeswithdrawalof the weaker input (Colman et al 1997 Kopp et al 2000) While itis unknown if larger synaptic strength at the MHCI-deficient NMJ isrelated to deficits in synapse elimination stronger inputs may beless vulnerable to competition-induced synaptic weakening In thisway loss of MHCI might prevent the development of sufficient dis-parity in synaptic strength to trigger functional silencing and phys-ical elimination in the most closely-matched competitions
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Interestingly paired increases in synapse strength and density likethose we observe at the MHCI-deficient NMJ are also seen in thecortex (Glynn et al 2011) and lateral geniculate nucleus (LGN)(Lee et al 2014) of MHCI-deficient mice Additional studies inMHCI-deficient mice found increases in synapse density in cortex(Adelson et al 2014) and increased synapse density (Dixon-Salazar et al 2014) and strength (Fourgeaud et al 2010) in hip-pocampus While the present study identifies a role for MHCI insynapse elimination at the NMJ it is as yet unknown if MHCI inhi-bits synapse formation andor promotes synapse elimination incortex or hippocampus
It will be important to determine if MHCI expression is regu-lated by activity at the developing NMJ as it is in the visual system(Corriveau et al 1998) to determine if it is specifically responsiveto the levels patterns andor synchronization of activity and tofully define the cellular and molecular networks linking MHCI tochanges in the strength and connectivity of synapses Animmunoreceptor for MHCI PirB has been detected in the CNS(Syken et al 2006) and at the adult NMJ (Thams et al 2009)
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

8 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
but knocking out PirB does not replicate the failure of retino-geniculate pruning seen in MHCI-deficient mice (Datwani et al2009 Huh et al 2000 Lee et al 2014 Syken et al 2006) suggest-ing that in the retino-geniculate projection MHCI promotes devel-opmental synapse elimination via a PirB-independent mechanismIt remains an open question whether PirB is involved in MHCI-dependent changes in synapse number at the NMJ or in otherregions of the CNS such as hippocampus and cortex Overall adetailed molecular mechanistic understanding of MHCI signalingin the nervous system is still lacking The current study opensthe door to advance this critical research at the NMJ a large acces-sible synapse that is uniquely amenable to manipulation and imag-ing in vivo
In MHCI-deficient animals aging-related denervation of EDLmuscle fibers is decreased The characteristic fragmentation orabsence of pre- andor post-synaptic staining at some NMJs in agedanimals is unlikely to be due to incomplete labeling for severalreasons (1) similar discontinuities are never observed in youngermuscle (2) in 2-year-old muscle most postsynaptic sites are fullylabeled and (3) the percentage of denervated muscle fibers mea-sured in WTs corresponds well with published values for this mus-cle (eg (Chai et al 2011 Valdez et al 2012)) The relativeabsence of denervation in MHCI-deficient animals cannot be fullyexplained by the persistence of supernumary inputs and preferen-tial loss of these excess inputs with aging because developmentalsynapse elimination while impaired is still largely complete atP15 in the EDL in MHCI-deficient animals
In adults MHCI levels increase in situations where muscle fiberdenervation occurs including nerve injury neuromuscular disor-ders and normal aging (Edstrom et al 2004 Emslie-Smith et al1989 Thams et al 2009) MHCI expression is notably low inMNs innervating the extra-ocular muscles (EOMs) (Linda et al1999) which are resistant to aging-associated denervation(Valdez et al 2012) Here we provide evidence that MHCI proteinlevels rise at the aging NMJ and that MHCI promotes denervationof aging muscle Recent studies also identified a role for MHCI andthe MHCI light chain b2m in cognitive aging (Smith et al 2015)While we find that MHCI is selectively required for the final stagesof synapse elimination at the developing NMJ denervation of agingmuscle is almost completely prevented in MHCI-deficient micesuggesting that vulnerability to aging-related denervation largelydepends on MHCI Together the current results show that MHCIpromotes weakening and elimination of surplus synapses at thedeveloping NMJ and suggest that aberrant reactivation of MHCI-dependent synapse elimination mechanisms may contribute topathological denervation of adult muscle They also identify MHCIas a therapeutic target for the prevention of aging-related motorimpairments
4 Materials and methods
41 Animal subjects
All experimental protocols were conducted according to theNational Institutes of Health (NIH) guidelines for animal researchand approved by the Institutional Animal Care and Use Committee(IACUC) at the University of California San Diego andor PrincetonUniversity The following mouse strains were used (1) C57B16wild-type (WT HarlanJackson Labs) (2) mice lacking expressionof the MHCI light chain b2m as well as the transporter for antigenprocessing TAP1 which show greatly reduced stable cell-surfaceexpression of most but not all MHCI proteins (b2mTAPcourtesy of D Raulet and CJ Shatz) (Dorfman et al 1997Ljunggren et al 1995) (3) mice which 128 and lack expressionof both of the classical MHCI genes present in C57Bl6 H-2K and
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
H-2D (K bDb courtesy of CJ Shatz) (Vugmeyster et al1998) (4) mice which lack expression of single classical MHCIgenes H-2K or H-2D (K b and Db Taconic) (Perarnau et al1999) and (5) mice which overexpress the classical MHCI H-2Db
under the control of the neuron-specific enolase promoter (NSE-Db courtesy of MBA Oldstone) (Rall et al 1995)
42 Visualization of neuromuscular junctions usingimmunohistochemistry and rhodamine-conjugated a-bungarotoxin(a-btx)
Whole mounts of mouse diaphragm muscle were immunos-tained at P0 P7 P15 P29-50 12ndash13 months or 22ndash24 monthsand mouse EDL was stained at P15 and 22ndash24 months Animalswere deeply anesthetized using inhaled isoflurane and tissue wasdrop-fixed with 4 PFA (Electron Microscopy Systems) in phos-phate buffered saline (PBS Sigma) for 30 min Diaphragm and EDLmuscles were dissected rinsed in PBS incubated in 01 glycine(Fischer) for 12 h at 4 C and then blocked in a solution containing3 goat serum albumin (GSA Jackson) and 01 Triton X-100 in PBSfor 12 h at 4 C both on rockers Presynaptic MN terminals werevisualized by treating muscles for 48ndash72 h at 4 C with a primaryantibody cocktail consisting of one or both of the following anti-neurofilament (Invitrogen 1500) and anti-SV2 (developed by KM Buckley under the auspices of the NICHD maintained by TheUniversity of Iowa Department of Biology obtained from the Devel-opmental Studies Hybridoma Bank 1100) Tissues were rinsed inPBS for 3x2hrs and then incubated in Alexa-488-conjugated goatanti-mouse IgG (Jackson 1100) for 18ndash24 h at 4 C Tissues wererinsed in PBS for 3 2 h and treated with rhodamine-conjugateda-btx (Molecular Probes 10 lg100 lL) to label postsynaptic nico-tinic acetylcholine receptors for 30 min After a final rinse in PBStissues were mounted on slides using fluorescence-stabilizingmounting media (Vectashield hard set mount) and microscopywas performed using a Leica (Wetzlar Germany) DMI6000 invertedmicroscope outfitted with a Yokogawa (Tokyo Japan) spinning diskconfocal head and an Orca ER high resolution BampW cooled CCDcamera (Hamamatsu Sewickley PA) This approach yielded robustimmunostaining that allowed clear discrimination of motor neuronaxons and presynaptic motor end plates To ensure maximal anti-body penetration we examined surface fibers at approximatelythe same depth in all tissues
Importantly the appearance of presynaptic immunofluores-cence is expected to differ qualitatively from presynaptic fluores-cence in mouse lines expressing YFP in a subset of motor neuronswhich have been used in many influential studies of the develop-ment and denervation of the NMJ (eg (Valdez et al 2010)) Thisis because antibodies gradually penetrate the tissue to label specificpresynaptic structures (SV2 synaptic vesicles NF neurofilament)while YFP is a soluble protein that when overexpressed in motorneurons diffuses throughout the presynaptic cytoplasm
To assess multiple innervation montages of Z-serial images(lt20 lm total thickness per junction) were collected from intactmuscles under a 63 oil immersion lens at 16 magnificationZ-stacks of at least 50 terminals from each hemidiaphragm orEDL muscle (two per animal each animal was considered an inde-pendent n) were examined from each animal at each age by anobserver blind to genotype Junctions were uniformly sampledover the entire dorsalndashventral axis Innervation was assessed bycounting the number of anatomically distinct MN axons contactingeach postsynaptic junction by tracing them back as far as possiblein unflattened confocal stacks (54ndash27 lm total depth) In mostcases preterminal axons could be followed 50ndash100 lm from thesynapse Examination of unflattened confocal Z-stacks from multi-ple depths and angles is necessary to unambiguously distinguishindividual MN axons innervating a single postsynaptic site and
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 9
to determine if two axons innervate one postsynaptic site or sim-ply two postsynaptic sites in different focal planes Junctions inwhich multiple innervation could not be unambiguously deter-mined (eg due to extensive axon fasciculation variation in label-ing intensity proximity of other nAChR clusters or depth of axons)were scored as monoinnervated to ensure a conservative estimateof multiple innervation Postsynaptic perimeter and area wereassessed and quantified using ImageJ software (NIH)
For denervation studies in aged EDL montages of Z-serialimages (lt20 lm total thickness per junction) were collected fromintact muscles under a 63 oil immersion lens at 16 magnifica-tion Z-stacks of at least 50 terminals from each EDL (two EDLs peranimal each animal was considered an independent n) wereexamined from each animal by an observer blind to genotypeJunctions were uniformly sampled over the entire muscle Innerva-tion of a muscle fiber was defined as colocalization of green SV2-labeled presynaptic axon terminals with a-btx-labeled postsynap-tic nAChRs Both partial and full denervation were scored as iden-tified as nAChR labeling not fully apposed to presynaptic SV2labeling the same criteria used in (Valdez et al 2010) (Fig 4)Most denervated NMJs also showed one or more additionalaging-associated changes including (1) nAChR fragmentation(gt5 nAChR islands abnormal nAChR cluster shape) and decreasednAChR density (noticeably dimmer a-btx labeling compared toother muscle fibers in the same confocal plane) (2) retraction ofthe MN axon multiple innervation and extra-junctional MNsprouting and (3) axonal varicosities proximal to the NMJ and axo-nal atrophy (thinning of the preterminal or terminal axon)
43 MHCI and synaptophysin immunohistochemistry
Anesthetized P7 WT and b2mTAP animals were perfusedtranscardially with 4 PFA Diaphragms were dissected and storedat 4 C overnight Muscles were cleaned of connective tissue andblocked in a solution containing 5 bovine serum albumin 001Triton X-100 and 01 sodium azide (Fischer) for one hour Muscleswere treated with primary antibodies against synaptophysin(SYPH Santa Cruz 1100) and MHCI (OX18 AbdSerotec 1100)overnight at 4 C OX18 is a mouse monoclonal antibody that rec-ognizes a monomorphic epitope in the a3 region of the rat MHCIRT-1A (Fukumoto et al 1982) and reacts with mouse MHCI inWestern blots and immunohistochemistry (Corriveau et al 1998Datwani et al 2009 Dixon-Salazar et al 2014 Goddard et al2007 Huh et al 2000) The suspected mouse orthologue of ratRT-1A H-2K1 is 849 identical and 892 similar to RT-1A atthe amino acid level in the a3 region the site of the OX-18 epitopeSeveral lines of evidence support OX-18rsquos ability to bind to MHCI inthe mouse nervous system First genetically deleting the MHCIlight chain b2m reduces the amount of MHCI that reaches the cellsurface (Dorfman et al 1997 Zijlstra et al 1989) and cell surfaceOX-18 immunofluorescence is greatly attenuated in both b2m
(Needleman et al 2010) and b2mTAP (Goddard et al2007) neurons in vitro Second OX-18 recognizes proteins of theexpected molecular weight in western blots of adult mouse brain(Corriveau et al 1998 Dixon-Salazar et al 2014 Huh et al2000) and similar labeling is seen in rat brain using a rabbit poly-clonal antibody that recognizes a distinct epitope of MHCI(Needleman et al 2010) The secondary antibodies we used todetect MHCI cause some nonspecific background labeling whichcan be clearly distinguished from specific labeling in two ways(1) it is still present when labeling with an isotype-control anti-body and (2) it is not abolished in MHCI-deficient animals(Fig S1) Using these criteria specific MHCI labeling is present atthe postsynaptic site while other muscle labeling is nonspecificTaken together these results provide strong support for the speci-ficity of OX-18 in recognizing MHCI at the mouse NMJ
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Incubation in primary antibody was followed by 3 40 minrinses in PBS and a four hour treatment with the appropriate cock-tail of Alexa-488-conjugated donkey anti-mouse IgG (Invitrogen11000) andor Cy5-conjugated donkey anti-goat IgG (Jackson1100) in block After rinsing 3 40 min muscles were treatedfor 30 min with rhodamine-conjugated a-bungarotoxon (a-btx)diluted 110 in ddH2O Samples were rinsed in 3 20 min in PBSmounted on glass slides and coverslipped using Vectashield hard-set mount Coverslipped slides were sandwiched between twopieces of paper towel and compressed under a lead brick overnightedges sealed with clear nail polish and stored at 4 C until imagingZ-serial images (05 lm per step up to 20 lm total depth per junc-tion) were collected from intact muscles using a Leica (WetzlarGermany) DMI6000 inverted microscope outfitted with a Yoko-gawa (Tokyo Japan) spinning disk confocal head and an Orca ERhigh resolution BampW cooled CCD camera (Hamamatsu SewickleyPA) under a 63 oil immersion lens at 16 magnification WTand b2mTAP samples were imaged under identical condi-tions during the same microscopy session
44 Western blotting
EDL muscles were dissected as above from P15 and aged (2 yearold) WT mice (2 animals per age) Pooled tissue for each age wassuspended in 500 ll of lysis buffer (50 mMNaH2PO4 300 mMNaCl10 mM imidazole 1 NP-40 05 TritonX-100 Halt protease andphosphatase inhibitor (Pierce Rockland IL) and flash-frozen in liq-uid nitrogen Frozen tissue was placed in pre-cooled 50 ml grindingjars along with a 20 mm stainless steel ball bearing and subjectedto cryogenic grinding in liquid nitrogen for 20 min at 25 Hz on aRetsch CryoMill until rendered into a fine powder Total proteinconcentration in the samples was determined using a NanoDropND-1000 UVndashVis spectrophotometer from Thermo (absorbance(A280) was measured) Samples (40 80 or 160 lglane) were sub-jected to SDSndashPAGE using 4ndash20 Mini-PROTEAN TGXTM gradientgels from BIORAD and transferred to a PVDF membrane
Membranes were blocked for 1 h at room temperature (RT25 C) in 5 organic dry milk in TBST Primary antibodies (anti-MHCI [p8 1500 purified rabbit polyclonal raised against exon 8of H-2K GeneScript] or anti-GAPDH [MAB374 15000 purifiedmouse monoclonal raised against rabbit GAPDH EMD Millipore])were diluted in blocking buffer and incubated with the membranesovernight at 4 C (p8) or 1 h at RT (GAPDH) Membranes werewashed 12 5 min in TBST followed by 1 h incubation at RT withthe appropriate HRP-conjugated secondary antibody (for MHCIgoat anti-rabbit HRP [15000] from Jackson ImmunoResearch Lab-oratories product 111035144 for GAPDH goat anti-mouseHRP [110000] from ThermoFisher product A24512) Mem-branes were washed 12 5 min in TBST and labeling detectedusing either Pierce ECL Western Blotting Substrate (Pierce Rock-land IL catalog 32106) (for GAPDH) or SuperSignal West PicoChemiluminescent Substrate (Pierce Rockland IL catalog 34080) (for MHCI) and used to expose HyBlot CL AutoradiographyFilm (Denville Scientific Metuchen NJ) Blots were first probed forMHCI and then stripped and reprobed for GAPDH Blots werestripped using a mild stripping solution (1L of water containing15 g glycine 1 g SDS 10 ml Tween 20 pH 22) heated to 55 CBlots were incubated in 20 ml of stripping solution for 10 minwashed 12 5 min in TBST and blocked and processed as abovefor the second antibody (anti-MHCI)
45 Electrophysiology
Hemidiaphragm-phrenic nerve preparations were used fromWT and b2mTAP animals at P14-16 and adult (P29-P45) agesMuscles were dissected from anesthetized animals under cold
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

10 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
oxygenated (95 CO25 O2) Rees normal mammalian ringers(NMR (Rees 1978)) consisting of (in mM) 135 NaCl 5 KCl 15NaHCO3 1 Na2HPO4 1 MgSO4 25 Ca gluconate and 11 glucosepH 74 Contraction blockade which is necessary to permit micro-electrode recordings from postsynaptic muscle fibers wasachieved via 1 h pretreatment of diaphragms in oxygenated 2 lMl-conotoxin (Bachem) (adult muscles) or 3ndash6 lM tubocurare(P15) Muscles were pinned to a Sylgard-lined recording chamberand superfused (1 mLmin) with oxygenated NMR at 22ndash24 CIntracellular recordings of muscles fibers were performed usingsharp recording electrodes (30ndash69 MX) filled with 3 M KCl Endplate potentials (EPPs) were evoked via suprathreshold stimulationof the phrenic nerve with a suction electrode (square pulses 02 msduration at 05 Hz) Electrodes were visually guided to NMJs underoblique illumination Responses with rise times of more than15 ms represent electrode placements relatively distant from thejunction and were discarded Membrane potentials were continu-ously recorded and only muscle fibers in which potentials werelt55 mV and stable within plusmn5 mV were included in analysisMembrane responses were amplified by an Axoclamp 2A amplifier(Axon Instruments) low-pass filtered at 1 kHz and digitized andrecorded using WinWCP software (John Dempster)
Levels of innervation were assessed electrophysiologically byapplying graded stimulation to the phrenic nerve stump andcounting the number of elicited inputs of differential amplitudeabove normal quantal variability Miniature end-plate potentials(MEPPS) were recorded in gap-free mode from singly-innervatedjunctions and analyzed offline using MiniAnalysis software (JustinLee Synaptosoft) For quantal content analysis EPPS were evokedand MEPPs were recorded for two seconds following stimulation atsingly-innervated junctions Quantal content was estimated as theratio of the mean EPP amplitude to the mean MEPP amplitude(Wood and Slater 2001)
46 Antibody injections
P7 WT mice received daily intraperitoneal injections for 7 dayswith 10 lgg body weight of either panspecific anti-MHCI antibody(OX18 AbdSerotec) or isotype-matched mouse IgG1 control (Abd-Serotec) While injecting one specific antibody against H-2d caninduce a model of transfusion-induced lung injury in rats(Looney et al 2006) nine other anti-MHCI antibodies includingOX18 do not induce this model (Kelher et al 2009 Strait et al2011) P15 mice were perfused and diaphragms were eitherimmunostained for neuromuscular morphology as describedabove or treated with Alexa 488-conjugated goat anti-mouseIgG1 (Invitrogen 1100) for four hours to visualize OX18 or IgG1binding
47 Data analysis
All immunohistochemical experiments were collected and ana-lyzed blind to genotype Data are presented as mean plusmn standarderror of the mean p Values determined by unpaired one-tailed Stu-dentrsquos t-test Unless otherwise noted asterisks indicate p lt 005and n = number of animals
Acknowledgments
We thank E OrsquoDriscoll K K Frietze R Wyatt and JJ Park fortechnical support R Balice-Gordon for protocols and discussionG Patrick and M Scanziani for loaned equipment D Raulet andC Shatz for transgenic mice and A Ghosh and C Tyler forcomments Electrophysiological experiments piloted in UCSDTeaching Labs with L Lopez Supported by 1F30AG046044-01A1the Princeton Department of Molecular Biology and the Princeton
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Neuroscience Institute (MMT) the UCSD School of Medicine(MAM) the New Jersey Commission on Brain Injury Research(KKF) the Alfred P Sloan Foundation the Whitehall Foundationand a Princeton Neuroscience Institute Innovation Award (LMB)
Appendix A Supplementary data
Supplementary data associated with this article can be found inthe online version at httpdxdoiorg101016jbbi201601008
References
Adelson JD Barreto GE Xu L Kim T Brott BK Ouyang YB Naserke TDjurisic M Xiong X Shatz CJ et al 2012 Neuroprotection from stroke in theabsence of MHCI or PirB Neuron 73 1100ndash1107 S0896-6273(12)00138-9 [pii]101016jneuron201201020
Adelson JD Sapp RW Brott BK Lee H Miyamichi K Luo L Cheng S DjurisicM Shatz CJ 2014 Developmental sculpting of intracortical circuits by MHCClass I H2-Db and H2-Kb Cereb Cortex httpdxdoiorg101093cercorbhu243
Armati PJ Pollard JD Gatenby P 1990 Rat and human Schwann cells in vitrocan synthesize and express MHC molecules Muscle Nerve 13 106ndash116 httpdxdoiorg101002mus880130204
Berg A Zelano J Thams S Cullheim S 2013 The extent of synaptic stripping ofmotoneurons after axotomy is not correlated to activation of surrounding gliaor downregulation of postsynaptic adhesion molecules PLoS One 8 e59647httpdxdoiorg101371journalpone0059647
Bjartmar L Huberman AD Ullian EM Renteria RC Liu X Xu W Prezioso JSusman MW Stellwagen D Stokes CC et al 2006 Neuronal pentraxinsmediate synaptic refinement in the developing visual system J Neurosci 266269ndash6281 httpdxdoiorg101523JNEUROSCI4212-052006
Blom WM De Bont HJ Meijerman I Kuppen PJ Mulder GJ Nagelkerke JF1999 Interleukin-2-activated natural killer cells can induce both apoptosis andnecrosis in rat hepatocytes Hepatology 29 785ndash792 S0270913999001202 [pii]101002hep510290303
Bogdanik LP Chapman HD Miers KE Serreze DV Burgess RW 2012 AMusD retrotransposon insertion in the mouse Slc6a5 gene causes alterations inneuromuscular junction maturation and behavioral phenotypes PLoS One 7e30217 httpdxdoiorg101371journalpone0030217 PONE-D-11-20438[pii]
Brown MC Jansen JK Van Essen D 1976 Polyneuronal innervation of skeletalmuscle in new-born rats and its elimination during maturation J Physiol 261387ndash422
Buffelli M Busetto G Cangiano L Cangiano A 2002 Perinatal switch fromsynchronous to asynchronous activity of motoneurons link with synapseelimination Proc Natl Acad Sci U S A 99 13200ndash13205 httpdxdoiorg101073pnas202471199
Buffelli M Burgess RW Feng G Lobe CG Lichtman JW Sanes JR 2003Genetic evidence that relative synaptic efficacy biases the outcome of synapticcompetition Nature 424 430ndash434 httpdxdoiorg101038nature01844nature01844 [pii]
Busetto G Buffelli M Tognana E Bellico F Cangiano A 2000 Hebbianmechanisms revealed by electrical stimulation at developing rat neuromuscularjunctions J Neurosci 20 685ndash695
Chai RJ Vukovic J Dunlop S Grounds MD Shavlakadze T 2011 Strikingdenervation of neuromuscular junctions without lumbar motoneuron loss ingeriatric mouse muscle PLoS One 6 e28090 httpdxdoiorg101371journalpone0028090 PONE-D-11-11057 [pii]
Colman H Nabekura J Lichtman JW 1997 Alterations in synaptic strengthpreceding axon withdrawal Science 275 356ndash361
Corriveau RA Huh GS Shatz CJ 1998 Regulation of class I MHC geneexpression in the developing and mature CNS by neural activity Neuron 21505ndash520
Datwani A McConnell MJ Kanold PO Micheva KD Busse B Shamloo MSmith SJ Shatz CJ 2009 Classical MHCI molecules regulate retinogeniculaterefinement and limit ocular dominance plasticity Neuron 64 463ndash470 S0896-6273(09)00844-7 [pii] 101016jneuron200910015
Del Castillo J Katz B 1954 Quantal components of the end-plate potential JPhysiol 124 560ndash573
Dixon-Salazar TJ Fourgeaud L Tyler CM Poole JR Park JJ Boulanger LM2014 MHC class I limits hippocampal synapse density by inhibiting neuronalinsulin receptor signaling J Neurosci 34 11844ndash11856 httpdxdoiorg101523JNEUROSCI4642-122014
Dorfman JR Zerrahn J Coles MC Raulet DH 1997 The basis for self-toleranceof natural killer cells in beta2-microglobulin- and TAP-1- mice J Immunol 1595219ndash5225
Eddinger TJ Moss RL Cassens RG 1985 Fiber number and type composition inextensor digitorum longus soleus and diaphragm muscles with aging in Fisher344 rats J Histochem Cytochem 33 1033ndash1041
Edstrom E Kullberg S Ming Y Zheng H Ulfhake B 2004 MHC class I beta2microglobulin and the INF-gamma receptor are upregulated in agedmotoneurons J Neurosci Res 78 892ndash900
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx 11
Emslie-Smith AM Arahata K Engel AG 1989 Major histocompatibility complexclass I antigen expression immunolocalization of interferon subtypes and Tcell-mediated cytotoxicity in myopathies Hum Pathol 20 224ndash231
Fourgeaud L Davenport CM Tyler CM Cheng TT Spencer MB Boulanger LM 2010 MHC class I modulates NMDA receptor function and AMPA receptortrafficking Proc Natl Acad Sci USA 107 22278ndash22283 0914064107 [pii]101073pnas0914064107
Fukumoto T McMaster WR Williams AF 1982 Mouse monoclonal antibodiesagainst rat major histocompatibility antigens Two Ia antigens and expression ofIa and class I antigens in rat thymus Eur J Immunol 12 237ndash243
Glynn MW Elmer BM Garay PA Liu XB Needleman LA El-Sabeawy FMcAllister AK 2011 MHCI negatively regulates synapse density during theestablishment of cortical connections Nat Neurosci 14 442ndash451 nn2764 [pii]101038nn2764
Goddard CA Butts DA Shatz CJ 2007 Regulation of CNS synapses by neuronalMHC class I Proc Natl Acad Sci U S A 104 6828ndash6833
Hayashi-Takagi A Araki Y Nakamura M Vollrath B Duron SG Yan Z KasaiH Huganir RL Campbell DA Sawa A 2014 PAKs inhibitors ameliorateschizophrenia-associated dendritic spine deterioration in vitro and in vivoduring late adolescence Proc Natl Acad Sci U S A 111 6461ndash6466 httpdxdoiorg101073pnas1321109111
Hedden T Gabrieli JD 2004 Insights into the ageing mind a view from cognitiveneuroscience Nat Rev Neurosci 5 87ndash96 httpdxdoiorg101038nrn1323
Huh GS Boulanger LM Du H Riquelme PA Brotz TM Shatz CJ 2000Functional requirement for class I MHC in CNS development and plasticityScience 290 2155ndash2159
Je HS Yang F Ji Y Potluri S Fu XQ Luo ZG Nagappan G Chan JPHempstead B Son YJ et al 2013 ProBDNF and Mature BDNF as Punishmentand Reward Signals for Synapse Elimination at Mouse NeuromuscularJunctions J Neurosci 33 9957ndash9962 33249957 [pii] 101523JNEUROSCI0163-132013
Karpati G Pouliot Y Carpenter S 1988 Expression of immunoreactive majorhistocompatibility complex products in human skeletal muscles Ann Neurol23 64ndash72 httpdxdoiorg101002ana410230111
Kelher MR Masuno T Moore EE Damle S Meng X Song Y Liang XNiedzinski J Geier SS Khan SY et al 2009 Plasma from stored packed redblood cells and MHC class I antibodies causes acute lung injury in a 2-eventin vivo rat model Blood 113 2079ndash2087 httpdxdoiorg101182blood-2008-09-177857
Keller-Peck CR Walsh MK Gan WB Feng G Sanes JR Lichtman JW 2001Asynchronous synapse elimination in neonatal motor units studies using GFPtransgenic mice Neuron 31 381ndash394 S0896-6273(01)00383-X [pii]
Kopp DM Perkel DJ Balice-Gordon RJ 2000 Disparity in neurotransmitterrelease probability among competing inputs during neuromuscular synapseelimination J Neurosci 20 8771ndash8779
Krey JF Pasca SP Shcheglovitov A Yazawa M Schwemberger R RasmussonR Dolmetsch RE 2013 Timothy syndrome is associated with activity-dependent dendritic retraction in rodent and human neurons Nat Neurosci 16201ndash209 nn3307 [pii] 101038nn3307
Lee YI Mikesh M Smith I Rimer M Thompson W 2011 Muscles in a mousemodel of spinal muscular atrophy show profound defects in neuromusculardevelopment even in the absence of failure in neuromuscular transmission orloss of motor neurons Dev Biol 356 432ndash444 S0012-1606(11)00982-1 [pii]101016jydbio201105667
Lee H Brott BK Kirkby LA Adelson JD Cheng S Feller MB Datwani AShatz CJ 2014 Synapse elimination and learning rules co-regulated by MHCclass I H2-D Nature 509 195ndash200 httpdxdoiorg101038nature13154
Letellier M Willson ML Gautheron V Mariani J Lohof AM 2008 Normaladult climbing fiber monoinnervation of cerebellar Purkinje cells in micelacking MHC class I molecules Dev Neurobiol 68 997ndash1006
Li MX Jia M Yang LX Jiang H Lanuza MA Gonzalez CM Nelson PG 2004The role of the theta isoform of protein kinase C (PKC) in activity-dependentsynapse elimination evidence from the PKC theta knock-out mouse in vivo andin vitro J Neurosci 24 3762ndash3769 httpdxdoiorg101523JNEUROSCI3930-032004 24153762 [pii]
Linda H Hammarberg H Piehl F Khademi M Olsson T 1999 Expression ofMHC class I heavy chain and beta2-microglobulin in rat brainstemmotoneurons and nigral dopaminergic neurons J Neuroimmunol 101 76ndash86
Ljunggren HG Van Kaer L Sabatine MS Auchincloss Jr H Tonegawa S PloeghHL 1995 MHC class I expression and CD8+ T cell development in TAP1beta 2-microglobulin double mutant mice Int Immunol 7 975ndash984
Looney MR Su X Van Ziffle JA Lowell CA Matthay MA 2006 Neutrophilsand their Fc gamma receptors are essential in a mouse model of transfusion-related acute lung injury J Clin Invest 116 1615ndash1623 httpdxdoiorg101172JCI27238
McConnell MJ Huang YH Datwani A Shatz CJ 2009 H2-K(b) and H2-D(b)regulate cerebellar long-term depression and limit motor learning Proc NatlAcad Sci USA 106 6784ndash6789 0902018106 [pii] 101073pnas0902018106
Misgeld T Burgess RW Lewis RM Cunningham JM Lichtman JW Sanes JR2002 Roles of neurotransmitter in synapse formation development ofneuromuscular junctions lacking choline acetyltransferase Neuron 36 635ndash648
Needleman LA Liu XB El-Sabeawy F Jones EG McAllister AK 2010 MHCclass I molecules are present both pre- and postsynaptically in the visual cortexduring postnatal development and in adulthood Proc Natl Acad Sci USA 10716999ndash17004 1006087107 [pii] 101073pnas1006087107
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Nelson PA Sage JR Wood SC Davenport CM Anagnostaras SG Boulanger LM 2013 MHC class I immune proteins are critical for hippocampus-dependentmemory and gate NMDAR-dependent hippocampal long-term depressionLearn Mem 20 505ndash517
Nguyen QT Lichtman JW 1996 Mechanism of synapse disassembly at thedeveloping neuromuscular junction Curr Opin Neurobiol 6 104ndash112 S0959-4388(96)80015-8 [pii]
Nguyen QT Parsadanian AS Snider WD Lichtman JW 1998Hyperinnervation of neuromuscular junctions caused by GDNFoverexpression in muscle Science 279 1725ndash1729
Noutel J Hong YK Leu B Kang E Chen C 2011 Experience-dependentretinogeniculate synapse remodeling is abnormal in MeCP2-deficient miceNeuron 70 35ndash42 S0896-6273(11)00167-X [pii] 101016jneuron201103001
OrsquoBrien RA Ostberg AJ Vrbova G 1978 Observations on the elimination ofpolyneuronal innervation in developing mammalian skeletal muscle J Physiol282 571ndash582
Oliveira AL Thams S Lidman O Piehl F Hokfelt T Karre K Linda HCullheim S 2004 A role for MHC class I molecules in synaptic plasticity andregeneration of neurons after axotomy Proc Natl Acad Sci USA 101 17843ndash17848
Pennefather P Quastel DM 1981 Relation between subsynaptic receptorblockade and response to quantal transmitter at the mouse neuromuscularjunction J Gen Physiol 78 313ndash344
Perarnau B Saron MF Reina San Martin B Bervas N Ong H Soloski MJSmith AG Ure JM Gairin JE Lemonnier FA 1999 Single H2Kb H2Db anddouble H2KbDb knockout mice peripheral CD8+ T cell repertoire and anti-lymphocytic choriomeningitis virus cytolytic responses Eur J Immunol 291243ndash1252 httpdxdoiorg101002(SICI)1521-4141(199904)2904lt1243AID-IMMU1243gt30CO2-A
Personius KE Balice-Gordon RJ 2001 Loss of correlated motor neuron activityduring synaptic competition at developing neuromuscular synapses Neuron 31395ndash408 S0896-6273(01)00369-5 [pii]
Personius K Chang Q Bittman K Panzer J Balice-Gordon R 2001 Gapjunctional communication among motor and other neurons shapes patterns ofneural activity and synaptic connectivity during development Cell CommunAdhes 8 329ndash333
Personius KE Chang Q Mentis GZ OrsquoDonovan MJ Balice-Gordon RJ 2007Reduced gap junctional coupling leads to uncorrelated motor neuron firing andprecocious neuromuscular synapse elimination Proc Natl Acad Sci USA 10411808ndash11813
Personius KE Karnes JL Parker SD 2008 NMDA receptor blockade maintainscorrelated motor neuron firing and delays synapse competition at developingneuromuscular junctions J Neurosci 28 8983ndash8992 28368983 [pii] 101523JNEUROSCI5226-072008
Pfeiffer BE Zang T Wilkerson JR Taniguchi M Maksimova MA Smith LNCowan CW Huber KM 2010 Fragile X mental retardation protein is requiredfor synapse elimination by the activity-dependent transcription factor MEF2Neuron 66 191ndash197 S0896-6273(10)00188-1 [pii] 101016jneuron201003017
Rafuse VF Polo-Parada L Landmesser LT 2000 Structural and functionalalterations of neuromuscular junctions in NCAM-deficient mice J Neurosci 206529ndash6539 20176529 [pii]
Rall GF Mucke L Oldstone MB 1995 Consequences of cytotoxic T lymphocyteinteraction with major histocompatibility complex class I-expressing neuronsin vivo J Exp Med 182 1201ndash1212
Redfern PA 1970 Neuromuscular transmission in new-born rats J Physiol 209701ndash709
Rees D 1978 A non-phosphate-buffered physiological saline for in vitroelectrophysiological studies on the mammalian neuromuscular junction[proceedings] J Physiol 278 8Pndash9P
Roche SL Sherman DL Dissanayake K Soucy G Desmazieres A Lamont DJPeles E Julien JP Wishart TM Ribchester RR et al 2014 Loss of glialneurofascin155 delays developmental synapse elimination at theneuromuscular junction J Neurosci 34 12904ndash12918 httpdxdoiorg101523JNEUROSCI1725-142014
Rolleke U Flugge G Plehm S Schlumbohm C Armstrong VW Dressel RUchanska-Ziegler B Ziegler A Fuchs E Czeh B et al 2006 Differentialexpression of major histocompatibility complex class I molecules in the brain ofa New World monkey the common marmoset (Callithrix jacchus) JNeuroimmunol 176 39ndash50 httpdxdoiorg101016jjneuroim200604015
Rudolf R Khan MM Labeit S Deschenes MR 2014 Degeneration ofneuromuscular junction in age and dystrophy Front Aging Neurosci 6 99httpdxdoiorg103389fnagi201400099
Samuel NM Mirsky R Grange JM Jessen KR 1987 Expression of majorhistocompatibility complex class I and class II antigens in human Schwann cellcultures and effects of infection with Mycobacterium leprae Clin Exp Immunol68 500ndash509
Sanes JR Lichtman JW 1999 Development of the vertebrate neuromuscularjunction Annu Rev Neurosci 22 389ndash442 httpdxdoiorg101146annurevneuro221389
Sleigh JN Grice SJ Burgess RW Talbot K Cader MZ 2014 Neuromuscularjunction maturation defects precede impaired lower motor neuron connectivityin Charcot-Marie-Tooth type 2D mice Hum Mol Genet 23 2639ndash2650 httpdxdoiorg101093hmgddt659
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008
12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

12 MM Tetruashvily et al Brain Behavior and Immunity xxx (2016) xxxndashxxx
Smith LK He Y Park JS Bieri G Snethlage CE Lin K Gontier G Wabl RPlambeck KE Udeochu J et al 2015 Beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis Nat Med 21932ndash937 httpdxdoiorg101038nm3898
Smits KM Kuppen PJ Eggermont AM Tamatani T Miyasaka M Fleuren GJ1994 Rat interleukin-2-activated natural killer (A-NK) cell-mediated lysis isdetermined by the presence of CD18 on A-NK cells and the absence of majorhistocompatibility complex class I on target cells Eur J Immunol 24 171ndash175
Stevens B Allen NJ Vazquez LE Howell GR Christopherson KS Nouri NMicheva KD Mehalow AK Huberman AD Stafford B et al 2007 Theclassical complement cascade mediates CNS synapse elimination Cell 1311164ndash1178
Strait RT Hicks W Barasa N Mahler A Khodoun M Kohl J Stringer K WitteD Van Rooijen N Susskind BM et al 2011 MHC class I-specific antibodybinding to nonhematopoietic cells drives complement activation to inducetransfusion-related acute lung injury in mice J Exp Med 208 2525ndash2544httpdxdoiorg101084jem20110159
Syken J Grandpre T Kanold PO Shatz CJ 2006 PirB restricts ocular-dominanceplasticity in visual cortex Science 313 1795ndash1800
Tang G Gudsnuk K Kuo SH Cotrina ML Rosoklija G Sosunov A Sonders MS Kanter E Castagna C Yamamoto A et al 2014 Loss of mTOR-dependentmacroautophagy causes autistic-like synaptic pruning deficits Neuron 831131ndash1143 httpdxdoiorg101016jneuron201407040
Tapia JC Wylie JD Kasthuri N Hayworth KJ Schalek R Berger DRGuatimosim C Seung HS Lichtman JW 2012 Pervasive synaptic branchremoval in the mammalian neuromuscular system at birth Neuron 74 816ndash829 S0896-6273(12)00385-6 [pii] 101016jneuron201204017
Tetruashvily MA Melson JW Park JJ Peng X Boulanger LM Expression andalternative splicing of classical and nonclassical MHCI genes in thehippocampus and neuromuscular junction Mol Cell Neurosci in press
Thams S Brodin P Plantman S Saxelin R Karre K Cullheim S 2009 Classicalmajor histocompatibility complex class I molecules in motoneurons newactors at the neuromuscular junction J Neurosci 29 13503ndash13515 294313503 [pii] 101523JNEUROSCI0981-092009
Thompson W 1983 Synapse elimination in neonatal rat muscle is sensitive topattern of muscle use Nature 302 614ndash616
Please cite this article in press as Tetruashvily MM et al MHCI promotes devtebrate neuromuscular junction Brain Behav Immun (2016) httpdxdoiorg
Thompson WJ 1985 Activity and synapse elimination at the neuromuscularjunction Cell Mol Neurobiol 5 167ndash182
Thompson W Kuffler DP Jansen JK 1979 The effect of prolonged reversibleblock of nerve impulses on the elimination of polyneuronal innervation of new-born rat skeletal muscle fibers Neuroscience 4 271ndash281
Tsai NP Wilkerson JR Guo W Maksimova MA DeMartino GN Cowan CWHuber KM 2012 Multiple autism-linked genes mediate synapse eliminationvia proteasomal degradation of a synaptic scaffold PSD-95 Cell 151 1581ndash1594 S0092-8674(12)01425-0 [pii] 101016jcell201211040
Valdez G Tapia JC Kang H Clemenson Jr GD Gage FH Lichtman JW SanesJR 2010 Attenuation of age-related changes in mouse neuromuscularsynapses by caloric restriction and exercise Proc Natl Acad Sci USA 10714863ndash14868 1002220107 [pii] 101073pnas1002220107
Valdez G Tapia JC Lichtman JW Fox MA Sanes JR 2012 Shared resistance toaging and ALS in neuromuscular junctions of specific muscles PLoS One 7e34640 httpdxdoiorg101371journalpone0034640 PONE-D-12-03398[pii]
Vugmeyster Y Glas R Perarnau B Lemonnier FA Eisen H Ploegh H 1998Major histocompatibility complex (MHC) class I KbDb deficient mice possessfunctional CD8+ T cells and natural killer cells Proc Natl Acad Sci USA 9512492ndash12497
Wareham AC Morton RH Meakin GH 1994 Low quantal content of theendplate potential reduces safety factor for neuromuscular transmission in thediaphragm of the newborn rat Br J Anaesth 72 205ndash209
Wood SJ Slater CR 2001 Safety factor at the neuromuscular junction ProgNeurobiol 64 393ndash429 S0301008200000551 [pii]
Xu K Jha S Hoch W Dryer SE 2006 Delayed synapsing muscles are moreseverely affected in an experimental model of MuSK-induced myastheniagravis Neuroscience 143 655ndash659 httpdxdoiorg101016jneuroscience200609022
Yamamoto Y Henderson CE 1999 Patterns of programmed cell death inpopulations of developing spinal motoneurons in chicken mouse and ratDev Biol 214 60ndash71 httpdxdoiorg101006dbio19999413
Zijlstra M Li E Sajjadi F Subramani S Jaenisch R 1989 Germ-linetransmission of a disrupted beta 2-microglobulin gene produced byhomologous recombination in embryonic stem cells Nature 342 435ndash438
elopmental synapse elimination and aging-related synapse loss at the ver-101016jbbi201601008

















