merged_document
43
Zoology Moderatorship Thesis 2016 The Effects of the Acanthocephalan Polymorphus minutus on the Cannibalistic Behaviour of Gammarus duebeni Laura Matthews
-
Upload
laura-matthews -
Category
Documents
-
view
13 -
download
0
Transcript of merged_document

Zoology Moderatorship Thesis
2016
The Effects of the Acanthocephalan Polymorphus minutus on the Cannibalistic
Behaviour of Gammarus duebeni
Laura Matthews

Declaration I, Laura Matthews declare that this thesis is my own work except where stated through
references or in the Acknowledgements and that it is 7010 words in length.
Signed:
Date:

Abstract Parasites may often mediate behavioural changes in their hosts to better serve
themselves, often at a cost to the hosts’ fitness or survivorship. Cannibalism of juvenile
or smaller conspecifics is one such behaviour that has previously been shown to be
altered by parasitic infection in many parasites and hosts. Here, the effects of the
acanthocephalan parasite Polymorphus minutus on the cannibalistic activity of its
intermediate host, Gammarus duebeni, are investigated. Individual infected and
uninfected adults were isolated and offered juvenile same-species prey, and average
amount of cannibalistic activity and cannibalistic individuals per group were assessed.
Infection was shown to alter the cannibalistic behaviour by creating a more uniform
behaviour in their hosts, whereas uninfected individuals displayed a wide range of
behavioural habits.

Acknowledgements I would first like to thank Professor Celia Holland, thank you so very much for your
advice, your guidance and for being nothing less than a fantastic supervisor. You
always seemed happy to see or hear from me, and always had a solution to any issue I
had. From hyphenating words to kick-sampling, you were greatly appreciated and
instrumental in my finishing of this project in one piece. You are one of the kindest and
most pragmatic people I have ever met, and working with you was a delight.
Maureen Williams, thank you so much for listening to every silly problem I ever had, for
being patient, and for turning up at 7 in the morning so frequently. You made the
experience so much simpler and more fun. Best of luck with your PhD, I will miss
working with you!
Thank you to Lauren Redmond and Regan Drennan for coming with me to collect my
samples. Your kicking skills were greatly appreciated, along with your friendship this
year. Thank you to Regan again, and also Tom Murphy, Dan McDermott and anyone
else I’ve forgotten for always giving me a hand when R was being temperamental.
Thank you to Paula Tierney, for helping me sort, for taking my extra Gammarus off my
hands and for just generally being the person I could moan about my experiments to,
you’re the only one that really got it. Best of luck with your sweet Gamms.
Thank you to Sam, Jack, Siki, Chloe and Caoimhe, Celia’s other supervisees, for
always being optimistic, for keeping me interested in our group meetings, and for
sharing the experience this year. Best of luck to all of you with your own projects!
Special thanks to my mum, who, while not understanding a thing about what I was
doing, was always eager to ask me how my ‘Grammadus’ were doing.
Finally, and with sincerity, thank you to all of my Gammarus who gave their lives for this
study, particularly to Wilbert (Inf16, shown in plate 5). Even in death you were the best-
behaved little friend.

Table of Contents Declaration ..................................................................................................................i Abstract ......................................................................................................................ii Acknowledgements ..................................................................................................iii Table of Contents .....................................................................................................iv Index of Figures .........................................................................................................v Index of Tables ........................................................................................................vi Index of Plates .........................................................................................................vii 1. Introduction ............................................................................................................1 2. Materials and Methods ..........................................................................................7
2.1 Site Selection ............................................................................................7 2.2 Pilot Study .................................................................................................8
2.2.1 Collection of Hosts ........................................................................8
2.2.2 Processing of animals for experimentation ...................................8 2.2.3 Experimental design .....................................................................9 2.2.4 Data Analysis ..............................................................................10
2.3 Main Experiment .....................................................................................11 2.3.1 Collection of hosts .......................................................................11
2.3.2 Processing of animals for experimentation .................................11
2.3.3 Experimental design ...................................................................12
2.3.4 Dissection and parasitological procedures .................................13
2.3.5 Statistical Analysis ......................................................................14
3. Results ..................................................................................................................15 3.1 Pilot Study ...............................................................................................15 3.2 Main Experiment .....................................................................................16 3.3 Cystacanth identification .......................................................................21
4. Discussion .......................................................................................................... 23 5. References ...........................................................................................................27 Appendix ..................................................................................................................33

Index of Figures: Figure 1: Life cycle of Polymorphus spp, adapted from Mehlhorn and Aspöck, 2007......3
Figure 2: Map of Lough Lene. Sampling site is represented by the red star....................7
Figure 3: Boxplot of the relationship between infection status and log 10 transformed
number of juveniles consumed over a period of 24 hours..............................................16
Figure 4: Boxplot of the relationship between infection status and the Log 10
transformed number of juveniles consumed over a 48-hour period...............................18
Figure 5: Plot of the cumulative mean juveniles dead in each group at each time
interval............................................................................................................................19
Figure 6: Mean juveniles eaten at each 12-hour interval................................................19
Figure 7: Scatter plot of weight of adults (g) against number of juveniles consumed in
a 48-hour period.............................................................................................................20
Figure 8: Scatter plot of lengths of adults (mm) against number of juveniles consumed
in the 48-hour experimental period.................................................................................21

Index of Tables: Table 1: Summary statistics of juvenile mortality in the pilot study...................................5
Table 2: Comparison of length and weights between the two groups of Gammarus.......8
Table 3: Summary statistics of juvenile mortality in the main experiment........................8

Index of Plates:
Plate 1: Gammarus duebeni infected with a cystacanth of Polymorphus minutus, as
indicated by the orange dot in the body cavity. ............................................................... 5
Plate 2: Collection site, Lough Lene, County Westmeath, on day of collection for the
pilot study. ....................................................................................................................... 8
Plate 3: Sampling Site for Main Experiment, as for pilot study. ..................................... 11
Plate 4: View of a jar from above at the 12 hour interval. One juvenile had been eaten
by an uninfected adult. Adult is shown by the arrow. ................................................... 12
Plate 5: Infected adult prepared for dissection. Cystacanth can be seen as an orange
dot at the lower portion of the body. .............................................................................. 14
Plate 6: Activated Polymorphus minutus viewed under the microscope. Hooks can be
seen on the proboscis, as indicated by the arrow. ........................................................ 22
Plate 7: Closer viewing of proboscis. Characteristic hook pattern can be seen.............22

viii

9

1
1. Introduction: Cannibalism, or intraspecies predation (defined as the catching, killing and devouring of
an animal by a conspecific), is a widespread behaviour in the animal kingdom with at
least 3000 species, across 900 families subscribing at least partially to this method of
feeding (Fox, 1975; Polis, 1981). Cannibalism can be very advantageous. It is an
effective method of feeding, with the prey’s nutrient content matching closely what is
required of the predator, thus removing the danger of a nutritional mismatch (Bobisud,
1976). For example, arthropods tend to be limited in somatic growth by abundances of
specific rare minerals, and so, predating conspecifics eliminates the need to search for
such substances (Denno and Fagan, 2003). Cannibalism also has an indirect fitness
advantage of removing competitors, thus increasing potential resources. For example,
female three-spined sticklebacks cannibalise the eggs of conspecifics even when
alternative food is superabundant (Fitzgerald, 1992).
Cannibalism increases during periods of food scarcity for a number of reasons; the lack
of food leaves organisms weak and vulnerable to cannibalism, thus creating a ‘kill or be
killed’ scenario (Elgar and Crespi, 1992). Laws of optimal foraging theory show that
under conditions of food scarcity, individuals expand their diet into previously
unacceptable foodstuffs of high cost or low energy gain (Fox, 1975). Such predation is
common in colonial species, where the availability of same-species prey is high, or in
areas where other species to predate on are in low densities (Claessen et al., 2004).
Cannibalism is also more likely to occur in populations that have overlapping
generations in time and space, and also have notable differences in size (Wissinger
1992). It is a well-accepted fact that smaller (younger) individuals in a population are
usually cannibalized by larger (older) individuals, known as size asymmetric
cannibalism. Therefore, newborns are particularly vulnerable to attack, with such
predation by conspecifics found in over 80 species (Hardy, 1977). As such, cannibalism
can be a major source of juvenile mortality, and the danger of cannibalism decreases
with age (Polis 1981).
While cannibalism can present many advantages, and for many species may just be
considered an extension of normal predation behaviours (Hardy, 1977), it can also often
be disadvantageous. Filial cannibalism can create a potential net decrease in inclusive
fitness if the organism is likely to eat close relatives, although it may also increase the
survivorship and fitness of the remaining offspring and adult (Manica, 2002). Cannibals

2
and their conspecific prey, or their preys guardians (Sherman, 1981), may be evenly
matched in fighting ability, and the likelihood of injury to the cannibal may be high
(Dawkins, 1976). Cannibalism and intraspecific necrophagy (the eating of dead
conspecifics, a behaviour closely linked with cannibalism (Rudolph, 2007)) can also
increase the likelihood of disease or parasite transmission (Pfenning et al., 1998;
Rudolph & Antonovics, 2007). For example, cannibalistic tiger salamanders
(Ambystoma tigrinium nebulosum) that ate diseased conspecifics were found less likely
to survive to metamorphosis (Pfenning et al., 1998), and that the frequency of
cannibalistic tiger salamanders is negatively correlated with the bacterial density in their
habitat, to avoid the predation of infected conspecifics (Pfenning, 1991).
Parasites are important to almost all ecosystems and individuals within them and
greatly influence ecosystem properties (Bush et al., 1997; Hatcher et al., 2012; Hatcher
et al., 2014). Parasites may also have profound effect on populations of species through
manipulation of their host’s phenotype (Dobson, 1988; Hatcher and Dunn, 2011).
Parasites may have complex life-cycles, requiring transmission through many hosts to
complete their life cycle (Parker et al., 2003; Auld and Tinsley, 2014). Manipulative
parasites can affect all aspects of their hosts’ phenotype; their behaviour, reproduction,
morphology and physiology, to further serve themselves in their survival and
transmission to the next host in their life-cycle, or to propagate throughout a particular
population of one species (Poulin, 1995; Poulin, 2010). Increased transmission of
creates an increase in fitness for the parasite (Lafferty, 1999). Specifically, the larvae of
many parasites can alter the behaviour or appearance of intermediate hosts in ways
that increase the likelihood of transmission to the next host, by increased susceptibility
to predation (Brown and Thompson, 1986). The parasite-induced change in behaviour
can increase transmission and the frequency with which the definitive host may be
infected (Lafferty, 1999).
Acanthocephalans are one such phylum of obligate intestinal parasites. Often known as
‘spiny-headed’ worms, they are recognized by their proboscis, which they use to hook
into the intestines of their hosts (Petroschenko, 1958). They have a highly conserved,
yet complex life-cycle, culminating in a definitive vertebrate host, with an intermediate
arthropod host (Near, 2002). Cystacanths (the fully developed larva of an
acanthocephalan) have the ability to manipulate the phenotypic behaviour of a wide
range of arthropod intermediate hosts (Bethel and Holmes, 1977; Cézilly et al., 2011),
and acanthocephalans use this to take advantage of trophic interactions between their

3
intermediate and definitive hosts (Poulin, 1994). Polymorphus minutus is a common
acanthocephalan parasite culminating in a variety of aquatic bird species that act as
definitive hosts, with various members of the Gammarus family used as intermediate
hosts (Nicholas and Hynes, 1958).
Figure 1: Life cycle of Polymorphus spp, adapted from Mehlhorn and Aspöck, 2007.
Gammarus are crustaceans belonging to the order Amphipoda, a widely distributed and
speciose genus (Costa et al., 2009). The majority dwells in marine or freshwater
settings, although some species are semi-terrestrial, residing in sand or moist leaf litter
(Sexton, 1928). Gammarus are usually found in substrates that can provide both shelter
and a food source, such as in gravel or under dead vegetation (Fitter and Manuel, 1997;
McGrath et al., 2007). These freshwater amphipods are an ecologically significant
species, due to their wide distribution, the significant amount of biomass they constitute
in their habitats, and due to their role as shredders of organic material. Often, they are
the dominant macroinvertebrate in their habitat, and densities can reach up to
thousands of individuals per metre squared (Kelly and Dick 2005). Gammarus spp. are
considered detritivors, that usually feed on a thin layer of biotic material of
allochthonous leaf litter, comprised primarily of microorganisms (Nelson, 2011), and the
majority of the literature state that they form part of a herbivorous functional feeding

4
group (Hatcher and Dunn, 2011). However, some studies show them to be carrion
feeders (MacNeil, Dick and Elwood 2007), and Gammarus are also often found to prey
upon conspecifics or other closely related species.
Cannibalistic species often share similar characteristics in terms of their ecology and
life-history; they live in large groups and are poor dispersers, they are continuous
breeders with overlapping generations, and live in areas with poor abundances of
alternate prey. As a result, such species often manifest cannibalism of conspecifics.
Cannibalism among Gammarus has been reported in many laboratory studies, the first
mention being by Sexton (1928). For example, Ward (1986) noted that in G. pulex,
males often eat females after copulation and once the females exoskeleton has been
shed. Size asymmetric cannibalism is particularly common in all species of amphipods,
with juveniles often acting as prey for adults (MacNeil, Dick and Elwood, 2007). Sexton
(1928) notes that individuals do not usually prey on conspecifics unless they are at
some disadvantage, namely a smaller size or are weakened from a recent moult. Many
things, such as food availability or habitat, can mediate cannibalism in Gammarus. For
example, poor quality habitats can mediate the incidence of cannibalism, with adult
Gammarus having to turn to cannibalism due to lack of food and juveniles having to
forage more widely to find food, leaving them open to predation (MacGrath et al, 2007).
While cannibalism can be disadvantageous, as mentioned above, Gammarus have
evolved methods of avoiding these negative effects. A temporal behavioural change
occurs in relation to brood stage in female G. pulex, whereby the instances of
cannibalism are significantly reduced concurrent to the time their own eggs are
hatching, to prevent the likelihood of eating their own young (Lewis et al., 2010). It is
unknown if other phenotypic recognition cues are utilized in this instance, or whether
males subscribe to a similar temporal avoidance of cannibalism. As mentioned above,
arthropods commonly subscribe to asymmetric cannibalism, avoiding potential injury in
cannibalistic attacks.
Juvenile Gammarus have developed behaviours to avoid being predated upon by
conspecifics. Like in many cannibalistic species, juveniles tend not to adhere to natal
philopatry; remaining in or returning to natal territory (Pearce, 2007), and quickly
expand outwards from their native zone, to create a separation between the vulnerable
juveniles and cannibalistic adults. Gammarus tigrinis, Gammarus mucronatus and
Gammarus lawrencianus exhibit changes in phototaxic behaviour at an age that

5
correlates to a reduction in vulnerability to predation. This causes a partial separation of
adults and juveniles that reduces the likelihood of predation (Hunte and Myers, 1984).
Smaller individuals shift habitats to minimize the risk of predation by conspecifics. With
larger conspecifics in absentia, juveniles of G. pulex will use larger pores in substrates
to hide in whereas when there is predation danger, they only select smaller pores,
regardless of food availability (MacGrath et al, 2007).
Larval P. minutus lives in the body cavity of its intermediate host, Gammarus, and the
cystacanth is clearly visible through the cuticle as an orange dot. Besides this obvious
morphological change in colouration, which aids transmission of the parasite to its next
host (Bakker et al, 1997), it has other significant implications for Gammarus. Like many
parasites, acanthocephalans can negatively affect the survivorship and reproductive
abilities of their hosts; for example, Pomphorynchus laevis increases sensitivity to
heavy metal contamination in its intermediate Gammeridean host (Chen et al, 2015).
Severe infection of P. minutus in G. lacustris can cause castration of the females and
can slow down the secondary sexual characteristics development in males, leaving
them less fit than their conspecifics (Ward, 1986).
Plate 1: Gammarus duebeni infected with a cystacanth of Polymorphus minutus, as
indicated by the orange dot in the body cavity.
G. pulex infected with P. laevis showed a lower motivation to feed, and increased
refuge use than uninfected conspecifics, thus impairing optimal foraging and food intake

6
(Dianne et al, 2014). Individuals are found to become highly photophilic when infected
with P.laevis (Bauer et al, 2000). Changes in behaviour tend to only occur when it is of
benefit to the parasite, i.e. when it is suitable for transmission to the next host. P.
paradoxus reverses the photonegative tendencies of infected G. lacustris, but only at
the cystacanth stage of development which is infective to the definitive host (Bethel and
Holmes, 1974).
The clear deleterious effects that parasites have on these hosts indicate that avoidance
of these parasites would be a beneficial behavioural trait of uninfected individuals. As
such, avoidance has been indicated in some studies on parasitized amphipods. In G.
lacustris, those infected with a cystacanth of P. minutus are less likely to be found in a
precopulatory mate-guarding formation. It is suggested that this is an active avoidance
of infected individuals by the uninfected individuals in the population (Ward, 1986).
As parasitism and cannibalism both play significant roles in the ecology and population
dynamics of many animals (as shown above), it is likely that infection by parasites will
influence cannibalism. Parasitism can affect the rate of foraging behaviour in many
animals, for example an increase in food intake has been recorded in G.pulex infected
with P. minutus due to a suggested increase in metabolic requirements mediated by the
parasite (Crompton 1970). It is therefore likely that it can have an effect on the rate or
propensity of cannibalism, as has been indicated in recent studies. The
acanthocephalan parasite Echinorynchus truttae mediates a reduction in intraguild
predation (the killing and consumption of closely related species, a behaviour closely
linked with cannibalism) between G. duebeni and G. pulex, and thus promotes species
coexistence (MacNeil et al., 2003). Conversely, it has been recently shown by Bunke et
al. (2015), that cannibalistic activity is increased among Gammarus duebeni celticus
infected with the microsporidian Pleistophora mulleri. However, different parasites
manipulate their hosts in a variety of ways and it is not necessarily true that this
behavioural change will be the case for all parasitized Gammarus.
As such, this project seeks to investigate the relationship between cannibalism and
infection by a macroparasite, the acanthocephalan P. minutus in Gammarus duebeni,
both host and parasite being widespread in aquatic systems. The aim of this project is
to investigate whether the incidences of cannibalism will be altered in the parasitised
individuals compared with that of the uninfected individuals.

7
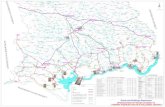
2. Materials and Methods 2.1 Site Selection The selected site for collecting the samples for this study was Lough Lene, a lake in the
upper Boyne catchment in County Westmeath. This site was selected due to a known
presence of Gammarus duebeni (Sutcliffe, 2010) and a comparatively high prevalence
of the acanthocephalan parasite Polymorphus minutus, found to be 6% from a
collection of Gammarus on February 12th 2015 (personal communication, Maureen
Williams). It is a limestone lake, of which the surface area is 4.16km2 (Shilland et al.,
2009). Mean depth is 4m and the maximum depth is 20m, which is considered a deep
lake (Inland Fisheries Ireland). It has been classed as a mesotrophic lake
(Environmental Protection Agency, 2008). The lake has been designated a special area
of conservation (SAC), and as such, conditions are carefully monitored. However, no
license was needed to remove a sample of Gammarus from the lake.
Figure 2: Map of Lough Lene. Sampling site is represented by the red star.

8
2.2 Pilot Study 2.2.1 Collection of hosts
Kick sampling for the pilot study began at approximately 8.15 a.m. at Lough Lene, on
the 3rd of November 2015. Sampling occurred in the shallow lake water close to a boat
jetty (Co-ordinates 53.660532, -7.195431), as shown in figure 2, above and plate 2,
below. Kick sampling was performed by three people using 20cm x 20cm sweep net for
twenty minutes providing one hour of sampling effort. Contents of the net were placed in
one of three 10L collection buckets filled with lake water. A 20L container of lake water
was also collected, for use later. Fallen leaves were collected as a food source. The
specimens were brought back to Trinity College, and left in the 14 degrees Celsius cold
room to acclimatize for three hours, with airlines inserted into the buckets to prevent
anoxia. The collected water was also stored in the cold room.
Plate 2: Collection site, Lough Lene, County Westmeath, on day of collection for the
pilot study.
2.2.2 Processing of animals for experimentation
The buckets were processed one at a time in the aquatic laboratory; this involved
sorting into parasitised adults, non-parasitised adults and non-parasitised juveniles.
Parasitised juveniles were not required, as cannibalistic adults have a propensity to
avoid parasitised juveniles, to avoid infection (MacNeil et al., 2003), and the use of

9
parasitised individuals could be a confounding variable. Sorting was achieved by
pouring small amounts of the Gammarus and water into a white tray, picking the
selected individual up with a disposable pipette with the tip cut off and placing in the
correct labeled bucket. A second pipette was also adapted, by partially cutting off the
bulb at the back, to create a scoop that could be used to pick up the particularly large
Gammarus individuals, or Gammarus pairs in precopula formation.
Each group of animals was sorted into three separate containers full of lake water.
Juveniles were defined as being under 6mm in length, as per Bunke et al. (2015), and
suitably large adults were defined as being over 7mm in length. Adults and juveniles
were selected by eye. Parasitized individuals were recognized by their distinctive
orange dot in their body cavity and each potentially infected individual was viewed
under a dissecting microscope to confirm that they were parasitized, as some non-
parasitized Gammarus can have slightly orange colouration. Only animals with a single
dot were selected, to eliminate duel infection as a possible confounding variable. Any
parasitized juveniles found were added to the ‘discard’ bucket, as they were not
required for this experiment. Any specimens of intermediate size, between the sizes for
adult or juvenile, as judged by eye, were also put in the discard bucket, as only the
largest adult individuals were required. After looking at the parasitized adult samples, it
was decided that eight of the adults were a suitably large size and could be utilized in
the pilot study. Eight non-parasitized adults of approximately similar sizes were also
selected for experimentation. Adult were not weighed or accurately measured for the
pilot study.
2.2.3 Experimental design
Twenty-four 750ml jars were washed in an industrial glass cleaner and filled with
approximately 600mls of lake water. Jars were labeled as follows: 1-8 control (to have
no adult Gammarus in them), 9-16 non-parasitised, and 17-24 parasitised adult. Each
jar was assigned the appropriate adult, and the controls were left empty of adults. The
jars were then brought on trays to the 14 degrees Celsius cold room. These jars were
covered over with a petri dish to prevent any spillage during transport from the lab, and
to prevent any Gammarus from escaping. The placement of the jars on the trays was
randomised, to prevent any bias that may occur by placement. Over the course of the
study, the subjects were kept in an appropriate light and dark cycle. The Gammarus

10
were then left to starve for a duration of 24 hours. The non-parasitized juvenile
Gammarus were returned to a large bucket with a food source for this period.
The following day, twenty-four petri dishes, each with a small amount of lake water,
were filled with 7 juveniles, using the same scoop and pipette, to be added to each of
the jars. A total of 168 juveniles (24 x 7) were used. As size discrimination was done by
eye, only the smallest of the juveniles were selected for introduction into the jars to
ensure they were in fact juveniles (below 6mm in length), and the remainders were
returned to the ‘discard’ bucket. These petri dishes were transported downstairs to the
14 degrees Celsius cold room and one petri dish of juveniles were added to each of the
jars at 4 pm, carefully ensuring that no jar had accidentally received more than one petri
dish of juveniles. The petri dish lid of each jar was quickly removed and the contents of
the petri dish were poured in. The lid was then immediately replaced.
Twenty-four hours later the jars were examined. The number of juveniles still present in
each jar was counted, by holding the jar up in front of a white piece of paper to make it
easier to see. Missing or dead juveniles were noted, and from this, the number of
cannibalistic activities was calculated.
2.2.4 Data Analysis
Data on the amount of cannibalistic activity was calculated by subtraction of the
remaining alive juveniles from the number of introduced juveniles (7). Data sets of
infection status and number of missing juveniles were created on Microsoft Excel.
Statistical analysis was done using the statistical programme R (version 3.2.3 app GUI
1.65 (6833) x86_64-apple-darwin10.8.0). A chi-squared test was used to assess the
difference in the amount of cannibalistic individuals in each infection group. Normality of
distribution of the amount of juveniles eaten was tested using a QQ plot, and the logged
amount of juveniles eaten in each group was analysed using a t-test of the log-
transformed data. Summary statistics were collated onto a table for further study if
necessary. A box plot was created showing visually the difference between the
cannibalistic activity of the infected and non-infected animals.

11
2.3 Main Experiment 2.3.1 Collection of hosts
The study began on the 24th of November 2015, in the same location as the pilot study.
Conditions were slightly more windy and rainy, meaning that the organisms were
already unsettled. Four people began kick sampling at approximately 9.30 and lasted
for one hour, totaling 4 hours of sampling effort to ensure there was a more sizeable
sample of parasitized adults. Four buckets were collected in total; along with two more
buckets of lake water, and another 30-litre container of lake water. The samples were
brought back to Trinity College and each bucket was poured through a splitting funnel
into two buckets of the same type. This was done to half the amount of Gammarus in
each bucket, to prevent anoxia and overcrowding. Each bucket was then topped up with
lake water, and some detritus, leaves and stones were added to each of the buckets. A
total of eight buckets were created in this way and left in the 14 degrees Celsius cold
room to acclimatize overnight. Airlines were places in the buckets to prevent anoxia.
Plate 3: Sampling Site for Main Experiment, as for pilot study.
2.3.2 Processing of animals for experimentation
The following day sorting of the Gammarus began. As with the pilot study, Gammarus
were sorted as follows - uninfected adults, uninfected juveniles and parasitised adults.
The pilot study indicated that there was a possible link between size of the adult and its
propensity to cannibalise juveniles, and so, it was decided to increase the size of the
suitable adults to 8mm. Once the sorting was complete, the suitably sized (now 8mm+)
parasitized adults were selected. Ten were deemed suitable. As the pilot study

12
indicated that weight might have a role in the decision to cannibalise, the individuals
were weighed, to investigate this possible correlation. These were individually weighed
using a fine scale and wet weight (mass in g) was determined to four decimal places.
These individuals were returned to a jar. Using the range of these weights as a guide,
size-matched uninfected adults were selected.
A total of 4143 Gammarus were processed in this way. Of these, 135 were parasitised,
giving a prevalence of 3.3%. Adults constituted 3138 of the total amount, and juveniles
accounted for 1005. 20 adults (10 infected and 10 uninfected) were utilized in this
experiment, along with 210 uninfected juveniles.
2.3.3 Experimental design
Each adult was placed in a jar with 300 mls of lake water, providing a total of 20 jars. 10
control jars with no adult Gammarus were also used. The volume of water was
decreased from 600 to 300 mls to increase the density of the juveniles, as an increase
in density results in an increase in encounter and frequency of aggressive responses
such as cannibalism (Polis, 1981). No extra food was put in the jars, to increase the
necessity to cannibalise, and no extra detritus was added, as juveniles use these to
hide from cannibalistic adults (McGrath et al., 2007). Each jar was labeled and placed in
the 14 degrees Celsius cold room, with their positions on the trays randomised. Adult
Gammarus were starved for a period of 48 hours. During the experiment, one infected
adult died and was removed.
After 48 hours of starvation, 7 juveniles were added to each jar, as in the pilot study.
The jars were examined every 12 hours for a period of 48 hours to record any
cannibalistic activity. Remaining juveniles were counted and any moults or remnants of
juveniles were removed at 12, 24, 36 and 48 hours post-starvation, to prevent any
alternative food sources. The temperature in each jar was also noted at each time
interval. The trial ran for 48 hours. Upon completion, photographs were taking of the
remaining juveniles so they could be measured if needed.

13
2.3.4 Dissection and parasitological procedures
Once the trial was complete, adult Gammarus were dissected to confirm their infection
status (parasitized or non-parasitised). Individual adults were stretched out to full length
and body length (mm) measured with an electronic vernier calipers to two decimal
places. Cysts were removed from the infected individuals by dissection.
For P. minutus the activating agent is bile salts (Lackie, 1974), so, as described by Graff
and Kitzman (1965), the activation protocol for acanthocephalan cystacanths was
carried out; the cystacanths were added to an individual labeled test tube with a
0.25mM sodium taurocholate solution (created by dissolving 0.0625g per 25 ml of
deionized water) to stimulate activation and were left in a 37 degrees Celsius incubator
for 72 hours. Both adult and juvenile Gammarus were preserved in 70% alcohol with
10% glycerol, in case further data collection was needed.
Once the cystacanths had activated, they were fixed in a 70% alcohol and 10% glycerol
solution. The parasites were mounted on microscope slides using polyvinyl lactophenol
and were left overnight in a fume cupboard to dry. Each slide was labeled and observed
under a microscope. Each parasite was identified to species level on the basis of their
proboscis morphology and numbers of rows of hooks (McDonald, 1988; Petroshenko
1958).
Plate 4: View of a jar from above at the 12 hour interval. One juvenile had been eaten
by an uninfected adult. Adult is shown by the arrow.

14
2.3.5 Statistical Analysis
Data sets were complied on Microsoft Word and were analyzed on R (version 3.2.3 app
GUI 1.65 (6833) x86_64-apple-darwin10.8.0). Difference in cannibalistic individuals in
each infection status was investigated using a chi-squared test. QQ normality plots
were created to determine the normality of the distribution of the lengths, of weights and
of juvenile deaths in each group. As size of the adult was suggested to be a factor in an
individual’s propensity to cannibalise, t-tests were preformed to determine whether the
weights or lengths of the infected and uninfected Gammarus were significantly different.
T-tests were conducted to investigate the difference in the overall number of
cannibalistic events in each infection group. Graphs were produced of the cumulative
mean deaths per jar of each infection status, and of the mean deaths per time period
(each 12 hour interval). Scatter plots were created to compare the infection status and
weight of the adults with the amount of juveniles eaten, and the length and infection
status with the amount of juveniles eaten.
Plate 5 Infected adult prepared for dissection. Cystacanth can be seen as an orange dot
at the lower portion of the body.

15
3. Results: 3.1 Pilot Study
The pilot study was conducted to ensure that there was some relationship between
infection status and cannibalism, and establish baselines of cannibalistic behaviour for
the main experiment. Only one juvenile in the control jars died, which was not attributed
to cannibalism, as the corpse was recovered from the jar at the end of the experiment.
This represents one random death; therefore it was likely that any death in the infected
or uninfected jars were from cannibalism, and not from any other cause. All the 16
adults (eight infected and eight uninfected) survived the experiment. Of the infected
individuals, 2 of the 8 adults showed cannibalistic tendencies (25%). In the uninfected
individuals, six of the 8 adults (75%) showed cannibalistic tendencies. A chi-squared
test showed that there was a significant difference in the number of cannibalistic
individuals between the two groups (X-squared = 4, df = 1, p-value = 0.0455) and that
cannibalistic individuals were more common among the uninfected than in the infected.
Summary statistics were determined (see Table 1, below).
Table 1: Summary statistics of juvenile mortality in the pilot study.
Infection Status Uninfected Infected Control
Deaths 15 3 1
Range of deaths 0-4 0-2 0-1
Mean deaths per jar 1.875 0.375 0.125
Standard Deviation 1.64 0.74 0.35
Deaths as Percentage 28.6% 5.3% 1.8%
Median 1.5 0 0
A QQ normality plot showed that the number of juveniles consumed was not distributed
normally. A t-test comparing the log transformed amount of juveniles consumed by the
infected and uninfected individuals was performed and the difference was found to be
statistically significantly different (t=-2.3977, df=12.086, p=0.03353), indicating that
uninfected adults consumed more juveniles on average than the infected did (see
Figure 4).

16
Figure 3: Boxplot of the relationship between infection status and log 10 transformed
number of juveniles consumed over a period of 24 hours.
3.2 Main Experiment Of the 20 adults used, 1 died mid-experiment (jar 12, infected). This animal was
removed from the analysis. QQ normality plots indicated that the distribution of weights
and lengths of the adult Gammarus were normally distributed. A t-test compared the
weight (g) of infected and uninfected individuals post-study and the difference was not
found to be significant (t = 1.395, df=17, p=0.181). Similarly, the mean lengths (mm) of
the infected and uninfected adults were compared and not found to differ significantly (t
=1.3897 df=17 p=0.1825)(see Table 2). Therefore, the size of the individuals was not
considered to be a potentially confounding variable of cannibalistic behaviour between
infected and uninfected adults.
Infected Uninfected
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Infection Status
Log
of J
uven
iles
Con
sum
ed

17
Temperature was measured, and found not to have changed between jars or between
sampling times (see Appendix).
Table 2: Comparison of length and weights between the two groups of Gammarus.
Infection Status Infected Uninfected
n 9 10
Range of Length (g) 0.069-0.153 0.072-0.16
Mean Weight (g) 0.1047 0.1204
S.D of Weight 0.032 0.03
Range of length (mm) 8.65-12.92 8.9-12.23
Mean Length (mm) 10.894 10.027
S.D of Length 1.35 1.37
Two juveniles in the control jars died. The deaths in the control jars were not attributed
to cannibalism, as their bodies were recovered from the jar, and represent two random
deaths. Therefore, the death in the infected and uninfected jars was attributed to
cannibalism. Of the ten uninfected adults, 6 showed a propensity to cannibalise (60%),
whereas in the infected specimens, 7 out of 9 (78%) showed cannibalism. Difference in
amount of cannibalistic individuals was assessed using a chi-squared test and was
found to not be significant (X-squared = 2.0387, df = 1, p-value = 0.1533) (see Table 3).
A Mann-Whitney test of the unlogged data showed that there was also no significant
difference between cannibalism events in the infected and uninfected groups (W=49, P-
value= 0.764). Summary statistics were determined and are shown below (Table 3)
Table 3: Summary statistics of juvenile mortality in the main experiment.
Infection Status Uninfected Infected Control
Deaths 16 10 2
Range of deaths per jar 0-4 0-3 0-1
Mean deaths per jar 1.6 1.11 0.2
Standard Deviation 1.65 0.78 0.43
Deaths as Percentage 11.2% 7.77% 1.4%
Median 1.5 1 0

18
A boxplot was created to compare the difference in cannibalistic activity between the
infected and non-infected adults (figure 5, below). The boxplot shows a clear difference
in the instances of cannibalistic activity.
Figure 4: Boxplot of the relationship between infection status and the Log 10
transformed number of juveniles consumed over a 48-hour period.
A cumulative plot of juveniles eaten over time was produced, and the average juveniles
eater per time interval was determined. Means were used, as the amount of adults used
in each infection status was not equal, and means would remove this unevenness.
Uninfected individuals were shown to eat more juveniles over the 48-hour period than
the infected individuals did, but did not eat more over every time interval. For two of the
four time-intervals, infected individuals ate more juveniles than the uninfected did.
infected uninfected
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Infection Status
Log
of J
uven
iles
Con
sum
ed

19
Figure 5: Plot of the cumulative mean juveniles dead in each group at each time
interval.
Figure 6: Mean juveniles eaten at each 12-hour interval.
0.0
0.5
1.0
1.5
Time (Hours)
Cum
ulat
ive
Mea
n ju
veni
les
dead
0 12 24 36 48
UninfectedInfectedControl
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Time (Hours)
Mea
n ju
veni
les
dead
0 12 24 36 48
UninfectedInfectedControl

20
Weight of adults was plotted against the amount of juveniles consumed (Figure 8). The
graph shows no correlation between adult weight and the propensity to cannibalise.
Figure 7: Scatter plot of weight of adults (g) against number of juveniles consumed in a
48-hour period.
Similarly, length of adults was plotted against the amount of juveniles eaten (Figure 9)
and no obvious correlation between adult length and their propensity to cannibalise was
detected.
0.05 0.10 0.15 0.20
01
23
45
Weight (g)of Adults
Juve
nile
s ea
ten
UninfectedInfected

21
Figure 8: Scatter plot of lengths of adults (mm) against number of juveniles consumed
in the 48-hour experimental period.
3.3 Cystacanth identification Of the adult Gammarus deemed to be infected at the beginning of the experiment, the
remaining nine were infected upon dissection at the end of the experiment. All had a
single cyst infection, eliminating duel infection as a possible confounding variable. All
the cysts were activated in the sodium taurochlorate solution after the 48-hour
incubation period. All the activated cystacanths looked similar upon inspection under
the microscope and were all identified by their characteristic proboscis and pattern of
hooks as the acanthocephalan parasite P. minutus.
8 9 10 11 12 13 14
01
23
45
Length (mm)of Adults
Juve
nile
s ea
ten
UninfectedInfected

22
Plate 7 Closer viewing of proboscis. Characteristic hook pattern can be seen
Plate 6 Activated Polymorphus minutus viewed under the microscope. Hooks can be
seen on the proboscis, as indicated by the arrow.
Plate 1Closer view of proboscis and hooks.

23
4. Discussion The pilot study indicated that there was a significant difference in the incidence of
cannibalistic activity between parasitised and unparasitised adults of G. duebeni.
However, it also indicated that there was a likely confounding variable present; that size
of the adult may play a role in the propensity to cannibalise. In some cannibalistic
species, it is the largest morphs that are found to have cannibalistic tendencies, as is
the case in Tiger Salamanders (Pfenning, 1991) and largemouth bass (Clady, 1974). Of
the uninfected individuals, the two that did not cannibalize juveniles were comparatively
small in relation to the other selected Gammarus, and the two infected individuals that
did cannibalise were notably larger than their other infected conspecifics. This prompted
the decision to weigh and measure the lengths of the adults accurately, and more
carefully size match the adults being used in the main experiment. Lengths and weights
were measured in the main experiment, but neither parameter over the range of the
adults selected was found to be a significant factor in their propensity to cannibalise.
However, the selected infected adults were larger in the main experiment than they
were in the pilot study, as there were more suitable adults to select from. The
percentage of those infected that showed cannibalistic activity rose from 25% in the
pilot study to 78% in the main experiment.
There was also a decrease in the amount of juveniles eaten by the uninfected adults,
which may also have been mediated by the change in size range. Many of the
uninfected adults used in the main experiment were smaller than those used in the pilot
study, as instead of using the largest ones, they were size matched to the range of the
infected adults. Cannibalistic individuals fell slightly, from 75% to 60% between the pilot
study and the main experiment. There is an indication therefore, that there is some link
with size in an individuals propensity to cannibalise. However the range of sizes
selected was very narrow in this experiment, and therefore it was not possible to
conclusively determine the relationship between size and cannibalistic behaviour.
There was uncertainty as to whether the absolute size of the individual adult, or the
relative size compared to its prey is the factor that mediates cannibalistic activity.
Studies of cannibalistic tendencies in a wide range of other species indicate that there is
a ‘cannibalism threshold’ i.e. that an individual should be a certain percentage larger
than its prey to cannibalize it (Polis, 1981), for example, conspecifics are only eaten by
piscivorous fish if the predator-to-prey ratio exceeds the threshold of 80-100% (Popova,
1967). It is well known that asymmetric predation is common in amphipods (MacNeil,

24
Dick and Elwood, 2007) and efforts were made to ensure that there was a notable size
difference between adults and their prey, by selecting from the smallest juveniles and
largest adults. However, in some instances, the ‘juveniles’ were relatively quite large,
and some of the parasitized adults were relatively small. This could have a significant
effect on the rate of cannibalism, if the cannibalism threshold had not been reached.
Therefore, in further studies, the size of the juveniles relative to the size of the adult
should be taken into account, as this could be considered a confounding variable.
The main experiment indicated that there was a marked difference in variance of
cannibalistic activity between infected and uninfected individuals. The majority of
infected adults ate one juvenile, whereas there was a wide range of numbers (0-4) of
juveniles consumed by the uninfected adults. This clearly indicates difference between
the behaviours of infected and uninfected adults, with a wide amount of variation in the
uninfected adults, and uniformity in the infected individuals. This has been suggested in
the literature on animal behaviour. In some species it has been noted that there are
‘cannibal specialists’, who are more predisposed to cannibalism than other adult
conspecifics indicating some sort of predisposition to cannibalism (Polis, 1981). This is
a kind of behavioural polyphenism or difference in ‘animal personality’, defined as the
differences in behaviour among individuals that are consistent over time and situations.
Behavioural traits may not always be beneficial when uniformly used by a whole
population, and so, populations of species may develop multiple phenotypic traits
(Reale et al., 2010). This may have an effect on propensity to cannibalise, and an effect
on the ability of the parasite to alter behaviour (Poulin, 2013). For example, in a study of
a colony of 900 herring gulls, intraspecific oophagy (the eating of eggs) and juvenile
cannibalism are common, and 23.3% of all eggs and chicks were eaten by conspecifics.
Of this, four individuals were responsible for 2-5% of this figure (Parsons, 1971). Such
specialism in the propensity to cannibalise is also found in bass (Clady, 1974), newts
(Kaplan, 1980), chimpanzees (Goodall, 1977), and many other species. There is a
similarly evident genetic component to cannibalism in some species. For example, in
eight separate strains of laboratory mice, rates of litter cannibalism remain the same for
at least 13 generations, implying a genetic link to cannibalism preferences (Hauschka,
1952). Therefore, it may have been likely that some of the selected uninfected adults
were genetically disposed to cannibalise or exhibited some sort of behavioural
polyphenism, as indicated by some uninfected individuals expansive feeding on
conspecifics, and some who were not predisposed to cannibalism. This is evident in the

25
wide variation in the amount of juveniles consumed by each uninfected adult; some ate
no juveniles, and some ate more than half. Similarly, some infected individuals may be
particularly adept at avoiding the parasites manipulative abilities, as exhibited in the
infected individuals that did not cannibalise, or cannibalised more than others did.
Poulin (2012) suggests that animals personality is a barrier to parasitic infection, where
some phenotypes of a polyphenotypic animals are more easy to manipulate, and some
presenting a greater difficulty.
Alternatively, the increase in cannibalism by the infected adults may have been
mediated by altered prey density. The reduction in water from 600mls per jar to 300mls
per jar increased the density from 1.14 juveniles per 100mls to 2.28 juveniles per
100mls. Increased rates of intraspecific predation have been linked to high densities or
overcrowding in the wild, as an increased density of prey will increase the probability of
encounter and the frequency of aggressive behaviours (Fox, 1975; Polis, 1981) such as
cannibalism. Therefore, the increase in density of juveniles may have affected the
instances of cannibalism in the infected adults.
Cannibalism activity may have been reduced in some of the adults, due to the temporal
shift in cannibalism activity found in recently brooding females (Lewis et al., 2010). This
would have reduced the incidence of cannibalism in the uninfected group. While it may
be beneficial to only utilize males in further studies to remove this potentially
confounding variable, it is difficult to tell the sexes apart without extensive investigation
under the microscope. Future experiments may possibly utilize males exclusively to
avoid this.
Upon inspection of the surviving juveniles, it was discovered that 2 of the remaining
juveniles were in fact infected (0.011%), as indicated by the characteristic bright orange
cyst on their sides. One was found in an ‘uninfected’ jar and one in an ‘infected’ jar.
While there is evidence of an avoidance of infected individuals by uninfected individuals
(Ward, 1986), and that more specifically, uninfected adults avoid predating infected
juveniles (MacNeil et al., 2003), this would not have affected the results as food sources
(i.e the juveniles) were already in excess, and cannibalism had still occurred in both the
of the jars these juveniles were found in. Infected adults do not discriminate between
infected and uninfected juveniles (Bunke et al., 2015), and so this would not have had
any effect in the infected jars.

26
Many parasitised animals, including humans, exhibit a reduction in food intake
compared to that of uninfected conspecifics (Crompton, 1984; Kyriazakis, 1998) and it
is possible that this macroparasite alters the individual in such a way that they only
consume the minimum necessary to survive. In many of the infected jars at each
interval, large portions of deceased juveniles were removed from the floor of the jar,
whereas in the uninfected jars, where the juvenile was cannibalised, the majority of the
body had been eaten, with the exception of some limbs.
The presentation of cannibalism in this experiment was different to that found in other
similar studies. For example, G. pulex infected with P. minutus were found to have an
increased rate of foraging for food (Crompton 1970). However, in this experiment, G.
duebeni infected with P. minutus presented with a reduced intake of same-species prey
(i.e. a reduction in rate of foraging). Bunke et. al. (2015) found that for G. duebeni
infected with P. mulleri, the incidence of cannibalism increased significantly. This is
contrary to the findings in the present study, where the variation in the infected
individuals was greatly reduced. However, P. mulleri microsporidian parasite, with a
single host-species life cycle. The propagation of P. mulleri occurs exclusively through
cannibalism (MacNeil et al., 2003), and it is likely that infection by this species may
have an effect on the rate of cannibalism. The results of this study has been mirrored by
the results found in Gammarus spp. infected with a fellow acanthocephalan parasite; E.
truttae. In this case, infection mediates a reduction in intraguild predation between G.
duebeni and G. pulex (MacNeil et al., 2003).
Overall, the alterations in cannibalistic behaviour by parasitic infection have presented
differently in this experiment than in that of comparable studies, though this is not to say
that this study has produced contrary or unhelpful information. Parasites are
exceedingly complex creatures and it is evidently not possible to have a rule of thumb
for any behavioural or morphological alteration, let alone for one so complex as
cannibalism. This project has gone some way into exploring the potentially vast and
varied effects parasites may have on different hosts foraging and cannibalistic
behaviour. In the future, further studies and larger analyses are required for this vast,
yet fascinating subject.

27
5. References: 1. Auld, S KJR, and M C Tinsley. "The Evolutionary Ecology Of Complex Lifecycle
Parasites: Linking Phenomena With Mechanisms". Heredity 114.2 (2014): 125-
132. Web.
2. Bakker, T. C. M., Mazzi, Z. & Zala, S. "Parasite-Induced Changes In Behavior
And Color Make Gammarus Pulex More Prone To Fish Predation". Ecology 78.4
(1997): 1098.
3. Bauer, A., Trouvé, S., Grégoire, A., Bollache, L. C. & Cézilly, F. Differential
influence of Pomphorhynchus laevis (Acanthocephala) on the behaviour of native
and invader gammarid species. International Journal for Parasitology, 30, (2000)
1453- 1457.
4. Bethel, W. M., and Holmes, J. C., "Correlation Of Development Of Altered
Evasive Behavior In Gammarus lacustris (Amphipoda) Harboring Cystacanths Of
Polymorphus paradoxus (Acanthocephala) With The Infectivity To The Definitive
Host". The Journal of Parasitology 60.2 (1974): 272-274. Print.
5. Bethel, W. M. and Holmes, J. C. (1977). Increased vulnerability of amphipods to
predation owing to altered behavior induced by larval
acanthocephalans. Canadian Journal of Zoology 55, 110–115.
6. Bobisud, L. Cannibalism as an evolutionary strategy. Bulletin of Mathematical
Biology (1976) 38:359-68
7. Brown, A. F., and Thompson D. B. A. "Parasite Manipulation Of Host Behaviour:
Acanthocephalans And Shrimps In The Laboratory". Journal of Biological
Education 20.2 (1986): 121-127.
8. Bunke M, Alexander ME, Dick JTA, Hatcher MJ, Paterson R and Dunn AM.
(2015) Eaten alive: cannibalism is enhanced by parasites. R. Soc. open sci. 2:
140369.
9. Bush, A. O. et al. "Parasitology Meets Ecology On Its Own Terms: Margolis Et Al.
Revisited". The Journal of Parasitology 83.4 (1997): 575.
10. Cezily, F. et al. "Host-Manipulation By Parasites With Complex Life Cycles:
Adaptive Or Not?". Trends in Parasitology 26.6 (2011): 311-317. Print.
11. Chen, Hui-Yu et al. "Effects Of The Acanthocephalan Polymorphus Minutus And
The Microsporidian Dictyocoela Duebenum On Energy Reserves And Stress
Response Of Cadmium Exposed Gammarus Fossarum". PeerJ 3 (2015): e1353.

28
12. Clady, M. Food habits of yellow perch, smallmouth bass and largemouth bass in
two unproductive lakes in northern Michigan. American Midlands Nature. (1974).
91:453-59
13. Claessen D, de Roos AM, Persson L. Population dynamic theory of size-
dependent cannibalism. Proc. R. Soc. London. (2004) B 27, 333–340.
(doi:10.1098/rspb.2003.2555)
14. Costa, F. O. et al. "Probing Marine Gammarus (Amphipoda) Taxonomy With
DNA Barcodes". Systematics and Biodiversity 7.4 (2009): 365-379.
15. Crompton, D.W.T. An ecological approach to acanthocephalan physiology.
(1970)
16. Crompton, D.W. "Influence Of Parasitic Infection On Food Intake.". Federation
Proceedings 43 (1984): 239-245. Print.
17. Dawkins, R. The Selfish Gene. Oxford: Oxford University Press, 1989. Print.
18. Denno, R. F., and Fagan W. F. 'Might Nitrogen Limitation Promote Omnivory
Among Carnivorous Arthropods?' Ecology 84.10 (2003): 2522-2531.
19. Dezfuli, Bahram S, Giari, L, and Poulin, R. "Species Associations Among Larval
Helminths In An Amphipod Intermediate Host". International Journal for
Parasitology 30.11 (2000): 1143-1146.
20. Dianne, L. et al. "Parasite-Induced Alteration Of Plastic Response To Predation
Threat: Increased Refuge Use But Lower Food Intake In Gammarus Pulex
Infected With The Acanothocephalan Pomphorhynchus Laevis". International
Journal for Parasitology 44.3-4 (2014): 211-216.
21. Dobson, A. P. "The Population Biology Of Parasite-Induced Changes In Host
Behavior". The Quarterly Review of Biology 63.2 (1988): 139-165. Web.
22. Elgar MA, Crespi BJ. 1992 Cannibalism: ecology and evolution among diverse
taxa. Oxford, UK: Oxford University Press.
23. Environmental Protection Agency. Water Quality In Ireland 2004 - 2006.
Wexford: EPA, (2008). Print.
24. Fitter, R., Richmond, S., and Manuel, R. Lakes, Rivers, Streams & Ponds Of
Britain & North-West Europe. : HarperCollins, 1994. Print.
25. FitzGerald, G. J. Egg Cannibalism By Sticklebacks: Spite Or Selfishness?.
Behavioral Ecology and Sociobiology 30.3-4 (1992): 201-206. Web.
26. Fox L. R. Cannibalism in natural Populations. Annual Review of Ecological
Systems. 1975 6:87-106

29
27. Goodall, J. Infant killing and cannibalism in free-living chimpanzees. Folia
Primatol. 1977 28:259-82
28. Graff, D. J., and Kitzman W. B. "Factors Influencing The Activation Of
Acanthocephalan Cystacanths". The Journal of Parasitology 51.3 (1965): 424.
Web.
29. Hardy, D. B. Infanticide as a primary reproductive strategy. American
Scientist 1977 65 (1): 40–49
30. Hatcher MJ, Dunn AM. 2011 Parasites in ecological communities: from
interactions to ecosystems. Cambridge, UK: Cambridge University Press.
31. Hatcher MJ, Dick JTA, Dunn AM. 2014 Parasites that change predator or prey
behaviour can have keystone effects on community composition. Biology Letters.
10, 20130879. (doi:10.1098/rsbl.2013.0879)
32. Hatcher, M. J., Dick J. T. A., and Dunn, A. M. "Diverse Effects Of Parasites In
Ecosystems: Linking Interdependent Processes". Frontiers in Ecology and the
Environment 10.4 (2012): 186-194.
33. Hauschka, T. Mutilation patterns and hereditary cannibalism in mice. Journal of
Heredity. (1952), 43:117-23
34. Hunte, W., and R.A. Myers. "Phototaxis And Cannibalism In Gammaridean
Amphipods". Marine Biology 81.1 (1984): 75-79. Print
35. Inland Fisheries Ireland, Lough Lene. Inland Fisheries Ireland, 2010. Print.
Sampling Fish For The Water Framework Directive.
36. Kelly, D. W., and DICK J. T. A., "Introduction Of The Non-Indigenous Amphipod
Gammarus Pulex Alters Population Dynamics And Diet Of Juvenile Trout Salmo
Trutta". Freshwater Biology 50.1 (2005): 127-140.
37. Kyriazakis, I., Tolkamp, B.J., and Hutchings, M.R. "Towards A Functional
Explanation For The Occurrence Of Anorexia During Parasitic Infections". Animal
Behaviour 56.2 (1998): 265-274. Web.
38. Lackie, A. M. The activation of cystacanths of Polymorphus minutus
(acanthocaphala) in vitro. Parasitology, 68: 135-46
39. Lafferty, K.D. "The Evolution Of Trophic Transmission". Parasitology Today 15.3
(1999): 111-115. Web.
40. Lewis, S. E. et al. "Avoidance Of Filial Cannibalism In The Amphipod Gammarus
Pulex". Ethology 116.2 (2010): 138-146.
41. Manica, A. "Filial Cannibalism In Teleost Fish". Biology Review 77.2 (2002): 261-
277.

30
42. McDonald, M. E. Key to Acanthocephala reported in waterfowl. (1988) DTIC
Document.
43. McGrath, K. E. et al. "Habitat-Mediated Cannibalism And Microhabitat Restriction
In The Stream Invertebrate Gammarus Pulex". Hydrobiologia 589 (2007): 155-
164.
44. MacNeil, C. et al. "Parasite Transmission And Cannibalism In An Amphipod
(Crustacea)". International Journal for Parasitology 33.8 (2003): 795-798.
45. MacNeil, C., Dick, J.T.A and Elwood, R, W,. "The Trophic Ecology of Freshwater
Gammarus spp. (Crustacea: Amphipoda): Problems and perspectives
concerning the functional feeding group concept” Biological Reviews 72.3 (2007):
349-364.
46. Mehlhorn, Heintz, and Horst Aspöck. Encyclopedia Of Parasitology. Berlin:
Springer, 2008. Print.
47. Near, T. J. "Acanthocephalan Phylogeny And The Evolution Of Parasitism".
Integrative and Comparative Biology 42.3 (2002): 668-677.
48. Nelson, D. 'Gammarus-Microbial Interactions: A Review'. International Journal of
Zoology 2011 (2011): 1-6.
49. Nicholas W. L., Hynes H. B. N. 1958: Studies on Polymorphus minutus (Goeze,
1782) (Acanthocephala) as a parasite of the domestic duck. Ann. Trop. Med.
Parasitol. 52: 36-47.
50. Parker, Geoff A. et al. "Evolution Of Complex Life Cycles In Helminth Parasites".
Nature 425.6957 (2003): 480-484. Web.
51. Parsons, J. Cannibalism in herring gulls. British Birds (1971) 64:528-37
52. Pearce, J.M. "Philopatry: A Return to Origins". (2007) The Auk 124(3): 1085–
1087.
53. Petroschenko, V. (1958) Acanthocephala of Domestic and Wild Animals, Vol. II.
Akademiya. 2-44
54. Pfennig, D. W., Loeb M. L. G., and Collins J. P. "Pathogens As A Factor Limiting
The Spread Of Cannibalism In Tiger Salamanders". Oecologia 88.2 (1991): 161-
166. Web.
55. Pfenning, D.W., Ho, S.G., and Hoffman, E. A. "Pathogen Transmission As A
Selective Force Against Cannibalism". Animal Behaviour 55 (1998): 1255-1261.
Print.
56. Polis, G A. "The Evolution And Dynamics Of Intraspecific Predation". Annual
Review of Ecological Systems. 12.1 (1981): 225-251. Web.

31
57. Popova, O. The ‘predator prey’ relationship among fishes. The Biological Basis
of Freshwater Fish Production, ed. S. Gerking, pp 359-376. (1967) Oxford:
Blackwell. 495 pp.
58. Poulin R Meta-analysis of parasite-induced behavioural changes. Animal
Behaviour (1994) 48:137–146
59. Poulin, R. ‘Adaptive’ changes in the behaviour of parasitized animals: a critical
review. Int. J. Parasitol. (1995)25, 1371–1383
60. Poulin, R., Parasite Manipulation of Host Behavior: An Update and Frequently
Asked Questions. In H. Jane Brockmann, editor: Advances in the Study of
Behavior, Vol. 41, Burlington: Academic Press, 2010, pp. 151-186. ISBN: 978-0-
12-380892-9
61. Poulin, R. "Parasite Manipulation Of Host Personality And Behavioural
Syndromes". Journal of Experimental Biology 216.1 (2012): 18-26. Web.
62. Reale, D. et al. "Evolutionary And Ecological Approaches To The Study Of
Personality". Philosophical Transactions of the Royal Society B: Biological
Sciences 365.1560 (2010): 3937-3946. Web.
63. Rudolf V. H. 2007 Consequences of stage-structured predators: cannibalism,
behavioral effects, and trophic cascades. Ecology 88, 2991–3003.
(doi:10.1890/07-0179.1)
64. Rudolf, V. H.W, and J. Antonovics. "Disease Transmission By Cannibalism: Rare
Event Or Common Occurrence?". Proceedings of the Royal Society B: Biological
Sciences 274.1614 (2007): 1205-1210.
65. Sexton, E. W. "On The Rearing And Breeding Of Gammarus In Laboratory
Conditions". Journal of the Marine Biological Association of the United Kingdom
15.01 (1928): 33.
66. Sherman, P. W. "Kinship, Demography, And Belding's Ground Squirrel
Nepotism". Behavioral Ecology and Sociobiology 8.4 (1981): 251-259. Web.
67. Shilland, P. et al. Revised Risk Assessment Policy For Surface Water
Abstractions From Lakes. Dublin City Council, 2009. Print. Eastern River Basin
District Project Abstractions- National POM/ Standards Study.
68. Smyth, J. D, and Wakelin, D. Introduction To Animal Parasitology. Cambridge,
Eng.: Cambridge University Press, 1994. Print.
69. Sutcliffe, David. Subspecies, Morphs And Clines In The Amphipod Gammarus
Duebeni From Fresh And Saline Waters. Freshwater Biological Association,
2010. Print.

32
70. Tarascheweski, H. "Host-Parasite Interactions In Acanthocephala: A
Morphological Approach". Advanced Parasitology 46.1-179 (2000): n. pag. Print.
71. Thompson, S. N., and M. Kavaliers. "Physiological Bases For Parasite-Induced
Alterations Of Host Behaviour". Parasitology 109.S1 (1994): S119-S138. Web.
72. Ward, P. I. "A Comparative Field Study Of The Breeding Behaviour Of A Stream
And A Pond Population Of Gammarus Pulex (Amphipoda)". Oikos 46.1 (1986):
29.
73. Wissinger, S. A. "Niche Overlap And The Potential For Competition And
Intraguild Predation Between Size-Structured Populations". Ecology 73.4 (1992):
1431

33
Appendix Results Table 4 Temperatures of jar during experimentation period.
Jar Number
Infection Status
Temperature in Degrees Celsius 0 Hour 12 Hour 24 Hour 36 Hour 48 Hour
1 Uninfected 14.0 14.0 14.1 14.0 14.0 2 Uninfected 14.4 14.3 14.4 14.3 14.4 3 Uninfected 14.0 14.0 14.2 14.1 14.0 4 Uninfected 14.1 14.0 14.0 14.1 14.2 5 Uninfected 14.0 14.0 14.0 14.0 14.0 6 Uninfected 14.0 14.1 14.1 14.1 14.1 7 Uninfected 14.3 14.3 14.1 14.1 14.2 8 Uninfected 14.0 14.0 14.0 14.0 14.0 9 Uninfected 14.0 14.1 14.2 14.2 14.0 10 Uninfected 14.1 14.2 14.0 14.0 14.0 11 Infected 14.0 14.0 14.0 14.0 14.0 12 N/A x x x x x 13 Infected 14.2 14.1 14.1 14.1 14.1 14 Infected 14.3 14.4 14.2 14.3 14.3 15 Infected 14.0 14.1 14.0 14.0 14.0 16 Infected 14.4 14.3 14.0 14.1 14.0 17 Infected 14.0 14.0 14.2 14.0 14.0 18 Infected 14.0 14.0 14.1 14.0 14.0 19 Infected 14.1 14.1 14.1 14.0 14.0 20 Infected 14.1 14.1 14.1 14.1 14.1 21 Control 14.0 14.1 14.2 14.0 14.0 22 Control 14.1 14.0 14.1 14.0 14.0 23 Control 14.1 14.1 14.3 14.1 14.0 24 Control 14.0 14.1 14.2 14.1 14.0 25 Control 14.1 14.2 14.1 14.0 14.3 26 Control 14.4 14.2 14.2 14.4 14.1 27 Control 14.0 14.2 14.0 14.0 14.2 28 Control 14.2 14.0 14.0 14.2 14.0 29 Control 14.2 14.0 14.0 14.0 14.0 30 Control 14.1 14.0 14.0 14.0 14.1
Temperature was found to not deviate significantly from 14 degrees Celsius.
Temperatures deviates the most at the 0 hour period, as the door had been opened and
closed often just previous to temperature reading, during experimental set up. Slightly
warmer jars tended to be closer to the door. The adult in jar 12 died before
experimentation began, and jar was removed.