
MEMBRANE-BOUND ELECTRON TRANSFER AND ATP …webspace.qmul.ac.uk/rwjanes/basic_19_16_web.pdf ·...
51
MEMBRANE-BOUND ELECTRON TRANSFER AND ATP SYNTHESIS (taken from Chapter 18 of Stryer)
Transcript of MEMBRANE-BOUND ELECTRON TRANSFER AND ATP …webspace.qmul.ac.uk/rwjanes/basic_19_16_web.pdf ·...

MEMBRANE-BOUND
ELECTRON TRANSFER AND
ATP SYNTHESIS (taken from
Chapter 18 of Stryer)

FREE ENERGY – MOST USEFUL
THERMODYNAMIC CONCEPT IN BIOCHEMISTRY
Living things require an input of free energy for 3 major
purposes
1. Mechanical – Muscle contraction and other cellular
movement
2. Active transport of molecules and ions
3. Synthesis of macromolecules and other biomolecules
from simple precursors

First law of thermodynamics
“Energy can be neither created nor
destroyed”
But, it can be converted from one
form into another

Free energy for these processes
comes from the environment
Phototrophs - obtained by trapping light energy
Chemotrophs – energy by oxidation of foodstuffs
Free energy donor for most energy requiring processes is
Adenosine triphosphate (ATP)

ATP
Large amounts of free energy is liberated when ATP is
hydrolysed to ADP + Pi or AMP + PPi
ATP is continuously formed and consumed
Phototrophs harness the free energy in light to generate ATP -
Photosynthesis
Chemotrophs form ATP by oxidation of fuel molecules –
Oxidative phosphorylation

OXIDATIVE
PHOSPHORYLATION
Glucose is converted to pyruvate
And under aerobic conditions undergoes oxidative
decarboxylation to form AcCoA which is then oxidised to
CO2 by the citric acid cycle

Stages of Catabolism
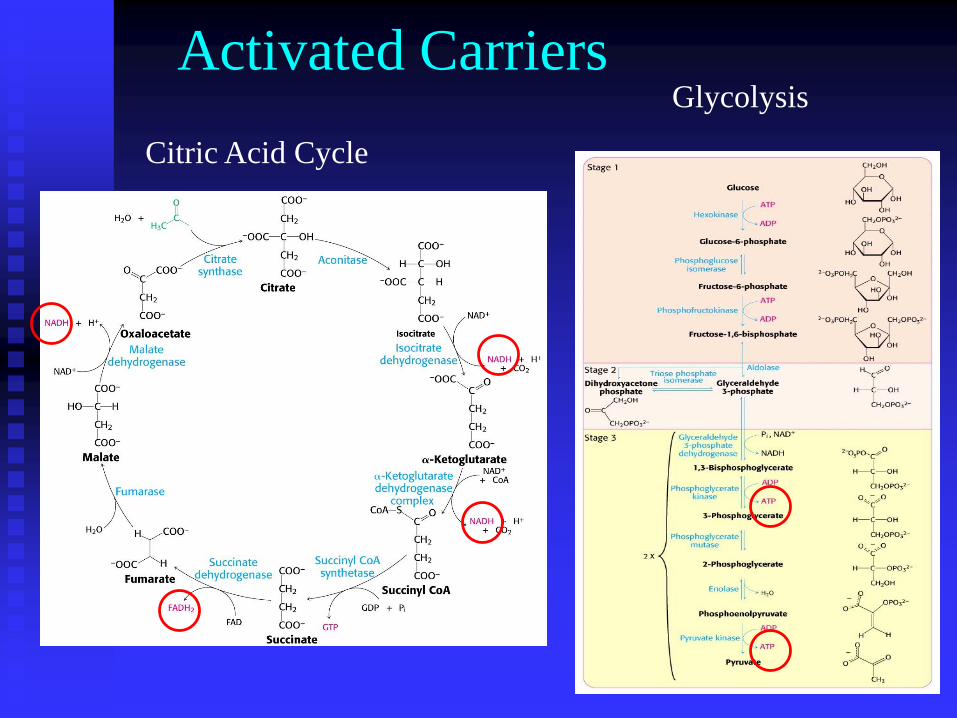
Citric Acid Cycle
Glycolysis
Activated Carriers

These pathways along with fatty acid oxidation produce
energy rich molecules NADH and FADH2 as well as small
amounts of ATP
Chemotrophs derive energy from oxidation of fuel molecules
and in aerobic organisms the ultimate electron acceptor is O2
Electron is not transferred directly
Electron is transferred through special carriers, Pyridine
nucleotides
Electron acceptor Electron donor
NAD+ NADH
FAD FADH2
Activated Carriers

Respiratory electron transfer is the transfer of electrons
from the NADH and FADH2 (formed in glycolysis, fatty
acid oxidation and the citric acid cycle) to molecular
oxygen, releasing energy.
Oxidative phosphorylation is the synthesis of ATP from
ADP and Pi using this energy.
Both processes are located on the IMM
Electron transfer and
oxidative phosphorylation

Mitochondrion

Outer membrane•Permeable (12000da)
•Porin – 30-35kd pore forming protein
Inner membrane•Impermeable all ions and polar molecules
•Possess family of transporter molecules (for
ATP/ADP , Pi , pyruvate, citrate , etc.).
•Matrix side (N-negative), cytosolic side (P-
postive)
Mitochondrion

Mitochondria are the result of an
Endosymbiotic event
Organelles contain their own DNA which encode 13
respiratory chain proteins
Many proteins encoded by cell nuclear DNA
Cells depend on organelle for oxidative phosphorylation ,
mitochondrion depend on cell for their very existence
Suggested that all extant mitochondria are derived from
bacterial Rickettsia prowazekii

Oxidative phosphorylation is conceptually
simple and mechanistically complex.
Flow of electrons from NADH and FADH2 to
O2 occurs via protein complexes located in the
IMM
Leads to the pumping of protons from the
matrix to the cytosol across the IMM.
ATP is synthesised when protons flow back
into the matrix via a protein complex in the
IMM.
Oxidative phosphorylation
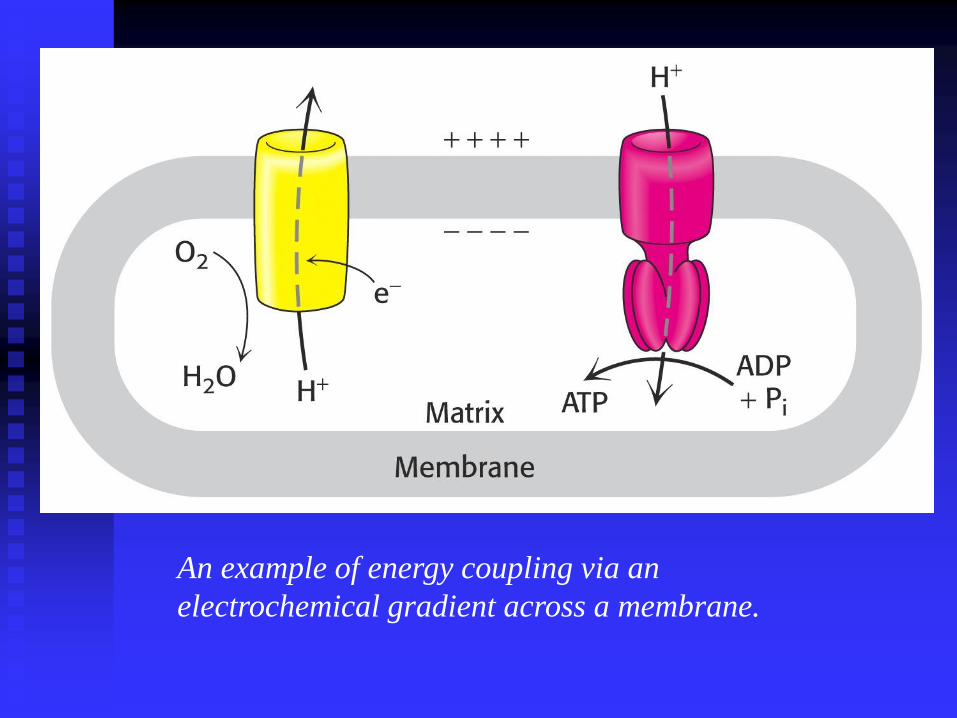
An example of energy coupling via an
electrochemical gradient across a membrane.

REDOX POTENTIAL AND
FREE ENERGY CHANGES
The energy stored in ATP is expressed as the phosphoryl
transfer potential which is given by Go for hydrolysis of
ATP (-7.3kcal/mol)
The electron transfer potential of NADH is represented as Eo
the redox potential ( or reduction potential or oxidation-
reduction potential) which is an electrochemical concept.
Redox potential is measured relative to the H+: H2 couple
which has a defined redox potential of 0V (Volts).

Redox couples
X- + H+ X + 1
2H2
X- X + e-
H+ + e- 1
2H2

A negative redox potential means that a substance has a lower
affinity for electrons than H2 .
A positive redox potential means a substance has a higher
affinity for electrons than H2.
NAD+/ NADH at -0.32V is a strong reducing agent and
poised to donate electrons
1/2 O2/ H2O at +0.82V is a strong oxidising reagent and
poised to accept electrons.
1/2 O2 + NADH + H+ H2O + NAD+
The difference (Eo = 1.14V) is equivalent to -52.6
kcal/mole.
Redox potential


Electrons can be transferrred between groups that are
not in contact
Distance between electron carrying
groups

THE RESPIRATORY ELECTRON TRANSFER
CHAIN CONSISTS OF THREE PROTON PUMPS
LINKED BY TWO MOBILE ELECTRON
CARRIERS
Electrons are
transferred from
NADH to O2 by a
chain of three large
transmembrane
respiratory chain
protein complexes
I
II
III
IV

These are
a) Complex I also known as
NADH-Ubiquinone (UQ) oxidoreductase
NADH-Q reductase
b) Complex III also known as
Ubiquinol (UQH2)-Cytochrome c oxidoreductase
Cytochrome reductase
c) Complex IV also known as
Cytochrome c- Oxygen oxidoreductase
Cytochrome oxidase
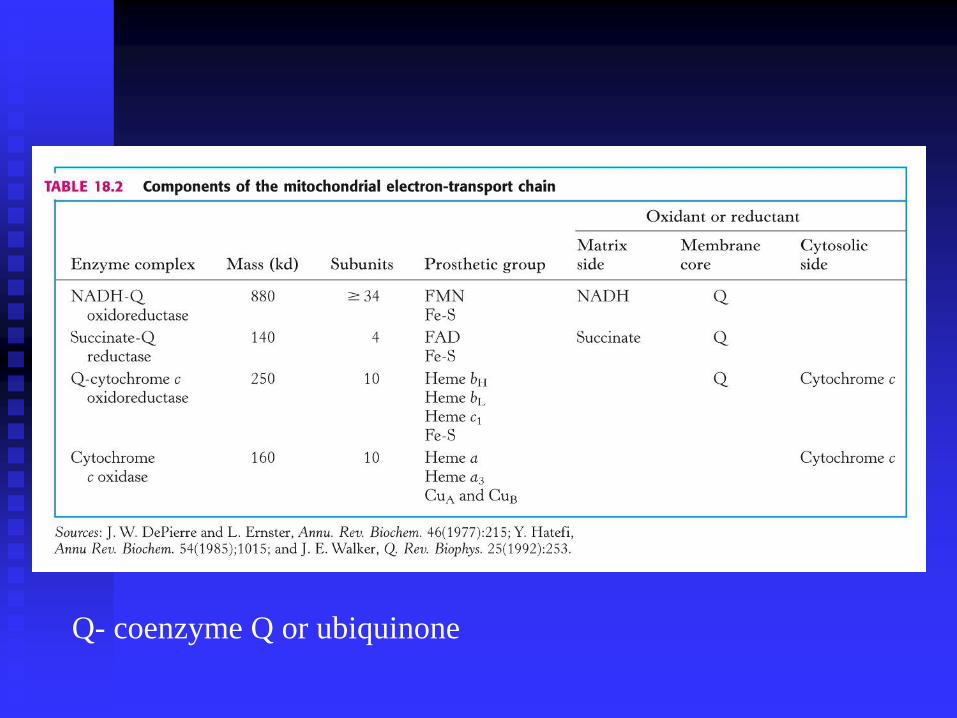
Q- coenzyme Q or ubiquinone

NADH-Q Oxidoreductase
x 2e-

NADH-Q reductase
NADH transfer of e- to flavin
mononucleotide to produce FMNH2
e- from FMNH2 transferred to iron sulfur
clusters
e- from iron sulfur (Fe-S) clusters shuttle to
coenzyme Q (ubiquinone)
Results in pumping of 4 H+ out of matrix
NADH + Q + 5H+matrix NAD+ +QH2 + 4H+
cytosol

Succinate Q reductase
FADH2 already part of complex, transfers
electrons to Fe-S centres and then to Q
This transfer does not result in transport of
protons

Q-cytochrome c Oxidoreductase

Q-cytochrome c Oxidoreductase
Transfers e- from QH2 (2 e- ) cytochrome c
(1 e- ) via heme
Mechanism known as Q cycle
QH2 + 2Cyt cox + 2H+matrix Q +2Cyt cred + 4H+
cytosol

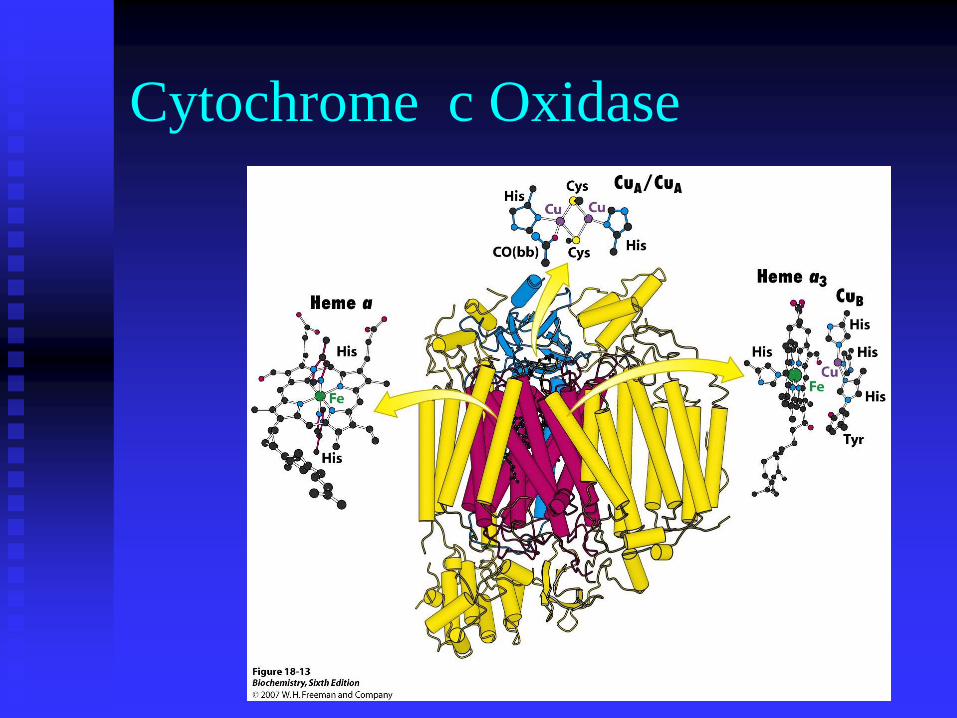
Cytochrome c Oxidase

Cytochrome c Oxidase

Proton transport by cytochrome c
oxidase


Electrons are carried from Complex I to Complex III
by UQH2, the hydrophobic quinol (reduced quinone)
diffuses rapidly within the IMM.
Electrons are carried from Complex III to Complex IV
by cytochrome c, a small hydrophilic peripheral
membrane protein located on the cytosolic or P side of
the IMM.
Complex II (Succinate-UQ oxidoreductase) is
membrane bound and contains the FADH2 as a
prosthetic group . So electrons from FADH2 feed in to
UQH2.
These respiratory chain complexes contain redox
groups to carry the electrons being transferred through
them. These are flavins, iron-sulfur clusters, haems
and copper ions.

PROTON PUMPS AND THE
ATP SYNTHASE
The free energy change of the reactions catalysed by
Complexes I, III and IV is large enough for them to
pump protons from the matrix or N side of the IMM
to the cytosolic or P side of the IMM.
There is not enough energy released in Complex II,
so no proton pumping occurs in this complex.

OXIDATION AND
PHOSPHORYLATION ARE
COUPLED BY A PROTON-
MOTIVE FORCEThis is the chemiosmotic hypothesis put forward by
Peter Mitchell in 1961.
Transfer of electrons from NADH (or FADH2) to
oxygen leads to the pumping of protons to the
cytosolic side of the IMM.
The H+ concentration (pH) becomes higher (lower
pH) on the cytosolic side, and an electrical
potential (membrane potential) with the cytosolic
side of the IMM positive is generated


So a proton-motive force (p) is generated which consists
of both a pH and a .
Mitchell proposed that this proton-motive force drives the
synthesis of ATP by another transmembrane protein
complex, as the protons return back across the IMM
through this protein complex.
This protein complex is called the ATPase (because like any
enzyme it is reversible and was first discovered by it’s
ability to hydrolyse ATP)
It’s preferred name is the ATP synthase.
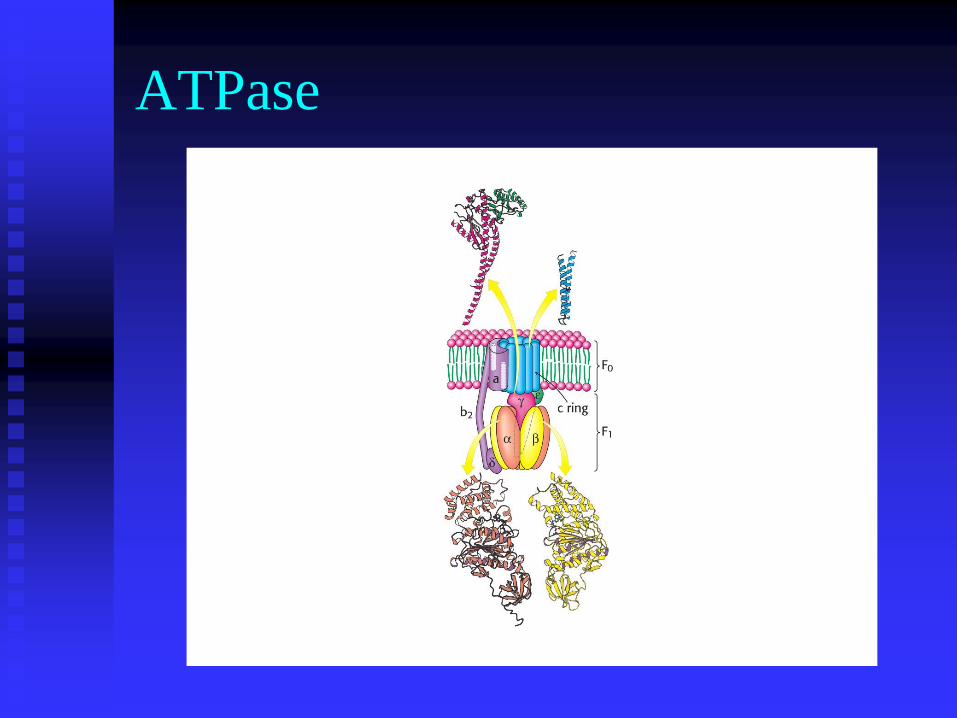
ATPase
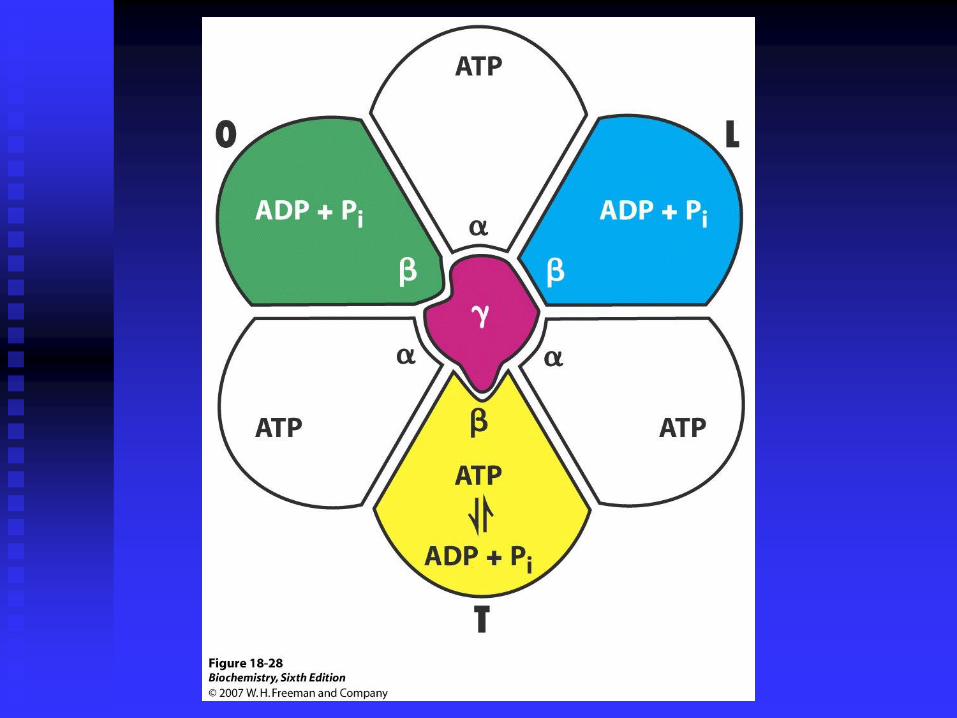


It is now thought that the proton-motive force induces a
conformational change in the ATP synthase, which allows
the release of tightly bound ATP (the product) from the
enzyme, and thus catalyses ATP synthesis.
So this is an example of energy coupling via an activated
protein conformation.

C-ring


Rotates 360º
Requires 3 protons
per molecule of
ATP formed
10 subunits
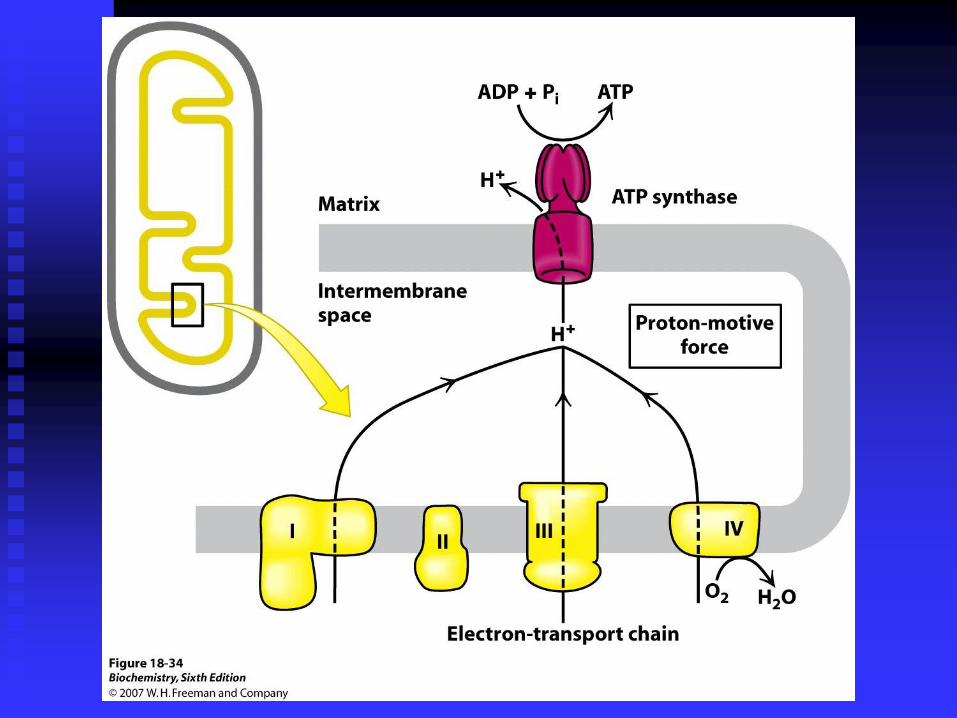

THE
COMPLETE
OXIDATION OF
GLUCOSE
YIELDS ABOUT
30 ATP

Net Yield per glucose
Glycolysis 2 ATP
Citric Acid cycle 2 ATP (GTP)
Oxidative phosphorylation ~26 ATP
Most of the ATP is generated by oxidative phosphorylation
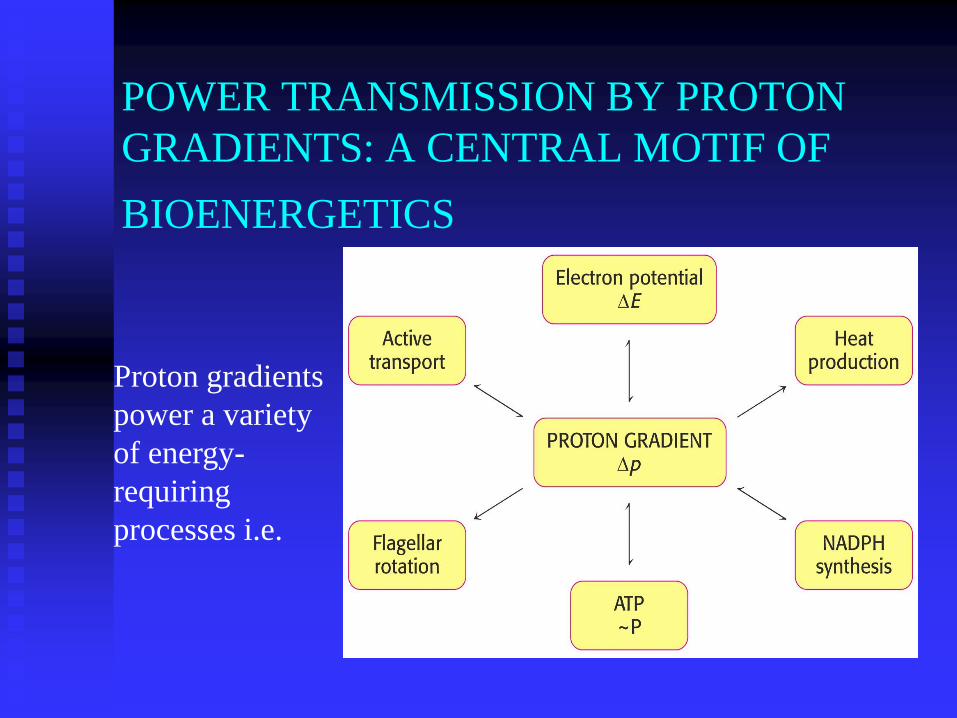
POWER TRANSMISSION BY PROTON
GRADIENTS: A CENTRAL MOTIF OF
BIOENERGETICS
Proton gradients
power a variety
of energy-
requiring
processes i.e.

IT IS EVIDENT THAT PROTON GRADIENTS ARE A
CENTRAL INTERCONVERTIBLE CURRENCY OF FREE
ENERGY IN BIOLOGICAL SYSTEMS.
THE RATE OF OXIDATIVE PHOSPHORYLATION IS
DETERMINED BY THE NEED FOR ATP
Under most physiologic conditions, electron transfer is tightly
coupled to phosphorylation. Electrons do not usually flow
through the electron transfer chain unless ADP is
simultaneously phosphorylated to ATP.
Oxidative phosphorylation and thus electron transfer require a
supply of
NADH
O2
ADP and Pi

Regulation
The most important factor controlling the rate of oxidative
phosphorylation is the level of ADP
Regulated by the energy charge.
This regulation of the rate of oxidative phosphorylation by the
ADP level is called respiratory control.


















