
Viruses as a Model System for Studies of Eukaryotic mRNA Processing

1
Short title: Mechanisms of long-distance mRNA movement 1
2
*Corresponding Authors: 3
Cankui Zhang 4
Department of Agronomy 5
Purdue University 6
West Lafayette, IN 47907 7
Phone: +1 765-496-6889 8
Email: [email protected] 9
Zhangjun Fei 10
Boyce Thompson Institute 11
Cornell University 12
Ithaca, NY 14853, USA 13
Phone: +1 607-254-3234 14
Email: [email protected] 15
16
Plant Physiology Preview. Published on May 2, 2018, as DOI:10.1104/pp.17.01836
Copyright 2018 by the American Society of Plant Biologists
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
Article title: Elucidation of the mechanisms of long-distance mRNA movement in a Nicotiana 17
benthamiana/tomato heterograft system 18
19
Chao Xia1,2,3†
, Yi Zheng4†
, Jing Huang1, Xiangjun Zhou
1, Rui Li
5, Manrong Zha
1, Shujuan 20
Wang1, Zhiqiang Huang
6, Hai Lan
2,3, Robert Turgeon
7, Zhangjun Fei
4,8*, and Cankui Zhang
1,9* 21
22
1Department of Agronomy, Purdue University, West Lafayette, IN, 47907, USA 23
2Maize Research Institute, Sichuan Agricultural University, Chengdu, Sichuan 611130, China 24
3Key Laboratory of Biology and Genetic Improvement of Maize in Southwest Region, Ministry 25
of Agriculture, China 26
4Boyce Thompson Institute, Cornell University, Ithaca, NY 14853, USA 27
5College of Agronomy, Sichuan Agricultural University, Chengdu, Sichuan 611130, China 28
6Biost Technology Co., Ltd, Beijing 102206, China 29
7Plant Biology Section, School of Integrative Plant Science, Cornell University, Ithaca, NY 30
14853, USA 31
8USDA-ARS Robert W. Holley Center for Agriculture and Health, Ithaca, NY 14853, USA 32
9Purdue Center for Plant Biology, Purdue University, West Lafayette, IN, 47907, USA 33
34
One-sentence summary: Messenger RNA is degraded as it moves through the tomato phloem 35
and the mobility of a transcript cannot be reliably predicted based on its abundance in the 36
Nicotiana benthamiana leaf or on whether it harbors a tRNA-like structural motif. 37
Author Contributions: C.Z., R.T., C.X., and X.Z. conceived and designed the experiments; 38
C.X., J.H., M.Z., and S.W. performed N. benthamiana/tomato grafting in hydroponic 39
experiments; C.X. and R.L. performed transgenic potato grafting experiments; C.X. performed 40
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

3
long-stem grafting, split shoot grafting, and canola/Arabidopsis grafting; Z.Y., Z.F., C.X., and 41
Z.H. devised and implemented the bioinformatics methodology and analysis; C.X. and C.Z. 42
wrote the manuscript; C.Z., Z.F., R.T., and H.L. revised and finalized the manuscript. 43
Funding Information: This work was supported by funds from Purdue University as part of 44
AgSEED Crossroads funding to support Indiana’s Agriculture and Rural Development (to C. Z.), 45
National Key Research and Development Project of China (2016YFD0101206), and the US 46
National Science Foundation (IOS-1339287 and IOS-1539831). 47
†These authors contributed equally to the article. 48
The author responsible for distribution of materials integral to the findings presented in this 49
article in accordance with the Journal policy described in the Instructions for Authors 50
(http://www.plantphysiol.org) is: Cankui Zhang ([email protected]). 51
*Corresponding Authors: 52
Cankui Zhang 53
Email: [email protected] 54
Zhangjun Fei 55
Email: [email protected] 56
57
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

4
ABSTRACT 58
Recent heterograft analyses showed that large-scale mRNA movement takes place in the phloem, 59
but the number of mobile transcripts reported varies widely. However, our knowledge of the 60
mechanisms underlying large-scale mRNA movement remains limited. In this study, using a 61
Nicotiana benthamiana/tomato (Solanum lycopersicum) heterograft system and a transgenic 62
approach involving potato (Solanum tuberosum), we found that: 1) the overall mRNA abundance 63
in the leaf is not a good indicator of transcript mobility to the root; 2) increasing the expression 64
levels of non-mobile mRNAs in the companion cells does not promote their mobility; 3) mobile 65
mRNAs undergo degradation during their movement; and 4) some mRNAs arriving in roots 66
move back to shoots. These results indicate that mRNA movement has both regulated and 67
unregulated components. The cellular origins of mobile mRNAs may differ between herbaceous 68
and woody species. Taken together, these findings suggest that the long-distance movement of 69
mRNAs is a complex process and elucidating the physiological roles associated with this 70
movement is challenging but remains an important task for future research. 71
72
Keywords: heterograft, phloem, mobile mRNA, long-distance, mRNA degradation, mRNA 73
cycling, TLS motif, RNA-Seq 74
75
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

5
INTRODUCTION 76
Higher plants have evolved a communication system that enables the coordination of 77
developmental and environmental cues between different organs (Spiegelman et al., 2013; 78
Turnbull and Lopez‐Cobollo, 2013). This communication is achieved by long-distance signaling 79
that takes place in the vasculature tissue (Banerjee et al., 2006). Phloem, one of the major 80
components in the vasculature system, has long been recognized as a tissue that transports 81
carbohydrates and amino acids. In recent years, it has been found that this tissue is also a conduit 82
for signals, e.g., mRNAs, small RNAs, proteins, small peptides, and hormones (Turgeon and 83
Wolf, 2009). 84
Although mRNAs have been traditionally viewed as local intermediate components between the 85
genomic DNA and the protein in a cell, a handful of classical studies have shown that some 86
mRNAs are able to traffic from source to sink tissues via the phloem and exert important 87
physiological functions in the distal recipient organs. For example, the development of sink 88
tissues, such as young leaves (Kim et al., 2001), tubers (Banerjee et al., 2006), and roots 89
(Notaguchi et al., 2012), are partially controlled by the long-distance mobile mRNAs generated 90
in the source tissues. 91
A recent study on the interaction between the parasitic Cuscuta pentagona and its host indicated 92
that a large amount of bidirectional mRNA exchange occurred via the vasculature between the 93
two species (Kim et al., 2014). In recent years, a few heterograft systems, in which one 94
species/genotype was used as a scion and another as a rootstock, were used to demonstrate that 95
large-scale migration of mRNAs occurs from leaf to root, or vice versa. Notaguchi et al. (2015) 96
determined that 138 transcripts moved from the rootstock of Arabidopsis thaliana to the 97
Nicotiana benthamiana scion via the phloem. Using two different ecotypes of Arabidopsis, 98
Thieme et al. (2015) determined that 2,006 mRNAs could move from the shoot to the root, or 99
vice versa, under either normal or mineral-deficient conditions. In a grapevine (Vitis vinifera) 100
heterograft system, a total of 3,333 mRNAs were found to be transmissible (Yang et al., 2015). 101
In another heterograft system, an analysis of the response to early phosphate deficiency revealed 102
that 3,546 mRNAs moved from cucumber (Cucumis sativus) source leaves to watermelon 103
(Citrullus lanatus) sink tissues via the phloem (Zhang et al., 2016). 104
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

6
This variability, with over an order of magnitude differences in the number of mRNAs 105
transported, suggests a great deal of species specificity and that not all the mobile mRNAs are 106
functional. Zhang et al. (2016) discovered that among the large number of mobile mRNAs 107
identified in the Arabidopsis heterografts, 11.4% of them harbored a tRNA-like structure (TLS). 108
Further experimental evidence supported the hypothesis that not only does this motif impart 109
stability and mobility to mRNAs, but also that mRNAs with this structure can be translated into 110
proteins after transport (Zhang et al., 2016). However, how this mechanism can be applied to 111
other plant heterograft systems remains unknown. A computational analysis conducted by 112
Calderwood et al. (2016) on the same set of mobile mRNAs suggested that most of the identified 113
mRNA species are mobile as a consequence of local abundance in companion cells. Similar 114
correlation between abundance and long-distance mobility has been noted by other researchers as 115
well (Kim et al., 2014; Yang et al., 2015). The extent to which this correlation, drawn from either 116
mathematical modeling methods or computational analysis, applies to natural plant systems 117
needs further experimental verification. 118
In this study, we developed a heterograft system using Nicotiana benthamiana and tomato 119
(Solanum lycopersicum) to investigate shoot-to-root mRNA movement. Compared to most 120
published heterograft systems, the phylogenetic distance between these two species is relatively 121
large, which allowed for more accurate distinction of scion mRNAs from those in the rootstock. 122
In addition, the ease of transformation of the two partners is advantageous in future molecular 123
and physiological studies. We used this system to test if: 1) the abundance of mRNAs in the leaf 124
is a determining factor for mRNA mobility; 2) the movement of mRNAs is a regulated or 125
unregulated process; 3) mobile mRNAs undergo degradation during their movement from shoots 126
to roots; and 4) shoot-to-root-to-shoot cycling movement occurs. In addition, a transgenic 127
approach involving potato (Solanum tuberosum) was used to test the relationship between the 128
abundance of companion cell transcripts and their shoot-to-root mobility. 129
130
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

7
RESULTS 131
mRNA migration from shoot to root 132
A heterograft system, in which N. benthamiana was used as the scion and tomato as the 133
rootstock, was established to identify the shoot-to-root long-distance movement of mRNAs 134
through the phloem (Fig. 1A). We compared the morphology of leaves and roots in both the 135
heterografts and the non-grafted counterparts. No visible phenotypic differences were observed, 136
indicating that the heterografts had largely maintained the endogenous physiology of the non-137
grafted plants (data not shown). 138
A total of 183 N. benthamiana mRNAs were determined to move into the tomato root in 139
heterografts grown in a hydroponic system (Supplementary Table 1). Gene ontology (GO) 140
analysis of these mobile mRNAs indicated that certain biological processes, e.g., translation 141
(19.1%), transport (14.2%), and biosynthetic process (14.8%), were over-represented. Within the 142
cellular component category, ribosome (15.3%), membrane (28.4%), and intracellular (14.8%) 143
were over-represented. Moreover, nucleotide binding (20.8%), transporter activity (16.9%), and 144
RNA binding (7.1%) were over-represented in the molecular function category (Fig. 1B and 145
Supplementary Table 2). 146
Orthologues of some of the mRNAs identified from this work were also reported in previous 147
studies in which traditional phloem collection methods were used (Ivashikina et al., 2003; 148
Doering-Saad et al., 2006; Omid et al., 2007; Deeken et al., 2008; Kanehira et al., 2010). For 149
example, IAA1, the mRNA encoding an auxin-responsive protein, was identified as mobile in our 150
system. A previous analysis discovered a member of the Aux/IAA family encoding a 151
transcriptional regulator in auxin signaling in the phloem sap of melon (Cucumis melo) (Omid et 152
al., 2007). IAA18 and IAA28, two additional auxin signaling-related transcripts, were found to be 153
expressed in the vasculature tissue of mature Arabidopsis leaves, and transported to the root tip 154
to regulate lateral root formation (Notaguchi et al., 2012). In addition, a number of mRNAs 155
encoding essential translation machinery components, e.g., ribosomal protein, elongation factor, 156
and eukaryotic translation initiation factor, were found to be mobile. Previous proteome studies 157
have discovered that many proteins related to translation exist in the phloem (Lin et al., 2009; 158
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

8
Ma et al., 2010; Ham and Lucas, 2014) even though the translational machinery is absent in 159
sieve elements. 160
Abundance of mRNA is not associated with mobility 161
To determine whether more abundant leaf mRNAs are prone to move, we collected mature 162
leaves from the N. benthamiana scions in the N. benthamiana/tomato heterograft and sequenced 163
the mRNAs. The abundances of the transcripts in leaves corresponding to the 183 mobile 164
mRNAs were compared with the abundances of the same 183 mRNAs in the heterograft roots. 165
No correlation (r2 = 0.0037) was found between the shoot-to-root mobility of the mRNAs and 166
their abundances in the leaves, the origin of these mobile mRNAs (Fig. 2A and Supplementary 167
Table 3). Further detailed analysis showed that none of the 100 most abundant leaf mRNAs 168
moved from shoots to roots; among the 183 mobile mRNAs, only 3 transcripts were ranked 169
among the 183 most abundant leaf mRNAs (Fig. 2B). On the contrary, some mRNAs with low 170
abundance in the leaf were able to move to the rootstock. The abundances of approximately half 171
(91 out of 183) of the mobile mRNAs were ranked lower than 30,000th
among the entire leaf 172
transcriptome. For example, the abundance of Niben101Scf00291g02025.1, a mRNA ranked the 173
8th
most mobile, was ranked 30,982th
in abundance in the leaf; Niben101Scf05425g00016.1 was 174
ranked 16th
in the mobile list but the 33,512th
most abundant in the leaf transcriptome. Although 175
most of the shoot-to-root mobile mRNAs had higher abundances in the scion than in the 176
rootstock, it is noteworthy that the abundances of 27 mobile mRNAs were higher in the rootstock 177
than in the leaf. For example, Niben101Scf01796g01005.1, an mRNA that was ranked the 17th
178
most mobile, had a reads per kilobase of exon per million fragments mapped (RPKM) value of 179
1.14 in the rootstock but only 0.20 in the leaf (the 40,445th
most abundant in the leaf 180
transcriptome) (Supplementary Table 4). 181
Overexpressing phloem immobile mRNAs in the companion cells does not promote 182
mobility 183
The lack of correlation between the mobility of mRNAs and their abundances in the leaf 184
indicated that using whole leaf tissues is not informative enough to predict the mobility of 185
mRNAs. One possibility is that only cells directly connected to the sieve tubes act as sources of 186
mobile mRNAs. In apoplastic loading species, such as N. benthamiana, the phloem is isolated 187
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

9
from surrounding cells and it is generally believed that the components moving in the sieve tubes 188
originate in the associated companion cells (King and Zeevaart, 1974; Stadler et al., 2005). Using 189
a simple computational model, Calderwood et al. analyzed a series of mobile RNA datasets 190
associated with Arabidopsis and postulated that mRNAs with higher abundance in the 191
companion cells are more prone to move (Mustroph et al., 2009; Calderwood et al., 2016). To 192
test this hypothesis, we investigated whether increasing the abundance of a phloem-immobile 193
mRNA will make it mobile. We overexpressed the Arabidopsis dual-affinity nitrate transporter 194
gene (AtCHL1) and the Arabidopsis ammonium transporter gene (AtAMT1;2) in the phloem of 195
potato driven by the strong companion-cell-specific sucrose transporter 2 (SUC2) promoter. 196
Neither of the homologous mRNA transcripts of these two genes were detected to be mobile in 197
our heterograft system. In addition, the phloem immobility of homologues of these two 198
transcripts in previously published heterograft systems was further confirmed by exploring the 199
PlaMoM database (Guan et al., 2016). Previous analysi had shown that the size of the shoot-to-200
root mobile mRNAs can be as large as 4 kb (Notaguchi et al., 2015; Thieme et al., 2015; Zhang 201
et al., 2016). Therefore, the length, i.e., 1,545 bp and 1,773 bp, respectively, of AtAMT1;2 and 202
AtCHL1, should not pose a negative effect to the movement of the individual mRNAs from the 203
companion cell to the sieve tube. Multiple transgenic lines were generated and the transgenic 204
potato plant with the highest expression of either AtCHL1 or AtAMT1;2 in the companion cells 205
was used as the scion that was grafted onto the wild-type potato rootstock (Fig. 3A and 206
Supplementary Fig. 1). The lack of amplification via PCR of these genes in the rootstock of the 207
heterografts indicated that the increased abundances of these phloem-immobile mRNAs 208
transcribed in the companion cells did not promote shoot-to-root movement (Fig. 3B). 209
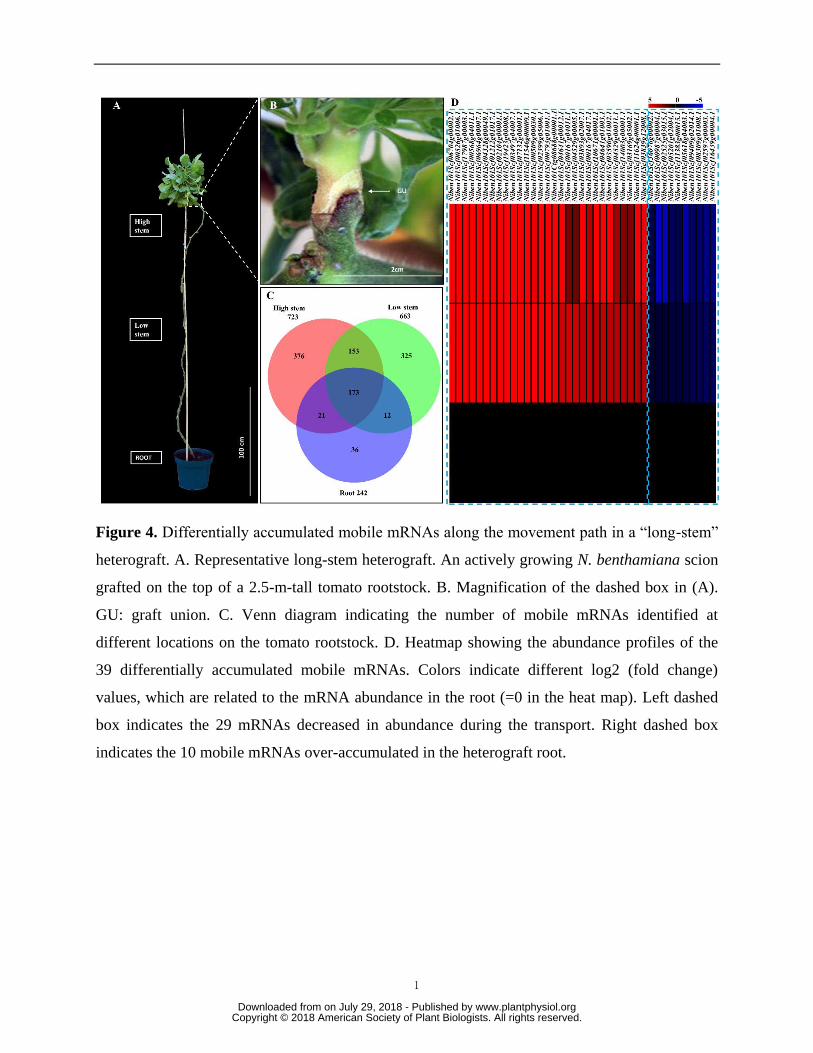
Mobile mRNAs differentially accumulate along the movement path 210
In addition to the cell specificity that might confer the mobility of an mRNA, transport from 211
shoot to root could depend on other factors, such as sequence structure and half-life (Calderwood 212
et al., 2016; Zhang et al., 2016). In the Arabidopsis/Cuscuta system, three host-born mRNAs 213
were chosen as representatives to investigate the fate of mobile mRNAs once entering the 214
parasitic plant. It was found that the accumulation patterns, i.e., increased or decreased 215
abundance, of these mobile mRNAs differed during their transport path (LeBlanc et al., 2013). 216
To understand whether mobile mRNAs accumulate differently once passing the graft joint on a 217
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

10
more global scale, N. benthamiana scions were grafted onto the tops of 2.5-m tall tomato plants 218
(Fig. 4A and B). RNAs were extracted from three locations, high stem (10-20 cm below the 219
grafting joint), low stem (120-130 cm below the grafting joint) and root, and sequenced. The 220
analysis revealed that a total of 1,096 mRNAs from the N. benthamiana scion passed the grafting 221
joint and moved downward to the tomato rootstock. However, 78% (854 transcripts) of the 222
mobile mRNAs did not arrive in the root. The total number of mobile mRNAs decreased from 223
the “high stem” (723) to the “low stem” (663) and the root (242) (Fig. 4C and Supplementary 224
Table 5, 6, and 7). Although the abundance of most mRNAs decreased during the shoot-to-root 225
transport, 10 mRNAs showed higher abundance in the root than in the “high stem” and “low 226
stem” (Fig. 4D, Table 1 and Supplementary Table 8). It is noteworthy that two of these root-227
accumulated mobile mRNAs were annotated as casein kinase I, which has been reported to be 228
involved in plant hormonal metabolism and establishment of root architecture (Liu et al., 2003). 229
Another interesting observation is that some of the mobile mRNAs identified in “low stem” or 230
root were not detected in tissues located above it. For example, 325 mRNAs detected in the “low 231
stem” were not detectable in the “high stem” site; similarly, 36 mRNAs detected in root were not 232
detectable in either location in the stem. A similar phenomenon was observed in the Arabidopsis 233
heterograft, where some of the rootstock mRNAs identified in the scion flowers were not 234
detectable in the inflorescence stem, a tissue mediating the mRNA transport (Thieme et al., 235
2015). One possible explanation for this is that the abundances of these mobile mRNAs are so 236
low that they cannot be readily detected in those tissues using the RNA-Seq analyses. 237
mRNAs move from scion to rootstock to scion 238
In addition to the shoot-to-root movement of mRNAs (Thieme et al., 2015; Yang et al., 2015; 239
Zhang et al., 2016), Arabidopsis and grapevine heterograft systems indicate that mRNAs 240
transcribed in roots can also move from roots to shoots (Thieme et al., 2015; Yang et al., 2015). 241
In these previous studies, two separate groups of mRNAs, originating either from leaves or roots, 242
were demonstrated to be mobile. We asked whether mRNAs in the rootstock that had arrived 243
from the scion can be transported back to the shoot. In contrast to previous analysis, our interest 244
was only focused on the same set of mRNAs that are transcribed in the leaf. To test this, we 245
developed a system in which potato and N. benthamiana were individually grafted onto two 246
separate stems emerging from the same tomato rootstock (Fig. 5A). The potato scion was first 247
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

11
grafted onto one of the two stems on the tomato rootstock (Fig. 5B). Once the first heterograft 248
was established and the potato scion was actively growing, the young sink leaves on the potato 249
shoot were removed and a N. benthamiana scion was grafted onto the second stem of the tomato 250
rootstock (Fig. 5C). Since only mature source leaves of the potato scion remained, N. 251
benthamiana mRNAs identified in the potato leaves would have to be imported either via the 252
phloem, against source-to-sink bulk flow, or via the non-vasculature cell-to-cell based rootstock-253
to-scion transport approach mediated by plasmodesmata (Lucas et al., 1995). A total of 58 N. 254
benthamiana mobile mRNAs were detected in the mature leaves of the potato scions 255
(Supplementary Table 9). 256
Functional analysis of the 58 scion-to-rootstock-to-scion mobile mRNAs revealed that diverse 257
biological processes were over-represented including those related to translation, transport, and 258
biosynthetic processes (Supplementary Table 10). Within the cellular component category, 259
intracellular, membrane, and thylakoid were over-represented. Moreover, structural molecule 260
activity, transporter activity, and hydrolase activity were over-represented in the molecular 261
function category. 262
Mobile mRNAs are highly system specific 263
Although large-scale movement of mRNAs has been demonstrated, it is not clear how many of 264
the mobile sequences are functional. If the mRNAs do play physiological roles, one would 265
predict sequence conservation between different systems. Identification of the “core” mRNAs 266
shared by these heterograft systems and functional characterization of these mRNAs will shed 267
light on their physiological roles. However, most species involved in these previous systems are 268
not easy to transform, which is a pre-requisite for future in-depth functional analyses of the 269
mobile mRNAs. To address this issue, we developed another heterograft system using canola 270
(Brassica napus) and Arabidopsis, both of which are transformable, as scion and rootstock, 271
respectively (Fig. 6A). Although canola and Arabidopsis are both Brassicaceae species, there is 272
considerable variation in their genome sequences, which facilitates accurate identification of 273
species-specific mRNAs. Surprisingly, RNA-Seq analysis of the Arabidopsis roots only led to 274
the identification of 23 shoot-to-root mobile mRNAs derived from canola (Supplementary Table 275
11). In previous heterograft studies, 1,698, 1,963, and 1,593 shoot-to-root mobile mRNAs were 276
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

12
detected in Arabidopsis, grapevine, and cucumber systems, respectively. In our N. 277
benthamiana/tomato system, 1,163 mRNAs, when counts from all experimental systems were 278
combined, were detected to be mobile from shoot to root (Supplementary Table 12). 279
To explore whether growth condition and physiological status can cause the movement of 280
different sets of mRNAs, we compared the 183 mobile mRNAs identified in the “regular stem” 281
N. benthamiana/tomato heterograft grown in a hydroponic condition to the 242 mobile mRNAs 282
identified in the “long stem” N. benthamiana/tomato heterograft grown in soil. Forty-one and 283
100 mRNAs were found to be “short stem + hydroponic-” and “long stem + soil-” specific, 284
respectively (Supplementary Table 13). The large difference in the number and identity of 285
mobile mRNAs was also observed in the Cuscuta system, where a total of 9,518 or 347 host 286
mRNAs were detected in Cuscuta when the host species was either Arabidopsis or tomato, 287
respectively (Kim et al., 2014). Together, these findings suggest that the movement of mobile 288
mRNAs could be determined by the species involved in the systems as well as other factors such 289
as the growth environment. 290
To detect whether a “core” group of shoot-to-root mobile mRNAs exists in various systems, we 291
compared the identified mobile mRNAs from the N. benthamiana/tomato heterografts with those 292
identified in Arabidopsis, grape, and cucumber heterograft systems (Thieme et al., 2015; Yang et 293
al., 2015; Zhang et al., 2016). Through orthology analysis, we found that only one mobile 294
mRNA, chlorophyll a/b binding protein 8 (Niben101Scf01328g01012), was shared among these 295
four systems (Fig. 6B). A physiological role for the chlorophyll a/b binding protein in the stem 296
and root has not been reported, although previous localization studies have shown that this gene 297
is lowly expressed in the stem and root (Sakamoto et al., 1991; Tada et al., Song et al., 2007). 298
Future functional study specifically targeted to the movement of this transcript will provide more 299
insight into the regulation of the transport of this transcript. A similar analysis was carried out 300
with the canola/Arabidopsis heterograft system and those in Arabidopsis, grape, and cucumber, 301
and no mRNA was found to be shared (Fig. 6C). 302
Harboring TLS motif does not necessarily lead to high mobility 303
Recently, Zhang et al. demonstrated that a 70-73-nt long transfer RNA (tRNA)-like structure 304
(TLS) motif confers shoot-to-root or root-to-shoot mobility of mRNAs and that approximately 305
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

13
11.4% and 7.5% of the mobile mRNAs identified from the Arabidopsis Col/Arabidopsis PED 306
and the grapevine heterografts, respectively, harbor this motif (Zhang et al., 2016). We used 307
TLSfinder in the PlaMoM database to scan the mobile transcripts identified in the N. 308
benthamiana/tomato system (Guan et al., 2016). Among the 1,163 mobile mRNAs, 126 (10.8%) 309
harbor at least one TLS motif in the coding sequence or untranslated region, which was 310
significantly enriched (p < 0.05, hypergeometric test) when compared to the genome background 311
(5,066 TLS-containing genes out of 57,140 total genes; 8.9%). We then asked to what extent this 312
positive relationship is reflected in individual mRNAs. To our surprise, among the top 100 most 313
abundant mRNAs in leaves of our N. benthamiana/tomato heterograft system, 18 harbored the 314
TLS motif but none of them were mobile; among the top 2,000 most abundant mRNAs, 174 315
harbored the TLS motif but only 11 were mobile. In addition, in the long-stem N. 316
benthamiana/tomato heterograft system, we found that the majority of the mobile mRNAs 317
harboring the TLS motif did not arrive in the tomato root. Only 23 of the 122 mobile transcripts 318
having this motif that successfully passed the graft junction were detected in the roots. It is 319
important to note that our discovery on the lack of correlation between mobility and the 320
harboring of the TLS motif does not negate the importance of this motif because it is possible 321
that under certain circumstances, e.g., low minerals, some of the immobile mRNAs containing 322
this motif could become mobile (Thieme et al., 2015; Zhang et al., 2016). 323
324
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

14
DISCUSSION 325
Previous studies using EDTA-facilitated exudation, laser capture micro-dissection (LMC), 326
fluorescence-activated cell sorting (FACS), or cucurbit exudation identified large numbers of 327
mRNAs in the phloem translocation stream (Ivashikina et al., 2003; Doering-Saad et al., 2006; 328
Omid et al., 2007; Deeken et al., 2008; Kanehira et al., 2010). However, the authenticity of these 329
mRNAs was often questioned (Turgeon and Wolf, 2009; Liu et al., 2012; Zhang et al., 2012; 330
Zhang et al., 2014). Recent studies using different heterograft systems strongly suggest the 331
existence of large-scale, long-distance movement of mRNAs via the phloem (Notaguchi et al., 332
2015; Thieme et al., 2015; Yang et al., 2015; Zhang et al., 2016). In addition, several earlier 333
studies on the interaction of the host plants Arabidopsis and tomato with the parasite Cuscuta 334
pentagona have demonstrated that large numbers of host plant mRNAs move to the parasitic 335
species via the phloem (LeBlanc et al., 2013; Kim et al., 2014). These discoveries collectively 336
indicate that movement of mRNAs via the phloem is a common physiological process. In this 337
study, we established a N. benthamiana/tomato heterograft system to identify shoot-to-root 338
phloem-mobile mRNAs. The relatively large differences in genome sequence between the two 339
species allowed confident and exhaustive identification of mobile mRNAs. In addition, we took 340
advantage of this system to study the accumulation pattern of shoot-born mRNAs during their 341
movement to the root. 342
Regulated and unregulated movement of mRNAs 343
It has been demonstrated that some phloem-mobile mRNAs play physiological roles in recipient 344
sink organs such as young leaves, roots, and tubers (Kim et al., 2001; Banerjee et al., 2006; 345
Notaguchi et al., 2012). However, the identification of large numbers of mobile mRNAs 346
(hundreds to thousands) from recent plant-parasitic or heterograft experimental systems and high 347
variation across different systems have raised concerns as to the percentage of these mobile 348
mRNAs associated with physiological functions. Is mRNA transport a regulated physiological 349
and developmental process or due primarily to the leakage of leaf mRNAs into the sieve tubes? 350
In the grapevine study, the number of mobile mRNAs identified from the field-grown 351
heterografts was much smaller than that in the in vitro-grown grafts (Yang et al., 2015). One 352
explanation is that more mobile mRNAs were degraded before reaching the recipient organs in 353
the field-grown grafts due to the longer distance than in the in vitro-grown grafts (Yang et al., 354
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

15
2015). Similar tendencies for RNA movement were reported for both the Arabidopsis grafts and 355
the parasitic plant and its host plant (LeBlanc et al., 2013; Kim et al., 2014; Thieme et al., 2015). 356
Our long-stem N. benthamiana/tomato experiments showed that a total of 1,096 mRNAs passed 357
the graft joint, but 854 of them disappeared, including some of those with the TLS motif, during 358
their movement from shoot-to-root. The disappearance of mRNAs during their movement can be 359
attributed to two reasons. One is that companion cells along the transport phloem in the stem 360
could specifically arrest certain types of mRNAs from the sieve tube for translation or 361
degradation. The other is that there is an RNase in the sieve tube which can degrade mRNAs 362
lacking protection. Although it is generally believed that the phloem is devoid of RNase and 363
therefore RNA degradation is unlikely (Morris, 2018), the small molecular weights (e.g., 25-29 364
kDa; Bariola et al., 1999) of some of the RNase enzymes are certainly an advantage for their 365
movement from the companion cell to the sieve element. Previous studies have shown that the 366
size exclusion limit between companion cells and sieve elements is large enough to allow the 367
transit of a 67-kDa green fluorescent protein (GFP) in Arabidopsis (Stadler et al., 2005). The 368
lack of detection of the RNase activity in the phloem may be due to inadequate sensitivity of the 369
chemical kits used for the assay (Sasaki et al., 1998; Doering‐Saad et al., 2002). It is reasonable 370
to assume that the movement of mRNAs undergoing degradation in the phloem is not regulated, 371
and therefore it is less likely that there are physiological functions associated with them. 372
However, evidence of a regulated process for mRNA movement exists. It was demonstrated that 373
the long-distance trafficking of StBEL5 in potato is regulated and plays a physiological role in 374
tuberization (Banerjee et al., 2006). A recent study by Zhang et al. (2016) demonstrated that a 375
TLS motif confers mobility for mRNAs. In the grapevine study, some mRNAs were transmitted 376
at rates higher than random transmission (Yang et al., 2015). In our N. benthamiana/tomato 377
grafts, a number of mRNAs had higher abundances in the rootstock than in the leaf, which is the 378
origin of the synthesis of these mobile mRNAs (Table 1; Supplemental Table 4). A mRNA 379
encoding casein kinase I was discovered to be mobile and accumulated in both the regular and 380
the “long-stem” N. benthamiana/tomato heterografts. In rice (Oryza sativa), casein kinase I is 381
involved in auxin metabolism and root development. Arabidopsis plants with downregulated 382
casein kinase I have fewer lateral and adventitious roots as well as shortened primary roots (Liu 383
et al., 2003). It is intriguing to study whether the long-distance movement of the casein kinase 384
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

16
mRNA plays a role in the establishment of root architecture. Transcript abundance in the 385
companion cells was proposed as one of the mechanisms related to mRNA mobility 386
(Calderwood et al., 2016). However, concerns associated with the mathematical model used to 387
draw the conclusion have been raised (Morris, 2018). Our transgenic potato experiment showed 388
that the abundance itself is not enough to lead to the movement of a mRNA because 389
overexpressing two phloem-immobile mRNAs, AtCHL1 or AtAMT1;2, in the companion cells 390
did not promote their shoot-to-root movements (Fig. 3A and B). This result is in agreement with 391
a recent study in which the overexpression of GFP in the companion cells did not lead to 392
movement of the transcript from the leaf to the root (Paultre et al., 2016). It is important to note 393
that the result from the transgenic potato experiment does not negate the importance of transcript 394
abundance and cell specificity to phloem mobility. They rather indicate that other unidentified 395
regulatory mechanisms, e.g., motif structure, length, interaction with RNA binding proteins or 396
the plasmodesmata between the companion cell and sieve element, etc., in addition to transcript 397
abundance, may also be involved in conferring mobility. 398
Cell origin: apoplastic and symplastic loading species may differ 399
When we compared transcript abundance in the entire leaf versus phloem mobility in the N. 400
benthamiana/tomato heterografts we did not observe any correlation (Fig. 2A), indicating that 401
more abundant transcripts in the leaf do not necessarily lead to higher probability of shoot-to-402
root movement. This lack of correlation is in conflict with the grapevine graft system in which 403
17 of the highly abundant 33 leaf mRNAs were mobile (Yang et al., 2015). This large difference 404
between the two systems led us to hypothesize whether leaf ultrastructure could play a role in 405
this physiological process. N. benthamiana, the scion in our heterograft system, is an apoplastic 406
loading species. The phloem companion cell-sieve element (CC-SE) complex is almost entirely 407
isolated from its surrounding cell types, including the mesophyll, while in grapevine, a woody 408
species and a symplastic loader, the plasmodesmatal connections between the mesophyll cells 409
and the CC-SE complex are very abundant. Therefore, when mRNAs extracted from the entire 410
leaf are used for abundance analysis, the most abundant mRNAs in an apoplastic loader may not 411
have the highest mobility unless they are transcribed in the phloem companion cells. Transcripts 412
generated in the mesophyll cells cannot move to the CC-SE due to the lack of plasmodesmata 413
between them. In contrast, in a symplastic loading species, such as grapevine, mRNAs can 414
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

17
potentially move directly to the translocation stream sieve element from surrounding cells such 415
as mesophyll. To explore the origin of the mobile mRNAs identified from our heterograft system 416
in which the scion, N. benthamiana, is an apoplastic species, we compared the 1,163 mobile 417
mRNAs derived from the three types of N. benthamiana/tomato heterografts (Supplementary 418
Table 12) to the phloem mRNAs identified with either phloem sap collection or the laser capture 419
microdissection (LCM) method (Ivashikina et al., 2003; Doering-Saad et al., 2006; Omid et al., 420
2007; Deeken et al., 2008; Kanehira et al., 2010). A total of 128 mobile mRNAs were found to 421
be detected in previous studies. A hypergeometric test showed that these mobile mRNAs were 422
over-represented in the mRNAs identified from previous studies (p < 0.05; Supplementary Table 423
14). Previous studies have shown that mRNAs can move from cell to cell via the plasmodesmata 424
(Lucas et al., 1995; Roberts and Oparka, 2003). Hence, the abundance of mRNAs in the leaf 425
could be a better indicator of mRNA mobility in symplastic loading species than in apoplastic 426
species. This hypothesis could also be used to explain why a large number of transcripts 427
harboring the TLS motif in the leaf of the N. benthamiana/tomato heterograft did not move to the 428
root. It is highly possible that this motif could confer mobility only when the mRNAs are 429
transcribed in the companion cells. 430
Scion-to-rootstock-to-scion mRNA cycling 431
While the xylem mainly serves to conduct water and minerals from roots to shoots, the phloem 432
serves as a conduit to transport carbohydrates, amino acids, and a number of signaling molecules. 433
Previous studies in different heterograft systems indicate that mRNAs not only move from 434
shoots to roots via the phloem, but also move from roots to shoots (Thieme et al., 2015; Yang et 435
al., 2015). It has been suggested that the root-to-shoot movement of mobile mRNAs can take 436
place either in the phloem against the source to sink bulk flow or via the cell-to-cell-based root-437
to-shoot transport system (Thieme et al., 2015; Ham and Lucas, 2017). The attentions of these 438
prior studies were either on shoot- or root-derived mRNAs. To find out whether root-arriving, 439
shoot-born mRNAs can move back to the shoot, we designed a “split shoot” system involving 440
tripartite grafting. To our surprise, 58 N. benthamiana mRNAs were found in the potato shoot. 441
To fulfill this scion-to-rootstock-to-scion movement of mRNAs, the scion-to-rootstock mobile 442
mRNAs from N. benthamiana need to be unloaded from the phloem to the apoplast/xylem area 443
in tomato and then are loaded to either an adjacent shoot-ward transport phloem or to other root 444
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

18
cells, as discussed above, before the rootstock-to-scion movement takes place. How the scion-to-445
rootstock-transmitted mRNAs are released from the phloem to the apoplast/xylem area and 446
exchanged to other cells is a fascinating question to be addressed in the future. It has been known 447
for a long time that the exchange of carbon containing compounds and minerals between the 448
apoplast/xylem and phloem happens in plants, and membrane transporters play important roles in 449
this process (van Bel, 1990; Zhang and Turgeon, 2009; White, 2012). However, there are no 450
mRNA transporters that have been discovered to efflux mRNAs. An alternative explanation is 451
that the scion-to-rootstock-arriving mRNAs are initially unloaded from the phloem in the stele 452
region via plasmodesmata, as indicated by a recent study in which the shoot-to-root-arriving 453
proteins can be unloaded symplastically (Paultre et al., 2016), and then by an unknown 454
mechanism, the unloaded mRNAs are transferred to cells, e.g., phloem and/or cortical, and move 455
to the shoot. 456
Conclusions 457
Heterografting is a powerful system to identify long-distance transport of mobile mRNAs. 458
However, the large variation in the number and identity of the mobile mRNAs derived from 459
different systems indicates that a more general understanding of the mechanisms of the 460
movement of these transcripts is needed before large-scale functional characterizations on 461
individual mRNAs is pursued. Endogenous physiological difference among plant species may be 462
one of the reasons for the large diversity of identified mobile mRNAs. Different heterograft 463
combinations and the degree of the distortion associated with the involved species may be 464
another reason leading to the large variation of identified mobile transcripts. Identifying the core 465
mobile mRNAs across species and dissecting the physiological functions associated with them 466
are challenging but remain an important task for future research. 467
468
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

19
MATERIALS AND METHODS 469
Plant materials and growth conditions 470
Seedlings of Nicotiana benthamiana, tomato (Solanum lycopersicum cv. Heinz 1706-BG), and 471
potato (Solanum tuberosum L. cv. Désirée) were grown in Propagation mix (Sun Gro 472
Horticulture, Agawam, MA) in a greenhouse with a day/night temperature of 28°C /25°C and a 473
light intensity of approximately 400-600 µmol m-2
s-1
on a 14-h:10-h light regime. Canola 474
(Brassica napus cv. Westar) and Arabidopsis (Arabidopsis thaliana ecotype Columbia) were 475
grown in a growth chamber with a day/night temperature of 26oC/23
oC and a light intensity of 476
250 µmol m-2
s-1
on a 12-h:12-h light regime. 477
Grafting and tissue sampling 478
To produce heterografts between N. benthamiana and tomato, 3-week-old tomato and N. 479
benthamiana seedlings were used as rootstock and scion, respectively. A “V”-shaped wedge of 480
an N. benthamiana stem was inserted into a slit in a tomato stem. Both the scion and the graft 481
joint were kept in a transparent plastic bag and the heterografts were placed beneath a bench with 482
dim light. After one week, the plastic bag was gradually opened over a period of 3 days and the 483
established heterografts were transferred to full-strength hydroponic growth solution and grown 484
in a growth chamber with a day/night temperature of 28°C /25°C and a light intensity of 400-600 485
µmol m-2
s-1
on a 14-h:10-h regime. At this stage, visible flower buds from the N. benthamiana 486
scion were pinched off to avoid contaminating the tomato root tissue with pollen. 487
To overexpress non-mobile mRNAs in the phloem companion cells of potato, two representative 488
Arabidopsis genes, nitrate transporter 1 (AtCHL1) and ammonium transporter 1;2 (AtAMT1;2), 489
were cloned and individually fused behind the promoter of the Arabidopsis sucrose transporter 2 490
(AtSUC2), a phloem companion-cell-specific gene. Primers flanked with restriction sites used to 491
amplify both promoters and genes are listed in Supplementary Table 15. pRI 101-AN was used 492
as the binary vector. Transgenic potato plants were produced by Agrobacterium-mediated 493
transformation following the protocol described in Zhou et al. (Zhou et al., 2017). Grafting 494
transgenic potato onto wild-type potato followed the same procedure described above. These 495
plants were grown in a greenhouse under the aforementioned conditions. 496
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

20
To study whether the accumulation pattern of mobile mRNAs differs during their movement, the 497
shoot tips of 2.5-m-tall (5-month-old) tomato plants were severed and N. benthamiana scions 498
were grafted onto the tops of these tomato plants. The established heterografts were allowed to 499
grow for 6 weeks in a greenhouse in the conditions described above. 500
To study whether scion-to-rootstock mobile mRNAs can move back to the scion after they arrive 501
in the root, tomato seedlings were trained to have two major stems during their first 3 weeks of 502
growth. A potato scion was grafted on one of the two stems to form the first heterograft. After 3 503
weeks, the young sink leaves on the actively growing potato scion were pinched off and a N. 504
benthamiana scion was grafted onto the second stem of the tomato to create a “split-shoot” plant. 505
These tripartite heterografts were grown in the same greenhouse in the conditions described 506
above for another 3 weeks before the mature leaves on the potato scion were harvested. 507
To produce the canola/Arabidopsis heterograft, shoots of 2-week-old canola plants were grafted 508
onto the severed inflorescence stems of 4-week-old Arabidopsis. The procedure was similar to 509
that used for the N. benthamiana/tomato system. The heterografts were allowed to grow for 6 510
weeks in a chamber with a day/night temperature of 26°C /23°C and a light intensity of 250 µmol 511
m-2
s-1
in a 12-h:12-h regime to produce several actively growing mature canola leaves. At 5 512
days prior to harvesting the Arabidopsis roots, the Arabidopsis leaves were covered by aluminum 513
foil to facilitate the source-to-sink bulk flow from the canola leaves to the Arabidopsis roots. 514
RNA isolation and RT-PCR 515
Total RNA was isolated from tissues using the E.Z.N.A. Total RNA Kit I (Omega Bio-tek, 516
Norcross, GA) following the manufacturer's instructions. One microgram of total RNA was 517
treated with RQ1 DNase (Promega, Madison, WI) for 30 min to remove genomic DNA, and then 518
converted into cDNA using iScript Reverse Transcription Supermix (Bio-Rad, Hercules, CA). 519
Gene-specific primers used in RT-PCR are listed in Table S15. Ex Taq DNA Polymerase 520
(Takara Bio USA, Mountain View, CA) was used to amplify the PCR products. The PCR cycle 521
consisted of 95°C for 3 min, followed by 40 cycles of 95°C for 30 s, 60°C for 30 s, and 72°C for 522
30 s. 523
RNA sequencing and RNA-Seq data processing 524
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

21
Strand-specific RNA-Seq libraries were constructed from 4 μg total RNA using the Illumina 525
TruSeq Stranded mRNA Library Prep Kit (Illumina, San Diego, CA) following the 526
manufacturer’s protocol. Three independent biological replicates for either leaf or root from the 527
different heterograft combinations described above were prepared and sequenced on an Illumina 528
NextSeq 500 system at the Genomics Resources Core Facility of Weill Cornell Medical College. 529
Libraries from different species were sequenced on different lanes of NextSeq 500 to avoid cross 530
contaminations between samples. 531
Raw RNA-Seq reads were first processed to trim the adaptor and low-quality sequences using 532
Trimmomatic (Bolger et al., 2014) and trimmed reads shorter than 40 bp were discarded. The 533
remaining high-quality reads were first aligned to a ribosomal RNA (rRNA) database using 534
Bowtie (Langmead et al., 2009), allowing up to three mismatches. These rRNA-mapped reads 535
were excluded from subsequent analyses. The remaining cleaned RNA-Seq reads were mapped 536
to the corresponding genomes using HISAT (Kim et al., 2015). Following alignments, raw 537
counts for each gene model were derived and then normalized using the RPKM (reads per 538
kilobase of exon per million fragments mapped) method (Mortazavi et al., 2008). 539
Graft-transmissible mRNA identification 540
To identify mRNAs transmitted from the N. benthamiana scion to the tomato rootstock, the 541
cleaned reads from the tomato rootstock were first mapped to the tomato genome (Tomato 542
Genome, 2012) using HISAT allowing up to two edit distances. The unmapped reads were 543
compared with the RNA-Seq reads from the non-grafted tomato plants, and those having perfect 544
matches were discarded. The remaining reads were further mapped to the N. benthamiana 545
genome (Bombarely et al., 2012) using HISAT, allowing up to one edit distance. Reads mapped 546
to the N. benthamiana genome were regarded as transmitted. The graft-transmissible mRNAs 547
were identified if the corresponding reads were detected in at least two out of the three biological 548
replicates. Similar procedures were followed to identify mobile mRNAs for the tripartite N. 549
benthamiana/tomato/potato system and the canola/Arabidopsis system. GO terms enriched in the 550
mobile mRNAs were identified using GO::TermFinder (Boyle et al., 2004). 551
Differential accumulation analysis in long-stem N. benthamiana/tomato heterografts 552
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

22
Plant tissues from “high stem” (a region approximately 10-20 cm below the grafting joint), “low 553
stem” (a region approximately 120-130 cm below the grafting joint) and root were collected for 554
RNA-Seq analysis. Identification of the mobile mRNAs followed the same procedure as 555
described above. Raw counts were then fed to edgeR (Robinson et al., 2010) to identify 556
differentially accumulated genes. The Benjamini and Hochberg method (Benjamini and 557
Hochberg, 1995) was used to adjust raw P-values with a confidence threshold of false discovery 558
rate (FDR) <0.01. The abundance of each mRNA in “high stem”, “low stem”, and root was 559
divided by that in the root to obtain relative fold changes along the movement path. 560
TLS structure analysis 561
tRNA-like structures (TLS) for either the entire N. benthamiana leaf transcriptome or all of the 562
identified mobile mRNAs were scanned using TLSfinder in the PlaMoM database (Guan et al., 563
2016) with default parameter settings. 564
Orthology analysis 565
Complete proteome sequences of N. benthamiana were compared to those of Arabidopsis, 566
grapevine, and cucumber using BLASTP with an e-value cutoff of 1e-05
. Orthologous pairs 567
between N. benthamiana and Arabidopsis, grapevine, or cucumber were identified based on the 568
reciprocal best hits of BLASTP. The orthologous pairs that shared the same N. benthamiana 569
genes were identified as those present in all four species. The same procedure was used to 570
identify orthologous pairs between canola and Arabidopsis, grapevine, or cucumber. Mobile 571
mRNAs identified from the different heterograft systems and presented in the four-way 572
orthology were used for the comparisons. 573
Accession Numbers 574
The sequence data were deposited in the NCBI sequence read archive (SRA) under the accession 575
number SRP111187. 576
SUPPLEMENTARY MATERIALS 577
The following materials are available in the online version of this article. 578
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

23
Supplementary Figure 1. Companion cell-specific overexpression of two phloem immobile 579
mRNAs in potato. 580
Supplementary Table 1. N. benthamiana mRNAs detected in the tomato root samples in the N. 581
benthamiana/tomato heterografts 582
Supplementary Table 2. Enrichment of GO slim terms in the 183 mobile mRNAs detected from 583
the N. benthamiana/tomato hetergrafts 584
Supplementary Table 3. Expression of genes in the leaf of N. benthamiana collected from the N. 585
benthamiana/tomato heterografts 586
Supplementary Table 4. Abundance of mobile mRNAs in the scion or rootstock in the N. 587
benthamiana/tomato heterografts 588
Supplementary Table 5. Mobile mRNAs detected from the root samples of tomato in the long-589
stem N. benthamiana/tomato heterografts 590
Supplementary Table 6. Mobile mRNAs detected from the "low stem" region of the tomato 591
rootstock in the long-stem N. benthamiana/tomato heterografts 592
Supplementary Table 7. Mobile mRNAs detected from the "high stem" region of the tomato 593
rootstock in the N. benthamiana/tomato heterografts 594
Supplementary Table 8. Differentially expressed mobile mRNAs detected in the "high-stem", 595
"low-stem", and root regions in the "long-stem" N. benthamiana/tomato heterografts 596
Supplementary Table 9. Mobile mRNAs detected from mature potato leaves in the split-shoot 597
grafting system 598
Supplementary Table 10. GO terms enriched in the shoot-to-root-to-shoot mobile mRNAs 599
Supplementary Table 11. Mobile mRNAs detected in the Arabidopsis root collected from the 600
canola/Arabidopsis heterografts 601
Supplementary Table 12. A combined list of shoot-to-root mobile mRNAs in the different 602
heterograft systems 603
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

24
Supplementary Table 13. Mobile mRNAs identified in the roots from different N. 604
benthamiana/tomato heterografts 605
Supplementary Table 14. Comparison of the 1,163 mobile mRNAs identified from the three N. 606
benthamiana/tomato heterografts with phloem mRNAs identified from previous studies 607
Supplementary Table 15. Primers used in this study 608
609
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

25
ACKNOWLEDGEMENTS 610
We are grateful to the Tomato Genetics Resource Center at the University of California, Davis, 611
for providing the tomato seeds and the Plant Gene Resources of Canada (PGRC) for supplying 612
the Brassica napus, Westar seeds. 613
614
Table 1. Top 10 most highly accumulated mobile mRNAs in the roots of long-stem heterografts 615
mRNA accessions Annotation
Niben101Scf36076g00002.1 Elongation factor 1-alpha 1
Niben101Scf00987g00004.1 Casein kinase I
Niben101Scf02525g03015.1 Casein kinase I
Niben101Scf05201g02004.1 Magnesium transporter NIPA2
Niben101Scf11383g00015.1 40S ribosomal protein S24-2
Niben101Scf05618g04003.1 tRNA (guanine(26)-N(2))-dimethyltransferase
Niben101Scf09409g02014.1 Choline/ethanolaminephosphotransferase 1
Niben101Scf08309g01008.1 Homogentisate 1,2-dioxygenase
Niben101Scf02597g00003.1 30S ribosomal protein S8
Niben101Scf16439g00004.1 30S ribosomal protein S8
616
617
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

26
Figure legends 618
Figure 1. Shoot-to-root mobile mRNAs identified from the N. benthamiana/tomato heterografts 619
grown in a hydroponic condition. A. Representative heterograft established between N. 620
benthamiana and tomato. The white box indicates the graft union (GU). Image to the right shows 621
an enlargement of the region boxed in the left image. Bar: 2cm. B. GO terms enriched in the 183 622
mobile mRNAs. Columns indicate the percentages of genes that were enriched in the GO terms. 623
“All genes” refers to the whole genome background. 624
Figure 2. Correlation analysis of the abundances of leaf mRNAs and their mobility. Expression 625
levels of the graft-transmissible mRNAs in leaf and root tissues were ranked by mean RPKM 626
values derived from heterografts (n=3) grown with full-strength nutrients. A. Correlation of 627
abundances for the 183 mobile mRNAs in the scion and rootstock. Red dashed is the trend line. 628
B. Venn diagram showing the number of mobile mRNAs among the most abundant 100 or 183 629
mRNAs in the scion. Blue color represents the number of the most abundant mRNAs in the N. 630
benthamiana scion. Red color represents the number of the mobile mRNAs in the tomato 631
rootstock. 632
Figure 3. Mobility of non-mobile mRNAs overexpressed in transgenic potato phloem 633
companion cells. A. Representative SUC2prom::AtCHL1 transgenic potato/WT potato 634
heterograft. White box indicates the graft union (GU). Image to the right shows an enlargement 635
of the region boxed in the left image. Bar: 2cm. B. RT-PCR analysis of AtAMT1;2 and AtCHL1 636
in transgenic potato grafts. Both AtAMT1;2 and AtCHL1 are phloem immobile mRNAs. 637
Transgenic potato plants with overexpressed AtAMT1;2 or AtCHL1 was grafted onto WT and the 638
expression of these genes in the rootstock was compared with that in the homografts in which the 639
scion and the rootstock were the same genotype (WT or transgenic). For the transgenic/WT 640
grafts, three shoots excised from the same transgenic potato plant with either overexpressed 641
AtAMT1;2 or AtCHL1 were grafted on the tomato rootstock, respectively. 642
Figure 4. Differentially accumulated mobile mRNAs along the movement path in a “long-stem” 643
heterograft. A. Representative long-stem heterograft. An actively growing N. benthamiana scion 644
grafted on the top of a 2.5-m-tall tomato rootstock. B. Magnification of the dashed box in (A). 645
GU: graft union. C. Venn diagram indicating the number of mobile mRNAs identified at 646
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

27
different locations on the tomato rootstock. D. Heatmap showing the abundance profiles of the 647
39 differentially accumulated mobile mRNAs. Colors indicate different log2 (fold change) 648
values, which are related to the mRNA abundance in the root (=0 in the heat map). Left dashed 649
box indicates the 29 mRNAs decreased in abundance during the transport. Right dashed box 650
indicates the 10 mobile mRNAs over-accumulated in the heterograft root. 651
Figure 5. Scion-to-rootstock-to-scion mRNA cycling revealed by a split-shoot heterograft 652
system. A. Representative “split-shoot” heterograft in which the scion on the right is N. 653
benthamiana, the scion on the left is potato, and the rootstock is tomato. The white boxes 654
indicate the graft union (GU). B. Close-up view of the heterograft union between potato and 655
tomato. C. Close-up view of the heterograft union between N. benthamiana and tomato. Bar: 2 656
cm. 657
Figure 6. Heterograft established between canola and Arabidopsis and comparison of mobile 658
mRNAs identified from different heterograft systems. A. Representative canola/Arabidopsis 659
heterograft grown in a hydroponic solution. White box indicates the graft union (GU). Bar: 2cm. 660
B. Venn diagram of orthologous mobile mRNAs identified in the N. benthamiana/tomato system 661
and other heterograft systems. C. Venn diagram of orthologous mobile mRNAs identified in the 662
canola/Arabidopsis system and other heterograft systems. Mobile mRNAs having orthologs in 663
the proteomes of the other three systems based on reciprocal BLASTP hits were included in the 664
comparison. 665
666
667
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

28
REFERENCES 668
Banerjee AK, Chatterjee M, Yu Y, Suh SG, Miller WA, Hannapel DJ (2006) Dynamics of a 669
mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell 18: 670
3443-3457 671
Bariola PA, MacIntosh GC, Green PJ (1999) Regulation of S-like ribonuclease levels in 672
Arabidopsis. Antisense inhibition of RNS1 or RNS2 elevates anthocyanin accumulation. 673
Plant Physio1 119: 331-42 674
Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful 675
approach to multiple testing. J R Stat Soc Ser B 57: 289–300 676
Bolger AM, Lohse M, Usadel B (2014) Trimmomatic: a flexible trimmer for Illumina sequence 677
data. Bioinformatics 30: 2114-2120 678
Bombarely A, Rosli HG, Vrebalov J, Moffett P, Mueller LA, Martin GB (2012) A draft 679
genome sequence of Nicotiana benthamiana to enhance molecular plant-microbe biology 680
research. Mol Plant Microbe In 25: 1523-1530 681
Boyle EI, Weng S, Gollub J, Jin H, Botstein D, Cherry JM, Sherlock G (2004) GO:: 682
TermFinder—open source software for accessing Gene Ontology information and finding 683
significantly enriched Gene Ontology terms associated with a list of genes. 684
Bioinformatics 20: 3710-3715 685
Calderwood A, Kopriva S, Morris RJ (2016) Transcript Abundance Explains mRNA Mobility 686
Data in Arabidopsis thaliana. Plant Cell 28: 610-615 687
Deeken R, Ache P, Kajahn I, Klinkenberg J, Bringmann G, Hedrich R (2008) Identification 688
of Arabidopsis thaliana phloem RNAs provides a search criterion for phloem-based 689
transcripts hidden in complex datasets of microarray experiments. Plant J 55: 746–759 690
Doering‐Saad C, Newbury HJ, Bale JS, Pritchard J (2002) Use of aphid stylectomy and RT‐691
PCR for the detection of transporter mRNAs in sieve elements. J Exp Bot 53: 631-637 692
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

29
Doering-Saad C, Newbury HJ, Couldridge CE, Bale JS, Pritchard J (2006) A phloem-693
enriched cDNA library from Ricinus: insights into phloem function. J Exp Bot 57: 3183-694
93 695
Guan D, Yan B, Thieme C, Hua J, Zhu H, Boheler KR, Zhao Z, Kragler F, Xia Y, Zhang S 696
(2016) PlaMoM: a comprehensive database compiles plant mobile macromolecules. 697
Nucleic Acids Res : 45, D1021-D1028 698
Ham BK, Lucas WJ (2014) The angiosperm phloem sieve tube system: a role in mediating 699
traits important to modern agriculture. J Exp Bot 65: 1799-1816 700
Ham BK, Lucas WJ (2017) Phloem-Mobile RNAs as Systemic Signaling Agents. Annu Rev 701
Plant Biol 68: 173-195 702
Ivashikina N, Deeken R, Ache P, Kranz E, Pommerrenig B, Sauer N, Hedrich 703
R (2003) Isolation of AtSUC2 promoter–GFP-marked companion cells for patch–clamp 704
studies and expression profiling. Plant J 36: 931–945 705
Kanehira A, YamadaeIwaya T, Tsuwamoto R, Kasai A, Nakazono M, Harada T (2010) 706
Apple phloem cells contain some mRNAs transported over long distances. Tree Genet 707
Genomes 6: 635–642 708
Kim D, Langmead B, Salzberg SL (2015) HISAT: a fast spliced aligner with low memory 709
requirements. Nat Methods 12: 357-360 710
Kim G, LeBlanc ML, Wafula EK, Depamphilis CW, Westwood JH (2014) Genomic-scale 711
exchange of mRNA between a parasitic plant and its hosts. Science 345: 808-811 712
Kim M, Canio W, Kessler S, Sinha N (2001) Developmental changes due to long-distance 713
movement of a homeobox fusion transcript in tomato. Science 293: 287-289 714
King R, Zeevaart J (1974) Enhancement of phloem exudation from cut petioles by chelating 715
agents. Plant Physiol 53: 96-103 716
Langmead B, Trapnell C, Pop M, Salzberg SL (2009) Ultrafast and memory-efficient 717
alignment of short DNA sequences to the human genome. Genome Biol 10: R25 718
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

30
LeBlanc M, Kim G, Patel B, Stromberg V, Westwood J (2013) Quantification of tomato and 719
Arabidopsis mobile RNAs trafficking into the parasitic plant Cuscuta pentagona. New 720
Phytol 200: 1225-1233 721
Lin MK, Lee YJ, Lough TJ, Phinney BS, Lucas WJ (2009) Analysis of the pumpkin phloem 722
proteome provides insights into angiosperm sieve tube function. Mol Cell Proteomics 8: 723
343-356 724
Liu DD, Chao WM, Turgeon R (2012) Transport of sucrose, not hexose, in the phloem. J Exp 725
Bot 63: 4315-4320 726
Liu W, Xu Z-H, Luo D, Xue H-W (2003) Roles of OsCKI1, a rice casein kinase I, in root 727
development and plant hormone sensitivity. Plant J 36: 189-202 728
Lucas WJ, Bouché-Pillon S, Jackson DP, Nguyen L (1995) Selective trafficking of 729
KNOTTED1 homeodomain protein and its mRNA through plasmodesmata. Science 270: 730
1980 731
Ma Y, Miura E, Ham BK, Cheng HW, Lee YJ, Lucas WJ (2010) Pumpkin eIF5A isoforms 732
interact with components of the translational machinery in the cucurbit sieve tube system. 733
Plant J 64: 536-550 734
Morris RJ (2018) On the selectivity, specificity and signalling potential of the long-distance 735
movement of messenger RNA. Curr Opin Plant Biol 43: 1-7 736
Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B (2008) Mapping and quantifying 737
mammalian transcriptomes by RNA-Seq. Nat Methods 5: 621-628 738
Mustroph A, Zanetti ME, Jang CJ, Holtan HE, Repetti PP, Galbraith DW, Girke T, 739
Bailey-Serres J (2009) Profiling translatomes of discrete cell populations resolves 740
altered cellular priorities during hypoxia in Arabidopsis. Proc Natl Acad Sci USA 106: 741
18843-18848 742
Notaguchi M, Higashiyama T, Suzuki T (2015) Identification of mRNAs that move over long 743
distances using an RNA-Seq analysis of Arabidopsis/Nicotiana benthamiana heterografts. 744
Plant Cell Physiol 56: 311-321 745
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

31
Notaguchi M, Wolf S, Lucas WJ (2012) Phloem-mobile Aux/IAA transcripts target to the root 746
tip and modify root architecture. J Integr Plant Biol 54: 760-772 747
Omid A, Keilin T, Glass A, Leshkowitz D, Wolf S (2007) Characterization of phloem-sap 748
transcription profile in melon plants. J Exp Bot 58: 3645-3656 749
Paultre DS, Gustin MP, Molnar A, Oparka KJ (2016) Lost in transit: long-distance 750
trafficking and phloem unloading of protein signals in Arabidopsis homografts. Plant Cell 751
28, 2016-2025 752
Roberts A, Oparka K (2003) Plasmodesmata and the control of symplastic transport. Plant, Cell 753
Environ 26: 103-124 754
Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bioconductor package for 755
differential expression analysis of digital gene expression data. Bioinformatics 26: 139-756
140 757
Sakamoto M, Sanada Y, Tagiri A, Murakami T, Ohashi Y, Matsuoka M (1991) Structure 758
and Characterization of a Gene for Light-Harvest Chl a/b Binding Protein from Rice. 759
Plant Cell Physiol 32: 385-393 760
Sasaki T, Chino M, Hayashi H, Fujiwara T (1998) Detection of Several mRNA Species in 761
Rice Phloem Sap. Plant Cell Physiol 39: 895-897 762
Song G, Honda H, Yamaguchi K (2007) Expression of a Rice Chlorophyll a/b Binding Protein 763
Promoter in Sweetpotato. J Amer Soc Hort Sci 132: 551-556 764
Spiegelman Z, Golan G, Wolf S (2013) Don't kill the messenger: Long-distance trafficking of 765
mRNA molecules. Plant Sci 213: 1-8 766
Stadler R, Wright KM, Lauterbach C, Amon G, Gahrtz M, Feuerstein A, Oparka KJ, 767
Sauer N (2005) Expression of GFP-fusions in Arabidopsis companion cells reveals non-768
specific protein trafficking into sieve elements and identifies a novel post-phloem domain 769
in roots. Plant J 41: 319-331 770
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

32
Tada Y, Sakamoto M, Matsuoka M, Fujimura T (1991) Expression of a monocot LHCP 771
promoter in transgenic rice. EMBO J 10: 1803–1808 772
Thieme CJ, Rojas-Triana M, Stecyk E, Schudoma C, Zhang W, Yang L, Minambres M, 773
Walther D, Schulze WX, Paz-Ares J, Scheible WR, Kragler F (2015) Endogenous 774
Arabidopsis messenger RNAs transported to distant tissues. Nat Plants 1: 15025 775
Tomato Genome C (2012) The tomato genome sequence provides insights into fleshy fruit 776
evolution. Nature 485: 635-641 777
Turgeon R, Wolf S (2009) Phloem transport: cellular pathways and molecular trafficking. Annu 778
Rev Plant Biol 60: 207-221 779
Turnbull CG, Lopez‐Cobollo RM (2013) Heavy traffic in the fast lane: long‐distance 780
signalling by macromolecules. New Phytol 198: 33-51 781
van Bel A (1990) Xylem-Phloem Exchange Via the Rays: The Undervalued Route of 782
Transport. J Exp Bot 41: 631–644 783
White PJ (2012) Long-distance transport in the xylem and phloem. In Marschner's Mineral 784
Nutrition of Higher Plants (Third Edition). Academic Press, pp 49-70 785
Yang Y, Mao L, Jittayasothorn Y, Kang Y, Jiao C, Fei Z, Zhong GY (2015) Messenger RNA 786
exchange between scions and rootstocks in grafted grapevines. BMC Plant Biol 15: 251 787
Zhang C, Turgeon R (2009) Downregulating the sucrose transporter VpSUT1 in Verbascum 788
phoeniceum does not inhibit phloem loading. Proc Natl Acad Sci USA 106: 18849-18854 789
Zhang C, Han L, Slewinski TL, Sun J, Zhang J, Wang Z-Y, Turgeon R (2014) Symplastic 790
phloem loading in poplar. Plant Physiol 166: 306-313 791
Zhang C, Yu X, Ayre BG, Turgeon R (2012) The origin and composition of cucurbit “phloem” 792
exudate. Plant Physiol 158: 1873-1882 793
Zhang W, Thieme CJ, Kollwig G, Apelt F, Yang L, Winter N, Andresen N, Walther D, 794
Kragler F (2016) tRNA-Related Sequences Trigger Systemic mRNA Transport in Plants. 795
Plant Cell 28: 1237-1249 796
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

33
Zhang Z, Zheng Y, Ham BK, Chen J, Yoshida A, Kochian LV, Fei Z, Lucas WJ (2016) 797
Vascular-mediated signalling involved in early phosphate stress response in plants. Nat 798
Plants 2: 16033 799
Zhou X, Zha M, Huang J, Li L, Imran M, Zhang C (2017) StMYB44 negatively regulates 800
phosphate transport by suppressing expression of PHOSPHATE1 in potato. J Exp Bot 68: 801
1265-1281 802
803
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1 www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
Figure 1. Shoot-to-root mobile mRNAs identified from the N. benthamiana/tomato heterografts
grown in a hydroponic condition. A. Representative heterograft established between N.
benthamiana and tomato. The white box indicates the graft union (GU). Image to the right shows
an enlargement of the region boxed in the left image. Bar: 2cm. B. GO terms enriched in the 183
mobile mRNAs. Columns indicate the percentages of genes that were enriched in the GO terms.
“All genes” refers to the whole genome background.
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 2. Correlation analysis of the abundances of leaf mRNAs and their mobility. Expression
levels of the graft-transmissible mRNAs in leaf and root tissues were ranked by mean RPKM
values derived from heterografts (n=3) grown with full-strength nutrients. A. Correlation of
abundances for the 183 mobile mRNAs in the scion and rootstock. Red dashed is the trend line.
B. Venn diagram showing the number of mobile mRNAs among the most abundant 100 or 183
mRNAs in the scion. Blue color represents the number of the most abundant mRNAs in the N.
benthamiana scion. Red color represents the number of the mobile mRNAs in the tomato
rootstock.
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 3. Mobility of non-mobile mRNAs overexpressed in transgenic potato phloem
companion cells. A. Representative SUC2prom::AtCHL1 transgenic potato/WT potato
heterograft. White box indicates the graft union (GU). Image to the right shows an enlargement
of the region boxed in the left image. Bar: 2cm. B. RT-PCR analysis of AtAMT1;2 and AtCHL1
in transgenic potato grafts. Both AtAMT1;2 and AtCHL1 are phloem immobile mRNAs.
Transgenic potato plants with overexpressed AtAMT1;2 or AtCHL1 was grafted onto WT and the
expression of these genes in the rootstock was compared with that in the homografts in which the
scion and the rootstock were the same genotype (WT or transgenic). For the transgenic/WT
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
grafts, three shoots excised from the same transgenic potato plant with either overexpressed
AtAMT1;2 or AtCHL1 were grafted on the tomato rootstock, respectively.
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
1
Figure 4. Differentially accumulated mobile mRNAs along the movement path in a “long-stem”
heterograft. A. Representative long-stem heterograft. An actively growing N. benthamiana scion
grafted on the top of a 2.5-m-tall tomato rootstock. B. Magnification of the dashed box in (A).
GU: graft union. C. Venn diagram indicating the number of mobile mRNAs identified at
different locations on the tomato rootstock. D. Heatmap showing the abundance profiles of the
39 differentially accumulated mobile mRNAs. Colors indicate different log2 (fold change)
values, which are related to the mRNA abundance in the root (=0 in the heat map). Left dashed
box indicates the 29 mRNAs decreased in abundance during the transport. Right dashed box
indicates the 10 mobile mRNAs over-accumulated in the heterograft root.
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 5. Scion-to-rootstock-to-scion mRNA cycling revealed by a split-shoot heterograft
system. A. Representative “split-shoot” heterograft in which the scion on the right is N.
benthamiana, the scion on the left is potato, and the rootstock is tomato. The white boxes
indicate the graft union (GU). B. Close-up view of the heterograft union between potato and
tomato. C. Close-up view of the heterograft union between N. benthamiana and tomato. Bar: 2
cm.
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 6. Heterograft established between canola and Arabidopsis and comparison of mobile
mRNAs identified from different heterograft systems. A. Representative canola/Arabidopsis
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
heterograft grown in a hydroponic solution. White box indicates the graft union (GU). Bar: 2cm.
B. Venn diagram of orthologous mobile mRNAs identified in the N. benthamiana/tomato system
and other heterograft systems. C. Venn diagram of orthologous mobile mRNAs identified in the
canola/Arabidopsis system and other heterograft systems. Mobile mRNAs having orthologs in
the proteomes of the other three systems based on reciprocal BLASTP hits were included in the
comparison.
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Parsed CitationsBanerjee AK, Chatterjee M, Yu Y, Suh SG, Miller WA, Hannapel DJ (2006) Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell 18: 3443-3457
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bariola PA, MacIntosh GC, Green PJ (1999) Regulation of S-like ribonuclease levels in Arabidopsis. Antisense inhibition of RNS1 orRNS2 elevates anthocyanin accumulation. Plant Physio1 119: 331-42
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat SocSer B 57: 289–300
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bolger AM, Lohse M, Usadel B (2014) Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30: 2114-2120Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bombarely A, Rosli HG, Vrebalov J, Moffett P, Mueller LA, Martin GB (2012) A draft genome sequence of Nicotiana benthamiana toenhance molecular plant-microbe biology research. Mol Plant Microbe In 25: 1523-1530
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Boyle EI, Weng S, Gollub J, Jin H, Botstein D, Cherry JM, Sherlock G (2004) GO:: TermFinder—open source software for accessingGene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 20:3710-3715
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Calderwood A, Kopriva S, Morris RJ (2016) Transcript Abundance Explains mRNA Mobility Data in Arabidopsis thaliana. Plant Cell 28:610-615
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Deeken R, Ache P, Kajahn I, Klinkenberg J, Bringmann G, Hedrich R (2008) Identification of Arabidopsis thaliana phloem RNAsprovides a search criterion for phloem-based transcripts hidden in complex datasets of microarray experiments. Plant J 55: 746–759
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Doering‐Saad C, Newbury HJ, Bale JS, Pritchard J (2002) Use of aphid stylectomy and RT‐PCR for the detection of transporter mRNAsin sieve elements. J Exp Bot 53: 631-637
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Doering-Saad C, Newbury HJ, Couldridge CE, Bale JS, Pritchard J (2006) A phloem-enriched cDNA library from Ricinus: insights intophloem function. J Exp Bot 57: 3183-93
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Guan D, Yan B, Thieme C, Hua J, Zhu H, Boheler KR, Zhao Z, Kragler F, Xia Y, Zhang S (2016) PlaMoM: a comprehensive databasecompiles plant mobile macromolecules. Nucleic Acids Res : 45, D1021-D1028
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ham BK, Lucas WJ (2014) The angiosperm phloem sieve tube system: a role in mediating traits important to modern agriculture. J ExpBot 65: 1799-1816
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Ham BK, Lucas WJ (2017) Phloem-Mobile RNAs as Systemic Signaling Agents. Annu Rev Plant Biol 68: 173-195Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ivashikina N, Deeken R, Ache P, Kranz E, Pommerrenig B, Sauer N, Hedrich R (2003) Isolation of AtSUC2 promoter–GFP-markedcompanion cells for patch–clamp studies and expression profiling. Plant J 36: 931–945
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kanehira A, YamadaeIwaya T, Tsuwamoto R, Kasai A, Nakazono M, Harada T (2010) Apple phloem cells contain some mRNAstransported over long distances. Tree Genet Genomes 6: 635–642
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kim D, Langmead B, Salzberg SL (2015) HISAT: a fast spliced aligner with low memory requirements. Nat Methods 12: 357-360Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kim G, LeBlanc ML, Wafula EK, Depamphilis CW, Westwood JH (2014) Genomic-scale exchange of mRNA between a parasitic plant andits hosts. Science 345: 808-811
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kim M, Canio W, Kessler S, Sinha N (2001) Developmental changes due to long-distance movement of a homeobox fusion transcript intomato. Science 293: 287-289
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
King R, Zeevaart J (1974) Enhancement of phloem exudation from cut petioles by chelating agents. Plant Physiol 53: 96-103Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Langmead B, Trapnell C, Pop M, Salzberg SL (2009) Ultrafast and memory-efficient alignment of short DNA sequences to the humangenome. Genome Biol 10: R25
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
LeBlanc M, Kim G, Patel B, Stromberg V, Westwood J (2013) Quantification of tomato and Arabidopsis mobile RNAs trafficking into theparasitic plant Cuscuta pentagona. New Phytol 200: 1225-1233
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lin MK, Lee YJ, Lough TJ, Phinney BS, Lucas WJ (2009) Analysis of the pumpkin phloem proteome provides insights into angiospermsieve tube function. Mol Cell Proteomics 8: 343-356
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu DD, Chao WM, Turgeon R (2012) Transport of sucrose, not hexose, in the phloem. J Exp Bot 63: 4315-4320Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu W, Xu Z-H, Luo D, Xue H-W (2003) Roles of OsCKI1, a rice casein kinase I, in root development and plant hormone sensitivity. PlantJ 36: 189-202
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lucas WJ, Bouché-Pillon S, Jackson DP, Nguyen L (1995) Selective trafficking of KNOTTED1 homeodomain protein and its mRNAthrough plasmodesmata. Science 270: 1980
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ma Y, Miura E, Ham BK, Cheng HW, Lee YJ, Lucas WJ (2010) Pumpkin eIF5A isoforms interact with components of the translationalmachinery in the cucurbit sieve tube system. Plant J 64: 536-550 www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Morris RJ (2018) On the selectivity, specificity and signalling potential of the long-distance movement of messenger RNA. Curr OpinPlant Biol 43: 1-7
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B (2008) Mapping and quantifying mammalian transcriptomes by RNA-Seq. NatMethods 5: 621-628
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mustroph A, Zanetti ME, Jang CJ, Holtan HE, Repetti PP, Galbraith DW, Girke T, Bailey-Serres J (2009) Profiling translatomes ofdiscrete cell populations resolves altered cellular priorities during hypoxia in Arabidopsis. Proc Natl Acad Sci USA 106: 18843-18848
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Notaguchi M, Higashiyama T, Suzuki T (2015) Identification of mRNAs that move over long distances using an RNA-Seq analysis ofArabidopsis/Nicotiana benthamiana heterografts. Plant Cell Physiol 56: 311-321
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Notaguchi M, Wolf S, Lucas WJ (2012) Phloem-mobile Aux/IAA transcripts target to the root tip and modify root architecture. J IntegrPlant Biol 54: 760-772
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Omid A, Keilin T, Glass A, Leshkowitz D, Wolf S (2007) Characterization of phloem-sap transcription profile in melon plants. J Exp Bot58: 3645-3656
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Paultre DS, Gustin MP, Molnar A, Oparka KJ (2016) Lost in transit: long-distance trafficking and phloem unloading of protein signals inArabidopsis homografts. Plant Cell 28, 2016-2025
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Roberts A, Oparka K (2003) Plasmodesmata and the control of symplastic transport. Plant, Cell Environ 26: 103-124Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bioconductor package for differential expression analysis of digital geneexpression data. Bioinformatics 26: 139-140
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sakamoto M, Sanada Y, Tagiri A, Murakami T, Ohashi Y, Matsuoka M (1991) Structure and Characterization of a Gene for Light-HarvestChl a/b Binding Protein from Rice. Plant Cell Physiol 32: 385-393
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sasaki T, Chino M, Hayashi H, Fujiwara T (1998) Detection of Several mRNA Species in Rice Phloem Sap. Plant Cell Physiol 39: 895-897Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Song G, Honda H, Yamaguchi K (2007) Expression of a Rice Chlorophyll a/b Binding Protein Promoter in Sweetpotato. J Amer Soc HortSci 132: 551-556
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Spiegelman Z, Golan G, Wolf S (2013) Don't kill the messenger: Long-distance trafficking of mRNA molecules. Plant Sci 213: 1-8Pubmed: Author and Title www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

CrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stadler R, Wright KM, Lauterbach C, Amon G, Gahrtz M, Feuerstein A, Oparka KJ, Sauer N (2005) Expression of GFP-fusions inArabidopsis companion cells reveals non-specific protein trafficking into sieve elements and identifies a novel post-phloem domain inroots. Plant J 41: 319-331
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tada Y, Sakamoto M, Matsuoka M, Fujimura T (1991) Expression of a monocot LHCP promoter in transgenic rice. EMBO J 10: 1803–1808
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Thieme CJ, Rojas-Triana M, Stecyk E, Schudoma C, Zhang W, Yang L, Minambres M, Walther D, Schulze WX, Paz-Ares J, Scheible WR,Kragler F (2015) Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat Plants 1: 15025
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tomato Genome C (2012) The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485: 635-641Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Turgeon R, Wolf S (2009) Phloem transport: cellular pathways and molecular trafficking. Annu Rev Plant Biol 60: 207-221Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Turnbull CG, Lopez‐Cobollo RM (2013) Heavy traffic in the fast lane: long‐distance signalling by macromolecules. New Phytol 198: 33-51
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
van Bel A (1990) Xylem-Phloem Exchange Via the Rays: The Undervalued Route of Transport. J Exp Bot 41: 631–644Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
White PJ (2012) Long-distance transport in the xylem and phloem. In Marschner's Mineral Nutrition of Higher Plants (Third Edition).Academic Press, pp 49-70
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yang Y, Mao L, Jittayasothorn Y, Kang Y, Jiao C, Fei Z, Zhong GY (2015) Messenger RNA exchange between scions and rootstocks ingrafted grapevines. BMC Plant Biol 15: 251
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang C, Turgeon R (2009) Downregulating the sucrose transporter VpSUT1 in Verbascum phoeniceum does not inhibit phloemloading. Proc Natl Acad Sci USA 106: 18849-18854
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang C, Han L, Slewinski TL, Sun J, Zhang J, Wang Z-Y, Turgeon R (2014) Symplastic phloem loading in poplar. Plant Physiol 166: 306-313
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang C, Yu X, Ayre BG, Turgeon R (2012) The origin and composition of cucurbit “phloem” exudate. Plant Physiol 158: 1873-1882Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang W, Thieme CJ, Kollwig G, Apelt F, Yang L, Winter N, Andresen N, Walther D, Kragler F (2016) tRNA-Related Sequences TriggerSystemic mRNA Transport in Plants. Plant Cell 28: 1237-1249
Pubmed: Author and TitleCrossRef: Author and Title www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Zhang Z, Zheng Y, Ham BK, Chen J, Yoshida A, Kochian LV, Fei Z, Lucas WJ (2016) Vascular-mediated signalling involved in earlyphosphate stress response in plants. Nat Plants 2: 16033
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhou X, Zha M, Huang J, Li L, Imran M, Zhang C (2017) StMYB44 negatively regulates phosphate transport by suppressing expressionof PHOSPHATE1 in potato. J Exp Bot 68: 1265-1281
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon July 29, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
















![Paraquat Modulates Alternative Pre-mRNA Splicing by ...muehlemann.dcb.unibe.ch/publications/Vivarelli.pdf · alternative pre-mRNA splicing (AS) [2]. Pre-mRNA splicing is a crucial](https://static.fdocuments.us/doc/165x107/606212ed67e7345b4269ee34/paraquat-modulates-alternative-pre-mrna-splicing-by-alternative-pre-mrna-splicing.jpg)

