Mec hanistic Diver sity of Cytokine Receptor … · m a in s o f th e re c e p to rs .T h e k in a...
17
www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 1 Circulating cytokines bind to specific receptors on the cell outer surface to evoke responses inside the cell. Binding of cytokines alters the association between re- ceptor molecules that often cross the membrane only once in a single alpha-helical segment. As a conse- quence, association of protein domains on the inside of the membrane are also altered. Increasing evidence suggests that an initial “off-state” of associated recep- tors is perturbed, and brought to an activated state that leads to intracellular signaling and eventually effects a change in DNA transcription. The initial detection event that transduces the change in receptor association is sensitive to both proximity and orientation of the recep- tors, and probably also to the time that the activated state or receptor association is maintained. Ultimately, a cascade of phosphorylation events is triggered. The ini- tial kinases are sometimes part of the intracellular do- mains of the receptors. The kinases can also be sepa- rate proteins that may be pre-associated with intracellu- lar domains of the receptors, or can be recruited after the intracellular association of the activated receptors. We focus here on each of the cases for which structures of the activated cytokine-receptor complexes are known, in a search for underlying mechanisms.The vari- ations in modes of association, stoichiometries of re- ceptors and cytokines, and orientations before and after activation of these receptors are almost as great as the number of complexes themselves.The principles uncov- ered nevertheless illustrate the basis for high specificity and fidelity in cytokine-mediated signaling. The membranes of all cells contain receptors that harmonize external and internal environments. Reversible associations of proteins with consequent changes in structure are dominant themes in signal transduction pathways both inside and outside the cell. This Review briefly introduces some of the general principles of receptor-mediated signaling and then focuses on one of the most highly characterized classes of receptors, the cytokine-receptor superfamily. The structures of the cytokine receptors show in atomic detail how they are activated and how they utilize a diversity of protein folds and oligomeric struc- tures to transmit signals. Fundamentals of Receptor Structure Most receptors are activated by binding a ligand on one side of the membrane; they then initiate a response on the other side. Most ligands do not pass through the membrane, but associate with extracellular binding sites within a single receptor molecule, or through bridging interactions between multiple re- ceptors. These ligands or effectors may be as small as a photon, in the case of photoreceptors, or as large as globular proteins, such as the cytokines. Changes in the relative positions of transmembrane seg- ments and their associated protein domains bring about signal transduction across the membrane. This can occur within a sin- gle multi-spanning transmembrane protein, in a mechanism that we describe as “vertical signaling,” or by changing lateral asso- ciations of receptors, which we term “horizontal” signaling (Fig. 1). These two signaling paradigms have different etiolo- gies, structures, and functions (Table 1). Vertical Versus Horizontal Signaling Vertical signaling receptors are found in the membranes of all organisms from bacteria through higher eukaryotes. Here, the Mechanistic Diversity of Cytokine Receptor Signaling Across Cell Membranes Robert M. Stroud 1 and James A. Wells 2 (Published 4 May 2004) R EVIEW 1 Department of Biochemistry and Biophysics, UCSF Genentech Hall, 600 16th Street, University of California, San Francisco, CA 94143–2240, USA. 2 Sunesis Pharmaceuticals, 3696 Haven Avenue, Suite C, Redwood City, CA 94063, USA. Fig. 1. The characteristics of horizontal and vertical receptor sig- naling mechanisms. Vertical receptor signaling (right) takes place within a single pre-assembled protein or oligomer of protein sub- units that constitute a transmembrane receptor. Signaling re- quires that a conformational change be relayed from outside to inside the cell by means of the trans-bilayer elements. Horizontal receptor signaling ( left ) depends on diffusion of receptor molecules within the membrane plane. Increasing evidence fa- vors the idea of receptor associations existing in an “off-state” be- fore activation (far left). The ligand is in yellow. The bilayer is in- dicated as spheres that represent phosphatidyl head groups. Lig- and binding to one receptor causes dissociation of inactive com- plexes and induces further association that involves at least one other receptor. The oligomeric state and orientation is imposed on the intracellular components of the receptors (below the mem- brane) where associations of kinases lead to their activation.
Transcript of Mec hanistic Diver sity of Cytokine Receptor … · m a in s o f th e re c e p to rs .T h e k in a...

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 1
Circulating cytokines bind to specific receptors on thecell outer surface to evoke responses inside the cell.Binding of cytokines alters the association between re-ceptor molecules that often cross the membrane onlyonce in a single alpha-helical segment. As a conse-quence, association of protein domains on the inside ofthe membrane are also altered. Increasing evidencesuggests that an initial “off-state” of associated recep-tors is perturbed, and brought to an activated state thatleads to intracellular signaling and eventually effects achange in DNA transcription. The initial detection eventthat transduces the change in receptor association issensitive to both proximity and orientation of the recep-tors, and probably also to the time that the activatedstate or receptor association is maintained. Ultimately, acascade of phosphorylation events is triggered. The ini-tial kinases are sometimes part of the intracellular do-mains of the receptors. The kinases can also be sepa-rate proteins that may be pre-associated with intracellu-lar domains of the receptors, or can be recruited afterthe intracellular association of the activated receptors.We focus here on each of the cases for which structuresof the activated cytokine-receptor complexes areknown, in a search for underlying mechanisms.The vari-ations in modes of association, stoichiometries of re-ceptors and cytokines, and orientations before and afteractivation of these receptors are almost as great as thenumber of complexes themselves.The principles uncov-ered nevertheless illustrate the basis for high specificityand fidelity in cytokine-mediated signaling.
The membranes of all cells contain receptors that harmonizeexternal and internal environments. Reversible associations ofproteins with consequent changes in structure are dominantthemes in signal transduction pathways both inside and outsidethe cell. This Review briefly introduces some of the generalprinciples of receptor-mediated signaling and then focuses onone of the most highly characterized classes of receptors, thecytokine-receptor superfamily. The structures of the cytokinereceptors show in atomic detail how they are activated and howthey utilize a diversity of protein folds and oligomeric struc-tures to transmit signals.
Fundamentals of Receptor Structure Most receptors are activated by binding a ligand on one side ofthe membrane; they then initiate a response on the other side.
Most ligands do not pass through the membrane, but associatewith extracellular binding sites within a single receptormolecule, or through bridging interactions between multiple re-ceptors. These ligands or effectors may be as small as a photon,in the case of photoreceptors, or as large as globular proteins,such as the cytokines.
Changes in the relative positions of transmembrane seg-ments and their associated protein domains bring about signaltransduction across the membrane. This can occur within a sin-
gle multi-spanning transmembrane protein, in a mechanism thatwe describe as “vertical signaling,” or by changing lateral asso-ciations of receptors, which we term “horizontal” signaling(Fig. 1). These two signaling paradigms have different etiolo-gies, structures, and functions (Table 1).
Vertical Versus Horizontal Signaling Vertical signaling receptors are found in the membranes of allorganisms from bacteria through higher eukaryotes. Here, the
Mechanistic Diversity of Cytokine ReceptorSignaling Across Cell Membranes
Robert M. Stroud1 and James A. Wells2
(Published 4 May 2004)
R E V I E W
1Department of Biochemistry and Biophysics, UCSF Genentech Hall,600 16th Street, University of California, San Francisco, CA94143–2240, USA. 2Sunesis Pharmaceuticals, 3696 Haven Avenue,Suite C, Redwood City, CA 94063, USA.
Fig. 1. The characteristics of horizontal and vertical receptor sig-naling mechanisms. Vertical receptor signaling (right) takes placewithin a single pre-assembled protein or oligomer of protein sub-units that constitute a transmembrane receptor. Signaling re-quires that a conformational change be relayed from outside toinside the cell by means of the trans-bilayer elements. Horizontalreceptor signaling (left) depends on diffusion of receptormolecules within the membrane plane. Increasing evidence fa-vors the idea of receptor associations existing in an “off-state” be-fore activation (far left). The ligand is in yellow. The bilayer is in-dicated as spheres that represent phosphatidyl head groups. Lig-and binding to one receptor causes dissociation of inactive com-plexes and induces further association that involves at least oneother receptor. The oligomeric state and orientation is imposed onthe intracellular components of the receptors (below the mem-brane) where associations of kinases lead to their activation.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 2
initial ligand binding event produces changes in the receptorconformation vertically through the membrane. These receptorsoften respond to small nonprotein ligands that cause a transientand reversible change in membrane potential or in cellularmetabolism. In the case of ligand or voltage-gated ion-conduct-ing receptors, the response-mediated gating controls are alsofound within the transmembrane region. The preassembled andgenerally unimolecular nature of these receptors allow them torespond rapidly, as is the case for neuronal signaling.
The vertical receptors include the seven-transmembrane receptorsuperfamily, light-sensing receptors, ion channels, and nutrient-sensing receptors, among others. These receptors undergo an in-tramolecular structural change in response to ligand binding (Fig. 1)(1, 2). The structurally best characterized of seven-transmembranemembrane proteins are rhodopsin (3, 4) and bacteriorhodopsin(which is structurally homologous to rhodopsin). Changes in re-sponse to activation by light have been detected in the cytoplasmicregions of rhodopsin (5-7). Electron crystallography (8-10) and x-ray crystallography (11) describe structural changes that open ashort channel on the cytoplasmic surface of bacteriorhodopsin in re-sponse to activation by light; this process may be prophetic of otherchanges in the heterotrimeric guanine nucleotide–binding protein(G protein)–coupled receptors (GPCRs). Vertical receptors also in-clude the chemotactic bacterial receptors for aspartate that are pre-assembled as dimeric complexes before activation. Ligand bindinginduces a change that is detected across the membrane (12).
Horizontal receptors are found principally in multicellularorganisms. They are generally activated by initial binding of amono- or multimeric protein ligand; this binding inducesoligomerization, or an ordered association of receptors on thecell surface. These receptors generally control longer-term or ir-reversible changes in cells, such as transcription, translation,replication, or apoptosis.
The three-dimensional structures have been determined foralmost a dozen ligands in complex with the extracellular por-tions of their horizontal class receptors. The vast majority ofthese involve cytokine-receptor complexes, which can be divid-ed into four major groups (I-IV) on the basis of the preponder-ance of α-helical, β sheet, mixed α/β, or mosaic substructureswithin the cytokine component of the complex (13). Receptoroligomerization is a common theme in horizontal signaling, yetthere is great diversity in the binding sites, folds, and stoi-chiometries of these signaling complexes (Fig. 2).
The distinction between horizontal and vertical receptors hasbeen blurred by recent discoveries that a number of single-passreceptors exist in inactive but preassociated states. In some cas-es, these are poised to bind cytokine, as for members of the tu-mor necrosis factor (TNF) receptor (TNFR) class (14). In othercases, such as the erythropoietin (EPO) receptor (EPOR) (15,16), the receptors are associated in an “off-state” that requiresthem to dissociate before a productive complex can be made.
Conversely, there are examples of receptors in the vertical classof seven-transmembrane receptors (1), such as the monocytechemoattractant protein MCP-1 (17), that appear to form activemultimers during the signaling process.
Mechanisms of Horizontal Signaling ComplexesAs horizontal receptors are drawn together on one side of themembrane, domains on the other side are oriented to initiate sig-naling events. In many cases, the intracellular domains of recep-tors are preassociated with protein kinases, either noncovalently,as is the case with EPORs (18), or by gene fusion, as in the caseof epidermal growth factor (EGF)–class of receptors [reviewedin (19)]. In other cases, the intracellular domains associate with
R E V I E W
γIFN
Fig. 2. The oligomeric forms of cytokine signaling complexesshow a rich variety of multimeric horizontal assemblies that aredetected and translated into pathways such as kinase cascadesinside the cell.
Horizontal receptors Vertical receptors
Typical ligand Protein Small molecule
Signaling induced by Change in oligomeric state Structural change in pre-associated receptor;
no change in quaternary state
Response time Slow Fast
Typical cellular response Change in transcription, Reversible metabolic change
translation, replication or apoptosis
Evolutionary origin Eukaryotes (recent) All species (old)
Example Cytokine receptors G protein–coupled, ligand-gated ion channels
Table 1. Comparison of “vertical” and “horizontal” signaling paradigms.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 3
kinases after they bind hormone, as is the case for human growthhormone (hGH) receptors (hGHRs) (20). Once the receptors areappropriately oriented, the kinases act intermolecularly totransphosphorylate each other. These in turn act as docking sitesfor binding and activating other signaling factors.
Cytokine-receptor complexes include those in which twoidentical receptor molecules are dimerized by binding to twodifferent sites on a single cytokine to produce a 2:1 complex, asseen for hGH (21) and EPO (22). Some cytokines form com-plexes in which two cytokine molecules bind two identical re-ceptors such as in the gp130:interleukin-6 (IL-6) complex, orthe granulocyte colony-stimulating factor and its receptor (G-CSF:G-CSFR) complex (23) and the fibroblast growth factorand its receptor (FGF:FGFR) complexes (24). Still other cy-tokines bind two different receptors simultaneously, as is seenwith IL-4 and interferon-gamma (IFN-γ). Still higher ordercomplexes are seen for the TNFR class.
Group 1, Four-α-Helix CytokineshGH and EPO receptors. HGH was the first cytokine-receptorcomplex for which structure and mechanism were elucidated.HGH is a four-α-helix bundle protein with up-up-down-downtopology. The cytokine is characteristic of the long-chain sub-family of the group I cytokines. The extracellular portion of thereceptor contains two domains that each have a seven-strandedfibronectin III-like topology [reviewed in (25)]. The domainsare termed D1 and D2 (typically, domains are numbered from Nto C terminus in direction, and the N terminus is generally far-thest from the membrane). hGH binds the first receptor througha higher-affinity site (site 1 on hGH), and binds a second identi-cal receptor through a lower-affinity site (site 2 on hGH) (21)(Fig. 3). The cytokine-binding sites are formed at the outside ofthe elbow bend formed between D1 and D2 domains, and in-volve six loops (L1-L6) between β strands from each of the do-mains, L1 to L3 from D1, and L4 to L6 from D2. The secondreceptor is rotated by 157° about the normal to the membraneplane relative to the first, as it binds to the opposite side of thecytokine.
Mutational and biophysical experiments show that thehGHRs associate sequentially with hGH, binding hGH first atsite 1 and then at site 2 on the hormone (26). This second step inthe reaction, which occurs in the plane of the membrane, is sub-sequently stabilized by receptor-receptor contacts, as well as bythe hormone-receptor interactions at site 2 [reviewed in (27)].
Alanine-scanning mutagenesis on both sides of the interfacewith site 1 show that binding affinity is dominated by a smallgroup of mostly hydrophobic side-chain contacts near the centerof the contact interface. In the case of the hGHR, these sidechains are centered around Trp104 and Trp169 (28, 29). Such “hotspots” seem to be present at many protein-protein interfaces, andthey can be used to bind multiple protein partners. Although inmany cases hot spots are hydrophobic, there are some that in-clude polar and charged residues as well. It is remarkable that thereceptor uses virtually the same set of contact residues to interactwith two very different contact surfaces on hGH (site 1 and site2). Moreover, the receptor-receptor contacts are not homotypic,but involve two different sets of residues from each receptorstem. The receptor achieves this by slight domain movements andlarger shifts in the surface side chains involved in binding.
Not only does the hGHR adapt to two different surfaces onhGH, but hGH can interact with multiple receptors. Complexes
of hGH with the prolactin (PRL) or placental lactogen receptorconform to the same paradigm (30). Virtually the same set ofcontact residues in site 1 of hGH mediate interaction with eitherto the hGHR or PRL receptor. Interestingly, hGH requires Zn2+
to bind the PRLR, but not to bind the hGHR. Although the con-tact residues are the same for binding either hGHR or PRLR,the residues that constitute the hot spot for binding on the re-ceptor differ (28). This functional and structural plasticity al-lows cytokines to cross-talk to multiple receptors.
EPO-EPOR complexes. The EPOR complexes provide newinsights into the structural requirements for turning signaling onand off. EPO is a four-α-helix group I cytokine with the samecharacteristic up-up-down-down topology seen in hGH. EPORs
are composed of a short N-terminal α-helix, two seven-strandedfibronectin III-like domains (D1 and D2) that are structurallyhomologous to those in hGHR, a single 22–amino acid trans-membrane domain, and a 237–amino acid cytoplasmic domain(31, 32). EPO dimerizes two receptors on the cell surface in asequential manner; site 1 of EPO binds a first receptor withhigh affinity (Kd ~ 1 nM), then EPO binds a second receptorthrough site 2 with lower affinity (Kd ~ 1 µM ) (33, 34).
The structure of the EPO:EPOR2 complex shows that EPO isbound at the outside of the elbow formed by the D1 and D2
R E V I E W
N
C
C
N
Site 2Site 2
Site 1
hGHhGH
hGHR hGHR
D2 domain
D1 domain
Fig. 3. In this and the next five figures of group 1 “four α-helical”cytokine:receptor complexes, the receptor chains are shown inribbon rendition in blue colors. Different shades of blue indicatedifferent receptor chains in the complex. The cytokines arecloaked in a molecular surface, shown in greens, above ribbondiagrams in purple colors. Slight variations of color distinguishdifferent cytokine chains. The N- and C-terminal ends of receptorchains are indicated. In general, the C-terminal ends are orientedtoward the transmembrane region, although intermediate do-mains are sometimes interposed between the N terminus and themembrane. Specifically, as illustrated in this figure and Fig. 4,hGH and EPO are long-chain four α-helical bundle cytokines.Both cytokines interact with two identical copies of their receptorsusing two completely different interaction sites on the cytokine,termed Site 1 and Site 2. Interaction Sites on each of the two re-ceptors use many of the same residues.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 4
R E V I E W
A
B C
NN
C C
N
N
C C
N
N
C
C
Site 2Site 1
EPOEPO
Peptide
EPOR
EPOR2
EPOR
D2 domain
D2 domain
D1 domainomainD1 do
Fig. 4. Activating and inhibitory complexes of EPO receptors. In these structures, the EPOR molecules were truncated at the C-termi-nal positions (C) to remove the single-spanning transmembrane segment and the intracellular portions of the receptor. Thus the posi-tions labeled C indicate the approximate separation of the single membrane crossing helices. (A) The EPO:EPOR2 complex shows a40° difference in orientation (about the vertical axis) of the receptor that binds to Site 2 relative to that seen in the hGH:hGHR2 complex. The Site 1 interactions (right side) produce a similar orientation of the cytokine:receptor interface in both the EPO:EPOR andhGH:hGHR complexes. Thus the interaction at Site 2 produces a very different angle of the Site 2-binding receptor around the per-pendicular to the membrane surface. The EPO:EPOR2 complex also differs from the hGH:hGHR2 complex in that there are no recep-tor-receptor contacts between EPORs. (B) The structure of EPOR2 brought about by binding so-called “EPO-mimetic” peptides. Dis-covered by selection from bacteriophage-displayed peptides, these peptides themselves dimerize to bring receptors close together, al-though in a completely different manner than that induced by actual EPO binding [as in (A)]. The relative orientation of one receptorchain to the other in this complex is 180° around the vertical axis perpendicular to the membrane. Although the peptide shown (EMP1)activates the EPO response, other similarly discovered peptides can be activating or inactivating for EPOR. (C) The structure of theEPOR2 dimeric complex may represent a means of ensuring that EPOR transmembrane domains are held apart so that EPORs donot come close enough together to become activated by chance. Shown is a view looking down onto the plane of the membraneplane. Thus, the greater distance between the positions of the C-termini (labeled C, top left, bottom right) may be a factor in prevent-ing the association of the transmembrane and intracellular domains inside the cell.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 5
domains (22, 35), as in the hGH:hGHR2 complex. However, thesecond receptor chain in the EPO:EPOR2 complex is rotated by110° perpendicular to the membrane relative to the first, which is40° less than that seen in the hGH:hGHR2 complex. TheEPO:EPOR2 complex also differs from the hGH:hGHR2 complexin that there are no receptor-receptor contacts. These crystal struc-tures show there is a difference in the way the receptors for groupI cytokines can be oriented for signaling to occur (Fig. 4A).
As for the hGHR, virtually the same residues of the EPORchain are buried on binding at both site 1 and site 2 of EPO,even though the epitopes on EPO to which they bind are entire-ly different from one another. Binding to the two different sitesby the same residues of EPOR is achieved by a small twist ofthe D1 and D2 domains relative to one another. The side chainsinvolved in binding EPO all derive from the loops L1 to L6.Phe93, the structural counterpart of Trp104 in hGH, has a similar-ly large effect on affinity when mutated (36). The hot spot onEPOR is remarkable, because it also provides the binding sitefor synthetic peptides (see below) and the interacting surfacefor unliganded receptor-receptor dimers that may form an off-state of receptors (15).
Evidence suggests that bringing the two extracellular por-tions of the EPO receptors close together can activate the intra-cellular pathway. For example, point mutations that introducecysteine residues into any of three positions in the EPOR(residue numbers 129, 131, and 132) form disulfide-linked in-termolecular dimers between the cysteines introduced into eachreceptor and the same cysteine on the other receptor and causeconstitutive signaling (37-40). These cysteines are arrayedaround the D2 domains near the membrane spanning region.Disulfide bonds here would place the receptors about 8 Å closertogether than in the EPO complex with EPOR, and rotated by atleast 60° from their normal EPO-activated position in the mem-brane plane. As a second example, a construct of two EPOmolecules linked from C-terminal of the first to N-terminal ofthe second, and both mutated to abrogate the weaker site bind-ing at site 2, leads to activation of the EPOR response, and withalmost the same efficiency as for wild-type EPO (41). Third,some but not all antibodies to EPOR can also lead to activation(42). Fourth, several “EPO-mimetic” peptides (EMPs) havebeen discovered by selection to bind, dimerize, and activate theEPOR (43, 44) (Fig. 4B).
These data, along with the structure of the EPO:EPOR2complex, suggest that close proximity of receptors is neces-sary for activation. However, the efficiency of activation ishighly dependent on the way in which receptors are dimer-ized, and on the length of time that they are held together. Forexample, the median effective dose (ED50) for EPO activationis 10 pM, just 5.6% of its median inhibitory concentration(IC50), implying that only about 60 receptors are dimerizedfor a 50% response in cells. The ED50 for EMP is typicallygreater than the IC50, where 600 receptors would be dimerizedfor a 50% response (22, 35). Some dimerizing peptides, forexample, clearly antagonize the response (45, 46), showingthat dimerization is not sufficient for activation of the recep-tor. This data and others showing that covalently dimerizedpeptides are 1000-fold more active (46) suggest that activa-tion depends on correct orientation and residence time.
Residence time has also been shown to be important for sig-naling in the hGH receptor. For example, a systematic set ofmutations in hGH that decease the residency time at site 1 be-
low a half-life (t1/2) of about 1 min proportionately diminishsignaling as detected by Janus kinase 2 (JAK-2) phosphoryla-tion and cell proliferation (47). Residency times longer than t1/2 ~ 1 min do not further enhance signaling, presumably be-cause the signal is maximal. Interestingly, the residency time forthe wild-type hormone is about 30 min, which is more than am-ple for maximal signaling.
The intracellular domain of the EPOR contains eight tyro-sine residues that can be phosphorylated after EPO binds. Thesephosphorylation sites allow the intracellular domains to interactwith cytosolic signaling factors that include signal transducerand activator of transcription 5 (STAT-5), the adaptor proteinGrb-2, and phosphatidylinositol (PI) kinase (48). Mutagenesisshows that no particular phosphotyrosine is essential for signal-ing (49). Indeed, a single tyrosine is sufficient to support differ-entiation of the erythroid lineage (50). This suggests that any ofthe phosphotyrosine sites can serve as a docking site for STAT-5. Once bound to the phosphorylated EPO receptor, STAT-5 isphosphorylated by JAK-2 protein kinase, which leads to STAT-5activation through dimerization and transport to the nucleus(51). Thus, regulated oligomerization reactions appear to be acommon theme both at the level of the membrane receptor andat the level of downstream effectors.
“Off-state” of receptors. The cell must have a means ofdistinguishing receptors associated in response to cytokinebinding, as opposed to random collisions. Recent x-ray studiessuggest that the extracellular domains of the EPOR mightdimerize in an inactive antiparallel fashion in the absence of ac-tivating ligand (15). In this state, the predicted distance betweenthe membrane insertion points would be about 70 Å, whichwould prevent the dimerization of intracellular domains. Such areceptor off-state would provide an active means of reducingnonspecific “noise” by preventing ligand-less signaling throughrandom receptor association.
To test this model, a fusion protein was constructed that con-tained complementary portions of the enzyme dihydrofolate reductase (DHFR) that were inactive when separate but becameactive when brought together. These portions were fused to thetransmembrane segment on the cytoplasmic side of EPOR (16).Addition of EPO was required for DHFR activity. However,when long glycine-rich linkers (lengths of 5, 10, or 30 aminoacids) were placed between the EPOR membrane-spanning andDHFR fragments, the receptors could be activated without EPO.This suggests that, without EPO, the intracellular domains ofthe EPORs are held apart, possibly in the manner seen in thestructure of the [EPOR]2 dimeric complex (15) (Fig. 4C).
Dimerization or oligomerization of a “resting state” of EPORhas also been established by immunofluorescence (52). Mutage-nesis shows that the correct orientation of L253, I257, and W258,located in the juxtamembrane region of the transmembrane α-helix in EPOR, is required for activation of the receptors. Thetransmembrane- (52) and ectodomain-associated dimers mayrepresent redundant mechanisms for ensuring that free receptorsremain in their off state but available in a cluster. When the EPOligand binds, it changes the orientation to an active state.
Recently, a peptide was discovered that activates the EPO re-sponse but binds to a noncompeting, therefore different, regionof the EPOR than does EPO (53). The site is close to the mem-brane surface on the extracellular portion of EPOR, and appearsto bind to EPOR in a 1:1 ratio. Amazingly, the peptide has thesame sequence as the sequence in EPOR that it binds to. One
R E V I E W

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 6
plausible function for this peptide is that it may antagonize anEPOR-EPOR (twofold symmetric) inactive dimer. However, itsbinding site on EPOR differs from that seen in EPOR-EPORdimers by crystallography, so any interference in that associa-tion may be indirect.
Other receptors can exist in preassociated states. Fluores-cence techniques have suggested preassociated oligomericforms for IL-1 receptors (IL-1Rs)(54), IL-2Rs (55), erbB2 receptors(56, 57), and EGF receptors(EGFRs). Crystallographic studiesshow that TNFR can form paralleland antiparallel dimers (58-60). Theantiparallel dimer would block TNFbinding, as is seen in the EPOR2complex.
Peptide mimetics and receptor hotspots. Several investigators have discov-ered small peptides from phage displaylibraries that were optimized for bind-ing receptors other than the EPOR; in-terestingly, they do so at the same hotspots used by the natural ligand. Onepeptide selected for binding to the ex-tracellular domain of the thrombopoi-etin receptor (TPOR) was found to haveagonist activity (61), and when cova-lently dimerized, it produced an agonistas potent as TPO itself (62). More po-tent agonists have been developed forEPOR (63) and G-CSFR by similarroutes. In another case, a 19-residuepeptide was selected for binding anFGFR. When the peptide was dimer-ized by fusion to the c-Jun leucine zip-per, it activated the FGFR in presenceof a synergistic binder, heparin, with apotency similar to that of FGF (64).Thus, small mimetics can be found for the cytokine receptors.
Peptides that compete with the normal ligand have also beendiscovered by phage display for vascular endothelial receptor(VEGFR) (65, 66), FGFR (67), gp120 (68), and the Fc region ofIgGs (69). The fact that all these phage-derived peptides targetthe natural ligand-binding sites on these receptors suggests thatthe surface chemistry of these receptor hot spots is somehowprimed for binding. Generally, these sites are hydrophobic, wellexposed on the surface, and positioned at hinge regions in thestructure, allowing adaptability, or conformational promiscuitysuch that they are the sites most suited to binding ligands (69).
Heterodimeric complexes in IL-4 signaling. IL-4 illus-trates how a helical bundle can heterodimerize two receptors.Many cytokines bind two entirely different receptors in theirsignaling complex. IL-4 is a group I cytokine belonging to theshort-chain subfamily. It binds first to the IL-4αR, then to amultipurpose receptor termed the common gamma chain (γC),which is common to several other cytokine complexes, such asthose containing IL-2, IL-7, and IL-9 (70). The use of differentreceptor chains opens the way to broader intersections of differ-ent signaling pathways.
There are important differences in the way that IL-4 binds itsIL-4αR, compared to the way that hGH and EPO bind their
high-affinity receptors. The high-affinity site on IL-4 that bindsIL-4αR (Kd = 150 pM) (71) corresponds structurally to theweaker site 2 interface of the hGH or EPO complexes (Fig. 5).The association rate of IL-4 for the IL-4αR shows an unusuallyhigh on-rate (that is the rate at which IL-4 associates with IL-4R) that can be ascribed to a positively charged surface that isattracted by the negative surface on IL-4αR (72, 73). Mutation-
al analysis of the IL-4αR shows thatit has two independent binding re-gions. Each region has a singlecharged side chain, important forbinding IL-4, that forms hydrogen-bonded pairs with IL-4; these re-gions are positioned between Asp72
and Arg88, and between Tyr183 andGlu9, respectively (74). The bindingsite on the receptor has been furthermapped by alanine-scanning muta-genesis to identify and evaluate thecontributions of four hydrophobicside chains (Ile100, Leu102, Tyr103,and Leu208) that interact with simi-larly hydrophobic side chains on thecytokine (75). Interestingly, IL-13, aclose homolog of IL-4, binds first tothe IL-13αR and then recruits theIL-4αR for signaling. Presumably,IL-13 uses sites similar to those usedby IL-4, but in opposite order ofbinding.
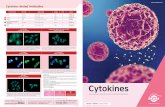
The 2:2 G-CSF:G-CSFR andIL-6:gp130 complexes. AlthoughG-CSF is structurally similar to EPOand hGH, it activates its receptorthrough a higher stoichiometry(2:2), and by binding on the “back”side of the receptor. G-CSF is a fourα-helical bundle cytokine that first
binds one receptor, and then forms a dimeric complex with an-other 1:1 G-CSF:receptor complex (23). In the 2:2 complexgenerated by receptor ectodomains that lack their N-terminal Igdomains, the receptors make contact, but there is no interactionbetween the two G-CSF molecules (Fig. 6). Each G-CSF formsa receptor contact of about 820 Å2 at the site 2 interface (helicesA and C) similar to that for IL-4. The interface also involves po-lar amino acid side chains. Incorporated into the interface arewater molecules that are proposed to fill cavities and reduce thepenalty of desolvating a charged protein interface. A secondsmaller interface of about 384 Å2 forms between the normallyflexible N-terminal regions of G-CSF in one of the 1:1 com-plexes, and the D2 domain of the second receptor, in a novelsecond interface.
The region in G-CSF that corresponds to site 1 in hGH is notinvolved in initial receptor binding, and it remains exposed inthe 2:2 complex. However, alanine-scanning mutagenesisthroughout this region suggests that site 1 does play a role in thebiological response (76). Higher-order complexes up to 4:4 stoi-chiometry also may form (77), suggesting that higher ordercomplexes may play a role in signaling. It is possible that thevacant site 1 region on G-CSF seen in the 2:2 complex is usedin these higher order complexes. The structure of the extracellu-
R E V I E W
N
C
IL-4
IL-4∝R
R88
D72
E9
Y185
Fig. 5. IL-4 is also a group 1 four α-helical cytokine,although in the short-chain subclass. The IL-4:IL-4αR structure mimics the Site 2 interface of the hGHor EPO complexes, and is one of the tightest bindingcytokine-receptor interfaces, assisted by oppositelycharged interfaces.
www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 7
lar portion of the G-CSFR contains additional domains not included in the x-ray structure, including an N-terminal Ig domain that stabilizes the signaling complex (78) and three additional fibronectin III domains that lie between D2 and thetransmembrane segment. Thus the exact nature of the active G-CSF:receptor complex still needs to be elucidated.
The structure of the complex formed between IL-6 and the full-length ectodomain of gp130 with the N-terminal Ig-like domain hasbeen determined (79). Two gp130:IL-6 complexes associate so thatthe N-terminal Ig domains of gp130 also bind to a third site on theopposite IL-6 of the complex to make a twofold symmetric “ring”of gp130:IL-6:gp130:IL-6:. Signaling may depend on bridging oftwo receptors by the ligand, but differently than for EPO or hGH.
The 2:2:2 IFN-γ receptor complex. The IFN-γ receptorcomplex is structurally analogous to two hGH-receptor complex-es glued together by domain swapping between two ligands.IFN-γ is composed of six helices and forms a V-shaped, inter-locked dimer in solution. The fifth and sixth helices of onemonomer pack along with helices 1 to 4 of the twofold relatedmonomer to form a “domain-swapped” dimer (80). The cytokinedimer binds with very high affinity (Kd ~ 10-10 M) to two IFN-γRα to form a 2:2 complex (81). This 2:2 complex then binds totwo accessory β-receptor chains from the group II cytokine re-ceptor family that are specific to the type of responding cell, ac-tivating a 2:2:2 signaling complex (82) (Fig. 7).
IFN-γRα contains two fibronectin III-like domains, D1 andD2, that are related in sequence to those of hGHR or EPOR. Incomplex with IFN-γ, it utilizes the loops between β strands ofD1 and D2, as in the hGH:receptor complex, and buries a sur-face area of 960Å2 in the interface, of which one-third is pro-vided by three aromatic side chains on the receptor (Tyr52,Trp210, and Trp85). One notable aspect of the interface is the
conservation of charged residues that pack up against trypto-phan and tyrosine side chains, where they account for a signifi-cant portion of the binding energy. Helices A, B, and F, and theA-B loop form the IFN-γ surface in an interface region that isvery different from that of the EPO or hGH complexes. In mak-ing the complex, the A-B loop in the cytokine undergoes a con-formational change, as is seen for G-CSF (83).
The twofold symmetric IFN-γ dimer evokes a twofold sym-metric dimerization of the receptors such that the D1 domainsof the receptor are rotated ~40° from corresponding positions in
R E V I E W
N N
C C
IFN-γ dimer
IFN-γR∝
1Domain 1
Domain 2A-B loop
Fig. 7. IFN-γRα is a short-chain group 1 four α-helical cytokinethat is intimately dimerized in a domain-swapped form. The dimerrecruits a 2:2:2 signaling complex.
AN
NN
C C
C
C
B
Site 2
Helix C
Helix A
Helix A
G-CSFRdimer
G-CSF molecules
Helix C
Site 1
Fig. 6. G-CSF is also a short-chain, group 1 four α-helical cytokine. A dimer of G-CSF binds to two receptors using two identical inter-actions that are each the mimic of site 2 interfaces of the hGH or EPO complexes. (A) A side view that shows the lack of contact be-tween G-CSF molecules. (B) A top view (looking down on the membrane) that shows the lack of contact between G-CSF receptors orG-CSF molecules.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 8
the hGH:hGHR2 complex. The are two identical sites for bind-ing two IFN-γRα molecules in the 2:2 complex. However, thisleaves the D2 domains of the IFN-γRα receptor at the mem-brane entry point, distant from each other.
The structure of the complex of IFN-γ with IFN-γRβ is need-ed to reveal the atomic details of the active signaling complex.Mutagenesis of IFN-γ suggests that it uses determinants close tothe C terminus of the IFN-γ molecule to bind to the IFN-γRβ. A3:1 IFN-γRα:IFN-γ complex seen crystallographically suggestsa binding site for IFN-γRβ. In this complex, the crystallographyshows a third IFN-γRα that binds to the spot that is expected tobind IFNRβ, and places the C-terminal regions of the two chainsclose together on the cytoplasmic side of the membrane (84).This notion is supported by an experiment in which IFN-γ wasmade into a continuous single-chain dimer by gene fusion, andthen mutated to remove just one of the binding sites for IFN-γRα. This 2:2:2 complex activates both JAK-1 and JAK-2 pro-tein kinases. This construct shows that the 1(IFN-γ dimer):1:1complex can still evoke a cellular response achieved through sin-gle copies of the receptor-bound JAK-1 and JAK-2. This tran-sient IFN-γ (dimer):IFN-γRα:IFN-γRβ complex, by analogywith the IL-4:IL-13αR:IL-4αR complex, indicates that thecross-phosphorylation steps are very efficiently carried out, pre-sumably by just one side of the complex. Ligand binding releas-es receptors from any restraint such as that brought about by in-active oligomeric states of the receptors (85).
The 2:2:2 IL-10:IL-10R1:IL-10R2 complex. Structurally, theIL-10:receptor complex is closely analogous to the IFN-γcomplex. IL-10 is a natural immune suppressant, and so is hi-jacked by viral mimetics of IL-10 to anaesthetize the immune sys-tem. Interestingly, the ways that the viral mimetics activate IL-10so as to enhance viral infectivity are completely different in eachcase, and produce different spectrums of downstream signaling(86).
IL-10 is a six-helix cytokine that forms a twofold symmetric,intertwined dimer in which four helices (A-D) from onemonomer conjugate with two (E and F) from the other to form a12-helix twofold symmetric bundle. IL-10 and IFN-γ containseveral conserved residues in their dimeric cores, which sug-gests a possible “fingerprint” for detection of other members ofthis dimerized family (87, 88). The IL-10 dimer first forms a
twofold symmetric dimer with high affinity (Kd ~ 8 nM) for IL-10R1 (Fig. 8), and then recruits the second, low-affinity site onIL-10R2 that activates the signaling pathway mediated by JAK1or TYK2 protein kinases and STATs.
There are viral homologs of IL-10, for example, in cy-tomegalovirus (CMV) and Epstein-Barr virus (EBV). Theirstructurally distinct scheme is paralleled by biological differences that highlight receptor orientation in the activatingcomplex (86). EBV, though it presents a homolog of IL-10, isunable to stimulate thymocyte production or mast cell prolifer-ation, for example, whereas human IL-10 (hIL-10) can. UnlikehIL-10, in CMV IL-10 the dimer is crosslinked by a cysteinebridge. Complexes of CMV IL-10 with IL-10R1s show that theCMV IL-10 and human IL-10 have a 40° difference (viewedparallel to the membrane plane) in their intramolecular angleas a dimer (86). The structures of 2:2:2 activating complexeshave not yet been determined. But the markedly different acti-vation spectra emphasize the evolutionary tendency to raise themost appropriate of pleiotropic responses, and they show thataspects of the orientation of IL10-R1/R2 receptors after cy-tokine dimer binding are key to effector signaling. Differentialactivation of various signaling pathways is presumably due todifferent complexes formed on the cell surface; the key is thateach complex is distinct enough in geometry and residencetime that it induces varied downstream signaling.
Group 2, β Sheet–Rich CytokinesTNF:receptor complexes and the formation of higher orderoligomers. TNF and its family members step up the degree ofoligomerization to trimeric structures, where the ligand serves asthe bridging scaffold, with no receptor-receptor contacts. TNF be-longs to the group 2 (β-sheet rich) cytokines and is implicated inmany inflammatory processes and apoptosis (89). TNFα is a 17-kD “β-jellyroll” that forms a wedge-shaped homotrimer (90, 91).Signaling is mediated by either of two TNF receptors, TNFR1 (55kD) and TNFR2 (75 kD). The structure of the extracellular por-tion of the TNFR1 has been solved by crystallography and showsthat it is composed of four cysteine-rich domains. The extracellu-lar domains of the TNFR1 crystallize as a dimer [reviewed in(92)].
Upon binding the TNF trimer, the receptor trimerizes in an ac-tive configuration (92). TNFR-associated factors (TRAFs) are afamily of cytoplasmic proteins that assist in transducing signalsfor the TNFR superfamily inside the cell (93). These proteins arealso trimeric structures in isolation (94). Thus, the TNF trimer it-self dictates the assembly of the TNF:TNFR trimer (59) (Fig. 9A)that is detected by a “trimer detection” system in the TRAFs.
Several forms of the dimeric receptor complex have been de-termined. At pH 7.5, the elongated receptor chains dimerize inboth a parallel manner and an antiparallel association. In the an-tiparallel dimer, the TNF-binding interfaces are buried, suggestingthat this may be the means of maintaining receptors in their off-state (58), which is reminiscent of the case with EPORs (Fig. 9B).In the parallel off-state form of the receptor dimer, the TNF bind-ing site is exposed and open to bind the cytokine, suggesting thatthis mode may promote clustering of TNF:TNFR complexes onthe cell surface.
The activity of TNF receptors is regulated in several ways;this is essential because overexpression of the intracellular do-mains alone leads to activation, mediated by an intrinsic aggre-gation mechanism (95). First, the extracellular concentration of
R E V I E W
N N
CC
IL-10 dimer
IL-10R
Fig. 8. IL-10 is a long-chain group 1 four α-helical cytokine thatfirst recruits IL-10R1 and then recruits the second, low-affinitysite on IL-10R2 that leads to activation.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 9
TNF is reduced by the presence of the cleaved extracellular do-main of TNFR in plasma. Second, the TNFRs themselves ag-gregate in at least two ways, as described above. Third, afterTNFRs are activated on the cell surface, the complexes are en-docytosed and the signal is truncated. The structure of TNFRdimers at pH 3.5 shows that they form dimers that share a largearea of contact (2900Å2), and compete directly for the TNFbinding surface, suggesting that the low-pH endosome can ter-minate signaling by this “competitive” dimerization mechanism(96). Fourth, silencer of death domains (SODD) associate withthe cytoplasmic “death domains” of TNFR1s. Binding of TNFreleases SODD from TNFR1, permitting recruitment of pro-teins such as TNFR-associated death domain (TRADD) andTRAF2 to the active TNFR1 signaling complex. SODD associ-ation may exemplify a general mechanism for preventing spon-
taneous signaling by death domain-containing receptors (95).The receptors for FGF, nerve growth factor (NGF), VEGF,
insulin, and EGF include intrinsic tyrosine kinase domains onthe cytoplasmic side of their receptor chains.
FGF receptor and the role of extracellular matrix. FGF il-lustrates how a polysaccharide can facilitate binding betweenligand and receptor. Crystal structures of the FGF:receptorcomplexes show that heparan sulfate chains in the extracellularmatrix enhance the binding between FGF and its receptor. Thissynergy between carbohydrate and protein has important rolesin development and organ homeostasis. The FGF growth factorfamily consists of 23 cytokine isotypes. There are four receptorisotypes, each with a different specificity for the FGFs. FiveFGFs have both a hydrophobic receptor-binding epitope and asomewhat variable heparin-binding surface that localizes the
R E V I E W
N
N
C C C
N
A B
C
N
N
C
C
N1N2
C2
C1
TNFR trimer
Fig. 9. Group 2 “β-sheet rich” cytokines. (A) TNF is inthe β-jellyroll subclass. A TNF trimer recruits three TNFRs to activate the response. (B) Several forms ofthe dimeric receptor complex show both a parallel (left)and an antiparallel association (right). The parallelform of the receptor dimer may promote clustering ofTNF:TNFR complexes on the cell surface. (C) The an-tiparallel mode may be the means of maintaining re-ceptors in their off-state.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 10
FGF to the extracellular matrix (97). The receptors are com-posed of two or three ~110 amino acid IgG-like domains (D1-D3), each containing seven β strands. The D2 domain bindsheparin with high affinity (98).
FGF2 forms a twofold symmetric 2:2 complex with the re-ceptor FGFR1 (24, 99), as is also found for a FGF1:FGFR2complex (100). Each monomeric FGF cytokine binds the D2and D3 domains of one FGFR, which induces a change in theFGFR that then leads to (FGF:FGFR):(FGF:FGFR) dimeriza-tion mediated primarily by the D2 domains within the FGFRs.The dimer interaction area between receptors is surprisinglysmall, about 300 Å2, and the FGF molecules have no contactwith each other in the complex. In contrast, the FGF:FGFR in-teraction surface is large (2700 Å2). The cytokine’s contactswith the D2 domain of the receptor are primarily hydrophobic,but also have important polar interactions. For example, Arg250
in the D2-D3 linker is oriented by neighboring residues to maketwo conserved hydrogen bonds to the cytokine. The preponder-ance of central hydrophobic interactions, along with key polarinteractions to encode specificity, are reminiscent of the EPOand hGH complexes with their receptors.
Dimerization of the receptor also depends on the heparin of ex-tracellular matrix. An optimal concentration exists for heparin toassist in cross-linking receptors (92). Higher than saturating con-centrations of heparin compete for all the sites and antagonizecrosslinkage. Thus, heparin can either promote or prevent the pro-liferation and migration of cells involved in vascular development.
The structure of the initial 2:2 complex between FGF andFGFR shows a basic canyon that extends from FGF2, across theD2 dimer, and across the D2-D3 linker of the FGFR, that couldaccommodate polyanionic heparin and stabilize the dimer (24). A crystal structure of the ternary complex ofFGF:FGFR:FGFR:FGF with a decasaccharide bound shows
how an activating heparin analog actually binds to the 2:2 com-plex (101). Surprisingly, two decasaccharides bind predomi-nantly to the FGF molecules (perhaps reflecting the FGF asso-ciation with the matrix before activation), and increase theaff inity of each FGF for its receptor in 1:1:1 FGF:FGFR:heparin complexes (Fig. 10A). Each decasaccharide adopts ahelical structure like that seen by fiber diffraction of heparinalone. The helix is generated by repeating D-glucosamine(GlcN)-L-iduronic acid (IdoA) disaccharide units, each onelinked to the next by a α 1-4-glycosidic bond. Each disaccha-ride unit is sulfated at three positions, and the sulfate and car-boxylate groups form the negatively charged edges of the hep-arin helix that appear every 17 to 19 Å on each side of the helix.Heparin polysaccharides have a nonreducing end (O4) and a re-ducing end (O1). The decasaccharides bind with their nonre-ducing ends toward the center of canyon, preserving the overalltwofold symmetry of the complex. These are each bound by 30hydrogen bonds. Of these, 25 are found within one FGF:FGFRpair (16 from EGF, nine from EGFR), and five more help bindthe neighboring FGFR, augmenting the cross-linking and hencestability of the quaternary 2:2:2 complex. These include saltbridges from positive charges of five lysines on each D2 of theFGFRs to sulfates of heparin.
The structure of the 2:2:2 FGF:FGFR:heparin complexshows how heparin draws together the protein components andincreases affinity of FGF for its receptor (101). It explains howsmaller oligosaccharides, including hexasaccharides, can bindand dimerize FGFRs in spite of not being able to reach fromone FGF:FGFR complex to the other, and do so even whilelacking the five-hydrogen bond saccharide-FGFR contact. It al-so can explain why excess heparin can antagonize the receptorsthrough competition for the secondary five-hydrogen bondedsite on the “other” receptor. If this model is correct, the optimal
R E V I E W
NN NN
N
N
A B
Heparin
Heparin
FFG
FGF
FGFR
FGFR
Fig. 10. Other examples of group 2 “β-sheet rich” class cytokines. (A) FGF is in the β-trefoil subclass. The 1:1:1 FGF-FGFR-heparincomplex illustrates how intracellular matrix contributes to the activation of the FGFR complexes. Heparin is rendered in stick modelcolored red. This is a side view; (B) is a top view onto the membrane.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 11
ratio of oligosaccharide to receptor would be 1:1. Anotherstructure of the complex formed with a bound decasaccharideconfirms a role for the heparin molecules in drawing the com-plex together (102). The conformational flexibility of IdoA, acrucial saccharide that becomes sulfated and is part of the lig-and, must play a role in specific recognition of various FGFs orFGFRs that are variable in sequence in this portion of the hep-arin site. The model suggests that heparin serves to bind in theinterface between FGF and FGFR in a 1:1:1 complex to bringabout a conformational change that leads to productive dimer-ization of FGFR, and hence to intracellular autophosphorylationas the first step in the signaling cascade (103) (Fig. 10A).
Another model proposes a preexisting inactive complex oftwo FGFRs. When FGF binds to this complex. it alters the con-formation of the preformed, inactive dimer to initiate signaling(97, 104, 105). Heparin, through its positional determination inthe intracellular matrix, ensures fibroblast growth once suchcells are in close contact with matching neighboring cells.
VEGF: a simpler symmetric paradigm. VEGF represents asimpler symmetrical complex where binding occurs at subunitinterfaces. VEGF is a member of the “cystine knot” subclass ofgroup 2 cytokines that all act as homodimers. Three subfamiliesare recognized that are distinguished by their different ways offorming homodimers. VEGF stimulates growth of endothelialcells (106). It is usually attached to proteoglycans in the intracel-lular matrix by heparan sulfate, and it binds to two different ty-rosine kinase-containing receptors, Flt-1 and KDR, to effect sig-naling. These receptors are in the PDGF receptor family, and arecomposed of five or seven Ig-like domains. In the VEGF-Flt-1complex, only Ig-like domains 2 and 3 provide the binding inter-face from the receptor side. Based on sequence analysis, theplatelet-derived growth factor receptor (PDGFR) family of re-ceptors may all use domains 2 and 3 in a similar way, with do-main 4 adding stability by receptor-receptor contacts. The inter-face from the hormone is rich in leucine and isoleucine residues,which constitute nine of 47 interface residues (107) (Fig. 11).
The NGF-TrkA complex suggests convergent evolution ingroup 2 cytokines. Like VEGF, NGF is in the dimeric “cystineknot” subgroup of the group 2 cytokine family. It is bound bytwo receptors of different types, TrkA and p75. Like the VEGFR chains, TrkA is composed of multiple Ig-like domains(five in total), although only the one closest to the membrane is
required for binding NGF. Binding occurs with little conforma-tion change (66).
p75 is a member of the TNFR class, and it promotes apoptosiswhen NGF binds in the absence of TrkA. Despite the many similari-ties to the VEGF:receptor complexes that reflect underlying homol-ogy, the NGF ligands dimerize in quite different ways, and the bind-ing domains of the NGF versus TNF receptors are at almost oppo-site poles of the receptor molecules (Fig. 12), suggesting convergentevolution of ligand induced dimerization, but mediated by a differ-ent domain in the receptor, and in a different way. In the NGFR, thecomplex brings membrane-proximal domains of the receptor chainswithin 20 Å to effect signaling, whereas in the VEGFR, several do-mains of undetermined function intervene between ligand-bindingdomains and the membrane.
BMP-2, TGF-β, and their serine-threonine kinase recep-tors. The cystine knot hormones form hetero-oligomeric recep-
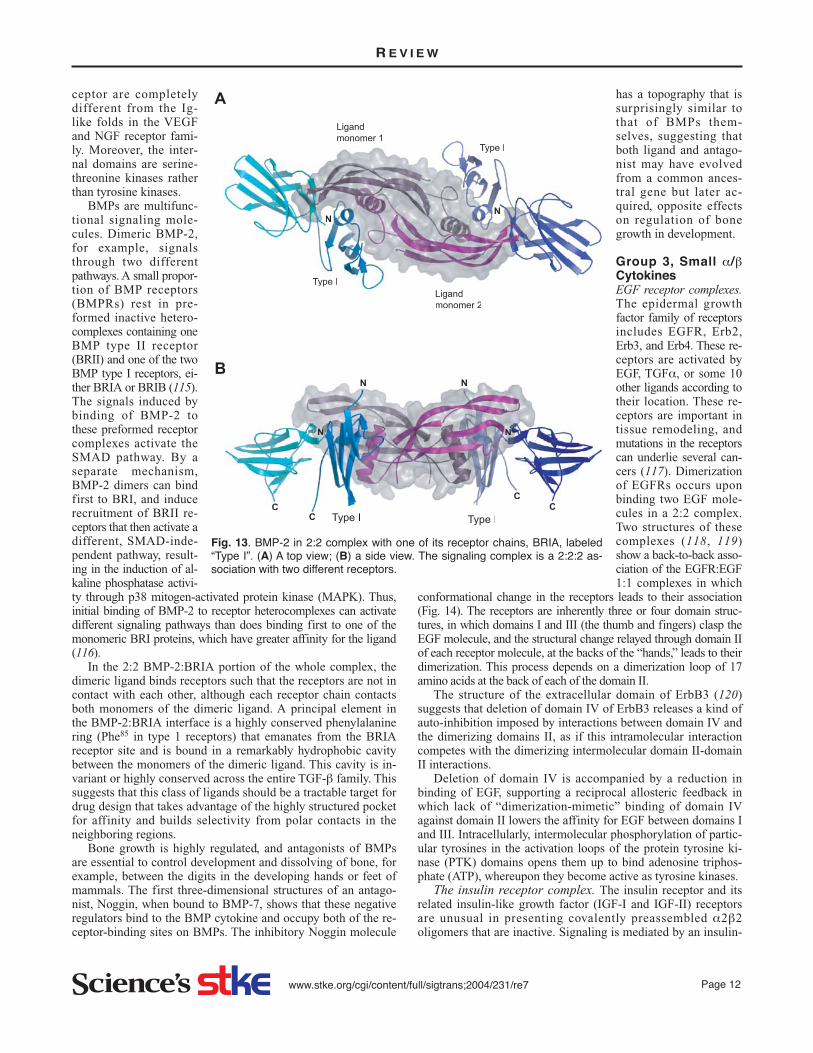
tor complexes with receptors that have unique folds. This thirdsubfamily of the group 2 β-sheet dimeric cystine knot cytokinesalso includes bone morphogenic growth factors (BMPs) and tis-sue growth factors (TGFs) that control proliferation and differ-entiation of bone and connective tissue vis-á-vis their position.These are dimeric proteins where each monomer is based on anine-stranded β sheet and one α helix. These cytokines act ashomodimers, and recruit two pairs of heterodimeric receptors toform a 2:2:2 signaling complex. The BMP-2 in 2:2 complexwith only one of its receptor chains, BRIA (108) (Fig. 13), andthe transforming growth factor TGFβ3 in complex with TGFβR-IIec (109), illustrate these quaternary interactions.
Unlike the cytokine receptors, these receptors have bindingdomains resembling neurotoxins that bind with high affinity tothe acetylcholine receptor (110-113) and are like domainsfound in the low-density lipoprotein (LDL) receptor (114).Each domain is composed of six extended strands that form athree-finger-like motif stabilized by six cystine bridges in aknot at one end of the molecule. The cystine bridges hold theextensively β-sheet structures together to present a large bind-ing surface in the form of a groove that is about 15 Å by 5 Å by5 Å in dimension. Although the BMP-2 ligand is similar toVEGF and NGF, the neurotoxin-like binding domains in the re-
R E V I E W
N N
CC
Flt-1R
VEGF
Fig. 11. VEGF is in the cystine knot subclass of group 2 cy-tokines. The 1:1 VEGF-Flt-1 complex is shown. Another receptorchain, KDR, is required for activation.
N N
C C
Fig. 12. NGF-TrkA uses just the membrane-proximal Ig-like domain and must bind a second receptor p75 to activate.
www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 12
ceptor are completelydifferent from the Ig-like folds in the VEGFand NGF receptor fami-ly. Moreover, the inter-nal domains are serine-threonine kinases ratherthan tyrosine kinases.
BMPs are multifunc-tional signaling mole-cules. Dimeric BMP-2,for example, signalsthrough two differentpathways. A small propor-tion of BMP receptors(BMPRs) rest in pre-formed inactive hetero-complexes containing oneBMP type II receptor(BRII) and one of the twoBMP type I receptors, ei-ther BRIA or BRIB (115).The signals induced bybinding of BMP-2 tothese preformed receptorcomplexes activate theSMAD pathway. By aseparate mechanism,BMP-2 dimers can bindfirst to BRI, and inducerecruitment of BRII re-ceptors that then activate adifferent, SMAD-inde-pendent pathway, result-ing in the induction of al-kaline phosphatase activi-ty through p38 mitogen-activated protein kinase (MAPK). Thus,initial binding of BMP-2 to receptor heterocomplexes can activatedifferent signaling pathways than does binding first to one of themonomeric BRI proteins, which have greater affinity for the ligand(116).
In the 2:2 BMP-2:BRIA portion of the whole complex, thedimeric ligand binds receptors such that the receptors are not incontact with each other, although each receptor chain contactsboth monomers of the dimeric ligand. A principal element inthe BMP-2:BRIA interface is a highly conserved phenylalaninering (Phe85 in type 1 receptors) that emanates from the BRIAreceptor site and is bound in a remarkably hydrophobic cavitybetween the monomers of the dimeric ligand. This cavity is in-variant or highly conserved across the entire TGF-β family. Thissuggests that this class of ligands should be a tractable target fordrug design that takes advantage of the highly structured pocketfor affinity and builds selectivity from polar contacts in theneighboring regions.
Bone growth is highly regulated, and antagonists of BMPsare essential to control development and dissolving of bone, forexample, between the digits in the developing hands or feet ofmammals. The first three-dimensional structures of an antago-nist, Noggin, when bound to BMP-7, shows that these negativeregulators bind to the BMP cytokine and occupy both of the re-ceptor-binding sites on BMPs. The inhibitory Noggin molecule
has a topography that issurprisingly similar tothat of BMPs them-selves, suggesting thatboth ligand and antago-nist may have evolvedfrom a common ances-tral gene but later ac-quired, opposite effectson regulation of bonegrowth in development.
Group 3, Small α/βCytokinesEGF receptor complexes.The epidermal growthfactor family of receptorsincludes EGFR, Erb2,Erb3, and Erb4. These re-ceptors are activated byEGF, TGFα, or some 10other ligands according totheir location. These re-ceptors are important intissue remodeling, andmutations in the receptorscan underlie several can-cers (117). Dimerizationof EGFRs occurs uponbinding two EGF mole-cules in a 2:2 complex.Two structures of thesecomplexes (118, 119)show a back-to-back asso-ciation of the EGFR:EGF1:1 complexes in which
conformational change in the receptors leads to their association(Fig. 14). The receptors are inherently three or four domain struc-tures, in which domains I and III (the thumb and fingers) clasp theEGF molecule, and the structural change relayed through domain IIof each receptor molecule, at the backs of the “hands,” leads to theirdimerization. This process depends on a dimerization loop of 17amino acids at the back of each of the domain II.
The structure of the extracellular domain of ErbB3 (120)suggests that deletion of domain IV of ErbB3 releases a kind ofauto-inhibition imposed by interactions between domain IV andthe dimerizing domains II, as if this intramolecular interactioncompetes with the dimerizing intermolecular domain II-domainII interactions.
Deletion of domain IV is accompanied by a reduction inbinding of EGF, supporting a reciprocal allosteric feedback inwhich lack of “dimerization-mimetic” binding of domain IVagainst domain II lowers the affinity for EGF between domains Iand III. Intracellularly, intermolecular phosphorylation of partic-ular tyrosines in the activation loops of the protein tyrosine ki-nase (PTK) domains opens them up to bind adenosine triphos-phate (ATP), whereupon they become active as tyrosine kinases.
The insulin receptor complex. The insulin receptor and itsrelated insulin-like growth factor (IGF-I and IGF-II) receptorsare unusual in presenting covalently preassembled α2β2oligomers that are inactive. Signaling is mediated by an insulin-
R E V I E W
NN
Type I
Ligandmonomer 2
Ligandmonomer 1
Type I
N N
N N
CC
CC
Type I Type I
A
B
Fig. 13. BMP-2 in 2:2 complex with one of its receptor chains, BRIA, labeled“Type I”. (A) A top view; (B) a side view. The signaling complex is a 2:2:2 as-sociation with two different receptors.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 13
induced conformationalchange within the pre-formedcomplex of two α subunits thatprovide for binding of insulin,and two β subunits that spanthe membrane and include in-tracellular tyrosine kinase do-mains. The two α chains aredisulfide cross-linked together,and each α chain is in turndisulfide-linked to a β chain.The insulin receptor is an ex-ample of a horizontal receptorthat has evolved vertical recep-tor characteristics; it is preas-sociated and, for the most part,regulates reversible changes incell metabolism. The three-di-mensional (3D) profile struc-ture of the complete covalentheterodimeric insulin receptorcomplex in the membrane,bound to a single insulin, wasvisualized to limited resolutionby electron cryomicroscopy (121, 122). The 3D profile wasthen fitted with available known high-resolution domain sub-structures to deduce the structure of the ligand-bound, activatedreceptor complex. The structure of a fragment of the homolo-gous IGF-IR that includes the two homologous L1 and L2 do-mains and the intervening cysteine-rich domain, comprisingabout 450 amino acids of the ectodomain of the α chain (123,124), and the imaging analysis of Fab fragments bound to spe-cific regions of the receptor (125), provide a sound basis for as-sessing and improving this model.
The insulin family is based on cysteine-rich, class III, α/βligands. α and β chains of the receptor complex bind insulin, andthe β chains each carry a single membrane-crossing sequenceand an intracellular kinase domain. These cytoplasmic domainshave intrinsic activity that can transfer the γ-phosphate of ATP totyrosine residues in the neighboring domain, once brought to-gether by the insulin-induced conformation change (121). Thebinding of insulin by the entire α2β2 complex is negatively co-operative, with one high-affinity site, and a second of low affini-ty (126, 127). Binding to a single αβ monomer occurs only withlow affinity and without cooperativity, implying that the four-chain complex is required to present the high-affinity site.
Peptides selected for binding to the insulin receptor, opti-mized by phage display, bound to any of three different sites onthe receptor (128). All competed with insulin, with Kd values inthe high nanomolar to low micromolar range. Some peptidesthat bound to site 1 alone activated the tyrosine kinase activityof the receptor, suggesting that site 1 corresponds to a regioninvolved in insulin-induced activation of the insulin receptor.Peptides that bound to site 2 or site 3 acted as antagonists inphosphorylation assays and assays of glucose incorporation intolipids in adiopocytes.
The structure of both the inactive intracellular insulin recep-tor tyrosine kinase domains (129) and the activated phosphory-lated forms, resolved to 1.9 Å (130, 131), define the changesthat activate the kinase. In the active state structure, the kinasedomains are dimerized in a well-ordered structure and bound to
a peptide substrate and an ATPanalog. This dimer shows howthe activation loop (the A-loop) of the kinase undergoesa major conformationalchange upon autophosphory-lation. Three tyrosines withinthe loop are phosphorylated:Tyr1158, Tyr1162 and Tyr1163.The resulting structuralchange then pulls the tyrosineresidues away from the activesite and so allows unrestrictedaccess of the kinase active siteto peptide substrates and ATP.Phosphorylated Tyr1163
(pTyr1163) plays the centralrole in determining the con-formation of the tri-phospho-rylated A-loop. pTyr1158 isthen fully exposed to solvent,suggesting a possible role ininteracting with other proteinsdownstream in the signaling
process. The structure of the IGF-IR kinase domain activated byphosphorylation shows that the degree of phosphorylation con-trols the degree of activation in successive stages (132, 133).
In the insulin receptor kinase, the peptide substrate forms anantiparallel β strand with the C-terminal portion of the activat-ing A-loop. The two methionine side chains in the YMXM sub-strate sequence are bound in two hydrophobic pockets on the C-terminal lobe of the kinase. The phosphotyrosine sites either en-hance catalytic efficiency of the kinase domain or provide a tar-get for recruitment of other signaling proteins downstream inthe signaling pathway. The structure thus reveals the molecularbasis for autophosphorylation of the insulin kinase and moregenerally suggests the basis for target specificity in the tyrosinekinases as well as the mechanism of phosphotransfer from ATP.
Receptor oligomerization in the family. Some of the earli-est biochemical evidence for the importance of receptor cluster-ing came from studies of the EGFR family (134). The receptorsfor erbB2 (HER2, Neu) are believed to form both homodimersand heterodimers with the EGF and heregulin receptors (erbB1or EGFR; and erbB3-4). A tremendous amount of biochemicaland cell biological evidence indicates that oligomerization iscritical for signaling; however, because of their size and com-plexity, it has been difficult to study these receptors structurallyand thereby discover their mode of oligomerization.
The structure of the entire HER3 EGFR (ERB3) (128) showsfour domains structurally homologous to those found in the type Iinsulin-like growth factor receptor (IGFR). Contacts between do-mains II and IV constrain the orientations of ligand-binding do-mains and provide a structural basis for understanding both mul-tiple-affinity forms of EGFRs and conformational changes in-duced in the receptor by ligand binding during signaling.
“Inside-out” receptors in the “RGD-activated” integrin fam-ilies as spring-loaded heterodimers. αβ heterodimeric integrinsprovide connections between cells and between cells and the ex-tracellular matrix. These proteins signal bidirectionally and arecoupled to intracellular signaling pathways through activation oftyrosine kinases and through connections to the cytoskeleton.
R E V I E W
NN
CC
Fig. 14. Group 3 “small α/β” cytokines. Dimerization of EGFRsoccurs on binding two EGF molecules to form a 2:2 complex in aback-to-back association of the EGFR-EGF 1:1 complexes inwhich conformational change in the receptors leads to their asso-ciation.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 14
The extracellular domains ofthe integrins bind an RGD se-quence in the ligand, config-ured as a β turn (135). The inte-grins are αβ heterodimers com-posed of two single-transmem-brane receptors in which the αchain is typically composed offour or five domains. The β do-main is composed of about fiveunique domains, followed byabout five EGF-like domains.Both chains end with a trans-membrane helix and a short in-tracellular platform for assem-bly of intracellular factors.These termini are seeminglysufficient to hold the α and βchains together (136). In bidi-rectional signaling, they relayinformation from the extracellu-lar environment of the cell tointracellular pathways (137),and in the reverse direction, in-tracellular connections to thesubmembranous cytoskeletonmodulate external binding tothe extracellular matrix.
Association of dimers in themembrane plane is sufficientto activate intracellular signaling by integrins (138). Other ki-nase-associated membrane proteins, called syndecans andtetraspans, sometimes augment the activation effect (139). Thecrystal structure of the αVβ3 integrin ectodomain (140) isswitched back on itself and is presumed to be the inactive con-formation. Upon binding the RGD sequences as would be pre-sented by a ligand, the integrins also undergo a large, spring-loaded extension of several external domains (141) that trans-form them from a low-affinity state to a high-affinity state forligand binding.
At the hairpin bend of the resting-state integrin β chain is adomain that in its structure resembles the α subunit of G pro-teins. This domain is tightly bound to a “seven-bladed propeller”domain on the integrin α chain that in its structure resembles aG protein β subunit (Fig. 15). These two domains each bind fibronectin (142). In structural terms, this extracellular interac-tion mimics the interaction between intracellular α and β sub-units of G proteins. In the integrin case, the G protein homologis not a GTPase; instead, the site corresponding to the GTP-binding site provides the invariant binding site at which the RGDsequence of the integrin ligand interacts with the integrin βchain. Binding of the RGD sequence is mediated by a Mg2+ met-al ion-binding site (a MIDAS domain). Dissociation of the Gβ-like and Gα-like domains triggers the response. In a curiousway, this pairing of domains that resemble intracellular matesthat transmit intracellular signals from the vertical receptors—but in the case of integrins occur on the outside of the cell—re-flects the bidirectional nature of integrin signaling. The bindingof GTP normally dissociates Gα proteins from the propeller do-main in Gβ. One of several proposals is that binding of the RGDligand may dissociate the integrin α and β chains, leading to the“switchblade” activation of the β chain (143).
SummaryThe structures of cytokinesand their receptors reveal a eu-karyotic system of intercellu-lar communication that is in-creasingly in focus, as re-vealed by the growing numberof signaling paradigms seen inthree-dimensional structuresof both the functional and theoff-state receptor complexes.Central principles are thatthere is control over lateral as-sociations of receptors in theplane of the membrane thatcan activate inside the cellcascades of intracellular en-zymes that are predominantlyprotein kinases. In general, thefunctionally important surfaceareas of the binding epitopeson cytokines span the range of700 to 1200 Å2.
The diversity of mecha-nisms relies on three princi-ples. First, many receptors aremaintained in some orderedoff-state in which their trans-membrane domains are heldapart. Second, binding of lig-
ands leads to activation by changing the way homo- or het-erodimers of receptors are formed as partners on the outsidesurface of the cell. Third, these structural changes in proximityand orientation of the outside domains are detected by associat-ed intracellular kinases whose actions are determined by theirproximity and orientation.
The diverse interactions that can result from different combi-nations of ligands and co-receptors allows integration of multi-ple incoming signals at the level of receptor activation. Uncov-ering the keys to this diversity in protein-protein interactions re-mains one of the most exciting challenges in modern biology,and it promises a rich field for intervention in human disease bytargeting of receptor-protein interactions.
References and Notes1. P. G. Strange, G-protein coupled receptors: Conformations and states.
Biochem. Pharmacol. 58, 1081-1088 (1999).2. T. Iiri, Z. Farfel, H. R. Bourne, G-protein diseases furnish a model for
the turn-on switch. Nature 394, 35-38 (1998).3. A. Krebs, C. Villa, P. C. Edwards, G. F. Schertler, Characterisation of an
improved two-dimensional p22121 crystal from bovine rhodopsin. J.Mol. Biol. 282, 991-1003 (1998).
4. K. Palczewski, T. Kumasaka, T. Hori, C. A. Behnke, H. Motoshima, B. A.Fox, I. Le Trong, D. C. Teller, T. Okada, R. E. Stenkamp, M. Yamamoto,M. Miyano, Crystal structure of rhodopsin: A G protein-coupled recep-tor. Science 289, 739-745 (2000).
5. C. Altenbach, K. Yang, D. L. Farrens, Z. T. Farahbakhsh, H. G. Kho-rana, W. L. Hubbell, Structural features and light-dependent changes inthe cytoplasmic interhelical E-F loop region of rhodopsin: A site-directedspin-labeling study. Biochemistry 35, 12470-12478 (1996).
6. C. Altenbach, J. Klein-Seetharaman, J. Hwa, H. G. Khorana, W. L.Hubbell, Structural features and light-dependent changes in the se-quence 59-75 connecting helices I and II in rhodopsin: A site-directedspin-labeling study. Biochemistry 38, 7945-7949 (1999).
7. J. Klein-Seetharaman, J. Hwa, K. Cai, C. Altenbach, W. L. Hubbell, H.G. Khorana, Single-cysteine substitution mutants at amino acid posi-tions 55-75, the sequence connecting the cytoplasmic ends of helices I
R E V I E W
N
1 domainCalf-1
Calf-2
Thigh domain
β-propeller
βTDβA-domain
CC
N
Fig. 15. Integrin signaling involves a large change in conforma-tion upon activation, which activates binding to the intracellularmatrix in a mechanism of bidirectional signaling. (A) The restingstate complex. (B) A model—made to match electron images—that indicates the extent and type of change involved when acti-vation is triggered to generate the binding site.

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 15
and II in rhodopsin: Reactivity of the sulfhydryl groups and their deriva-tives identifies a tertiary structure that changes upon light-activation.Biochemistry 38, 7938-44 (1999).
8. S. Subramaniam, M. Lindahl, P. Bullough, A. R. Faruqi, J. Tittor, D. Oester-helt, L. Brown, J. Lanyi, R. Henderson, Protein conformational changes inthe bacteriorhodopsin photocycle. J. Mol. Biol. 287, 145-61 (1999).
9. F. M. Hendrickson, F. Burkard, R. M. Glaeser, Structural characteriza-tion of the L-to-M transition of the bacteriorhodopsin photocycle. Bio-phys. J. 75, 1446-1454 (1998).
10. S. Subramaniam, R. Henderson, Molecular mechanism of vectorial pro-ton translocation by bacteriorhodopsin. Nature 406, 653-657 (2000).
11. T. Oka, H. Kamikubo, F. Tokunaga, J. K. Lanyi, R. Needleman, M.Kataoka, Conformational change of helix G in the bacteriorhodopsinphotocycle: Investigation with heavy atom labeling and x-ray diffraction.Biophys. J. 76, 1018-1023 (1999).
12. K. M. Ottemann, T. E. Thorgeirsson, A. F. Kolodziej, Y. K. Shin, D. E.Koshland Jr., Direct measurement of small ligand-induced conforma-tional changes in the aspartate chemoreceptor using EPR. Biochemistry 37, 7062-7069 (1998).
13. J. F. Bazan, Emerging families of cytokines and receptors. Curr. Biol. 3,603-606 (1993).
14. F. K. Chan, H. J. Chun, L. Zheng, R. M. Siegel, K.L. Bui, M. J. LenardoA domain in TNF receptors that mediates ligand-independent receptorassembly and signaling. Science 288, 2351-2354 (2000).
15. O. Livnah, E. A. Stura, S. A. Middleton, D. L. Johnson, L. K. Jolliffe, I. A.Wilson, Crystallographic evidence for preformed dimers of erythropoi-etin receptor before ligand activation. Science 283, 987-990 (1999).
16. I. Remy, I. A. Wilson, S. W. Michnick, Erythropoietin receptor activationby a ligand-induced conformation change. Science 283, 990-993(1999).
17. J. M. Rodriguez-Frade, A. J. Vila-Coro, A. M. de Ana, J. P. Albar, A. C.Martinez, M. Mellado, The chemokine monocyte chemoattractant protein-1 induces functional responses through dimerization of its re-ceptor CCR2. Proc. Natl. Acad. Sci. U.S.A. 96, 3628-3633 (1999).
18. J. Cohen, H, Altaratz, Y. Zick, U. Klingmuller, D. Neumann, Phosphory-lation of erythropoietin receptors in the endoplasmic reticulum by per-vanadate-mediated inhibition of tyrosine phosphatases. Biochem. J.327, 391-397 (1997).
19. A. Ullrich, J. Schlessinger, Signal transduction by receptors with tyro-sine kinase activity. Cell 61, 203-212 (1990).
20. L. S. Argetsinger, G. S. Campbell, X. Yang, B. A. Witthuhn, O. Silven-noinen, J. N. Ihle, C. Carter-Su, Identification of JAK2 as a growth hor-mone receptor-associated tyrosine kinase. Cell 74, 237-244 (1993).
21. A. M. De Vos, M. Ultsch, A. A. Kossiakoff, Human growth hormone andextracellular domain of its receptor: Crystal structure of the complex.Science 255, 306-312 (1992).
22. R. S. Syed, S. W. Reid, C. Li, J .C. Cheetham, K. H. Aoki, B. Liu, H.Zhan, T. D. Osslund, A. J. Chirino, J. Zhang, J. Finer-Moore, S. Elliott,K. Sitney, B. A. Katz, D. J. Matthews, J. J. Wendoloski, J. Egrie, R. M.Stroud, Efficiency of signalling through cytokine receptors depends criti-cally on receptor orientation. Nature 395, 511-516 (1998).
23. M. Aritomi, N. Kunishima, T. Okamoto, R. Kuroki, Y. Ota, K. Morikawa,Atomic structure of the GCSF-receptor complex showing a new cytokine-receptor recognition scheme. Nature 401, 713-717 (1999).
24. A. N. Plotnikov, J. Schlessinger, S. R. Hubbard, M. Mohammadi, Structural basis for FGF receptor dimerization and activation. Cell 98,641-650 (1999).
25. A. A. Kossiakoff, A. M. De Vos, Structural basis for cytokine hormone-receptor recognition and receptor activation. Adv. Protein Chem. 52,67-108 (1998).
26. B. C. Cunningham, M. Ultsch, A. M. De Vos, M. G. Mulkerrin, K. R.Clauser, J. A. Wells, Dimerization of the extracellular domain of the hu-man growth hormone receptor by a single hormone molecule. Science254, 821-825 (1991).
27. J. A. Wells, A. M. de Vos, Structure and function of human growth hor-mone: Implications for the hematopoietins. Annu. Rev. Biophys. Biomol.Struct. 22, 329-351 (1993).
28. B. C. Cunningham, J. A. Wells, Comparison of a structural and a func-tional epitope J. Mol. Biol. 234, 554-563 (1993) [published erratum ap-pears in J. Mol. Biol. 237, 513 (1994)].
29. T. Clackson, J. A. Wells, A hot spot of binding energy in a hormone-receptor interface. Science 267, 383-386 (1995).
30. H. W. Christinger, P. A. Elkins, Y. Sandowski, E. Sakal, A. Gertler, A. A.Kossiakoff, A. M. de Vos, Crystallization of ovine placental lactogen in a1:2 complex with the extracellular domain of the rat prolactin receptor.Acta Crystallogr. D Biol. Crystallogr. 54, 1408-1411 (1998).
31. M. J. Koury, M. C. Bondurant, The molecular mechanisms of erythro-poietin action. Eur. J. Biochem. 210, 649-663 (1992).
32. H. Youssoufian, G. Longmore, D. Neumann, A. Yoshimura, H. F.Lodish, Structure, function, and activation of the erythropoietin receptor.
Blood 81, 2223-2236 (1993).33. J. S. Philo, K. H. Aoki, T. Arakawa, L. O. Narhi, J. Wen, Dimerization of
the extracellular domain of the erythropoietin (EPO) receptor by EPO:One high-affinity and one low-affinity interaction. Biochemistry 35,1681-1691 (1996).
34. D. J. Matthews, R. S. Topping, R. T. Cass, L. B. Giebel, A sequentialdimerization mechanism for erythropoietin receptor activation. Proc.Natl. Acad. Sci. U.S.A. 93, 9471-9476 (1996).
35. H. Zhan, B. Liu, S. W. Reid, K. H. Aoki, C. Li, R. S. Syed, C. Karkaria,G. Koe, K. Sitney, K. Hayenga, F. Mistry, L. Savel, M. Dreyer, B. A.Katz, J. Schreurs, D. J. Matthews, J. C. Cheetham, J. Egrie, L. B.Giebel, R. M. Stroud, Engineering a soluble extracellular erythropoietinreceptor (EPObp) in Pichia pastoris to eliminate microheterogeneity,and its complex with erythropoietin. Protein Eng. 12, 505-513 (1999).
36. S. A. Middleton, D. L. Johnson, R. Jin, F. J. McMahon, A. Collins, J. Tul-lai, R. H. Gruninger, L. K. Jolliffe, L. S. Mulcahy, Identification of a criti-cal ligand binding determinant of the human erythropoietin receptor. Ev-idence for common ligand binding motifs in the cytokine receptor family.J. Biol. Chem. 271, 14045-14054 (1996).
37. A. Yoshimura, A. D. D’Andrea, H. F. Lodish, Friend spleen focus-form-ing virus glycoprotein gp55 interacts with the erythropoietin receptor inthe endoplasmic reticulum and affects receptor metabolism. Proc. Natl.Acad. Sci. U.S.A. 87, 4139-4143 (1990).
38. S. S. Watowich, A. Yoshimura, G. D. Longmore, D. J. Hilton, Y.Yoshimura, H. F. Lodish, Homodimerization and constitutive activationof the erythropoietin receptor. Proc. Natl. Acad. Sci. U.S.A. 89, 2140-2144 (1992).
39. S. S. Watowich, D. J. Hilton, H. F. Lodish, Activation and inhibition oferythropoietin receptor function, role of receptor dimerization. Mol. CellBiol. 14, 3535-3549 (1994).
40. G. D. Longmore, P. N. Pharr, H. F. Lodish, A constitutively activated ery-thropoietin receptor stimulates proliferation and contributes to transfor-mation of multipotent, committed nonerythroid and erythroid progenitorcells. Mol. Cell Biol. 14, 2266-2277 (1994).
41. H. Qiu, A. Belanger, H. W. Yoon, H. F. Bunn, Homodimerization re-stores biological activity to an inactive erythropoietin mutant. J. Biol.Chem. 273, 11173-11176 (1998).
42. H. Schneider, W. Chaovapong, D. J. Matthews, C. Karkaria, R. T. Cass,H. Zhan, M. Boyle, T. Lorenzini, S. G. Elliot, L. B. Giebel, Homodimer-ization of erythropoietin receptor by a bivalent monoclonal antibody trig-gers cell proliferation and differentiation of erythroid precursers. Blood89, 473-482 (1997).
43. N. C. Wrighton, F. X. Farrell, R. Chang, A. K. Kashyap, F. P. Barbone, L.S. Mulcahy, D. L. Johnson, R. W. Barrett, L. K. Jolliffe, W. J. Dower,Small peptides as potent mimetics of the protein hormone erythropoi-etin. Science 273, 458-464 (1996).
44. O. Livnah, E. A. Stura, D. L. Johnson, S. A. Middleton, L. S. Mulcahy, N.C. Wrighton, W. J. Dower, L. K. Jolliffe, I. A. Wilson, Functional mimicryof a protein hormone by a peptide agonist: The EPO receptor complexat 2.8 A. Science 273, 464-471 (1996).
45. O. Livnah, D. L. Johnson, E. A. Stura, F. X. Farrell, F. P. Barbone, Y.You, K. D. Liu, M. A. Goldsmith, W. He, C. D. Krause, S. Pestka, L. K.Jolliffe, I. A. Wilson, An antagonist peptide-EPO receptor complex sug-gests that receptor dimerization is not sufficient for activation. Nat.Struct. Biol. 5, 993-1004 (1998).
46. D. L. Johnson, F. X. Farrell, F. P. Barbone, F. J. McMahon, J. Tullai, D.Kroon, J. Freedy, R. A. Zivin, L. S. Mulcahy, L. K. Jolliffe, Amino-termi-nal dimerization of an erythropoietin mimetic peptide results in in-creased erythropoietic activity. Chem. Biol. 4, 939-950 (1997).
47. K. H. Pearce Jr., B. C. Cunningham, G. Fuh, T. Teeri, J. A. Wells,Growth hormone binding affinity for its receptor surpasses the require-ments for cellular activity. Biochemistry 38, 81-89 (1999).
48. S. Gobert, S. Chretien, F. Gouilleux, O. Muller, C. Pallard, I. Dusanter-Fourt, B. Groner, C. Lacombe, S. Gisselbrecht, P. Mayeux, Identifica-tion of tyrosine residues within the intracellular domain of the erythro-poietin receptor crucial for STAT5 activation. EMBO J. 15, 2434-2441(1996).
49. U. Klingmuller, S. Bergelson, J. G. Hsiao, H. F. Lodish, Multiple tyrosineresidues in the cytosolic domain of the erythropoietin receptor promoteactivation of STAT5. Proc. Natl. Acad. Sci. U.S.A. 93, 8324-8328(1996).
50. M. Socolovsky, I. Dusanter-Fourt, H. F. Lodish, The prolactin receptorand severely truncated erythropoietin receptors support differentiationof erythroid progenitors. J. Biol. Chem. 272, 14009-14012 (1997).
51. M. Socolovsky, A. E. Fallon, S. Wang, C. Brugnara, H. F. Lodish, Fetalanemia and apoptosis of red cell progenitors in Stat5a-/-5b-/- mice: a di-rect role for Stat5 in Bcl-XL induction. Cell 98, 181-191 (1999).
52. S. N. Constantinescu, T. Keren, M. Socolovsky, H. Nam, Y. I. Henis, H.F. Lodish, Ligand-independent oligomerization of cell-surface erythro-poietin receptor is mediated by the transmembrane domain. Proc. Natl.
R E V I E W

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 16
Acad. Sci. U.S.A. 98, 4379-4384 (2001).53. T. Naranda, R. I. Kaufman, J. Li, K. Wong, A. Boge, D. Hallen, K. Y.
Fung, M. W. Duncan, N. Andersen, A. Goldstein, L. Olsson, Activationof erythropoietin receptor through a novel extracellular binding site. Endocrinology 143, 2293-2302 (2002).
54. C. Guo, K. E. Georgiadis, S. K. Dower, D. Holowka, B. A. Baird, Inter-leukin 1 (IL-1) causes changes in lateral and rotational mobilities of IL-1type I receptors. Biochemistry 38, 1618-1625 (1999).
55. S. Damjanovich, L. Bene, J. Matko, A. Alileche, C. K. Goldman, S.Sharrow, T. A. Waldmann, Preassembly of interleukin 2 (IL-2) receptorsubunits on resting Kit 225 K6 T cells and their modulation by IL-2, IL-7,and IL-15: A fluorescence resonance energy transfer study. Proc. Natl.Acad. Sci. U.S.A. 94, 13134-13139 (1997).
56. P. Nagy, A. Jenei, A. K. Kirsch, J. Szollosi, S. Damjanovich, T. M. Jovin,Activation-dependent clustering of the erbB2 receptor tyrosine kinasedetected by scanning near-field optical microscopy. J. Cell Sci. 112,1733-1741 (1999).
57. P. Nagy, L. Bene, M. Balazs, W. C. Hyun, S. J. Lockett, N. Y. Chiang, F.Waldman, B. G. Feuerstein, S. Damjanovich, J. Szollosi, EGF-inducedredistribution of erbB2 on breast tumor cells: Flow and image cytomet-ric energy transfer measurements. Cytometry 32, 120-131 (1998).
58. J. H. Naismith, T. Q. Devine, B. J. Brandhuber, S. R. Sprang, Crystallo-graphic evidence for dimerization of unliganded tumor necrosis factorreceptor. J. Biol. Chem. 270, 13303-13307 (1995).
59. J. H. Naismith, B. J. Brandhuber, T. Q. Devine, S. R. Sprang, Seeingdouble: Crystal structures of the type I TNF receptor. J. Mol. Recognit.9, 113-117 (1996).
60. J. H. Naismith, T. Q. Devine, T. Kohno, S. R. Sprang, Structures of theextracellular domain of the type I tumor necrosis factor receptor. Structure 4, 1251-1262 (1996).
61. T. Kimura, H. Kaburaki, S. Miyamoto, J. Katayama, Y. Watanabe, Dis-covery of a novel thrombopoietin mimic agonist peptide. J. Biochem.(Tokyo) 122, 1046-1051 (1997).
62. S. E. Cwirla, P. Balasubramanian, D. J. Duffin, C. R. Wagstrom, C. M.Gates, S. C. Singer, A. M. Davis, R. L. Tansik, L. C. Mattheakis, C. M.Boytos, P. J. Schatz, D. P. Baccanari, N. C. Wrighton, R. W. Barrett, W.J. Dower, Peptide agonist of the thrombopoietin receptor as potent asthe natural cytokine. Science 276, 1696-1699 (1997).
63. S. J. McConnell, T. Dinh, M. H. Le, S. J. Brown, K. Becherer, K.Blumeyer, C. Kautzer, F. Axelrod, D. G. Spinella, Isolation of erythropoi-etin receptor agonist peptides using evolved phage libraries. Biol.Chem. 379, 1279-1286 (1998).
64. M. D. Ballinger, V. Shyamala, L. D. Forrest, M. Deuter-Reinhard, L. V.Doyle, J. Wang, L. Panganiban-Lustan, J. R. Stratton, G. Apell, J. A.Winter, M. V. Doyle, S. Rosenberg, W. M. Kavanaugh, Semirational de-sign of a potent, artificial agonist of fibroblast growth factor receptors.Nat. Biotechnol. 17, 1199-1204 (1999).
65. W. J. Fairbrother, H. W. Christinger, A. G. Cochran, G. Fuh, C. J.Keenan, C. Quan, S. K. Shriver, J. Y. Tom, J. A. Wells, B. C. Cunning-ham, Novel peptides selected to bind vascular endothelial growth factortarget the receptor-binding site. Biochemistry 37, 17754-17764 ((1998).
66. C. Weismann, M. H. Ultsch, S. H. Bass, A. M. de Voss, Crystal struc-ture of nerve growth factor in complex with the ligand-binding domain ofthe TrkA receptor. Nature 401, 184-188 (1999).
67. S. J. McConnell, V. J. Thon, D. G. Spinella, Isolation of fibroblastgrowth factor receptor binding sequences using evolved phage displaylibraries. Comb. Chem. High Throughput Screen. 2, 155-163 (1999).
68. M. Ferrer, S. C. Harrison, Peptide ligands to human immunodeficiencyvirus type 1 gp120 identified from phage display libraries. J. Virol. 73,5795-5802 (1999).
69. W. L. DeLano, M. H. Ultsch, A. M. de Vos, J. A. Wells, Convergent solu-tions to binding at a protein-protein interface. Science 287, 1279-1283(2000).
70. S. Y. reLai, W. Xu, S. L. Gaffen, K. D. Liu, G. D. Longmore, W. C. Greene,M. A. Goldsmith, The molecular role of the common γC subunit in signaltransduction reveals functional asymmetry within multimeric cytokine recep-tor complexes. Proc. Natl. Acad. Sci. U.S.A. 93, 231-235 (1996).
71. T. Hage, W. Sebald, P. Reinemer, Crystal structure of the interleukin-4/receptor α chain complex reveals a mosaic binding inter-face. Cell 97, 271-281 (1999).
72. B. J. Shen, T. Hage, W. Sebald, Global and local determinants for thekinetics of interleukin-4/interleukin-4 receptor α chain interaction. Abiosensor study employing recombinant interleukin-4-binding protein.Eur. J. Biochem. 240, 252-261 (1996).
73. Y. Wang, B. J. Shen, W. Sebald, A mixed-charge pair in human inter-leukin 4 dominates high-affinity interaction with the receptor α chain.Proc. Natl. Acad. Sci. U.S.A. 94, 1657-1662 (1997).
74. J. L. Zhang, I. Simeonowa, Y. Wang, W. Sebald, The high-affinity inter-action of human IL-4 and the receptor α chain is constituted by two in-dependent binding clusters. J. Mol. Biol. 315, 399-407 (2002).
75. J. L. Zhang, M. Buehner, W. Sebald, Functional epitope of common γchain for interleukin-4 binding. Eur. J. Biochem. 269, 1490-1499 (2002).
76. J. F. Reidhaar-Olson, J. A. De Souza-Hart, H. E. Selick, Identification ofresidues critical to the activity of human granulocyte colony-stimulatingfactor. Biochemistry 35, 9034-9041 (1996).
77. O. Hiraoka, H. Anaguchi, Y. Ota, Formation of 1:1 complex of the cy-tokine receptor homologous region of granulocyte colony-stimulatingfactor receptor with ligand. Biosci. Biotechnol. Biochem. 59, 2351-2354(1995).
78. O. Hiraoka, H. Anaguchi, A. Asakura, Y. Ota, Requirement for the im-munoglobulin-like domain of granulocyte colony-stimulating factor re-ceptor in formation of a 2:1 receptor-ligand complex. J. Biol. Chem.270, 25928-25934 (1995).
79. D. Chow, X. He, A. L. Snow, S. Rose-John, K. C. Garcia, Structure ofan extracellular gp130 cytokine receptor signaling complex. Science291, 2150-2155 (2001).
80. S. E. Ealick, W. J. Cook, S. Vijay-Kumar, M. Carson, T. L. Nagab-hushan, P. P. Trotta, C. E. Bugg, Three-dimensional structure of recom-binant human interferon-γ. Science 252, 698-702 (1991).
81. M. R. Walter, W. T. Windsor, T. L. Nagabhushan, D. J. Lundell, C. A.Lunn, P. J. Zauodny, S. K. Narula, Crystal structure of a complex be-tween interferon-γ and its soluble high-affinity receptor. Nature 376,230-235 (1995).
82. E. A. Bach, M. Aguet, R. D. Schreiber, The IFN γ receptor: A paradigm forcytokine receptor signaling. Annu. Rev. Immunol. 15, 563-591 (1997).
83. T. Li, T. Horan, T. Osslund, G. Stearns, T. Arakawa, Conformationalchanges in G-CSF/Receptor complex as investigated by isotope-editedFTIR spectroscopy. Biochemistry 36, 8849-8857 (1997).
84. D. J. Thiel, M. H. le Du, R. L. Walter, A. D’Arcy, C. Chene, M. Foun-toulakis, G. Garotta, F. K. Winkler, S. E. Ealick, Observation of an unex-pected third receptor molecule in the crystal structure of human interfer-on-γ receptor complex. Structure Fold Des. 8, 927-936 (2000).
85. M. Randal, A. A. Kossiakoff, The structure and activity of a monomericinterferon-γ:α-chain receptor signaling complex. Structure (Camb.) 9,155-163 (2001).
86. B. C. Jones, N. J. Logsdon, K. Josephson, J. Cook, P. A. Barry, M. R.Walter, Crystal structure of human cytomegalovirus IL-10 bound to sol-uble human IL-10R1. Proc. Natl. Acad. Sci. U.S.A. 99, 9404-9409(2002).
87. M. R. Walter, T. L. Nagabhushan, Crystal structure of interleukin 10 re-veals an interferon γ-like fold. Biochemistry 34, 12118-12125 (1995).
88. J. Finer-Moore, J. F. Bazan, J. Rubin, R. M. Stroud, Identification ofmembrane proteins and soluble protein secondary structural elements,domain structure, and packing arrangements by Fourier-transform am-phipathic analysis, In Prediction of Protein Structure and the Principlesof Protein Conformation, G. Fasman, Ed. (Plenum Press, New York,1989), pp. 719–759.
89. L. A. Tartaglia, D. V. Goeddel, Two TNF receptors. Immunol. Today. 13,151-153 (1992).
90. M. J. Eck, S. R. Sprang, The structure of tumor necrosis factor-α at2.6Å resolution. Implications for receptor binding. J. Biol. Chem. 264,17595-17605 (1989).
91. E. Y. Jones, D. I. Stuart, N. P. Walker, Structure of tumour necrosis fac-tor. Nature 338, 225-228 (1989).
92. J. H. Naismith, S. R. Sprang, Tumor necrosis factor receptor superfami-ly. J. Inflamm. 47, 1-7 (1995).
93. M. Rothe, S. C. Wong, W. J. Henzel, D. V. Goeddel, A novel family ofputative signal transducers associated with the cytoplasmic domain ofthe 75 kDa tumor necrosis factor receptor. Cell 78, 681-692 (1994).
94. Y. C. Park, V. Burkitt, A. R. Villa, L. Tong, H. Wu, Structural basis forself-association and receptor recognition of human TRAF2. Nature 398,533-538 (1999).
95. Y. Jiang, J. D. Woronicz, W. Liu, D. V. Goeddel, Prevention of constitu-tive TNF receptor 1 signaling by silencer of death domains. Science283, 543-546 (1999) [published erratum appears in Science 283, 1852(1999)].
96. J. H. Naismith, S. R. Sprang, Modularity in the TNF-receptor family.Trends Biochem. Sci. 23, 74-79 (1998).
97. S. Ye, Y. Luo, W. Lu, R. B. Jones, R. J. Linhardt, I. Capila, T. Toida, M.Kan, H. Pelletier, W. L. McKeehan, Structural basis for interaction ofFGF-1, FGF-2, and FGF-7 with different heparan sulfate motifs. Biochemistry 40, 14429-14439 (2001).
98. F. Uematsu, M. Kan, F. Wang, J. H. Jang, Y. Luo, W. L. McKeehan, Lig-and binding properties of binary complexes of heparin and im-munoglobulin-like modules of FGF receptor 2. Biochem. Biophys. Res.Commun. 272, 830-836 (2000).
99. A. N. Plotnikov, S. R. Hubbard, J. Schlessinger, M. Mohammadi, Crys-tal structures of two FGF-FGFR complexes reveal the determinants ofligand-receptor specificity. Cell 101, 413-424 (2000).
100. D. J. Stauber, A. D. DiGabriele, W. A. Hendrickson, Structural interactions of
R E V I E W

www.stke.org/cgi/content/full/sigtrans;2004/231/re7 Page 17
fibroblast growth factor receptor with its ligands. Proc. Natl. Acad. Sci.U.S.A. 97, 49-54 (2000).
101. J. Schlessinger, A. N. Plotnikov, O. A. Ibrahimi, A. V. Eliseenkova, B. K.Yeh, A. Yayon, R. J. Linhardt, M. Mohammadi, Crystal structure of aternary FGF-FGFR-heparin complex reveals a dual role for heparin inFGFR binding and dimerization. Mol. Cell 6, 743-750 (2000).
102. L. Pellegrini, D. F. Burke, F. von Delft, B. Mulloy, T. L. Blundell, Crystalstructure of fibroblast growth factor receptor ectodomain bound to lig-and and heparin. Nature 407, 1029-1034 (2000).
103. M. Mohammadi, J. Schlessinger, S. R. Hubbard, Structure of the FGFreceptor tyrosine kinase domain reveals a novel autoinhibitory mechanism. Cell 86, 577-587 (1996).
104. W. L. McKeehan, X. Wu, M. Kan, Requirement for anticoagulant hep-aran sulfate in the fibroblast growth factor receptor complex. J. Biol.Chem. 274, 21511-21514 (1999).
105. M. Kan, X. Wu, F. Wang, W. L. McKeehan, Specificity for fibroblastgrowth factors determined by heparan sulfate in a binary complex withthe receptor kinase. J. Biol. Chem. 274, 15947-15952 (1999).
106. Y. A. Muller, B. Li, H. W. Christinger, J. A. Wells, B. C. Cunningham, A.M. de Vos, Vascular endothelial growth factor: Crystal structure andfunctional mapping of the kinase domain receptor binding site. Proc.Natl. Acad. Sci. U.S.A. 94, 7192-7197 (1997).
107. C. Weismann, G. Fuh, H. W. Christinger, C. Eigenbrot, J. A. Wells, A.M. de Voss, Crystal Structure at 1.7Å Resolution of VEGF in complexwith domain 2 of the Flt-1 receptor. Cell 91, 695-704 (1997).
108. T. Kirsch, W. Sebald, M. K. Dreyer, Crystal structure of the BMP-2-BRIAectodomain complex. Nat. Struct. Biol. 7, 492-496 (2000).
109. P. J. Hart, S. Deep, A. B. Taylor, Z. Shu, C. S. Hinck, A. P. Hinck, Crys-tal structure of the human TβR2 ectodomain—TGF-β3 complex. Nat.Struct. Biol. 9, 203-208 (2002).
110. R. A. Love, R. M. Stroud, The crystal structure of α-bungarotoxin at2.5Å resolution: Relation to solution structure and binding to acetyl-choline receptor. Protein Eng. 1, 37-46 (1986).
111. P. A. Kosen, J. Finer-Moore, M. P. McCarthy, V. J. Basus, Structuralstudies of α-bungarotoxin. 3. Corrections in the primary sequence andX-ray structure and characterization of an isotoxic α-bungarotoxin. Biochemistry 27, 2775-2781 (1988).
112. R. M. Stroud, M. P. McCarthy, M. Shuster, Nicotinic acetylcholine recep-tor superfamily of ligand-gated ion channels. Biochemistry 29, 11009-11023 (1990).
113. M. Harel, R. Kasher, A. Nicolas, J. M. Guss, M. Balass, M. Fridkin, A. B.Smit, K. Brejc, T. K. Sixma, E. Katchalski-Katzir, J. L. Sussman, S.Fuchs, The binding site of acetylcholine receptor as visualized in the X-ray structure of a complex between α-bungarotoxin and a mimotopepeptide. Neuron 32, 265-275 (2001).
114. D. Fass, S. Blacklow, P. S. Kim, J. M. Berger, Molecular basis of familialhypercholesterolaemia from structure of LDL receptor module. Nature388, 691-693 (1997).
115. L. Gilboa, A. Nohe, T. Geissendorfer, W. Sebald, Y. I. Henis, P. Knaus,Bone morphogenetic protein receptor complexes on the surface of livecells: A new oligomerization mode for serine/threonine kinase receptors. Mol. Biol. Cell 11, 1023-1035 (2000).
116. A. Nohe, S. Hassel, M. Ehrlich, F. Neubauer, W. Sebald, Y. I. Henis, P.Knaus, The mode of bone morphogenetic protein (BMP) receptoroligomerization determines different BMP-2 signaling pathways. J. Biol.Chem. 277, 5330-5338 (2002).
117. Z. Lu, G. Jiang, P. Blume-Jensen, T. Hunter, Epidermal growth factor-induced tumor cell invasion and metastasis initiated by dephos-phorylation and downregulation of focal adhesion kinase. Mol. Cell Biol.21, 4016-4031 (2001).
118. H. Ogiso, R. Ishitani, O. Nureki, S. Fukai, M. Yamanaka, J. Kim, K.Saito, A. Sakamoto, M. Inoue, M. Shirouzu, S. Yokoyama, Crystal struc-ture of the complex of human epidermal growth factor and receptor ex-tracellular domains. Cell 110, 775-787 (2002).
119. T. P. Garrett, N. M. McKern, M. Lou, T. C. Elleman, T. E. Adams, G. O.Lovrecz, H. J. Zhu, F. Walker, M. J. Frenkel, P. A. Hoyne, R. N. Joris-sen, E. C. Nice, A. W. Burgess, C. W. Ward, Crystal structure of a trun-cated epidermal growth factor receptor extracellular domain bound totransforming growth factor α. Cell 110, 763-773 (2002).
120. H. S. Cho, D. J. Leahy, Structure of the extracellular region of HER3 re-veals an interdomain tether. Science 297, 1330-1333 (2002).
121. R. Z. Luo, D. R. Beniac, A. Fernandes, C. C. Yip, F. P. Ottensmeyer,Quaternary structure of the insulin-insulin receptor complex. Science285, 1077-1080 (1999).
122. F. P. Ottensmeyer, D. R. Beniac, R. Z. Luo, C. C. Yip, Mechanism oftransmembrane signaling: Insulin binding and the insulin receptor. Biochemistry 39, 12103-12112 (2000).
123. T. P. Garrett, N. M. McKern, M. Lou, M. J. Frenkel, J. D. Bentley, G. O.Lovrecz, T. C. Elleman, L. J. Cosgrove, C. W. Ward, Crystal structure ofthe first three domains of the type-1 insulin-like growth factor receptor.Nature 394, 395-399 (1998).
124. C. W. Ward, T. P. Garrett, N. M. McKern, M. Lou, L. J. Cosgrove, L. G. Spar-row, M. J. Frenkel, P. A. Hoyne, T. C. Elleman, T. E. Adams, G. O. Lovrecz,L. J. Lawrence, P. A. Tulloch, The three dimensional structure of the type Iinsulin-like growth factor receptor. Mol. Pathol. 54, 125-132 (2001).
125. P. A. Tulloch, L. J. Lawrence, N. M. McKern, C. P. Robinson, J. D. Bent-ley, L. Cosgrove, N. Ivancic, G. O. Lovrecz, K. Siddle, C. W. Ward, Single-molecule imaging of human insulin receptor ectodomain and itsFab complexes. J. Struct. Biol. 125, 11-18 (1999).
126. P. De Meyts, The structural basis of insulin and insulin-like growth fac-tor-I receptor binding and negative co-operativity, and its relevance tomitogenic versus metabolic signalling. Diabetologia 37 (suppl. 2), S135-S148 (1994).
127. C. T. Christoffersen, K. E. Bornfeldt, C. M. Rotella, N. Gonzales, H. Vissing,R. Shymko, M. J. ten Hoeve, J. Groffen, N. Heisterkamp, P. De Meyts, Negative cooperativity in the insulin-like growth factor-I receptor and achimeric IGF-I/insulin receptor. Endocrinology 135, 472-475 (1994).
128. R. C. Pillutla, K. C. Hsiao, J. R. Beasley, J. Brandt, S. Ostergaard, P. H.Hansen, J. C. Spetzler, G. M. Danielsen, A. S. Andersen, R. E. Bris-sette, M. Lennick, P. W. Fletcher, A. J. Blume, L. Schaffer, N. I. Gold-stein, Peptides identify the critical hotspots involved in the biological ac-tivation of the insulin receptor. J. Biol. Chem. 277, 22590-22594 (2002).
129. S. R. Hubbard, L. Wei, L. Ellis, W. A. Hendrickson, Crystal structure ofthe tyrosine kinase domain of the human insulin receptor. Nature 372,746-754 (1994).
130. S. R. Hubbard, Crystal structure of the activated insulin receptor tyro-sine kinase in complex with peptide substrate and ATP analog. EMBOJ. 16, 5572-5581 (1997).
131. S. R. Hubbard, Structural analysis of receptor tyrosine kinases. Prog.Biophys. Mol. Biol. 71, 343-358 (1999).
132. S. Favelyukis, J. H. Till, S. R. Hubbard, W. T. Miller, Structure and au-toregulation of the insulin-like growth factor 1 receptor kinase. Nat.Struct. Biol. 8, 1058-1063 (2001).
133. A. Pautsch, A. Zoephel, H. Ahorn, W. Spevak, R. Hauptmann, H. Nar,Crystal structure of bisphosphorylated IGF-1 receptor kinase: Insight in-to domain movements upon kinase activation. Structure (Camb.) 9,955-965 (2001).
134. Y. Schechter, L. Hernadez, J. Schlessinger, P. Cuatrecasas, Epidermalgrowth factor: Local aggregation of hormone receptor complexes is re-quired for activation. Nature 278, 835-838 (1979).
135. B. K. Blackburn, A., Lee, M. Baier, B. Kohl, A. G. Olivero, R. Mata-moros, K. D. Robarge, R. S. McDowell, From peptide to non-peptide. 3.Atropisomeric GPIIbIIIa antagonists containing the 3,4-dihydro-1H-1,4-benzodiazepine-2,5-dione nucleus. J. Med. Chem. 40, 717-729 (1997).
136. A. M. Weljie, P. M. Hwang, H. J. Vogel, Solution structures of the cyto-plasmic tail complex from platelet integrin α IIb- and β 3-subunits. Proc.Natl. Acad. Sci. U.S.A. 99, 5878-5883 (2002).
137. M. A. Schwartz, M. H. Ginsberg, Networks and crosstalk: Integrin sig-nalling spreads. Nat. Cell Biol. 4, E65-E68 (2002).
138. N. Hogg, B. Leitinger, Shape and shift changes related to the function ofleukocyte integrins LFA-1 and Mac-1. J. Leukoc. Biol. 69, 893-898 (2001).
139. A. Woods, J. R. Couchman, Integrin modulation by lateral association.J. Biol. Chem. 275, 24233-24236 (2000).
140. J. P. Xiong, T. Stehle, B. Diefenbach, R. Zhang, R. Dunker, D. L. Scott,A. Joachimiak, S. L. Goodman, M. A. Arnaout, Crystal structure of theextracellular segment of integrin αVβ3. Science 294, 339-345 (2001).
141. N. Beglova, S. C. Blacklow, J. Takagi, T. A. Springer, Cysteine-richmodule structure reveals a fulcrum for integrin rearrangement upon activation. Nat. Struct. Biol. 9, 282-287 (2002).
142. A. P. Mould, J. A. Askari, S. Aota, K. M. Yamada, A. Irie, Y. Takada, H. J.Mardon, M. J. Humphries, Defining the topology of integrin α5β1-fi-bronectin interactions using inhibitory anti-α5 and anti-β1 monoclonalantibodies. Evidence that the synergy sequence of fibronectin is recog-nized by the amino-terminal repeats of the α5 subunit. J. Biol. Chem.272, 17283-17292 (1997).
143. R. R. Hantgan, C. Paumi, M. Rocco, J. W. Weisel, Effects of ligand-mimeticpeptides Arg-Gly-Asp-X (X = Phe, Trp, Ser) on αIIbβ3 integrin conformationand oligomerization. Biochemistry 38, 14461-14474 (1999).
144. R.M.S. is supported by the National Institutes of Health (GM24485).
Citation: R. M. Stroud, J. A. Wells, Mechanistic diversity of cytokine re-ceptor signaling across cell membranes. Sci. STKE 2004, re7 (2004).
R E V I E W