
Meat batter proteins—Effect of chemical modification on structure
8
J Sci Food Agric 1995,68,451-464 Meat Batter Proteins-Effect of Chemical Modification on Structure A Gordon and S Barbut* Animal and Poultry Science, University of Guelph, Guelph, Ontario, Canada, N1G 2W1 (Received 14 June 1994; revised version received 6 December 1994; accepted 15 March 1995) Abstract: The effects of five chemicals (H,O,, /?-mercaptoethanol, EDTA, urea and Tween 80) on the structure of meat protein extracts from commercial-type poultry meat batters were studied. Disulphide bonds appeared to affect protein conformation by influencing the interactions in which hydrophobic residues were involved. The spectrophotometric data for EDTA suggested that one of the mechanisms by which it caused meat batter protein aggregation was by salt- bridge formation through sulphydryl residues. Urea caused exposure of the hydrophobic residues in the meat batters’ proteins. Tween 80 did not appear to directly affect the protein-protein interaction. Key words : meat proteins, fluorescence, UV difference spectra, chemical modifi- cation. INTRODUCTION Meat batters (ie comminuted muscle tissue, fat, water and salt) are multiphase systems (Hermansson 1986) in which proteins are present in three different phases : the protein matrix, the aqueous phase and the interfacial protein film (IPF) around fat globules (Swift et a1 1961; Jones 1984). The types and amounts of proteins present in each of these phases affect the stability (ie fat and water binding) and textural properties of the cooked product (Schut 1978; Montejano et a1 1984; Herma- nsson 1986). It has been reported that the levels of some of the major proteins in the aqueous phase are related to their levels in the protein matrix (Gordon and Barbut 1992a). Hence, information about the state of the pro- teins within a meat batter can be obtained by examin- ing the structure and type of proteins remaining in the aqueous phase after comminution. The conformation of the proteins in a system deter- mines the number and type of residues which are exposed to the environment and which later become involved in molecular interactions (Nakai 1983; Jian et al 1988). Specific structural alignments involving dis- ulphide bonds, hydrogen bonds and electrostatic or hydrophobic interactions affect protein functionality attributes such as emulsification, gelation and water- * To whom correspondence should be addressed. binding (Borejdo 1983; Li-Chan et a1 1984; Stark 1970). Protein conformation largely determines functionality. Hence, studies on the overall functionality of meat pro- teins, which have undergone chemical modification of their spatial conformation, can assist in understanding the molecular events involved in meat batter stabilisa- tion. The effects of chemical treatments on protein confor- mation can be studied using spectroscopic techniques (Means and Feeney 1971). Fluorimetry has been used quite frequently because of its high absolute sensitivity to changes in the molecular conformation of proteins, and its flexibility with regard to sample preparation (Kronman and Holmes 1971; Brown 1980). Measure- ments can be made with the use of molecular probes which bind to specific residues (Kato and Nakai 1980; Li-Chan et a1 1984; Wicker et a1 1986); alternatively, the intrinsic fluorescence of samples may be measured (Yamagishi et a1 1981; Ma and Harwalker 1988). Ultra- violet (UV) absorbance spectroscopy is also used, often in addition to fluorimetry (Yamagishi et a1 1981), because it gives specific information about the state of several chromophores within protein molecules (Brown 1980). There is very little published work on protein extracts from commercial-type meat batters and the ones which exist mainly discuss quantification of their protein content (Acton and Dick 1984; Whiting 1987, 1988; 457 J Sci Food Agric 0022-5142/95/$09.00 0 1995 SCI. Printed in Great Britain
Transcript of Meat batter proteins—Effect of chemical modification on structure

J Sci Food Agric 1995,68,451-464
Meat Batter Proteins-Effect of Chemical Modification on Structure A Gordon and S Barbut* Animal and Poultry Science, University of Guelph, Guelph, Ontario, Canada, N1G 2W1 (Received 14 June 1994; revised version received 6 December 1994; accepted 15 March 1995)
Abstract: The effects of five chemicals (H,O,, /?-mercaptoethanol, EDTA, urea and Tween 80) on the structure of meat protein extracts from commercial-type poultry meat batters were studied. Disulphide bonds appeared to affect protein conformation by influencing the interactions in which hydrophobic residues were involved. The spectrophotometric data for EDTA suggested that one of the mechanisms by which it caused meat batter protein aggregation was by salt- bridge formation through sulphydryl residues. Urea caused exposure of the hydrophobic residues in the meat batters’ proteins. Tween 80 did not appear to directly affect the protein-protein interaction.
Key words : meat proteins, fluorescence, UV difference spectra, chemical modifi- cation.
INTRODUCTION
Meat batters (ie comminuted muscle tissue, fat, water and salt) are multiphase systems (Hermansson 1986) in which proteins are present in three different phases : the protein matrix, the aqueous phase and the interfacial protein film (IPF) around fat globules (Swift et a1 1961; Jones 1984). The types and amounts of proteins present in each of these phases affect the stability (ie fat and water binding) and textural properties of the cooked product (Schut 1978; Montejano et a1 1984; Herma- nsson 1986). It has been reported that the levels of some of the major proteins in the aqueous phase are related to their levels in the protein matrix (Gordon and Barbut 1992a). Hence, information about the state of the pro- teins within a meat batter can be obtained by examin- ing the structure and type of proteins remaining in the aqueous phase after comminution.
The conformation of the proteins in a system deter- mines the number and type of residues which are exposed to the environment and which later become involved in molecular interactions (Nakai 1983; Jian et al 1988). Specific structural alignments involving dis- ulphide bonds, hydrogen bonds and electrostatic or hydrophobic interactions affect protein functionality attributes such as emulsification, gelation and water-
* To whom correspondence should be addressed.
binding (Borejdo 1983; Li-Chan et a1 1984; Stark 1970). Protein conformation largely determines functionality. Hence, studies on the overall functionality of meat pro- teins, which have undergone chemical modification of their spatial conformation, can assist in understanding the molecular events involved in meat batter stabilisa- tion.
The effects of chemical treatments on protein confor- mation can be studied using spectroscopic techniques (Means and Feeney 1971). Fluorimetry has been used quite frequently because of its high absolute sensitivity to changes in the molecular conformation of proteins, and its flexibility with regard to sample preparation (Kronman and Holmes 1971; Brown 1980). Measure- ments can be made with the use of molecular probes which bind to specific residues (Kato and Nakai 1980; Li-Chan et a1 1984; Wicker et a1 1986); alternatively, the intrinsic fluorescence of samples may be measured (Yamagishi et a1 1981; Ma and Harwalker 1988). Ultra- violet (UV) absorbance spectroscopy is also used, often in addition to fluorimetry (Yamagishi et a1 1981), because it gives specific information about the state of several chromophores within protein molecules (Brown 1980).
There is very little published work on protein extracts from commercial-type meat batters and the ones which exist mainly discuss quantification of their protein content (Acton and Dick 1984; Whiting 1987, 1988;
457 J Sci Food Agric 0022-5142/95/$09.00 0 1995 SCI. Printed in Great Britain

458 A Gordon, S Barbut
Gordon and Barbut 1992a). In addition, most of the studies on the structure of meat proteins have used pure proteins (Samejima et al 1981). Thus, the objective of this study was to investigate conformational differences between proteins from chemically modified, commercial-type meat batters using intrinsic fluores- cence and difference derivative UV absorption spectros- COPY.
MATERIALS AND METHODS
Batter preparation
Six batches (750 g each) of chicken breast meat (650 g kg-') and pork back fat (250 g kg-') batters were prepared (by comminution) with NaCl(25 g kg- ') and added water (100 g kg-') as previously described (Gordon and Barbut 199 1). The proximate composition (AOAC 1980) of the trimmed chicken breast meat was as follows: moisture 731 g kg-', fat 5 g kg-', protein 255 g kg-' ash 9 g kg-'; the composition of the pork back fat was: moisture 283 g kg-', fat 674 g g-', protein 41 g kg-', ash 2 g kg-'. One of the NaCl batters served as a control and the other five were treated with the following chemical agents: H,Oz (3 g litre- '), B-mercaptoethanol (B-ME-2.5 g litre- ') ethylenediaminetetraacetic acid (EDTAb-2 g litre-' urea (45 l i tre- '4.75 M) and Tween 80 (6.6 g litre-'). The levels used have been shown to affect meat protein conformation, as intended (Gordon and Barbut 1991, 1992b). All chemicals were obtained from Fisher Chemi- cals (Toronto, Ontario, Canada), except mercaptoetha- no1 (Sigma Chemicals, St Louis, MO, USA). The experiment was repeated three times and was based on a complete randomized block design.
Protein extraction and protein content determination
The rapid protein extraction procedure previously developed (Gordon and Barbut 1992a) was modified to incorporate a single extraction step (Gordon and Barbut 1995). Briefly, the meat batters were mixed 1 : 1 with the corresponding chemical solution for 30 s using a Stomacher Lab Blender (United Laboratories, London, UK). The slurry was centrifuged at 30000 x g for 20 min, 2 ml of the saline phase was diluted five times with 0.6 M NaCl, 10 mM phosphate buffer (pH 7.2) and filtered through a 0.45 pm filter (Millex-HV, Millipore, Bedford, MA, USA) to remove any remaining fat. The protein content of the extracts was determined at 595 nm using the Coomassie blue dye binding method (Peterson 1983).
Fluorescence emission spectroscopy
Fluorescence emission spectra were determined with a Shimadzu Spectrofluorimeter RF540 (Shimadzu Co,
Tokyo, Japan) equipped with data-processing capabil- ities. Protein extracts standardized to 2.5 pg/ml were excited at 280 nm and emission spectra were recorded from 280 to 430 nm at right angles to the excitation. The excitation slit width was 10 nm. Difference spectra were generated by subtracting the emission spectra of the control WaCl (25 g litre-') protein extracts] from each of those of the chemically treated meat batters.
Ultraviolet (UV) spectrophotometry
UV spectra were obtained using a Shimadzu UV Visible Recording Spectrophotometer UV-260 (Shimadzu Co, Tokyo, Japan). All samples were standardized to a con- centration of 125 pg/ml protein before their spectra were determined. Difference spectra were obtained by placing the treated and the control protein extracts in the sample and reference compartments, respectively, and scanning from 190 to 340 nm. Second-order deriv- atives were generated from the original difference spectra. The derivative wavelength difference shift in the major absorbance peaks, as a result of differentiation, was 2 nm.
RESULTS AND DISCUSSION
Protein profiles of meat batter extracts
The modified protein extraction procedure used here is very convenient and takes only a few hours (Gordon and Barbut 1995). It produced relatively clean fractions which could be used directly for spectrophotometry without a further clean-up step. It is important to note that this procedure is designed to recover proteins present in the aqueous phase of meat batters without extracting any additional proteins from meat. These extracts are therefore representative of the proteins in the aqueous phase of the meat batters.
The spectral properties of proteins reflect the primary attributes of the individual residues acting independent- ly of each other and are only affected by interactions with their interest neighbours in the amino acid sequence (Tanford 1968). Spectroscopic methods are therefore well suited to provide detailed information about the molecular environment of selected amino acids. Fluorimetric studies are very useful but, because interpretations involve considerable speculation, they should be supported by other techniques (Penzer 1980). UV absorption spectroscopy was also used because it provides specific information about the conformation of proteins and is therefore a suitable technique to com- plement fluorimetry (Balestrieri er a1 1978). Difference second-order derivative absorption spectroscopy was particularly useful for obtaining information about changes in the environment of aromatic amino acid residues.

Meat batter proteins 459
Fluorescence spectra
Fluorescence spectra were generated for each of the extracts from the chemically treated batters (Fig 1). Dif- ference fluorescence spectra were also generated, the values presented being the differences from the NaCl (control) treatment (Fig 2). Fluorimetry is based on the emission of energy quanta as photons of light in specific regions of the electromagnetic spectrum depending on the chromophore involved (Kronman and Holmes 1971 ; Bashford and Harris 1987). The sample is excited by incident light at a pre-determined wavelength and the proteins within it emit some of the incident energy as photons at specific wavelengths (A) depending on their amino acid composition and conformation. The major fluorescence peak (Amax) for most proteins is due to tryp- tophan (Trp) fluorescence and is located at 350 nm when the sample is in aqueous solution (Kronman and
Holmes 1971; Ma and Harwalker 1988). A minor peak in the region of 305-315nm may also be present due to tyrosine (Tyr) fluorescence (Kronman and Holmes 1971; Bashford and Harris 1987).
The NaCl batter aqueous phase protein extract had a fluorescence maximum at 360 nm with a relative inten- sity of 68% (Fig 1, line a). This indicates that the Trp peak in this treatment was shifted to 360 nm (from 350 nm). Shifts in fluorescence spectra to higher wavelengths are called ‘red shifts’ and indicate that the structure of the protein is more open. These results, therefore, show that NaCl does cause an opening of the structure of myofibrillar proteins. This has been shown in several other studies (Hamm 1970; Hermansson 1986; Lewis et al 1986; Belton et a1 1987).
The oxidation of free sulphydryls to form disulphide bonds caused a large reduction in the fluorescence intensity (FI) of the protein extracts from the meat
60
20
wavelength (nm) Fig 1. Fluorescence emission spectra of the protein extracts from chemically treated meat batters. (a) NaCl batter; (b) H,O,- treated batter; (c) fl-mercaptoethanol-treated batter; (d) EDTA-treated batter; (e) urea-treated batter; (0 Tween 80-treated batter.
100 I
.- 0 (0 c W C
-w .- W u L 2 Lo 0) I 0 3
L L -
60
20
I I I I I I I I
I
300 340 380 wavelength (nm)
Fig 2. Difference fluorescence spectra of the protein extracts from chemically treated meat batters. (i) H,O,-treated batter; (ii) j-mercaptoethanol-treated barrier; (iii) EDTA-treated batter; (iv) urea-treated batter; (v) Tween 80-treated batter.

460 A Gordon, S Barbut
batters (Fig 1, line b; Fig 2, line i) but does not affect the position of the I,,,. A reduction in the FI is known as ‘quenching’. Quenching of fluorescence is due to non- radiative mechanisms which convert some of the inci- dent energy to thermal (or vibrational) energy more effectively than it is converted to photons of light by the chromophores within the system (Penzer 1980). This gives a good indication of changes in the environment of specific chromophores although the relationship between the degree of quenching and the amount of conformational change is not linear (Bashford and Harris 1987). Hence, the results for the H,O,-treated batter suggest a greater quenching of the Trp fluores- cence. This may be because of the increased efficiency of the non-radiative mechanisms due to the greater number of disulphide bonds which were present in the proteins (Penzer 1980; Ma and Harwalkar 1988).
The protein extracts from the b-ME treatment pro- duced a fluorescence spectrum which was indicative of the reduction of disulphide bonds (Fig 1, line c). The L,,, was 360 nm with a relative intensity of 52%. The reduction in the FI was less than that which occurred in the oxidised batter (Fig 2, line ii versus line i) and may be due to quenching of the fluorescence of the hydro- phobic residues by the solvent (Penzer 1980) as opposed to quenching by other non-radiative means such as interactions with the disulphide bonds (Kronman and Holmes 1971). This could have been caused by the opening up of the structure of the proteins in this treat- ment as a result of disulphide bond reduction. However, because the reduction in intensity was relatively small and no significant shift of the emission maximum occurred (Fig 1, line c ; Fig 2, line ii), it appears that the number of disulphide bonds involved in the structure of these proteins is relatively small. Alternatively, it may be that the exposure of more hydrophobic regions fol- lowing disulphide bond reduction was followed by hydrophobic interactions which reduced the number of hydrophobic groups that were exposed to the environ- ment. This would correspond with the very diffuse gel matrix which was shown to be formed in raw meat batters treated with P-ME (Gordon and Barbut 1991).
The EDTA-treated batter produced an aqueous phase in which the fluorescence maximum was shifted to 354 nm, a lower wavelength (blue shift), and showed a small increase in intensity compared to the control (Fig 1, line d ; Fig 2, line iii). This may be due to a reduction in the quenching of the fluorescence of the hydrophobic residues in the protein extract from this batter and suggests that the hydrophobic residues may have been less exposed (Bashford and Harris 1987). Cheung (1969) showed that at high pH, the hydropho- bic sites in myosin become more polar. This could imply that the reverse occurs in low pH environments, which was the case in this treatment (pH 5.30 versus 5.90 of the control and the other chemical treatments) and could therefore lead to protein aggregation and a
more closed structure. Such a structure was shown (by TEM) to be formed in raw batters treated with EDTA (Gordon Barbut 1991). This is also supported by the major blue shift recorded in the fluorescence spectra (Figs 1 and 2).
Urea resulted in a protein extract in which there was a reduction in the FI but not significant shift in the Imax (Fig 1, line e). This implies that the hydrophobic pockets within the recovered proteins were more exposed than those in the control. Urea is known to disrupt protein structure by increasing the solubility of hydrophobic groups and disrupting hydrogen bonds (O’Riordan et a1 1989).
Several workers have shown that protein denatur- ation causes a reduction in the fluorescence intensity as a result of increased quenching of the exposed residues (Kronman and Holmes 1971; Yamagishi et a1 1981). Cheung (1969) found a reduction in the fluorescence of myosin in the presence of urea using a probe method. Ma and Harwalkar (1988) also reported similar observ- ations with oat globulin. However, they observed a sig- nificant red shift in the emission maxima which was not found here (Fig 2, line iv). Protein denaturation is gen- erally accompanied by a red shift in emission maxima (Kronman and Holmes 1971). It may be that this was not observed with the extracts from the urea treatment because most of the proteins (which initially had a higher percentage of exposed hydrophobic residues) would have remained in the protein matrix as part of the dense structure which has been shown to exist in the matrices of these batters (Gordon and Barbut 1991).
The Tween 80-treated batter produced a protein extract which showed a significant blue shift in the Imax of its fluorescence spectrum and a large increase in the FI compared to the control (Fig 1, line f; Fig 2, line v). The Imax was shifted from 360 to 350 nm and the FI was 98%. This indicates a drastic change in the struc- ture of the proteins recovered from this batter. This sug- gests that the hydrophobic groups in these proteins (and therefore in some of the matrix proteins) were removed from the polar environment in the extract (ie ‘hidden’). The increase in the fluorescence intensity was probably due to the mechanism (previously discussed) of a reduction in fluorescence quenching due to confor- mational changes which favour such an increase (Penzer 1980; Yamagushi et a1 1981). This is consistent with the highly aggregated microstructure of the raw meat batters treated with Tween 80, as reported by Gordon and Barbut (1991).
Difference UV spectra and difference second derivative UV spectra
Difference UV spectra were generated for the five chemically treated batters based on the control (NaCI)

Meat batter proteins 461
"I
I - . m . i . i . m s i r
WAVELENGTH
(nm)
A=H202
B=Mercaptoethanol
C-EDTA
D=Urea
E=Tween 80
.p;*.. ,.' ...A. , ... ........ ....., * * a . . *. . .. *.
I 1 I I o ~ y m a m
u u
-
Fig 3. Difference UV absorption spectra for protein extracts from chemically treated meat batters. (a) H,O,-treated batter; (b) B-mercaptoethanol-treated batter; (c) EDTA-treated batter; (d) urea-treated batter; (e) Tween 80-treated batter. Difference spectra
were determined versus the control (NaCI).
treatment (Fig. 3). Difference spectra give an idea of changes occurring in the environment of certain resi- dues, especially the aromatic amino acids (Yamagishi et a1 1981). The net sign of the spectra is a measure of the extent to which the hydrophobic areas are exposed: a positive curve indicates increased exposure while a negative (downward) curve indicates a more closed structure (Brown 1980). The location of maxima can be used to identify which residues are involved in the changes observed in the spectra. Absorptions in the range of 240-270 nm are generally dominated by phenylalanine (Phe) although Tyr has some strong absorption bands in this region (Yamagishi et a1 1981). Phe has maxima between 250 and 270 nm, notably at 253, 259 and 265 nm (Balestrieri et a1 1978; Ma and Harwalkar 1988); Tyr between 270 and 300 nm, notably at 274, 280 and 288 nm (Brown 1980; Yamagishi et a1 1981); and Trp has a characteristic absorption maximum at 291-292 nm which is due to the transfer of its indole chromophore from the interior of a protein into an aqueous environment (Yamagishi et a1 1981; Ma and Harwalkar 1988). Other absorption maxima of interest are those of cysteine (Cys) at 235 nm and cystine at 250 nm (Brown 1980).
The difference second-order derivative spectra for each of the chemical treatments are shown in Fig 4. Dif-
ferences in the derivative difference intensity (d2AA/dA2; where A is absorbance) are an indication of differences in the degree of exposure between residues (Ma and Harwalkar 1988). Because of the differentiation process, peaks (maxima) in the original absorption spectra appear as troughs (minima) in the second order deriv- ative spectra and troughs appear as peaks (Balestrieri et a1 1978).
The difference spectrum for the H,O,-treated meat batter (Fig 3, line a) shows an increase in intensity which was mainly due to a major peak at 223 nm. A minor peak at 253 nm was also evident. The locations of peak maxima were ascertained from the first order derivative spectra of these difference spectra. The peak at 253 nm was most probably due to the increased exposure of Phe residues although Cys may also show a weak absorption in this range. The second order deriv- ative spectrum of the H,O,-treated sample showed a large peak at 234 nm along with minor troughs at 251- 253 nm, 285 and 291 nm (Fig 4, line a). This confirmed that there was a slight increase in the exposure of the Phe residues and also showed that the hydrophobic regions involving other amino acids were also slightly more exposed in this treatment. The very strong peak for Cys indicated a large reduction in the exposure of this amino acid in the oxidised batter as compared to
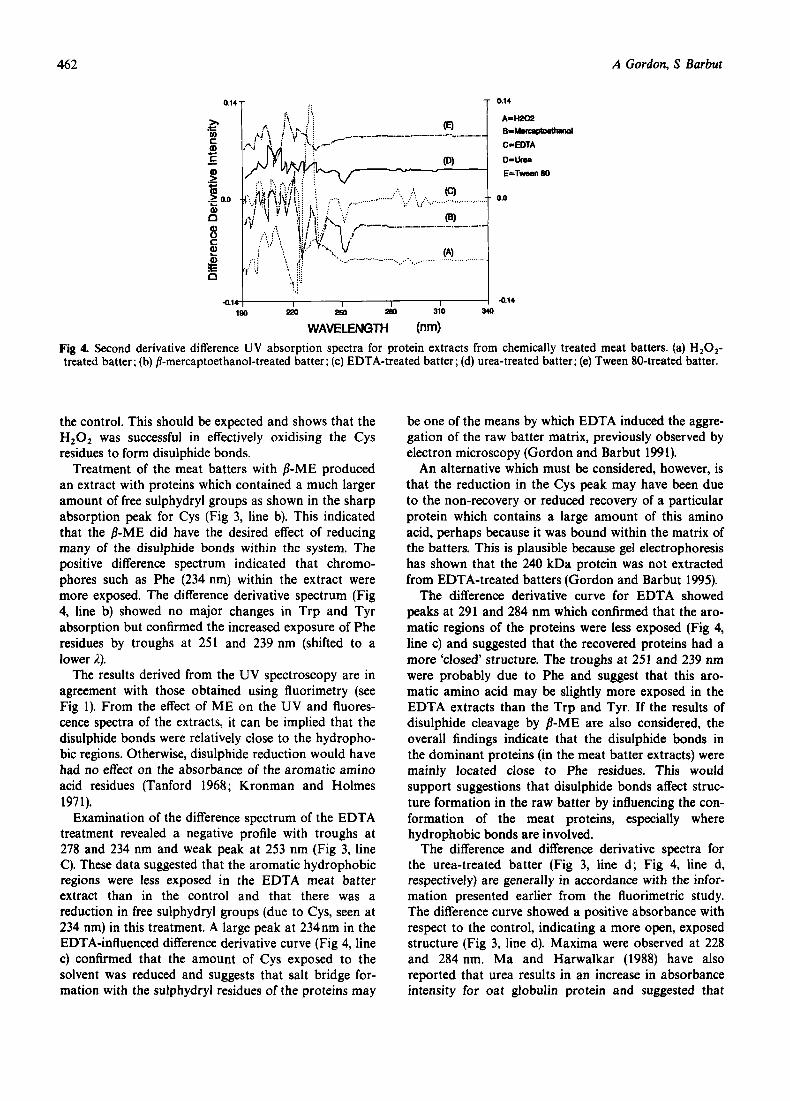
462 A Gordon, S Barbut
E v) C Q C
a, .2 la a,
Q 0 K
Q
c. - 4-
.g PO n
!!! E n
0.0
-0.14 1 I -0.14 I I I I 190 220 250 aB0 310 340
WAVELENGTH (nm) Fig 4. Second derivative difference UV absorption spectra for protein extracts from chemically treated meat batters. (a) H,O,- treated batter; (b) /I-mercaptoethanol-treated batter; (c) EDTA-treated batter; (d) urea-treated batter; (e) Tween 80-treated batter.
the control. This should be expected and shows that the H,0 , was successful in effectively oxidising the Cys residues to form disulphide bonds.
Treatment of the meat batters with /?-ME produced an extract with proteins which contained a much larger amount of free sulphydryl groups as shown in the sharp absorption peak for Cys (Fig 3, line b). This indicated that the /?-ME did have the desired effect of reducing many of the disulphide bonds within the system. The positive difference spectrum indicated that chromo- phores such as Phe (234 nm) within the extract were more exposed. The difference derivative spectrum (Fig 4, line b) showed no major changes in Trp and Tyr absorption but confirmed the increased exposure of Phe residues by troughs at 251 and 239 nm (shifted to a lower i).
The results derived from the UV spectroscopy are in agreement with those obtained using fluorimetry (see Fig 1). From the effect of ME on the UV and fluores- cence spectra of the extracts, it can be implied that the disulphide bonds were relatively close to the hydropho- bic regions. Otherwise, disulphide reduction would have had no effect on the absorbance of the aromatic amino acid residues (Tanford 1968; Kronman and Holmes 197 1).
Examination of the difference spectrum of the EDTA treatment revealed a negative profile with troughs at 278 and 234 nm and weak peak at 253 nm (Fig 3, line C). These data suggested that the aromatic hydrophobic regions were less exposed in the EDTA meat batter extract than in the control and that there was a reduction in free sulphydryl groups (due to Cys, seen at 234 nm) in this treatment. A large peak at 234nm in the EDTA-influenced difference derivative curve (Fig 4, line c) confirmed that the amount of Cys exposed to the solvent was reduced and suggests that salt bridge for- mation with the sulphydryl residues of the proteins may
be one of the means by which EDTA induced the aggre- gation of the raw batter matrix, previously observed by electron microscopy (Gordon and Barbut 1991).
An alternative which must be considered, however, is that the reduction in the Cys peak may have been due to the non-recovery or reduced recovery of a particular protein which contains a large amount of this amino acid, perhaps because it was bound within the matrix of the batters. This is plausible because gel electrophoresis has shown that the 240 kDa protein was not extracted from EDTA-treated batters (Gordon and Barbut 1995).
The difference derivative curve for EDTA showed peaks at 291 and 284 nm which confirmed that the aro- matic regions of the proteins were less exposed (Fig 4, line c) and suggested that the recovered proteins had a more 'closed' structure. The troughs at 251 and 239 nm were probably due to Phe and suggest that this aro- matic amino acid may be slightly more exposed in the EDTA extracts than the Trp and Tyr. If the results of disulphide cleavage by /?-ME are also considered, the overall findings indicate that the disulphide bonds in the dominant proteins (in the meat batter extracts) were mainly located close to Phe residues. This would support suggestions that disulphide bonds affect struc- ture formation in the raw batter by influencing the con- formation of the meat proteins, especially where hydrophobic bonds are involved.
The difference and difference derivative spectra for the urea-treated batter (Fig 3, line d ; Fig 4, line d, respectively) are generally in accordance with the infor- mation presented earlier from the fluorimetric study. The difference curve showed a positive absorbance with respect to the control, indicating a more open, exposed structure (Fig 3, line d). Maxima were observed at 228 and 284 nm. Ma and Harwalkar (1988) have also reported that urea results in an increase in absorbance intensity for oat globulin protein and suggested that

Meat batter proteins 463
this was the result of the more open structure of the denatured protein.
The main feature of the derivative curve in the spec- tral range of interest was a trough at 245 nm (Fig 4, line d). This indicated that Phe residues were at least slightly more exposed by the urea than in the control treatment. As noted earlier, the results for the urea extract may be due to the non-recovery of most of the ‘denatured’ pro- teins from the meat batters since they may already have participated in structure formation (Gordon and Barbut 1991).
Tween 80 produced a meat batter extract in which the proteins had a less open structure than those from NaCl treatment (Fig 3, line e). The overall absorption profile was negative with 2 major troughs at 223 and 281 nm. The trough at 281 was probably due to Tyr absorption and indicated Tyr was less accessible to the solvent in these extracts. The derivative difference spec- trum (Fig 4, line e) showed no major changes in the hydrophobic residues compared to the control but had a trough at 235 nm. This may suggest that more free sulphydryl groups are available in the proteins within this extract as compared to the control (NaCl) extract. The results agree with the findings of the fluorimetric examination of this extract.
In summary, fluorimetric and difference UV spectro- photometric studies showed that proteins recovered from the aqueous phase of chemically treated meat batters showed conformations suggesting that the treat- ments applied had the expected effects on the batters. It appears that disulphide bonds are not directly involved in raw batter formation. They appear to influence protein conformation especially with respect to the hydrophobic residues. They affect the strength of the interactions in which these residues are involved during raw batter preparation, thereby influencing the struc- ture formed. The UV absorption curves for EDTA suggest that one of the mechanisms by which it causes meat batter protein aggregation is salt-bridge formation through the sulphydryl residues. For urea, spectro- photometric data indicate that it may act to facilitate the formation of a meat batter gel matrix by exposing the hydrophobic residues which interact to form a cohe- sive structure. Hence, little of the proteins containing exposed hydrophobic residues were present in the aqueous phase.
REFERENCES
Acton J C, Dick R L 1984 Protein-protein interactions in pro- cessed meats. Recip Meat Conf Proc 37 36-43.
AOAC 1980 OQicial Methods of Analysis (13th edn). Associ- ation of Official Analytical Chemists, Washington, DC, USA.
Balestrieri C, Colonna G, Giovane A, Irace G, Servillo L 1978 Second derivative spectroscopy of proteins. A method for
the quantitative determination of aromatic amino acid resi- dues in proteins. Eur J Biochem 90 433-440.
Bashford C L, Harris D A 1987 Spectrophotometry and Spectrojluorimetry : A Practical Approach. IRL Press Ltd, Eynsham, UK.
Belton P S, Packer K J, Southon T E 1987 35C1 nuclear mag- netic resonance studies of the interaction of chloride ions with meat in the presence of tripolyphosphate. J Sci Food Agric 40,267-275.
Borejdo J 1983 Mapping of hydrophobic sites on the surface of myosin and its fragments. Biochem 22 1182-1 187.
Brown S B 1980 Ultraviolet and visible spectroscopy. In An Introduction to Spectroscopy for Biochemists, ed Brown S. B. Academic Press Inc, London, UK, pp 14-69.
Cheung H C 1969 Conformation of myosin. Effects of sub- strates and modifiers. Biochem Biophys Acta 194 478-485.
Gordon A, Barbut S 1991 Effect of chemical modification on the microstructure of raw meat batters. Food Struct 10 241- 253.
Gordon A, Barbut S 1992a The effect of chloride salts on protein extraction and interfacial protein film formation in meat batters. J Sci Food Agric 58 227-238.
Gordon A, Barbut S 1992b Effect of chemical modification on the stability, texture and microstructure of cooked meat batters microstructure. Food Struct 11 133-142.
Gordon A, Barbut S 1995 Meat batters: effect of chemical modifications on protein recovery and functionality. Food Res Int (in press).
Hamm R 1970 Interactions between phosphates and meat proteins. In Symposium: Phosphates in Food Processing, eds Deman J M & Melnychyn P. AVI Publishing Co, West- port, CT, USA.
Hermansson A M 1986 Water and fat holding. In Functional Properties of Food Macromolecules, eds Mitchell J R & Ledward D A. Elsevier Publishing Co, London, UK, pp
Jiang S T, Hwang D C, Chen C S 1988 Effect of storage tem- peratures on the formation of disulphides and denaturation of milkfish actomyosin (Chanos chanos). J Food Sci 53
Jones K W 1984 Protein-lipid interactins in processed meats. Proc Ann Recip Meat Conf 37 52-57.
Kato A, Nakai S 1980 Hydrophobicity determined by a fluo- rexcent probe method and its correlation with surface properties of proteins. Biochem Biophys Acta 624 13-20.
Kronman M J, Holmes L G 1971 The fluorescence of native, denatured and reduced-denatured proteins. Photochem Pho- tobiol 14 113-134.
Lewis D F, Groves K H M, Holgate J H 1986 Action of poly- phosphates in meat products. Food Microstruct 5 53-62.
Li-Chan E, Nakai S, Wood D F 1984 Hydrophobicity and solubility of meat proteins and their relationship to emul- sifying properties. J Food Sci 49 345-350.
Ma C, Harwalkar V R 1988 Study of thermal denaturation of oat globulin by ultraviolet and fluorescence spectroscopy. J Agric Food Chem 36 155-160.
Means G E, Feeney R E 1971 In: Chemical ModiJcation of Proteins. Holden Day Inc, San Francisco, CA, USA, pp
Montejano J G, Hamann D D, Lanier T C 1984 Thermally induced gelatin of selected comminuted muscle systems- Rheological changes during processing, final strengths and microstructure. J Food Sci 49 1496-1505.
Nakai S 1983 Structure-function relationships of food pro- ducts with an emphasis on the importance of food hydro- phobicity. J Agric Food Chem 31 676-682.
ORiordan D, Mulvihill D M, Morriddey P A, Kinsella J E 1989 Study of the molecular forces involved in the gelation
273-314.
1333-1335,1386.
4-50.

464 A Gordon, S Barbut
of plasma proteins at alkaline pH. J Food Sci 54, 1202- 1205.
Penzer G 1980 Molecular emission spectroscopy (fluorescence and phosphorescence). In : An Introduction to Spectroscopy for Biochemists, ed Brown S B. Academic Press Inc, London, UK, pp 70-1 14.
Peterson G L 1983 Determination of total protein. Meth Enzymol91 95-119.
Samejima K, Ishioroshi M, Yasui T 1981 The relative role of the head and tail portions of the molecule in the gelation of myosin. J Food Sci 46 1412-1418.
Schut J 1978 Basic meat emulsion technology. In Meat Industry Res Conf Proc p. 1-15.
Stark G R 1970 Recent developments in chemical modifi- cation and sequential degradation of proteins. Adv Prot Chem 24 261-308.
Swift C E, Lockett C, Fryar A J 1961 Comminuted meat
emulsions. The capacity of meat for emulsifying fat. Food Technoll5 468-473.
Tanford C 1968 Protein Deneturation, Part C. Adu Protein Chem 24 2-97.
Whiting R C 1987 Influence of various salts and water soluble compounds on the water and fat exudation and gel strength of meat batters. J Food Sci 52 1130-1 158.
Whiting R C 1988 Solute-protein interactions in a meat batter. Recipr Meat Conf Proc 41 53-56.
Wicker L, Lanier T C, Hamann D D, Akahane T 1986 Thermal transitions in myosin-ANS fluorescence and gel rigidity. J Food Sci 51 154-1543, 1560.
Yamagishi T, Yamuchi F, Shibasaki K 1981 State of aromatic amino acid residues in soybean 11s globulin heated in the presence of n-ethylmaleimide by derivative spectrophotom- etry. Agric Biol Chem 45 459-467.


















