
Mar. Vol. American for in U.S.A. Isolation and ...iai.asm.org/content/7/3/315.full.pdf · Isolation...
7
INFZCTION AND IMMUNITY, Mar. 1973, p. 315-321 Copyright © 1973 American Society for Microbiology Vol. 7, No. 3 PXrinted in U.S.A. Isolation and Characterization of the Antigen of Neisseria gonorrhoeae MICHAEL A. APICELLA AND JAMES C. ALLEN Department of Medicine, State Urniversity of New York at Buffalo, Buffalo, New York 14215 Received for publication 21 August 1972 The antigen of Neisseria gonorrhoeae has been isolated from the alkaline- extracted gonococcal endotoxin by ion exchange, molecular sieve, and powder block electrophoretic chromatography. Hemagglutination inhibition studies indi- cate that the preparation is essentially free of common enterobacterial and gono- coccal a antigen. Analysis of the isolated antigen by immunodiffusion and acryl- amide gel electrophoresis reveal only one detectable component. Chemical studies indicate that the antigen is an acidic glycoprotein composed primarily of four major amino acids: alanine, glutamic acid, glycine, and proline. The antigen has an s5o,w of 8.55, and spectral analysis in the ultraviolet and visible range reveals a single absorption peak at 217 nm. During the past 10 years the spread of gonococ- cal infections has reached epidemic proportions. This has renewed interest in the isolation of specific gonococcal antigens for the detection of infection in symptomatic and asymptomatic individuals. Maeland defined two major antigenic determinants in the alkaline-extracted endotoxin of Neisseria gonorrhoeae (17, 18, 21). These determinants have been called the a and ,B determinants. It has been assumed that they arise from gonococcal endotoxin and exist as part of a single molecule (19, 20). The a determinant is considered to be a polysaccharide and has serological multispecificity (22, 23). It is periodate sensitive but Pronase resistant. In the gonococcal alkaline extract which has been used in human serological tests, the principal determinant is the ,B antigen (18). This determinant is assumed to be a protein since it is Pronase sensitive and per- iodate resistant. In addition, it has been demon- strated that the ,B determinant is resistant to trypsin digestion (18). Serological testing with this ,3-antigen-rich extract has been unreliable because of a high rate of false positive and nega- tive reactions (15, 17). In order to standardize this test system and to evaluate the specificity of the 13 antigen in the human serological response to the gonococcus, we undertook the isolation of this antigen from the crude extract. In this paper, we will describe the isolation and chemical char- acteristics of 13 antigen of N. gonorrhoeae. MATERIALS AND METHODS Organisms. N. gonorrhoeae was obtained from fresh patient isolates and grown for antigen prepa- ration within 72 h of initial isolation. All organisms were unequivocally N. gonorrhoeae by gram stain, cultural characteristics, oxidase reactivity, and sugar fermentations. Organisms were maintained on chocolate agar at 35 C in a moist CO2 incubator and preserved by lyophilization in 5% bovine serutn albumin-5% monosodium glutamate. Antisera. Antisera to the whole gonococcus were produced by two injections of 1 ml of 109 organisms emulsified 1:2 with complete Freund adjuvant in the posterior cervical region of male albino rabbits at 10-day intervals. On days 21 and 23 after the onset of immunization, the animals were injected intravenously with 0.1 ml of 109 organisms. The rabbits were bled for antisera 7 days later. The organisms used for production of antisera were collected from 20 fresh isolates either by scraping chocolate plates with a glass spatula or by growth in supplemented Mueller- Hinton broth. Organisms were washed three times in distilled water before use as antigens. Antisera were also produced to purified ,B-antigen prepara- tions in rabbits. This was accomplished by footpad injection of 250 Ag of antigen emulsified in an equal volume of Freund adj uvant at weekly intervals for 28 days. Animals were bled for anti- body 7 days after the final injection. Growth of N. gonorrhoeae in liquid me- dium. Organisms were grown for antigen isolation in 2-liter screw-capped culture flasks in 1 liter of Mueller-Hinton broth containing modified Lang- ford supplement. Attempts were made to insure that all organisms were colonial type I or II of Kellogg et al. (13) before being used as iilocula in the broth. Multiple vials of fresh individual (type I or II) isolates were lyophilized. Continued availability of these colonial forms from the same organism for growth in liquid medium was main- tained by reconstitution of these lyophilized 315 on July 27, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Mar. Vol. American for in U.S.A. Isolation and ...iai.asm.org/content/7/3/315.full.pdf · Isolation...

INFZCTION AND IMMUNITY, Mar. 1973, p. 315-321Copyright © 1973 American Society for Microbiology
Vol. 7, No. 3PXrinted in U.S.A.
Isolation and Characterization of the Antigen ofNeisseria gonorrhoeae
MICHAEL A. APICELLA AND JAMES C. ALLENDepartment of Medicine, State Urniversity of New York at Buffalo, Buffalo, New York 14215
Received for publication 21 August 1972
The antigen of Neisseria gonorrhoeae has been isolated from the alkaline-extracted gonococcal endotoxin by ion exchange, molecular sieve, and powderblock electrophoretic chromatography. Hemagglutination inhibition studies indi-cate that the preparation is essentially free of common enterobacterial and gono-coccal a antigen. Analysis of the isolated antigen by immunodiffusion and acryl-amide gel electrophoresis reveal only one detectable component. Chemical studiesindicate that the antigen is an acidic glycoprotein composed primarily of fourmajor amino acids: alanine, glutamic acid, glycine, and proline. The antigen hasan s5o,w of 8.55, and spectral analysis in the ultraviolet and visible range reveals asingle absorption peak at 217 nm.
During the past 10 years the spread of gonococ-cal infections has reached epidemic proportions.This has renewed interest in the isolation ofspecific gonococcal antigens for the detection ofinfection in symptomatic and asymptomaticindividuals. Maeland defined two major antigenicdeterminants in the alkaline-extracted endotoxinof Neisseria gonorrhoeae (17, 18, 21). Thesedeterminants have been called the a and ,Bdeterminants. It has been assumed that they arisefrom gonococcal endotoxin and exist as part of asingle molecule (19, 20). The a determinant isconsidered to be a polysaccharide and hasserological multispecificity (22, 23). It is periodatesensitive but Pronase resistant. In the gonococcalalkaline extract which has been used in humanserological tests, the principal determinant is the,B antigen (18). This determinant is assumed to bea protein since it is Pronase sensitive and per-iodate resistant. In addition, it has been demon-strated that the ,B determinant is resistant totrypsin digestion (18). Serological testing withthis ,3-antigen-rich extract has been unreliablebecause of a high rate of false positive and nega-tive reactions (15, 17). In order to standardizethis test system and to evaluate the specificity ofthe 13 antigen in the human serological responseto the gonococcus, we undertook the isolation ofthis antigen from the crude extract. In this paper,we will describe the isolation and chemical char-acteristics of 13 antigen of N. gonorrhoeae.
MATERIALS AND METHODSOrganisms. N. gonorrhoeae was obtained from
fresh patient isolates and grown for antigen prepa-
ration within 72 h of initial isolation. All organismswere unequivocally N. gonorrhoeae by gram stain,cultural characteristics, oxidase reactivity, andsugar fermentations. Organisms were maintainedon chocolate agar at 35 C in a moist CO2 incubatorand preserved by lyophilization in 5% bovineserutn albumin-5% monosodium glutamate.
Antisera. Antisera to the whole gonococcuswere produced by two injections of 1 ml of 109organisms emulsified 1:2 with complete Freundadjuvant in the posterior cervical region of malealbino rabbits at 10-day intervals. On days 21 and23 after the onset of immunization, the animalswere injected intravenously with 0.1 ml of 109organisms. The rabbits were bled for antisera 7days later. The organisms used for production ofantisera were collected from 20 fresh isolateseither by scraping chocolate plates with a glassspatula or by growth in supplemented Mueller-Hinton broth. Organisms were washed three timesin distilled water before use as antigens. Antiserawere also produced to purified ,B-antigen prepara-tions in rabbits. This was accomplished by footpadinjection of 250 Ag of antigen emulsified in anequal volume of Freund adj uvant at weeklyintervals for 28 days. Animals were bled for anti-body 7 days after the final injection.Growth of N. gonorrhoeae in liquid me-
dium. Organisms were grown for antigen isolationin 2-liter screw-capped culture flasks in 1 liter ofMueller-Hinton broth containing modified Lang-ford supplement. Attempts were made to insurethat all organisms were colonial type I or II ofKellogg et al. (13) before being used as iilocula inthe broth. Multiple vials of fresh individual (typeI or II) isolates were lyophilized. Continuedavailability of these colonial forms from the sameorganism for growth in liquid medium was main-tained by reconstitution of these lyophilized
315
on July 27, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

APICELLA AND ALLEN
organisms. The flasks were inoculated directlywith a loopful of organisms from a 24-h chocolateplate. The flasks were placed on a gyratory shakerplatform with a shaking rate of 180 per min. Afterinoculation and prior to incubation, each flask wasgassed with a filtered mixture of 10% CO2 and airfor 30 s. Cultures were grown for 16 h, subcultured,and placed at 4 C for 48. h to insure death oforganisms. After confirmation that the flasks con-tained pure cultures of N. gonorrhoeae, the or-ganisms were spun free in a Sorvall continuous-flow centrifuge at 6,000 X g, washed once withdistilled water and once with phosphate-bufferedsaline (PBS), and were then ready for antigenextraction. Approximately 1.8 g (wet weight) oforganisms was obtained per liter of broth.HA and HAI. Both hemagglutination (HA) and
hemagglutination inhibition (HAI) were used todetect the presence of the (3 antigen in the isolationprocedures. All studies were done by using micro-titer techniques. The HA system was similar tothat described by Edwards and Driscoll (10) formeningococcal polysaccharides. The HAI systememployed antisera either to whole organisms orisolated antigen as the agglutinator. The erythro-cyte-coating antigen was an alkaline-extractedpreparation made from 20 individual gonococcalisolates. The concentration of antisera and cell-coating antigen for the inhibition system were de-termined by a block titration with the final con-centration of each reagent being one dilution lessthan the respective end point. The erythrocyte-coating antigen did contain some a-antigen ac-tivity but, at the dilutions of antisera and antigenemployed, the only antigen detected was the (antigen. In addition, absorption of the antiserawith as much as 5 mg of a antigen per ml had noeffect on the inhibition titers of ,B antigens. AnHAI system was also established to gonococcala antigen by a method identical to that describedabove for gonococcal ,B antigen. Analysis of thepurified preparation for common enterobacterialantigen was performed by Erwin Neter (35).
Immunodiffusion was done by the method ofOuchterlony in 2% noble agar containing 0.05 Mbarbital buffer, pH 8.6 (24). Immunoelectro-phoresis was done with the same agar and bufferconditions using the method of Scheidegger (25).Ion exchange chromatography. Ion exchange
chromatography was performed by using CM-Sephadex 50 equilibrated with 0.01 M acetatebuffer, pH 5.2. To determine the initial elutionpattern, a Buchler Varigrad was used to make anionic-strength gradient. Wells 1, 2, 3, 5, and 8 con-tained the starting buffer; wells 6, 7, and 9 con-tained 0.1M acetate, pH 5.2. Well 4 contained90%0 starting buffer and 10% final buffer. Each wellcontained 40 ml of buffer. A 6-ml sample was ap-plied to the column, and the elution volumes were7 ml. At the termination of the gradient the.column was washed with 0.5M NaCI. After theelution pattern was established, batch prepara-tions were used subsequently in antigen isolation.Molecular sieve chromatography. Molecular
sieve chromatography was performed with P-200(Bio-Rad) in a glass column (100 by 2.5 cm) byupward flow. The column was equilibrated with a0.2M acetate buffer, pH 5.2. Sample volume was10 ml, and the elution volume was 4 ml.Pevikon block electrophoresis. Powder block
electrophoresis was performed by the method ofKunkel (14) using Pevikon. The origin was placedin the center of the block, and the sample waselectrophoresed for 6 h at 400 V and 4 C.Enzyme digestions. During isolation of the
antigen, the crude alkaline extract was dialyzedagainst PBS and treated with Ribonuclease A(Worthington Biochemical) 10 ugm/ml at 37 C for16 h. Pronase digestion was performed by theaddition of Pronase (Calbiochem) in PBS at 37 Cfor 24 h. Trypsin digestion was accomplished bythe addition of trypsin (Worthington Biochemical)(2%, wt/wt) to antigen previously dialyzed against0.01 M Tris (pH 8.1), 0.005M CaCl2 at 37 C for 24h. Periodation of the antigen was accomplished byadding equal volumes of 0.04 M sodium meta-periodate to 3 antigen in distilled water (12). Thiswas allowed to stand at 20 C for 48 h. Sodiummetapeiiodate was removed by dialysis againstPBS at 4 C.Preparation of phenol-extracted endotoxin
and a determinant. Preparation of the phenol-extracted endotoxin was achieved by the methodof Tauber and Russell (31) and treated withPronase as suggested by Maeland and Kristoffer-sen (22) to make it specific for a antigen.Chemical studies. Moisture content was de-
termined by drying, total nitrogen was deter-mined by Galbraith Analytic Laboratories, andtotal and inorganic phosphorus were determinedby the method of Chen (5). Determination ofprotein content was done by the Biuret methodand Folin-Ciocalteau reaction with human serumalbumin used as standard (4, 16). Amino acidanalysis was performed by the method of Spack-man, Stein and Moore, using a Beckman model120C amino acid analyzer (27). Samples werehydrolyzed at 24-, 48-, and 72-h intervals in 6 NHCI at 110 C in vacuo. All analyses were performedin duplicate, and the results are expressed as amean of these values.
Total carbohydrate content was determined bythe phenol sulfuric acid method (9) and totalhexoses by the anthrone method (26). Methylpentoses and pentoses were determined by themethod of Dische (7) and Dische and Shettles (8),and 2-keto 3-deoxy sugar acids were determined bythe method o£Warren (34).
Qua1tative analysis of the antigen for lipid wasperforme&- by thin-layer chromatography withsilica gel plates in- a benzene-methanol (85:15)solvent system. The antigen was subjected toalkaline hydrolysis fot 16 h ihi methanolic potas-sium hydroxide. Lipids were revealed on theplates by spraying with a 0.05% solution ofRhodamine B in ethanol and scanning with anultraviolet lamp.
Spectral analysis in the visible range (700 to
316 INFECT. IMMUNITY
on July 27, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

L7ANTIGEN OF N. GONORRHOEAE
400 nm) and ultraviolet range (400 to 180) wasperformed with a Cary model 14 recording spec-trophotometer. A 1-cm light path was used, andsamples were dissolved in PBS, pH 7.2.
Calculation of s2o, w was performed by using amodel E analytical ultracentrifuge with schlierenoptics (6). Samples were spun at 52,640 rpm in aAN-D rotor in PBS, pH 7.2.
Acrylamide gel electrophoresis was performedby a modification of the method of Hilborn andAnastassiadis (11). Samples were electrophoresedat 6 mA per gel for 45 min and stained for carbo-hydrate with 0.5% Alcian Blue in 7% acetic acidor for protein with 0.5% Coomassie Blue in 10%trichloracetic acid. Gels were constructed in glasstubes (5 by 60 mm), and the sample applied was20 pliters of a 2-mg/ml antigen solution.
RESULTSAn outline of the method of isolation of the ,3
antigen of N. gonorrhoeae is presented in Fig. 1.The alkaline-extracted endotoxin studied byMaeland includes steps 1 through 7. Ultravioletspectral analysis of this crude alkaline extract(at step 7) revealed the presence of significantquantities of ribonucleic acid. Ribonuclease A
Step1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
digestion proved to be the most efficient methodof removing this contaminant. The preparationwas then dialyzed against 0.01 M acetate buffer(pH 5.2) and placed on a CM-50 Sephadexcolumn equilibrated with the same buffer. The, antigen under these buffer conditions wasexcluded from the CM sephadex, whereas asubstantial portion of other substances in thealkaline extract did adhere. The antigen elutedat the void volume of the column in the startingbuffer. In the salt concentrations produced bythe 0.01 to 0.1 M sodium acetate gradient, noother peaks eluted. When the column was washedwith 0.5 M NaCl, a second peak eluted which wasdevoid of , antigen. After the a-antigen-con-taining peak was concentrated and dialyzedagainst 0.2 M acetate buffer (pH 5.2), it wasapplied to a P-200 column (2.5 by 100 cm). Theelution profile of this column is shown in Fig. 2.The # antigen eluted just after the void volumeof the column. A second peak could be detectedby 280-nm absorption and was devoid of , anti-gen. Immunization of rabbits with eluate con-taining the , antigen resulted in the production of
N. gonorrhoeae organisms
0.1 M NaOH-glycine, pH 10.6
Dialysis PBS, pH 7.0
Ethanol 4 volumesF- Discard superilataint
Precipitate
Dissolve in PBS
Ultracentrifugation(50,000-60,000 X g)
- Discard precipitate (wash 3 times)Ribonuclease A
Concentration anddialysis 0.01 M acetate, pH 5.2
CM-Sephadex
Concentration anddialysis 0.2 M, pH 5.2
P-200
Concentration anddialysis 0.05 M Barbital, pHl 8.6
1'evikoni block electrophoresis
Dialysis against distillced water
Lyophilization
FIG. 1. Schematic outline of the isolation of 6 antigen from gonococcal alkaline-extracted endotoxin.
317VOL. 7, 1973
on July 27, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
APICELLA AND ALLEN
.7
.6
.5
.40 . D.280nWi .3
.2 / I
10 20 30 40 50 60 70 80 90 100
tube #
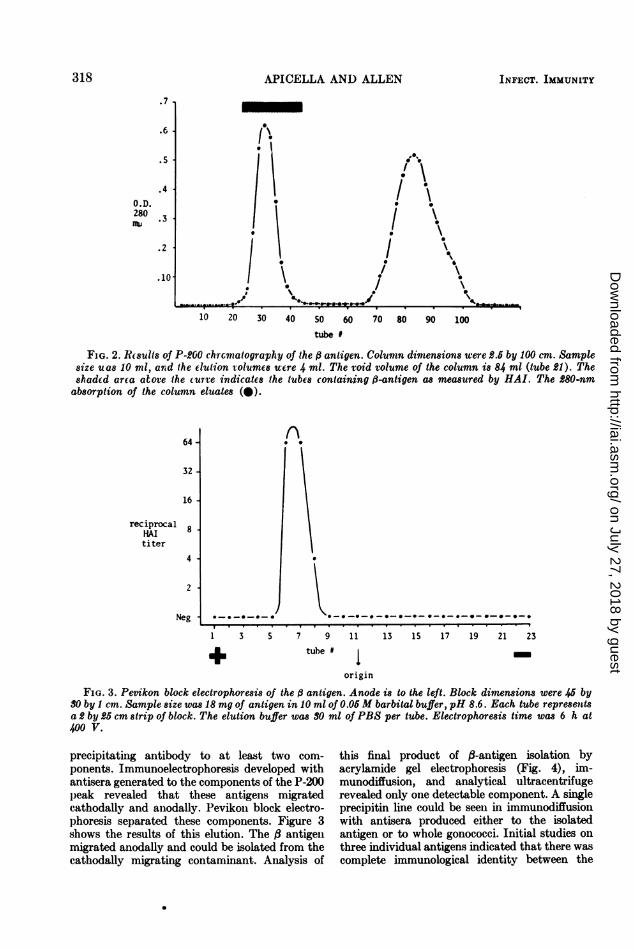
FIG. 2. R(sults of P-200 chrcrnatography of the , antigen. Column dimensions were 2.5 by 100 cm. Samplesize uas 10 ml, arid the elution volumes were 4 ml. The void volume of the column is 84 ml (tube 21). Theshaded area atove the curve indicates the tubes containing i-antigen as measured by HAI. The 280-nmabsorption of the column eluates (@).
7 9 11 13 15 17 19 21 23tube 1m
origin
FIG. 3. Pevikon block electrophoresis of the ,3 antigen. Anode is to the left. Block dimensions were 45 by30 by 1 cm. Sample size was 18 mg of antigen in 10 ml of 0.05 M barbital buffer, pH 8.6. Each tube representsa 2 by 25 cm strip of block. The elution buffer was 30 ml of PBS per tube. Electrophoresis time was 6 h at400 V.
precipitating antibody to at least two com-ponents. Immunoelectrophoresis developed withantisera generated to the components of the P-200peak revealed that these antigens migratedcathodally and anlodally. Pevikon block electro-phoresis separated these components. Figure 3shows the results of this elution. The # antigenmigrated anodally and could be isolated from thecathodally migrating contaminant. Analysis of
this final product of ,-antigen isolation byacrylamide gel electrophoresis (Fig. 4), im-munodiffusion, and analytical ultracentrifugerevealed only one detectable component. A singleprecipitin line could be seen in immunodiffusionwith antisera produced either to the isolatedantigen or to whole gonococci. Initial studies onthree individual antigens indicated that there wascomplete immunological identity between the
64 -
32-
16
reciprocalHAItiter
8-
4.
2
NegI . . .
1 3 S
318 INFECT. IMMUNITY
0 0
0
i0-0-*-O-O
on July 27, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

V1ANTIGEN OF N. GONORRHOEAE
Betaait "
FIG. 4. Acrylamide gel electrophoresis of the(3 antigen in a 6% cyanogumn gel. The buffer was0.01 M formate-phosphate, pH 11.6. Sample waselectrophoresed at 6 mA per gel for 45 min andstained with 0.5% Alcian Blue in 7% acetic acid.
TABLE 1. Results of enzymatic and oxidativetreatment of isolated, antigen as measured by
the ability of the treated antigen toinhibit hemagglutination
Treatment Inhibiting Concn of inhibitorTreatment titer (Iug/mnl)B Antigen control 1:2,048 0.49Pronase-treated (3 Negative >1,000antigen
Periodate-treated 1:1,024 0.98( antigen
Trypsin-treated 1:1,024 0.98antigen
anitigens studied. HAI studies failed to detecta antigen in the preparation, and studies forcommon enterobacterial antigen by Erwin Neterrevealed only trace amounts of this antigen byHAI.The effects of enizyme and oxidative treatments
of the anitigenl are shown iln Table 1. Pronasedestroyed all antiggenic activity, whereas perioda-tioni and trypsin had little effect on the antigenicdeterminiant. The senisitivity of the HAI test wasalso demonstrated with 0.49 jug of , antigendetected by this method. From the isolation
procedures described above, the best yield ofantigen was 8 mg of lyophilized antigen from50 g (wet weight) of organisms or 750 mg ofcrude dry alkaline-extracted endotoxin. Theisolated ,-antigen determinant appeared to beimmunologically identical to that present in thecrude alkaline extract; however, one majoralteration had occurred during isolation. Thecrude antigen hemabsorbed directly to erythro-cytes, whereas the purified # antigen did not.Physicochemical studies. Chemical analyses
of the , antigen indicated that it was a glyco-protein. After lyophilization, the moisture con-tent of the anitigen was 28.8% as determined bydrying. All chemical analyses were based on theweight of the antigen free of water. The totalnitrogen was 11.2% of the dry weight, whereasthe total phosphorus was less than 0.5% of thedry weight of the antigen. The protein portionaccounted for 61.2 to 63.7% dry weight ofantigen as determined by the Biuret reaction. Itis of interest that protein measurement by themodified Folin-Ciocalteau method in antigenconcentrations as high as 1 mg/ml produced nocolor development. This may be related eitherto a paucity of aromatic amino acids or to thehigh carbohydrate content of the antigen (1, 2,33). The results of amino acid analysis are shownin Table 2. The major amino acids present werealanine, glutamic acid, glycine, and proline.These four amino acids accounted for between 74and 79% of the total micromoles of amino
TABLE 2. Amino acid analysis of the (3 antigen"
Amino acids
Lysine .............Histidine ..........Arginine ...........Aspartic acid......Threonine.......Serine .............Glutamic acid......Proline ............Glycine ............Alanine ............Valine .............Methionine ........Isoleucine..........Leucine ............Tyrosine ...........Phenylalaniine ......
Antigen (umoles/100 mg)
24 hb
6.701.101.755.276.076.4734.6218.419.1569.543.592.043.805.262.552.12
a All samples were hyvacuo at 110 C.
b Hours of hydrolysis.
48 hb
6.801.341.465.496.826.29
37.7127.7511.1986.253.351.522.394.121.152.65
72 hb
6.361.77NP4.945.114.5428.4715.838.53
62.143.86NP2.283.310.891.12
rdrolyzed in 6 N HCI in
VOL. 7, 1973 319
on July 27, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

APICELLA AND ALLEN
acids in the protein portion, depending on thehydrolysis time. The optimal hydrolysis timevaried for each amino acid, but the 48-h timegave the greatest yield of the four major aminoacids. The antigen contained small amounts oftyrosine and phenylalanine. Amino acid analysisof ,B antigen isolated from two other organismsgave similar results. Direct measurement oftryptophan had not been performed, but theultraviolet absorption spectrum would indicatethat this amino acid, if present, was in lowconcentrations. Spectral analysis revealed noabsorption peaks in the visible light range (400 to700 nm) and a single absorption peak at 217 nmin the ultraviolet range (399 to 180 nm).
Carbohydrate content by the phenol-sulfuricacid method was approximately 27.6 to 29.8%.Hexose content was between 6 and 7% by theanthrone method. Methyl pentoses, pentoses,and 3-keto 2-deoxy sugar acids were not de-tectable colorimetrically. More detailed studies ofthe carbohydrate component of this antigen arepresently in progress. Lipids could not be detectedin this antigen by qualitative analysis on thin-layer chromatography. By our methods ofanalysis, we could account for between 88.8 and93.5% of the dry weight of antigen as proteinand carbohydrate. Finally, ultracentrifugationstudies indicated that the antigen sedimented as asingle component in the analytical ultracentrifugewith an ss%o, of 8.55.
DISCUSSIONIn 1950, Thomas and Mennie (32) first sug-
gested the use of erythrocytes sensitized by cellwall components of the gonococcus to study theimmunological response of gonococcal infectionin man. Chanarin (3) expanded this techniqueand described in detail methods for preparingsensitized erythrocytes with an alkaline extractof the gonococcus. Tauber showed that thealkaline-extracted endotoxin, a preparation iden-tical to that studied by Chanarin, was composedprimarily of protein with small amounts of lipidand polysaccharide (29). This is in contrast to thephenol-extracted endotoxin which contains pri-marily polysaccharide and lipid with smallamounits of protein (30, 31). Maeland studiedcharacteristics of these two endotoxin prepara-tions and aqueous ether endotoxin (17, 23). Thealkaline extract was devoid of endotoxin activitybut contained antigenic determinants present inthe two preparations with endotoxin activity(18). Two major antigenic determinants in thesepreparations were designated a and ,B. The as-sumption has also been made that these twoantigens are a part of the lipid A-lipid B-protein
polysaccharide structure of gonococcal endotoxin(18). To more precisely define the chemical andserological nature of this antigen, isolation fromalkaline-extracted endotoxin has been achievedby serial chromatographic and electrophoreticprocedures. By these methods, the yield of I3antigen from lyophilized alkaline-extracted endo-toxin is approximately 1% of the dry weight ofstarting extract, demonstrating the high degree ofcontamination of this determinant in the crudeextract. The B antigen is an acidic glycoprotein.Previous studies on crude extracts (30) haveindicated the carbohydrate content to be ap-proximately 3%. The purified antigen contains aconsiderably higher carbohydrate content, in therange of 30% of the dry weight of antigen. Itcontains no cysteine and very small amounts ofhistidine, methionine, tyrosine, and phenylala-nine. Since antisera to whole gonococci are capableof reacting with this isolated B antigen in bothimmunodiffusion and HAI systems, it would ap-pear that the antigenic nature of the determinanthas not been significantly altered by isolation.One major change has occurred during isolation,that is, the antigen will no longer directly hemab-sorb as in the crude extract. Whether this isrelated to loss of a portion of the molecule con-taining a lipid or carbohydrate moiety or due toloss of other charged groups is not known. Thelocation of the antigen in the gonococcus is un-certain; the studies of Maeland strongly suggestit is in the cell wall (18). Whether this antigen isa component of endotoxin is unknown, but theabsence of 2-keto 3-deoxy sugar acids or heptosesmake this unlikely. The relationship betweenthis antigen and gonococcal pili is unknown (28).At the present time, investigations are under-
way to determine the components of the carbo-hydrate portion of the antigen and the serologicalspecificity of the 13 antigen in animal and humanserological systems.
ACKNOWLEDGMENTSWe thank Gail Feser, Phyllis Rosenberg, and
Nicholas Gagliardi for expert technical assistance;Erwin Neter for assay of our antigen for common en-terobacterial antigen; Nicholas Calvanico for helpfulcomments; and Thomas B. Tomasi for use of equipmentin his laboratory.
This investigation was supported in part by PublicHealth Service grant 1-RO1-CC-00588-01 from theCenter for Disease Control, Atlanta, Ga.
LITERATURE CITED1. Berg, D. H. 1971. Hexosamine interference with the
determination of protein by the Lowry procedure.Anal. Biochem. 42:505-508.
2. Bonitali, J., W. B. Elliot, and P. G. Miles. 1969.Interference by carbohydrate and other sub-stances in the estimation of protein with the
320 INFECT. IMMUNITY
on July 27, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

,8 ANTIGEN OF N. GONORRHOEAE
Folin-Ciocalteau reagent. Anal. Biochem. 31:399-404.
3. Chanarin, I. 1954. An investigation of Neiseriagonorrheae by a red cell sensitization technique.J. Hyg. 52:425-443.
4. Chase, M. W., and C. A. Williams. 1968. Proteinanalysis, p. 271-275. In C. A. Williams andM. W. Chase (ed.), Methods in immunology andimmunochemistry, vol. 2. Academic Press Inc.,New York.
5. Chen, P. S., T. Y. Toribara, and H. Warner. 1956.Microdetermination of phosphorus. Chemistry28:1756-1758.
6. Chervenka, C. H. 1969. A manual of methods forthe analytical ultracentrifuge. Beckman Instru-ments Inc., Palo Alto, Calif.
7. Dische, Z. 1949. Spectrophotometric method for thedetermination of free pentose and pentose in nu-cleotides. J. Biol. Chem. 181:379-392.
8. Dische, Z., and L. B. Shettles. 1948. A specific colorreaction of methyl pentose and a spectropho-tometric micromethod for their determination.J. Biol. Chem. 175:595-602.
9. Dubois, M., K. A. Gilles, J. K. Hamilton, P. A.Rebers, and F. Smith. 1956. Colorimetric methodfor determination of sugars and related sub-stances. Anal. Chem. 28:350-356.
10. Edwards, E. A., and W. S. Driscoll. 1967. Group-specific hemagglutination test for Nei.seria men-inaitidis antibodies. Proc. Soc. Exp. Biol. Med.126:876-879.
11. Hilborn, J. C., and P. A. Anastassiadis. 1971. Esti-mation of the molecular weights of acidic muco-polysaccharides by polyacrylamide gel electro-phoresis. Anal. Biochem. 39:88-92.
12. Kabat, E. A., and M. M. Meyer. 1961. Experimen-tal immunochemistry, p. 546. Charles C Thomas,Publisher, Springfield, Ill.
13. Kellogg, D. S., I. R. Cohen, L. C. Norins, A. L.Schroeter, and G. Reising. 1968. Neisseria gonor-rheae. II. Clonal variation and pathogenicity dur-ing 35 months in vitro. J. Bacteriol. 96:596-605.
14. Kunkel, H. G. 1954. Zone electrophoresis, p. 141.In D. Glicke (ed.), Methods of biochemical anal-ysis, vol. 1. Interscience, New York.
15. Logan, L. C., P. M. Cox, and L. C. Norins. 1970.Reactivity of two gonococcal antigens in an auto-mated microhemagglutination procedure. Appl.Microbiol. 20:907-909.
16. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, andR. J. Randall. 1951. Protein measurement withthe Folin phenol reagent. J. Biol. Chem. 193:265-275.
17. Maeland, J. A. 1966. Antibodies in human seraagainst antigens in gonococci demonstrated by apassive hemolysis test. Acta Pathol. Microbiol.Scand. 67:102-110.
18. Maeland, J. A. 1968. Antigenic properties of variouspreparations of Neisseria gonorrheae endotoxin.Acta Pathol. Microbiol. Scand. 73:413-422.
19. Maeland, J. A. 1969. Antigenic determinants ofaqueous ether extracted endotoxin from Neisseriagonorrheae. Acta Pathol. Microbiol. Scand. 76:475-483.
20. Maeland, J. A. 1969. Immunochemical character-ization of aqueous ether extracted endotoxin fromNei88eria gonorrheae. Acta Pathol. Microbiol.Scand. 76:484-492.
21. Maeland, J. A. 1969. Serological cross-reactions ofaqueous ether extracted endotoxin from Neisseriagonorrheae strains. Acta Pathol. Microbiol. Scand.77:505-517.
22. Maeland, J. A., and T. Kristoffersen. 1971. Immu-nochemical ihvestigations on Neisseria gonorrheaeendotoxin. I. Characterization of phenol-waterextracted endotoxin and comparison with aqueousether preparations. Acta Pathol. Microbiol.Scand. 79B:226-232.
23. Maeland, J. A., T. Kristoffersen, and T. Hofstead.1971. Immunochemical investigations on Nei8-seria gonorrheac endotoxin. II. Serological multi-specificity and other properties of phenol waterpreparations. Acta Pathol. Microbiol. Scand.79B:233-238.
24. Ouchterlony, 0. 1962. Diffusion-in-gel methods forimmunological analysis. Progr. Allergy 6:30-154.
2.5. Scheidegger, J. J. 1955. Une micro-methode del'immunoelectrophorese. Int. Arch. Allergy 7:103-110.
26. Scott, T. A., and E. H. Melvin. 1953. Determinationof dextran with anthrone. Anal. Chem. 25:1656-1661.
27. Spachman, D. H., W. H. Stein, and S. Moore.1958. Automatic recording apparatus for usein the chromatography of amino acids. Anal.Chem. 30:1190-1206.
28. Swanson, J., S. J. Kraus, and E. C. Gotschlich.1971. Studies on gonococcus infection. 1. Pili andzones of adhesions. Their relation to gonococcalgrowth patterns. J. Exp. Med. 134:886-906.
29. Tauber, H., and W. Garson. 1957. Preparation andsome properties of Neisseria gonorrheae endotoxin.Proc. Soc. Exp. Biol. Med. 95:669-672.
30. Tauber, H., and W. Garson. 1959. Isolation of lipo-polysaccharide endotoxin. J. Biol. Chem. 234:1391-1393.
31. Tauber, H., and H. Russell. 1960. Amino com-pounds in lipopolysaccharides. J. Biol. Chem.235:961-964.
32. Thomas, J. C., and A. T. Mennie. 1950. Bacterialpolysaccharides in the diagnosis of infections.The polysaccharide lysis test. Lancet 2:745-746.
33. Toennies, G., and Feng, F. 1965. Measurementand characterization of proteins by color reac-tions. Anal. Biochem. 11:411-417.
34. Warren, L. 1959. The thiobarbituric acid assay ofsialic acids. J. Biol. Chem. 234:1971-1974.
35. Whang, H. Y., M. E. Heller, and E. Neter. 1972.Production by Aeromonas of common enterobac-terial antigen and its possible taxonomic signifi-cance. J. Bacteriol. 110:161-164.
321VOL. 7, 1973
on July 27, 2018 by guesthttp://iai.asm
.org/D
ownloaded from


















