Macroscopic Structure of the Fin-Rays and TheirAnnuli in ...
24
Macroscopic Structure of the Fin-Rays and Their Annuli in Pectoral and Pelvic Fins of Chinook Salmon, Oncorhynchus tshawytscha LEONOR C. G. FERREIRA, 1 RICHARD J. BEAMISH, 2 AND JOHN H. YOUSON 1 * 1 Department of Zoology and Division of Life Sciences, University of Toronto at Scarborough, Scarborough, Ontario M1C 1A4 Canada 2 Department of Fisheries and Oceans, Fisheries Research Branch, Pacific Biological Station, Nanaimo, British Columbia V9R 5K6 Canada ABSTRACT Light microscopy, enzyme clearing, and staining techniques were used to describe the structure of fin-rays in pectoral and dorsal fins of adult and juvenile chinook salmon,Oncorhynchus tshawytscha. In addition, several decalcification agents, fixatives, and staining methods were employed to demonstrate and determine the nature of the fin-ray annulus (yearly growth ring). Etched, transverse sections of fin-rays were examined by scan- ning electron microscopy (SEM). A description is provided of specific morpho- logical regions along the length of fin-rays in both fins. Each ray consists of parallel halves, medial and lateral hemisegments, which diverge proximally and embrace underlying radial cartilages and are segmented distally. Fibro- cartilage masses are associated with bone and hyaline cartilage in the proximal regions. There is evidence of periosteal, and perhaps endochondral, osteogenesis in the fin-rays. A novel fixation-staining method is described to depict the fin ray annulus in adult chinook salmon. Transverse sections of rays of adults, but not juveniles, demonstrate alternating narrow-dark and wide-light purple bands following prolonged decalcification/fixation in Bouin’s fluid only with subsequent hematoxylin-eosin staining. The dark and light bands correspond to translucent (annuli) and opaque zones, respectively, in dried, unstained sections in transmitted light and in backscatter SEM. In conjunction with different light microscopic staining procedures, etched, transverse sections of fin-ray bone in SEM suggest that annuli are a manifes- tation of an ordered regionalization of the extracellular matrix of the bone in the fin-rays. J. Morphol. 239:297–320, 1999. r 1999 Wiley-Liss, Inc. KEY WORDS: fin-ray; Oncorhynchus tshawytscha; annuli The ability to age fish is important in fisheries management and calcified struc- tures (i.e., osseochronometry), such as scales, otoliths, opercular bones, vertebrae, dorsal spines, and fin-rays, are useful tools. In all these osseous structures, age determination is dependent upon recognizable yearly growth rings, referred to as annuli. Many definitions exist to describe exactly what the annulus and its accompanying structures represent, but most recently Casselman (’87) clearly explained that age determination should be expressed as a systematic, yet subjective interpretation of various ‘‘checks,’’ ‘‘breaks’’ or ‘‘changes in spacing,’’ or ‘‘zones’’ that, based on their translucency, are opti- cally different. Checks or zones, considered to be associated with yearly deposited annu- lar rings, referred to as ‘‘annuli,’’ are then enumerated to determine age. Like mammals, teleosts possess both com- pact and spongy bone formed from either Contract grant sponsor: Department of Fisheries and Oceans, Canada; Contract grant sponsor: Natural Science and Engineer- ing Research Council of Canada (NSERC). *Correspondence to: J.H. Youson, Department of Zoology and Division of Life Sciences, University of Toronto at Scarborough, Scarborough, Ontario M1C 1A4 Canada. E-mail: [email protected] JOURNAL OF MORPHOLOGY 239:297–320 (1999) r 1999 WILEY-LISS, INC.
Transcript of Macroscopic Structure of the Fin-Rays and TheirAnnuli in ...

Macroscopic Structure of the Fin-Rays and Their Annuliin Pectoral and Pelvic Fins of Chinook Salmon,Oncorhynchus tshawytscha
LEONOR C. G. FERREIRA,1 RICHARD J. BEAMISH,2AND JOHN H. YOUSON1*1Department of Zoology and Division of Life Sciences, University of Torontoat Scarborough, Scarborough, Ontario M1C 1A4 Canada2Department of Fisheries and Oceans, Fisheries Research Branch, PacificBiological Station, Nanaimo, British Columbia V9R 5K6 Canada
ABSTRACT Light microscopy, enzyme clearing, and staining techniqueswere used to describe the structure of fin-rays in pectoral and dorsal fins ofadult and juvenile chinook salmon,Oncorhynchus tshawytscha. In addition,several decalcification agents, fixatives, and staining methods were employedto demonstrate and determine the nature of the fin-ray annulus (yearlygrowth ring). Etched, transverse sections of fin-rays were examined by scan-ning electron microscopy (SEM). A description is provided of specific morpho-logical regions along the length of fin-rays in both fins. Each ray consists ofparallel halves, medial and lateral hemisegments, which diverge proximallyand embrace underlying radial cartilages and are segmented distally. Fibro-cartilage masses are associated with bone and hyaline cartilage in theproximal regions. There is evidence of periosteal, and perhaps endochondral,osteogenesis in the fin-rays. A novel fixation-staining method is described todepict the fin ray annulus in adult chinook salmon. Transverse sections ofrays of adults, but not juveniles, demonstrate alternating narrow-dark andwide-light purple bands following prolonged decalcification/fixation in Bouin’sfluid only with subsequent hematoxylin-eosin staining. The dark and lightbands correspond to translucent (annuli) and opaque zones, respectively, indried, unstained sections in transmitted light and in backscatter SEM. Inconjunction with different light microscopic staining procedures, etched,transverse sections of fin-ray bone in SEM suggest that annuli are a manifes-tation of an ordered regionalization of the extracellular matrix of the bone inthe fin-rays. J. Morphol. 239:297–320, 1999. r 1999 Wiley-Liss, Inc.
KEY WORDS: fin-ray; Oncorhynchus tshawytscha; annuli
The ability to age fish is important infisheries management and calcified struc-tures (i.e., osseochronometry), such as scales,otoliths, opercular bones, vertebrae, dorsalspines, and fin-rays, are useful tools. In allthese osseous structures, age determinationis dependent upon recognizable yearlygrowth rings, referred to as annuli. Manydefinitions exist to describe exactly what theannulus and its accompanying structuresrepresent, but most recently Casselman (’87)clearly explained that age determinationshould be expressed as a systematic, yetsubjective interpretation of various ‘‘checks,’’‘‘breaks’’ or ‘‘changes in spacing,’’ or ‘‘zones’’
that, based on their translucency, are opti-cally different. Checks or zones, consideredto be associated with yearly deposited annu-lar rings, referred to as ‘‘annuli,’’ are thenenumerated to determine age.
Like mammals, teleosts possess both com-pact and spongy bone formed from either
Contract grant sponsor: Department of Fisheries and Oceans,Canada; Contract grant sponsor: Natural Science and Engineer-ing Research Council of Canada (NSERC).
*Correspondence to: J.H. Youson, Department of Zoologyand Division of Life Sciences, University of Toronto atScarborough, Scarborough, Ontario M1C 1A4 Canada. E-mail:[email protected]
JOURNAL OF MORPHOLOGY 239:297–320 (1999)
r 1999 WILEY-LISS, INC.

lamellar or woven forms and there is histo-logical evidence of osseous resorption andredeposition found in both types (Moss, ’61a).Although teleost bone lacks true osteons, itdoes nonetheless have numerous vascularchannels. These channels, referred to as pri-mary osteons, commonly anastomose witheach other through lateral branching canalsthat appear to be homologous to the Volk-mann’s canals of mammals (Moss,’61a). Theorganic components and the inorganic/or-ganic ratio of piscine bone is similar to thatdescribed for other vertebrates (Moss andPosner, ’60).
In contrast to the situation in higher verte-brates, however, teleost bone is unique inthat it may be totally devoid of osteocytes(Parenti, ’86). Based on the presence or ab-sence of enclosed osteocytes, piscine bone isidentified as osteocytic (cellular) or anosteo-cytic (acellular), respectively (Weiss, ’78). Insome extreme instances the endoskeletal el-ements of fish may contain a mixture ofcellular and acellular types (Moss, ’61a). Pis-cine osteogenesis is described as either peri-osteal or chondroidal (Moss, ’61b, ’63). Chon-droidal osteogenesis is a process unique toteleosts and enables the direct transfor-mation of chondroid (a nonvascular, cellulartissue). Periosteal osteogenesis is the for-mation of bone through metaplasia of fibro-blasts and their differentiation into osteo-blasts within the periosteum.
Descriptions of the histological structureof the mineralized tissues used in ageing ofteleosts have mainly focused on otoliths. Al-though it has been demonstrated that an-nuli are an integral part of the internalcalcified components of the fish skeleton, ourcurrent knowledge of ageing does not in-clude a detailed study of annulus composi-tion and structure. Given the importance ofageing to fisheries management and thepresent concern of declining commercial fishstocks, there is a need to correct this defi-ciency in our knowledge of ageing.
The overall goal of our study is to eluci-date the internal constituents of the fin-rayannulus in chinook salmon. However, to datethere has not been a study of this type car-ried out on fin-rays for any fish species. It isnot the focus of this study to determine ageby the enumeration of annuli, but instead toelucidate the actual structure and composi-tion of these yearly deposited incrementsusing light microscopy, scanning electron mi-croscopy, and transmission electron micros-
copy. The availability and the ease withwhich fin-rays are removed and their useful-ness in determining age of long-lived speciesmakes these structures an ideal choice ofstudy. The present report is the first of a twopart series. In this first part, the objectivesare:(1) to document in detail the macro-scopic structure and arrangement of fin-raysin both the pectoral and dorsal fins by theuse of light microscopy and enzyme-clearingand staining techniques; (2) to describe andcompare the presence of the annulus in juve-nile and adult salmon using light micros-copy; and (3) to use scanning electron micro-scopic techniques in order to describe thethree-dimensional appearance and relativeamounts of inorganic salts present withinthe differing regions of the fin-ray.
MATERIALS AND METHODSAnimal and tissue collection
Chinook salmon, Oncorhynchus tshaw-ytscha, were collected by fishermen troll-fishing in various areas throughout the Straitof Georgia, British Columbia, during themonths of September to May and in Febru-ary. Upon capture of the animals, theirlength, weight, and sex were recorded. Allfish were killed by a sharp blow to the headand the immediate removal of both the leftand right pectoral fins and the dorsal finfollowed. Randomly, fins were either placedin small envelopes and air dried, or in solu-tions of either Bouin’s fixative, 10% bufferedformalin, or 2.5% glutaraldehyde in a phos-phate buffer, pH 7.4 (Warshawsky andMoore, ’67) for further processing at a latertime. In conjunction with the above proce-dures, scales were also taken from desig-nated areas on the same fish, as outlined inan earlier report by the International NorthPacific Fisheries Commission (’59). All finsto be analyzed for light and electron micros-copy were shipped to the laboratory at theUniversity of Toronto at Scarborough, fromthe Pacific Biological Station in Nanaimo,B.C. Dried fins and scales remained at theStation for analysis.
In order to compare fin-ray annuli of olderand younger individuals, the Ministry ofNatural Resources of Ontario at Ringwood,Ontario, provided juvenile chinook salmon(approximately 130–150 days old).
Enzyme clearing and stainingThis technique was modified from that of
Dingerkus and Uhler (’77). Formalin-fixed
298 L.C.G. FERREIRA ET AL.

samples were rinsed in two changes of dis-tilled water for 5 min each followed by com-plete immersion of the fin in an alcian bluestaining solution for 24–48 hr or until thetissue was light blue in color. This stain wasspecific for the identification of cartilage.Fins were then hydrated in a graded seriesof ethanols for 30 min each and subse-quently bleached in a 1:9 part solution of 3%hydrogen peroxide to 1% potassium hydrox-ide under direct light to enhance and acceler-ate the bleaching procedure. Clearing anddigestion of adhering tissues were accom-plished in a trypsin-buffer (3% solution oftrypsin) solution ten times the volume of thespecimen, and subsequently stained with a1% solution of alizarin red in sodium borate.This latter stain is specific for bone. Extrane-ous dye was removed in one to two changesof 1% potassium hydroxide and glycerine.Fins were stored in a 100% glycerine solu-tion with thymol crystals added.
Light microscopyA total of 20 fins, representing samples
from 11 fish (35–70 cm) were processed forroutine light microscopy. With most of thesamples used, both fixation and decalcifica-tion of entire fins were achieved by storingindividual samples in Bouin’s fixative (Hu-mason, ’67) for 6 months to 1 year. In somecases, however, fins were fixed in 10% buff-ered formalin and were decalcified in solu-tions of 1N nitric acid, 4N formic acid, or0.2M ethylene diamine tetraacetic acid(EDTA, Sigma Chemical Co., St. Louis, MO).Decalcification of fins varied between hoursto days and was dependent upon the solu-tion and the size of the sample used. A simpleexperiment was carried out to determinewhich decalcifying solution best demineral-ized the fin-ray bone in the shortest timepossible. Complete decalcification of bonewas indicated using an ammonium oxalatesolution (Clayden, ’52). A few drops of thedecalcifying solution were placed into theammonium oxalate. If a precipitate formed,the decalcifying solution was replaced. If theammonium oxalate solution remained clear,however, this indicated decalcification wascomplete. All tissues were then transferredinto 70% ethanol for storage. In thosesamples exposed to Bouin’s for prolongedperiods of time, however, lithium carbonate(Fisher Scientific, Fairlawn, NJ) was addedto the ethanol to remove excess picric acid.
Following fixation, tissues were dehy-drated in a graded series of ethanols, cleared
in Histoclear (National Diagnostics, Man-ville, NJ, or Diamed Lab Supplies Inc., Mis-sissauga, Ont.) and infiltrated and embed-ded in Tissue Prep paraffin (m.p. 56.6°C,Fisher Scientific). Blocks of tissue were seri-ally sectioned in transverse and horizontalplanes, 5–8 µm in thickness. All sectionswere mounted on pre-cleaned slides (Basergaand Malamud, ’69) with dilute egg albumen.Dried slides were then deparaffinized in His-toclear, rehydrated and stained withEhrlich’s hematoxylin, and counterstainedwith eosin (Sheehan and Hrapchak, ’80),dehydrated in ethanol, cleared in Histoclear,and mounted with Permount (Fisher Scien-tific). In order to determine the tissue con-stituents of the fin-ray annulus, a number ofother staining procedures were also em-ployed such as Feulgen’s, Masson’s tri-chrome, periodic acid-Schiff, and MalloryHeidenhain (Sheehan and Hrapchak, ’80).
Scanning electron microscopyPectoral fins of trout, Oncorhynchus
mykiss, were used for preliminary studies toestablish proper techniques for scanningelectron microscopy. At that time, individualrays were dissected and as much adherentsoft tissue as possible was removed. Eachcross-sectional area was then filed andetched. Filing was achieved by the use ofsandpaper that varied in degrees of gritthickness. Etching was accomplished by theuse of a 5–10% solution of EDTA followed bya 5–10 % solution of sodium hypochlorite(NaOCl, Mallinckrodt, Paris, KY), an or-ganic dissolving agent. Rays remained inthese solutions at varying times between1–20 min. Following the etching procedure,rays were dehydrated in a graded series ofethanols and allowed to air dry for 24–48 hr.In some cases samples were dehydrated andexposed to a drying agent, hexamethyldisila-zane (HMDS) according to Nation (’83) andthen allowed to air dry for 15 min to 1 hr.Small samples were dehydrated in a gradedseries of acetones and critical point dried incarbon dioxide.
A modification of the above procedure wasused on fins of chinook salmon, in order tomaintain the integrity of the bone matrixand the surrounding connective tissues.Three to five fin-rays were left intact and cutaway from the remaining fin along the lengthof the ray. Each sample was then dehy-drated in a graded series of ethanols, post-fixed in 2% osmium tetroxide in a phosphatebuffer (pH 7.4) for 1 hr, and completely infil-
SALMON FIN-RAY ANNULUS 299

trated and embedded in a modified Epon/Araldite resin (Luft, ’61). To embed fin-raysfor further processing, disposable 10-cc sy-ringes were used (Becton-Dickinson, Ruther-ford, NJ). These were filled with resin andthe fin-ray placed parallel to the length ofthe syringe.
Once polymerization of the plastic wascomplete, fin-rays were cut in cross sectionusing a Buehler Isomet diamond saw (Tec-Met Inc., Toronto, Ont) to approximately 1.9mm in thickness. Cutting and subsequentpreparation were carried out at the Depart-ment of Geology, University of Toronto. Sur-face preparation was accomplished initiallyusing 9-µm diamond spray on Pan-W paperfor 1 hr at approximately 30 revolutions permin (R.P.M.) on a Buehler 8-inch, reciprocal,variable-speed polisher. Subsequently, 1 and0.025 µm diamond paste were placed onPan-W paper for 1 hr and 30 min, respec-tively, at 15 R.P.M. Following each polishingprocedure, surfaces were cleaned with 95%ethanol. Etching of fin-ray cross-sectionalsurfaces was then accomplished by the im-mersion of fin-ray tissues in various solu-tions such as EDTA, Baxter decalcifying so-lution (Baxter, McGraw Park, IL), or Decal(National Diagnostics, Manville, NJ).
Fin-rays to be examined using the HitachiS-530 scanning electron microscope weremounted on aluminum specimen mounts andwere gold coated using a Polaron PS3 Sput-ter Coater. To examine differences in densi-ties within the fin-rays, some samples wereexamined using a JOEL (Peabody, MA) 820scanning electron microscope at The Hospi-tal for Sick Children, Toronto. In this case,secondary electrons were used and sampleswere coated with carbon using the Dentonbench top Turbo Carbon Coater.
Preparation of dried fin-rayand scale samples
The following procedures were carried outby the technicians in a special Fish AgeingLaboratory located at the Pacific BiologicalStation, Department of Fisheries and Oceansin Nanaimo, B.C. The techniques explainedbelow can be found in manuals written byMacLellan (’87) and York (’87) and in anarticle by Chilton and Beamish (’82).
Fins were removed using bone cutters,ensuring that the fin-ray bases were in-cluded. Prior to air drying, fins were placedin 14.5 kg Kraft ungummed envelopes mak-ing certain the cut edge of each fin wasplaced at the open end of the envelope while
the cut surface remained perpendicular tothe long axis of the rays.
Depending upon ambient temperature andhumidity, fins were left to dry for up to 3weeks. Fins were then trimmed and excessrays and adhering tissues removed. Epoxy(Dexter Corporation, Vancouver, B.C.) wasthen applied smoothly up to approximately3.5 cm from the basal area of the fin toprovide rigid support for sectioning. Epoxiedfins were allowed to dry for 3–4 days, afterwhich time they were sectioned using a Bron-mill sectioning machine to approximately1.3–1.5 mm in thickness. Sections were thenplaced on glass slides and covered with Flo-texx liquid mounting medium (Lerner Labo-ratories, Pittsburgh, PA.) Dried fin-raysamples were analyzed using either re-flected or transmitted light in compound orstereo-microscopes.
As mentioned previously, all scales wereremoved from fish according to a designatedarea outlined by the International NorthPacific Fisheries Commission (’59). This des-ignated area is usually above the lateral lineon a diagonal from the posterior insertion ofthe dorsal fin toward the origin (anteriorinsertion) of the anal fin. Upon removal,scales were checked for deformation and re-generation. If these latter features were pres-ent, the scale was discarded and anotherscale removed. Scales, with their rough sidesfacing upwards, were then placed in specificscale booklets possessing pages with an ad-hesive coating. All information was then re-corded in this booklet. Scales were then ex-amined using an Eberbach microprojectorwith a 40X magnification or on a microfichereader-printer.
Interpretation of fish age estimates wasbased upon the identification of an annulus.In dried fin-ray sections this interpretationis based upon the presence of translucent orhyaline zones and in scales by areas ofcrowded circuli. All ages were determinedfrom the scales by highly trained individualsat the ageing laboratory in Nanaimo. Alongwith the data on scale ages, slides of dried,fin-ray samples were also sent to the labora-tory at the University of Toronto at Scarbor-ough and photographed according to themethods described above for light micros-copy.
RESULTSTerminology
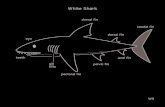
Figure 1 defines the terms presented inthis section and how they refer to the posi-tion and location of fin-rays within the pecto-
300 L.C.G. FERREIRA ET AL.

ral and dorsal fins of the chinook salmon.These terms include anterior, posterior,proximal, distal, and medial and lateral. Tofurther clarify these, ‘‘anterior’’ and ‘‘poste-
rior’’ refer to the rays only, while the terms‘‘rostral’’ and ‘‘caudal’’ refer to the fish as awhole. The terms ‘‘proximal’’ and ‘‘distal’’will indicate the region of the ray closest to,
Fig. 1. Diagrammatic representation of the terms used with reference to the position andlocation of the fin-rays within the pectoral (lower enlargement) and dorsal (upper enlargement)fins of the chinook salmon, Oncorhynchus tshawytscha. (Modified from Geerlink, ’89.)
SALMON FIN-RAY ANNULUS 301

or farthest from, respectively, the point ofarticulation of the fin to the body cavity.Rays are numbered starting from their mostanterior position within the fin. The terms‘‘medial’’ and ‘‘lateral’’ are defined in the fol-lowing section on fin-ray morphology.
In scales, regular configurations or concen-tric ridges are referred to as circuli. Thesecirculi are arranged in such a way as to formtwo distinct areas consisting of alternatingwide and closely spaced bands. Zonation inother structures such as otoliths, dried fin-rays, vertebrae, and opercular bones refersto a difference in the degree of translucencyor opacity of the particular tissue whenviewed with specific forms of light. Translu-cent zones when observed using transmittedlight appear bright in section (dark in reflec-tive light) in contrast to opaque zones thatappear darker (bright in reflective light).Annuli or yearly deposited rings are identi-fied by abrupt changes associated with therelatively uniform appearance of the calci-fied tissue being examined. In scales, theannulus appears associated with an obviousbreak in the continuity in the configurationof circuli, and has often been referred to as acheck. The annulus of other calcified struc-tures is associated with the translucent zone.Checks or zones are usually formed prior toperiods of reduced growth or cessation ofgrowth (winter growth) and are separatedby annuli from what is referred to as growthzones. These zones are associated with fasterperiods of growth (summer growth) and, inscales, appear as continuous, widely spacedcirculi and in other calcified tissue as opaquezones (Casselman, ’74, ’83, ’87). Since theannulus in this particular investigation isconsidered to correlate with a yearly event,the term band will be used to describe aconcentric mark or feature deposited in thebone matrix.
General morphology and arrangementof fin-ray components
The number of fin-rays varied between12–19 for the pectoral fin and 13–17 for thedorsal fin. A single fin-ray is actually com-posed of two parallel bony elements (Fig.2A), often referred to by previous investiga-tors as hemitrichia, hemisegments, or demi-rays (Lanzing, ’76; Geerlink, ’89). We usehemisegments with reference to the bonyelements of both the pectoral and dorsalfins. Hemisegments are apposed to one an-other and are referred to as ‘‘medial’’ or
‘‘lateral’’ depending upon their position inthe fin (Fig. 2A).
The following description includes dia-grams and complementary light micrographsof tissue sections along the fin-ray (Figs.2A–D, 3, 4). Hemisegments are constructedso that their shafts have an unsegmentedproximal part and a flexible, segmented dis-tal region. The latter area is composed ofplate-like segments, which, upon macro-scopic examination, appear as horizontalstriations along the greater part of the lengthof each ray. Proximal and distal units jointogether to form a single supporting rod, ahemisegment (Fig. 2A). When the fin wasviewed in cross section, a channel of connec-tive tissue was found between the concavesurfaces of the crescent-shaped medial andlateral elements of each fin-ray. This region,referred to as the intersegmental region(Lanzing, ’76), is characterized by the pres-ence of nerve bundles, blood vessels, andfibrous connective tissue (Fig. 2A,D). Nervebundles observed in this region consist ofboth myelinated and unmyelinated fibersenclosed by the cytoplasm of Schwann cells.Nearby blood vessels were identified as arter-ies and veins.
At their most proximal extremity, the pec-toral rays are supported by the radial bonesof the pectoral girdle. The distal scapularradial supports the anterior-most ray di-rectly and four large radials support theremainder of the rays (Fig. 3). Hemiseg-ments of the dorsal fin diverge at the distal
Fig. 2. A: Diagrammatic representation of medial(mh) and lateral (lh) dorsal hemisegments in Oncorhyn-chus tshawytscha (modified from Lanzing, ’76). Theshafts of the hemisegments have an unsegmented proxi-mal part that embraces the radial cartilage (c) and aflexible segmented distal region viewed as horizontalstriations (arrows). The latter area is composed of plate-like segments (*). Transverse sections of the fin-rayreveal a channel of connective tissue, the intersegmen-tal region (I). Horizontal lines (b,c,d) are approximateregions represented in subsequent micrographs (B–D).B: Transverse section through the most distal aspect ofthe radials shown in A. Note the presence of adiposetissue (A) surrounded by bone (arrows). ct, fibrous con-nective tissue. C: Chondrogenic tissue (C) representa-tive of the radial hyaline cartilage characterizes region Iof the pectoral and dorsal fins. Bone tissue (arrows) isreduced from that demonstrated in B. M, muscle. D:Transverse section through the intersegmental regionlocated between the concave surfaces of the crescent-shaped medial (mh) and lateral (lh) pectoral hemiseg-ments. This region is characterized by the presence ofnerve bundles (arrows), arteries (a), veins (v), and con-nective tissue (ct). Scale bars 5 100 µm for B, C, and D.
302 L.C.G. FERREIRA ET AL.

Figure 2
SALMON FIN-RAY ANNULUS 303

Fig. 3. Diagrammatic representation of the pectoral fin of the chinook salmon, Oncorhyn-chus tshawytscha, and its association with the underlying radials (r). Views of transversesections of the fin (I–VI) along their long axis and the various shapes and types of tissuecomponents are depicted.
304 L.C.G. FERREIRA ET AL.

Fig. 4. Diagrammatic representation of the dorsal fin of the chinook salmon, Oncorhynchustshawytscha, and its association with underlying radials (r). Views of transverse sections of thefin (I–V) along their long axis and the various shapes and types of tissue components aredepicted.
SALMON FIN-RAY ANNULUS 305

margin of the endoskeletal radials (Figs. 2A,4). Following digestion of both fins with buff-ered trypsin, pectoral and dorsal hemiseg-ments were observed embracing cartilagi-nous tissue overlying the radials (Figs. 3, 4).The radials articulate with the pectoral anddorsal hemisegments and they also articu-late with the underlying pectoral girdle andvertebral column, respectively.
The general shape of hemisegments is vari-able along the length of the pectoral anddorsal fins. For this reason, both fins weredivided into regions along their long axis.Each transverse serial section could then beclassified into a specific region that wouldindicate to the reader from which area alongthe fin the section was taken. The reader isreferred to a series of diagrams, presented inFigures 3 and 4. Figure 3 illustrates six(I–VI) regionalized areas of the pectoral fin,whereas, five (I–V) regions are demonstratedin the dorsal fin (Fig. 4). Light micrographs(Figs. 2B, 5B–8B) corresponding to the spe-cific regions represented in the diagrams(Figs. 3 and 4), are used to clarify the compo-nents characteristic of each area. Diagramsand micrographs are used in conjunctionwith one another to describe these regions indetail below.
Region I of the pectoral fin is presenteddiagrammatically in Figure 3 (3I), and isrepresented in Figures 2B and C. Distinct,centrally positioned, groups of hyaline carti-lage are characteristic of this region. In thecorresponding stained tissue sections, thematrix of this cartilage appears strongly ba-sophilic and chondrocytes form isogenouscell groups. Spicules of bone in the periph-eral sections of this chondrogenic tissue rep-resent the most distal aspects of the underly-ing radials (Fig. 2B). Region I of the dorsalfin represents the same area just above thepoint where rays embrace the cartilaginouspads of tissue belonging to the dorsal radi-als. The absence of bony tissue in this regionof the dorsal fin (Fig. 4I) simply indicatesthat the level of sectioning of this fin beganmore distally than that of the pectoral fin.Cartilage was displayed as discrete groupsof chondrogenic tissue bordered on eitherside by muscle (Fig. 4I).
Region II of the dorsal and pectoral finsdemonstrates similar tissue components toregion I and is representative of the raysjust above their most proximal extremity.Pads of cartilaginous tissue are still presentin these regions (Figs. 3II, 4II). Adjacent to
the cartilaginous pads, are areas of denseconnective tissue, of an uncertain nature,but that appear to be composed of fibroblastsand small groups of chondrocytes amongnumerous collagen fibers in both the dorsal(Fig. 5A,B) and pectoral (Fig. 5C,D) fins.Their appearance is characteristic of fibro-cartilage tissue found in other vertebrates.For this reason, it was referred to as a ‘‘fibro-cartilage mass.’’ Sometimes an area of con-densed, bony tissue is present within thefibrocartilage mass (Fig. 6A,B). We believethis bony element represents the most proxi-mal portion of the hemisegment. In the sameregion, dorsal fin hemisegments first appearon the medial aspect of the fin as irregularlyshaped structures (Fig. 4II) and progressdistally to an elongated form similar to thatof the pectoral fins (Fig. 6C). It seems thatthe appearance of the medial hemisegmentprior to those on the lateral side is due tolength differences between the two hemiseg-ments. The most consistent feature of regionII in both fins is the presence of centrallylocated groups of hyaline cartilage, denseconnective tissue areas that, more distally,appear to give rise to bony structures, andthe continuously changing shape of fin-raysthat are already visible.
Cross-sectioned, medial fin-ray elementsof the pectoral fin, in regions III to V areroughly horseshoe shaped (Fig. 3III, IV, V)when compared with corresponding lateralhemisegments.Although, the latter vary con-siderably in their form, they are, for themost part, roughly crescent-shaped. We re-fer to extensions of the main body of eachhemisegment as ‘‘arms.’’ Arms are furtherdescribed as ‘‘major’’ or ‘‘minor’’ reflectingdifferences in size (Fig. 6D). The pectoral
Fig. 5. A: A transverse section through region II ofthe dorsal fin of an adult salmon, Oncorhynchus tshaw-ytscha, representing the most distal aspect of the under-lying radial cartilage. Directly adjacent to the pad ofcartilage (C), are dense areas of connective tissue consist-ing of discrete masses of fibrocartilage (fm). B: Highmagnification of three fibrocartilage masses (fm) ob-served in A, showing their compact and hyalinized na-ture compared to the surrounding fibrous connectivetissue (ct). C, hyaline cartilage. C: Transverse section ofregion II of the pectoral fin representing the distalextremity of the radials and showing an area of fibrocar-tilage (fm) tissue adjacent to the radial cartilage (C),muscle (M), adipose tissue (A), and bone (arrows). D:High magnification of fibrocartilage (fm) observed in C,revealing cells (arrows) among numerous collagen fibers(arrowheads). A, adipose tissue; C, hyaline cartilage; M,muscle. Scale bars for A to D 5 100 µm.
306 L.C.G. FERREIRA ET AL.

Figure 5
SALMON FIN-RAY ANNULUS 307

Figure 6
308 L.C.G. FERREIRA ET AL.

arrangement of the two apposing hemiseg-ments is asymmetric in these regions, sincemedial hemisegments are always larger thanlateral ones (Figs. 3III to VI, 7A–D). Themajor and minor arms of the medial hemiseg-ment are more clearly defined than in thecorresponding lateral hemisegments. In re-gion III of the pectoral fin, a cone-shaped,well-delimited structure attaches to the outermedial hemisegment of each fin-ray (Fig.6D). This tissue appears similar to the ‘‘fibro-cartilage mass’’ previously identified in re-gion II. The fibrocartilage tissue consists ofan extensive collagenous network surround-ing small areas of hyaline cartilage and at-taches to the medial hemisegment along aline where collagenous fibers penetrated thebone from the periosteal area (Fig. 7A,B).This basophilic mass appears to originate inthe dermis at the proximal end of the fin anddiminishes gradually towards the distal tipof the ray (Figs. 3IV, 7C,D). The intersegmen-tal region is not well developed in region IIIof the pectoral fins since the loss of cartilagi-nous elements results in a large space be-tween apposing hemisegments (Fig. 3III).
The main difference in hemisegments inregion IV of the pectoral fin from those ofregion III is a slight change in their shape.Rays are considerably larger in region IVand the intersegmental region between theapposing hemisegments is well vascularizedwith arteries and veins and contains bundlesof nerve fibers, collagen fibers, and adipose
tissue. In regions V and VI of the pectoralfin, the reduction in size of the fibrocartilagemass and the intersegmental region is appar-ent (Figs. 3V,VI, 7C,D). The medial hemiseg-ments of these regions are less attenuatedand smaller while the lateral hemisegmentsare elongated and crescent-shaped.
Region III of the dorsal fin is character-ized by well-formed medial hemisegmentsand adjacent connective tissue masses (Figs.4III, 6A). Medial hemisegments of the dorsalfin, as in the pectoral fin, are also horseshoe-shaped. In addition, condensed tissue, likelybone, is apparent (Fig. 6B). In this region aswell, a large space exists where cartilagi-nous elements had been present in moreproximal areas (Fig. 4III), probably a conse-quence of tissue displacement during section-ing (Fig. 4III).
In region IV of the dorsal fin, hemiseg-ments are symmetric in distribution andshape (Fig. 8A,B).Although nerves and bloodvessels are present within the intersegmen-tal regions, this area appears less vascular-ized than the corresponding region in thepectoral fin. Collagen fibers are also presentin the areas surrounding the hemisegmentsbut the fibrocartilage mass observed in re-gions III and IV of the pectoral fin is notpresent. Region V is characterized by reduc-tion in size and shape of hemisegments withthe lateral and medial hemisegments hav-ing a distinct U-shaped or arched appear-ance (Fig. 4IV,V).
Fin-ray annuliLight microscopic preparations of trans-
versely sectioned rays from adult tissueclearly demonstrate the presence of narrowdark (purple) bands alternating with widelight (purple) bands. This pattern was onlyvisible when fins were fixed and decalcifiedsimultaneously with Bouin’s fixative andsubsequently stained with Ehrlich’s hema-toxylin and an eosin counterstain (Fig. 8A,B).Initially various decalcifying agents weretested to determine which solution best de-calcified the bone tissue in the least amountof time but still might express an alternatebanding pattern. All efforts were made toensure the size of the tissues were similar (3mm3), that temperature remained constant,and that solutions were changed frequently.Complete decalcification was determined ac-cording to Clayden (’52) (see Materials andMethods) with the exception of those tissuesdemineralized in Bouin’s fixative. In this
Fig. 6. A: A transverse section taken from the dorsalfin of an adult salmon, Oncorhynchus tshawytscha. Thisarea represents the most distal aspect of region II andthe most proximal aspect of region III. The medialhemisegments (mh) are characteristically horseshoeshaped while the lateral hemisegments (lh) are observedas sites of condensed tissue. A space (s) is now in the areaformerly occupied by the radial cartilages. A, adiposetissue; ct, fibrous connective tissue. B: High magnifica-tion of a lateral hemisegment in A. From the site ofcondensed tissue a definitive lateral hemisegment willdevelop through the process of periosteal osteogenesis.Note the presence of osteoblasts (arrowheads) in theperiosteum (p) and osteocytes (arrows) surrounded bynewly formed bone matrix (b). ct, fibrous connectivetissue. C: Region II of the pectoral fin has radial carti-lages (C), elongate medial hemisegments of bone (mh)and fibrocartilage (fm). ct, fibrous connective tissue. D:A transverse section through the medial hemisegmentin region III of the pectoral fin from an adult salmon.Note the major (MA) and minor (ma) arms and thecone-shaped, well-delimited fibrocartilage mass (fm) at-tached to the outer portion of the hemisegment. A,adipose tissue; p, periosteum; arrows, osteocytes. Scalebars for A to D 5 100 µm.
SALMON FIN-RAY ANNULUS 309

Figure 7
310 L.C.G. FERREIRA ET AL.

latter case, the endpoint of decalcificationwas determined mechanically by piercingthe bone with a pin. The data showed thatthe major factor responsible for the demon-stration of these bands (when all tissueswere stained with hematoxylin and eosin)was the Bouin’s fixative and the length oftime tissues spent in this solution. Bandswere present in different sized adult tissues(3 mm3, single fin-ray, entire fin) fixed inBouin’s fluid for 60, 93, and 275 days, respec-tively. No bands were found in stained sec-tions following fixation in 10% buffered for-malin and decalcification in either nitric acid,formic acid, EDTA, Von Ebners, or Richman-Gelfand-Hill solutions. It is also noteworthythat staining of discrete bands was not ob-served with any of the juvenile samples ex-amined, even after extended exposure to theBouin’s fixative.
Alternating dark and light bands in adultsamples corresponded to narrow translu-cent and wide opaque zones, respectively,when dried unstained sections were exam-ined through transmitted light (Figs. 8C,D,9A,B). Since the annulus had been associ-ated with the translucent zone of driedpreparations (Casselman, ’78) and the darkbanding of the fixed, stained tissue coin-cided with this translucent area, the darkbands were recognized as annuli. In mostcases, several dark bands, grouped togetherin stained preparations corresponded to asingle translucent zone. Bands were presentin the form of opaque and translucent zoneswhen back-scatter SEM was used (Fig. 9C).In general, the opaque zone was approxi-mately four times greater than adjacenttranslucent zones.
Staining methods, other than hematoxy-lin and eosin, were utilized in an attempt toelucidate the structural composition of theannulus. Sections of bone decalcified inBouin’s fixative were stained specifically forthe presence of DNA using Feulgen’s stain,collagen using Masson’s trichrome stain, andsimple polysaccharides of the ground sub-stance (glycosaminoglycans) using the peri-odic acid-Schiff technique. Only tissuesstained with hematoxylin and eosin demon-strated a banded appearance, indicating thatthese bands were not composed of polysac-charides or DNA. Although banding was ab-sent with the Masson’s trichrome procedure,the entire matrix stained homogeneously forcollagen.
Banding of hemisegments appeared alongthe entire length of the ray throughout allthe regions outlined above but often thestaining intensity of these bands varied. Inmany instances, the central (core body) andperipheral (beneath the periosteum) bandsstained darker than bands found in otherregions of the matrix (Fig. 10A). Not onlywas staining inconsistent, but the number ofbands also varied between individuals andbetween the regions examined. Banding waspresent in the most proximal aspect of therays in regions II and III. As the distalaspects of the hemisegments were ap-proached in regions IV, V, and VI of thepectoral fins and in regions IV and V of thedorsal fins, the number of bands was re-duced. In many cases, only the core body wasobserved in these regions, appearing as abilaterally curved structure close to theperiphery of the ray (Fig. 10B,C). Region IVof the medial hemisegment in the pectoralfin displayed the greatest number of bandsthroughout the entire ray. Up to twelve bandswere counted on the major arm of one par-ticular fin-ray from a four-year old salmon(Fig. 10C). In general, the number of bandswas greater in the medial hemisegments ofthe pectoral fin than in the medial hemiseg-ments of the dorsal fin. In many cases, thevariance in band number may have beendue to the differing ages of the individualswhen the fins were removed. An individualthat was determined as being 2 years oldusually exhibited a number of bands groupedtogether (annuli), usually in the central andperipheral areas of the ray. This banding inturn, corresponded to two translucent zonesin dried fins (Fig. 9A,B). A fish that was 3years of age displayed an additional series of
Fig. 7. Fins of chinook salmon, Oncorhynchus tshaw-ytscha. A: High magnification of the cone-shaped massobserved in Figure 6D. This tissue resembles the fibro-cartilage masses described in region II and consists of anextensive network of collagen (arrows), fibroblasts (f),and chondrocytes (arrowheads). B: Light micrograph ofthe horseshoe shaped medial (mh) and the crescentshaped lateral (lh) hemisegments in region IV of thepectoral fin. A dense cord of fibrocartilage tissue (arrow)attaches to the outer edge of the medial ray and anintersegmental region (I) is present between the hemiseg-ments. C: A transverse section through region V of thepectoral fin showing a less attenuated medial hemiseg-ment (mh), more elongate lateral hemisegments (lh),and a reduced intersegmental region (I). D: Region VI ischaracteristic of the distal portion of the pectoral fin andshows elongated medial (mh) and lateral (lh) hemiseg-ments with an intervening intersegmental region. Scalebars for A 5 50 µm, for B– D 5 100 µm.
SALMON FIN-RAY ANNULUS 311

Figure 8
312 L.C.G. FERREIRA ET AL.

bands when compared to 2-year-old individu-als (Fig. 8C,D). In this case, dried fin samplesexhibited a third translucent zone (Fig. 8D).Even when rays were sectioned longitudi-nally or obliquely, they continued to demon-strate groups of dark bands whose numberswere indicative of the age of the animal.
DISCUSSIONMorphology of the fin-ray
of the chinook salmonThis study demonstrates that the number
and arrangement of rays in both pectoraland dorsal fins, and their association withthe underlying radials are consistent withprevious descriptions (Hikita, ’62) on fin-raymorphology in the genus Oncorhynchus (Sal-monidae). Many of the morphological fea-tures documented in studies of the three-dimensional organization of fin-rays withinthe fins of various chordate species (Arita,’71; Lanzing, ’76; Potthoff and Kelley, ’82;Geerlink, ’89; Holland and Holland, ’91),were also observed in the fins of the chinooksalmon. However, the present study pro-vides new insight into the histological struc-ture of the salmon fin-ray.
The cartilaginous areas that appeared inregions I and II of the pectoral and dorsalfins have also been described in other spe-cies of Pacific salmon as a pad of subcartilagi-nous tissue intercalated between the bonesof the radials and the proximal ends of thefin-rays (Hikita, ’62). In the current study,the diverging halves of each ray were ob-served to embrace this pad of tissue at theextremities of the scapula and the four radi-
als of the pectoral fin, and the radials of thedorsal fin. This same arrangement of fin-rays to radial cartilages exists in the pecto-ral fins of blennies (Brandstatter et al., ’90).The pectoral fins of Coris formosa also havean elongated fibrocartilage pad into whichthe distal ends of the radials fit and eachbase of the medial and lateral hemiseg-ments articulate (Geerlink, ’83). The latterfibrocartilage structure resembles the sub-cartilaginous pad described earlier insalmon. In the dorsal fins of tuna and sword-fish, similar cartilaginous elements of theunderlying radials have been described asarticulating with the proximal ends of theoverlying hemisegments (Potthoff, ’74;Potthoff and Kelley, ’82).
The fin-rays of chinook salmon are con-structed so that each hemisegment has anunsegmented proximal part and a flexibledistal region, consisting of small plate-likesegments. This structural composition hasbeen observed in the fin-rays of many fishes(Arita, ’71; Lanzing, ’76; Brandstatter et al.,’90) and is consistent with the definition of asoft ray (Prince and Pulos, ’83). It is believedthat this construction permits bending ofthe rays (Geerlink and Videler, ’87).
The fibrocartilage mass, attached to theouter edge of the medial hemisegment, is anunusual feature present only in regions IIIand IV of the pectoral fin. A similar struc-ture, referred to as the lepidotrichal cord inblennies, is also located along the free edgeof the outer medial hemisegment graduallydiminishing towards the distal tip (Brand-statter et al., ’90). The lepidotrichal cord iscomposed of fibrocytes, collagenous fibers,elastic fibers, and an alcian blue-positiveextracellular matrix. It has been suggestedthat it contributes to the stiffness of the bonyfin-rays. Since in chinook salmon the fibro-cartilage mass occurs only in the proximalportions of the medial ray, this structuremay also impart stiffness to this region ofthe fin-rays. That these masses are not pres-ent in dorsal fins may be a reflection offunctional differences in the various fins. Ithas been documented that pectoral and pel-vic fins act as swim paddles whereas thedorsal fin may act only to align the fish in itsmovements (Romer and Parsons, ’86).
The histology and ultrastructure of mam-malian bone have been well documented(Cameron, ’72; Robinson et al., ’77), provid-ing most of the information that exists onthe structure of vertebrate bone. Prior to the
Fig. 8. Fins of chinook salmon, Oncorhynchus tshaw-ytscha. A: Transverse section of the dorsal fin represen-tative of region IV. Note the symmetrical shape of me-dial (mh) and lateral (lh) hemisegments and the presenceof an intersegmental (I) region. B: High magnification ofa region in A demonstrating two opposing hemiseg-ments. The intersegmental region (I) consists of nervebundles (n), collagen fibers (arrows), and adipose tissue(A). At the tips of the major arm (MA) the periosteum (p)is visible and annuli (arrowheads) are also present. ma,minor arm. C: Light micrograph of a medial hemiseg-ment removed from the pectoral fin of a 3-year-oldsalmon and stained with Ehrlich’s hematoxylin and aneosin counterstain. The presence of three distinct, darklystained bands (annuli) are observed (a1, a2, a3). The corebody (cb) is located in the central region of the ray. D:Dried preparation of a medial hemisegment correspond-ing to C. Both rays were removed from the same fish.Note the presence of alternating wide opaque and nar-row translucent bands. The latter type represents an-nuli. Three annuli (a1, a2, a3) are visible. cb, core body.Scale bars for A 5 200 µm, for B– D5 100 µm.
SALMON FIN-RAY ANNULUS 313

Figure 9
314 L.C.G. FERREIRA ET AL.

present study on the nature of demineral-ized bone of chinook salmon fin-rays, fewstudies had focused on the histological struc-ture of teleost bone or on its mode of develop-ment. Moss (’61a,b, ’63), and more recentlyWeiss (’78), provided the most comprehen-sive studies on teleost bone. Histologicalstudies of teleost bone have demonstratedthat bone forms beneath periosteal tissueand endosteal tissue within a cartilaginousmatrix (Norris et al., ’63). These sites of bonedevelopment coincide with three distinctanatomic types of bone, namely, primarybone (non-lamellar), endosteal or secondarybone (lamellar), and endochondral or tra-becular bone. Our companion study usesetching techniques and extracellular matrixdigestion in conjunction with both scanningand transmission electron microscopy to de-scribe the nature of the bone in the fin-raysof chinook salmon. In addition, the compan-ion study shows that the bone in fin-rays ofchinook salmon forms through periosteal os-teogenesis and that the presence of enclosedosteocytes within the matrix, may suggestthat this piscine bone is a cellular (osteo-cytic) type. However, the present study alsoprovides some evidence of periosteal osteo-genesis and also of bone formation from pre-chondrogenic precursors in region I of thepectoral fins in the chinook salmon. Thisarea likely represents the most distal aspectof the cartilaginous joints of the underlyingradials. The identity of cartilage was clearlydemonstrated in whole fins from enzymedigestion, clearing techniques, and alcianblue. Previously, Geraudie and Landis (’82),when describing the embryonic develop-ment of the pelvic fin of trout, stated thatradials develop from an underlying cartilagi-nous girdle that stains with alcian blue. This
cartilaginous girdle is considered analogousto the subcartilaginous tissue described inother Pacific salmon (Hikita, ’62) and to thecartilage tissue observed in region I of thepectoral and dorsal fins of the chinooksalmon.
In the current study, the presence of hyper-trophic chondrocytes within the cartilagi-nous matrix and the surrounding bone tis-sue in region I of the pectoral fins impliesthat radials develop through a process resem-bling endochondral ossification. A similardevelopmental process, chondroid osteogen-esis, has also been described in the articularareas of the head in bony fishes (Moss, ’61b).These areas are considered similar to mam-malian epiphyseal cartilage where mineral-ization is achieved through the deposition ofcalcium salts following cell hypertrophy ofchondroid tissue. The resulting tissue hasbeen described as intermediate in form be-tween notochordal tissue and hyaline carti-lage.
Similar areas of cartilaginous tissue arepresent in region I of the dorsal fin; however,unlike region I of the pectoral fin, spicules ofbone tissue, representing the underlying ra-dials, were not observed. The different pro-files in region I of the pectoral and dorsalfins may be explained by the way in whichthe dorsal fin was removed from the adja-cent body cavity or simply may be due to thelevel at which sectioning of the fin began.
Hemisegments were first observed in re-gion II of the pectoral and dorsal fins adja-cent to the cartilaginous joints of the under-lying radials in dense masses of whatappeared to be fibrocartilage tissue. Al-though there is no documented evidence sug-gesting that bone formation occurs from fi-brocartilage, these areas nevertheless appearto offer the appropriate environment for theinitial sites of hemisegment development.The cytoplasmic basophilia of cells surround-ing small spicules of bone in region II of thepectoral fin and regions II and III of thedorsal fins implies osteoblastic activity insynthesis of protein necessary for the proteo-glycan component of bone. This activitywould result in the initial formation of rayelements through periosteal osteogenesis.Since teleost bone of periosteal origin is re-ferred to as primary or fibrillar bone (Norriset al., ’63), this view of periosteal osteogen-esis is reinforced by the presence of an abun-dance of collagenous fibrils within these re-gions. According to Moss (’61b), during
Fig. 9. A: Light micrograph of a medial hemiseg-ment, removed from the dorsal fin of a 2-year-old salmon,Oncorhynchus tshawytscha, demonstrating alternatingdark and light bands following hematoxylin and eosinstaining. The core body (cb) and two annuli (a1, a2) arepresent. B: This micrograph demonstrates a dried, un-stained medial ray in transverse section, viewed throughtransmitted light. Narrow translucent and wide opaquezones correspond to dark and light bands viewed in A,respectively. The core body (cb) and two annuli (a1, a2)are visible. Scale bar 5 C: Backscatter scanning elec-tron micrograph demonstrating translucent and opaquezones within the cross-sectional surface of a hemiseg-ment. Translucent areas (arrowheads) were consideredto be analogous to the annuli described in other prepara-tions of fin-ray bone. cb, corebody. Scale bars for A andB 5 100 µm, for C 5 25 µm.
SALMON FIN-RAY ANNULUS 315

Figure 10
316 L.C.G. FERREIRA ET AL.

periosteal osteogenesis in fish, osteoblastsaid in the production of consecutive layers ofbone by the deposition of collagen fibrils.
Periosteal osteogenesis is considered themajor mode of bone development in teleosts(Moss, ’61b). Evidence of this type of bonedevelopment has also been documented inembryological studies on the pectoral, pel-vic, and caudal fins of rainbow trout, On-corhynchus mykiss. Fin-ray ontogenesis oc-curs by direct mineralization of thecollagenous basal lamella of the skin with-out passage through a cartilaginous precur-sor (Geraudie and Landis, ’89). In an earlierembryological study of pelvic fin develop-ment in trout, hemisegments were stainedwith alizarin red but not with alcian blue(Geraudie and Landis, ’82). Since alizarinred stains specifically for bone, they con-cluded that these dermal structures arosewithout an endochondral sequence. Thesereports, when considered in conjunction withour present observations, strongly suggestthat endochondral ossification or chondroidosteogenesis does not play a role in the devel-opment of fin-rays in chinook salmon, sincewe observed fin-rays arising from the activ-ity of osteoblasts. Thus, the mode of fin-raydevelopment differs from that of radial for-mation, since the latter appears to arisefrom cartilaginous tissue.
According to Landis and Geraudie (’91),after fin-ray development commences in thepectoral fin, elongation of the bony rays is
achieved by the successful addition of newcalcified segments at the distal end. Otherinvestigators have documented this mode ofdevelopment in the dorsal fins of fish (Pot-thoff, ’74; Potthoff and Kelley, ’82). Thus, thefirst ray elements to be formed would beobserved in the most proximal areas of thefin.
AnnulusWe demonstrated alternating dark and
light purple bands following tissue exposureto hematoxylin and eosin. If we relate thesebands to the optical zonation pattern demon-strated in prepared dried fin tissue, a num-ber of dark bands correspond to a translu-cent zone and each light band to an opaquezone. Although oxytetracycline labelling ofchinook salmon fin-rays has not been per-formed, the number of translucent zonescorrespond to the age of fish as determinedby the time they were tagged as smolts(Beamish, unpublished data). Therefore, thegroups of dark bands we revealed by thisstaining procedure each represent an annu-lus.
The presence of collagen in the annuli wasverified using Masson’s trichrome stain.Since fin-ray tissue failed to react with Feul-gen’s stain and the periodic acid-Schiffmethod, we believe that nucleic acids andpolysaccharides, respectively, are not compo-nents of the annulus. In routine hematoxy-lin and eosin preparations, collagen is nor-mally acidophilic, i.e., eosinophilic, in nature.However, often after long-term exposure ofadult salmon fin-rays to Bouin’s fluid, hema-toxylin stained specific portions (annuli) ofthe fin-ray intensely as compared to theintervening areas. This staining pattern maybe due to the orientation of the fibers in theannulus and their closely packed arrange-ment. Possibly, ions of the dye molecule (he-matoxylin) become trapped between the fi-bers of this area and are not easily removedduring subsequent washing procedures. Thishematoxylin staining would result in thefailure of the eosin to stain the collagen.Unlike the core body of adults, the core bodyof juveniles, however, did not stain withhematoxylin and eosin. This differenceamong developmental stages may suggestthat the above postulate of fiber orientationaffecting staining properties and intensity isnot entirely the case. An alternative view isthat calcium has not been totally removedfrom the bone of the adult fin-ray. With hema-toxylin and eosin stains, most calcium salts
Fig. 10. A: Light micrograph from region IV of amedial pectoral hemisegment, demonstrating the dis-tinct banding pattern of fin-rays in the salmon, On-corhynchus tshawytscha. The core body (cb) follows theshape of the ray and is surrounded by a number ofclosely packed bands representing the first annulus (a1).Another group of bands is directly beneath the perios-teum (p) and represents the second annulus (a2). Finefracture lines extend from the core body to the perios-teum (arrowheads). MA, major arm; ma, minor arm. B:Region V of a lateral hemisegment from the dorsal fin.This tissue was etched with EDTA followed by NaOCl.SEM. Note the double-arched shape of the core body(cb). I, intersegmental region; arrowheads, lacunae ofosteocytes. C: Region IV of a medial hemisegment fromthe dorsal fin. SEM. This ray was etched with Baxterdecalcifying solution.The core body (cb) appears as abilaterally curved structure. p, periosteum; I, interseg-mental region; arrowheads, core body fibers. D: A trans-verse section through region IV of a medial hemiseg-ment in the pectoral fin. Pectoral rays examined fromthis region consistently displayed the greatest numberof bands throughout the fin. Twelve bands (arrowheads)were counted on the major arm of this 4-year-old salmon.Scale bars for A5 100 µm, for B,D 5 50 µm, and for C 525 µm.
SALMON FIN-RAY ANNULUS 317

are colored a deep blue-purple (Sheehan andHrapchak, ’80). Since X-ray microanalysisinfers that translucent areas possess rela-tively more calcium (Casselman, ’78), theseareas would stain more intensely with hema-toxylin than adjacent regions. Similar find-ings have also been documented in teeththat are decalcified and stained with hema-toxylin and eosin (Schour and Hoffman, ’49).This latter study assumed that hematoxylinstained well-calcified tissue more intenselyin decalcified preparations. This hypothesismight better explain the failure of the corebody of juveniles to stain. Animals were stillquite young and possibly there was less cal-cium per unit area than was found in adultfin-ray bone. Calcium ions, therefore, mayhave been easily removed and, thus, bone ofthe juvenile fin-rays would not stain asreadily with hematoxylin.
Staining and differentiation were also in-fluenced to a great extent by the fixation/decalcification method. Differential stain-ing, i.e., banding, did not occur with tissuesdecalcified with EDTA, nitric acid or formicacid, suggesting that the prolonged decalcifi-cation of fin-ray tissue in Bouin’s fixativemay have accounted for the banding patternobserved in the current study. Fixationcauses an increased affinity of proteins suchas collagen for stains because as peptidechains unfold, secondary groups becomeavailable for the reaction with dye molecules(Padykula, ’88). It is possible that the pro-longed exposure of fins of adult salmon toBouin’s fixative and more specifically picricacid may have altered the isoelectric point ofcollagen to such an extent that it reactedwith an affinity to the cationic ions of hema-toxylin. Although, juveniles were also placedin Bouin’s fixative for prolonged periods oftime, the period was not as protracted asthat of adult fins and this variable mayaccount for the discrepancy in staining ofbone between the two life cycle intervals.
The trends postulated above are con-firmed when they are related to the resultsobtained directly from the backscatter analy-sis of the fin-ray tissue. Resembling imagesof microradiographs, darker-grey level areasrepresent tissue that contains a lower min-eral content and lighter-grey level tissuethat contains a higher mineral content (Reidand Boyde, ’87; Grynpas and Holmyard, ’88;Boyce et al., ’90; Skedros et al., ’91). Thelighter-grey level areas are represented asnarrow bands located between much wider
darker-grey level areas. These areas corre-late well with the narrow translucent (an-nuli) and wide opaque zones of dried fin-raytissue and, therefore, these areas almostcertainly represent regions of high and lowlevels of calcium, respectively.
Calcium values determined by line-scananalysis across cross-sectional slices of codotoliths also indicate that the calcium con-tent is higher in the translucent than in theopaque zone. Mugiya (’66) reported that dur-ing the period of opaque zone formation inflatfish, the diffusible calcium in otolith fluiddecreases to 65.4% of the total calcium, butreaches a maximum of 79.1% during theperiod of translucent zone formation (citedby Casselman, ’78). In acid-etched otoliths oflaboratory fish, the translucent zones corre-spond with well-defined rings while theopaque zones are poorly etched (Taubert andCoble,’77). Similar findings were observedwhen EDTA was used for the etching(Gauldie and Nelson, ’88). These results cor-relate well with the observations of the pres-ent study as we noted that elevations offin-ray tissue are located in regions thatcorrespond to translucent areas of dried tis-sues.
CONCLUSIONS
The present light microscopic study pro-vides a detailed morphological description ofthe fin-ray of chinook salmon and indicatesthat transverse, stained sections of the fin-rays are useful indicators of age. In addition,the nature of the response to various decalci-fication agents and subsequent staining hasprovided some indication of a differentialorganization of extracellular matrix compo-nents between lamellae of the bone, namelyfibrous and inorganic components. We sus-pect that this varied organization accountsfor the marked appearance of annuli rela-tive to intervening lamellae. Fine structuralobservations are required to confirm thissupposition.
ACKNOWLEDGMENTS
This study was supported by a ScienceSubvention Grant from the Department ofFisheries and Oceans, Canada, and theNatural Science and Engineering ResearchCouncil of Canada (NSERC) and an NSERCoperating grant to J.H.Y. The authors appre-ciate the technical help provided by D. Holm-yard, R. Or, R. Roopsingh, G. Taylor, and S.Zaks. C. Neville and A. Thompson assistedin animal and tissue collection.
318 L.C.G. FERREIRA ET AL.

LITERATURE CITED
Arita GS. 1971. A re-examination of the functional mor-phology of the soft-rays in teleosts. Copeia 1971:691–697.
Baserga R, Malamud D. 1969. Modern methods in experi-mental pathology. Autoradiography. techniques andapplications. New York: Harper and Row, p 129–146.
Boyce TM, Bloebaum RD, Bachus KN, Skedros JG.1990. Calibration of the backscattered electron signalfor bone, biomaterial and implant research. SixteenthAnnual Meeting of the Society for Biomaterials. May20–23. Charleston, SC. p 188.
Brandstatter R, Misof B, Pazmandi C, Wagner GP. 1990.Micro-anatomy of the pectoral fin in blennies (Blen-niini, Blennioidea, Teleostei). J Fish Biol 37:729–743.
Cameron DA. 1972. The ultrastucture of bone. In: BourneGH, editor. The biochemistry and physiology of bone,Vol I. New York: Academic Press, p 191–121.
Casselman JM. 1974. Analysis of hard tissue of pike,Esox lucius L. with special reference to age and growth.In: Bagenal TB, editor. The ageing of fish: proceedingsof an international symposium. London: Unwin Broth-ers Ltd, p 13–27.
Casselman JM. 1978. Calcified tissue and body growthof northern pike, Esox lucius Linnaeus. PhD thesis,University of Toronto, Toronto Ontario. 782 p.
Casselman JM. 1983. Age and growth assessment fromcalcified structures: techniques and tools. In: Proceed-ings of the international workshop on age determina-tion of oceanic pelagic fishes: tunas, billfishes, andsharks. Miami, February 15–18, 1982. US Dept Com-merce, NOAA Tech Rep NMFS 8:1–17.
Casselman JM. 1987. Determination of age and growth.In: Weatherley AH, Gill HS, editors. The biology of fishgrowth. London: Academic Press, p 209–242.
Chilton DE, Beamish RJ. 1982.Age determination meth-ods for fishes studied by the groundfish program at thePacific Biological Station. Can Spec Pub Fish AquatSci 60:102 p.
Clayden EC. 1952. A discussion on the preparation ofbone sections by the paraffin wax method with speciealreference to the control of decalcification. J Med LabTechnol 10:103–123.
Dingerkus G, Uhler LD. 1977. Enzyme clearing of alcianblue stained whole small vertebrates for demonstra-tion of cartilage. Stain Tech 52:229–232.
Gauldie RW, Nelson DGA. 1988. Aragonite twinning andneuroprotein secretion are the cause of daily growthrings in fish otoliths. Comp Biochem Physiol 90:501–509.
Geerlink PJ. 1983. Pectoral fin kinematics of Cronsisformosa (Labridae, Teleostei). Neth J Zool 33:515–531.
Geerlink PJ. 1989. Pectoral fin morphology: a simplerelation to movement pattern? Neth J Zool 39:166–193.
Geerlink PJ, Videler JJ. 1987. The relation betweenstructure and bending properties of the teleost finrays. Neth J Zool 37:59–80.
Geraudie J, Landis WJ. 1982. The fine structure of thedeveloping pelvic fin skeleton in the trout, Salmogairdneri. Am J Anat 163:141–156.
Geraudie J, Landis WJ. 1989. Mineral deposition in thedeveloping pelvic fin bud of the trout, Salmo gaird-neri. Connect Tissue Res 22:224–230.
Grynpas MD, Holmyard D. 1988. Changes in quality ofbone mineral on aging and in disease. Scan Microsc2:1045–1054.
Hikita T. 1962. Ecological and morphological studies ofthe genus, Oncorhynchus (Salmonidae) with particu-
lar consideration on phylogeny. Sci Repts HokkaidoSalmon Hatchery 17:1–99.
Holland ND, Holland LZ. 1991. The histochemistry andfine structure of the nutritional reserves in the finrays of the lancet, Branchiostoma lanceolatum(Cephalocordata5Acrania). Acta Zool (Stockholm) 72:203–207.
Humason GL. 1967. Animal tissue techniques, 2nd ed.San Fransico: W.H. Freeman and Co.
International North Pacific Fisheries Commission 1959.Proceeding of the Annual Meeting 1958. 73 p.
Landis WJ, Geraudie J. 1991. Organization and develop-ment of the mineral phase during early ontogenesis ofthe bony fin rays of the trout, Oncorhynchus mykiss.Anat Rec 228:383–391.
Lanzing WJR. 1976. The fine structure of fins and fin-rays of Tilapia mossambica (Peters). Cell Tissue Res173:349–356.
Luft JH. 1961. Improvements in epoxy resin embeddingmethods. J Biophys Biochem Cytol 9:409–414.
MacLellan SE. 1987. Guide for sampling structures usedin age determination of pacific salmon. Nanaimo, BC:Dept. of Fisheries and Oceans, Fisheries ResearchBranch. Pacific Biological Station. p 1–27.
Moss ML. 1961a. Studies of the acellular bone of teleostfish. I. Morphological and systematic variations. ActaAnat 46:343–362.
Moss ML. 1961b. Osteogenesis of acellular teleost fishbone. Am J Anat 108: 99–110.
Moss ML. 1963. The biology of acellular teleost bone.Ann NY Acad Sci 109:337–350.
Moss ML, Posner AS. 1960. X-ray diffraction study ofacellular teleost bone. Nature 188:1037–1038.
Mugiya Y. 1966. Calcification in fish and shell-fish VI.Seasonal change in calcium and magnesium concentra-tions of the otolith fluid in some fish, with specialreference to the zone formation of their otolith. BullJpn Soc Sci Fish 32:549–557.
Nation JL. 1983. A new method using hexamethyldisila-zane for preparation of soft tissue insect tissues forscanning electron microscopy. Stain Technol 58:347–351.
Norris WP, Charin W, Lombard LS. 1963. Studies ofcalcification in the marine teleost. Ann NY Acad Sci109:312–336.
Padykula HA. 1988. Histochemistry and cytochemistry.In: Weiss L, editor. Cell and tissue biology. a textbookof histology. Baltimore: Urban and Schwartzenberg,Inc. p 93–113.
Parenti L. 1986. The phylogenetic significance of bonetypes in euteleost fishes. Zool J Linn Soc Lond 87:37–51.
Potthoff T. 1974. Osteological development and varia-tion in young tuna, genus Thunnus (Pisces, Scombri-dae) from the Atlantic Ocean. Fish Bull US 72:563–588.
Potthoff T, Kelley S. 1982. Development of the vertebralcolumn, fins and fin supports, branchiostegal ray andsquamation in the swordfish, Xiphias Gladius. FishBull 80:161–186.
Prince EC, Pulos LM, editors. 1983. Proceedings of theinternational workshop on age determination of oce-anic pelagic fishes: tunas, billfishes, and sharks. Mi-ami, February 15–18, 1982. US Dept. of Commerce,NOAA Tech Rep NMFS 8. 211 p.
Reid SA, Boyde A. 1987. Changes in mineral densitydistrubution in human bone with age: image analysisusing backscattered electrons in the scanning electronmicroscope. J Bone Min Res 2:13–22.
Robinson RA, Poly SB, Cooper RR. 1977. Electron micros-copy of mammlian bone. In: Zipkin I, editor. Biological
SALMON FIN-RAY ANNULUS 319

mineralization. New York: John Wiley and Sons. p257–298.
Romer AS, Parsons TS. 1986. The vertebrate body. Phila-delphia: WB Saunders.
Schour I, Hoffman MM. 1949. Studies in tooth develop-ment. J Dent Res 15:91–102.
Sheehan DC, Hrapchak BB. 1980. Theory and practiceof histotechnology. St. Louis: CV Mosby.
Skedros JG, Bloebaum RD, Boyce TM. 1991. Relation-ship of graylevels in backscattered electron images ofstimulated bone tissue to atomic number, density andmineral content. Sixteenth Annual Meeting of theSociety for Biomaterial. May 20–23, Charleson, SC. p 53.
Taubert BD, Coble DW. 1977. Daily rings in otoliths ofthree species of Lepomis and Tilapia mossambica. JFish Res Board Can 34:332–340.
Warshawsky H, Moore G. 1967. A technique for thefixation and decalcification of rat incisors for electronmicroscopy. J Histochem Cytochem 15:542–549.
Weiss RE. 1978. The comparative biology of osteogenesisand resorption of fish bone. PhD Dissertation, Univer-sity of South Carolina, Columbia, SC. 180 p.
York B. 1987. A guide to processing fin rays for ageingpurposes. Nanaimo, BC: Dept. of Fisheries and Oceans,Fisheries Research Branch. Pacific Biological Station.p 1–17.
320 L.C.G. FERREIRA ET AL.