
MacrophagesDiscriminateGlycosylationPatternsof...
9
Macrophages Discriminate Glycosylation Patterns of Apoptotic Cell-derived Microparticles * □ S Received for publication, June 17, 2011, and in revised form, November 2, 2011 Published, JBC Papers in Press, November 10, 2011, DOI 10.1074/jbc.M111.273144 Rostyslav O. Bilyy ‡§ , Tanya Shkandina ‡ , Andriy Tomin ‡ , Luis E. Muñoz § , Sandra Franz § , Volodymyr Antonyuk ‡ , Yuriy Ya. Kit ‡ , Matthias Zirngibl § , Barbara G. Fürnrohr § , Christina Janko § , Kirsten Lauber ¶ , Martin Schiller , Georg Schett § , Rostyslav S. Stoika ‡ , and Martin Herrmann §1 From the ‡ Institute of Cell Biology, National Academy of Sciences of Ukraine, 79005-Lviv, Ukraine, the § Department of Internal Medicine-3, Institute for Clinical Immunology and Rheumatology, University of Erlangen-Nuremberg, 91054 Erlangen, Germany, the ¶ Department of Radiation Oncology, University of Munich, 81377 Munich, Germany, and the Department of Medicine-V, University of Heidelberg, 69120 Heidelberg, Germany Background: Apoptotic cells release vesicles, which expose “eat-me” signals. Results: Vesicles originated from endoplasmic reticulum expose immature glycoepitopes and are preferentially phagocytosed by macrophages. Conclusion: Immature surface glycoepitopes serve as “eat-me” signals for the clearance of apoptotic vesicles originated from endoplasmic reticulum. Significance: Understanding the distinction by macrophages of apoptotic blebs may provide new insights into clearance-related diseases. Inappropriate clearance of apoptotic remnants is considered to be the primary cause of systemic autoimmune diseases, like systemic lupus erythematosus. Here we demonstrate that apo- ptotic cells release distinct types of subcellular membranous particles (scMP) derived from the endoplasmic reticulum (ER) or the plasma membrane. Both types of scMP exhibit desialy- lated glycotopes resulting from surface exposure of immature ER-derived glycoproteins or from surface-borne sialidase activ- ity, respectively. Sialidase activity is activated by caspase-depen- dent mechanisms during apoptosis. Cleavage of sialidase Neu1 by caspase 3 was shown to be directly involved in apoptosis- related increase of surface sialidase activity. ER-derived blebs possess immature mannosidic glycoepitopes and are prioritized by macrophages during clearance. Plasma membrane-derived blebs contain nuclear chromatin (DNA and histones) but not components of the nuclear envelope. Existence of two immuno- logically distinct types of apoptotic blebs may provide new insights into clearance-related diseases. During apoptotic death the surfaces of apoptotic remnants, including apoptotic blebs, are modified due to oxidative pro- cesses resulting in the appearance of immunologically “novel” autoantigens (1). An inefficient clearance of apoptotic cell material (shrunken apoptotic cells, apoptotic bodies or subcel- lular membranous particles (scMP) 2 ) will result in the accumu- lation of secondary necrotic remnants that release modified and potentially pro-inflammatory contents into the extracellu- lar milieu. It is common belief that inappropriate clearance of apoptotic cells can lead to the accumulation of modified autoantigens in tissues that foster autoimmune diseases (2). Unlike necrosis reportedly eliciting inflammatory responses (3), apoptosis mainly displays anti-inflammatory and immuno- suppressive properties (4). Apoptotic cells consecutively expose “eat-me” signals like phosphatidylserine (5) and altered glycoepitopes (6), important for early and late clearance, respectively. Artificially desialylated lymphocytes mimic a late apoptotic glycocalyx and are effectively cleared by human macrophages (7). The mechanism(s) modifying the glycocalyx are still elusive. We propose that (i) apoptozing cells may change the glycosyl- ation of de novo synthesized glycoconjugates. (ii) internal mem- branes containing immature glycoproteins (GP) may become exposed and (iii) mature GP may get modified by glycosidases (e.g. sialidases) in situ. To avoid confusion we introduced the term scMP for blebs that obviously had been released into the supernatant from the surfaces of apoptotic cells. The membranous structure formed by late apoptotic cells will be referred to as the big membranous bubble (bMB) (supplemental Fig. S1). Here we demonstrate that apoptotic cells release distinct types of scMP derived from endoplasmic reticulum (ER) or plasma membrane. Both exhibit desialylated glycotopes result- ing from surface exposure of immature ER-derived GP or from surface-borne sialidase activity, respectively. We addressed molecular mechanisms of GP redistribution during apoptosis * This work was supported by German-Ukrainian Grant UKR08/035 (to R. B. and M. H.), Grants from NASU, WUBMRC, and the President of Ukraine (to R. B.), by an intramural grant ELAN M3– 09.03.18.1 (to L. E. M.), by the DFG (GRK-SFB643), and the K&R Wucherpfennigstiftung (to M. H.). Confocal microscopy was done at the Nikon Imaging Center at the University of Heidelberg. □ S This article contains supplemental Figs. S1–S6. 1 To whom correspondence should be addressed. Department of Internal Medicine 3, University of Erlangen-Nuremberg, Krankenhausstrasse 12, D-91054 Erlangen, Germany. Tel.: 49-9131-85-36990; Fax: 49-9131-85- 35776; E-mail: [email protected]. 2 The abbreviations used are: scMP, subcellular membranous particle; ER, endoplasmic reticulum; GP, glycoprotein; PMN, polymorpho-nuclear leu- kocytes; PM, plasma membrane; RBC, red blood cells; bMB, big membra- nous blebs; PBMC, peripheral blood mononuclear cells. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 1, pp. 496 –503, January 2, 2012 © 2012 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. 496 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 1 • JANUARY 2, 2012 by guest on June 5, 2018 http://www.jbc.org/ Downloaded from
Transcript of MacrophagesDiscriminateGlycosylationPatternsof...

Macrophages Discriminate Glycosylation Patterns ofApoptotic Cell-derived Microparticles*□S
Received for publication, June 17, 2011, and in revised form, November 2, 2011 Published, JBC Papers in Press, November 10, 2011, DOI 10.1074/jbc.M111.273144
Rostyslav O. Bilyy‡§, Tanya Shkandina‡, Andriy Tomin‡, Luis E. Muñoz§, Sandra Franz§, Volodymyr Antonyuk‡,Yuriy Ya. Kit‡, Matthias Zirngibl§, Barbara G. Fürnrohr§, Christina Janko§, Kirsten Lauber¶, Martin Schiller�,Georg Schett§, Rostyslav S. Stoika‡, and Martin Herrmann§1
From the ‡Institute of Cell Biology, National Academy of Sciences of Ukraine, 79005-Lviv, Ukraine, the §Department of InternalMedicine-3, Institute for Clinical Immunology and Rheumatology, University of Erlangen-Nuremberg, 91054 Erlangen, Germany,the ¶Department of Radiation Oncology, University of Munich, 81377 Munich, Germany, and the �Department of Medicine-V,University of Heidelberg, 69120 Heidelberg, Germany
Background: Apoptotic cells release vesicles, which expose “eat-me” signals.Results: Vesicles originated from endoplasmic reticulum expose immature glycoepitopes and are preferentially phagocytosedby macrophages.Conclusion: Immature surface glycoepitopes serve as “eat-me” signals for the clearance of apoptotic vesicles originated fromendoplasmic reticulum.Significance:Understanding the distinction bymacrophages of apoptotic blebsmay provide new insights into clearance-relateddiseases.
Inappropriate clearance of apoptotic remnants is consideredto be the primary cause of systemic autoimmune diseases, likesystemic lupus erythematosus. Here we demonstrate that apo-ptotic cells release distinct types of subcellular membranousparticles (scMP) derived from the endoplasmic reticulum (ER)or the plasma membrane. Both types of scMP exhibit desialy-lated glycotopes resulting from surface exposure of immatureER-derived glycoproteins or from surface-borne sialidase activ-ity, respectively. Sialidase activity is activated by caspase-depen-dent mechanisms during apoptosis. Cleavage of sialidase Neu1by caspase 3 was shown to be directly involved in apoptosis-related increase of surface sialidase activity. ER-derived blebspossess immaturemannosidic glycoepitopes and are prioritizedby macrophages during clearance. Plasma membrane-derivedblebs contain nuclear chromatin (DNA and histones) but notcomponents of the nuclear envelope. Existence of two immuno-logically distinct types of apoptotic blebs may provide newinsights into clearance-related diseases.
During apoptotic death the surfaces of apoptotic remnants,including apoptotic blebs, are modified due to oxidative pro-cesses resulting in the appearance of immunologically “novel”autoantigens (1). An inefficient clearance of apoptotic cellmaterial (shrunken apoptotic cells, apoptotic bodies or subcel-
lular membranous particles (scMP)2) will result in the accumu-lation of secondary necrotic remnants that release modifiedand potentially pro-inflammatory contents into the extracellu-lar milieu. It is common belief that inappropriate clearance ofapoptotic cells can lead to the accumulation of modifiedautoantigens in tissues that foster autoimmune diseases (2).Unlike necrosis reportedly eliciting inflammatory responses
(3), apoptosis mainly displays anti-inflammatory and immuno-suppressive properties (4). Apoptotic cells consecutivelyexpose “eat-me” signals like phosphatidylserine (5) and alteredglycoepitopes (6), important for early and late clearance,respectively. Artificially desialylated lymphocytes mimic a lateapoptotic glycocalyx and are effectively cleared by humanmacrophages (7).The mechanism(s) modifying the glycocalyx are still elusive.
We propose that (i) apoptozing cells may change the glycosyl-ation of de novo synthesized glycoconjugates. (ii) internalmem-branes containing immature glycoproteins (GP) may becomeexposed and (iii) mature GP may get modified by glycosidases(e.g. sialidases) in situ.To avoid confusion we introduced the term scMP for blebs
that obviously had been released into the supernatant from thesurfaces of apoptotic cells. The membranous structure formedby late apoptotic cells will be referred to as the bigmembranousbubble (bMB) (supplemental Fig. S1).Here we demonstrate that apoptotic cells release distinct
types of scMP derived from endoplasmic reticulum (ER) orplasma membrane. Both exhibit desialylated glycotopes result-ing from surface exposure of immature ER-derived GP or fromsurface-borne sialidase activity, respectively. We addressedmolecular mechanisms of GP redistribution during apoptosis
* This work was supported by German-Ukrainian Grant UKR08/035 (to R. B.and M. H.), Grants from NASU, WUBMRC, and the President of Ukraine (toR. B.), by an intramural grant ELAN M3– 09.03.18.1 (to L. E. M.), by the DFG(GRK-SFB643), and the K&R Wucherpfennigstiftung (to M. H.). Confocalmicroscopy was done at the Nikon Imaging Center at the University ofHeidelberg.
□S This article contains supplemental Figs. S1–S6.1 To whom correspondence should be addressed. Department of Internal
Medicine 3, University of Erlangen-Nuremberg, Krankenhausstrasse 12,D-91054 Erlangen, Germany. Tel.: 49-9131-85-36990; Fax: 49-9131-85-35776; E-mail: [email protected].
2 The abbreviations used are: scMP, subcellular membranous particle; ER,endoplasmic reticulum; GP, glycoprotein; PMN, polymorpho-nuclear leu-kocytes; PM, plasma membrane; RBC, red blood cells; bMB, big membra-nous blebs; PBMC, peripheral blood mononuclear cells.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 1, pp. 496 –503, January 2, 2012© 2012 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
496 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 1 • JANUARY 2, 2012
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from
and observed that sialidase activity is activated by caspase-de-pendent mechanisms. Plasma membrane-derived bMB andscMP contain chromatin but not the nuclear envelope andER-derived scMP are prioritized by macrophages during clear-ance. The understanding of the immunological distinction bymacrophages of ER and plasma membrane derived apoptoticblebs may provide new insights into clearance-related diseases.
EXPERIMENTAL PROCEDURES
Cell Culture and Isolation of Cells—We employed humanleukemia Jurkat T-cells, human HeLa cells, and CD95 positivehuman breast cancer cells MCF-7, as well as primary humanpolymorphonuclear leukocytes (PMN) and monocyte-derivedmacrophages from healthy volunteers for our experiments.Peripheral blood mono-nuclear cells (PBMC) were isolated fromperipheral anticoagulated blood by LymphoPrep� gradient cen-trifugation according to the manufacturer’s recommendations.Plasticadherent PBMC were then cultured for 7 days in the pres-ence of GM-CSF (100 units/ml) and autologous serum (days 1, 3,and5) togeneratemonocyte-derivedmacrophages.After 7daysofdifferentiation, these phagocytes usually contain more than 85%phagocytes positive for CD11b, CD14, and CD89.Induction and Inhibition of Apoptosis—Cell viability was
controlled by AnnexinV/PI staining. Apoptosis was induced byetoposide (1�M, 24 h; Jurkat cells), by irradiationwith ultravioletlight typeB (UV-B; 180mJ/cm2within60s; Jurkat andHeLa), orbyaging (PMN). Caspase inhibitors zVAD-FMK (zVAD, carboben-zoxy-valyl-alanyl-aspartyl-[O-methyl]-fluoro-methyl-ketone,Bachem AG, Bubendorf), zDEVD-FMK, zIETD-FMK andzLEHD-FMK (R&DSystems,Minneapolis) were applied as rec-ommended by the manufacturers.Cytochemistry—Cytochemistry was performed with stand-
ard laboratory methods (8). Analyses employing fluorescence-labeled lectins (9) were performed using an EPICS XLTM(Coulter, Hialeah) or FACS scan flow cytometer (BD Biosci-ences). Tunicamycin (1 �M), monensin (1 �M), 2-deoxy-D-glu-
cose (100 mM), cycloheximide (15 �g/ml), and actinomycin D(5 �g/ml) (all from Sigma) were applied according to the man-ufacturer’s recommendations.Transfection of Cells—Transfection of HeLa cells with KDEL
receptor-GFP constructwas carried out as described previously(10). Transduction with GFP-linked N-acetylgalactosaminyl-transferase-2 and SV40 nuclear translocation signal were per-formed using Organelle LightsTM (Invitrogen) Golgi andnuclear envelope GFP kits, respectively.Staining with Lectins—LectinsNPL (Narcissus pseudonarcis-
sus lectin), PNA (peanut agglutinin), RCA-I (Ricinus communisagglutinin), PSL (Pisum sativum lectin), were from LectinotestLaboratory (Ukraine) and VAA (Viscum album agglutinin 1),CEL (Canavalia ensiformis lectin), MAA-II (Maackia amuren-sis agglutinin II) (a2,3-sialil specific), UEA (Ulex EuropeausAgglutinin I) (specific to terminal fucose) were from VectorLaboratories. For the imaging of bubbling apoptotic cells, 10�g/ml lectins were added to cultured cells and immediatelyimaged to achieve optimal S/N ratios (11), non-permebializedcells were used to demonstrate surface-related signals.Measurement of Sialidase Activity—Sialidase activity was
visualized using 5 �M final concentration of the enzymatic sub-strate 2�-(4-methylumbelliferyl)-�-D-N-acetylneuraminic acid(4-MUNA) providing a fluorescent product upon cleavage (12,13). PM was stained with Vybrant DiI (Invitrogen), ER wasstained with ER-TrackerTM Green (BODIPY� FL glibencl-amide) (Invitrogen) prior to induction of apoptosis.Microscopy—Confocal microscopy was performed using
PerkinElmer Ultra-View spinning disc confocal on NikonTE2000-E Ti inverted microscope using �100 oil-immersionlens (excitation at 488, 568, 647 nm, detection at 650 nm, shownin red and 488 nm, shown in green). Fluorescent microscopywas performed using Zeiss AxioImager A1with Zeiss AxioCamMRm digital camera. Some images were deconvolved usingAutoQuant X2 (MediaCybernetics Inc). ImageJ (NIH) software
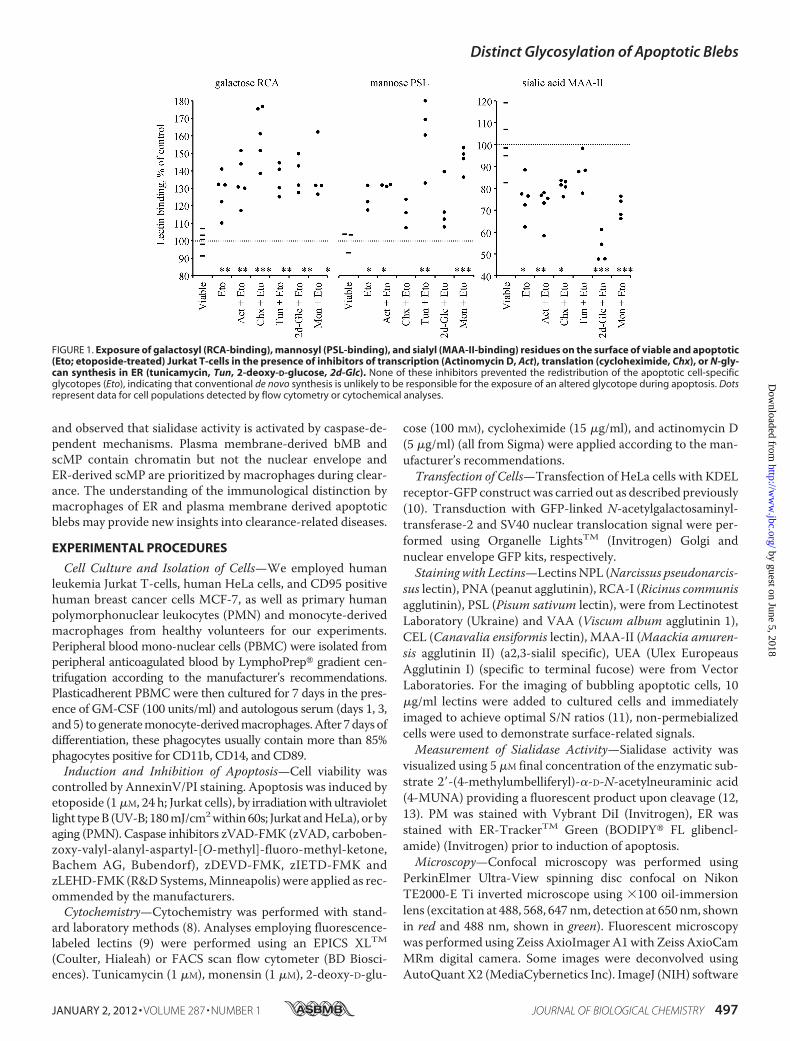
FIGURE 1. Exposure of galactosyl (RCA-binding), mannosyl (PSL-binding), and sialyl (MAA-II-binding) residues on the surface of viable and apoptotic(Eto; etoposide-treated) Jurkat T-cells in the presence of inhibitors of transcription (Actinomycin D, Act), translation (cycloheximide, Chx), or N-gly-can synthesis in ER (tunicamycin, Tun, 2-deoxy-D-glucose, 2d-Glc). None of these inhibitors prevented the redistribution of the apoptotic cell-specificglycotopes (Eto), indicating that conventional de novo synthesis is unlikely to be responsible for the exposure of an altered glycotope during apoptosis. Dotsrepresent data for cell populations detected by flow cytometry or cytochemical analyses.
Distinct Glycosylation of Apoptotic Blebs
JANUARY 2, 2012 • VOLUME 287 • NUMBER 1 JOURNAL OF BIOLOGICAL CHEMISTRY 497
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

was used for co-localization analysis and image processing. In-focus rendering of DIC images was donewithHelicon Focus. Inall cases, microscopy was done observing at least 5 Petri dishes(9 cm) with cells grown to 70–90% confluency for each variant.For quantification at least 1000 cells were counted. Experi-ments were repeated at least 3 times.Determination of Trans-sialidase and Sialidase Activity—
Cell trans-sialidase activity toward RBCs was determined usinga test based on RBC agglutination with PNA lectin describedelsewhere (14) and by their co-incubation at 37 °C for 3 h withcells/conditioned medium or Clostridium perfringens (Sigma)sialidase as controls and standard, respectively.Caspase inhibitors zVAD-FMK (zVAD, carbobenzoxy-valyl-
alanyl-aspartyl-[O-methyl]-fluoro-methyl-ketone, Bachem
AG), zDEVD-FMK, zIETD-FMK and zLEHD-FMK (R&D Sys-tems) were applied according to the manufacturers’ recom-mendations. Monoclonal anti-Neu2 and polyclonal anti-Neu1antibodies were from Abnova, monoclonal anti-Neu2, poly-clonal anti-Neu1 and anti-Neu3were fromSantaCruz Biotech-nology, and polyclonal anti-Neu4 antibody was from Protein-tech Group. Isoelectric focusing was performed according toRef. 15.Phagocytosis Assays—Phagocytosis of scMP was assessed by
incubation of PMN-derived scMP with human monocyte-de-rived macrophages and uningested scMP stained with lectin(s)were analyzed by flow cytometry.In silico analysis for transmembrane regions was done with
HMMTOP and TMHMM. For the prediction of cleavage sites
FIGURE 2. Apoptotic cells produce distinct types of scMP. a, apoptotic cells form scMP that are derived from the PM (red) or from the ER (green). 120� afterirradiation with UV-B, only PM-derived blebs are generated. Later, ER-derived membranes can be observed on the surface of the cell (arrow) and on scMP(arrowhead), respectively. Finally a big PM-derived bubble (bMB) is formed from PM. b, HeLa cells were stained with ER-tracker™, and apoptosis was induced;after 3 h apoptotic cells were counterstained for immature ER-related glycotopes with CEL. scMP, positive for both ER-tracker and CEL, are indicated by arrows.During apoptosis, ER-related glycoconjugates are exposed on the cell surfaces. The intensely stained cell is a secondary necrotic one, exposing internalglycoepitopes. c, apoptotic cells (like B) were incubated in the medium containing the sialidase substrate 4-MUNA. A scMP positive for ER and negative forsialidase activity is indicated by arrows; a scMP negative for ER and positive for sialidase activity is indicated by arrowheads. During apoptosis sialidase activityis present on the PM and on PM-related scMP, but not on ER-related scMP. d, bMB arises from the PM (DiI staining, red, left panel) and is filled with chromatin-derived antigens (here: histone 2B (middle panel)) in most of the transfected cells. The nuclear envelope, visualized by GFP-linked nesprin-1�, is excluded formthe bMB (five preparations utilizing 1000 cells each; right panel). a-d, non-permeabilized non-fixed cells were imaged.
Distinct Glycosylation of Apoptotic Blebs
498 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 1 • JANUARY 2, 2012
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

by caspases of sialidases, we employed GrabCas, CASVM, Pep-tideCutter, and CasCleave and set the cut-off scores �5,0(GrabCas).Statistics—Statistical significance was assessed by Student’s t
test. Three levels of significance were used: *, p � 0.05; **, p �0.01; ***, p � 0.001.
RESULTS
The Exposure of Galactose/Mannose on the Surfaces of Apo-ptotic Cells Is Independent of Proteinneogenesis—Toanalyze themechanisms modifying the glycocalyx we blocked the synthesispathway for theN-glycans of GP and gangliosides at several steps.Inhibitors of transcription [actinomycinD], translation [cyclohex-imide],N-glycan synthesis in theER [tunicamycin, 2-deoxy-D-glu-cose], and export fromGolgi to plasmamembrane (PM), [monen-sin] did not prevent the increased exposure of galactose/mannoseand decreased exposure of sialic acid (Fig. 1).ER-derived Membranes Are Exposed in Late Apoptosis—
Next, we transfectedHeLa cells with an ER-residing KDEL recep-tor-GFP fusion construct and stained the PMwith the fluorescenthydrophobic dye DiI. When we induced apoptosis with UVB,blebbing of the PM started 120 min after irradiation. After 240min, the PM constricted and formed a polar conglomerate stillconnected to the cells’ bodies. Concomitantly, ERmembranes gotexposed and started blebbing. In the supernatant of these cells,distinct scMPwere detected that had originated fromPM (red) orfrom ER (green), respectively (Fig. 2a). After 360 min, a bMB wasformed that originated from the former PM (red). All blebbingcells produced PM-derived scMP, and most (�90,0%) of the lateapoptotic cells demonstrated ER-derived scMP.However, analyz-ing about 1000 cells from seven independent preparations, wenever observed double positive scMP in all our microscopic stud-ies. ER-trackerpositiveblebsboundNPL (Narcissuspseudonarcis-sus lectin) andCEL (Canavalia ensiformis lectin), which preferen-tially recognize ER-related high mannose N-glycans andER-specific terminally glucosylated N-glycan intermediates (16),respectively (Fig. 2b). Both NPL and CEL signal were co-localizedwith ER-tracker signal, as shown for permeabilized cells in supple-mental Fig. S2.
Sialidase Activity and ER-Markers Are Mutually Exclusive—Wescreened the cells for sialidase activity and observed a reducedsurface sialylationandan increasedexposureof galactosyl residueson the surfaces of apoptotic cells and their surface-derived scMP.The non-penetrating sialidase substrate 4-MUNA detected siali-dase activity on the surfaces of apoptotic cells and of some of thescMP. Sialidase activity and ER-tracker staining of the scMPweremutually exclusive (Fig. 2c). Chromatin-derived histone 2B-GFPfusionproteinswere translocated to the cytoplasmduring apopto-sis and was finally located inside the bMB (Fig. 2d) originatingfrom the former PM (observed in most of the apoptotic cell rem-nants). Moreover, during late apoptosis nuclear components canmainly be included in apoptotic scMP, (observed in some few ofthe blebbing cells (supplemental Fig. S3)). After induction withetoposide of apoptosis the trans-sialidase activity of Jurkat cellstoward redblood cell surface,GPwas increasedmore than20-fold(Fig. 3).The Increase of Sialidase Activity in Apoptotic Cells Is
Caspase-dependent—Treatment with caspases 3 or 8 of celllysates from viable cells increased the sialidase activities to a levelsimilar to cell lysates generated from apoptotic cells (Fig. 4a). Incontrast, the caspase inhibitor zVAD inhibited the generation ofsialidase activity in Jurkat cells treated with etoposide or in agedPMN (Fig. 4b). Caspase 3-deficient MCF-7 cells displayed noincreased surface sialidase activity during apoptosis (induced byUV-B, etoposideoranti-CD95Ab;Fig. 4c).TransfectionofMCF-7cells with caspase 3 restored the increase during apoptosis of siali-dase activity (Fig. 4d). MCF-7 cells, lacking caspase-3 andincreased sialidase activity during apoptosis produced ER-derivedscMP upon treatment with UV-B, They also produce two distinctpopulations of scMP that were positive for ER-tracker or Dil,respectively (Fig. 4e and supplemental Fig. S4).In silico analysis revealed transmembrane regions and a pre-
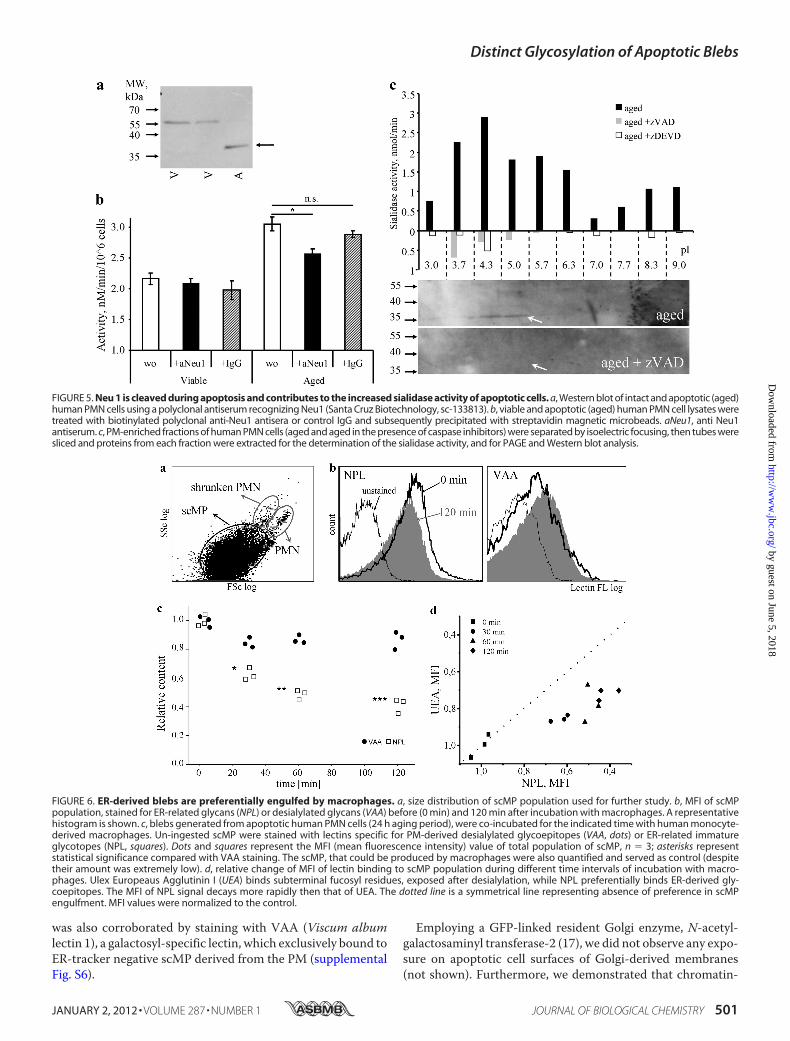
dicted caspase 3 cleavage sites for both Neu1/4 and Neu1 (Uni-Prot: Q5JQI0, cleavage at Asp-135with score 12,0 byGrabCas),respectively. As Neu2 is exclusively expressed in muscle andNeu3/4 has no high-score cleavage site for executer caspaseswefocused on Neu1 as the best fitting candidate among the fourknown human sialidases. Western blot analysis of the lysatesfrom viable or apoptotic PMN showed a band most likely rep-resenting a cleavage product of the Neu1 protein in the latter(Fig. 5a). A similar fragment was to be observed after in vitrotreatment of cell lysates with caspase 3 (not shown). The deple-tion of Neu1 abrogated the increased sialidase activity of apo-ptotic human PMN (Fig. 5b).Isoelectric focusing of PM-enriched fractions revealed a
caspase-dependent sialidase activity in the acidic fractions. Thepeak of the sialidase activity contained a polypeptide with themolecular mass (�35 kDa) compatible with the C-terminalfragment of Neu1 (136–415; predicted pI-5.61), which wasreactive with a polyclonal antiserum against Neu1 (Fig. 5c).Macrophages Prefer ER-derived scMP—To analysis of phag-
ocytosis of the distinct types of scMP we employed humanmonocyte-derived macrophages as a validated model for clear-ance studies (7). scMP derived from aged human PMN servedas “prey.” We detected the staining (judged by MFI) of“uncleared” scMP, since it is less reliable to evaluate the amountof ingested scMP, which instantly become degraded inside the
FIGURE 3. Sialidase activity of apoptotic cells. Apoptotic cells possessincreased sialidase activity trans-acting toward RBC. Viable and apoptotic Jurkatcells or their conditioned medium were co-incubated with RBC for 3 h at 37 °C.RBC were then treated with the lectin PNA (specific for desialylated gly-coepitopes) and agglutination was evaluated. Solid and open dots representactivity of intact and apoptotic cells, respectively. Neuraminidase from C. perfrin-gens served as standard. Presented data are mean values of three independentexperiments.
Distinct Glycosylation of Apoptotic Blebs
JANUARY 2, 2012 • VOLUME 287 • NUMBER 1 JOURNAL OF BIOLOGICAL CHEMISTRY 499
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

phagocytes. The scMP, that are produced bymacrophageswerealso quantified and served as control (despite their amount wasextremely low). scMPgated by FSc/SSc data as shownonFig. 6awere analyzed for fluorescence intensity after staining with lec-tins. When we co-incubated apoptotic ER- and PM-derivedscMP from aged human PMN with human macrophages, ER-derived scMP, endowed with immature ER-related oligoman-nosidic glycoepitopes, detected with the lectin NPL, were sig-nificantly faster cleared by macrophages than the PM-derivedones, exposing PM-related desialylated glycolepitopes charac-terized by terminal galactose or subterminal fucose residuesdetected with the lectins VAA (Fig. 6c) or UEA, respectively(Fig. 6d). Thiswas not due to the size difference asmacrophages
do not display anymarked size preference for the engulfment ofscMP (supplemental Fig. S5).
DISCUSSION
Blockage of the synthesis pathway for N-glycans of GP andgangliosides suggests that de novo synthesis is unlikely to beresponsible for the apoptosis-related surface-neoglycotopes.Fluorescence microscopy revealed the formation of distinctapoptotic scMP that originated from the PM or the ER andpossess characteristic patterns of glycosylation. PM- and ER-derived scMP expose desialylated and immature mannose-richglycotopes, respectively. We demonstrated that sialidase activ-ity is focused on the PM and on PM-derived scMP. This finding
FIGURE 4. Caspases are involved in sialidase activation during apoptosis. a, treatment of viable (closed dots) and apoptotic (open dots) Jurkat cell lysateswith caspase 3 and 8 resulted in apoptosis-like induction of sialidase activity. Simultaneous treatment with caspase inhibitors demonstrates specificity. Dottedline represents the mean value of viable cell lysates. V, viable; A, apoptotic; zV, zVAD; zD, zDEVD; zI, zIETD; C3 & C8, caspase 3 & 8, respectively. b, sialidase activityon the surfaces of human PMN after aging in the presence of the pan-caspase inhibitor zVAD. c, sialidase activity of cells either expressing (Jurkat) or lacking(MCF-7) caspase 3 at different time points after treatment with inducers of apoptosis such as UV-B irradiation (UV, 180mJ/cm2, 60s), anti-CD95 antibodies(CD95, 1 �g/ml), and etoposide (Eto, 1 �M). Caspase 3 is required for the increased sialidase activity during the execution of apoptosis. Receptor-dependent andinternal pathways are affected. Dotted lines represent the mean value of viable cells. d, transfection of MCF-7 cells with caspase 3 restored sialidase activationduring apoptosis induced by UV-B irradiation, but not during simultaneous treatment with UV-B irradiation and zVAD. The dotted line represents the meansialidase activity of the viable cell preparation. e, despite lacking detectable sialidase activity upon apoptosis, MCF-7 cells (not expressing caspase 3) produceER-derived scMP detected by staining with NPL in more than 30% of the blebbing cells. Apoptosis was induced by UV-B irradiation, and images were taken 4 hafter induction.
Distinct Glycosylation of Apoptotic Blebs
500 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 1 • JANUARY 2, 2012
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from
was also corroborated by staining with VAA (Viscum albumlectin 1), a galactosyl-specific lectin, which exclusively bound toER-tracker negative scMP derived from the PM (supplementalFig. S6).
Employing a GFP-linked resident Golgi enzyme, N-acetyl-galactosaminyl transferase-2 (17), we did not observe any expo-sure on apoptotic cell surfaces of Golgi-derived membranes(not shown). Furthermore, we demonstrated that chromatin-
FIGURE 5. Neu 1 is cleaved during apoptosis and contributes to the increased sialidase activity of apoptotic cells. a, Western blot of intact and apoptotic (aged)human PMN cells using a polyclonal antiserum recognizing Neu1 (Santa Cruz Biotechnology, sc-133813). b, viable and apoptotic (aged) human PMN cell lysates weretreated with biotinylated polyclonal anti-Neu1 antisera or control IgG and subsequently precipitated with streptavidin magnetic microbeads. aNeu1, anti Neu1antiserum. c, PM-enriched fractions of human PMN cells (aged and aged in the presence of caspase inhibitors) were separated by isoelectric focusing, then tubes weresliced and proteins from each fraction were extracted for the determination of the sialidase activity, and for PAGE and Western blot analysis.
FIGURE 6. ER-derived blebs are preferentially engulfed by macrophages. a, size distribution of scMP population used for further study. b, MFI of scMPpopulation, stained for ER-related glycans (NPL) or desialylated glycans (VAA) before (0 min) and 120 min after incubation with macrophages. A representativehistogram is shown. c, blebs generated from apoptotic human PMN cells (24 h aging period), were co-incubated for the indicated time with human monocyte-derived macrophages. Un-ingested scMP were stained with lectins specific for PM-derived desialylated glycoepitopes (VAA, dots) or ER-related immatureglycotopes (NPL, squares). Dots and squares represent the MFI (mean fluorescence intensity) value of total population of scMP, n � 3; asterisks representstatistical significance compared with VAA staining. The scMP, that could be produced by macrophages were also quantified and served as control (despitetheir amount was extremely low). d, relative change of MFI of lectin binding to scMP population during different time intervals of incubation with macro-phages. Ulex Europeaus Agglutinin I (UEA) binds subterminal fucosyl residues, exposed after desialylation, while NPL preferentially binds ER-derived gly-coepitopes. The MFI of NPL signal decays more rapidly then that of UEA. The dotted line is a symmetrical line representing absence of preference in scMPengulfment. MFI values were normalized to the control.
Distinct Glycosylation of Apoptotic Blebs
JANUARY 2, 2012 • VOLUME 287 • NUMBER 1 JOURNAL OF BIOLOGICAL CHEMISTRY 501
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

derived histone 2B-GFP fusion proteins are translocated to thecytoplasm during apoptosis and end up in a bMB (Fig. 2d) thatoriginated from the PM. During late stages of apoptosis mostchromatin is translocated into a bMB (supplemental Fig. S3).However, we did not observe scMP containing nesprin-1� themarker protein of the nuclear envelop.Analyzing the following organells: endoplasmic reticulum,
endosomes, Golgi, lysosomes, mitochondria, nuclear envelope,nucleus, and peroxisomes (all visualized via transfection withcorresponding fluorescent marker proteins) we observed thatonly endoplasmic reticulum, endosomes, and nuclei areredistributed into the scMP. Additionally, we have observedsome organelles (discriminated by DIC microscopy), insidethe bMB.Induction of apoptosis considerably increased a surface-as-
sociated sialidase activity, which was not detected in the cells’supernatants. This suggests the action of a cell surface-linkedsialidase like Neu-3 (18) or the reportedly surface associatedNeu-1 (19). Antisera directed against peptides of Neu-1 andNeu-3 diminished surface-associated sialidase activity (notshown).Immunoprecipitation studies suggest thatNeu-1may at least
partially be responsible for the increased sialidase surface activ-ity observed during apoptosis. As shown byWestern blot anal-yses, Neu-1 is cleaved by caspase 3. However, we do not yet
know how (directly or indirectly) this cleavage leads to the acti-vation of Neu-1.The ability of MCF-7 cells lacking caspase-3 (and thus lack-
ing increased sialidase activity during apoptosis) to produceboth ER- and PM-derived scMP, suggests distinct molecularmechanisms involved in the formation of ER-derived scMP andin desialylation of glycoepitopes of PM-derived scMP. The for-mer processwas previously reported to be dependent onROCK(10).A plethora of receptors and adaptor proteins are involved in
the uptake by macrophages of apoptotic cells and their subcel-lular particles (reviewed in Ref. 20). In general they either rec-ognize phosphatidylserine or altered carbohydrates. Justrecently the RAGE receptor has been added to the list of phos-phatidylserine-dependent receptors enhancing the efferocyto-sis of apoptotic cells (21).Both nucleotides (22) and lipids (23) were shown to act as
“eat-me” and “find-me” signals in promotion of apoptotic cellclearance. The surface exposure of the latter was shown to bedependent on caspase 3. Changes of the surface charge (24) aswell as carbohydrate content (25) are essential for the clearanceof apoptotic cells. Recently we have shown that artificial desia-lylation of viable cell surfaces creates an “eat-me” signal formacrophages (7).
FIGURE 7. The dual pathway model for altered N-glycan exposure during late apoptosis and formation of two distinct types of scMP (1). Conventionalde novo glycan synthesis is not active during apoptosis (2). ER membranes with immature glycans are exposed on the cell surface and in ER-derived scMP (3).Apoptosis is accompanied by a caspase-dependent activation of surface sialidase(s) leading to desialylation of pre-existing glycoepitopes on the cells surfacesand on PM-derived scMP.
Distinct Glycosylation of Apoptotic Blebs
502 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 1 • JANUARY 2, 2012
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

In phagocytosis assays, human macrophages cleared ER-de-rived scMP (possessing immature ER-related glycoepitopes)significantly faster than the PM-derived ones (possessing PM-related desialylated glycoepitopes) (Fig. 6). This may result in adifferential immunological processing of these subcellular par-ticles and their associated antigens, the size of scMP has had aminor effect on their clearance speed (supplemental Fig. S5).Intriguingly, nucleosomes, a prototypic autoantigen targeted
by sera of patients with systemic lupus erythematosus (SLE),were mainly concentrated in the bMB (Fig. 2D). This is of par-ticular importance since SLE patients have been reported tofrequently display a deficient clearance of apoptotic cells (2).Vaccinia and other viruses have been shown to mimic PM-
derived, phosphatidylserine (PS) exposing scMP, utilizing theability of surrounding cells for macropinocytosis of PS-expos-ing particles. This pathway augments the infectivity of theviruses, especially that for phagocytes (26, 27).In conclusion,we propose amechanism including alterations
of the glycocalyx during apoptosis and provide evidence thatreduced sialylation of cells undergoing apoptosis can be causedby both surface exposure of immature ER-derived GP and sur-face-bound sialidase activity cleaving mature GP in situ. Ourdata indicate that these processes aremutually executed by pre-viously undifferentiated types of apoptotic scMP (Fig. 7). ER-derived scMP are prioritized by macrophages during apoptoticcell clearance. Our data are in a good agreement with previousobservation where calreticulin exposed from ER during apo-ptosis has been found to dictate the immunogenicity of cancercell death (28).
Acknowledgments—We thank Dr. U. Engel andDr. C. Ackermann forexcellent technical support.
REFERENCES1. Casciola-Rosen, L. A., Anhalt, G., and Rosen, A. (1994) J. Exp. Med. 179,
1317–13302. Muñoz, L. E., Lauber, K., Schiller, M., Manfredi, A. A., and Herrmann, M.
(2010) Nat. Rev. Rheumatol. 6, 280–2893. Sancho, D., Joffre, O. P., Keller, A. M., Rogers, N. C., Martínez, D., Her-
nanz-Falcón, P., Rosewell, I., Reis, E., and Sousa, C. (2009) Nature 458,899–903
4. Voll, R. E., Herrmann, M., Roth, E. A., Stach, C., Kalden, J. R., and Girkon-
taite, I. (1997) Nature 390, 350–3515. Ravichandran, K. S. (2010) J. Exp. Med. 207, 1807–18176. Bilyy, R., and Stoika, R. (2007) Autoimmunity 40, 249–2537. Meesmann, H. M., Fehr, E. M., Kierschke, S., Herrmann, M., Bilyy, R.,
Heyder, P., Blank, N., Krienke, S., Lorenz, H. M., and Schiller, M. (2010)J. Cell Sci. 123, 3347–3356
8. Bilyy, R. O., and Stoika, R. S. (2003) Cytometry A 56, 89–959. Franz, S., Frey, B., Sheriff, A., Gaipl, U. S., Beer, A., Voll, R. E., Kalden, J. R.,
and Herrmann, M. (2006) Cytometry A 69, 230–23910. Franz, S., Herrmann, K., Führnrohr, B., Sheriff, A., Frey, B., Gaipl, U. S.,
Voll, R. E., Kalden, J. R., Jäck, H. M., and Herrmann, M. (2007) Cell Death.Differ. 14, 733–742
11. Kuno, A., Uchiyama, N., Koseki-Kuno, S., Ebe, Y., Takashima, S., Yamada,M., and Hirabayashi, J. (2005) Nat. Methods 2, 851–856
12. Warner, T. G., and O’Brien, J. S. (1979) Biochemistry 18, 2783–278713. Tomin, A., Shkandina, T., and Bilyy, R. (2011) Proc. SPIE 8087, 80876914. Nakano, V., Fontes Piazza, R. M., and Avila-Campos, M. J. (2006) Anaer-
obe 12, 238–24115. O’Farrell, P. H. (1975) J. Biol. Chem. 250, 4007–402116. Helenius, A., and Aebi, M. (2001) Science 291, 2364–236917. Keller, P., Toomre, D., Díaz, E., White, J., and Simons, K. (2001) Nat. Cell
Biol. 3, 140–14918. Azuma, Y., Sato, H., Higai, K., andMatsumoto, K. (2007) Biol. Pharm Bull
30, 1680–168419. Seyrantepe, V., Iannello, A., Liang, F., Kanshin, E., Jayanth, P., Samarani, S.,
Szewczuk, M. R., Ahmad, A., and Pshezhetsky, A. V. (2010) J. Biol. Chem.285, 206–215
20. Elliott, M. R., and Ravichandran, K. S. (2010) J. Cell Biol. 189, 1059–107021. Friggeri, A., Banerjee, S., Biswas, S., de Freitas, A., Liu, G., Bierhaus, A., and
Abraham, E. (2011) J. Immunol. 186, 6191–619822. Elliott, M. R., Chekeni, F. B., Trampont, P. C., Lazarowski, E. R., Kadl, A.,
Walk, S. F., Park, D., Woodson, R. I., Ostankovich, M., Sharma, P., Lysiak,J. J., Harden, T. K., Leitinger, N., and Ravichandran, K. S. (2009) Nature461, 282–286
23. Lauber, K., Bohn, E., Kröber, S. M., Xiao, Y. J., Blumenthal, S. G., Linde-mann, R. K., Marini, P., Wiedig, C., Zobywalski, A., Baksh, S., Xu, Y.,Autenrieth, I. B., Schulze-Osthoff, K., Belka, C., Stuhler, G., and Wessel-borg, S. (2003) Cell 113, 717–730
24. Savill, J. S., Henson, P. M., and Haslett, C. (1989) J. Clin. Investig. 84,1518–1527
25. Savill, J., Fadok, V., Henson, P., andHaslett, C. (1993) Immunol. Today 14,131–136
26. Mercer, J., and Helenius, A. (2008) Science 320, 531–53527. Mercer, J., and Helenius, A. (2009) Nat. Cell Biol. 11, 510–52028. Obeid, M., Tesniere, A., Ghiringhelli, F., Fimia, G. M., Apetoh, L., Perfet-
tini, J. L., Castedo, M., Mignot, G., Panaretakis, T., Casares, N., Métivier,D., Larochette, N., van Endert, P., Ciccosanti, F., Piacentini, M., Zitvogel,L., and Kroemer, G. (2007) Nat. Med. 13, 54–61
Distinct Glycosylation of Apoptotic Blebs
JANUARY 2, 2012 • VOLUME 287 • NUMBER 1 JOURNAL OF BIOLOGICAL CHEMISTRY 503
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

HerrmannJanko, Kirsten Lauber, Martin Schiller, Georg Schett, Rostyslav S. Stoika and Martin
ChristinaVolodymyr Antonyuk, Yuriy Ya. Kit, Matthias Zirngibl, Barbara G. Fürnrohr, Rostyslav O. Bilyy, Tanya Shkandina, Andriy Tomin, Luis E. Muñoz, Sandra Franz,
MicroparticlesMacrophages Discriminate Glycosylation Patterns of Apoptotic Cell-derived
doi: 10.1074/jbc.M111.273144 originally published online November 10, 20112012, 287:496-503.J. Biol. Chem.
10.1074/jbc.M111.273144Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2011/11/10/M111.273144.DC1
http://www.jbc.org/content/287/1/496.full.html#ref-list-1
This article cites 28 references, 9 of which can be accessed free at
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from


















