
Lymphatic Drainage Mice That Lack Dermal K14-VEGFR-3-Ig in ...
11
of April 16, 2018. This information is current as Lymphatic Drainage Mice That Lack Dermal K14-VEGFR-3-Ig in Impaired Humoral Immunity and Tolerance Melody A. Swartz Emma L. Kuan, Kari Alitalo, Gwendalyn J. Randolph and Susan N. Thomas, Joseph M. Rutkowski, Miriella Pasquier, http://www.jimmunol.org/content/189/5/2181 doi: 10.4049/jimmunol.1103545 July 2012; 2012; 189:2181-2190; Prepublished online 27 J Immunol Material Supplementary 5.DC1 http://www.jimmunol.org/content/suppl/2012/07/27/jimmunol.110354 References http://www.jimmunol.org/content/189/5/2181.full#ref-list-1 , 22 of which you can access for free at: cites 62 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2012 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 16, 2018 http://www.jimmunol.org/ Downloaded from by guest on April 16, 2018 http://www.jimmunol.org/ Downloaded from
-
Upload
nguyenphuc -
Category
Documents
-
view
213 -
download
0
Transcript of Lymphatic Drainage Mice That Lack Dermal K14-VEGFR-3-Ig in ...

of April 16, 2018.This information is current as
Lymphatic Drainage Mice That Lack DermalK14-VEGFR-3-Igin
Impaired Humoral Immunity and Tolerance
Melody A. SwartzEmma L. Kuan, Kari Alitalo, Gwendalyn J. Randolph and Susan N. Thomas, Joseph M. Rutkowski, Miriella Pasquier,
http://www.jimmunol.org/content/189/5/2181doi: 10.4049/jimmunol.1103545July 2012;
2012; 189:2181-2190; Prepublished online 27J Immunol
MaterialSupplementary
5.DC1http://www.jimmunol.org/content/suppl/2012/07/27/jimmunol.110354
Referenceshttp://www.jimmunol.org/content/189/5/2181.full#ref-list-1
, 22 of which you can access for free at: cites 62 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2012 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Impaired Humoral Immunity and Tolerance inK14-VEGFR-3-Ig Mice That Lack Dermal LymphaticDrainage
Susan N. Thomas,* Joseph M. Rutkowski,* Miriella Pasquier,* Emma L. Kuan,†
Kari Alitalo,‡ Gwendalyn J. Randolph,†,1 and Melody A. Swartz*,x
Lymphatic vessels transport interstitial fluid, soluble Ag, and immune cells from peripheral tissues to lymph nodes (LNs), yet the
contribution of peripheral lymphatic drainage to adaptive immunity remains poorly understood. We examined immune responses
to dermal vaccination and contact hypersensitivity (CHS) challenge in K14-VEGFR-3-Ig mice, which lack dermal lymphatic
capillaries and experience markedly depressed transport of solutes and dendritic cells from the skin to draining LNs. In response
to dermal immunization, K14-VEGFR-3-Ig mice produced lower Ab titers. In contrast, although delayed, T cell responses were
robust after 21 d, including high levels of Ag-specific CD8+ T cells and production of IFN-g, IL-4, and IL-10 upon restimulation.
T cell-mediated CHS responses were strong in K14-VEGFR-3-Ig mice, but importantly, their ability to induce CHS tolerance in
the skin was impaired. In addition, 1-y-old mice displayed multiple signs of autoimmunity. These data suggest that lymphatic
drainage plays more important roles in regulating humoral immunity and peripheral tolerance than in effector T cell
immunity. The Journal of Immunology, 2012, 189: 2181–2190.
It was suggested four decades ago that lymphatic drainage wasinvolved in local immune tolerance (1, 2), yet since then, therole of lymph drainage (i.e., transport of soluble components
from the periphery to lymph nodes (LNs)) in modulating adaptiveimmunity and tolerance has remained largely unexplored. There is agrowing appreciation that tolerance to inhaled and ingested Ags isinduced at the level of the draining LN (3) and that LN stromal cellsmediate tolerance to self-Ag by endogenous expression on MHCmolecules (4), but the importance of lymphatic drainage function,and soluble lymph delivery to the LN, is poorly understood.Altered immunity has been reported in primary lymphedema,
a congenital pathology of dysfunctional lymphatic drainage that
commonly leaves edematous limbs prone to infections (5). Fur-thermore, lymphangiogenesis often occurs in areas of chronicinflammation such as in Crohn’s disease (6) and in LNs draininginflamed areas or after vaccination (7, 8), yet the function of anexpanded lymphatic network in inflammation is unclear. In con-trast, inflammatory lymphangiogenesis has been correlated withgraft rejection in corneal, islet, and renal transplants (9–14), but ithas also been shown to promote resolution of experimental in-flammation (15) and in humans was correlated with long-termsurvival of renal transplants (16). In tumors, increased lymphaticdrainage is correlated with tumor progression (17, 18), and tumor-associated lymphangiogenesis was recently shown to promoteimmune tolerance (19, 20). Thus, lymphatic drainage likely playscomplex roles in modulating immunity.Adaptive immune responses can be initiated by either peripheral
dendritic cells (DCs) that pick up Ag in the periphery and migrateto the draining LNs or by LN-resident DCs that pick up soluble,lymph-borne Ag; both presumably educate T cells in the LN (21).In addition, lymph-borne Ag is taken up by subcapsular macro-phages for delivery to LN follicle-resident B cells (22) or directlyby the B cells themselves in the case of small Ags (23) to initiatehumoral responses. Interestingly, cytokine responses by LN-resident DCs that take up soluble Ag differ from those of Ag-bearing DCs that travel to the LN after being activated in theperiphery (24), suggesting the importance of lymph-borne Ag andtherefore lymphatic drainage in fine-tuning the immune response.In addition to facilitating adaptive immunity, the LN is an im-
portant site for the maintenance of self-tolerance (25). The stromalcells of the LN T cell zone form the structural network thatguides lymphocyte trafficking (26) through secretion of the CCR7ligands CCL21 and CCL19, which are essential to the positioningof CCR7+ regulatory T (TReg) cells, naive T cells, and APCs in theLN paracortex (27). Without such positioning, as in the case of pltmice that lack CCR7 ligands (and subsequently have impairedtrafficking of DCs and naive T cells into the LN), Ag-specificeffector T cell responses can be mounted in the spleen, leadingto potent although delayed T cell immunity (28). However, self-
*Institute of Bioengineering, School of Life Sciences, Swiss Federal Institute ofTechnology, Lausanne (EPFL), 1015 Lausanne, Switzerland; †Department of Geneand Cell Medicine, Immunology Institute, Mount Sinai School of Medicine, NewYork, NY 10029; ‡Molecular/Cancer Biology Laboratory, Biomedicum Helsinki,University of Helsinki, SF-00014 Helsinki, Finland; and xSwiss Institute for Exper-imental Research, Swiss Federal Institute of Technology, Lausanne (EPFL), 1015Lausanne, Switzerland
1Current address: Department of Pathology and Immunology, Washington UniversitySchool of Medicine, St. Louis, MO.
Received for publication December 13, 2011. Accepted for publication June 22,2012.
This work was supported by grants from the National Institutes of Health(HL096539), the European Research Council (206653-2), the Swiss National ScienceFoundation (310010), the U.S. Department of Defense Breast Cancer Research Pro-gram (BC086586), and the American Heart Association (0740052N). S.N.T. was anInternational Fellow of the Whitaker Foundation.
Address correspondence and reprint requests to Dr. Melody A. Swartz, Institute ofBioengineering and Swiss Institute for Experimental Cancer Research, School of LifeSciences, Ecole Polytechnique Federale de Lausanne, 1015 Lausanne, Switzerland.E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: aoo, acetone with 20% olive oil; CHS, contacthypersensitivity; DC, dendritic cell; dLN, dermal-draining lymph node; DNFB, dini-trofluorobenzene; DNTB, 5,59-dithiobis(2-nitrobenzoic acid); i.d., intradermal(ly);LN, lymph node; mLN, mesenteric lymph node; p.i., postimmunization; PNAd,peripheral lymph node addressin; TREG, regulatory T; WT, wild-type.
Copyright� 2012 by The American Association of Immunologists, Inc. 0022-1767/12/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1103545
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

tolerance mechanisms fail in CCR72/2 mice, which developgeneralized autoimmunity (29). This is because, in addition toCCR7 being required for the establishment of central tolerancewithin the thymus (29, 30), TReg cells require LN occupancy fortheir activation (25, 31–33). Furthermore, LN-resident stromalcells and lymphatic endothelium can present endogenous periph-eral Ag for the deletion of self-reactive CD8+ T cells (26, 34, 35).Thus, both APCs activated in the periphery, which then travel tothe LN, as well as APCs activated in the LN from lymph-bornesoluble Ag (including, importantly, B cells) contribute to theimmune regulatory balance (25).Hence, although the lymphatic system in immunity has largely
been regarded in its cell transport roles, providing physical routesfor immune cell trafficking from the periphery to the LN andchemokines for positioning these cells within the LN, the immuneimplications of fluid drainage by lymphatics from the periphery tothe LN remain poorly understood. Unfortunately, mouse models ofimpaired lymphatic drainage require causative gene defects thatmay directly affect immunity, and inflammation associated withsurgical or chemical disruption of lymphatic vessels also stronglyaffects immune responses. K14-VEGFR-3-Ig mice express solubleVEGFR-3-Ig via the keratin 14 promoter, which results in de-fective lymphatic growth that is restricted to the skin. As adults,they display a paucity of initial dermal lymphatic capillaries anddecreased fluid clearance from the skin, and no other knownphysiological manifestations have been observed (36, 37); themice survive to an advanced age (up to 2 y), the LNs are intact,and lymphatic vessels in other (nondermal) tissues appear normal(36). We examined the adaptive immune response to dermalversus peritoneal vaccination as well as acquired tolerance todermal contact hypersensitivity (CHS) in these mice. Our findingssupport the hypothesis that local lymphatic drainage is critical forhumoral immunity and acquired tolerance but less important foreffector T cell immunity after vaccination.
Materials and MethodsAnimals
K14-VEGFR-3-Ig mice are described earlier (36). Littermates lacking thetransgene were used as wild-type (WT) controls. Mice were used betweenages of 8 and 20 wk, except in studies examining autoimmune phenotypesin aged mice, which used mice at 12–18 mo. Notably, the mice weremaintained on a special chow diet that lacks any chicken-derived products(Diet 2918; Harlan Laboratories, Itingen, Switzerland). All procedureswere approved by the Office Veterinaire Cantonale Vaud (Switzerland).
Lymphatic uptake
The lymphatic uptake rate was determined as described previously (38).Briefly, mice were anesthetized (using an i.p. injection of 65 mg/kg ket-amine and 13 mg/kg xylazine), and a 30-gauge needle catheter containing0.9% NaCl with 2 mg/ml FITC–dextran 70 kDa (Invitrogen, Carlsbad,CA) was carefully placed intradermally (i.d.) into the tail tip. The catheterwas attached to a low-pressure reservoir that permitted 5-cm stepwisechanges from 40 to 60 cm H2O pressure. These infusion pressures allowedphysiologic uptake into the lymphatic capillaries (when present) andpostcapillary venules while minimizing potential swelling or tissue dam-age. By measuring the infusion flow rate together with the movement ofthe fluorescent dextran in the interstitial space (using a Leica MZ16 FAstereomicroscope) as functions of infusion pressure, we could estimatethe hydraulic conductivity and relative clearance from the tissue. Calcu-lations were made according to the theoretical framework outlined pre-viously (38). Results (representing percent volume of injected solutioncleared per time and pressure drop) were normalized relative to WT lit-termates.
Trypan blue drainage assay
Twenty microliters of 0.4% trypan blue (Invitrogen) was injected i.d. intothe front and back legs and tail. Ten minutes postinjection, mice weresacrificed, and LNs were removed.
FITC Painting
The backs of mice were shaved and painted with 8% FITC (type I isomers;Sigma-Aldrich, St. Louis, MO) diluted in a 1:1 mixture of acetone anddibutylphthalate. After 4 d, mice were sacrificed, and draining LNs wereanalyzed for FITC+CD11c+ DCs by flow cytometry (FACSCanto; BDBiosciences, San Diego, CA).
In vivo migration assay
Yellow-green polystyrene microspheres of 1 mm in diameter (Polysciences,Warrington, PA) diluted in PBS 1:20 were injected i.d. in each hind limb(40 ml/injection) and tail (20 ml). Twenty-four hours later, LNs wereharvested, treated for 1 h with collagenase D in HBSS (both from Sigma-Aldrich) at 37˚C, homogenized, and passed through a 70-mm filter (BDBiosciences) and analyzed by flow cytometry.
Immunization
Mice were immunized i.d. in each hind or fore limb (40 ml/injection) andtail (20 ml) with 0.1 mg/ml chicken OVA (Profos, Marburg, Germany) in 1)PBS, 2) a 1:1 emulsion of PBS and alum (ThermoFisher, Rockford, IL), or3) PBS with 25 mg Ultrapure 0111:B4 LPS (InvivoGen, Nunningen,Switzerland); solutions were injected i.d. using a 30-gauge needle. In se-lect experiments, mice were immunized i.p. (100 ml).
Flow cytometry
All Abs used for flow cytometry were from eBioscience (San Diego, CA),unless otherwise stated. Streptavidin–Alexa 649 was used in conjunctionwith biotinylated Abs. Abs were prepared in HBSS/0.5% BSA and addedto samples prior to incubation at 4˚C for 30 min in the dark. For detectionof the OVA H-2Kb MHCI peptide SIINFEKL, samples were stained withPE-conjugated SIINFEKL tetramer (ProImmune, Oxford, U.K.) diluted1:10 in HBSS/0.5% BSA for 10 min at room temperature in the dark.For intracellular staining of Langerin, surface-stained cells were fixed/permeabilized with Cytofix/Cytoperm (BD Biosciences) for 30 min at4˚C, followed by goat anti-mouse Langerin (Santa Cruz Biotechnology,Santa Cruz, CA) and anti-goat PE (Jackson ImmunoResearch Laboratories,New Market, U.K.) staining. Both Abs were diluted in perm/wash buffer(BD Biosciences). For Foxp3 staining, cells were fixed/permeabilizedusing Foxp3 Fixation/Permeabilization kit (eBioscience) overnight at 4˚Cand stained the following day in permeabilization buffer with anti-Foxp3.Data were acquired in a Dako CyAn flow cytometer (DakoCytomation,Glostrup, Denmark) with compensation using either calibration beads (BDBiosciences) or single-stained cells. Data analysis was performed usingFlowJo software (version 8.8; Tree Star, Ashland, OR).
Immunofluorescence
Frozen LNs were cryosectioned (8 mm) and subjected to standard immu-nofluorescence protocols using the following anti-mouse Abs: hamster anti-CD3ε (1:100; BD Pharmingen, San Diego, CA), biotinylated rat anti-B220(1:100; BD Biosciences), Alexa Fluor 647-conjugated CD11c (1:40; eBio-science), rat anti–ER-TR7 (1:50; Hycult Biotech, Uden, The Netherlands),rat anti-peripheral LN addressin (PNAd) (1:100; BioLegend, Uithoorn, TheNetherlands), rabbit anti–Lyve-1 (1:250; Teco Medical, Neufahrn, Ger-many), rat anti-CD21/CD35 (1:400; eBioscience), rat anti-CXCL13 (1:10;R&D Systems, Minneapolis, MN), goat anti-gp38 (1:50; R&D Systems),rabbit anti-collagen I (1:50; Santa Cruz Biotechnology), and biotinylatedrabbit anti-collagen IV (1:800; Abcam, Cambridge, U.K.); fluorescentlyconjugated secondary Abs and streptavidin were obtained from Invitrogen.Paraffin-embedded skin sections were cut into 8-mm-thick sections,blocked in goat serum, and stained with goat anti-mouse IgG-Alexa Fluor647. Alternatively, ear tissue samples were whole-mount stained usingstandard techniques (39). Frozen sections (8 mm) of organs of healthy WTmice were first incubated in donkey serum and then stained with 1% serafrom K14-VEGFR-3-Ig mice or age-matched WT littermates; after wash-ing, sections were incubated with donkey anti-mouse IgG-AF647 (1:200).Sections were counterstained with DAPI (Vector Laboratories, Burlin-game, CA) and imaged using a LSM 510 or 710 confocal microscope (CarlZeiss, Feldbach, Switzerland). Sections were scored in a blinded fashion.
Serum anti-OVA Ab titers
On day 21 postimmunization (p.i.) with a day 10 boost, mice were sacri-ficed, and their sera were analyzed by ELISA. Ninety-six-well plates (BDBiosciences) were coated overnight with 10 mg/ml OVA diluted in PBS.Plates were then blocked with 2.5% casein in PBS, and samples wereadded in blocking solution for 2 h, rinsed, and incubated with either HRP-conjugated anti-IgG, HRP-conjugated anti-IgG2c, or biotinylated anti-IgG1
2182 LYMPHATIC DRAINAGE IN REGULATING IMMUNE RESPONSE
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

(all from eBioscience). 3,39,5,59-Tetramethylbenzidine (GE Healthcare,Glattbrugg, Switzerland) was added and incubated 30 min room temper-ature in the dark. The reaction was quenched with 2 N H2SO4, andabsorbances were read on a plate reader (Tecan, Mannedorf, Switzerland).For IgG1 titers, an additional 30-min incubation at room temperature withstreptavidin-conjugated HRP (R&D Systems) diluted 1:200 in 2.5% caseinsolution was included between the secondary Ab incubation and 3,39,5,59-tetramethylbenzidine development.
In vitro recall response of T cells
On day 9 or 21 p.i., the draining LNs (popliteal, sciatic, and sacral) andspleens were harvested and digested to form cell suspensions as describedabove. They were plated at 105 cells/well of a 96-well plate in restimulationmedia (RPMI 1640 medium with 10% FBS, 50 mM 2-ME, and 1%penicillin/streptomycin; all from Life Technologies, Carlsbad, CA) with 0 or5 mg/ml Endograde OVA (Hyglos, Regensburg, Germany). One hundredten hours after initial plating, cell supernatants were harvested and stored at280˚C until IFN-g, IL-10, and IL-4 cytokine production was analyzed byELISA (eBioscience), according to the manufacturer’s protocol.
Adoptive transfer experiments
OT-I and OT-II splenocytes were treated with ammonium chloride potas-sium RBC lysis buffer and then positively selected for CD8+ cells (Ly-2magnetic beads; Miltenyi Biotec, Bergisch Gladbach, Germany) or CD4+
cells (L3T4 beads; Miltenyi Biotec), respectively. Cells were labeled with1 mM final concentration of CFSE (Invitrogen) prior to adoptive transfer of1 3 106 OT-I cells or 2 3 106 OT-II cells in IMDM by tail vein injectioninto naive mice. After 3 d, mice were immunized with OVA plus LPS i.d.in the back legs and tail. Three or 6 d p.i., dermal-draining LNs (dLNs) andspleens were harvested and processed to make single-cell suspensions.Cells were analyzed by flow cytometry. Because K14-VEGFR-3-Ig micewere on a C57BL/6 CD45.2 background, as were OT-I and OT-II mice, wecould only trace the OT-I and OT-II cells by CFSE labeling before adoptivetransfer. The percentages of unproliferated and proliferated OT cells wereestimated by summing the unproliferated (G0) cells and proliferating (G1–G6) cells, respectively, and normalizing to the total numbers of parentalcells they arose from. For days 9 and 12 p.i., the frequencies of unpro-liferated cells were normalized to the total CFSE+ cell counts on day 3 p.i.
Contact hypersensitivity
To establish contact sensitivity, 25 ml 0.5% dinitrofluorobenzene (DNFB)in acetone with 20% olive oil (aoo) was painted onto the shaved backs ofmice. Five days later, one ear was challenged with 0.3% DNFB. Forty-eight hours later, ear swelling was measured relative to the untreated ear. Inseparate experiments to determine tolerance to DNFB-mediated CHS, wefirst induced tolerance by treating the shaved stomachs of naive mice with100 ml 1% 5,59-dithiobis(2-nitrobenzoic acid) (DNTB) in aoo on day 0.We then sensitized the mice on day 7 by treating their shaved backs with25 ml 0.5% DNFB in aoo. Finally, mice were challenged on day 12 todetermine the extent of tolerance by treating one ear with 0.3% DNFB inaoo. Forty-eight hours later, ear thickness was measured, and ear speci-mens were harvested. Alternatively, resolution of ear swelling was deter-mined by measuring ear thickness for 10–14 d postchallenge.
Histological analysis
Paraffin-embedded ears and frozen spleens were cut into 4- or 8-mm-thicksections and H&E stained. Paraffin-embedded tail skin was cut into 8-mm-thick sections and stained with Miller’s stain. Samples were imaged witha Zeiss (Feldbach, Switzerland) MRc camera.
Serum isotype titers and serum dsDNA titers
Serum isotype values were determined using an ELISA kit (BD Bio-sciences), and dsDNA titers were determined by ELISA (VWR, Nyon,Switzerland), according to the manufacturer’s protocols.
Ex vivo B cell stimulation
Splenocytes were first treated with ammonium chloride potassium RBClysis buffer, and then splenocyte and dermal LN cell suspensions werenegatively selected for B cells (Ly-48, L3T4, and Ter-119 magnetic beads;Miltenyi Biotec). B cells were plated at 5 3 105 cells/well in 96-wellsuspension culture plates (Greiner Bio-one, Frickenhausen, Germany) inIMDM with 10% FBS, 50 mM 2-ME, and 1% penicillin/streptomycin (allfrom Life Technologies) and treated with 0.2 mg/ml Ultrapure LPS(InvivoGen). After 24 h, supernatants were harvested and stored at 280˚Cuntil analysis for IFN-g by ELISA (eBioscience). Cells were washed,
stained for B220 and B cell activation markers CD86 and IgM, and ana-lyzed by flow cytometry.
Statistical analysis
Data are represented as the mean with the SEM. Statistics were calculatedusing GraphPad Prism 5 (GraphPad, La Jolla, CA) software. Statisticalsignificance was defined as p , 0.05 following one-way ANOVA and posthoc analysis. When normality tests failed, Kruskal–Wallis tests wereperformed. For contingency analysis, Fisher’s exact test was used.
ResultsReduced lymphatic drainage and DC migration inK14-VEGFR-3-Ig mice
We first established the degree of impairment of lymphatic drainageand transport of peripheral DCs to the dLNs in K14-VEGFR-3-Igmice. Consistent with their lack of dermal lymphatic capillaries(36), lymphatic drainage from the skin of K14-VEGFR-3-Ig micewas severely impaired, as seen qualitatively by the lack of trypanblue in the dLN 10 min after i.d. injection (Supplemental Fig. 1A)and quantitatively using a previously established tail model ofquantitative microlymphangiography (Fig. 1A) (40).DC trafficking from the skin to the dLN was virtually abrogated
in these mice, as demonstrated by a lack of FITC+CD11c+ andFITC+langerin+CD11c+ cells in dLNs 4 d after application ofFITC-containing contact sensitizer to the epidermis (“FITCpainting”; Fig. 1B, 1C). This abolishment of DC trafficking toskin-draining LNs was confirmed by the absence of bead+CD11c+
cells in the dLNs 24 h after i.d. injection of 1-mm fluorescentmicrospheres (Fig. 1D), which can only reach the LN after beingtaken up by peripheral monocytes that differentiate into DCs (21).Total frequencies of Lang+CD11c+ cells in dLNs were also se-verely decreased (Fig. 1E). Importantly, dLNs of K14-VEGFR-3-Ig mice had normal amounts of CCL21, a chemokine that attractsDCs and naive T cells to the LN (Supplemental Fig. 1B), permilligram of total dLN protein, indicating that the impaired DCtrafficking was not due to decreased chemokine signaling in theLN. Thus, the severely impaired lymphatic drainage and DCtrafficking from skin to dLNs in K14-VEGFR-3-Ig mice waspresumably due to the lack of dermal lymphatic capillaries.
Altered LN organization in K14-VEGFR-3-Ig mice
We found disorganized B cell follicles in the dLNs but not mes-enteric LNs (mLN) of K14-VEGFR-3-Ig mice (Fig. 1F); specifi-cally, B cells were found scattered throughout the dLNs amongT cells and were less concentrated in follicles compared withthose in WT mice. Similarly, the distributions but not amounts ofCCL21 and CXCL13, a chemokine that directs B cell positioningwithin the LN, were abnormal in the dLNs of K14-VEGFR-3-Igmice (Supplemental Fig. 1B‑D). T cell zone fibroblastic reticularcells and reticular fibers, which regulate T cell migration (41), alsoappeared highly disorganized in dLN (Fig. 1G, Supplemental Fig.2A) but not mLN (Supplemental Fig. 2A) of transgenic mice.Finally, the lymphatic endothelial network and distribution offollicular DCs within the dLNs, but not mLNs, of transgenic miceappeared very disorganized (Supplemental Fig. 2B) and containedsignificantly smaller high endothelial venules (PNAd+) than inWT dLNs (Fig. 1H, 1I, Supplemental Fig. 2C).Interestingly, although naive dLNs of the transgenic mice were
smaller (36) and less organized than their WT counterparts (Fig.1F–H), they contained similar fractions (as percentage of CD45+
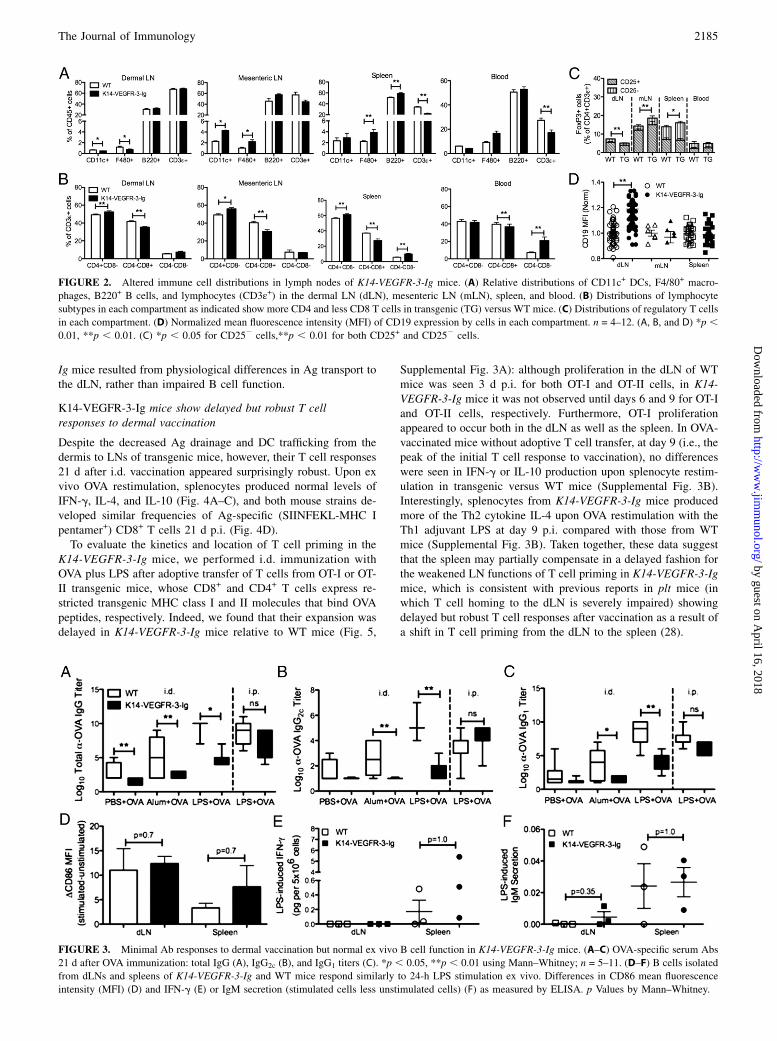
cells) of B cells (B220+) and T cells (CD3ε+) (Fig. 2A). Fractionsof DCs (CD11c+) and macrophages (F4/80+) were lower in dLNsbut higher in mLNs of transgenic mice (Fig. 2A). However, higherCD4/CD8 T cell ratios were found in both dLNs and mLNs aswell as the spleens (Fig. 2B) of K14-VEGFR-3-Ig mice compared
The Journal of Immunology 2183
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

with WT mice. Interestingly, there were fewer TReg cells (Foxp3+,
as the percentage of CD4+CD3ε+ cells) in the dLN and more in themLN of transgenic mice compared with WT mice (Fig. 2C),suggesting that deficiencies in TReg cell homing to the dLN inK14-VEGFR-3-Ig mice is compensated by homing to other LNs.The spleens and blood of transgenic mice contained higher
frequencies of double-negative (CD42CD8a2) CD3ε+ cells (Fig.2B), which may negatively regulate immune responses (42).However, no significant differences were seen in systemic TReg
cell frequencies other than a small increase in the adaptive TReg
cell compartment (CD252Foxp3+) (Fig. 2C). Another strikingdifference was the increased expression of B cell coreceptor CD19by cells in the dLNs, but not mLNs or spleens, of transgenic mice(Fig. 2D). Because CD19 increases B cell sensitivity to Ag-specific stimulation, its increased expression could be associatedwith autoantibody production (43) and autoimmunity (44).
Poor humoral responses to dermal, but not peritoneal,vaccination in K14-VEGFR-3-Ig mice
Lymph flow from peripheral tissues provides Ag to draining LNsand transports APCs to LNs to initiate immune responses (21). To
determine how adaptive immunity to dermal challenge was af-fected in K14-VEGFR-3-Ig mice, we compared immune respon-ses to the model Ag OVA (confirmed endotoxin-free beforeinjection) after i.d. or i.p. immunization with LPS (a Th1-promoting adjuvant) or i.d. immunization with alum (a Th2-promoting adjuvant). We note that mild responses to OVA plussaline in WT mice were expected because OVA is a foreign Ag,and the mice had been maintained on a chicken product-free chowdiet.On day 21 postimmunization (p.i.) with a day 10 boost, we found
drastically reduced anti-OVA Ab titers in K14-VEGFR-3-Ig micecompared with WT controls with both adjuvants upon i.d. chal-lenge (Fig. 3A–C). However, when immunized i.p., anti-OVA Abtiters in response to OVA plus LPS were normal (Fig. 3A–C). Poorhumoral responses in response to i.d. challenge were not appar-ently due to deficiency in the functionality of the B cells, becauseB cells isolated from dLN and spleens of transgenic miceresponded similarly to those from WT mice when stimulated exvivo with LPS in terms of maturation (Fig. 3D) as well as IFN-gand IgM production (Fig. 3E, 3F). Thus, our data suggest that thereduced Ab response to i.d. immunization seen in K14-VEGFR-3-
FIGURE 1. Reduced lymphatic drainage and dendritic cell transport to lymph nodes and abnormal architecture in skin-draining LNs of K14-VEGFR-3-Ig
mice. (A) Lymphatic uptake after i.d. injection into the tail representing volume of fluid drained per tissue volume per time and pressure gradient, nor-
malized to WT mice; n = 6–7. Representative flow cytometry plots (B) and quantification of DC trafficking from periphery to LN after FITC painting (C) or
i.d. injection of 1 mm yellow-green polystyrene microspheres (D); n = 4–7. (E) Steady-state Langerin+ cell numbers in the dLN. (F–H) Immunohisto-
chemistry of dLN (brachial and inguinal) and mesenteric-draining LN sections stained for B cells (B220, red), T cells (CD3ε, green), and DCs (CD11c,
blue) (F); (fibroblastic reticular cells [ER-TR7; (G) green; (H) red] (G, H) and high endothelial venules [PNAd; green]). Scale bars: (F) top, 200 mm; middle,
50 mm; bottom, 100 mm, (G) 300 mm, (H) 20 mm. (I) Relative expression of PNAd in high endothelial venules from immunostaining. *p , 0.05 by Mann–
Whitney.
2184 LYMPHATIC DRAINAGE IN REGULATING IMMUNE RESPONSE
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Ig mice resulted from physiological differences in Ag transport tothe dLN, rather than impaired B cell function.
K14-VEGFR-3-Ig mice show delayed but robust T cellresponses to dermal vaccination
Despite the decreased Ag drainage and DC trafficking from thedermis to LNs of transgenic mice, however, their T cell responses21 d after i.d. vaccination appeared surprisingly robust. Upon exvivo OVA restimulation, splenocytes produced normal levels ofIFN-g, IL-4, and IL-10 (Fig. 4A–C), and both mouse strains de-veloped similar frequencies of Ag-specific (SIINFEKL-MHC Ipentamer+) CD8+ T cells 21 d p.i. (Fig. 4D).To evaluate the kinetics and location of T cell priming in the
K14-VEGFR-3-Ig mice, we performed i.d. immunization withOVA plus LPS after adoptive transfer of T cells from OT-I or OT-II transgenic mice, whose CD8+ and CD4+ T cells express re-stricted transgenic MHC class I and II molecules that bind OVApeptides, respectively. Indeed, we found that their expansion wasdelayed in K14-VEGFR-3-Ig mice relative to WT mice (Fig. 5,
Supplemental Fig. 3A): although proliferation in the dLN of WTmice was seen 3 d p.i. for both OT-I and OT-II cells, in K14-VEGFR-3-Ig mice it was not observed until days 6 and 9 for OT-Iand OT-II cells, respectively. Furthermore, OT-I proliferationappeared to occur both in the dLN as well as the spleen. In OVA-vaccinated mice without adoptive T cell transfer, at day 9 (i.e., thepeak of the initial T cell response to vaccination), no differenceswere seen in IFN-g or IL-10 production upon splenocyte restim-ulation in transgenic versus WT mice (Supplemental Fig. 3B).Interestingly, splenocytes from K14-VEGFR-3-Ig mice producedmore of the Th2 cytokine IL-4 upon OVA restimulation with theTh1 adjuvant LPS at day 9 p.i. compared with those from WTmice (Supplemental Fig. 3B). Taken together, these data suggestthat the spleen may partially compensate in a delayed fashion forthe weakened LN functions of T cell priming in K14-VEGFR-3-Igmice, which is consistent with previous reports in plt mice (inwhich T cell homing to the dLN is severely impaired) showingdelayed but robust T cell responses after vaccination as a result ofa shift in T cell priming from the dLN to the spleen (28).
FIGURE 2. Altered immune cell distributions in lymph nodes of K14-VEGFR-3-Ig mice. (A) Relative distributions of CD11c+ DCs, F4/80+ macro-
phages, B220+ B cells, and lymphocytes (CD3ε+) in the dermal LN (dLN), mesenteric LN (mLN), spleen, and blood. (B) Distributions of lymphocyte
subtypes in each compartment as indicated show more CD4 and less CD8 T cells in transgenic (TG) versus WT mice. (C) Distributions of regulatory T cells
in each compartment. (D) Normalized mean fluorescence intensity (MFI) of CD19 expression by cells in each compartment. n = 4–12. (A, B, and D) *p ,0.01, **p , 0.01. (C) *p , 0.05 for CD252 cells,**p , 0.01 for both CD25+ and CD252 cells.
FIGURE 3. Minimal Ab responses to dermal vaccination but normal ex vivo B cell function in K14-VEGFR-3-Ig mice. (A–C) OVA-specific serum Abs
21 d after OVA immunization: total IgG (A), IgG2c (B), and IgG1 titers (C). *p , 0.05, **p , 0.01 using Mann–Whitney; n = 5–11. (D–F) B cells isolated
from dLNs and spleens of K14-VEGFR-3-Ig and WT mice respond similarly to 24-h LPS stimulation ex vivo. Differences in CD86 mean fluorescence
intensity (MFI) (D) and IFN-g (E) or IgM secretion (stimulated cells less unstimulated cells) (F) as measured by ELISA. p Values by Mann–Whitney.
The Journal of Immunology 2185
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

K14-VEGFR-3-Ig mice mount robust CHS reactions but fail toinduce experimental CHS tolerance
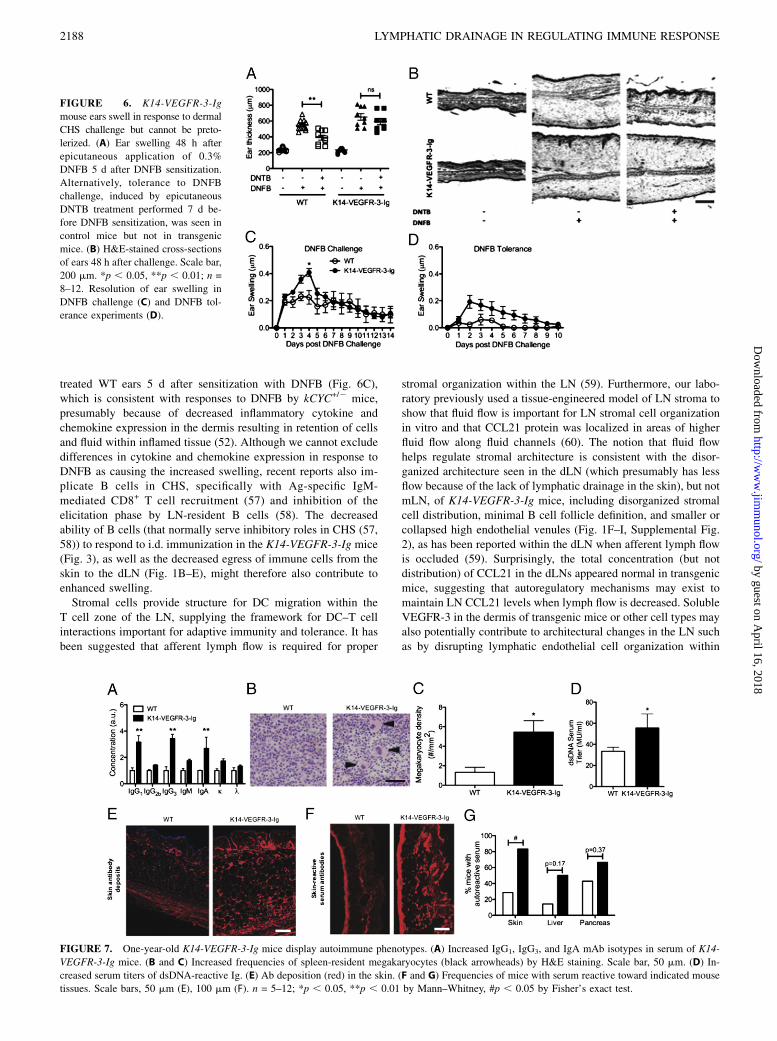
To determine whether T cell responses to dermal immunization inK14-VEGFR-3-Ig mice would be recapitulated in another dermalimmune challenge, we used a model of CHS. In this model, miceare sensitized to DNFB, and ear swelling occurs upon DNFB re-exposure as a result of T cell-mediated immune responses (45,46). In dermal CHS experiments, ears of transgenic mice swelledmore robustly than ears of WT mice for the first 4 d after DNFBchallenge (Fig. 6A–C) but resolved on a similar timescale (Fig.6C). These data support the concept of robust T cell immunity intransgenic mice.In contrast, transgenic mice failed a well-established test of
acquired tolerance to CHS. In WT mice, but not K14-VEGFR-3-Igmice, pretreatment with DNTB (a tolerizing agent to DNFB)prevented DNFB-induced ear swelling (Fig. 6A, 6B, 6D). Thissuggests that transgenic mice are deficient in mechanisms of ac-quired tolerance against skin-encountered Ag and corroboratesolder studies in skin transplants that failed tolerance tests whenlymphatic drainage was blocked (1, 2).In support of the concept that lymphatic drainage to the LN is
important in maintaining immune tolerance against peripheral Ag,we noted several hallmarks of autoimmunity in 1-y-old K14-VEGFR-3-Ig mice (Fig. 7). These included increased serum levelsof IgG1, IgG3, and IgA but not IgG2b, IgM, and K or L chain Abisotypes (indicating disruptions in B cell Ig class switching), thepresence of megakaryocytes in the spleen [associated with in-creased plasma cell survival (47)], elevated serum titers ofdsDNA, and Ab deposits in the skin as well as increased fre-quencies of mice with skin-reactive Abs in their serum (indicatingautoimmune reactions against skin). Finally, in the skin of K14-VEGFR-3-Ig mice lacking Lyve-1+ lymphatic capillaries, we ob-served a decrease in CCL21 levels (Supplemental Fig. 4A), nor-mal frequencies of skin-resident APCs (Supplemental Fig. 4B),and an increase in elastin (Supplemental Fig. 4C). Thus, althoughskin physiology appears altered in K14-VEGFR-3-Ig mice [con-sistent with earlier reports (25)], the normal densities of skin-resident APCs under steady-state conditions may suggest self-regulating mechanisms control APC density in skin, even whenegress into lymphatics is impaired.
DiscussionLNs support the priming of adaptive immune responses as well asthe maintenance of tolerance to autoantigens by orchestrating theinteractions between APCs and T cells (25), whereas lymphaticvessels sustain Ag delivery to the LN through both APC traf-
ficking and direct transport of peripheral lymph-soluble Ag (21).Our data in K14-VEGFR-3-Ig mice, which lack dermal lymphaticcapillaries (36, 37) and thus have reduced transport of soluble Agfrom the skin to the draining LNs (Fig. 1A), support the notionthat lymphatic drainage from the periphery to the draining LN iscritical for inducing humoral immunity and maintaining periph-eral tolerance.K14-VEGFR-3-Ig mice manifested profoundly impaired Ab
responses to i.d. but not i.p. challenge with Ag (Fig. 3A–C), in-dicating that Ag delivery to the LN is critical for adaptive humoralimmunity. This finding complements previous reports showingthat lymph-borne Ag is delivered to LN-resident B cells eitherdirectly through the conduits in the case of smaller Ags (23) or bytransfer from subcapsular macrophages for larger Ags (22). Hu-moral responses to dermal challenge in K14-VEGFR-3-Ig micemight also be impaired by the disorganized dLN architecturegiven recent reports of B cell priming and affinity maturation inresponse to s.c. challenge requiring the specialized topography ofthe LN (48), although B cell function appears robust in ectopiclymphoid structures that form in autoimmune-related disorders(49, 50). B cell function may also be impaired by the delay inCD4+ T cell priming, which plays a well-established role in ma-ture B cell immune responses (51). Regardless of the mechanism,our data are consistent with the notion that lymph flow—whichaffects LN architecture, T cell priming, and Ag transport to the LNconcurrently—plays an important role in humoral immunity.Besides transporting lymph-borne Ag, the lymphatics are
highways for DC homing to LNs in both stead-state and inflamedconditions (21). Although naive K14-VEGFR-3-Ig mice showednormal APC frequencies in the skin (Supplemental Fig. 4B), andDC frequencies in dLNs were only slightly lower than those incontrol mice (Fig. 2), DC migration to the dLN was severelyimpaired when challenged with FITC painting or injection of 1-mm beads (Fig. 1B–E). However, it is important to note that thetotal levels of CCL21 were normal in the dLN (Supplemental Fig.1B), even though CCL21+ lymphatic capillaries were mostly ab-sent in the skin (Supplemental Fig. 4A). In contrast, high endo-thelial venules were smaller in transgenic mice (Fig. 1H, 1I,Supplemental Fig. 2C), and the T cell zone fibroblastic reticularcells and fibers were also disrupted (Supplemental Fig. 2A), whichsuggest that the homing of naive and TReg cells to the LN shouldalso be impaired. Nevertheless, although the migration of APCsfrom the periphery to the draining LN in response to activatingstimuli is widely considered critical for the initiation of Ag-specific T cell immunity, K14-VEGFR-3-Ig mice were able tomount robust (Fig. 4) but delayed (Fig. 5) T cell responses, as
FIGURE 4. Robust Ag-specific T cell responses to
dermal immunization in K14-VEGFR-3-Igmice. Twenty-
one days p.i. with OVA, splenocytes were isolated and
restimulated with and without OVA. IFN-g (A), IL-4 (B),
and IL-10 (C) production by splenocytes with 110-h OVA
restimulation less cytokine production by unstimulated
cells. (D) Frequencies of Ag-specific (SIINFEKL pen-
tamer+) CD8+ splenic T cells. n = 5–12.
2186 LYMPHATIC DRAINAGE IN REGULATING IMMUNE RESPONSE
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

exhibited by normal cytokine production upon splenocyte Agrestimulation after 21 d and normal frequencies of Ag-specificeffector CD8 T cells with vaccination (Fig. 4), as well as robustear swelling in response to dermal DNFB challenge (Fig. 6),a CHS reaction mediated by CD4+ and CD8+ T cells (45, 46).These findings are complementary to recent observations ofdelayed but strong CHS reactions to DNFB challenge in miceheterozygous for the expression of Kaposi’s sarcoma-associatedherpesvirus latent-cycle gene k-cyclin under the control of theVEGFR-3 promoter (kCYC+/2) (52), which exhibit dermal edemaand diminished DC trafficking from the skin to dLN (52, 53).Furthermore, delayed but robust T cell-mediated immune re-sponses have also been observed in plt mice (28), another modelof impaired DC trafficking to the LNs, although by a differentmechanism (namely, the lack of CCR7 ligands that direct homingand positioning between APCs and naive T cells). Interestingly,splenectomized plt mice failed to mediate any T cell response toimmunization (28), and efficient T cell immune responses wereobserved in alymphoplasia (aly/aly) mice (characterized by
a complete lack of LNs and Peyer’s patches) reconstituted withfunctional T cells, although priming was supported in the spleenand liver (54). These data support our observations of T cellproliferation within the spleen of K14-VEGFR-3-Ig mice and in-dicate that lymphatic transport from the skin to the dLN afterimmunization is not essential for the initiation of CD8 T cellresponses.In dermal CHS experiments, ears of K14-VEGFR-3-Ig mice
swelled robustly in response to dermal DNFB challenge (Fig. 6),indicating an efficient cutaneous immune reaction to DNFB me-diated by Ag-specific T cells (45, 46) despite impaired transportfrom the skin to dLNs. Our observations contrast with the previ-ously reported mechanism of VEGFR-3+ corneal DC-dependent,lymphatic-independent delayed-type hypersensitivity reactions toallografts (55). However, DCs outside of the cornea do not expressVEGFR-3 (56); we therefore suppose that mechanisms of immuneresponse in the cornea, which lack lymphatic vessels under non-inflamed conditions, are very different from those in the dermis.Ears of transgenic mice swelled more robustly compared with
FIGURE 5. T cell priming after immunization in K14-VEGFR-3-Ig mice is delayed. The number of OT-I (A, B) and OT-II (C, D) cells responding to
immunization quantified as the total numbers of cells in each generation (G) (A, C) or as the percentage of unproliferated cells (determined by calculating
the number of cells in each generation, normalized by the number of divisions, to determine the total) postimmunization with OVA + LPS in the spleen and
dLNs (B, D). Initial OT-I and OT-II cell priming in WT (i, iii) and K14-VEGFR-3-Ig (ii, iv) mice, respectively. *p , 0.05, **p , 0.01 using two-way
ANOVA; n = 3.
The Journal of Immunology 2187
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
treated WT ears 5 d after sensitization with DNFB (Fig. 6C),which is consistent with responses to DNFB by kCYC+/2 mice,presumably because of decreased inflammatory cytokine andchemokine expression in the dermis resulting in retention of cellsand fluid within inflamed tissue (52). Although we cannot excludedifferences in cytokine and chemokine expression in response toDNFB as causing the increased swelling, recent reports also im-plicate B cells in CHS, specifically with Ag-specific IgM-mediated CD8+ T cell recruitment (57) and inhibition of theelicitation phase by LN-resident B cells (58). The decreasedability of B cells (that normally serve inhibitory roles in CHS (57,58)) to respond to i.d. immunization in the K14-VEGFR-3-Ig mice(Fig. 3), as well as the decreased egress of immune cells from theskin to the dLN (Fig. 1B–E), might therefore also contribute toenhanced swelling.Stromal cells provide structure for DC migration within the
T cell zone of the LN, supplying the framework for DC–T cellinteractions important for adaptive immunity and tolerance. It hasbeen suggested that afferent lymph flow is required for proper
stromal organization within the LN (59). Furthermore, our labo-ratory previously used a tissue-engineered model of LN stroma toshow that fluid flow is important for LN stromal cell organizationin vitro and that CCL21 protein was localized in areas of higherfluid flow along fluid channels (60). The notion that fluid flowhelps regulate stromal architecture is consistent with the disor-ganized architecture seen in the dLN (which presumably has lessflow because of the lack of lymphatic drainage in the skin), but notmLN, of K14-VEGFR-3-Ig mice, including disorganized stromalcell distribution, minimal B cell follicle definition, and smaller orcollapsed high endothelial venules (Fig. 1F–I, Supplemental Fig.2), as has been reported within the dLN when afferent lymph flowis occluded (59). Surprisingly, the total concentration (but notdistribution) of CCL21 in the dLNs appeared normal in transgenicmice, suggesting that autoregulatory mechanisms may exist tomaintain LN CCL21 levels when lymph flow is decreased. SolubleVEGFR-3 in the dermis of transgenic mice or other cell types mayalso potentially contribute to architectural changes in the LN suchas by disrupting lymphatic endothelial cell organization within
FIGURE 6. K14-VEGFR-3-Ig
mouse ears swell in response to dermal
CHS challenge but cannot be preto-
lerized. (A) Ear swelling 48 h after
epicutaneous application of 0.3%
DNFB 5 d after DNFB sensitization.
Alternatively, tolerance to DNFB
challenge, induced by epicutaneous
DNTB treatment performed 7 d be-
fore DNFB sensitization, was seen in
control mice but not in transgenic
mice. (B) H&E-stained cross-sections
of ears 48 h after challenge. Scale bar,
200 mm. *p , 0.05, **p , 0.01; n =
8–12. Resolution of ear swelling in
DNFB challenge (C) and DNFB tol-
erance experiments (D).
FIGURE 7. One-year-old K14-VEGFR-3-Ig mice display autoimmune phenotypes. (A) Increased IgG1, IgG3, and IgA mAb isotypes in serum of K14-
VEGFR-3-Ig mice. (B and C) Increased frequencies of spleen-resident megakaryocytes (black arrowheads) by H&E staining. Scale bar, 50 mm. (D) In-
creased serum titers of dsDNA-reactive Ig. (E) Ab deposition (red) in the skin. (F and G) Frequencies of mice with serum reactive toward indicated mouse
tissues. Scale bars, 50 mm (E), 100 mm (F). n = 5–12; *p , 0.05, **p , 0.01 by Mann–Whitney, #p , 0.05 by Fisher’s exact test.
2188 LYMPHATIC DRAINAGE IN REGULATING IMMUNE RESPONSE
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

dLNs. Regardless of the mechanism, it is likely that the disorga-nized LN stroma and B cell follicles in transgenic mice contributeto their impaired immune responses to dermal vaccination.Our findings of autoimmune outcomes (Fig. 7) as well as the
failure to induce CHS tolerance to DNFB with DNTB pretreat-ment (Fig. 6) in K14-VEGFR-3-Ig mice corroborate 40-y-oldreports (1, 2) that propose lymphatic drainage to a sentinel LNis essential to acquired peripheral tolerance. Those studies useda model of skin transplant where the blood, but not lymphatic,vasculature was connected to the surrounding tissue, and CHStolerance could not be induced. Our current model is not com-plicated by the factor of tissue damage and inflammation, whichcould also affect local immunity, present in the past model. Un-fortunately, those older studies were not followed up since themid-1970s, even though there is an emerging appreciation forcorrect LN function and organization in peripheral tolerance,particularly with regards to the positioning of B cells and traf-ficking of T cells (25). Autoimmune phenotypes have been ob-served in mice lacking either CCR7 or CXCR5 (29, 61), thereceptors for CCL21 and CXCL13 important in the segmentationof the T and B cell zones of the LN, respectively. Although thosereports highlighted the need for proper B cell and TReg cell po-sitioning in the LN, which are guided by CXCL13 and CCL21,our findings suggest that lymphatic drainage may help guide thedistribution and segmentation of these cytokines, because theirdistributions but not total amounts were abnormal in the dLNsof K14-VEGFR-3-Ig mice (Supplemental Fig. 1B). Moreover,CCL21 expression in the skin of transgenic mice appeared dras-tically reduced, mostly because their main source in the skin—lymphatic capillaries—were largely absent (Supplemental Fig.4A). Thus, the lack of lymphatic drainage from skin to the dLNcould promote autoimmunity in multiple ways. However, althoughwe see no evidence of impaired lymphatic drainage in other tis-sues of adult K14-VEGFR-3-Ig mice, altered lymphatics in organsother than the skin during development as characterized byMakinen and colleagues (36) may confound a strict interpretationof the observations of autoimmune phenotypes as being solely theresult of impaired dermal lymphatic drainage.In addition to the decreased availability of peripheral Ag, de-
creased DC trafficking, and disorganization of the dLN, the im-mune cell distributions within the dLN may contribute to thedevelopment of autoimmune reactions seen in 1-y-old K14-VEGFR-3-Ig mice (Fig. 7). For example, K14-VEGFR-3-Ig micedemonstrated higher systemic CD4/CD8 T cell ratios (Fig. 2B) butonly local disturbances (in the LNs) Foxp3+ T cell distributions(Fig. 2C); this is likely a result of disruptions in TReg cellhomeostasic migration to the dLNs via the skin draining lym-phatics (62). CD19, which is expressed by B cells, complexes withthe AgR of B cells to increase sensitivity to Ag-specific stimula-tion. Consequently, increased expression of CD19 is associatedwith autoimmunity (44), and overexpression of CD19 causes in-creased autoantibody production (43). Taken together with thedisrupted stromal cell organization and decreased frequencies ofTReg cells, the increased CD19 expression by cells in the LNs ofK14-VEGFR-3-Ig mice (Fig. 2D) suggests that the B cell regu-latory balance normally maintained by the LN (25) might bedysregulated in the transgenic mice.In addition, there is increasing evidence linking skin fibrosis to
aberrant B cell signaling and autoimmunity. TSK/+ mice, whichare heterozygous for a mutation in fibrillin-1 that results in cuta-neous hyperplasia, exhibit skin sclerosis and autoimmunity as wellas chronic B cell activation as a result of augmented CD19 sig-naling (43). Knockdown of CD19 in TSK/+ mice reduces hypo-dermal thickness and skin collagen deposition, suggesting CD19
involvement in regulating skin pathology. IL-4 deficiency alsorescues TSK/+ mice from skin fibrosis and autoantibody produc-tion (63). K14-VEGFR-3-Ig mice exhibit increased skin elastindeposition (Supplemental Fig. 4C). The correlating impairmentsin LN-resident B cell function, increased CD19 expression, andthe heightened IL-4 production following LPS immunization inK14-VEGFR-3-Ig mice suggest a strong coupling between CD19-mediated B cell autoimmunity and skin fibrosis with impaireddermal lymphatic drainage.In conclusion, this study demonstrates the pivotal role of dermal
lymphatic drainage in the organization and function of B cellsresident in the draining LN, efficient Ab responses to dermalvaccination, and postdevelopmental tolerance induction against Agencountered in the skin. In contrast, T cell responses to dermalchallenge appeared to be only delayed by impaired lymphatic flow,with the exception of possibly skewing responses toward a Th2phenotype. These findings implicate lymphatic drainage as criticalto regulating humoral immunity and peripheral tolerance.
AcknowledgmentsWe thank Patricia Corthesy-Henrioud, Veronique Borel, Didier Foretay,
Jackson Eby, Thomas McKee, Chiara Nembrini, and Andrew Platt for
technical assistance and advice.
DisclosuresThe authors have no financial conflicts of interest.
References1. Friedlaender, M. H., and H. Baer. 1972. Immunologic tolerance: role of the
regional lymph node. Science 176: 312–314.2. Friedlaender, M. H., F. V. Chisari, and H. Baer. 1973. The role of the inflam-
matory response of skin and lymph nodes in the induction of sensitization tosimple chemicals. J. Immunol. 111: 164–170.
3. Hintzen, G., L. Ohl, M. L. del Rio, J. I. Rodriguez-Barbosa, O. Pabst,J. R. Kocks, J. Krege, S. Hardtke, and R. Forster. 2006. Induction of tolerance toinnocuous inhaled antigen relies on a CCR7-dependent dendritic cell-mediatedantigen transport to the bronchial lymph node. J. Immunol. 177: 7346–7354.
4. Reynoso, E. D., J. W. Lee, and S. J. Turley. 2009. Peripheral tolerance inductionby lymph node stroma. In Crossroads between Innate and Adaptive Immunity II.S. P. Schoenberger, P. D. Katsikis, and B. Pulendran, eds. Springer, New York, p.113–127.
5. Baird, J. B., J. L. Charles, T. G. Streit, J. M. Roberts, D. G. Addiss, andP. J. Lammie. 2002. Reactivity to bacterial, fungal, and parasite antigens inpatients with lymphedema and elephantiasis. Am. J. Trop. Med. Hyg. 66: 163–169.
6. von der Weid, P. Y., S. Rehal, and J. G. P. Ferraz. 2011. Role of the lymphaticsystem in the pathogenesis of Crohn’s disease. Curr. Opin. Gastroenterol. 27:335–341.
7. Angeli, V., F. Ginhoux, J. Llodra, L. Quemeneur, P. S. Frenette, M. Skobe,R. Jessberger, M. Merad, and G. J. Randolph. 2006. B cell-driven lym-phangiogenesis in inflamed lymph nodes enhances dendritic cell mobilization.Immunity 24: 203–215.
8. Halin, C., N. E. Tobler, B. Vigl, L. F. Brown, and M. Detmar. 2007. VEGF-Aproduced by chronically inflamed tissue induces lymphangiogenesis in draininglymph nodes. Blood 110: 3158–3167.
9. Yin, N., N. Zhang, J. N. Xu, Q. X. Shi, Y. Z. Ding, and J. S. Bromberg. 2011.Targeting lymphangiogenesis after islet transplantation prolongs islet allograftsurvival. Transplantation 92: 25–30.
10. Kerjaschki, D., H. M. Regele, I. Moosberger, K. Nagy-Bojarski, B. Watschinger,A. Soleiman, P. Birner, S. Krieger, A. Hovorka, G. Silberhumer, et al. 2004.Lymphatic neoangiogenesis in human kidney transplants is associated withimmunologically active lymphocytic infiltrates. J. Am. Soc. Nephrol. 15: 603–612.
11. Ling, S., C. Qi, W. Li, J. Xu, and W. Kuang. 2009. Crucial role of corneallymphangiogenesis for allograft rejection in alkali-burned cornea bed. Clin.Experiment. Ophthalmol. 37: 874–883.
12. Dietrich, T., F. Bock, D. Yuen, D. Hos, B. O. Bachmann, G. Zahn, S. Wiegand,L. Chen, and C. Cursiefen. 2010. Cutting edge: lymphatic vessels, not bloodvessels, primarily mediate immune rejections after transplantation. J. Immunol.184: 535–539.
13. Hoffmann, F., E. P. Zhang, A. Mueller, F. Schulte, H. D. Foss, J. Franke, andS. E. Coupland. 2001. Contribution of lymphatic drainage system in cornealallograft rejection in mice. Graefes Arch. Clin. Exp. Ophthalmol. 239: 850–858.
14. Kerjaschki, D. 2006. Lymphatic neoangiogenesis in renal transplants: a drivingforce of chronic rejection? J. Nephrol. 19: 403–406.
The Journal of Immunology 2189
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

15. Huggenberger, R., S. S. Siddiqui, D. Brander, S. Ullmann, K. Zimmermann,M. Antsiferova, S. Werner, K. Alitalo, and M. Detmar. 2011. An important roleof lymphatic vessel activation in limiting acute inflammation. Blood 117: 4667–4678.
16. Stuht, S., W. Gwinner, I. Franz, A. Schwarz, D. Jonigk, H. Kreipe, D. Kerjaschki,H. Haller, and M. Mengel. 2007. Lymphatic neoangiogenesis in human renalallografts: results from sequential protocol biopsies. Am. J. Transplant. 7: 377–384.
17. Harrell, M. I., B. M. Iritani, and A. Ruddell. 2007. Tumor-induced sentinellymph node lymphangiogenesis and increased lymph flow precede melanomametastasis. Am. J. Pathol. 170: 774–786.
18. Ruddell, A., M. I. Harrell, and M. Furuya. 2009. Sentinel lymph node lym-phangiogenesis and increased lymph flow in murine tumor metastasis. Clin. Exp.Metastasis 26: 883–884.
19. Lund, A. W., F. V. Duraes, S. H. Hirosue, V. R. Raghavan, C. Nembrini,S. N. Thomas, A. Issa, S. Hugues, and M. A. Swartz. 2012. VEGF-C promotesimmune tolerance in B16 melanomas and cross-presentation of tumor antigen bylymph node lymphatics. Cell Reports 1: 191–199.
20. Swartz, M. A., and A. W. Lund. 2012. Lymphatic and interstitial flow in thetumour microenvironment: linking mechanobiology with immunity. Nat. Rev.Cancer 12: 210–219.
21. Randolph, G. J., V. Angeli, and M. A. Swartz. 2005. Dendritic-cell trafficking tolymph nodes through lymphatic vessels. Nat. Rev. Immunol. 5: 617–628.
22. Junt, T., E. A. Moseman, M. Iannacone, S. Massberg, P. A. Lang, M. Boes,K. Fink, S. E. Henrickson, D. M. Shayakhmetov, N. C. Di Paolo, et al. 2007.Subcapsular sinus macrophages in lymph nodes clear lymph-borne viruses andpresent them to antiviral B cells. Nature 450: 110–114.
23. Roozendaal, R., T. R. Mempel, L. A. Pitcher, S. F. Gonzalez, A. Verschoor,R. E. Mebius, U. H. von Andrian, and M. C. Carroll. 2009. Conduits mediatetransport of low-molecular-weight antigen to lymph node follicles. Immunity 30:264–276.
24. Itano, A. A., and M. K. Jenkins. 2003. Antigen presentation to naive CD4 T cellsin the lymph node. Nat. Immunol. 4: 733–739.
25. Forster, R., A. C. Davalos-Misslitz, and A. Rot. 2008. CCR7 and its ligands:balancing immunity and tolerance. Nat. Rev. Immunol. 8: 362–371.
26. Mueller, S. N., and R. N. Germain. 2009. Stromal cell contributions to the ho-meostasis and functionality of the immune system. Nat. Rev. Immunol. 9: 618–629.
27. Drayton, D. L., S. Liao, R. H. Mounzer, and N. H. Ruddle. 2006. Lymphoidorgan development: from ontogeny to neogenesis. Nat. Immunol. 7: 344–353.
28. Mori, S., H. Nakano, K. Aritomi, C. R. Wang, M. D. Gunn, and T. Kakiuchi.2001. Mice lacking expression of the chemokines CCL21-ser and CCL19 (pltmice) demonstrate delayed but enhanced T cell immune responses. J. Exp. Med.193: 207–218.
29. Davalos-Misslitz, A. C., J. Rieckenberg, S. Willenzon, T. Worbs, E. Kremmer,G. Bernhardt, and R. Forster. 2007. Generalized multi-organ autoimmunity inCCR7-deficient mice. Eur. J. Immunol. 37: 613–622.
30. Nitta, T., S. Nitta, Y. Lei, M. Lipp, and Y. Takahama. 2009. CCR7-mediatedmigration of developing thymocytes to the medulla is essential for negativeselection to tissue-restricted antigens. Proc. Natl. Acad. Sci. USA 106: 17129–17133.
31. Schneider, M. A., J. G. Meingassner, M. Lipp, H. D. Moore, and A. Rot. 2007.CCR7 is required for the in vivo function of CD4+CD25+ regulatory T cells. J.Exp. Med. 204: 735–745.
32. Menning, A., U. E. Hopken, K. Siegmund, M. Lipp, A. Hamann, and J. Huehn.2007. Distinctive role of CCR7 in migration and functional activity of naive- andeffector/memory-like Treg subsets. Eur. J. Immunol. 37: 1575–1583.
33. Ochando, J. C., A. C. Yopp, Y. Yang, A. Garin, Y. Li, P. Boros, J. Llodra,Y. Ding, S. A. Lira, N. R. Krieger, and J. S. Bromberg. 2005. Lymph node oc-cupancy is required for the peripheral development of alloantigen-specificFoxp3+ regulatory T cells. J. Immunol. 174: 6993–7005.
34. Lee, J. W., M. Epardaud, J. Sun, J. E. Becker, A. C. Cheng, A. R. Yonekura,J. K. Heath, and S. J. Turley. 2007. Peripheral antigen display by lymph nodestroma promotes T cell tolerance to intestinal self. Nat. Immunol. 8: 181–190.
35. Cohen, J. N., C. J. Guidi, E. F. Tewalt, H. Qiao, S. J. Rouhani, A. Ruddell,A. G. Farr, K. S. Tung, and V. H. Engelhard. 2010. Lymph node-resident lym-phatic endothelial cells mediate peripheral tolerance via Aire-independent directantigen presentation. J. Exp. Med. 207: 681–688.
36. Makinen, T., L. Jussila, T. Veikkola, T. Karpanen, M. I. Kettunen,K. J. Pulkkanen, R. Kauppinen, D. G. Jackson, H. Kubo, S. Nishikawa, et al.2001. Inhibition of lymphangiogenesis with resulting lymphedema in transgenicmice expressing soluble VEGF receptor-3. Nat. Med. 7: 199–205.
37. Rutkowski, J. M., C. E. Markhus, C. C. Gyenge, K. Alitalo, H. Wiig, andM. A. Swartz. 2010. Dermal collagen and lipid deposition correlate with tissueswelling and hydraulic conductivity in murine primary lymphedema. Am. J.Pathol. 176: 1122–1129.
38. Swartz, M. A., A. Kaipainen, P. A. Netti, C. Brekken, Y. Boucher,A. J. Grodzinsky, and R. K. Jain. 1999. Mechanics of interstitial-lymphatic fluidtransport: theoretical foundation and experimental validation. J. Biomech. 32:1297–1307.
39. Lammermann, T., B. L. Bader, S. J. Monkley, T. Worbs, R. Wedlich-Soldner,K. Hirsch, M. Keller, R. Forster, D. R. Critchley, R. Fassler, and M. Sixt. 2008.
Rapid leukocyte migration by integrin-independent flowing and squeezing.Nature 453: 51–55.
40. Swartz, M. A., D. A. Berk, and R. K. Jain. 1996. Transport in lymphaticcapillaries. I. Macroscopic measurements using residence time distributiontheory. Am. J. Physiol. 270: H324–H329.
41. Bajenoff, M., J. G. Egen, L. Y. Koo, J. P. Laugier, F. Brau, N. Glaichenhaus, andR. N. Germain. 2006. Stromal cell networks regulate lymphocyte entry, migra-tion, and territoriality in lymph nodes. Immunity 25: 989–1001.
42. Duncan, B., C. Nazarov-Stoica, J. Surls, M. Kehl, C. Bona, S. Casares, andT. D. Brumeanu. 2010. Double negative (CD3+4‑8‑) TCR ab splenic cells fromyoung NOD mice provide long-lasting protection against type 1 diabetes. PLoSOne 5: e11427.
43. Saito, E., M. Fujimoto, M. Hasegawa, K. Komura, Y. Hamaguchi, Y. Kaburagi,T. Nagaoka, K. Takehara, T. F. Tedder, and S. Sato. 2002. CD19-dependentB lymphocyte signaling thresholds influence skin fibrosis and autoimmunity inthe tight-skin mouse. J. Clin. Invest. 109: 1453–1462.
44. Sato, S., M. Hasegawa, M. Fujimoto, T. F. Tedder, and K. Takehara. 2000.Quantitative genetic variation in CD19 expression correlates with autoimmunity.J. Immunol. 165: 6635–6643.
45. Akiba, H., J. Kehren, M. T. Ducluzeau, M. Krasteva, F. Horand, D. Kaiserlian,F. Kaneko, and J. F. Nicolas. 2002. Skin inflammation during contact hyper-sensitivity is mediated by early recruitment of CD8+ T cytotoxic 1 cells inducingkeratinocyte apoptosis. J. Immunol. 168: 3079–3087.
46. Wang, B., H. Fujisawa, L. Zhuang, I. Freed, B. G. Howell, S. Shahid,G. M. Shivji, T. W. Mak, and D. N. Sauder. 2000. CD4+ Th1 and CD8+ type 1cytotoxic T cells both play a crucial role in the full development of contacthypersensitivity. J. Immunol. 165: 6783–6790.
47. Chu, V. T., A. Beller, T. T. Nguyen, G. Steinhauser, and C. Berek. 2011. Thelong-term survival of plasma cells. Scand. J. Immunol. 73: 508–511.
48. Hofmann, J., M. Greter, L. Du Pasquier, and B. Becher. 2010. B-cells need aproper house, whereas T-cells are happy in a cave: the dependence of lymphocyteson secondary lymphoid tissues during evolution. Trends Immunol. 31: 144–153.
49. Schroder, A. E., A. Greiner, C. Seyfert, and C. Berek. 1996. Differentiation ofB cells in the nonlymphoid tissue of the synovial membrane of patients withrheumatoid arthritis. Proc. Natl. Acad. Sci. USA 93: 221–225.
50. Stott, D. I., F. Hiepe, M. Hummel, G. Steinhauser, and C. Berek. 1998. Antigen-driven clonal proliferation of B cells within the target tissue of an autoimmunedisease: the salivary glands of patients with Sjogren’s syndrome. J. Clin. Invest.102: 938–946.
51. MacLennan, I. C. 1994. Germinal centers. Annu. Rev. Immunol. 12: 117–139.52. Sugaya, M., Y. Kuwano, H. Suga, T. Miyagaki, H. Ohmatsu, T. Kadono,
H. Okochi, A. Blauvelt, K. Tamaki, and S. Sato. 2012. Lymphatic dysfunctionimpairs antigen-specific immunization, but augments tissue swelling followingcontact with allergens. J. Invest. Dermatol. 132: 667–676.
53. Sugaya, M., T. Watanabe, A. Yang, M. F. Starost, H. Kobayashi, A. M. Atkins,D. L. Borris, E. A. Hanan, D. Schimel, M. A. Bryant, et al. 2005. Lymphaticdysfunction in transgenic mice expressing KSHV k-cyclin under the control ofthe VEGFR-3 promoter. Blood 105: 2356–2363.
54. Greter, M., J. Hofmann, and B. Becher. 2009. Neo-lymphoid aggregates in theadult liver can initiate potent cell-mediated immunity. PLoS Biol. 7: e1000109.
55. Chen, L., P. Hamrah, C. Cursiefen, Q. Zhang, B. Pytowski, J. W. Streilein, andM. R. Dana. 2004. Vascular endothelial growth factor receptor-3 mediates in-duction of corneal alloimmunity. Nat. Med. 10: 813–815.
56. Hamrah, P., L. Chen, Q. Zhang, and M. R. Dana. 2003. Novel expression ofvascular endothelial growth factor receptor (VEGFR)-3 and VEGF-C on cornealdendritic cells. Am. J. Pathol. 163: 57–68.
57. Tsuji, R. F., M. Szczepanik, I. Kawikova, V. Paliwal, R. A. Campos, A. Itakura,M. Akahira-Azuma, N. Baumgarth, L. A. Herzenberg, and P. W. Askenase. 2002.B cell-dependent T cell responses: IgM antibodies are required to elicit contactsensitivity. J. Exp. Med. 196: 1277–1290.
58. Watanabe, R., M. Fujimoto, N. Ishiura, Y. Kuwano, H. Nakashima, N. Yazawa,H. Okochi, S. Sato, T. F. Tedder, and K. Tamaki. 2007. CD19 expression inB cells is important for suppression of contact hypersensitivity. Am. J. Pathol.171: 560–570.
59. Mebius, R. E., P. R. Streeter, J. Breve, A. M. Duijvestijn, and G. Kraal. 1991.The influence of afferent lymphatic vessel interruption on vascular addressinexpression. J. Cell Biol. 115: 85–95.
60. Tomei, A. A., S. Siegert, M. R. Britschgi, S. A. Luther, and M. A. Swartz. 2009.Fluid flow regulates stromal cell organization and CCL21 expression in a tissue-engineered lymph node microenvironment. J. Immunol. 183: 4273–4283.
61. Wengner, A. M., U. E. Hopken, P. K. Petrow, S. Hartmann, U. Schurigt,R. Brauer, and M. Lipp. 2007. CXCR5- and CCR7-dependent lymphoid neo-genesis in a murine model of chronic antigen-induced arthritis. Arthritis Rheum.56: 3271–3283.
62. Tomura, M., T. Honda, H. Tanizaki, A. Otsuka, G. Egawa, Y. Tokura,H. Waldmann, S. Hori, J. G. Cyster, T. Watanabe, et al. 2010. Activated regu-latory T cells are the major T cell type emigrating from the skin during a cuta-neous immune response in mice. J. Clin. Invest. 120: 883–893.
63. Kodera, T., T. L. McGaha, R. Phelps, W. E. Paul, and C. A. Bona. 2002. Dis-rupting the IL-4 gene rescues mice homozygous for the tight-skin mutation fromembryonic death and diminishes TGF-b production by fibroblasts. Proc. Natl.Acad. Sci. USA 99: 3800–3805.
2190 LYMPHATIC DRAINAGE IN REGULATING IMMUNE RESPONSE
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from


















