Liver-Specific miR-122 does Not Affect mRNA Levels of ... life span and experience a drastic decline...
8
ALTEX Proceedings, 1/12, Proceedings of WC8 165 1 Introduction Due to growing ethical concerns regarding animal testing in the EU and the implementation of new legislation, including the European Cosmetic Directive (2003/15/EC) and the REACH Regulation (EC No 1907/2006, Registration, Evaluation, Au- thorisation and restriction of CHemical substances), alternative testing methods, complying with the 3Rs concept of Russell and Burch (Refinement, Reduction and Replacement of ani- mal experimentation) (Russell and Burch, 1959), are becom- ing increasingly important (Vanhaecke et al., 2009; Adler et al., 2011). Since biotransformation of xenobiotics, such as pharma- ceuticals, mainly occurs in the liver and, more particularly in hepatocytes, hepatic in vitro models usually represent appropri- ate alternatives for pharmaco-toxicological testing. Primary hepatocyte cultures, considered the gold standard in the field of liver-based in vitro modelling, however, have a lim- ited life span and experience a drastic decline of liver-specific functionality, including biotransformation capacity (Vanhaecke and Rogiers, 2006; Elaut et al., 2006; Guguen-Guillouzo and Guillouzo, 2010). Therefore, the development of a phenotypi- cally stable primary hepatocyte model that will allow extrapo- lation of generated in vitro data to the in vivo situation is still considered an absolute priority, especially to meet the novel legislative changes (Paine and Andreakos, 2004; Adler et al., 2011; Vanhaecke et al., 2011). In this regard, different strategies have been introduced, including the re-establishment of the liver micro-environment in vitro, by adding soluble differentiation- promoting factors and restoring cell-matrix and cell-cell inter- actions, all with rather limited success (Vanhaecke and Rogiers, 2006; Elaut et al.,2006). Our group explores a more site-directed, anti-dedifferentiation approach by intervening at the epigenetic level through expo- sure of primary hepatocytes to epigenetic modifiers, such as his- tone deacetylase (HDAC) and DNA methyltransferase (DNMT) inhibitors or a combination of these. Thus far, we found that such a chromatin remodelling strategy significantly counteracts dedifferentiation in primary hepatocyte cultures, as evidenced by delayed cell death and enhanced liver-specific functionality (Fraczek et al., 2011; Henkens et al., 2007, 2011; Patent submis- sions WO2006/045331 1 and WO2008/145736 2 ). In the current study, we focused on another member of the epigenetic machinery while further optimizing our hepatocyte stabilization methodology, namely microRNA-related control. MicroRNAs are short (~22 nt) non-coding RNAs that post- transcriptionally modulate gene expression by mRNA cleav- Liver-Specific miR-122 does Not Affect mRNA Levels of Cytochrome P450 Enzymes and Nuclear Receptors in Primary Rat Hepatocyte Cultures Jennifer Bolleyn 1 , Joery De Kock 1 , Hélène Mertens 1 , Joanna Fraczek 1 , Mathieu Vinken 1 , Olivier Govaere 2 , Tania Roskams 2 , Vera Rogiers 1 , and Tamara Vanhaecke 1 1 Department of Toxicology, Center for Pharmaceutical Research, Vrije Universiteit Brussel (VUB), Brussels, Belgium; 2 Department of Pathology, Laboratory for Morphology and Molecular Pathology, Catholic University Leuven (KUL), Leuven, Belgium Summary Primary hepatocyte cultures suffer from the loss of xenobiotic biotransformation capacity as a function of culture time. Over the years, different strategies have been developed to phenotypically stabilize these cultures, but with limited success. In the present study, we evaluate the modulation of microRNA (miRNA, miR), which are major determinants of gene expression at the post-transcriptional level, as a novel stabilization approach in this respect. miR-122, an acknowledged liver-specific miRNA species, was either overexpressed or inhibited, and the outcome on mRNA levels of cytochrome P450 enzymes and nuclear receptors was investigated. However, no persistent effects could be observed, suggesting that miRNA species other than miR-122 should be tested for their potential to serve as a target for innovative stabilization efforts in cultures of primary hepatocytes. Keywords: microRNA, miR-122, primary hepatocytes, mimics, inhibitors 1 Patent submission WO2006/045331. Differentiation of stem cells and stabilization of phenotypical properties of primary cells. Applicant: VUB – l Inventors: Rogiers, V., Snykers, S., Papeleu, P., Vinken, M., Henkens, T., Elaut, G., Vanhaecke, T. 2 Patent submission WO2008/145736. Stabilisation of the phenotypic properties of isolated primary cells. Applicant: VUB – Inventors: Rogiers, V., Vanhaecke, T., De Rop, E., Fraczek, J.
Transcript of Liver-Specific miR-122 does Not Affect mRNA Levels of ... life span and experience a drastic decline...

Altex Proceedings, 1/12, Proceedings of WC8 165
1 Introduction
Due to growing ethical concerns regarding animal testing in the eU and the implementation of new legislation, including the european Cosmetic Directive (2003/15/eC) and the ReACH Regulation (eC No 1907/2006, Registration, evaluation, Au-thorisation and restriction of CHemical substances), alternative testing methods, complying with the 3Rs concept of Russell and Burch (Refinement, Reduction and Replacement of ani-mal experimentation) (Russell and Burch, 1959), are becom-ing increasingly important (Vanhaecke et al., 2009; Adler et al., 2011). Since biotransformation of xenobiotics, such as pharma-ceuticals, mainly occurs in the liver and, more particularly in hepatocytes, hepatic in vitro models usually represent appropri-ate alternatives for pharmaco-toxicological testing.
Primary hepatocyte cultures, considered the gold standard in the field of liver-based in vitro modelling, however, have a lim-ited life span and experience a drastic decline of liver-specific functionality, including biotransformation capacity (Vanhaecke and Rogiers, 2006; elaut et al., 2006; Guguen-Guillouzo and Guillouzo, 2010). therefore, the development of a phenotypi-cally stable primary hepatocyte model that will allow extrapo-lation of generated in vitro data to the in vivo situation is still
considered an absolute priority, especially to meet the novel legislative changes (Paine and Andreakos, 2004; Adler et al., 2011; Vanhaecke et al., 2011). In this regard, different strategies have been introduced, including the re-establishment of the liver micro-environment in vitro, by adding soluble differentiation-promoting factors and restoring cell-matrix and cell-cell inter-actions, all with rather limited success (Vanhaecke and Rogiers, 2006; elaut et al.,2006).
Our group explores a more site-directed, anti-dedifferentiation approach by intervening at the epigenetic level through expo-sure of primary hepatocytes to epigenetic modifiers, such as his-tone deacetylase (HDAC) and DNA methyltransferase (DNMt) inhibitors or a combination of these. thus far, we found that such a chromatin remodelling strategy significantly counteracts dedifferentiation in primary hepatocyte cultures, as evidenced by delayed cell death and enhanced liver-specific functionality (Fraczek et al., 2011; Henkens et al., 2007, 2011; Patent submis-sions WO2006/0453311 and WO2008/1457362).
In the current study, we focused on another member of the epigenetic machinery while further optimizing our hepatocyte stabilization methodology, namely microRNA-related control. MicroRNAs are short (~22 nt) non-coding RNAs that post-transcriptionally modulate gene expression by mRNA cleav-
Liver-Specific miR-122 does Not Affect mRNA Levels of Cytochrome P450 Enzymes and Nuclear Receptors in Primary Rat Hepatocyte Cultures Jennifer Bolleyn 1, Joery De Kock 1, Hélène Mertens 1, Joanna Fraczek 1, Mathieu Vinken 1, Olivier Govaere 2, Tania Roskams 2, Vera Rogiers 1, and Tamara Vanhaecke 1
1Department of toxicology, Center for Pharmaceutical Research, Vrije Universiteit Brussel (VUB), Brussels, Belgium; 2Department of Pathology, laboratory for Morphology and Molecular Pathology, Catholic University leuven (KUl), leuven, Belgium
SummaryPrimary hepatocyte cultures suffer from the loss of xenobiotic biotransformation capacity as a function of culture time. Over the years, different strategies have been developed to phenotypically stabilize these cultures, but with limited success. In the present study, we evaluate the modulation of microRNA (miRNA, miR), which are major determinants of gene expression at the post-transcriptional level, as a novel stabilization approach in this respect. miR-122, an acknowledged liver-specific miRNA species, was either overexpressed or inhibited, and the outcome on mRNA levels of cytochrome P450 enzymes and nuclear receptors was investigated. However, no persistent effects could be observed, suggesting that miRNA species other than miR-122 should be tested for their potential to serve as a target for innovative stabilization efforts in cultures of primary hepatocytes.
Keywords: microRNA, miR-122, primary hepatocytes, mimics, inhibitors
1 Patent submission WO2006/045331. Differentiation of stem cells and stabilization of phenotypical properties of primary cells. Applicant: VUB – l Inventors: Rogiers, V., Snykers, S., Papeleu, P., Vinken, M., Henkens, T., Elaut, G., Vanhaecke, T. 2 Patent submission WO2008/145736. Stabilisation of the phenotypic properties of isolated primary cells. Applicant: VUB – Inventors: Rogiers, V., Vanhaecke, T., De Rop, E., Fraczek, J.

Bolleyn et al.
Altex Proceedings, 1/12, Proceedings of WC8166
age or translational inhibition, depending on complementary binding to 3’-untranslated regions (3’-UtR) of target mRNAs (Ambros, 2004). Here we investigated whether modulation of the expression of miR-122, a well-known liver-specific miR-NA (Chang et al., 2004; Coulouarn et al., 2009, Filipowics and Grosshans, 2011), in primary hepatocyte cultures affects mR-NA levels of nuclear receptors known to drive liver-specific gene expression and cytochrome P450 (CYP) enzymes, the most important phase I xenobiotic biotransformation enzymes in hepatocytes.
2 Animals, materials and methods
Chemicals and reagentsl-glutamine, bovine insulin, and glucagon were purchased from Sigma-Aldrich (Belgium). Williams’ e medium and foetal bo-vine serum (FBS) were obtained from Gibco BRl (Belgium). All other chemicals and reagents were commercial products of the highest analytical grade available.
Isolation and cultivation of primary rat hepatocytesAll experiments were performed in accordance with the regu-lations of the local ethical committee of the Vrije Universiteit Brussel. Hepatocytes were isolated as previously described (Papeleu et al., 2005) by use of a two-step collagenase method from male outbred Sprague-Dawley OFA rats (200-300 g) pur-chased from Charles River laboratories (Belgium). After test-ing cell viability (≥80%) by trypan blue exclusion, the hepa-tocytes were seeded as a monolayer on a plastic culture dish (Henkens et al., 2005) in Williams’ e medium supplemented with 10% (v/v) FBS, 2 mM L-glutamine, 7 ng/ml glucagon, 7.3 IU/ml benzyl penicillin, 50 µg/ml streptomycin sulfate, 50 µg/ml kanamycin monosulfate and 10 µg/ml sodium ampi-cillin. Cell cultures were placed at 37°C in an atmosphere of 5% CO2 and 95% air at 100% relative humidity. After 4 h, the medium was renewed with serum-free medium supplemented with 25 µg/ml hydrocortisone sodium succinate and 0.5 µg/ml bovine insulin. twenty-four hours after seeding, the medium was renewed with serum-free culture medium. this was re-peated daily. From daily renewal of the medium (i.e., after 24 h cultivation) onwards, experimental conditions were as follows for 48 h: (i) untreated cells, (ii) cultures transfected with a non-cytotoxic concentration of 100 nM miR-122 mimic, and (iii) a non-cytotoxic concentration of 5 nM miR-122 inhibitor. After 48 h of transfection, the medium was renewed daily with serum-free culture medium. Sampling was performed at 24, 48, and 96 h after cessation of transfection.
Cell transfectiontransfections were carried out using DharmaFeCt 1 (Dhar-macon, USA) according to the manufacturer’s instructions. miRIDIAN miRNA Hairpin Inhibitors and Mimics from Dhar-macon (USA) were used for the specific inhibition and overex-pression of miR-122. the amount of small RNA was adjusted
to a final concentration of 5 nM hairpin inhibitor and 100 nM mimic, respectively. Cell viability tests (lactate dehydrogenase leakage) revealed that these concentrations were not cytotoxic (data not shown).
Cell morphologyCell morphology was analyzed by light microscopy using a Nikon eclipse ti phase contrast microscope (Nikon, Belgium) with a 100x magnification.
Expression of miR-122a)Isolation of small RNAFreshly isolated hepatocytes were directly pelleted, whereas cultured cells were harvested by scraping, washed twice with ice-cold phosphate buffered saline (PBS) and pelleted thereafter. Small RNA (≤200nt) was isolated using the mirVana™miRNA isolation kit (Ambion, Inc., USA) according to the manufac-turer’s instructions in an RNAse-free environment. Quantifica-tion of RNA in the samples was performed using a Nanodrop ND-1000 Spectrophotometer (NanoDrop technologies, USA).
b)Quantitative RT-PCR (qPCR) of miR-122After RNA quantification, copyDNA (cDNA) was synthetized from the isolated small RNA using a taqMan® MicroRNA Reverse transcription Kit (Applied Biosystems, Belgium) and a taqMan® MicroRNA Assay containing specific primers for miR-122 (assay ID: 002245) and U6 snRNA (assay ID: 001973) (Applied Biosystems, Halle, Belgium). the Rt-reaction mix-ture consisted of 0.15 µl 100 mM dNtPs (with dttP), 1 µl MultiScribe™ Reverse Transcriptase (50 U/µl), 1.5 µl 10x Re-verse transcription Buffer, 0.19 µl RNase Inhibitor (20 U/µl), 4.16 µl nuclease-free water, 5 µl total RNA and 3 µl of a miRNA specific RT-primer in a final volume of 15 µl. Reactions were incubated on a Biorad iCycler at 16°C for 30 min, 42°C for 30 min and 85°C for 5 min. Following the Rt step, PCR am-plification was performed by combining 2 µl of cDNA and 18 µl mastermix containing 7 µl nuclease-free water, 10 µl taq-Man® Universal PCR Master Mix, No Amperase® UNG and 1 µl of 20x taqman MicroRNA Assay. PCR conditions com-prised 10 min of enzyme activation at 95°C, followed by 40 cycles of 15 sec at 95°C and 60 sec at 60°C performed on a Bio-Rad iCycler iQ5 Multicolor Real-time PCR (Bio-Rad, Belgium). U6 snRNA was used to normalize values and relative changes in miRNA expression were calculated using the 2- ΔΔCt method (Schmittgen and livak, 2008).
Expression of cytochrome P450 enzymes and nuclear receptorsa)Isolation of mRNAFreshly isolated hepatocytes were directly pelleted, whereas cultured cells were harvested by scraping, washed twice with PBS, and pelleted thereafter. total RNA was isolated using tRIzol reagent (Invitrogen, Belgium) according to the manu-facturer’s instructions in a nuclease-free environment with the following modification: total RNAs were precipitated with isopropanol at 4°C overnight for better precipitation of small

Bolleyn et al.
Altex Proceedings, 1/12, Proceedings of WC8 167
RNAs. Quantification of RNA in the samples was performed using a Nanodrop ND-1000 Spectrophotometer (NanoDrop technologies, USA).
b)Quantitative RT-PCR (qPCR) Total RNA was reverse transcribed into cDNA using iScript™ cDNA Synthesis Kit (BioRad, Belgium). the Rt-reaction con-sisted of 8 µl 5x iScript™ reaction mix, 2 µl iScript™ Reverse transcriptase and 30 µl RNA mixture, containing 2 µg RNA diluted in nuclease-free water. Reactions were incubated on a Biorad iCycler at 25°C for 5 min, 42°C for 30 min and 85°C for 5 min. Following the RT step, cDNA purification was per-formed with the Genelute PCR clean up kit (Sigma-Aldrich, Belgium) according to the manufacturer’s instructions. cDNA products were used for quantitative amplification of the target genes. the primers used in this study are listed in table 1. All samples were run in duplicate and each run included two nega-tive controls (NtC) and a serial dilution of a pooled cDNA mix from all samples to calculate the efficiency curve. The qPCR reaction mix consisted of 12.5 μl TaqMan Universal Master Mix (Applied Biosystems, Belgium), 1.25 μl 20x Assay-on-Demand Mix (Applied Biosystems, Belgium) and 2 μl of cDNA in a 25 μl volume adjusted with nuclease-free water. qPCR condi-tions were as follows: enzyme activation for 2 min at 50°C, incubation for 10 min at 95°C, followed by 40 cycles of 15 sec denaturation at 95°C, annealing for 1 min at 60°C, performed on a Bio-Rad iCycler iQ5 Multicolor Real-time PCR (BioRad, Belgium).
c)Selection of stable reference genesAssessing gene expression stability of the selected reference genes (Ubiquitin C (UBC), beta-2-microglobulin (B2M), glycer-aldehyde 3-phosphate dehydrogenase (GADPH), hydroxymeth-ylbilane synthase (HMBS), eukaryotic 18S rRNA (18S), and actin β (ACTB)) was done using three different algorithms: geNorm, Normfinder, and Bestkeeper (Ceelen et al., 2011). geNorm, a visual basic application for Microsoft excel, calculates a gene expression stability measure M for each reference gene. Step-wise exclusion of the gene with the highest M value results in a combination of two constitutively expressed reference genes that have the most stable expression in the tested samples. Selection of the optimal number of stable reference genes was based on geNorm’s pairwise variation analysis between subsequent nor-malization factors using an approximate cut-off value of 0.15 for the inclusion of additional reference genes (Vandesompele et al., 2002). Normfinder, also a visual basic applet, identifies the most stable reference gene among a set of reference genes by assigning a stability value (Andersen et al., 2004). Finally, Bestkeeper, another excel-based application, determines the variability in expression of a set of reference genes by analyzing quantification cycle (Cq) values and classifying variability by the coefficient of variance (CV) and the standard deviation (SD). To define the most stable reference gene, the software generates a BestKeeper index which finally is compared to each candidate gene, resulting in a value for the Pearson correlation coefficient (r) and probability (p), which are then allocated to each candi-date reference gene (Pfaffl et al., 2004).
Tab. 1: Primers used for the evaluation of the gene expression of cytochrome P450 enzymes and nuclear receptorsAbbreviations: 18S: Eukaryotic 18S rRNA; AB: Applied Biosystems; ACTB: beta-actin; AhR: aryl hydrocarbon receptor; B2M: beta-2-microglobulin; CaR: constitutive androstane receptor; CYP: cytochrome P450; GAPDH: glyceraldehyde 3-phosphate dehydro-genase; HMBS: hydroxymethylbilane synthase; Nr: Nuclear hormone receptor; PxR: pregnane X receptor; UBC: ubiquitin C
Gene Assay-on-Demand ID Amplicon length (bp) Source
GADPH Rn01775763_g1 174 AB
B2M Rn00560865_m1 58 AB
UBC Rn01789812_g1 88 AB
ACTB Rn00667869_m1 91 AB
18S Hs99999901_s1 187 AB
HMBS Rn00565886_m1 99 AB
CYP1A1 Rn00487218_m1 120 AB
CYP1A2 Rn01422531_m1 136 AB
CYP2B1 Rn01457875_m1 89 AB
CYP2B2 Rn02786833_m1 119 AB
CYP3A2 Rn00756461_m1 116 AB
AhR Rn00682057_m1 67 AB
Nr1i2/PxR Rn00583887_m1 59 AB
Nr1i3/CaR Rn00576085_m1 63 AB
Bolleyn et al.
Altex Proceedings, 1/12, Proceedings of WC8168
Cell morphologyDuring the cultivation period, the morphology of the cultured cells was evaluated by light microscopic analysis. No visual dif-ferences were noticed between the three conditions (Fig. 2). All showed a typical progressive deterioration of their morphologi-cal appearance, evidenced by the manifestations of extensions, a more flattened morphology, and signs of cell death.
qPCR of cytochrome P450 enzymes and nuclear receptorsthe outcome of miR-122 mimic and inhibitor on the mRNA abundance of a number of selected genes, including phase I xe-nobiotic biotransformation enzymes and nuclear receptors, was tested over time. the expression of these genes was normalized using a pool of four reference genes, chosen out of six common reference genes, including UBC, B2M, GADPH, HMBS, 18S, and β-actin. The expression stability of the latter reference genes was first assessed by the use of three software programs. The Bestkeeper analysis showed that none of the reference genes was stably expressed, due to the presence of four outliers. Upon removal of those outliers, UBC was found to be the best refer-ence gene. this result was also obtained using the algorithm Normfinder, after exclusion of the outliers. Through gradual
Statistical analysisthe results are expressed as mean ± standard deviation (SD) of three independent experiments (n=3). Statistical analyses were performed using one-way analysis of variance (ANOVA), fol-lowed by a Student’s t-test. The significance level was set at 0.05.
3 Results
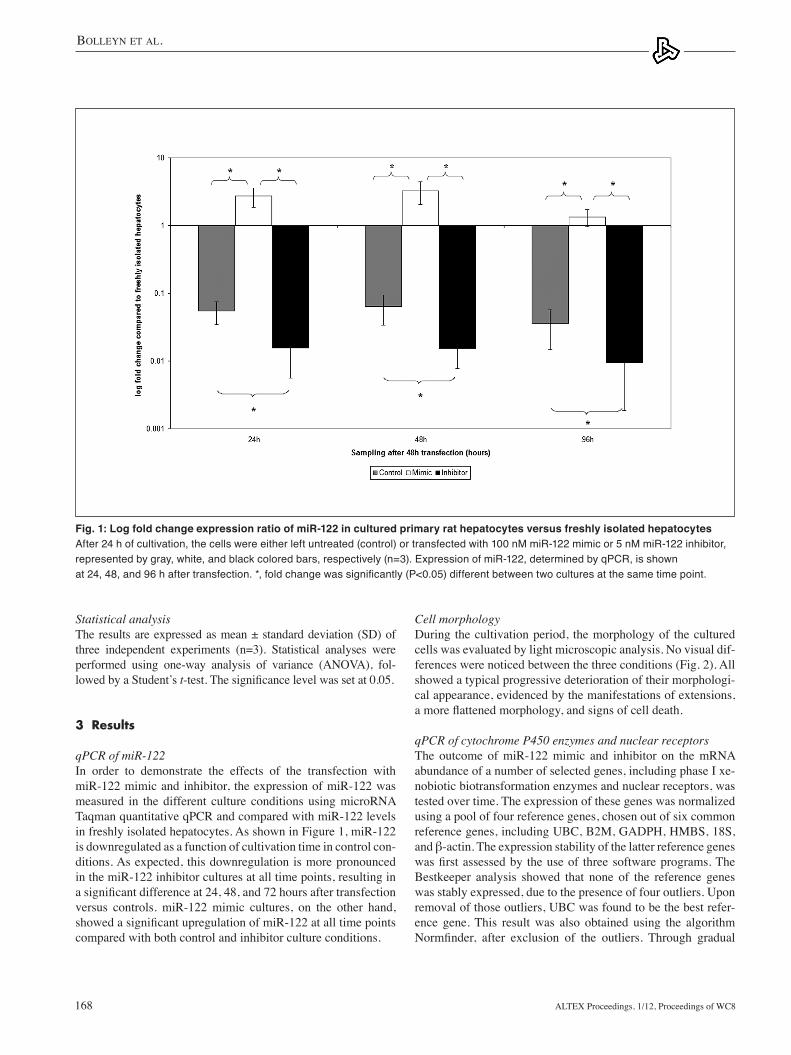
qPCR of miR-122In order to demonstrate the effects of the transfection with miR-122 mimic and inhibitor, the expression of miR-122 was measured in the different culture conditions using microRNA taqman quantitative qPCR and compared with miR-122 levels in freshly isolated hepatocytes. As shown in Figure 1, miR-122 is downregulated as a function of cultivation time in control con-ditions. As expected, this downregulation is more pronounced in the miR-122 inhibitor cultures at all time points, resulting in a significant difference at 24, 48, and 72 hours after transfection versus controls. miR-122 mimic cultures, on the other hand, showed a significant upregulation of miR-122 at all time points compared with both control and inhibitor culture conditions.
Fig. 1: Log fold change expression ratio of miR-122 in cultured primary rat hepatocytes versus freshly isolated hepatocytesAfter 24 h of cultivation, the cells were either left untreated (control) or transfected with 100 nM miR-122 mimic or 5 nM miR-122 inhibitor, represented by gray, white, and black colored bars, respectively (n=3). Expression of miR-122, determined by qPCR, is shown at 24, 48, and 96 h after transfection. *, fold change was significantly (P<0.05) different between two cultures at the same time point.

Bolleyn et al.
Altex Proceedings, 1/12, Proceedings of WC8 169
exclusion of reference genes with the least stable expression, and thus the highest M value, GADPH and HMBS were found to be the most suitable pair of reference genes, using geNorm. Next, selection of the optimal number of reference genes was based on a pairwise variation analysis between subsequent nor-malization factors. Using an approximate cut-off value of 0.15 (Vandesompele et al., 2002), it was shown that, for the current study, four reference genes were sufficient to normalize the ex-amined target genes. table 2 summarizes the four best perform-ing reference genes of each method, from which we selected UBC, GADPH, HMBS, and B2M to normalize the expression of the liver-specific genes. Subsequently, log fold changes were calculated and compared to freshly isolated hepatocytes. As shown in Figure 3, all tested phase I biotransformation genes, except CYP1A1, were downregulated towards the end stages of hepatic cultivation. this downregulation further illustrates the loss of metabolic capacity in primary hepatocyte cultures. Increasing the expression of miR-122 resulted in a significant
Fig. 2: Light microscopic analysis of the morphology of primary rat hepatocytesAfter 24 h of cultivation, the cells were either left untreated (control) or transfected with 100 nM miR-122 mimic or 5 nM miR-122 inhibitor. Pictures are shown 24, 48, and 96 h after transfection. Magnification: 100x.
Tab. 2: Summary and ranking of the four best reference genes for Bestkeeper, Normfinder, and geNormAbbreviations: B2M: beta-2-microglobulin; GAPDH: glyceraldehyde 3-phosphate dehydrogenase; HMBS: hydroxymethylbilane synthase; UBC: ubiquitin C
Ranking Bestkeeper Normfinder geNorm
1 UBC UBC GADPH
2 B2M HMBS HMBS
3 GADPH GADPH UBC
4 HMBS B2M Actin β

Bolleyn et al.
Altex Proceedings, 1/12, Proceedings of WC8170
Fig. 3: Regulation of phase I biotransformation genesRight panel: Log fold change expression ratio of (a) CYP1A1, (b) CYP1A2, (c) CYP2B1, (d) CYP2B2 in cultured primary rat hepatocytes versus freshly isolated hepatocytes. Left panel: Log fold change expression ratio of (e) CYP3A2, (f) AhR, (g) CaR, and (h) PxR in primary rat hepatocytes versus freshly isolated hepatocytes. After 24 h of cultivation, the cells were either left untreated (control) or transfected with 100 nM miR-122 mimic or 5 nM miR-122 inhibitor, represented by gray, white, and black colored bars, respectively (n=3). Expression of cytochrome P450 and nuclear receptors, determined by qPCR, is shown 24, 48, and 96 h after transfection *, fold change was significantly (P<0.05) different between two cultures at the same time point. Abbreviations: AhR: aryl hydrocarbon receptor; CaR: constitutive androstane receptor; CYP: cytochrome P450; PxR: pregnane X receptor; UBC: ubiquitin C

Bolleyn et al.
Altex Proceedings, 1/12, Proceedings of WC8 171
known role in other hepatic metabolic functions such as lipid metabolism (esau et al., 2006; Moore et al., 2011). either no full complementarity to the 3’UtR regions of the investigated CYP and nuclear receptor mRNAs exists, or other miRNAs or a combination of these could be responsible for directly controlling the mRNA expression of biotransformation-related genes.
Further research in this direction is needed and is anticipated to yield a novel anti-differentiation strategy.
ReferencesAdler, S., Basketter, D., Creton, S., et al. (2011). Alternative
(non-animal) methods for cosmetic testing: current status and future prospects – 2010. Arch. Toxicol. 85, 367-485.
Ambros, V. (2004). the functions of animal microRNAs. Na-ture 431, 350-355.
Andersen, C. l., Jensen, J. l., and Orntoft, t. F. (2004). Nor-malization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64, 5245-5250.
Bissell, D. M. (1976). Study of hepatocyte function in cell cul-ture. Prog. Liver Dis. 5, 69-82.
Ceelen, l., De Spiegelaere, W., David, M., et al. (2011). Criti-cal selection of reliable reference genes for gene expresison study in the HepaRG cell line. Biochem. Pharmacol. 81, 1255-1261.
Chang, J., Nicolas, e., Marks, D., et al. (2004). miR-122, a mammalian liver-specific microRNA, is processed from hcr mRNA and may downregulate the high affinity cationic ami-no acid transporter CAt-1. RNA Biol. 1, 106-113.
Chang, J., Guo, J. T., Jiang, D., et al. (2008). Liver-specific mi-croRNA miR-122 enhances the replication of hepatitis C vi-rus in nonhepatic cells. J. Virol. 82, 8215-8223.
Chuang, J. C. and Jones, P. A. (2007). epigenetics and microR-NAs. Pediatr. Res. 61, 24-29.
Coulouarn, C., Factor, V. M., Andersen, J. B., et al. (2009). loss of miR-122 expression in liver cancer correlates with sup-pression of the hepatic phenotype and gain of metastatic prop-erties. Oncogene 28, 3526-3536.
elaut, G., Henkens, t., Papeleu, P., et al. (2006). Molecular mechanisms underlying the dedifferentiation process of iso-lated hepatocytes and their cultures. Curr. Drug Metab. 7, 629-660.
esau, C., Davis, S., Murray, S. F., et al. (2006). miR-122 regula-tion of lipid metabolism revealed by in vivo antisense target-ing. Cell Metab. 3, 87-98.
european Commission (2003). Directive 2003/15/eC of the eu-ropean parliament and of the council of 27 February 2003 amending council directive 76/768/eeC on the approxima-tion of the laws of the member states relating to cosmetic products.
european Commission (2006). Regulation (eC) No 1907/2006 of the european parliament and of the council of 18 Decem-ber 2006 concerning the registration, evaluation, authorisa-
down- and upregulation of CYP1A2 and CYP2B1 at 24 and 96 hours, respectively. After 24 hours, a significant downregu-lation also was seen for CYP3A2 between control and inhibi-tor conditions. However, these significant differences were lost during further cultivation and were not observed at other time points. Aryl hydrocarbon receptor (AhR) remained consistent-ly stable for all conditions throughout the cultivation regime. By contrast, the expression of constitutive androstane recep-tor (CaR) was downregulated upon cultivation and pregnane x receptor (PxR) levels were slightly upregulated, followed by a decrease to the initial levels. No significant differences in expression of all nuclear receptors investigated were found be-tween the three experimental conditions at any time points.
4 Discussion
Since the liver is a primary target for toxicity, cultures of hepa-tocytes are envisaged as relevant in vitro models in the area of liver toxicology (elaut et al., 2006; Sahi et al., 2010). However, a major limitation to the use of primary hepatocyte cultures is that they rapidly lose their liver-specific phenotype, including their CYP P450 content (Bissell, 1976; Paine, 1990; Paine and Andreakos, 2004). In order to counteract or delay this dedif-ferentiation, our lab has developed various strategies, including epigenetic modification of the hepatocellular gene expression by interfering with histone acetylation and/or DNA methylation (Patent submissions WO2006/045331 and WO2008/145736, Henkens et al., 2007, 2011; Fraczek et al., 2011).
Another approach to modulate the hepatocellular gene ex-pression relies on the modulation of microRNA. microRNAs are noncoding RNAs that are key post-transcriptional regula-tors of gene expression and that also constitute the epigenome (Ambros, 2004; Coulouarn et al., 2009; Chuang and Jones, 2007; Rouhi et al., 2008; Sato et al., 2011). In this study, we have focused on the liver-specific miR-122 (Lagos-Quintana et al., 2002; Chang et al., 2004). Several reports have empha-sized the importance of miR-122 in liver homeostasis. Spe-cifically, it was shown to play an important role in fatty acid and cholesterol metabolism (Chang et al., 2004; Chang et al., 2008; esau et al., 2006). Moreover, Coulouarn and colleagues (2009) demonstrated that the loss of miR-122 in liver tumor cells is linked to the suppression of the hepatic differentiated phenotype. to our best knowledge, no studies have yet been performed to test the possible role of miR-122 in xenobiotic biotransformation capacity.
We modulated the miR-122 hepatic expression by transfect-ing either miR-122 mimic or miR-122 inhibitor into cultured primary rat hepatocytes. As expected, miR-122 showed a pro-gressive decline in its expression in conventional cultures, which can be associated with the overall loss of the hepatic phenotype. Our results further show that inhibition or overexpression of miR-122 has no significant effect on mRNA levels of the in-vestigated phase I CYP 450 genes and their nuclear receptors. Hence, miR-122 does not seem to be involved in direct mRNA cleavage of the former in primary hepatocytes, in contrast to its

Bolleyn et al.
Altex Proceedings, 1/12, Proceedings of WC8172
Russell, W. M. S. and Burch, R. l. (1959). The principles of hu-man experimental technique. london: Methuen & Co., ltd.
Sahi, J., Grepper, S., and Smith, C. (2010). Hepatocytes as a tool in drug metabolism, transport and safety evaluations in drug discovery. Curr. Drug Discov. Technol. 7, 188-198.
Sato, F., tsuchiya, S., Meltzer, S. J., and Shimizu, K. (2011). MicroRNA and epigenetics. FEBS J. 278, 1598-1609.
Schmittgen, t. D. and livak, K. J. (2008). Analyzing real-time PCR data by the comparative C(t) method. Nat. Protoc. 3, 1101-1108.
Vandesompele, J., De Preter, K., Pattyn, F., et al. (2002). Ac-curate normalization of real-time quantitative Rt-PCR data by geometric averaging of multiple internal controls. Genome Biol. 3, ReSeARCH0034.
Vanhaecke, t. and Rogiers, V. (2006). Hepatocyte cultures in drug metabolism and toxicological research and testing. Methods Mol. Biol. 320, 209-227.
Vanhaecke, t., Snykers, S., Rogiers, V., et al. (2009). eU re-search activities in alternative testing strategies: current status and future perspectives. Arch. Toxicol. 83, 1037-1042.
Vanhaecke, t., Pauwels, M., Vinken, M., et al. (2011). towards an integrated in vitro strategy for repeated dose toxicity test-ing. Arch. Toxicol. 85, 365-366.
Acknowledgementsthe authors would like to thank Mr P. Claes, Mr S. Branson, and Mrs A. Heymans for their dedicated technical assistance.
Financial supportThis work is financially supported by grants from the Fund for Scientific Research (FWO), Vlaanderen, Belgium, the Research Council (OZR) of the Vrije Universiteit Brussel, Belgium and the Johns Hopkins Center for Alternatives to Animal testing (CAAt), Baltimore, USA.
Correspondence toJennifer BolleynCenter for Pharmaceutical ResearchDepartment of toxicologyDermato-Cosmetology and PharmacognosyVrije Universiteit Brussel (VUB)laarbeeklaan 1031090 BrusselsBelgiumPhone: +32 2 477 45 17Fax: +32 2 477 45 823e-mail: [email protected]
tion and restriction of chemicals (ReACH), establishing a european chemicals agency, amending directive 1999/45/eC and repealing council regulation (eeC) No 793/93 and com-mission regulation (eC) No 1488/94 as well as council di-rective 76/769/eeC and commission directives 91/155/eeC, 93/67/eeC, 93/105/eC and 2000/21/eC.
Filipowics, W. and Grosshans, H. (2011). The liver-specific mi-croRNA miR-122: biology and therapeutic potential. Prog. Drug Res. 67, 221-238.
Fraczek, J. e., Vinken, M., trouwé, D., et al. (2011). Synergetic effects of DNA methylation and histone deacetylase inhibi-tion in primary rat hepatocytes. Invest. New Drugs, in press.
Guguen-Guillouzo, C. and Guillouzo, A. (2010). General re-view on in vitro hepatocyte models and their applications. Methods Mol. Biol. 640, 1-40.
Henkens, t., Vanhaecke, t., Papeleu, P., et al. (2005). Rat hepa-tocyte cultures: conventional monolayer and cocultures with rat liver epithelial cells. Methods Mol. Biol. 320, 239-246.
Henkens, t., Papeleu, P., Vinken, M., et al. (2007). trichostatin A, a critical factor in maintaining the functional differentia-tion of primary cultured rat hepatocytes. Toxicol. Appl. Phar-macol. 218, 64-71.
Henkens, t., Snykers, S., Vinken, M., et al. (2011). Preservation of hepatocellular functionality upon exposure to 4-Me2N-BAVAH, a hydroxamate-based HDAC-inhibitor. Toxicol. In Vitro 25, 100-109.
lagos-Quintana, M., Rauhut, R., Yalcin, A., et al. (2002). Iden-tification of tissue-specific microRNAs from mouse. Curr. Biol. 12, 735-739.
Moore, K. J., Rayner, K. J., Suárez, Y., and Fernández-Hernan-do, C. (2011). The role of microRNAs in cholesterol efflux and hepatic lipid metabolism. Annu. Rev. Nutr. 31, 49-63.
Paine, A. J. (1990). the maintenance of cytochrome P-450 in rat hepatocyte culture: some applications of liver cell cultures to the study of drug metabolism, toxicity and the induction of the P-450 system. Chem. Biol. Interact. 74, 1-31.
Paine, A. J. and Andreakos, e. (2004). Activation of signalling pathways during hepatocyte isolation: relevance to toxicol-ogy in vitro. Toxicol. In Vitro 18, 187-193.
Papeleu, P., Vanhaecke, t., Henkens, t., et al. (2005). Isolation of rat hepatocytes. Methods Mol. Biol. 320, 229-237.
Pfaffl, M. W., Tichopad, A., Prgomet, C., and Neuvians, T. P. (2004). Determination of stable housekeeping genes, differen-tially regulated target genes and sample integrity: BestKeeper – excel-based tool using pair-wise correlations. Biotechnol. Lett. 26, 509-515.
Rouhi, A., Mager, D. l., Humphries, R. K., and Kuchenbauer, F. (2008). MiRNAs, epigenetics, and cancer. Mamm. Genome 19, 517-525.