
Linking novelty seeking and harm avoidance personality traits to basal ganglia: volumetry and mean...
11
ORIGINAL ARTICLE Linking novelty seeking and harm avoidance personality traits to basal ganglia: volumetry and mean diffusivity Daniela Laricchiuta • Laura Petrosini • Fabrizio Piras • Debora Cutuli • Enrica Macci • Eleonora Picerni • Chiara Chiapponi • Carlo Caltagirone • Gianfranco Spalletta Received: 4 December 2012 / Accepted: 26 February 2013 Ó Springer-Verlag Berlin Heidelberg 2013 Abstract Novelty Seeking (NS) and Harm Avoidance (HA) temperamental traits are related to approaching or avoiding motivational circuits relying on the integrity and functionality of distributed brain areas implicated in arousal and action. The present study verified whether and how macro- and micro-structural variations of basal gan- glia are correlated with scores obtained in the NS and HA temperamental scales of the Temperament and Character Inventory by Cloninger. To this aim, 125 healthy adults aged 18–67 years of both sexes completed the Tempera- ment and Character Inventory and underwent a high-reso- lution T1-weighted magnetic resonance imaging and a diffusion tensor imaging using a 3T scanner. The scores obtained in the temperamental scales were associated with volumes, mean diffusivity and fractional anisotropy mea- sures of basal ganglia of both hemispheres separately, by using linear regression analyses. We found increased bilateral caudate and pallidum volumes associated with higher NS scores, as well as increased mean diffusivity in the bilateral putamen associated with higher HA scores. Macro- and micro-structural variations of basal ganglia regions contribute to explain the biological variance asso- ciated with NS or HA personality phenotype. The present findings evidencing some brain-temperament relationships highlight the importance of obtaining macro- and micro- structural measures in relation to individual differences. Keywords Individual differences Á Brain volumes Á Mean diffusivity Á Fractional anisotropy Á Grey matter Introduction The most qualified personality theories follow different approaches, as the trait adjective (Eysenck and Eysenck 1985), the affective disposition (Tellegen 1985; Watson and Clark 1993) and the motivational system (Gray 1987; Lang 1995) approaches. Elliot and Thrash (2002) showed that two latent factors, approach and avoidance temperaments, account for the shared variance among these approaches. Namely, approach/avoidance temperament is defined as a general neurobiological sensitivity to positive/negative stimuli respectively, accompanied by a perceptual vigilance for, an affective reactivity to, and a behavioral predisposition towards such stimuli (Elliot 2008). Temperamental traits determine approaching or avoiding disposition to attachment and to the early emotions of fear and anger as well as allow emitting different automatic responses to the stimuli of novelty, danger and reward. Namely, temperamental traits of Novelty Seeking (NS) and Harm Avoidance (HA), as defined in the Temperament and Character Inventory (TCI) by Cloninger (1986), are retained to be related to approaching or avoiding motivational circuits relying on the functionality of distributed areas implicated in arousal and Electronic supplementary material The online version of this article (doi:10.1007/s00429-013-0535-5) contains supplementary material, which is available to authorized users. D. Laricchiuta Á L. Petrosini Á F. Piras Á D. Cutuli Á E. Macci Á E. Picerni Á C. Chiapponi Á C. Caltagirone Á G. Spalletta I.R.C.C.S. Santa Lucia Foundation, Via Ardeatina 306, 00142 Rome, Italy D. Laricchiuta (&) Á L. Petrosini Á D. Cutuli Á E. Picerni Department of Psychology, Faculty of Medicine and Psychology, University ‘‘Sapienza’’ of Rome, Via dei Marsi 78, 00185 Rome, Italy e-mail: [email protected] C. Caltagirone Department of Neuroscience, Tor Vergata University, Via Montpellier 1, 00135 Rome, Italy 123 Brain Struct Funct DOI 10.1007/s00429-013-0535-5
-
Upload
fabrizio-piras -
Category
Documents
-
view
213 -
download
0
Transcript of Linking novelty seeking and harm avoidance personality traits to basal ganglia: volumetry and mean...

ORIGINAL ARTICLE
Linking novelty seeking and harm avoidance personality traitsto basal ganglia: volumetry and mean diffusivity
Daniela Laricchiuta • Laura Petrosini • Fabrizio Piras • Debora Cutuli •
Enrica Macci • Eleonora Picerni • Chiara Chiapponi • Carlo Caltagirone •
Gianfranco Spalletta
Received: 4 December 2012 / Accepted: 26 February 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract Novelty Seeking (NS) and Harm Avoidance
(HA) temperamental traits are related to approaching or
avoiding motivational circuits relying on the integrity and
functionality of distributed brain areas implicated in
arousal and action. The present study verified whether and
how macro- and micro-structural variations of basal gan-
glia are correlated with scores obtained in the NS and HA
temperamental scales of the Temperament and Character
Inventory by Cloninger. To this aim, 125 healthy adults
aged 18–67 years of both sexes completed the Tempera-
ment and Character Inventory and underwent a high-reso-
lution T1-weighted magnetic resonance imaging and a
diffusion tensor imaging using a 3T scanner. The scores
obtained in the temperamental scales were associated with
volumes, mean diffusivity and fractional anisotropy mea-
sures of basal ganglia of both hemispheres separately, by
using linear regression analyses. We found increased
bilateral caudate and pallidum volumes associated with
higher NS scores, as well as increased mean diffusivity in
the bilateral putamen associated with higher HA scores.
Macro- and micro-structural variations of basal ganglia
regions contribute to explain the biological variance asso-
ciated with NS or HA personality phenotype. The present
findings evidencing some brain-temperament relationships
highlight the importance of obtaining macro- and micro-
structural measures in relation to individual differences.
Keywords Individual differences � Brain volumes �Mean
diffusivity � Fractional anisotropy � Grey matter
Introduction
The most qualified personality theories follow different
approaches, as the trait adjective (Eysenck and Eysenck
1985), the affective disposition (Tellegen 1985; Watson and
Clark 1993) and the motivational system (Gray 1987; Lang
1995) approaches. Elliot and Thrash (2002) showed that two
latent factors, approach and avoidance temperaments,
account for the shared variance among these approaches.
Namely, approach/avoidance temperament is defined as a
general neurobiological sensitivity to positive/negative
stimuli respectively, accompanied by a perceptual vigilance
for, an affective reactivity to, and a behavioral predisposition
towards such stimuli (Elliot 2008). Temperamental traits
determine approaching or avoiding disposition to attachment
and to the early emotions of fear and anger as well as allow
emitting different automatic responses to the stimuli of
novelty, danger and reward. Namely, temperamental traits of
Novelty Seeking (NS) and Harm Avoidance (HA), as
defined in the Temperament and Character Inventory (TCI)
by Cloninger (1986), are retained to be related to
approaching or avoiding motivational circuits relying on the
functionality of distributed areas implicated in arousal and
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00429-013-0535-5) contains supplementarymaterial, which is available to authorized users.
D. Laricchiuta � L. Petrosini � F. Piras � D. Cutuli � E. Macci �E. Picerni � C. Chiapponi � C. Caltagirone � G. Spalletta
I.R.C.C.S. Santa Lucia Foundation, Via Ardeatina 306,
00142 Rome, Italy
D. Laricchiuta (&) � L. Petrosini � D. Cutuli � E. Picerni
Department of Psychology, Faculty of Medicine and
Psychology, University ‘‘Sapienza’’ of Rome,
Via dei Marsi 78, 00185 Rome, Italy
e-mail: [email protected]
C. Caltagirone
Department of Neuroscience, Tor Vergata University,
Via Montpellier 1, 00135 Rome, Italy
123
Brain Struct Funct
DOI 10.1007/s00429-013-0535-5

action (LeDoux 2000). Excessive tendency to NS or HA
predicts vulnerability to psychiatric disorders (Richter and
Brandstrom 2009). In particular, high levels of behavioral
inhibition, as in high-scored HA subjects, determine
increased risk for developing anxiety disorders and depres-
sion (Biederman et al. 2001; Muris et al. 2001) and con-
versely, high levels of impulsive behavior, as in high-scored
NS subjects, determine increased risk of exhibiting sub-
stance abuse and antisocial behavior (Meyer et al. 1999;
Mitchell and Nelson-Gray 2006). However, apart from
individuals with neuropsychiatric symptoms whose scores
fall at the extreme ends of the normal distribution for each
personality trait, NS and HA are part of not-dysfunctional
behaviors and contribute to adaptive functioning. In fact,
even in not-abnormal situations, the variance in the normal
range of expression of personality traits appears to be linked
to structural variance in specific brain structures. In partic-
ular, it has been demonstrated that NS scores positively
correlate with volumes of frontal and posterior cingulate
cortices, while HA scores negatively correlate with volumes
of orbito-frontal, occipital and parietal cortices (Gardini
et al. 2009). The strength of fiber tracts from hippocampus
and amygdala to striatum predicts individual differences in
NS (Cohen et al. 2009), while decreased micro-structural
integrity of white matter (WM) in cortico-limbic circuit is
associated with high HA scores (Westlye et al. 2011). Fur-
thermore, striatal activity is correlated with novelty-based
choices (Wittmann et al. 2008). Subjects characterized by
relatively low striatal dopaminergic receptor density are
reported to score lowest on NS and highest on HA (Montag
et al. 2010). Very recently, we reported that NS scores are
positively and HA scores are negatively associated with
cerebellar WM and cortex volumes (Laricchiuta et al.
2012a). Since personality traits are related to the motiva-
tional reactions that imply the involvement of a variety of
deep structures concerned in arousal and action (LeDoux
2000; Cohen et al. 2009; Westlye et al. 2011), it seemed
important to test the hypothesis that NS and HA personality
traits are reflected in structural variations in bilateral deep
gray matter (GM) structures. Assuming that variability in
NS and HA is normally distributed, the present research was
performed on a large sample of subjects without psychiatric
diagnosis to minimize the influence of disease-related and
environmental confounders, as recently suggested by
Westlye et al. (2011). In fact, the methodological approach
comparing the so-called normal control subjects to individ-
uals with specific neuro-psychiatric disorders might belie
empirical evidence and suggests that many human traits are
normally distributed. The exclusion of subjects with neuro-
psychiatric disorders is a valid approach, although contro-
versial in relation to specific state-dependent phenomena
associated with pathological conditions (Cannon et al. 2007;
Reimold et al. 2008; Selvaraj et al. 2011). A recent
‘oversampling’ study demonstrated that serotonin trans-
porter density was associated with the variance in character
and not in temperament in healthy individuals selected for
high or low HA scores (Tuominen et al. 2012). Thus,
although further studies on the relationships between brain
structure and function in relation to character personality
variations are needed, studying brain-behavior relations
within healthy subjects exhibiting normal temperamental
personality variations might provide critical insight into the
neural substrate of human behavior and psychopathology.
To this aim, in the same large cohort of healthy adults whose
TCI scores have been associated to cerebellar volumes
(Laricchiuta et al. 2012a), we investigated the associations
between NS and HA scores with variations in macro- (vol-
ume) and micro- (Mean Diffusivity, MD; Fractional
Anisotropy, FA) structural values in basal ganglia through a
high-resolution structural magnetic resonance imaging
(MRI) and a diffusion tensor imaging (DTI) scan protocol.
Noteworthy, in front of a very limited number of reports
describing DTI variations of WM in relation to personality
traits, the present research is the first report addressing GM
micro-structural data in relation to NS and HA individual
differences. DTI is sensitive to the direction and degree of
water displacement in biological tissues. Namely, MD is a
scalar measure of the total diffusion within a voxel and FA
measures anisotropy of water diffusion processes (Pierpaoli
et al. 1996). In physiological states, extracellular water
diffusion is influenced by different factors, such as pore
size between cells, cellular structure, density and surface
(Le Bihan 2007; Sykova and Nicholson 2008; Concha et al.
2010). In WM, water molecules are limited in the direc-
tions of diffusion, resulting in a high FA value. Conversely,
in GM water molecular diffusion exhibits significantly less
directional dependence, causing low FA values relative to
WM. Thus, although the link between information pro-
cessing and diffusion properties is not yet fully clarified,
diffusion parameter changes are suggested to affect the
efficacy of synaptic and extra-synaptic transmission (Sy-
kova 2004). Variations in water diffusion parameters could
be linked to variations in cognitive functions (Piras et al.
2010, 2011) and personality dimensions (Westlye et al.
2011; Bjørnebekk et al. 2012). In this view, DTI measures
represent a reliable research tool, supplying physiological
information not available on conventional MRI.
Methods
Participants
The same large cohort of healthy adults whose TCI scores
have been associated to cerebellar volumes in a recent study
(Laricchiuta et al. 2012a) was used in the present study. In
Brain Struct Funct
123

particular, the sample of 125 neurological intact subjects [52
males (42 %); mean age ± SD = 34.9 ± 12.4 years, range
18.2–67.4] was recruited from Universities, community
recreational centers and hospital personnel by local adver-
tisement. Education level ranged from an eighth grade
education to a post-graduate degree (mean educa-
tion ± SD = 15.5 ± 2.8 years, range 8–24). All partici-
pants were right-handed as assessed with the Edinburgh
Handedness Inventory (Oldfield 1971). The subjects were
submitted to MRI. The inclusion criteria were age between
18 and 70 years and suitability for MRI scanning. Exclusion
criteria included (1) suspicion of cognitive impairment or
dementia based on Mini Mental State Examination (Fol-
stein et al. 1975) score B24 (Measso et al. 1993), identi-
fying positive screening for cognitive deterioration in
Italian population and confirmed by clinical neuropsycho-
logical evaluation using the Mental Deterioration Battery
(Carlesimo et al. 1996) and NINCDS-ADRDA criteria for
dementia (McKhann et al. 1984); (2) subjective complaint
of memory difficulties or of any other cognitive deficits,
interfering or not with the daily living activities; (3) major
medical illnesses, e.g., diabetes (not stabilized), obstructive
pulmonary disease, or asthma; hematological and oncologic
disorders; pernicious anaemia; clinically significant and
unstable active gastrointestinal, renal, hepatic, endocrine, or
cardiovascular system disease; newly treated hypothyroid-
ism; (4) current or reported psychiatric, assessed by the
SCID-I and the SCID-II (First et al. 1997a, b) or neuro-
logical (assessed by a clinical neurological evaluation)
disorders (e.g., schizophrenia, mood disorders, anxiety
disorders, stroke, Parkinson disease, seizure disorder, head
injury with loss of consciousness, and any other significant
mental or neurological disorder), (5) known or suspected
history of alcoholism or drug dependence and abuse during
life-time, (6) MRI evidence of focal parenchymal abnor-
malities or cerebrovascular diseases: for each subject, a
trained neuroradiologist and a neuropsychologist expert in
neuroimaging co-inspected all the available clinical MRI
sequences (i.e. T1 and T2-weighted and FLAIR images) to
ensure that subjects were free from structural brain
pathology and vascular lesions (i.e. T2-weighted hyperin-
tensities or T1-weighted hypointensities). On the basis of
inclusion criteria, in a quality control previous to sample
definition we had excluded 28 subjects (the most elderly)
showing hyperintensities evident on T2-weighted MRI
sequences.
The same inclusion–exclusion criteria were used in our
previous study on cerebellar involvement in personality
differences (Laricchiuta et al. 2012a).
The study was approved by the Local Ethics Committee
of the I.R.C.C.S. Santa Lucia Foundation and written
consent was obtained from all participants after a full
explanation of study procedures.
Temperament and character inventory
Temperament and Character Inventory consists of 240
items comprising 7 dimensions, including 4 temperament
scales (NS, HA, Reward Dependence and Persistence) and
3 character scales (Self-directedness, Cooperativeness and
Self-transcendence) (Cloninger et al. 1993). We focused on
NS and HA stable traits with high heritability (Cloninger
1986; Cloninger et al. 1993; Stallings et al. 1996). NS
refers to a tendency to action behaviors and it is expressed
as the tendency to exploratory activity in response to
novelty, impulsive decision making, extravagant approach
to cues of reward and quick loss of temper. HA describes a
tendency to intensely respond to aversive stimuli, leading
to avoidance behavior. Individuals with high HA are
characterized as cautious, tense, fearful, worried, shy, and
easily fatigable. In the present sample NS mean scores
were 21.0 ± SD = 5.5, while HA mean scores were
13.4 ± SD = 6.3.
MRI acquisition and DTI analysis
All 125 participants underwent the same imaging protocol,
which included standard clinical sequences (FLAIR, DP-T2-
weighted), whole-brain T1-weighted and diffusion-weighted
scanning using a 3T Allegra MR imager (Siemens, Erlan-
gen, Germany) with a standard quadrature head coil. All
planar sequence acquisitions were obtained in the plane of
the anterior-posterior commissure line. Particular care was
taken to center the subjects in the head coil and to restrain
their movements with cushions and adhesive medical
tape. Diffusion-weighted volumes were acquired using
echo-planar imaging (TE/TR = 89/8,500 ms, bandwidth =
2,126 Hz/vx; matrix size: 128 9 128; 80 axial slices, voxel
size: 1.8 9 1.8 9 1.8 mm3) with 30 isotropically distributed
orientations for the diffusion-sensitizing gradients at a
b-value of 1,000 s/mm2 and six b = 0 images. Scanning
was repeated three times to increase the signal-to-noise ratio.
Whole-brain T1-weighted images were obtained in the
sagittal plane using a modified driven equilibrium Fourier
transform (MDEFT) sequence (TE/TR = 2.4/7.92 ms, flip
angle: 15�, voxel-size: 1 9 1 9 1 mm3).
Image processing was performed using FSL 4.1 (www.
fmrib.ox.ac.uk/fsl/). Image distortions induced by eddy
currents and head motion in the DTI data were corrected by
applying a 3D full affine (mutual information cost func-
tion) alignment of each image to the mean no-diffusion
weighting (b0) image. After these corrections, DTI data
were averaged and concatenated into 31 (1 b0 ? 30 b1000)
volumes. A diffusion tensor model was fitted at each voxel,
generating FA and MD maps. The FA maps were used to
obtain a better co-registration with T1-weighted images
because the spatial distribution of signal intensities was
Brain Struct Funct
123

similar in both image modalities, and MD values were used
as index of micro-structural integrity within the deep GM
nuclei. The FA maps created were registered to brain-
extracted whole-brain volumes from T1-weighted images
using a full affine (correlation ratio cost function) align-
ment with nearest-neighbor resampling. The calculated
transformation matrix was applied to the MD maps with
identical resampling options.
Anatomical T1-weighted images were processed with
the segmentation tool FIRST 1.1 integrated in the FSL
software. This is a model-based segmentation/registration
tool. The shape/appearance models used in FIRST are
constructed from manually segmented images provided by
the Center for Morphometric Analysis (CMA), MGH,
Boston, MA. The manual labels are parameterized as sur-
face meshes and modeled as a point distribution model.
Deformable surfaces are used to automatically parameterize
the volumetric labels in terms of meshes; the deformable
surfaces are constrained to preserve vertex correspondence
across the training data. Furthermore, normalized intensities
along the surface are sampled and modeled. The shape and
appearance model is based on multivariate Gaussian
assumptions. Shape is then expressed as a mean with modes
of variation (principal components). On the basis of the
learned models, FIRST searches through linear combina-
tions of shape modes of variation for the most probable
shape instance given the intensities observed in the T1
image. In other words, this tool is optimized to find the
optimal border and extent of the structures considered,
modeling these structures as surfaces.
This method of segmentation is particularly useful for
structures with a low contrast-to-noise ratio. For each sub-
ject and each hemisphere, the caudate (body), the putamen,
and the pallidum were segmented. For each subject, the
results of region of interest (ROI) segmentation and the co-
registered FA map were superimposed on the original T1-
weighted volume and the resulting images were visually
assessed to exclude misregistration or erroneous ROI
identification. For each subject and each hemisphere, we
calculated the volumes of the above-mentioned ROIs. Prior
to statistical analyses, to account for individual differences
in head size, we used the integrated tool SIENAX part of the
FSL software library (Smith et al. 2004) for automatic
evaluation of brain size, atrophy, and GM and WM vol-
umes. It corrects each volume with a multiplicative scaling
factor derived from an affine transform. This Atlas Scaling
Factor (ASF) was computed as the determinant of the affine
transform connecting each individual to the MNI standard
template. The ASF represents the whole-brain volume
expansion (or contraction) required to register each indi-
vidual to the template. In other words, we used for statistical
analyses the normalized volumes calculated as follows:
VolNormalized = ASF*VolReal.
The segmented ROIs defined the binary masks where
mean values of FA and MD were calculated for each
individual. For each subject, all available clinical MRI
sequences (i.e. T1- and T2-weighted and FLAIR images)
were visually assessed also to ensure that subjects were
free from structural brain pathology and vascular lesions.
Statistical analyses
Parametric associations between NS scores, HA scores,
volumes of bilateral structures (i.e., caudate, putamen and
pallidum), DTI (MD and FA) values of the same structures,
age and years of education were tested by using Pearson
product moment correlation (Fisher r to z). The effect of
sex was assessed by using independent-samples t test for
NS scores, HA scores, volumes or DTI values of bilateral
structures. Associations between NS or HA scores and
volumes or DTI values of bilateral structures were tested
by using linear regression analyses with the NS or HA
scores as dependent variable and age, sex, total GM vol-
ume and volumes (or MD or FA values) of bilateral
structures as independent variables. Sex was considered a
‘‘dummy variable’’ given its dichotomic nature.
As in the present study a large number of tests was run,
controlling for the alpha inflation was needed. The pro-
portion of type I errors among all rejected null hypotheses
was controlled by setting the False Discovery Rate (FDR)
to 0.05. The FDR was estimated through the procedure
described by Storey and Tibshirani (2003). The bootstrap
procedure was used to estimate the p0 parameter (Storey
et al. 2004). In our results, the 0.05 level of significance
corresponded to an FDR 0.04. Power analysis calculated on
the multiple linear regressions with 125 participants
showed high statistical power = 1.
Results
Relationships between NS, HA, macro- (volume)
or micro- (MD and FA) structural variations in deep
GM structures and years of education, sex or age
A negative correlation between age and years of education
(r = -0.32, p = 0.0005) was found.
No correlation between HA or NS scores and years of
education (NS: r = 0.08, p = 0.37; HA: r = -0.05,
p = 0.56) was found. Only the bilateral putamen was
positively correlated, as regard the volumes (right:
r = 0.22, p = 0.01; left: r = 0.20, p = 0.02), and nega-
tively correlated, as regard MD values (right: r = -0.32,
p = 0.0001; left: r = -0.29, p = 0.001) with years of
education. No correlation between FA values and years of
education was found in any basal structure.
Brain Struct Funct
123

Male and female participants had similar NS scores
(t = -0.52, p = 0.60). On the contrary, female partici-
pants showed higher HA scores than male participants
(t = 4.01, p = 0.0001). Furthermore, while female partic-
ipants showed smaller volumes for all bilateral structures
than male participants (caudate: right: t = 5.32, p =
0.0001; left: t = 5.02, p = 0.0001; putamen: right:
t = 5.37, p = 0.0001; left: t = 5.53, p = 0.0001; palli-
dum: right: t = 2.57, p = 0.01; left: t = 4.36, p =
0.0001), both females and males showed similar MD and
FA values for all bilateral structures.
No correlation between HA scores and age (r = 0.15,
p = 0.08) was found, while significant negative correla-
tions between NS scores and age (r = -0.33, p = 0.0002),
and between NS and HA scores (r = -0.37, p = 0.0001)
were found.
Negative correlations were found between age and
caudate (right: r = -0.27, p = 0.002; left: r = -0.31,
p = 0.0001), putamen (right: r = -0.39, p = 0.0001; left:
r = -0.40, p = 0.0001) and pallidum (right: r = -0.27,
p = 0.002; left: r = -0.30, p = 0.0001) volumes. Positive
correlations were found between age and bilateral caudate
(right: r = 0.49, p = 0.00001; left: r = 0.27, p = 0.002)
as well as right putamen (r = 0.22, p = 0.01) MD values.
Positive correlations were found between age and bilateral
putamen (right: r = 0.4, p = 0.0001; left: r = 0.4, p = 0.
0001), left caudate (r = 0.21, p = 0.02), as well as left
pallidum (r = 0.20, p = 0.04) FA values.
No correlation between volumes and MD or FA values
as well as between MD and FA values in any bilateral basal
structure was found.
Relationships between NS, HA and macro- (volume)
or micro- (MD and FA) structural variations in deep
GM structures
Results of linear regression analyses used to evaluate the
associations between NS or HA scores and macro- or
micro-structural variations of bilateral basal ganglia are
reported in Table 1 (volume values) and in Table 2 (MD
values). In particular, significant positive associations
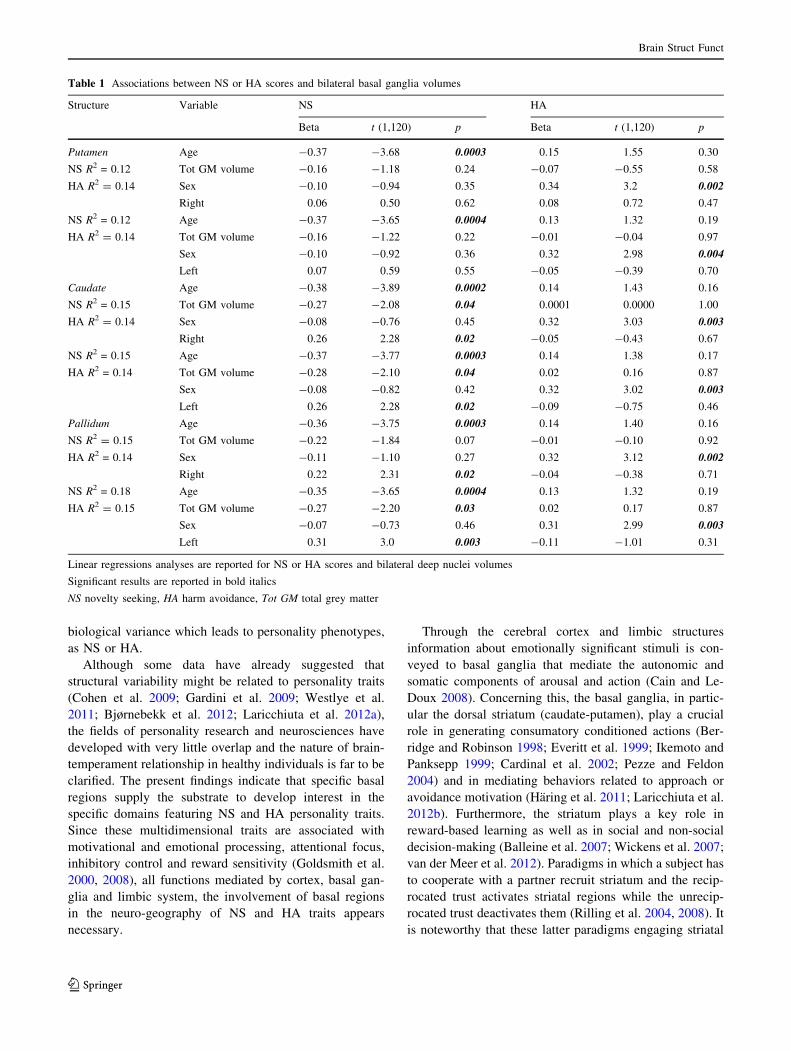
emerged between NS scores and left and right caudate as
well as left and right pallidum volumes (Fig. 1). No asso-
ciation was found between NS scores and bilateral putamen
volumes. NS scores resulted to be associated to total GM
values for both caudate nuclei and for left pallidum. Fur-
thermore, no significant association emerged between NS
scores and MD measures in all GM assemblies.
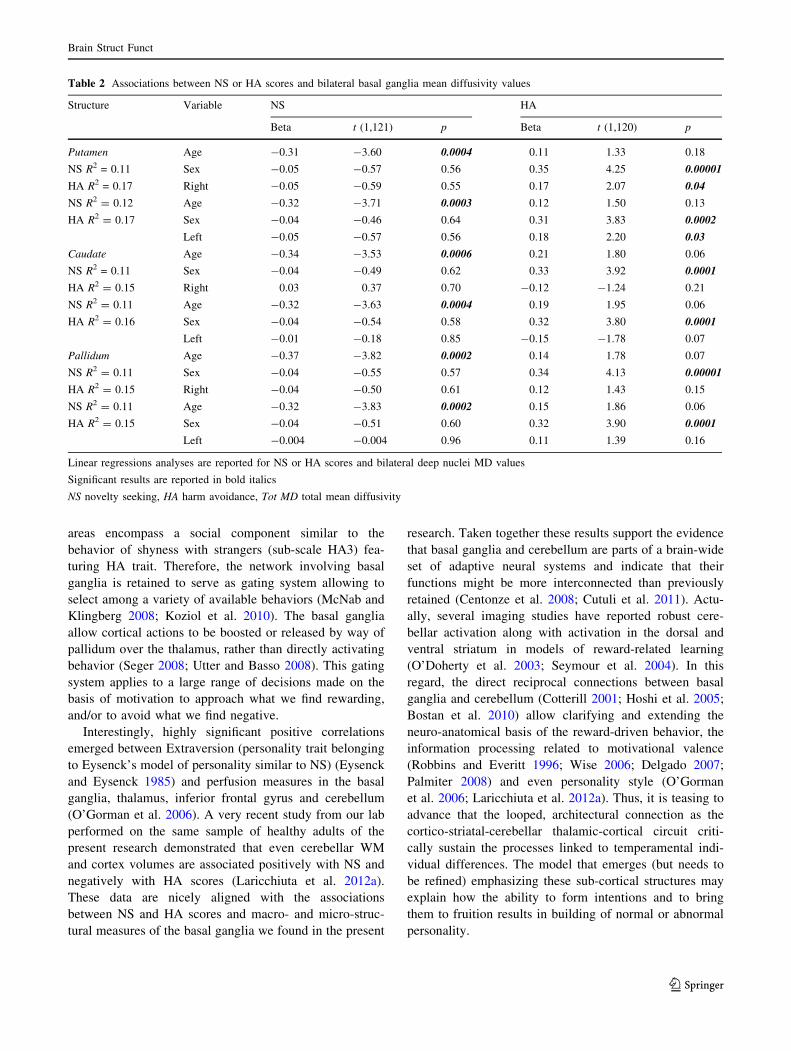
No significant associations were observed between HA
scores and bilateral basal ganglia volumes. Interestingly,
a significant positive association emerged between
HA scores and left and right putamen MD measures
(Fig. 2).
For both volumes and MD of bilateral basal structures,
NS and HA scores resulted to be associated to age and sex,
respectively.
As for FA analyses, no significant association between
NS or HA scores and bilateral basal structures was found
(details are reported in Online Resource Table 1).
Discussion
Novelty Seeking trait is defined as a heritable tendency to
exhibit exploratory activity in pursuit of reward and
avoidance of monotony and it is characterized by a pro-
clivity to impulsivity and risk-taking behaviors, linked to
approach tendencies and arousal regulation. HA trait is
defined as a heritable tendency to withdrawal and inhibi-
tion of behavior and it is characterized by sensitivity to
aversive and non-rewarding stimuli that evoke negative
emotions such as anticipatory worry, fear and anxiety as
well as avoidance behavior. Individuals with high NS
scores are exploratory, impulsive, fickle, excitable, quick-
tempered and extravagant, whereas those with high HA
scores are cautious, passive, fearful of uncertainty, shy and
easily fatigued (Cloninger 1986; Cloninger et al. 1993).
Individuals with neuropsychiatric symptoms such as
depression (Ono et al. 2002), suicidal behavior (Pompili
et al. 2008), bipolar mania (Loftus et al. 2008), schizo-
phrenia (Fresan et al. 2007), substance use disorders
(Conway et al. 2003), pathological gambling (Martinotti
et al. 2006) and anxiety disorders (Kashdan and Hofmann
2008) have scores which fall at the extreme tails of the
normal distribution for each personality trait. However, NS
and HA traits support the adaptive functioning to envi-
ronmental stimuli even in not-dysfunctional situations. In
fact, the associations between macro- and micro-structural
data of GM structures and NS and HA scores indeed
reflected the normal variability in personality traits and not
pathological states. In accord with other studies (Cloninger
et al. 1993; Fresan et al. 2011; Westlye et al. 2011), we
indicated that female participants had HA scores higher
than male participants and younger participants had NS
scores higher than older participants. Furthermore, we
found that the bilateral putamen was positively correlated
as regard the volumes, and negatively correlated as regard
MD values, with years of education.
Specific associations between NS and HA scores and
basal ganglia structure were demonstrated: bilateral cau-
date and pallidum volumes correlated positively with NS
trait, while bilateral putamen MD measures correlated
positively with HA trait. No association was found between
temperamental traits and FA measures. Thus, it may be
advanced that macro- (volume) and micro- (MD) structural
integrity of basal structures contributes to explain the
Brain Struct Funct
123
biological variance which leads to personality phenotypes,
as NS or HA.
Although some data have already suggested that
structural variability might be related to personality traits
(Cohen et al. 2009; Gardini et al. 2009; Westlye et al.
2011; Bjørnebekk et al. 2012; Laricchiuta et al. 2012a),
the fields of personality research and neurosciences have
developed with very little overlap and the nature of brain-
temperament relationship in healthy individuals is far to be
clarified. The present findings indicate that specific basal
regions supply the substrate to develop interest in the
specific domains featuring NS and HA personality traits.
Since these multidimensional traits are associated with
motivational and emotional processing, attentional focus,
inhibitory control and reward sensitivity (Goldsmith et al.
2000, 2008), all functions mediated by cortex, basal gan-
glia and limbic system, the involvement of basal regions
in the neuro-geography of NS and HA traits appears
necessary.
Through the cerebral cortex and limbic structures
information about emotionally significant stimuli is con-
veyed to basal ganglia that mediate the autonomic and
somatic components of arousal and action (Cain and Le-
Doux 2008). Concerning this, the basal ganglia, in partic-
ular the dorsal striatum (caudate-putamen), play a crucial
role in generating consumatory conditioned actions (Ber-
ridge and Robinson 1998; Everitt et al. 1999; Ikemoto and
Panksepp 1999; Cardinal et al. 2002; Pezze and Feldon
2004) and in mediating behaviors related to approach or
avoidance motivation (Haring et al. 2011; Laricchiuta et al.
2012b). Furthermore, the striatum plays a key role in
reward-based learning as well as in social and non-social
decision-making (Balleine et al. 2007; Wickens et al. 2007;
van der Meer et al. 2012). Paradigms in which a subject has
to cooperate with a partner recruit striatum and the recip-
rocated trust activates striatal regions while the unrecip-
rocated trust deactivates them (Rilling et al. 2004, 2008). It
is noteworthy that these latter paradigms engaging striatal
Table 1 Associations between NS or HA scores and bilateral basal ganglia volumes
Structure Variable NS HA
Beta t (1,120) p Beta t (1,120) p
Putamen Age -0.37 -3.68 0.0003 0.15 1.55 0.30
NS R2 = 0.12
HA R2 = 0.14
Tot GM volume -0.16 -1.18 0.24 -0.07 -0.55 0.58
Sex -0.10 -0.94 0.35 0.34 3.2 0.002
Right 0.06 0.50 0.62 0.08 0.72 0.47
NS R2 = 0.12
HA R2 = 0.14
Age -0.37 -3.65 0.0004 0.13 1.32 0.19
Tot GM volume -0.16 -1.22 0.22 -0.01 -0.04 0.97
Sex -0.10 -0.92 0.36 0.32 2.98 0.004
Left 0.07 0.59 0.55 -0.05 -0.39 0.70
Caudate Age -0.38 -3.89 0.0002 0.14 1.43 0.16
NS R2 = 0.15
HA R2 = 0.14
Tot GM volume -0.27 -2.08 0.04 0.0001 0.0000 1.00
Sex -0.08 -0.76 0.45 0.32 3.03 0.003
Right 0.26 2.28 0.02 -0.05 -0.43 0.67
NS R2 = 0.15
HA R2 = 0.14
Age -0.37 -3.77 0.0003 0.14 1.38 0.17
Tot GM volume -0.28 -2.10 0.04 0.02 0.16 0.87
Sex -0.08 -0.82 0.42 0.32 3.02 0.003
Left 0.26 2.28 0.02 -0.09 -0.75 0.46
Pallidum Age -0.36 -3.75 0.0003 0.14 1.40 0.16
NS R2 = 0.15
HA R2 = 0.14
Tot GM volume -0.22 -1.84 0.07 -0.01 -0.10 0.92
Sex -0.11 -1.10 0.27 0.32 3.12 0.002
Right 0.22 2.31 0.02 -0.04 -0.38 0.71
NS R2 = 0.18
HA R2 = 0.15
Age -0.35 -3.65 0.0004 0.13 1.32 0.19
Tot GM volume -0.27 -2.20 0.03 0.02 0.17 0.87
Sex -0.07 -0.73 0.46 0.31 2.99 0.003
Left 0.31 3.0 0.003 -0.11 -1.01 0.31
Linear regressions analyses are reported for NS or HA scores and bilateral deep nuclei volumes
Significant results are reported in bold italics
NS novelty seeking, HA harm avoidance, Tot GM total grey matter
Brain Struct Funct
123
areas encompass a social component similar to the
behavior of shyness with strangers (sub-scale HA3) fea-
turing HA trait. Therefore, the network involving basal
ganglia is retained to serve as gating system allowing to
select among a variety of available behaviors (McNab and
Klingberg 2008; Koziol et al. 2010). The basal ganglia
allow cortical actions to be boosted or released by way of
pallidum over the thalamus, rather than directly activating
behavior (Seger 2008; Utter and Basso 2008). This gating
system applies to a large range of decisions made on the
basis of motivation to approach what we find rewarding,
and/or to avoid what we find negative.
Interestingly, highly significant positive correlations
emerged between Extraversion (personality trait belonging
to Eysenck’s model of personality similar to NS) (Eysenck
and Eysenck 1985) and perfusion measures in the basal
ganglia, thalamus, inferior frontal gyrus and cerebellum
(O’Gorman et al. 2006). A very recent study from our lab
performed on the same sample of healthy adults of the
present research demonstrated that even cerebellar WM
and cortex volumes are associated positively with NS and
negatively with HA scores (Laricchiuta et al. 2012a).
These data are nicely aligned with the associations
between NS and HA scores and macro- and micro-struc-
tural measures of the basal ganglia we found in the present
research. Taken together these results support the evidence
that basal ganglia and cerebellum are parts of a brain-wide
set of adaptive neural systems and indicate that their
functions might be more interconnected than previously
retained (Centonze et al. 2008; Cutuli et al. 2011). Actu-
ally, several imaging studies have reported robust cere-
bellar activation along with activation in the dorsal and
ventral striatum in models of reward-related learning
(O’Doherty et al. 2003; Seymour et al. 2004). In this
regard, the direct reciprocal connections between basal
ganglia and cerebellum (Cotterill 2001; Hoshi et al. 2005;
Bostan et al. 2010) allow clarifying and extending the
neuro-anatomical basis of the reward-driven behavior, the
information processing related to motivational valence
(Robbins and Everitt 1996; Wise 2006; Delgado 2007;
Palmiter 2008) and even personality style (O’Gorman
et al. 2006; Laricchiuta et al. 2012a). Thus, it is teasing to
advance that the looped, architectural connection as the
cortico-striatal-cerebellar thalamic-cortical circuit criti-
cally sustain the processes linked to temperamental indi-
vidual differences. The model that emerges (but needs to
be refined) emphasizing these sub-cortical structures may
explain how the ability to form intentions and to bring
them to fruition results in building of normal or abnormal
personality.
Table 2 Associations between NS or HA scores and bilateral basal ganglia mean diffusivity values
Structure Variable NS HA
Beta t (1,121) p Beta t (1,120) p
Putamen Age -0.31 -3.60 0.0004 0.11 1.33 0.18
NS R2 = 0.11
HA R2 = 0.17
Sex -0.05 -0.57 0.56 0.35 4.25 0.00001
Right -0.05 -0.59 0.55 0.17 2.07 0.04
NS R2 = 0.12
HA R2 = 0.17
Age -0.32 -3.71 0.0003 0.12 1.50 0.13
Sex -0.04 -0.46 0.64 0.31 3.83 0.0002
Left -0.05 -0.57 0.56 0.18 2.20 0.03
Caudate Age -0.34 -3.53 0.0006 0.21 1.80 0.06
NS R2 = 0.11
HA R2 = 0.15
Sex -0.04 -0.49 0.62 0.33 3.92 0.0001
Right 0.03 0.37 0.70 -0.12 -1.24 0.21
NS R2 = 0.11
HA R2 = 0.16
Age -0.32 -3.63 0.0004 0.19 1.95 0.06
Sex -0.04 -0.54 0.58 0.32 3.80 0.0001
Left -0.01 -0.18 0.85 -0.15 -1.78 0.07
Pallidum Age -0.37 -3.82 0.0002 0.14 1.78 0.07
NS R2 = 0.11
HA R2 = 0.15
Sex -0.04 -0.55 0.57 0.34 4.13 0.00001
Right -0.04 -0.50 0.61 0.12 1.43 0.15
NS R2 = 0.11
HA R2 = 0.15
Age -0.32 -3.83 0.0002 0.15 1.86 0.06
Sex -0.04 -0.51 0.60 0.32 3.90 0.0001
Left -0.004 -0.004 0.96 0.11 1.39 0.16
Linear regressions analyses are reported for NS or HA scores and bilateral deep nuclei MD values
Significant results are reported in bold italics
NS novelty seeking, HA harm avoidance, Tot MD total mean diffusivity
Brain Struct Funct
123
Recently, increased HA scores have been associated
with increased MD and decreased FA measures in WM
cortico-limbic tracts (Westlye et al. 2011). To our knowl-
edge the current research is the first large-scale study
demonstrating association between increased HA scores
and increased MD in GM basal region of putamen. Inter-
estingly, both studies (Westlye et al. 2011 and the present
research) emphasize the relationship between anxiety-
related personality trait and DTI-derived indices of WM
and GM integrity. Given the described involvement of
putamen in inhibitory control (Rubia et al. 2006), the
present findings support the notion that a defective micro-
structural integrity of putamen may lead to dysfunctional
HA trait. Several studies have described the relationship
between repetitive behavior or inhibitory control deficits
and striatal abnormalities in individuals with various neu-
ropsychiatric disorders, such as obsessive compulsive dis-
order, Tourette’s syndrome and autism (Albin and Mink
2006; Langen et al. 2009; van den Heuvel et al. 2010).
Significant micro-structural abnormalities of WM tracts
originating from putamen and nucleus accumbens have
been described in adults with autism (Langen et al. 2012).
Furthermore, it has been reported that HA scores nega-
tively correlate with dopaminergic receptor availability in
the dorsal caudate and putamen (Kim et al. 2011).
Recently, a link between social reward dependency and
WM microstructure in a large healthy sample has been
described (Bjørnebekk et al. 2012).
Fig. 1 Relationship between caudate (upper panel, red color) and pallidum (lower panel, blue color) volumes and Novelty Seeking scores.
Scatterplots are separated for left and right volumes. Linear fit (solid black line) is also reported
Fig. 2 Relationship between putamen (green color) MD values and Harm Avoidance scores. Scatterplots are separated for left and right side.
Linear fit (solid black line) is also reported
Brain Struct Funct
123

The temperamental differences and their neural sub-
strates in healthy subjects may be relevant for under-
standing individual differences in resilience and
vulnerability to clinical psychiatric disorders. Our data
suggest that individuals with a micro-structure of putamen
characterized by higher MD values will be more vulnerable
in experiencing negative emotional states and tendencies to
withdrawal and inhibition. In contrast, individuals with
larger volumes of caudate and pallidum will be more vul-
nerable in experiencing positive emotional states and ten-
dencies to approach. The present findings concerning the
regional specificity of brain-temperament relationships
highlight on the importance of obtaining macro- and micro-
structural measures in the sub-cortical regions related to
motivational and emotional processing. Studies on healthy
and clinical subjects may establish whether the brain-
temperament relationship is maintained across non-patho-
logical variability and full-blown psychiatric disorders.
Acknowledgments The authors would like to thank Prof. Fabio
Ferlazzo for his kind and expert support in statistical analyses.
Conflict of interest The authors declare that they have no conflict
of interest.
References
Albin RL, Mink JW (2006) Recent advances in Tourette syndrome
research. Trends Neurosci 29:175–182
Balleine BW, Delgado MR, Hikosaka O (2007) The role of the dorsal
striatum in reward and decision making. J Neurosci 27:8161–8165
Berridge KC, Robinson TE (1998) What is the role of dopamine in
reward: hedonic impact reward learning or incentive salience?
Brain Res Rev 28:309–369
Biederman J, Hirshfeld-Becker DR, Rosenbaum JF, Herot C,
Friedman D, Snidman N, Kagan J, Faraone SV (2001) Further
evidence of association between behavioral inhibition and social
anxiety in children. Am J Psychiatry 158:1673–1679
Bjørnebekk A, Westlye LT, Fjell AM, Grydeland H, Walhovd KB
(2012) Social reward dependence and brain white matter
microstructure. Cereb Cortex 22:2672–2679
Bostan AC, Dum RP, Strick PL (2010) The basal ganglia commu-
nicate with the cerebellum. Proc Natl Acad Sci USA
107:8452–8456
Cain CK, LeDoux JE (2008) Emotional and motivational processing:
in search of brain mechanisms. In: Elliot AJ (ed) Handbook of
approach and avoidance motivation. Psychology Press Taylor &
Francis Group, New York, pp 17–34
Cannon DM, Ichise M, Rollis D, Klaver JM, Gandhi SK, Charney DS,
Manji HK, Drevets WC (2007) Elevated serotonin transporter
binding in major depressive disorder assessed using positron
emission tomography and [11C]DASB; comparison with bipolar
disorder. Biol Psychiatry 62:870–877
Cardinal RN, Parkinson JA, Hall J, Everitt BJ (2002) Emotion and
motivation: the role of the amygdala ventral striatum and
prefrontal cortex. Neurosci Biobehav Rev 26:321–352
Carlesimo G, Caltagirone C, Gainotti G (1996) The mental deteri-
oration battery: normative data diagnostic reliability and
qualitative analyses of cognitive impairment. The group for the
standardization of the mental deterioration battery. Eur Neurol
36:378–384
Centonze D, Rossi S, De Bartolo P, De Chiara V, Foti F, Musella A,
Mataluni G, Rossi S, Bernardi G, Koch G, Petrosini L (2008)
Adaptations of glutamatergic synapses in the striatum contribute
to recovery from cerebellar damage. Eur J Neurosci
27:2188–2196
Cloninger CR, Svrakic DM, Przybeck TR (1993) A psychobiological
model of temperament and character. Arch Gen Psychiatry
50:975–990
Cloninger CR (1986) A unified biosocial theory of personality and its
role in the development of anxiety states. Psychiatr Dev 4:167–226
Cohen MX, Schoene-Bake JC, Elger CE, Weber B (2009) Connec-
tivity-based segregation of the human striatum predicts person-
ality characteristics. Nat Neurosci 12:32–34
Concha L, Livy DJ, Beaulieu C, Wheatley BM, Gross DW (2010)
In vivo diffusion tensor imaging and histopathology of the
fimbria-fornix in temporal lobe epilepsy. J Neurosci 30:996–1002
Conway KP, Kane RJ, Ball SA, Poling JC, Rounsaville BJ (2003)
Personality substance of choice and polysubstance involvement
among substance dependent patients. Drug Alcohol Depend
71:65–75
Cotterill RM (2001) Cooperation of the basal ganglia cerebellum
sensory cerebrum and hippocampus: possible implications for
cognition consciousness intelligence and creativity. Prog Neu-
robiol 64:1–33
Cutuli D, Rossi S, Burello L, Laricchiuta D, De Chiara V, Foti F, De
Bartolo P, Musella A, Gelfo F, Centonze D, Petrosini L (2011)
Before or after does it matter? Different protocols of environ-
mental enrichment differently influence motor synaptic and
structural deficits of cerebellar origin. Neurobiol Dis 42:9–20
Delgado MR (2007) Reward-related responses in the human striatum.
Ann N Y Acad Sci 1104:70–88
Elliot AJ, Thrash TM (2002) Approach-avoidance motivation in
personality: approach and avoidance temperaments and goals.
J Pers Soc Psychol 82:804–818
Elliot AJ (2008) Approach and Avoidance Motivation. In: Elliot AJ
(ed) Handbook of approach and avoidance motivation. Psychol-
ogy Press Taylor & Francis Group, New York, pp 3–14
Everitt BJ, Parkinson JA, Olmstead MC, Arroyo M, Robledo P,
Robbins TW (1999) Associative processes in addiction and
reward. The role of amygdala-ventral striatal subsystems. Ann N
Y Acad Sci 877:412–438
Eysenck HJ, Eysenck MW (1985) Personality and individual
differences: a natural science approach. Plenum, New York
First MB, Gibbon M, Spitzer RL, Williams JB, Benjamin LS (1997a)
structured clinical interview for DSM-IV Axis II personality
disorders (SCID-II). American Psychiatric Press, Washington
DC
First MB, Spitzer RL, Gibbon M, Williams JB (1997b) Structured
clinical interview for DSM-IV axis I disorders (SCID-I).
Clinician version. American Psychiatric Press, Washington DC
Folstein M, Folstein S, McHugh P (1975) ‘‘Mini-mental state’’. A
practical method for grading the cognitive state of patients for
the clinician. J Psychiatr Res 12:189–198
Fresan A, Apiquian R, Nicolini H, Cervantes JJ (2007) Temperament
and character in violent schizophrenic patients. Schizophr Res
94:74–80
Fresan A, Robles-Garcıa R, Lopez-Avila A, Cloninger CR (2011)
Personality differences according to age and sex in a Mexican
sample using the temperament and character inventory-revised.
Compr Psychiatry 52:774–779
Gardini S, Cloninger CR, Venneri A (2009) Individual differences in
personality traits reflect structural variance in specific brain
regions. Brain Res Bull 79:265–270
Brain Struct Funct
123

Goldsmith HH, Lemery KS, Aksan N, Buss KA (2000) Tempera-
mental substrates of personality development. In: Molfese V,
Molfese D (eds) Temperamental substrates of personality
development. Lawrence Erlbaum, Mahwah NJ, pp 1–32
Goldsmith HH, Pollak SD, Davidson RJ (2008) Developmental
neuroscience perspectives on emotion regulation. Child Dev
Perspect 2:132–140
Gray JA (1987) The psychology of fear and stress, 2nd edn.
Cambridge University Press, New York
Haring M, Kaiser N, Monory K, Lutz B (2011) Circuit specific
functions of cannabinoid CB1 receptor in the balance of
investigatory drive and exploration. PLoS ONE 6:e26617
Hoshi E, Tremblay L, Feger J, Carras PL, Strick PL (2005) The
cerebellum communicates with the basal ganglia. Nat Neurosci
8:1491–1493
Ikemoto S, Panksepp J (1999) The role of nucleus accumbens
dopamine in motivated behavior: a unifying interpretation with
special reference to reward-seeking. Brain Res Rev 31:6–41
Kashdan TB, Hofmann SG (2008) The high-novelty-seeking impul-
sive subtype of generalized social anxiety disorder. Depress
Anxiety 25:535–541
Kim JH, Son YD, Kim HK, Lee SY, Cho SE, Kim YB, Cho ZH
(2011) Association of harm avoidance with dopamine D2/3
receptor availability in striatal subdivisions: a high resolution
PET study. Biol Psychol 87:164–167
Koziol LF, Budding DE, Chidekel D (2010) Adaptation expertise and
giftedness: towards an understanding of cortical subcortical and
cerebellar network contributions. Cerebellum 9:499–529
Lang PJ (1995) The emotion probe: studies of motivation and
attention. Am Psychol 50:372–385
Langen M, Leemans A, Johnston P, Ecker C, Daly E, Murphy CM,
Dell’acqua F, Durston S, AIMS Consortium, Murphy DG (2012)
Fronto-striatal circuitry and inhibitory control in autism: findings
from diffusion tensor imaging tractography. Cortex 48:183–193
Langen M, Schnack HG, Nederveen H, Bos D, Lahuis BE, de Jonge
MV, van Engeland H, Durston S (2009) Changes in the
developmental trajectories of striatum in autism. Biol Psychiatry
66:327–333
Laricchiuta D, Petrosini L, Piras F, Macci E, Cutuli D, Chiapponi C,
Cerasa A, Piperni E, Caltagirone C, Girardi P, Tamorri SM,
Spalletta G (2012a) Linking novelty seeking and harm avoidance
personality traits to cerebellar volumes. Hum Brain Mapp. doi:
10.1002/hbm.22174
Laricchiuta D, Rossi S, Musella A, De Chiara V, Cutuli D, Centonze
D, Petrosini L (2012b) Differences in spontaneously avoiding or
approaching mice reflect Differences in CB1-mediated signaling
of dorsal striatal transmission. PLoS ONE 7:e33260
Le Bihan D (2007) The ‘wet mind’: water and functional neuroim-
aging. Phys Med Biol 52:R57–R90
LeDoux JE (2000) Emotion circuits in the brain. Annu Rev Neurosci
23:155–184
Loftus ST, Garno JL, Jaeger J, Malhotra AK (2008) Temperament
and character dimensions in bipolar I disorder: a comparison to
healthy controls. J Psychiatr Res 42:1131–1136
Martinotti G, Andreoli S, Giametta E, Poli V, Bria P, Janiri L (2006)
The dimensional assessment of personality in pathologic and
social gamblers: the role of novelty seeking and self-transcen-
dence. Compr Psychiatry 47:350–356
McKhann G, Drachman D, Folstein M, Katzman R, Price D, Stadlan
E (1984) Clinical diagnosis of Alzheimer’s disease: report of the
NINCDS-ADRDA. Work Group under the auspices of Depart-
ment of Health and Human Services Task Force on Alzheimer’s
disease. Neurology 34:939–944
McNab F, Klingberg T (2008) Prefrontal cortex and basal ganglia
control access to working memory. Nat Neurosci 11:103–107
Measso G, Cavarzeran F, Zappala‘ G, Lebowitz B, Crook T,
Pirozzolo F, Amaducci L, Massari D, Grigoletto F (1993) The
mini-mental state examination: normative study of an Italian
random sample. Dev Neuropsychol 9:77–85
Meyer B, Johnson SL, Carver CS (1999) Exploring behavioral
activation and inhibition sensitivities among college students at
risk for bipolar spectrum symptomatology. J Psychopathol
Behav Assess 21:275–292
Mitchell JT, Nelson-Gray RO (2006) Attention-deficit/hyperactivity
disorder symptoms in adults: relationship to Gray’s behavioral
approach system. Pers Individ Differ 40:749–760
Montag C, Markett S, Basten U, Stelzel C, Fiebach C, Canli T, Reuter
M (2010) Epistasis of the DRD2/ANKK1 Taq Ia and the BDNF
Val66Met polymorphism impacts novelty seeking and harm
avoidance. Neuropsychopharmacology 35:1860–1867
Muris P, Merckelbach H, Schmidt H, Gadet BB, Bogie N (2001)
Anxiety and depression as correlates of self-reported behavioural
inhibition in normal adolescents. Behav Res Ther 39:1051–1061
O’Doherty JP, Dayan P, Friston K, Critchley H, Dolan RJ (2003)
Temporal difference models and reward-related learning in the
human brain. Neuron 38:329–337
O’Gorman RL, Kumari V, Williams SC, Zelaya FO, Connor SE,
Alsop DC, Gray JA (2006) Personality factors correlate with
regional cerebral perfusion. Neuroimage 31:489–495
Oldfield RC (1971) The assessment and analysis of handedness: the
Edinburgh inventory. Neuropsychologia 9:97–113
Ono Y, Ando J, Onoda N, Yoshimura K, Momose T, Hirano M,
Kanba S (2002) Dimensions of temperament as vulnerability
factors in depression. Mol Psychiatry 7:948–953
Palmiter RD (2008) Dopamine signaling in the dorsal striatum is
essential for motivated behaviors: lessons from dopamine-
deficient mice. Ann NY Acad Sci 1129:35–46
Pezze MA, Feldon J (2004) Mesolimbic dopaminergic pathways in
fear conditioning. Prog Neurobiol 74:301–320
Pierpaoli C, Jezzard P, Basser PJ, Barnett A, Di Chiro G (1996)
Diffusion tensor MR imaging of the human brain. Radiology
201:637–648
Piras F, Caltagirone C, Spalletta G (2010) Working memory
performance and thalamus microstructure in healthy subjects.
Neuroscience 171:496–505
Piras F, Cherubini A, Caltagirone C, Spalletta G (2011) Education
mediates microstructural changes in bilateral hippocampus. Hum
Brain Mapp 32:282–289
Pompili M, Rihmer Z, Akiskal HS, Innamorati M, Iliceto P, Akiskal
KK, Lester D, Narciso V, Ferracuti S, Tatarelli R, De Pisa E,
Girardi P (2008) Temperament and personality dimensions in
suicidal and nonsuicidal psychiatric inpatients. Psychopathology
41:313–321
Reimold M, Batra A, Knobel A, Smolka MN, Zimmer A, Mann K,
Solbach C, Reischl G, Schwarzler F, Grunder G, Machulla HJ,
Bares R, Heinz A (2008) Anxiety is associated with reduced
central serotonin transporter availability in unmedicated patients
with unipolar major depression : a [11C]DASB PET study. Mol
Psychiatry 13(606–613):557
Richter J, Brandstrom S (2009) Personality disorder diagnosis by
means of the temperament and character inventory. Compr
Psychiatry 50:347–352
Rilling JK, Goldsmith DR, Glenn AL, Jairam MR, Elfenbein HA,
Dagenais JE, Murdock CD, Pagnoni G (2008) The neural
correlates of the affective response to unreciprocated coopera-
tion. Neuropsychologia 46:1256–1266
Rilling JK, Sanfey AG, Aronson JA, Nystrom LE, Cohen JD (2004)
Opposing BOLD responses to reciprocated and unreciprocated
altruism in putative reward pathways. NeuroReport
15:2539–2543
Brain Struct Funct
123

Robbins TW, Everitt BJ (1996) Neurobehavioural mechanisms of
reward and motivation. Curr Opin Neurobiol 6:228–236
Rubia K, Smith AB, Woolley J, Nosarti C, Heyman I, Taylor E,
Brammer M (2006) Progressive increase of frontostriatal brain
activation from childhood to adulthood during event-related
tasks of cognitive control. Hum Brain Mapp 27:973–993
Seger CA (2008) How do the basal ganglia contribute to categori-
zation? Their roles in generalization response selection and
learning via feedback. Neurosci Biobehav Rev 32:265–278
Selvaraj S, Murthy NV, Bhagwagar Z, Bose SK, Hinz R, Grasby PM,
Cowen PJ (2011) Diminished brain 5-HT transporter binding in
major depression: a positron emission tomography study with
[11C]DASB. Psychopharmacology 213:555–562
Seymour B, O’Doherty JP, Dayan P, Koltzenburg M, Jones AK,
Dolan RJ, Friston KJ, Frackowiak RS (2004) Temporal differ-
ence models describe higher-order learning in humans. Nature
429:664–667
Smith SM, Jenkinson M, Woolrich MW, Beckmann CF, Behrens TE,
Johansen-Berg H, Bannister PR, De Luca M, Drobnjak I, Flitney
DE, Niazy RK, Saunders J, Vickers J, Zhang Y, De Stefano N,
Brady JM, Matthews PM (2004) Advances in functional and
structural MR image analysis and implementation as FSL.
Neuroimage 23:S208–S219
Stallings MC, Hewitt JK, Cloninger CR, Heath AC, Eaves LJ (1996)
Genetic and environmental structure of the tridimensional
personality questionnaire: three or four temperament dimen-
sions? J Pers Soc Psychol 70:127–140
Storey JD, Taylor JE, Siegmund D (2004) Strong control, conserva-
tive point estimation and simultaneous conservative consistency
of false discovery rates: a unified approach. J R Stat Soc
B66:187–205
Storey JD, Tibshirani RJ (2003) Statistical significance for genome-
wide studies. Proc Natl Acad Sci USA 100:9440–9445
Sykova E, Nicholson C (2008) Diffusion in brain extracellular space.
Physiol Rev 88:1277–1340
Sykova E (2004) Diffusion properties of the brain in health and
disease. Neurochem Int 45:453–466
Tellegen A (1985) Structures of mood and personality and their
relevance to assessing anxiety with an emphasis on self-report.
In: Tuma A, Maser J (eds) Anxiety and the anxiety disorders.
Erlbaum, Hillsdale NJ
Tuominen L, Salo J, Hirvonen J, Nagren K, Laine P, Melartin T,
Isometsa E, Viikari J, Cloninger CR, Raitakari O, Hietala J,
Keltikangas-Jarvinen L (2012) Temperament, character and
serotonin activity in the human brain: a positron emission
tomography study based on a general population cohort. Psychol
Med 31:1–14
Utter AA, Basso MA (2008) The basal ganglia: an overview of
circuits and function. Neurosci Biobehav Rev 32:333–342
van den Heuvel OA, van der Werf YD, Verhoef KM, de Wit S,
Berendse HW, Wolters EC, Veltman DJ, Groenewegen HJ
(2010) Frontal-striatal abnormalities underlying behaviours in
the compulsive-impulsive spectrum. J Neurol Sci 289:55–59
van der Meer M, Kurth-Nelson Z, Redish AD (2012) Information
processing in decision-making systems. Neuroscientist 18:
342–359
Watson D, Clark LA (1993) Behavioral disinhibition versus con-
straint: a dispositional perspective. In: Wegner D, Pennebaker J
(eds) Handbook of mental control. Prentice Hall, New York,
pp 506–527
Westlye LT, Bjørnebekk A, Grydeland H, Fjell AM, Walhovd KB
(2011) Linking an anxiety-related personality trait to brain white
matter microstructure: diffusion tensor imaging and harm
avoidance. Arch Gen Psychiatry 68:369–377
Wickens JR, Budd CS, Hyland BI, Arbuthnott GW (2007) Striatal
contributions to reward and decision making: making sense of
regional variations in a reiterated processing matrix. Ann NY
Acad Sci 1104:192–212
Wise RA (2006) Role of brain dopamine in food reward and
reinforcement. Philos Trans R Soc Lond B Biol Sci
361:1149–1158
Wittmann BC, Daw ND, Seymour B, Dolan RJ (2008) Striatal
activity underlies novelty-based choice in humans. Neuron
58:967–973
Brain Struct Funct
123


















