
Life-history effects of arsenic toxicity in clades of the earthworm Lumbricus rubellus
8
Life-history effects of arsenic toxicity in clades of the earthworm Lumbricus rubellus C.J. Anderson a, b , P. Kille b , A.J. Lawlor c , D.J. Spurgeon a, * a Centre of Ecology and Hydrology, MacLean Building, Benson Lane, Crowmarsh Gifford, Wallingford OX10 8BB, United Kingdom b Cardiff University, School of Biosciences, Cardiff CF10 3AT, United Kingdom c Centre of Ecology and Hydrology, Lancaster Environment Centre, Bailrigg, Lancaster LA1 4AP 8BB, United Kingdom article info Article history: Received 5 June 2012 Received in revised form 6 September 2012 Accepted 8 September 2012 Keywords: Population growth rate EC 50 Genotype Bioaccumulation abstract Exposures of Lumbricus rubellus to a series of arsenic concentrations in soil were used to assess life-stage (juvenile, adult) and genotype specific sensitivities, to calculate population growth rate (l) and to assess patterns of As accumulation. Significant mortality was seen in juveniles at 125 mg/kg As, while growth and maturation was affected from 36 mg/kg and above. In adults, cocoon production at the highest concentration (125 mg/kg) was significantly reduced. Phylogenetic analysis was performed by compar- ison of mitochondrial sequences to establish genotypic variation among juveniles. Three clades with more than 7.5% divergent were described, with 70% of earthworms belonging to a single clade. Date of and mass at maturation was significantly different between clades, but clades were not differentially As sensitive. Parameter l was reduced at 36 mg/kg As and was negative at 125 mg/kg As, suggesting impacts and population stability and potential extinction at environmentally relevant concentrations. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction The long-lasting, deleterious effects of metal contamination have been the subject of many ecotoxicological analyses (Walker et al., 2006). Arsenic (As) is one of several trace elements that has contributed towards aquatic and terrestrial contamination. Gener- ally, soil surface concentrations of As are relatively low (<11 mg/ kg As; Emmett et al., 2010), however, processes such as mineral extraction can lead to localisations that bare enormous concen- trations, ranging from between 2 and 3 orders of magnitude above background levels (Klinck et al., 2005). These soils are capable of enforcing a significant toxic burden upon natural populations. Consequently, As has been the subject of many environmental investigations and has more recently been recognised as a contemporary toxicant of concern for humans such as in Asia (Lu and Zhang, 2005; Stroud et al., 2011). Earthworms are sentinels for terrestrial systems on account of their definitive ecological roles. Consequently they have been widely used in ecotoxicological studies attempting to resolve the deleterious effects associated with metal exposure (Lowe and Butt, 2007). Previous investigations involving the species Lumbricus rubellus, have attempted to resolve the effects of As on specific life- history parameters (Langdon et al., 2003). Basic toxicological data has established that adult earthworms lose condition when exposed to 2000 mg/kg As (Langdon et al., 2009, 1999), contrib- uting to significant levels of mortality in laboratory exposures. Exposures to As-contaminated field soils have also been shown to cause weight loss in exposed earthworms (Shin et al., 2007). Such single endpoint analyses are vital for determining the short-term ecological implications of exposure, however, recognising multiple traits affected by perturbation can provide additional insight into population level impacts. Integration of toxic effects on multiple traits has been proposed as a more appropriate means of extrapolating endpoints to better describe population responses to exposure (Forbes et al., 2008). Population growth rate, in particular, has been found to provide good insight into long-term population level susceptibility (Klok et al., 2006; Kooijman, 2000). For earthworms, Spurgeon et al. (2003) developed an approach to effect assessment that allows for the comparison of toxic effects between different life-stages. Applying the stratified testing approach revealed that juveniles are particularly sensitive to cadmium and copper toxicity (Spurgeon et al., 2004). Furthermore, reduced cocoon production as a result of exposure has also been linked to delayed maturation (Spurgeon and Hopkin, 1996). These examples highlight the value of integrating sublethal endpoints within a demographic context. * Corresponding author. E-mail address: [email protected] (D.J. Spurgeon). Contents lists available at SciVerse ScienceDirect Environmental Pollution journal homepage: www.elsevier.com/locate/envpol 0269-7491/$ e see front matter Ó 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.envpol.2012.09.005 Environmental Pollution 172 (2013) 200e207
-
Upload
cj-anderson -
Category
Documents
-
view
212 -
download
0
Transcript of Life-history effects of arsenic toxicity in clades of the earthworm Lumbricus rubellus

at SciVerse ScienceDirect
Environmental Pollution 172 (2013) 200e207
Contents lists available
Environmental Pollution
journal homepage: www.elsevier .com/locate/envpol
Life-history effects of arsenic toxicity in clades of the earthworm Lumbricusrubellus
C.J. Anderson a,b, P. Kille b, A.J. Lawlor c, D.J. Spurgeon a,*
aCentre of Ecology and Hydrology, MacLean Building, Benson Lane, Crowmarsh Gifford, Wallingford OX10 8BB, United KingdombCardiff University, School of Biosciences, Cardiff CF10 3AT, United KingdomcCentre of Ecology and Hydrology, Lancaster Environment Centre, Bailrigg, Lancaster LA1 4AP 8BB, United Kingdom
a r t i c l e i n f o
Article history:Received 5 June 2012Received in revised form6 September 2012Accepted 8 September 2012
Keywords:Population growth rateEC50GenotypeBioaccumulation
* Corresponding author.E-mail address: [email protected] (D.J. Spurgeon).
0269-7491/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.envpol.2012.09.005
a b s t r a c t
Exposures of Lumbricus rubellus to a series of arsenic concentrations in soil were used to assess life-stage(juvenile, adult) and genotype specific sensitivities, to calculate population growth rate (l) and to assesspatterns of As accumulation. Significant mortality was seen in juveniles at 125 mg/kg As, while growthand maturation was affected from 36 mg/kg and above. In adults, cocoon production at the highestconcentration (125 mg/kg) was significantly reduced. Phylogenetic analysis was performed by compar-ison of mitochondrial sequences to establish genotypic variation among juveniles. Three clades withmore than 7.5% divergent were described, with 70% of earthworms belonging to a single clade. Date ofand mass at maturation was significantly different between clades, but clades were not differentially Assensitive. Parameter l was reduced at 36 mg/kg As and was negative at 125 mg/kg As, suggesting impactsand population stability and potential extinction at environmentally relevant concentrations.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The long-lasting, deleterious effects of metal contaminationhave been the subject of many ecotoxicological analyses (Walkeret al., 2006). Arsenic (As) is one of several trace elements that hascontributed towards aquatic and terrestrial contamination. Gener-ally, soil surface concentrations of As are relatively low (<11 mg/kg As; Emmett et al., 2010), however, processes such as mineralextraction can lead to localisations that bare enormous concen-trations, ranging from between 2 and 3 orders of magnitude abovebackground levels (Klinck et al., 2005). These soils are capable ofenforcing a significant toxic burden upon natural populations.Consequently, As has been the subject of many environmentalinvestigations and has more recently been recognised asa contemporary toxicant of concern for humans such as in Asia (Luand Zhang, 2005; Stroud et al., 2011).
Earthworms are sentinels for terrestrial systems on account oftheir definitive ecological roles. Consequently they have beenwidely used in ecotoxicological studies attempting to resolve thedeleterious effects associated with metal exposure (Lowe and Butt,2007). Previous investigations involving the species Lumbricus
All rights reserved.
rubellus, have attempted to resolve the effects of As on specific life-history parameters (Langdon et al., 2003). Basic toxicological datahas established that adult earthworms lose condition whenexposed to 2000 mg/kg As (Langdon et al., 2009, 1999), contrib-uting to significant levels of mortality in laboratory exposures.Exposures to As-contaminated field soils have also been shown tocause weight loss in exposed earthworms (Shin et al., 2007). Suchsingle endpoint analyses are vital for determining the short-termecological implications of exposure, however, recognisingmultiple traits affected by perturbation can provide additionalinsight into population level impacts.
Integration of toxic effects on multiple traits has been proposedas a more appropriate means of extrapolating endpoints to betterdescribe population responses to exposure (Forbes et al., 2008).Population growth rate, in particular, has been found to providegood insight into long-term population level susceptibility (Kloket al., 2006; Kooijman, 2000). For earthworms, Spurgeon et al.(2003) developed an approach to effect assessment that allowsfor the comparison of toxic effects between different life-stages.Applying the stratified testing approach revealed that juvenilesare particularly sensitive to cadmium and copper toxicity(Spurgeon et al., 2004). Furthermore, reduced cocoon production asa result of exposure has also been linked to delayed maturation(Spurgeon and Hopkin, 1996). These examples highlight the valueof integrating sublethal endpoints within a demographic context.

C.J. Anderson et al. / Environmental Pollution 172 (2013) 200e207 201
Here we utilise the approach of Spurgeon et al. (2003) to quantifythe toxic effects of As for a range of relevant life-cycle traits inL. rubellus.
Although demographic studies integrate effects more compre-hensively than studies concentrating on a single trait, they do notexplicitly acknowledge the role of genetic variation (Calow, 1996).Variation in sensitivity between individuals for a given endpointmay be indicative of genetically distinct cohorts within populationsthat respond differently to the selective pressure of the toxicant(Morgan et al., 2007). This is particularly relevant in L. rubellus, asrecent evidence shows that populations inhabiting the extensivelysurveyed former As and copper mine site at Devon Great Consols(DGC), UK, are able to maintain populations in soils containing inexcess of 8000 mg/kg As (Button et al., 2009a; Langdon et al., 2009,1999). Further, evidence has suggested a high degree of localisedgenetic differentiation that occurs in populations across metalcontaminated sites (Andre et al., 2010). This is suggestive of geno-type specificity in the ability to tolerate exposure and promotes theinclusion of genotypes to identify any possible sources of variationwithin ecotoxicological data (Calow, 1996; Morgan et al., 2007). Inthis study, the influence of genotype (as defined by sequencing ofan established mitochondrial marker) upon L. rubellus life-historyparameters relative to As sensitivity is examined.
2. Materials and methods
Established protocols were used to assess As effects on L. rubellus adult andjuvenile parameters and total tissue As, as well as phylogenetic analysis for juvenileworms. The exposure protocols were based on Spurgeon et al. (2003). The detailedprocedures used are provided in Supplementary Information and are summarisedbelow.
2.1. Test medium
Soils used for both adult and juvenile tests consisted of sieved commerciallyavailable clay loam soil (Broughton Loam, Kettering, UK), amended with 3% com-posted bark (final medium pH 7.1, organic matter content w5%) as previously usedand fullycharacterisedbyHooperet al. (2011) andSpurgeonet al. (2004). Inboth testssodium arsenate (Na2HAsO4$7H2O) was added to give soil concentrations of 0, 3, 12,36, and 125mg/kg, as selected based on literature values (Fischer andKoszorus,1992;Langdon et al., 2002;Meharg et al., 1998; Yeates et al., 1994). Soils were then left for 7days to allow initial speciation equilibration (Spurgeon et al., 2003).
2.2. Adult As exposure
For each of the five concentrations, five replicates, each including five adultworms taken from a culture established from a field-collected population, wereused. During the 28 days of the exposure, the containers were kept at 14 �C inconstant light and fed appropriately As-spiked horse manure as described in theSupplementary Information. At 14 days, mortality, weight change and foodconsumptionwas checked by counting and weighing the earthworms and weighingmanure left on the soil surface. After 28 d, these parameters were again assessed.Further any surviving worms were immediately snap frozen for later elementalanalysis (n.b. worms were not depurated to maintain the integrity of samples forlater metabolomic and gene expression studies (not reported here)). Cocoonproduction rates (cocoons/worm/day) were determined by sieving the test soil andcounting cocoons.
2.3. Juvenile As exposure
For each test concentration 25 replicates were used. Each replicate containeda single exposed hatchling worm. Each was kept for 280 days at 14 �C in constantlight. Survival, weight and developmental status (juvenile, subadult, adult) wererecorded every 28 days over this time. Following completion of the experiment,w10 mg of tissue was removed from the posterior of each individual for latergenotyping using a mitochondrial marker.
2.4. Juvenile genotyping
DNA was then extracted using the QIAamp DNA mini kit (Qiagen) prior topolymerase chain reaction (PCR) amplification according to Andre et al. (2010) usingCOII forward (TAGCTCACTTAGATGCCA) and reverse (GTATGCGGATTTCTAATTGT).PCR products were checked by gel electrophoresis and purified for sequencing using0.25 U each of Exonuclease I and Shrimp Alkaline Phosphatase incubated at 37 �C for
45 min and 80 �C for 15 min to inactivate the reagents. Purified PCR products werethen sequenced as in Andre et al. (2010), using ABI PRISM� BigDye v3.1 TerminatorSequencing technology (Applied Biosystems, USA).
Sequences were aligned and trimmed for tree construction using the MaximumLikelihood method using the General Time Reversible model with a gamma distri-bution Mega (v5.01). Tree topology was supported by bootstrap analyses over 1000iterations and the use of outlier sequences from the other lumbricid species.Sequences for L. rubellus associated with specific mitochondrial clades alreadydocumented in the UK were incorporated as anchor sequences.
2.5. Soil As analysis
Approximately 5gof soilwere removed fromfive replicates of each concentrationfrom both the adult and juvenile toxicity tests following termination of the exposure.Soilsweredried for 48h at 80 �Cbefore beingpassed througha2mmsieve. Soilswereanalysed for total As concentration as described in Spurgeon et al. (2011). Briefly,following aqua regia digestion using a microwave system, samples were quantifiedusing a Perkin Elmer Elan DRCII inductively coupled plasmamass spectrometer (ICP-MS) (Perkin Elmer 4300DV). Quality controlwithin the analyseswas conducted usingthe standard reference material ISE 192 (International Soil Exchange, TheNetherlands). Certified values for reference materials corroborated well with data inthis analysis, beingwithin 10% ofmean certified concentration for As concentrations.
2.6. Tissue metal analysis
Metal tissue analysis was conducted for worms exposed in the adult test usinghomogenised tissue derived from all surviving worms within each replicate. Wormswere not depurated following the exposure. These samples were analysed for totalAs concentration using 0.5 g of dried sample digested with 10 ml of 70% HNO3(Ultrapure) at 200 �C for 15 min within a microwave digestion vessel. Samples wererun on the Perkin Elmer DRCII ICP-MS. Each run included the certified referencematerials TORT-2 and DOLT-4 (National Research Council, Canada) for qualitycontrol. Certified values for reference materials corroborated well with data in thisanalysis, which were within 10% of certified values for As.
2.7. Statistics
Effects on mortality, weight change, cocoon production and juvenile maturationwere checked for normality using the KolmogoroveSmirnov statistic and thenfurther analysed using analysis of variance (ANOVA) in Minitab (v15) (a ¼ 0.05).Cocoon production was corrected for mortality events, thereby allowing cocoonproduction rate (cocoons/worm/week) estimation. Tukey’s testwasused for thepost-hoc pairwise comparison of cocoon production rates between the different treat-ments. An LC50 for juvenile L. rubellus exposed to Aswas calculated using the survivaldata at 280daysusing the trimmedSpearmaneKarbermethod (Hamilton et al.,1977).The EC50 for effects on cocoon production was calculated by fitting cocoon produc-tion rate data for each replicate using a logistic model in Sigmaplot (v10.0).
The patterns of juvenile growth relative to exposure concentration were fittedusing a Gompertz model in SigmaPlot 12.0 using meanmass values for the samplingintervals throughout the exposure. This model is summarised as below.
Wt ¼ Aþ Ce�e�Bðt�MÞ
where Wt is the weight at time t; A the lower asymptotic weight (g), C the upperasymptotic weight (g), M the age at the inflection point (d), and B is the Gompertzgrowth constant (days�1). From these parameters, maximum growth (at the point ofinflection) was calculated using the equation,
BC�e�1
The Gompertz model has been shown to provide a good description of thepatterns of earthworm growth following trace metal exposure (Spurgeon et al.,2004). The influence of haplotype variation on both the length of the juvenileperiod and size at maturation under different As treatments was assessed usinga General Linear Model (GLM), with sequenced haplotype and As concentration asthe fixed factors including the combined factor also within the analysis. Individualsthat did not mature were not included in this analysis.
2.8. Population model
To integrate the toxic effects of As on multiple life-cycle traits, life-history datawas used to predict the intrinsic rate of increase (l) using separately parameterisedmodels for each test concentration. Stage-structured matrix population modelswere used as they are mathematically manageable and carry enough biologicalinformation for risk assessment (Caswell, 2001; Forbes et al., 2011; Neubert andCaswell, 2000). A three-stage matrix population model (Fig. 1) was used. Thisincorporates (i) a cocoon stage (during which early development of the youngworms occurs) inwhich worms either stay in cocoons; hatch to become juveniles ordie; (ii) a juvenile stage during which worms may stay juvenile; mature to adults or

Fig. 1. A stage-structured life-cycle model for L. rubellus, where nodes 1, 2 and 3represent cocoon, juvenile and adult life-stages, respectively. Pi is the likelihood ofremaining as the current stage, Gi is the probability of progression to the next stage andF3 is cocoon production/worm/week, where subscipt i is life-stage. The probability ofcocoon survival and progression is fixed from literature data (see text).
Fig. 2. Cocoon production rate of earthworms (L. rubellus) exposed to a range ofconcentrations of As for 28 days. Points show cocoon production rate for individualreplicate containers (cocoons/worm/week), dashed line shows fitted logistic model.
C.J. Anderson et al. / Environmental Pollution 172 (2013) 200e207202
die; and (iii) an adult stage in which individuals either remain as adults producingcocoon at a given rate, or are removed from the population. A value for l can then becalculated as the dominant eigenvalue of this projection matrix.
To parameterise the models for each of the nominal concentrations used in thetoxicity test (0, 3, 12, 36,125mg/kg As), separate 3� 3matrices were populatedwithvalues describing these stage specific values and transitions based on prior knowl-edge and the experimental data. For the cocoon stage, experimental data was notavailable and so previous cocoon hatching information was used to provide defaultvalues to be used for all test concentrations. Cocoons were assumed to hatch after 56days, with 80% producing a single hatchling per cocoon (Kammenga et al., 2003).This simplification may result in underestimation of population growth rates if Asexposure has an effect on the developing juvenile.
For the remaining values, the matrices were parameterised using experimentaldata for each life-stage specific for each nominal soil As concentration in the adultand juvenile tests. The duration of adult exposure corresponded to the projectioninterval of the model (28 d), therefore adult survival and cocoon production rateswere used directly as parameters. Juvenile parameter values were derived from themeasurement of survival to adulthood and maturation time relative to exposure.Error measurements for l were estimated via parameter estimates and associatedstandard deviation data over 1000 iterations.
3. Results
3.1. Soil physico-chemistry
Background As concentrations in uncontaminated soil were16.5 � 0.6 mg/kg. Negating background concentrations by sub-tracting from totals indicated that measured values were within15% of nominal concentrations. For simplicity, concentrations arereferred to according to nominal concentrations throughout thistext, although measured concentrations were used for the LC50and cocoon production EC50 calculations.
3.2. Adult and juvenile mortality
There was good survival of worms in the Adult toxicity test, withno significant reductions found relative to exposure, as determinedby ANOVA (P > 0.05). In controls 88% of worms survived theexperiment, falling to only 80% among worms at the highestconcentration used (125 mg/kg). In the juvenile test, survival ratesover the experiment were much lower in comparison to those inthe adult test. ANOVA indicated a significant effect of As concen-tration on juvenile survival to 280 days (P < 0.001). This reflectsa stark contrast, whereby all worms in control conditions remainedalive throughout the experiment, but survival was reduced to 12%at 125 mg/kg As. Using survival data of juveniles at 280 days, anLC50 of 67mg/kg (95% confidence interval¼ 57.44e78.35) could beestimated for this life-stage.
3.3. Cocoon production
Cocoon production rates in control soils were in accordancewith the results of previous work. For example, Klok (2007) found
an overall mean of 0.51 � 0.07 cocoons/earthworm/day whenanalysing the effect of density upon reproduction in unpollutedsoils. This compares to the rates of 0.7 � 0.3 seen in the controlpopulation in this study. Reduced cocoon production was seen inadult worms exposed to the higher soil As concentrations (Fig. 2).ANOVA confirmed that cocoon production was significantlyaffected by As exposure (ANOVA P < 0.001), with post-hocassessment using Tukey’s test indicating that cocoon productionwas significantly reduced at 125 mg/kg, with all remaining treat-ments not significantly different from the control. The strongdecline in rates of cocoon production observed resulted in a logisticmodel that described a steep reduction in reproduction betweenthe two highest treatment concentrations. On the basis of this fit(R2 ¼ 0.57) an EC50 of 118 mg/kg As (95% CI not available) could beestimated.
3.4. Adult earthworm tissue As concentrations
Following log-transformation to ensure normality of the data,earthworm tissue As concentrations significantly increasedrelative to soil concentrations (ANOVA P < 0.001) (Table 1).Post-hoc comparison indicated that worms exposed to soil Asconcentrations of 12, 36, and 125 mg/kg had significantly highertissue As concentrations (2.0 � 0.5, 5.9 � 0.7, and 26.1 � 4.06,respectively) compared to worms exposed to the control soil(0.8 � 0.2 mg/kg As).
3.5. Growth and maturation
Juvenile L. rubellus mass was measured at 28 d intervals overa 280 d exposure. Fig. 3 demonstrates that higher soil As concen-trations resulted in reduced mean growth rate and final body size.Worms exposed to the higher concentrations were significantlyaffected (ANOVA P< 0.001), reaching only 0.084 g by the end of theexposure, as compared to 1.8 g in control worms. Post-hoc analysesfurther demonstrated that worms exposed to 36, as well as 125mg/kg, were significantly smaller after 280 days than worms in thecontrol and lower concentration treatments.
The temporal patterns of juvenile weight change were fittedusing Gompertz curves parameterised using mean mass values
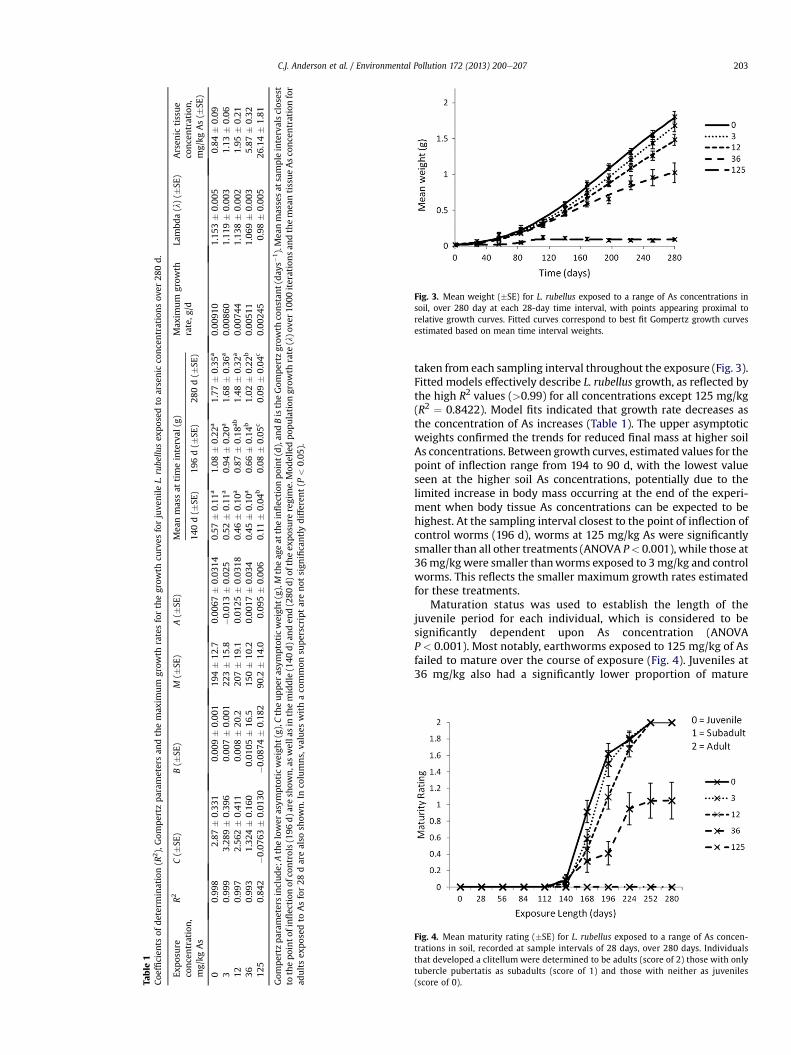
Fig. 3. Mean weight (�SE) for L. rubellus exposed to a range of As concentrations insoil, over 280 day at each 28-day time interval, with points appearing proximal torelative growth curves. Fitted curves correspond to best fit Gompertz growth curvesestimated based on mean time interval weights.
Table
1Coe
fficien
tsof
determination(R
2),Gom
pertz
param
etersan
dthemax
imum
grow
thratesforthegrow
thcu
rves
forjuve
nile
L.rube
llusex
posedto
arsenic
concentrationsov
er28
0d.
Exposure
concentration,
mg/kg
As
R2
C(�
SE)
B(�
SE)
M(�
SE)
A(�
SE)
Mea
nmassat
timeinterval
(g)
Max
imum
grow
thrate,g
/dLambd
a(l)(�
SE)
Arsen
ictissue
concentration,
mg/kg
As(�
SE)
140d(�
SE)
196d(�
SE)
280d(�
SE)
00.99
82.87
�0.33
10.00
9�
0.00
119
4�
12.7
0.00
67�
0.03
140.57
�0.11
a1.08
�0.22
a1.77
�0.35
a0.00
910
1.15
3�
0.00
50.84
�0.09
30.99
93.28
9�
0.39
60.00
7�
0.00
122
3�
15.8
�0.013
�0.02
50.52
�0.11
a0.94
�0.20
a1.68
�0.36
a0.00
860
1.11
9�
0.00
31.13
�0.06
120.99
72.56
2�
0.41
10.00
8�
20.2
207�
19.1
0.01
25�
0.03
180.46
�0.10
a0.87
�0.18
ab1.48
�0.32
a0.00
744
1.13
8�
0.00
21.95
�0.21
360.99
31.32
4�
0.16
00.01
05�
16.5
150�
10.2
0.00
17�
0.03
40.45
�0.10
a0.66
�0.14
b1.02
�0.22
b0.00
511
1.06
9�
0.00
35.87
�0.32
125
0.84
2�0
.076
3�
0.01
30�0
.087
4�
0.18
290
.2�
14.0
0.09
5�
0.00
60.11
�0.04
b0.08
�0.05
c0.09
�0.04
c0.00
245
0.98
�0.00
526
.14�
1.81
Gom
pertz
param
etersinclude;
Athelower
asym
ptoticweigh
t(g),C
theupper
asym
ptoticweigh
t(g),M
theag
eat
theinflection
point(d),an
dBistheGom
pertz
grow
thco
nstan
t(day
s�1).Mea
nmassesat
sample
intervalsclosest
tothepointo
finflection
ofco
ntrols(196
d)a
resh
own,aswella
sin
themiddle
(140
d)a
nden
d(280
d)o
ftheex
posure
regime.Mod
elledpop
ulation
grow
thrate
(l)o
ver10
00iterationsan
dthemea
ntissueAsco
ncentrationfor
adultsex
posed
toAsfor28
darealso
show
n.Inco
lumns,va
lues
withaco
mmon
superscriptarenot
sign
ificantlydifferent(P
<0.05
).
C.J. Anderson et al. / Environmental Pollution 172 (2013) 200e207 203
taken from each sampling interval throughout the exposure (Fig. 3).Fitted models effectively describe L. rubellus growth, as reflected bythe high R2 values (>0.99) for all concentrations except 125 mg/kg(R2 ¼ 0.8422). Model fits indicated that growth rate decreases asthe concentration of As increases (Table 1). The upper asymptoticweights confirmed the trends for reduced final mass at higher soilAs concentrations. Between growth curves, estimated values for thepoint of inflection range from 194 to 90 d, with the lowest valueseen at the higher soil As concentrations, potentially due to thelimited increase in body mass occurring at the end of the experi-ment when body tissue As concentrations can be expected to behighest. At the sampling interval closest to the point of inflection ofcontrol worms (196 d), worms at 125 mg/kg As were significantlysmaller than all other treatments (ANOVA P< 0.001), while those at36mg/kgwere smaller thanworms exposed to 3mg/kg and controlworms. This reflects the smaller maximum growth rates estimatedfor these treatments.
Maturation status was used to establish the length of thejuvenile period for each individual, which is considered to besignificantly dependent upon As concentration (ANOVAP < 0.001). Most notably, earthworms exposed to 125 mg/kg of Asfailed to mature over the course of exposure (Fig. 4). Juveniles at36 mg/kg also had a significantly lower proportion of mature
Fig. 4. Mean maturity rating (�SE) for L. rubellus exposed to a range of As concen-trations in soil, recorded at sample intervals of 28 days, over 280 days. Individualsthat developed a clitellum were determined to be adults (score of 2) those with onlytubercle pubertatis as subadults (score of 1) and those with neither as juveniles(score of 0).

Fig. 5. Maximum likelihood phylogenetic tree of the cytochrome oxidase II mito-chondrial gene of 63 Lumbricus rubellus juveniles exposed to arsenic. Three clades withgreater than 7.5% sequence variation are identifiable and the sequences belonging totwo previously identified genotypes, A and B, have been included as anchors and liewithin clades 3 and 2, respectively. Sequences from L. terrestris and L. castaneus wereused as outliers. The number of individuals pertaining to the same haplotype isidentifiable next to highlighted branches, with bootstrap support indicated above eachnode. Values lower than 70% were condensed.
C.J. Anderson et al. / Environmental Pollution 172 (2013) 200e207204
adults relative to lower exposure concentrations, with only 50% ofsurviving individuals developing a clitellum after 280 days.Earthworms exposed to 0, 3 and 12 mg/kg As were all mature bythe end of exposure, with average maturation times of 207, 211,and 223 days, respectively.
3.6. Effects of As on intrinsic rate of population increase
Values for adult survival and reproduction were taken directlyfrom the results obtained for each tested concentration in the adultexposure. Values for juvenile survival and maturation were calcu-lated from the juvenile test. Earthworms that failed to mature overthe duration of the exposure were assumed to have matured by theputative next sample interval (set at 308 days). This is almostcertainly an unrealistic assumption for worms at 125 mg/kg,therefore estimation of l at this concentration is likely to be lowerthan the given value. Based upon fittedmodels, control populationsare expected to increase with a l of 1.15 (see Table 1 for all values).Values at 3 and 12mg/kg As are similar to that for the controls (1.12and 1.14 respectively). Populations at 36 mg/kg As had a lowerpopulation growth rate (1.07) than controls, though it remainedabove 1, demonstrating reduced stability. At 125 mg/kg As, modelestimates suggest that the population will eventually go extinct,with l at 0.98. Given the severe effects on maturation and theassumption of maturation at 308 days, it may be expected thatextinction may in fact occur at rates greater than estimated by themodel.
3.7. Genetic variation, juvenile development and As sensitivity
Earthworm genotypes were determined by sequencing of COIImitochondrial markers. A Maximum Likelihood model con-structed from the obtained sequences resulted in identical cladeconstruction which identified three major clades demonstratedfurther by the posterior probabilities and bootstrap values (Fig. 5).Some discrepancies were seen within clade 2, although the othertwo clades were well supported (bootstrap values over 70%), withgreater than 7.5% genetic differentiation occurring betweenidentified clades (Hillis and Bull, 1993). Outlying sequences fromLumbricus terrestris and Lumbricus castaneus COII genes were usedto define potential misidentification of early stage juveniles, withan average sequence divergence of 20.2% and 19.6% respectively.Comparison of genetic variation between and within clades, aswell as the frequency of clades in exposure regimes are shown inTable 2. Anchor sequences of two previously recognised Britishlineages (using AFLP and COII sequences) were placed among twodistinct clades, though not among the most numerous clade foundin this study (Andre et al., 2010). Branches with Bootstrap valuesbelow 70% were collapsed.
Considering clade effects, GLM showed that the date at whichindividuals matured was significantly affected by the concentrationof As (F ¼ 3.99, P < 0.05). Further, haplotype was also found to havea significant effect on maturation time (F ¼ 10.06, P < 0.01). Therewas, however, no interaction between the two variables. Post-hoctesting indicated that worms in clade 2 matured at an earlier daterelative to worms from clade 1, which represent the majority ofworms in this experiment. Individuals belonging to clade 3 werenot included in the analysis as only four individuals, occurring ata single exposure concentration were present among the sampledworms. For size at maturation, there was no significant effect ofeither genotype or As exposure concentration (F ¼ 3.2 P > 0.05) onmass at maturation. The low frequency of individuals from clade 2(Table 2), however, indicate that further analysis is required tomorefully document genotype specific variation among life-historyparameters.
4. Discussion
Our analyses demonstrated both acute and sublethal toxicityeffects of As. Life-history stages appear to be differentially sensitive,with effects on traits seen at lower concentrations in tests withjuvenile worms than those occurring within the adult test. Thiseffect could, however, also relate to the shorter exposure durationused in the adult toxicity test. Integration of affected parameterswithin a simple demographicmodel indicated negative effects of Ason intrinsic rate of population increase, which will ultimately resultin population extinction when As is at high concentrations.Significant effects particularly seen in juvenile survival and matu-ration, as well as reduced cocoon production, are mainly respon-sible for this population level effect of As exposure.
In responding to the presence of elevated As concentrations insoil, the iteroparous nature of L. rubellus (Sims and Gerard, 1985) islikely to allow for optimisation for maximum reproductive outputto exploit contemporary circumstances. Lika and Kooijman (2003)stated that indeterminate growers (who keep growing aftermaturation), such as the earthworm, were likely to reduce alloca-tion of resources away from reproduction, towards maintenance incases where individual are exposed to sub-optimal conditions.Widdows and Johnson (1988) pioneered analyses to providea quantitative assessment of the energy status of animals relative togrowth rate, or rather “scope for growth”. Their seminal studydemonstrated reduced growth in the mussel, Mytilus edulis, understress imposed by copper and hydrocarbons, specifically demon-strating this diversion of resources. Dynamic energy budget (DEB)models have been used previously to explore population level
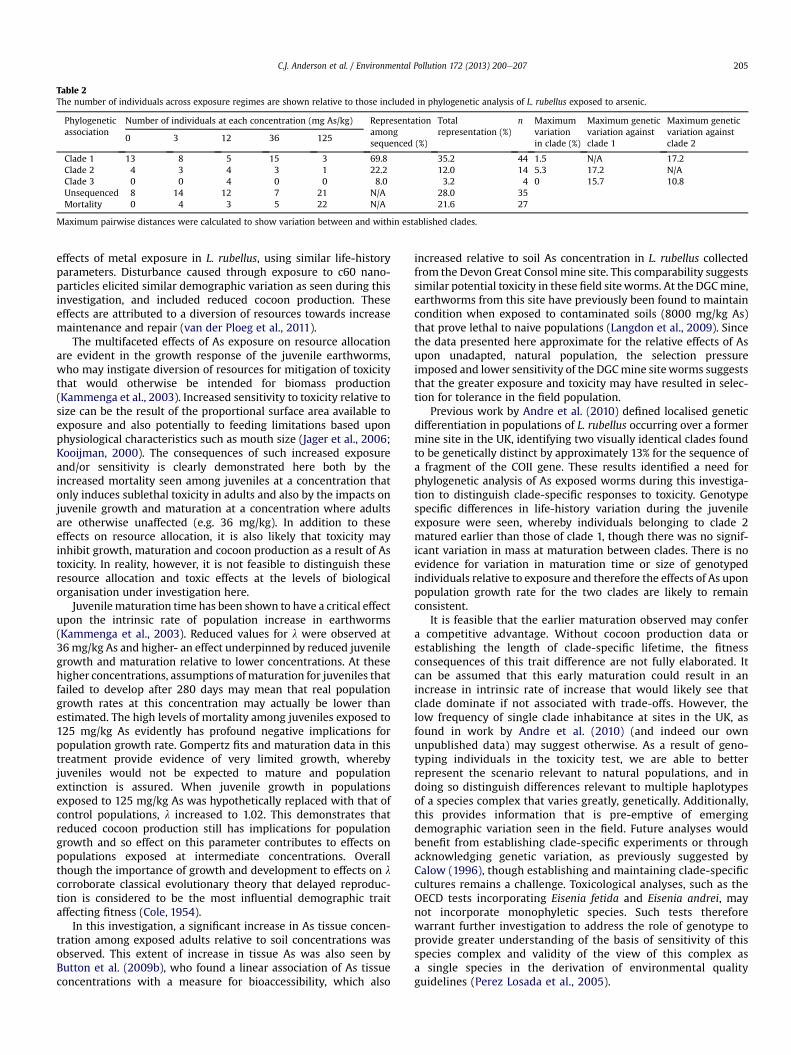
Table 2The number of individuals across exposure regimes are shown relative to those included in phylogenetic analysis of L. rubellus exposed to arsenic.
Phylogeneticassociation
Number of individuals at each concentration (mg As/kg) Representationamongsequenced (%)
Totalrepresentation (%)
n Maximumvariationin clade (%)
Maximum geneticvariation againstclade 1
Maximum geneticvariation againstclade 2
0 3 12 36 125
Clade 1 13 8 5 15 3 69.8 35.2 44 1.5 N/A 17.2Clade 2 4 3 4 3 1 22.2 12.0 14 5.3 17.2 N/AClade 3 0 0 4 0 0 8.0 3.2 4 0 15.7 10.8Unsequenced 8 14 12 7 21 N/A 28.0 35Mortality 0 4 3 5 22 N/A 21.6 27
Maximum pairwise distances were calculated to show variation between and within established clades.
C.J. Anderson et al. / Environmental Pollution 172 (2013) 200e207 205
effects of metal exposure in L. rubellus, using similar life-historyparameters. Disturbance caused through exposure to c60 nano-particles elicited similar demographic variation as seen during thisinvestigation, and included reduced cocoon production. Theseeffects are attributed to a diversion of resources towards increasemaintenance and repair (van der Ploeg et al., 2011).
The multifaceted effects of As exposure on resource allocationare evident in the growth response of the juvenile earthworms,who may instigate diversion of resources for mitigation of toxicitythat would otherwise be intended for biomass production(Kammenga et al., 2003). Increased sensitivity to toxicity relative tosize can be the result of the proportional surface area available toexposure and also potentially to feeding limitations based uponphysiological characteristics such as mouth size (Jager et al., 2006;Kooijman, 2000). The consequences of such increased exposureand/or sensitivity is clearly demonstrated here both by theincreased mortality seen among juveniles at a concentration thatonly induces sublethal toxicity in adults and also by the impacts onjuvenile growth and maturation at a concentration where adultsare otherwise unaffected (e.g. 36 mg/kg). In addition to theseeffects on resource allocation, it is also likely that toxicity mayinhibit growth, maturation and cocoon production as a result of Astoxicity. In reality, however, it is not feasible to distinguish theseresource allocation and toxic effects at the levels of biologicalorganisation under investigation here.
Juvenilematuration time has been shown to have a critical effectupon the intrinsic rate of population increase in earthworms(Kammenga et al., 2003). Reduced values for l were observed at36mg/kg As and higher- an effect underpinned by reduced juvenilegrowth and maturation relative to lower concentrations. At thesehigher concentrations, assumptions of maturation for juveniles thatfailed to develop after 280 days may mean that real populationgrowth rates at this concentration may actually be lower thanestimated. The high levels of mortality among juveniles exposed to125 mg/kg As evidently has profound negative implications forpopulation growth rate. Gompertz fits and maturation data in thistreatment provide evidence of very limited growth, wherebyjuveniles would not be expected to mature and populationextinction is assured. When juvenile growth in populationsexposed to 125 mg/kg As was hypothetically replaced with that ofcontrol populations, l increased to 1.02. This demonstrates thatreduced cocoon production still has implications for populationgrowth and so effect on this parameter contributes to effects onpopulations exposed at intermediate concentrations. Overallthough the importance of growth and development to effects on l
corroborate classical evolutionary theory that delayed reproduc-tion is considered to be the most influential demographic traitaffecting fitness (Cole, 1954).
In this investigation, a significant increase in As tissue concen-tration among exposed adults relative to soil concentrations wasobserved. This extent of increase in tissue As was also seen byButton et al. (2009b), who found a linear association of As tissueconcentrations with a measure for bioaccessibility, which also
increased relative to soil As concentration in L. rubellus collectedfrom the Devon Great Consol mine site. This comparability suggestssimilar potential toxicity in these field siteworms. At the DGCmine,earthworms from this site have previously been found to maintaincondition when exposed to contaminated soils (8000 mg/kg As)that prove lethal to naive populations (Langdon et al., 2009). Sincethe data presented here approximate for the relative effects of Asupon unadapted, natural population, the selection pressureimposed and lower sensitivity of the DGCmine siteworms suggeststhat the greater exposure and toxicity may have resulted in selec-tion for tolerance in the field population.
Previous work by Andre et al. (2010) defined localised geneticdifferentiation in populations of L. rubellus occurring over a formermine site in the UK, identifying two visually identical clades foundto be genetically distinct by approximately 13% for the sequence ofa fragment of the COII gene. These results identified a need forphylogenetic analysis of As exposed worms during this investiga-tion to distinguish clade-specific responses to toxicity. Genotypespecific differences in life-history variation during the juvenileexposure were seen, whereby individuals belonging to clade 2matured earlier than those of clade 1, though there was no signif-icant variation in mass at maturation between clades. There is noevidence for variation in maturation time or size of genotypedindividuals relative to exposure and therefore the effects of As uponpopulation growth rate for the two clades are likely to remainconsistent.
It is feasible that the earlier maturation observed may confera competitive advantage. Without cocoon production data orestablishing the length of clade-specific lifetime, the fitnessconsequences of this trait difference are not fully elaborated. Itcan be assumed that this early maturation could result in anincrease in intrinsic rate of increase that would likely see thatclade dominate if not associated with trade-offs. However, thelow frequency of single clade inhabitance at sites in the UK, asfound in work by Andre et al. (2010) (and indeed our ownunpublished data) may suggest otherwise. As a result of geno-typing individuals in the toxicity test, we are able to betterrepresent the scenario relevant to natural populations, and indoing so distinguish differences relevant to multiple haplotypesof a species complex that varies greatly, genetically. Additionally,this provides information that is pre-emptive of emergingdemographic variation seen in the field. Future analyses wouldbenefit from establishing clade-specific experiments or throughacknowledging genetic variation, as previously suggested byCalow (1996), though establishing and maintaining clade-specificcultures remains a challenge. Toxicological analyses, such as theOECD tests incorporating Eisenia fetida and Eisenia andrei, maynot incorporate monophyletic species. Such tests thereforewarrant further investigation to address the role of genotype toprovide greater understanding of the basis of sensitivity of thisspecies complex and validity of the view of this complex asa single species in the derivation of environmental qualityguidelines (Perez Losada et al., 2005).

C.J. Anderson et al. / Environmental Pollution 172 (2013) 200e207206
5. Conclusion
Soil As concentrations of 125 mg/kg are estimated to evokepopulation extinction in the earthworm, L. rubellus through affectson juvenile and adult life-history parameters. Significant mortalitywas observed among juveniles, although not among adults, whichproduced fewer cocoons in response to exposure. Juvenile growthand maturation was significantly affected at the highest Asconcentrations tested, notably at 36 mg/kg, suggesting increasedsensitivity of juvenile compared to adults, which were notadversely affected at this concentration for measured traits. Wealso suggest that deviation of resources to manage toxicity may beresponsible for the sublethal perturbation seen in this investiga-tion, corroborating with previous studies. Phylogenetic analyses ofjuveniles established three genetically distinct clades, two of whichwere found to vary in time to maturation, but did not showphenotypic variation relative to As exposure. This investigationfurther resolves genetic complexity of L. rubellus and providesphenotypic anchors for molecular endpoints. The clades presentwithin L. rubellus also indicate the presence of substantial geneticvariation that may be the basis of adaptive traits.
Acknowledgements
CA was supported by the Natural Environment ResearchCouncil Environment Doctoral Training Grant Reference: NE/G523571/1. We thank Dr. Claus Svendsen and Dr. Helen Hooperfor advice and assistance during the experimental design andtesting work.
Appendix A. Supplementary information
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.envpol.2012.09.005
References
Andre, J., King, R.A., Stürzenbaum, S.R., Kille, P., Hodson, M.E., Morgan, A.J., 2010.Molecular genetic differentiation in earthworms inhabiting a heterogeneousPb-polluted landscape. Environmental Pollution 158 (3), 883e890.
Button, M., Jenkin, G.R.T., Harrington, C.F., Watts, M.J., 2009a. Arsenic biotransfor-mation in earthworms from contaminated soils. Journal of EnvironmentalMonitoring 11 (8), 1484e1491.
Button, M., Watts, M.J., Cave, M.R., Harrington, C.F., Jenkin, G.T., 2009b. Earthwormsand in vitro physiologically-based extraction tests: complementary tools fora holistic approach towards understanding risk at arsenic-contaminated sites.Environmental Geochemistry and Health 31 (2), 273e282.
Calow, P., 1996. Variability: noise or information in ecotoxicology? EnvironmentalToxicology and Pharmacology 2 (2e3), 121e123.
Caswell, H., 2001. Matrix Population Models: Construction, Analysis, and Inter-pretation. Sinauer Associates, Sunderland, MA, USA.
Cole, L.C., 1954. The population consequences of life history phenomena. QuarterlyReview of Biology 29, 103e137.
Emmett, B.A., Reynolds, B., Chamberlain, P.M., Rowe, E., Spurgeon, D., Brittain, S.A.,Frogbrook, Z., Hughes, S., Lawlor, A.J., Poskitt, J., et al., 2010. 2010 CountrysideSurvey: Soils Report from 2007. Technical Report No. 9/07 NERC. Centre forEcology & Hydrology, 192 pp.
Fischer, E., Koszorus, L., 1992. Sublethal effects, accumulation capacities and elim-ination rates of As, Hg and Se in the manure worm, Eisenia-foetida (Oligochaeta,Lumbricidae). Pedobiologia 36 (3), 172e178.
Forbes, V.E., Calow, P., Grimm, V., Hayashi, T.I., Jager, T., Katholm, A., Palmqvist, A.,Pastorok, R., Salvito, D., Sibly, R., et al., 2011. Adding value to ecological riskassessment with population modeling. Human and Ecological Risk Assessment17 (2), 287e299.
Forbes, V.E., Calow, P., Sibly, R.M., 2008. The extrapolation problem and how pop-ulation modeling can help. Environmental Toxicology and Chemistry 27 (10),1987e1994.
Hamilton, M.A., Russo, R.C., Thurston, R.V., 1977. Trimmed SpearmaneKarbermethod for estimating median lethal concentrations in toxicity bioassays.Environmental Science & Technology 11 (7), 714e719.
Hillis, D.M., Bull, J.J., 1993. An empirical test of bootstrapping as a method forassessing confidence in phylogenetic analysis. Systematic Biology 42 (2),182e192.
Hooper, H.L., Jurkschat, K., Morgan, A.J., Bailey, J., Lawlor, A.J., Spurgeon, D.J.,Svendsen, C., 2011. Comparative chronic toxicity of nanoparticulate and ioniczinc to the earthworm Eisenia veneta in a soil matrix. Environment Interna-tional 37 (6), 1111e1117.
Jager, T., Heugens, E.H.W., Kooijman, S.A.L.M., 2006. Making sense of ecotoxico-logical test results: towards application of process-based models. Ecotoxicology15 (3), 305e314.
Kammenga, J.E., Spurgeon, D.J., Svendsen, C., Weeks, J.M., 2003. Explaining density-dependent regulation in earthworm populations using life-history analysis.Oikos 100, 89e95.
Klinck, B., Palumbo, B., Cave, M., Wragg, J., 2005. Arsenic Dispersal and Bio-accessibilty in Mine Contaminated Soils: A Case Study from an AbandonedArsenic Mine in Devon. British Geological Survey, UK. Nottingham, 52 pp.
Klok, C., 2007. Effects of earthworm density on growth, development, andreproduction in Lumbricus rubellus (Hoffm.) and possible consequences forthe intrinsic rate of population increase. Soil Biology & Biochemistry 39 (9),2401e2407.
Klok, C., Zorn, M., Koolhaas, J.E., Eijsackers, H.J.P., van Gestel, C.A.M., 2006. Doesreproductive plasticity in Lumbricus rubellus improve the recovery of pop-ulations in frequently inundated river floodplains? Soil Biology & Biochemistry38 (3), 611e618.
Kooijman, S.A.L.M., 2000. Dynamic Energy and Mass Budgets in Biological Systems.Cambridge University Press, Cambridge, UK.
Langdon, C.J., Meharg, A.A., Feldmann, J., Balgar, T., Charnock, J., Farquhar, M.,Piearce, T.G., Semple, K.T., Cotter-Howells, J., 2002. Arsenic-speciation inarsenate-resistant and non-resistant populations of the earthworm, Lumbricusrubellus. Journal of Environmental Monitoring 4 (4), 603e608.
Langdon, C.J., Morgan, A.J., Charnock, J.M., Semple, K.T., Lowe, C.N., 2009. As-resistance in laboratory-reared F1, F2 and F3 generation offspring of theearthworm Lumbricus rubellus inhabiting an As-contaminated mine soil. Envi-ronmental Pollution 157 (11), 3114e3119.
Langdon, C.J., Piearce, T.G., Black, S., Semple, K.T., 1999. Resistance to arsenic-toxicityin a population of the earthworm Lumbricus rubellus. Soil Biology & Biochem-istry 31 (14), 1963e1967.
Langdon, C.J., Piearce, T.G., Meharg, A.A., Semple, K.T., 2003. Interactions betweenearthworms and arsenic in the soil environment: a review. EnvironmentalPollution 124 (3), 361e373.
Lika, K., Kooijman, S., 2003. Life history implications of allocation to growth versusreproduction in dynamic energy budgets. Bulletin of Mathematical Biology 65(5), 809e834.
Lowe, C.N., Butt, K.R., 2007. Earthworm culture, maintenance and species selectionin chronic ecotoxicological studies: a critical review. European Journal of SoilBiology 43, S281eS288.
Lu, X.W., Zhang, X.L., 2005. Environmental geochemistry study of arsenic inWestern Hunan mining area, PR China. Environmental Geochemistry andHealth 27 (4), 313e320.
Meharg, A.A., Shore, R.F., Broadgate, K., 1998. Edaphic factors affecting the toxicityand accumulation of arsenate in the earthworm Lumbricus terrestris. Environ-mental Toxicology and Chemistry 17 (6), 1124e1131.
Morgan, A.J., Kille, P., Sturzenbaum, S.R., 2007. Microevolution and ecotox-icology of metals in invertebrates. Environmental Science & Technology 41(4), 1085e1096.
Neubert, M.G., Caswell, H., 2000. Density-dependent vital rates and theirpopulation dynamic consequences. Journal of Mathematical Biology 41 (2),103e121.
PerezLosada, M., Eiroa, J., Mato, S., Dominguez, J., 2005. Phylogenetic speciesdelimitation of the earthworms Eisenia fetida (Savigny, 1826) and Eiseniaandrei Bouche, 1972 (Oligochaeta, Lumbricidae) based on mitochondrial andnuclear DNA sequences. Pedobiologia 49 (4), 317e324.
Shin, K.H., Kim, J.Y., Kim, K.W., 2007. Earthworm toxicity test for the monitoringarsenic and heavy metal-containing mine tailings. Environmental EngineeringScience 24 (9), 1257e1265.
Sims, R.W., Gerard, B.M., 1985. In: Kermack, D.M., Barnes, R.S.K. (Eds.), Earthworms.Linnean Society and the Estuarine and Brackish-Water Sciences Association,London, 171 pp.
Spurgeon, D.J., Hopkin, S.P., 1996. Effects of metal-contaminated soils on thegrowth, sexual development, and early cocoon production of the earthwormEisenia fetida, with particular reference to zinc. Ecotoxicology and Environ-mental Safety 35 (1), 86e95.
Spurgeon, D.J., Lawlor, A., Hooper, H.L., Wadsworth, R., Svendsen, C., Thomas, L.D.K.,Ellis, J.K., Bundy, J.G., Keun, H.C., Jarup, L., 2011. Outdoor and indoor cadmiumdistributions near an abandoned smelting works and their relations to humanexposure. Environmental Pollution 159 (12), 3425e3432.
Spurgeon, D.J., Svendsen, C., Kille, P., Morgan, A.J., Weeks, J.M., 2004. Responses ofearthworms (Lumbricus rubellus) to copper and cadmium as determined bymeasurement of juvenile traits in a specifically designed test system. Ecotox-icology and Environmental Safety 57 (1), 54e64.
Spurgeon, D.J., Svendsen, C., Weeks, J.M., Hankard, P.K., Stubberud, H.E.,Kammenga, J.E., 2003. Quantifying copper and cadmium impacts on intrinsicrate of population increase in the terrestrial oligochaete Lumbricus rubellus.Environmental Toxicology and Chemistry 22 (7), 1465e1472.
Stroud, J.L., Norton, G.J., Islam, M.R., Dasgupta, T., White, R.P., Price, A.H.,Meharg, A.A., McGrath, S.P., Zhao, F.-J., 2011. The dynamics of arsenic infour paddy fields in the Bengal delta. Environmental Pollution 159 (4),947e953.

C.J. Anderson et al. / Environmental Pollution 172 (2013) 200e207 207
van der Ploeg, M.J.C., Baveco, J.M., van der Hout, A., Bakker, R., Rietjens, I.M.C.M., vanden Brink, N.W., 2011. Effects of C(60) nanoparticle exposure on earthworms(Lumbricus rubellus) and implications for population dynamics. EnvironmentalPollution 159 (1), 198e203.
Walker, C.H., Hopkin, S.P., Sibly, R.M., Peakall, D.B., 2006. Principles of Ecotoxicology.Taylor & Francis, London, UK, 314 pp.
Widdows, J., Johnson, D., 1988. Physiological energetics of Mytilus edulis e scope forgrowth. Marine Ecology-Progress Series 46 (1e3), 113e121.
Yeates, G.W., Orchard, V.A., Speir, T.W., Hunt, J.L., Hermans, M.C.C., 1994.Impact of pasture contamination by copper, chromium, arsenic timberpreservative on soil biological activity. Biology and Fertility of Soils 18 (3),200e208.


















