
Leonid S. Brown Department of Physics, University of Guelph, ON N1G 2W1, Canada How Biological...
23
Leonid S. Brown Department of Physics, University of Guelph, ON N1G 2W1, Canada How Biological Systems Use How Biological Systems Use Light Light Or What Photobiophysics Is Photobiophysics Is All About
-
Upload
sergio-lickert -
Category
Documents
-
view
216 -
download
0
Transcript of Leonid S. Brown Department of Physics, University of Guelph, ON N1G 2W1, Canada How Biological...

Leonid S. Brown
Department of Physics, University of Guelph, ON N1G 2W1, Canada
How Biological Systems Use LightHow Biological Systems Use Light Or What Photobiophysics IsPhotobiophysics Is All
About

What Is Light?
Electromagnetic Spectrum

Major Meaningful Ways in which Living Creatures Use Light
• Energy Source
• Information Source
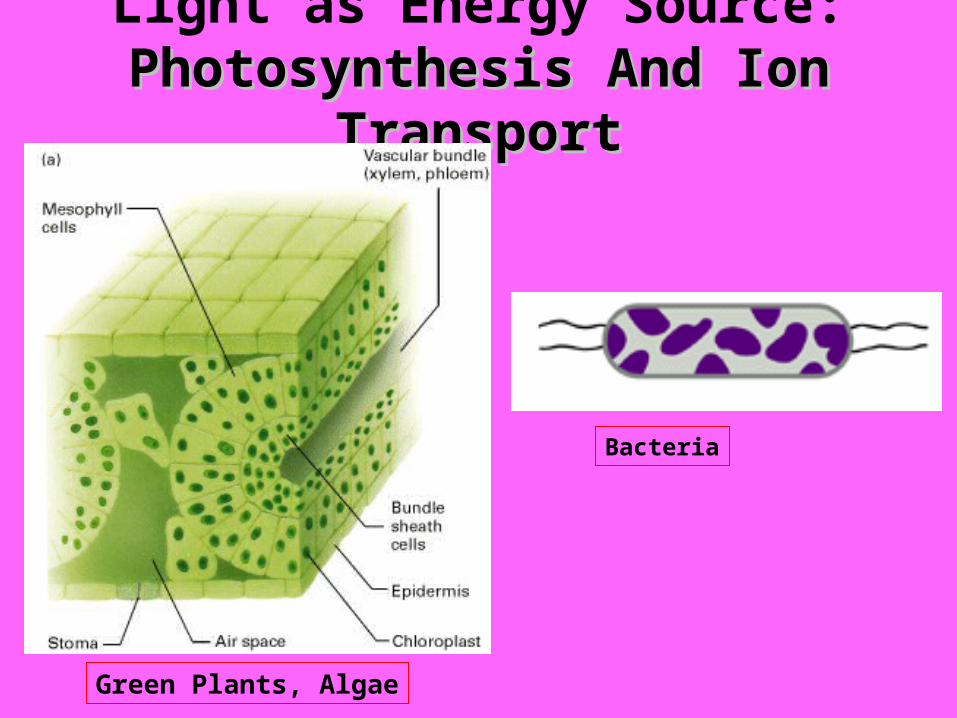
Light as Energy Source: Photosynthesis And Ion TransportPhotosynthesis And Ion Transport
Green Plants, Algae
Bacteria

Light as Information Source
• Phototaxis
• Phototropism
• Light-dependent Changes in Metabolism
• Photomorphogenesis
• Photoperiodism (circadian rhythms)
• Vision

Focus of Our Research at Focus of Our Research at GuelphGuelph
• To find out how very similar proteins can use light for completely different purposes, namely ion transport (ENERGY) or phototaxis (INFORMATION)
• To find and characterize biophysically (by means of visible and infrared spectroscopy) new photosensitive proteins

Description of the Current Project: New Eucaryotic Rhodopsins
Similar to the Haloarchaeal Ones
• Introduction to haloarchaeal rhodopsins and to the problem
• Our methods
• Representative results

Family of Haloarchaeal 7-TMH Retinal Proteins (Halobacterial Rhodopsins)
• Bacteriorhodopsin (BR) - H+ pump
• Halorhodopsin (HR) - Cl- pump
• Sensory rhodopsin I (SRI) - photosensor
• Sensory rhodopsin II (SRII) - photosensor
• These proteins bind all-trans retinal via lysine Schiff base, which photoisomerizes to 13-cis causing protein conformational changesFrom Spudich et al., Annu. Rev. Cell. Dev. Biol.,
2000, 16: 365
Retinal Schiff Base
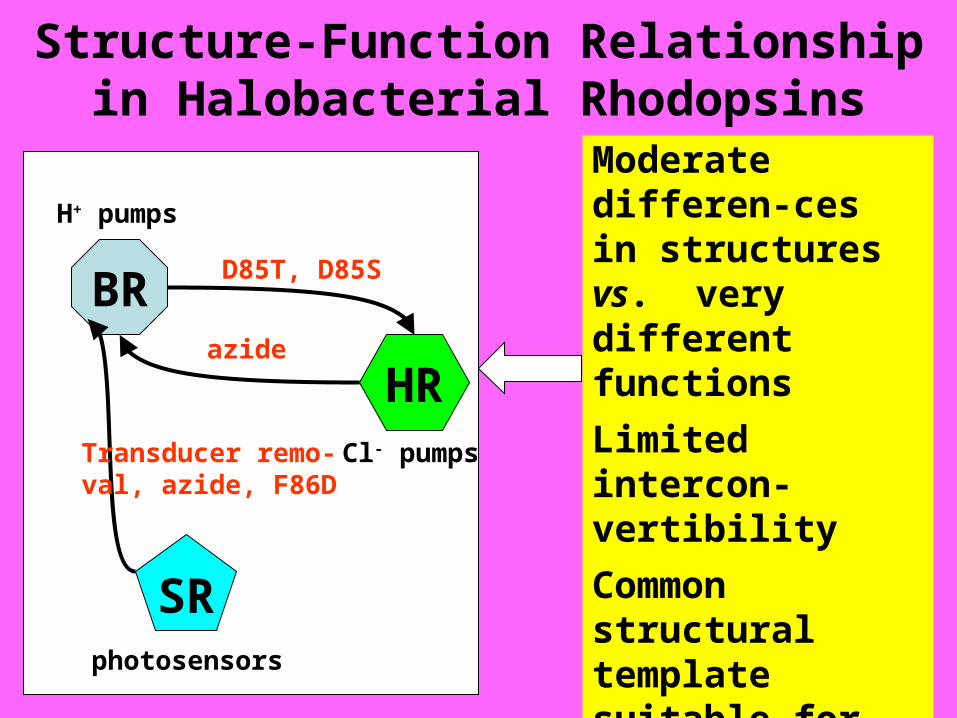
Structure-Function Relationship in Halobacterial Rhodopsins
Moderate differen-ces in structures vs. very different functions
Limited intercon-vertibility
Common structural template suitable for various functions?
How do the primary structures determine the functions?
BR
HR
SR
D85T, D85S
azide
Transducer remo-val, azide, F86D
Cl- pumps
H+ pumps
photosensors
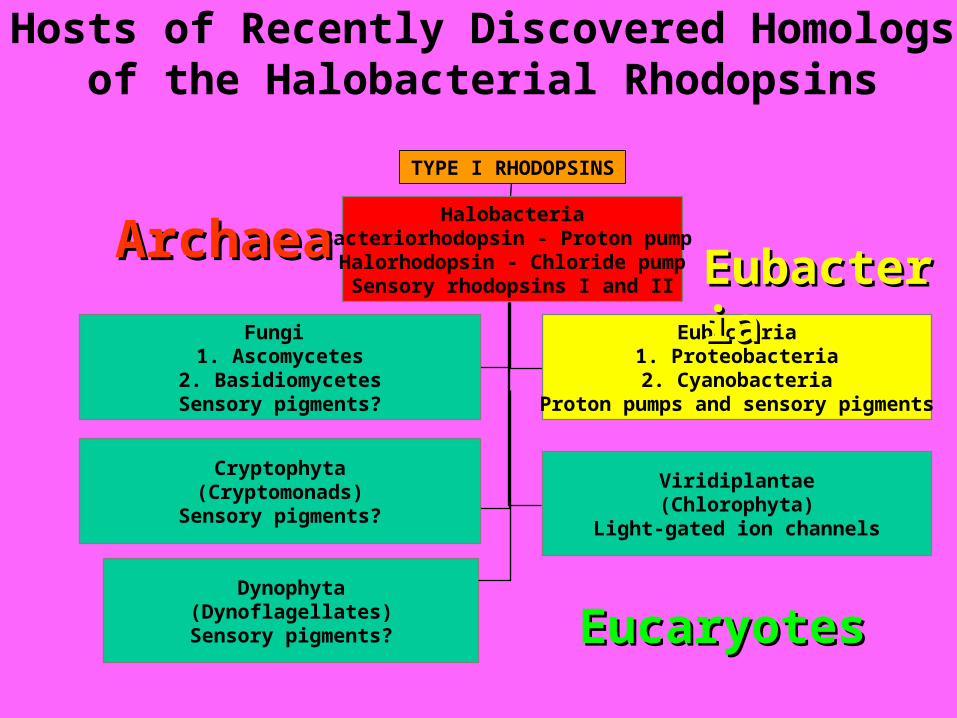
TYPE I RHODOPSINS
HalobacteriaBacteriorhodopsin - Proton pump
Halorhodopsin - Chloride pumpSensory rhodopsins I and II
Fungi 1. Ascomycetes
2. BasidiomycetesSensory pigments?
Eubacteria1. Proteobacteria2. Cyanobacteria
Proton pumps and sensory pigments
Cryptophyta(Cryptomonads)
Sensory pigments?
Viridiplantae(Chlorophyta)
Light-gated ion channels
ArchaeaArchaea
Dynophyta(Dynoflagellates)
Sensory pigments?
Hosts of Recently Discovered Homologs of the Halobacterial Rhodopsins
EucaryotesEucaryotes
EubacteriaEubacteria
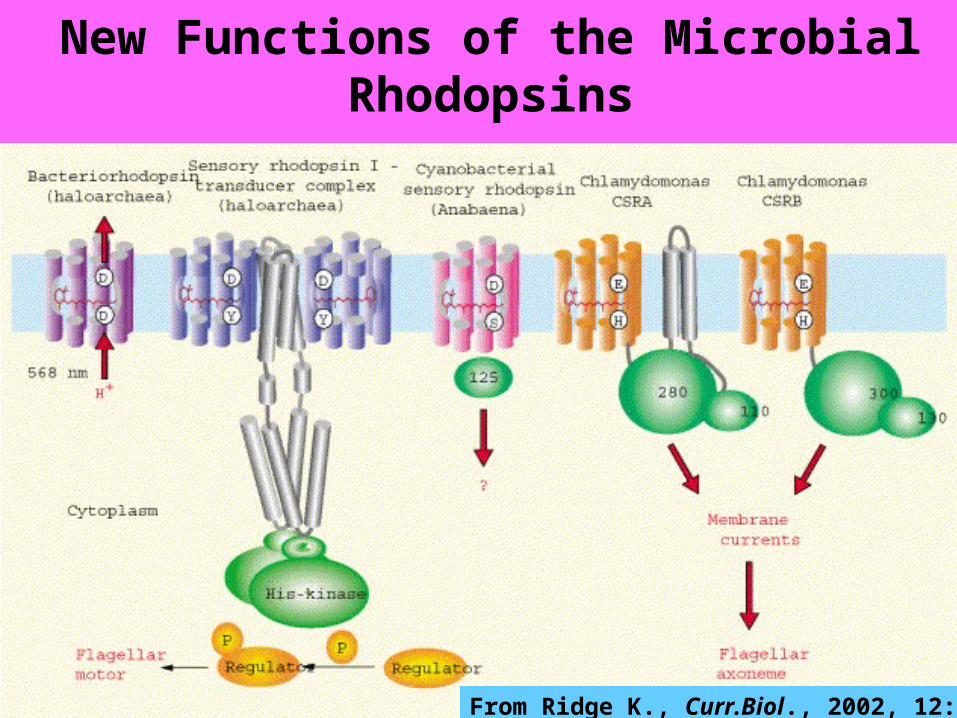
New Functions of the Microbial Rhodopsins
From Ridge K., Curr.Biol., 2002, 12: 588

Why Study Rhodopsins from Different Organisms?
• It is not clear why rhodopsins of a similar structure perform dramatically different functions. Accumulating various examples of pumping and sensory pigments from distant taxonomic groups can clarify how moderate changes in their primary structure translate into totally different functions
• From the other side, looking at natural amino acid sequence variability in the pigments of the same function should give clues about obligatory and optional elements of the pumping and sensory mechanisms in these rhodopsins

A New Fungal Rhodopsin Pipeline
Finding a new gene
Getting the DNA
OverexpressingVisible time-resolved and low-temperature
static spectroscopy
Raman
Time-resolved FTIRspectroscopy
His-tag purification, reconstitution in liposomes
(optional)
Low-temperature FTIRSpectroscopy (Nagoya)
Mutagenesis

Typical Characterization by Time-resolved Spectroscopy in the Visible
410 nm - follows protonation state of the retinal Schiff base
570 nm - follows disappe-arance of the initial state
660 nm - follows reisome-rization of the retinal and deprotonation of the primary proton acceptor
457 nm - in presence of pH-sensitive dye, follows H+ release and uptake
M
O
H+
N, BR
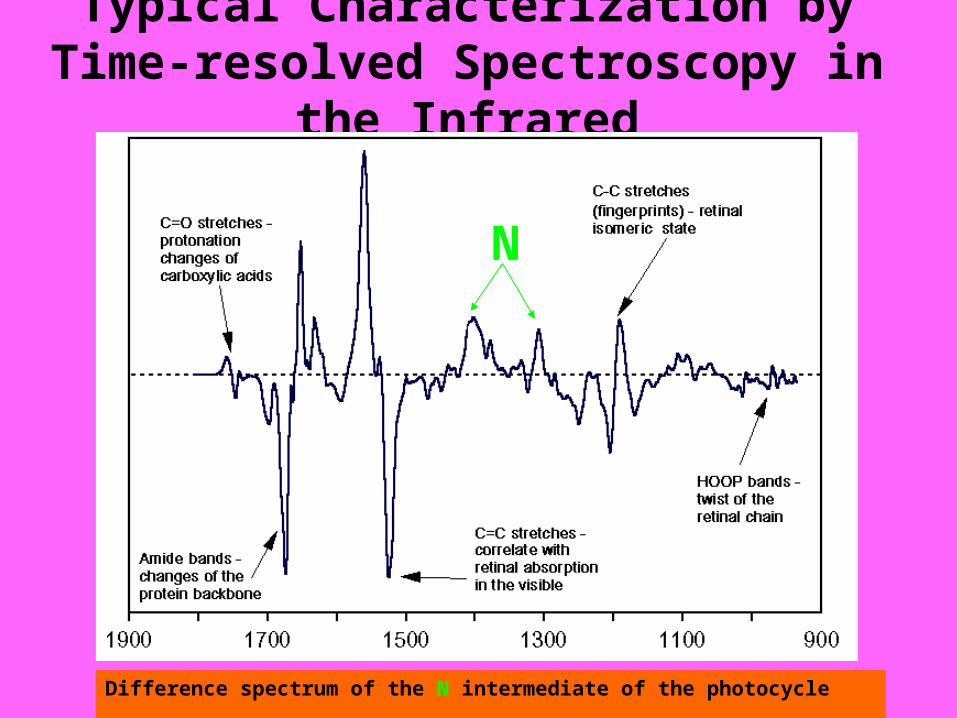
Typical Characterization by Time-resolved Spectroscopy in the Infrared
Difference spectrum of the N intermediate of the photocycle
N

NR - Rhodopsin from Neurospora
• Shows conservation of the residues important for H+ pumping in bacteriorhodopsin, including the primary H+ donor and acceptor
• When expressed heterologously, it binds retinal and has a photocycle similar to that of SRII (Bieszke et al, 1999) - photosensor or H+ pump? - No pumping detected directly!
From Spudich et al., Annu. Rev. Cell. Dev. Biol., 2000, 16: 365
E

Photocycle of NR vs H+ Pumps BR and PR
The intermediates known for BR are also found in NR, but its photocycle is much slower. A protein with such a slow photocycle would be a very inefficient H+ pump but an efficient photosensory pigment
(bacteriorhodopsin)(proteorhodopsin)
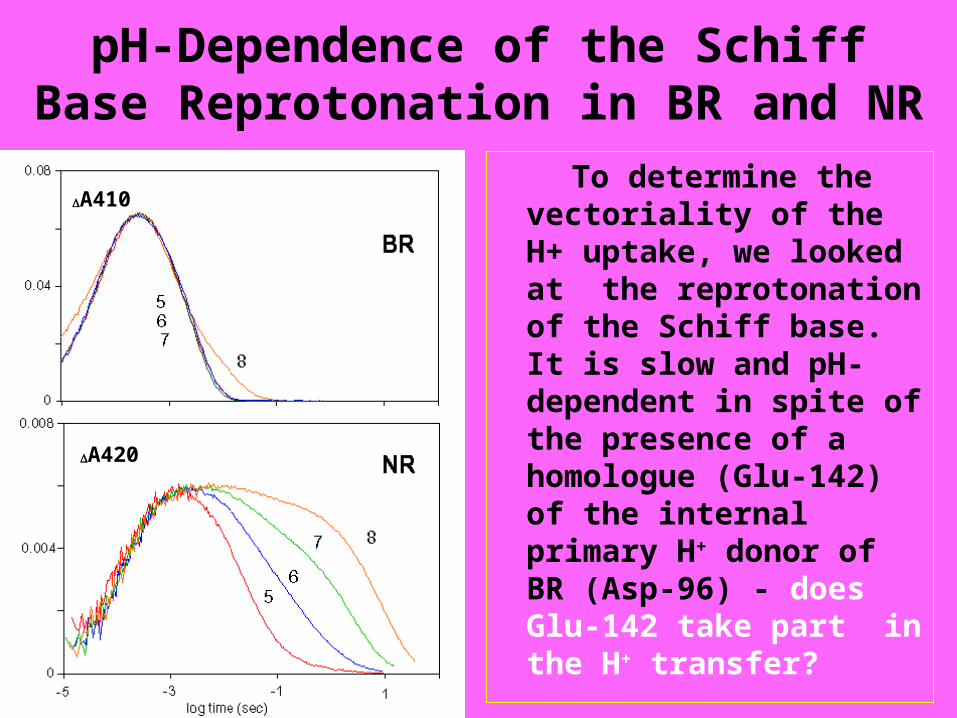
pH-Dependence of the Schiff Base Reprotonation in BR and NR
To determine the vectoriality of the H+ uptake, we looked at the reprotonation of the Schiff base. It is slow and pH-dependent in spite of the presence of a homologue (Glu-142) of the internal primary H+ donor of BR (Asp-96) - does Glu-142 take part in the H+ transfer?
A420
A410

Effect of Removing the Primary Proton Donor for the Schiff Base
Replacement of Glu-142 with Gln does not slow reprotonation of the Schiff base. This implies that the reprotonation (and proton uptake) may occur from the extracellular side, unlike in BR
and PR - no transport?

FTIR Difference Spectra of Late Photocycle Intermediates of NR and BR
• FTIR shows similar mixtures of the late photocycle intermediates (M, N, and O) in both proteins
• Main features include the protonation band of the primary proton acceptor (Asp-85 for BR, Asp-131 for NR), and bands of the isomerized (13-cis) chromophore

FTIR Spectra Confirm That Glu-142 Does Not Reprotonate the Schiff Base
FTIR spectra of the late inter-mediates of the NR photocycle are virtually unchanged when Glu-142 is replaced by Gln. COOH stretch region shows no depro-tonation (BR-like) or perturbation (PR-like) bands dependent on Glu-142 - is Glu-142 deprotonated in the unilluminated NR?
Instead, a new pH-dependent (correlated with reprotonation of the Schiff base) band is detected, which may belong to its H+ donor on the extracellular side of NR

FT-Raman Spectra Show Differences in Retinal Configuration between NR and BR
The proton transport may be absent because changes in the retinal conformation prevent the occurrence of the reprotonation switch (change in the Schiff base accessibility from the extracellular to the cytoplasmic side)

Acknowledgments
Andrei K. Dioumaev, Jennifer Shih, & Janos K. LanyiDept. of Physiology & Biophysics, Univ. of California, Irvine
Stefko Waschuk & Arandi Bezerra, Jr.Dept. of Physics, Univ. of Guelph
Yuji Furutani & Hideki KandoriNagoya Inst. of Technology
Elena N. Spudich & John L. SpudichDept. of Microbiology & Molecular Genetics, Univ. of Texas, Houston
Richard N. NeedlemanDept. of Biochemistry, Wayne State Univ., Detroit, MI


















