
Lens formation from cornea in the presence of the old lens in larval Xenopus laevis
6
THE JOURNAL OF EXPERIMENTAL ZOOLOGY 213:9-14 (1980) Lens Formation From Cornea in the Presence of the Old Lens in Larval Xenopus laevis LUIGI BOSCO, SERGIO FILONI, AND CARLA CIONI Istituto di Anatornm Cornparato, Uniuersita di Roma, 00161 Roma, Italy (L.B., S.F.), and Instituto di Zoologin e Anatomia Compamta, Uniuersita di Pisa, 56100 Pisa, Italy (C.C.) ABSTRACT Three different types of experiments were carried out to investi- gate the role of the lens in lens-forming transformations of the outer cornea of Xenopus laevis tadpoles (at stage 51, as defined by Nieuwkoop and Faber, '56): 1) simple lentectomy; 2) incision of outer and inner cornea in the region opposite the dorsal iris without lens removal; and 3) incision of outer and inner cornea and perforation of dorsal iris without lens removal. The results indicate that the outer cornea ofXenopus laevis larvae has lens-forming competence,even in the presence of the old lens, only when direct communication has been set up between the outer cornea and the environment of the vitreous chamber. These data indicate that the inhibition exerted by the lens on lens-forming transformations of the outer cornea is mechanical in nature. In some recent work (Filoni et al., '78a; Bosco et al., '79), we demonstrated that, once the outer cornea of lentectomized Xenopus laevis larvae is provided with sufficient nutritional substances, it will undergo lens-forming trans- formations, but only if it is reached by a fac- tor(s) present in the eye environment. In a normal eye, this factor is not capable of passing the barrier of the inner cornea. Campbell and Jones ('68) assert that the phenotypic stability of the outer cornea of a normal Xenopus laevis eye is probably due to an inhibiting action of a chemical nature exerted by the lens. According to them, this inhibiting actioncould be positive, in the sense that the lens may produce a sub- stance capable of inhibiting the lens-forming transformations of the outer cornea, or else it could be negative, in the sense that the lens may preferentially remove a substance neces- sary for regeneration. Recently, however, Reeve and Wild ('78), after removing the original lens and its re- placement in the pupillary space, observed cases of lens differentiation from the outer cornea or from fragments of outer cornea im- planted in the vitreous chamber, in the pres- ence of the lens. In most cases, the regrafted lenses had a normal histological appearance; in other cases, the old lenses were damaged, but there was no correlation between lens damage and lens formation from the cornea. However, since the operated eyes were examined some days after the operation, it is possible that damage occurred even in the lenses that ap- peared normal. Furthermore, the workers do not state whether lens replacement was carried out preserving the orignal polarity. Therefore, it appeared desirable to approach the problem of the inibitory role of the lens using experimental techniques that do not re- quire the lens to be handled, as well as insuring that its original polarity and morphological in- tegrity be maintained. If this is done, it be- comes possible to exclude the possibility that the presence of any regenerated lens is due to morphological and functional damage suffered by the original lens andlor to its reconstruction from the epithelio-capsular fragments. In a previous paper (Filoni et al., '801, we reported that after making an incision in the dorsal pericorneal integument and the under- lying eye wall, we found that the parts of pericorneal epidermis affected by the opera- tion, and thereby placed in direct communica- tion with the environment of the vitreous chamber, underwent lens-forming transforma- tions, even in the presence of the original lens. We found that these transformations were quantitatively similar to those found in the pericorneal epidermis of lentectomized eyes. Thus, we concluded that once direct communi- cation has been set up between this tissue hav- ing lens-regenerating competence and the vit- reous chamber, the posterior region of the lens is unable to neutralize or effectively remove the lens-forming factors(s) contained in the vitre- 0022-104X/80/2131-0009$01.40 IC, 1980 ALAN R. LISS, INC.
-
Upload
luigi-bosco -
Category
Documents
-
view
215 -
download
1
Transcript of Lens formation from cornea in the presence of the old lens in larval Xenopus laevis

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 213:9-14 (1980)
Lens Formation From Cornea in the Presence of the Old Lens in Larval Xenopus laevis
LUIGI BOSCO, SERGIO FILONI, AND CARLA CIONI Istituto di Anatornm Cornparato, Uniuersita di Roma, 00161 Roma, Italy (L.B., S.F.), and Instituto di Zoologin e Anatomia Compamta, Uniuersita di Pisa, 56100 Pisa, Italy (C.C.)
ABSTRACT Three different types of experiments were carried out to investi- gate the role of the lens in lens-forming transformations of the outer cornea of Xenopus laevis tadpoles (at stage 51, as defined by Nieuwkoop and Faber, '56): 1) simple lentectomy; 2) incision of outer and inner cornea in the region opposite the dorsal iris without lens removal; and 3) incision of outer and inner cornea and perforation of dorsal iris without lens removal. The results indicate that the outer cornea ofXenopus laevis larvae has lens-forming competence, even in the presence of the old lens, only when direct communication has been set up between the outer cornea and the environment of the vitreous chamber. These data indicate that the inhibition exerted by the lens on lens-forming transformations of the outer cornea is mechanical in nature.
In some recent work (Filoni et al., '78a; Bosco et al., '79), we demonstrated that, once the outer cornea of lentectomized Xenopus laevis larvae is provided with sufficient nutritional substances, it will undergo lens-forming trans- formations, but only if it is reached by a fac- tor(s) present in the eye environment. In a normal eye, this factor is not capable of passing the barrier of the inner cornea. Campbell and Jones ('68) assert that the phenotypic stability of the outer cornea of a normal Xenopus laevis eye is probably due to an inhibiting action of a chemical nature exerted by the lens. According to them, this inhibiting actioncould be positive, in the sense that the lens may produce a sub- stance capable of inhibiting the lens-forming transformations of the outer cornea, or else it could be negative, in the sense that the lens may preferentially remove a substance neces- sary for regeneration.
Recently, however, Reeve and Wild ('78), after removing the original lens and its re- placement in the pupillary space, observed cases of lens differentiation from the outer cornea or from fragments of outer cornea im- planted in the vitreous chamber, in the pres- ence of the lens. In most cases, the regrafted lenses had a normal histological appearance; in other cases, the old lenses were damaged, but there was no correlation between lens damage and lens formation from the cornea. However, since the operated eyes were examined some days after the operation, it is possible that
damage occurred even in the lenses that ap- peared normal. Furthermore, the workers do not state whether lens replacement was carried out preserving the orignal polarity.
Therefore, it appeared desirable to approach the problem of the inibitory role of the lens using experimental techniques that do not re- quire the lens to be handled, as well as insuring that its original polarity and morphological in- tegrity be maintained. If this is done, it be- comes possible to exclude the possibility that the presence of any regenerated lens is due to morphological and functional damage suffered by the original lens andlor to its reconstruction from the epithelio-capsular fragments.
In a previous paper (Filoni et al., '801, we reported that after making an incision in the dorsal pericorneal integument and the under- lying eye wall, we found that the parts of pericorneal epidermis affected by the opera- tion, and thereby placed in direct communica- tion with the environment of the vitreous chamber, underwent lens-forming transforma- tions, even in the presence of the original lens. We found that these transformations were quantitatively similar to those found in the pericorneal epidermis of lentectomized eyes. Thus, we concluded that once direct communi- cation has been set up between this tissue hav- ing lens-regenerating competence and the vit- reous chamber, the posterior region of the lens is unable to neutralize or effectively remove the lens-forming factors(s) contained in the vitre-
0022-104X/80/2131-0009$01.40 IC, 1980 ALAN R. LISS, INC.

10 L. BOSCO, S. FILONI, AND C. CIONI
ous chamber. Nevertheless, on the basis of these conclusions alone, it is not legitimate to assume that, even on the outer cornea left in situ, the lens does not exert any chemical-type inhibition and that this inhibition must thus be mechanical in nature. In fact, it is conceivable
in paraffin, and cut into 7-pm cross-sections. The serial sections were stained with hematox- ylin-eosin, using either the Mallory-Azan method or by treatment with PAS reaction. Three different kinds of experiments were ac- complished (Fig. 1).
that the lens has a differential activity at the two Doles.
-
Experiment Z--simple lentectomy (Fig. 1 ) 1 - ~ - ~ . .
The aim of the present research is to evaluate the function of the anterior region of the lens in lens-forming transformations of the outer cornea.
In this experiment, 35 Xenopus laevis tad- poles, from which the lens of the right eye had been removed, were used. The lentectomy techniaue used had been extensivelv emploved
MATERIALS AND METHODS by us ireviously (Filoni et al., '76; '$7; Bbscb et al., '79). After incising the outer and inner
The animals used in the present work con- sistedof llOXenopus laevis tadpoles a t stage 51 (according to the definition of Nieuwkoop and Faber, '56) obtained after ovulation induced by gonadotropic hormone (Pregnyl-Organon) and
cornea, the lens was removed by means of aspi- ration. The surviving animals were sacrificed at 3, 5 , 7, 10, and 15 days after the operation. Five additional larvae were sacrificed immedi- ately afterward to serve as controls.
mating. During the operations, the tadpoles were immersed in full-strength Holtfreter's solution, where they remained for 24 hours after the operation. The operated individuals were later gradually transferred to tap water and fed on boiled nettle powder. The larvae were then fixed in Bouin's solution, embedded
B d
C
Experiment ZZ--incision of outer and inner cornea at the dorsal iris without lens removal
(Fig. 1) Thirty-five larvae were operated upon. The
animals were subjected to an incision of the outer and inner cornea in the region opposite the dorsal iris, without removing the lens. The operated tadpoles were sacrificed at 3,5,7, 10, and 15 days after the operation (Table 1). Five additional animals were sacrificed immedi- ately after the operation to serve as controls.
Experiment ZfZ-incision of the outer and inner cornea, trnd perforation of dorsal iris
without lens removal (Fig. 1 ) Operations were performed on thirtv-five
I -
Fig. 1. Diagram of various types of operation. A) Simple lentectomy (experiment I). B) Incision of outer and inner cornea at dorsal iris without lens removal (experiment 11). C) Incision of the outer and inner cornea and perforation ofthe dorsal iris without lens removal (experiment 111).
larvae. All anirnals were subjected to an inci- sion of outer and inner cornea in the region Opposite the dorsal iris. Then a hole was made in the dorsal iris with iridectomy scissors. The
TABLE 1 . Summary of the results of experiment II and experiment III
No of Lens neoformation stage Days after No of cases cases deadi No of cases No of
Experiment operation operated discarded examined regenerates st. 2 st. 3 st. 4 st. 5
- - I1 3 5 - - 5 5 5 - - 5 7 5 - - 5 10 5 - - 5 15 10 1 9 - -
111 3 5 5 4 2 2
- -
- - - - - - - -
- - - -
- - -
- - - 5 5 1 4 3 - 1 2 -
- 3 - I 5 1 4 3 - 2 1 10 5 1 4 3
1 1 15 10 6 4 2 - - -
-
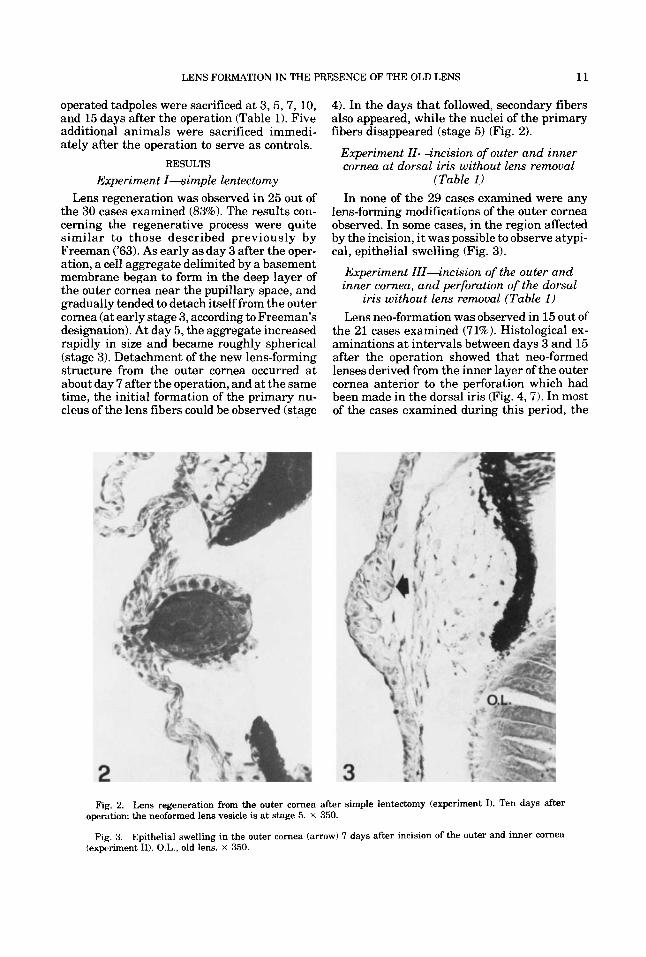
LENS FORMATION IN THE PRESENCE OF THE OLD LENS 11
operated tadpoles were sacrificed at 3,5,7, 10, and 15 days after the operation (Table 1). Five additional animals were sacrificed immedi- ately after the operation to serve as controls.
Experiment I-simple lentectomy Lens regeneration was observed in 25 out of
the 30 cases examined (83%). The results con- cerning the regenerative process were quite similar to those described previously by Freeman ('63). As early as day 3 after the oper- ation, a cell aggregate delimited by a basement membrane began to form in the deep layer of the outer cornea near the pupillary space, and gradually tended to detach itself from the outer cornea (at early stage 3, according to Freeman's designation). At day 5, the aggregate increased rapidly in size and became roughly spherical (stage 3). Detachment of the new lens-forming structure from the outer cornea occurred at about day 7 after the operation, and at the same time, the initial formation of the primary nu- cleus of the lens fibers could be observed (stage
RESULTS
4). In the days that followed, secondary fibers also appeared, while the nuclei of the primary fibers disappeared (stage 5) (Fig. 2).
Experiment Zl-incision of outer and inner cornea at dorsal iris without lens removal
(Table 1) In none of the 29 cases examined were any
lens-forming modifications of the outer cornea observed. In some cases, in the region affected by the incision, i t was possible to observe atypi- cal, epithelial swelling (Fig. 3).
Experiment ZIZ-incision of the outer and inner cornea, and perforation of the dorsal
iris without lens remoual (Table 1) Lens neo-formation was observed in 15 out of
the 21 cases examined (71%). Histological ex- aminations a t intervals between days 3 and 15 after the operation showed that neo-formed lenses derived from the inner layer of the outer cornea anterior to the perforation which had been made in the dorsal iris (Fig. 4, 7). In most of the cases examined during this period, the
Fig. 2. Lens regeneration from the outer cornea after simple lentectomy (experiment I). Ten days after operation: the neoformed lens vesicle is at stage 5. x 350.
Fig. 3. Epithelial swelling in the outer cornea (arrow) 7 days after incision of the outer and inner cornea (experiment 11). O.L., old lens. x 350.
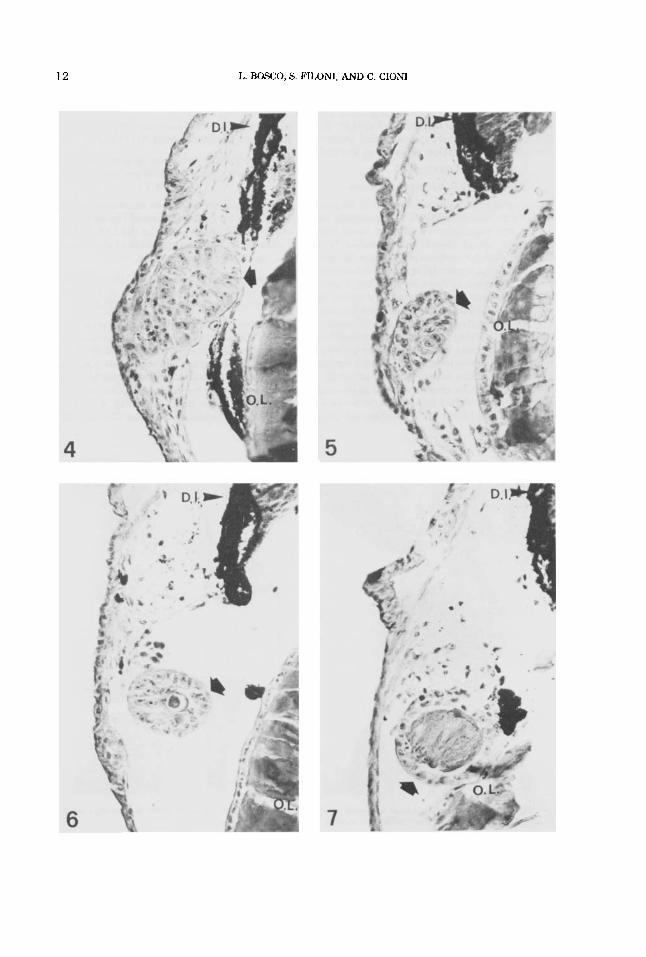
12 L. BOSCO, S. FILONI, AND C. CIONI

LENS FORMATION IN THE PRESENCE OF THE OLD LENS 13
new lens-forming structures were still attached to the outer cornea (Fig. 4, 5) .
Analysis of the stages involved in the process of lens neo-formation between days 3 and 10 after the operation revealed that these stages follow basically the same sequence as was found in experiment I (simple lentectomy), with a few differences. In fact, even in those cases in which the newly-formed lens exhibited a highly advanced stage of differentiation, its size was smaller than the regenerated lens in comparable individuals subjected to simple lentectomy. In no case did the regenerated lens, even when detached from the outer cornea, suc- ceed in reaching the vitreous chamber; instead, it remained in the space between the outer cornea (from which it was derived) and the dor- sal iris (Fig. 4, 5 , 6, 7).
Of the four cases examined after 15 days, a newly-formed lens was observed in two cases only. Furthermore, the volume of the lens was less, and it showed less differentiation than in those cases observed ten days after operation. In all of these cases, the neo-formed lenses were situated in the anterior chamber, and the wound in the dorsal iris had completely healed.
DISCUSSION
In 1arvalXenopus laevis, Campbell and Jones ('68) observed lens regeneration from outer cornea fragments cultivated in vitro, and they explain this result by attributing an intrinsic lens-forming capacity to the outer cornea. These authors assume that the phenotypic sta- bility of the outer cornea of a normal eye is insured by an inhibiting action of a chemical nature, probably exerted by the lens. However, Waggoner ('731, implanting the outer cornea in various regions of the body of Xenopus laevis tadpoles (lentectomized eyes, dorsal fins, am- putated hind limbs, or subcutaneously in the head region), observed that lenses formed from
the implants in the lentectomized eyes and in the amputated hind limbs. This author attri- butes the lens formation from the outer cornea in these two regions to the presence of factors capable of stimulating the cornea.
The results of the present investigation show that the outer cornea of the eye of Xenopus laevis tadpoles can also form a lens in the pres- ence of the old lens. However, this happens only when direct communication has been set up between the outer cornea and the environment of the vitreous chamber. In fact, in experiment I1 (incision of the outer and inner cornea), in which the tadpoles did not undergo perforation of the dorsal iris and the outer cornea was iso- lated from the vitreous chamber by the pres- ence of the old lens, the outer cornea did not undergo any lens-forming transformation, even though the inner cornea had been incised and the outer cornea was thereby in direct communication with the environment of the anterior chamber.
Lens-forming transformations of the outer cornea in the presence of the lens were found in experiment I11 (incision of the outer and inner cornea and subsequent perforation of the dorsal iris). In this case, since the dorsal iris had been perforated, the outer cornea was in communi- cation with the environment of the vitreous chamber. In particular, under these experi- mental conditions, the outer cornea formed a lens through the typical sequence of mor- phogenetic stages found in lentectomized eyes (experiment I).
These data demonstrate that the factor re- sponsible for the lens-forming transformations of the outer cornea comes from the vitreous chamber and that the lens exerts effective in- hibiting action only when it is able to provide an obstacle to communication between the an- terior chamber and the vitreous chamber. However, the lens cannot guarantee the pheno-
Fig. 4-7. Neoformation of the lens after incision of the outer and inner cornea, and perforation of the dorsal iris (experiment 111). In all figures the neoformed lens vesicle is situated in front of the wound of the dorsal iris. Stainingwas done by the Mallory-Azan method. x 300.
Fig. 4. Five days after operation: the neoformed lens vesicle is at stage 3 (arrow).
Fig. 5. Seven days after operation: the neoformed lens vesicle is at stage 3 (arrow).
Fig. 6. Ten days after operation: the neoformed lens vesicle is at stage 4 (arrow).
Fig. 7. Ten days after operation: the neoformed lens vesicle is at stage 5 (arrow). D.I., dorsal iris; O.L., old lens.

14 L. BOSCO, S. FILONI. AND C. CIONI
typic stability of the outer cornea whenever such communication has been set up on a large scale.
However, in experiment 111, the percentage of regeneration observed between day 3 and day 10 after operation (76%) was slightly lower than that found in experiment I (83%). Fur- thermore, the regenerated lenses observed after 15 days, besides being very small in vol- ume, displayed a lower degree of differentiation than those observed at day 10. The first of these results, i.e., the lower percentage regeneration, may be explained by reference to the results obtained in previous experiments which showed that the inner cornea forms an insur- mountable barrier to the eye factor(s) responsi- ble for the lens-forming transformations of the outer cornea (Filoni et al., '78a; Bosco et al., '79). Indeed, in experiment 111, the cut made in the inner cornea could not be as large as the cut in experiment I, or the old lens would have been forced into the anterior chamber or ejected from it under the pressure of the vitreous humor. Under the conditions of experiment 111, the inner cornea healed more quickly and the outer cornea was isolated from the eye environment in less time. The lenses' small size and the re- gression of the regenerated lenses observed at day 15 after operation may be explained by the fact that the factor(s) contained in the vitreous chamber is necessary not only to trigger the lens-forming transformations of the territories with lens-regenerating competence (in this case of the outer cornea), but also to stimulate the subsequent growth and conservation of the newly formed lens-forming structures (Filoni ek al., '78b; Bosco et al., '79). In fact, the incision made in the dorsal iris in experiment I11 heals very quickly, being closed completely by day 10 after operation.
In previous investigations (Filoni et al., '80), we showed that the posterior region of the lens is not capable of preventing the lens-forming transformations of the pericorneal epidermis when the latter is in direct communication with the environment of the vitreous chamber. These data all show that neither the posterior region of the lens nor the anterior one are capa- ble of effectively neutralizing or removing this factor(s1, which is present in the vitreous chamber.
Our results, in accordance with the results of Reeve and Wild ('78), indicate that the inhibi-
tion exerted by the lens on lens-forming trans- formations of the outer cornea is mechanical rather than chemical in nature. This is evi- denced by the fact that, since the lens occupies the pupillary space, it prevents the factor(s) from escaping from the vitreous chamber. In- asmuch as we have previously demonstrated that the inner cornea forms an insurmountable barrier for the factor(s) necessary for the lens- forming transformations of the outer cornea, quite apart from the lens (Filoni et al., '78a; Bosco et al., '79), it may be concluded that the phenotypic stability of the outer cornea of the normal eye of the Xenopus laevis tadpole is guaranteed by the presence of two eye struc- tures, i.e., the inner cornea and the lens, both of which prevent any direct contact between the outer cornea and the environment of the vitre- ous chamber.
LITERATURE CITED
Bosco, L., S. Filoni, and S. Cannata (1979) Relationships between eye factors and lens-forming transformations in the cornea and pericorneal epidermis of larval Xeizopus lueuis. J. Exp. Zool., 209t261-282.
Campbell, J.C., and K.W. Jones (1968) The in uitro develop- ment of lens from cornea oflarvalXempus laeuis. Develop. Biol., 17:l-15.
Filoni, S., L. Bosco, and C. Cioni (1976) I1 problema della rigenerazione del cristallino degli Anfibi Anuri negli stadi post-embrionali. Esperienze di asportazione del cristallino in larve diRana esculenta exenopus laeuis. Acta Embryol. Exper., 3:319-334.
Filoni, S., L. Bosco, and C. Cioni (1977) I1 problema della rigenerazione del cristallino degli M i b i Anuri negli stadi post-embrionali 11. Esperienze di asportazione del cristal- lino in larve di Discoglossuspictus. Acta Embryol. Exper., 2:155-162.
Filoni, S., L. Bosco, and S. Cannata (1978a) Primi dati speri- mentali sui fattori necessari per la trasformazione len- togena della cornea esterna di larve di Xenopus lueuis. Acta Embryol. Exper., 3:344.
Filoni, S., L. Bosco, and S. Cannata (1978b) Influenza dell'oc- chi0 sulla rigenerazione del cristallino in larve diXenopus laeuis. Acta Embryol. Exper., 2:179-195.
Filoni, S., L. Bosco, N. Paglioni, and C. Cioni (1980) Lens formation from pericorneal epidermis in presence of the old lens in larval Xenopus laeuis. J. Exp. 2001. (in press).
Freeman, G. (19631 Lens regeneration from cornea in Xenopus laeuis. J. Exp. Zool., 154:39-65.
Nieuwkoop, P.D., and J. Faber (1956) Normal table of Xempus laeuis (Daudin). North-Holland Publishing Com- pany, Amsterdam.
Reeve, J.G., and A.E. Wild (1978) Lens regeneration from cornea of larval Xenopus laeuis in the presence of the lens. J. Embryol. Exp. Morph., 48:205-214.
Waggoner, P.L. (1973) Lens differentiation from the cornea following lens exi.irpation or cornea transplantation in Xempus laeuis. J. Exp. Zool., 186:97-105.


















