Lens Antibodies and Eye Development - ARVO...
10
Lens antibodies and eye development Jan hangman and Harry Maisel When 32 hour chick embryos were treated with different lens antibodies, cellular degenera- tion and gross abnormalities, such as anencephalus, rachischisis, and anophthalmia, were observed only in embryos treated with antisera prepared against total lens and alpha crystallin, but not in those treated with beta or gamma crystallin antiserum. Since the restdts of treatment with total lens and alpha crystallin antibodies were identical in nature, it is concluded that the antibody effect is due exclusively to the interaction of alpha crystallin antibodies and their corresponding cellular anitgens. The cellular degeneration and gross abnormalities were restricted to the brain and eye, that is, those organs in which the presence of lens antigens was previously demonstrated by means of immunologic and fluorescent antibody techniques. None of the other tissues and organs in the abnormal embryos showed any effect of antibody treatment, thus indicating a high specificity of the alpha crystallin antigen-antibody reaction. Abnormalities found in the 24 hour series were identical to those obtained in the 32 hour series, indicating that in both cases the alpha crystallin antibodies had interfered with the same developmental processes. Treatment of 42 hour embryos was ineffective, suggesting that at this stage of development the antigen-antibody interaction either has become impossible or is no longer able to interfere with morphologic development. Since in many of the grossly abnormal embryos no indication of cellular degeneration was found, it seems likely that an antigen-antibody interaction does not necessarily lead to cellular degeneration and necrosis, but may also interfere with cell life in a less damaging manner, such as inhibition of growth and rechanneling of normal differentiation capacities. When rabbits were immunized with chick lens extracts before and during pregnancy, lens antibodies were demonstrated in the maternal circulation throughout pregnancy, in the yolk-sac fluid at the tenth day of develop- ment, and in the fetal circidation at the twenty-eighth day of development, by means of immunologic methods. None of the treated rabbit fetuses, however, showed any abnormalities. I t has recently been suggested that lens antibodies circulating in the maternal bloodstream may interfere with lens de- velopment in the mammalian embryo and cause cataracts based on an antigen-anti- body interaction. 14 Even more surprising is the observation of Barber, Willis, and From the Department of Anatomy, McGill Uni- versity, Montreal, Quebec. This work was supported by a Fight for Sight Research Fellowship from the National Council to Combat Blindness, Inc., New York, to one of us (H. M.), and by grants from the National Research Council and the National Cancer Institute of Canada. Afeman, 1 who immunized mice against brain extract and found congenital lens abnormalities as a result of this treatment. Careful analysis of these and other experi- ments with tissue antibodies 3 ' G shows that in some experiments the actual presence of circulating antibodies has not been demonstrated, whereas in others degenera- tion was found in tissues and organs which do not contain antigenic components cor- responding to the circulating antibodies at any stage of development. It is thus evi- dent that the results obtained in these experiments are difficult to explain in a logical manner. Since a careful analysis of an antigen- antibody interaction in mammalian em- 396 Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018
Transcript of Lens Antibodies and Eye Development - ARVO...

Lens antibodies and eye development
Jan hangman and Harry Maisel
When 32 hour chick embryos were treated with different lens antibodies, cellular degenera-tion and gross abnormalities, such as anencephalus, rachischisis, and anophthalmia, wereobserved only in embryos treated with antisera prepared against total lens and alpha crystallin,but not in those treated with beta or gamma crystallin antiserum. Since the restdts of treatmentwith total lens and alpha crystallin antibodies were identical in nature, it is concluded thatthe antibody effect is due exclusively to the interaction of alpha crystallin antibodies andtheir corresponding cellular anitgens. The cellular degeneration and gross abnormalities wererestricted to the brain and eye, that is, those organs in which the presence of lens antigenswas previously demonstrated by means of immunologic and fluorescent antibody techniques.None of the other tissues and organs in the abnormal embryos showed any effect of antibodytreatment, thus indicating a high specificity of the alpha crystallin antigen-antibody reaction.Abnormalities found in the 24 hour series were identical to those obtained in the 32 hourseries, indicating that in both cases the alpha crystallin antibodies had interfered with thesame developmental processes. Treatment of 42 hour embryos was ineffective, suggesting thatat this stage of development the antigen-antibody interaction either has become impossibleor is no longer able to interfere with morphologic development. Since in many of the grosslyabnormal embryos no indication of cellular degeneration was found, it seems likely that anantigen-antibody interaction does not necessarily lead to cellular degeneration and necrosis,but may also interfere with cell life in a less damaging manner, such as inhibition of growthand rechanneling of normal differentiation capacities. When rabbits were immunized withchick lens extracts before and during pregnancy, lens antibodies were demonstrated in thematernal circulation throughout pregnancy, in the yolk-sac fluid at the tenth day of develop-ment, and in the fetal circidation at the twenty-eighth day of development, by means ofimmunologic methods. None of the treated rabbit fetuses, however, showed any abnormalities.
I t has recently been suggested that lensantibodies circulating in the maternalbloodstream may interfere with lens de-velopment in the mammalian embryo andcause cataracts based on an antigen-anti-body interaction.14 Even more surprisingis the observation of Barber, Willis, and
From the Department of Anatomy, McGill Uni-versity, Montreal, Quebec.
This work was supported by a Fight for SightResearch Fellowship from the National Councilto Combat Blindness, Inc., New York, to oneof us (H. M.), and by grants from the NationalResearch Council and the National CancerInstitute of Canada.
Afeman,1 who immunized mice againstbrain extract and found congenital lensabnormalities as a result of this treatment.Careful analysis of these and other experi-ments with tissue antibodies3'G shows thatin some experiments the actual presenceof circulating antibodies has not beendemonstrated, whereas in others degenera-tion was found in tissues and organs whichdo not contain antigenic components cor-responding to the circulating antibodies atany stage of development. It is thus evi-dent that the results obtained in theseexperiments are difficult to explain in alogical manner.
Since a careful analysis of an antigen-antibody interaction in mammalian em-
396
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018

Volume 1Number 3 Lens antibodies and eye development 397
bryos is exceedingly difficult, it was decidedfirst to test this interaction in the chickembryo. Hence, 24, 32, and 42 hour chickembryos were treated with antisera pre-pared against total chick lens extract andwith antibodies prepared against theisolated lens proteins, alpha, beta, andgamma crystallin. In the second experi-ment, rabbits were immunized with chicklens extract before and during pregnancyand the effect of this treatment on theembryos was analyzed.
Materials and methodsWhite Leghorn eggs incubated at 38° C. for
24, 32, or 42 hours were cleaned with 70 percent alcohol. After a small hole was punctured inboth ends of the egg, an opening was sawed inthe shell and the membrane lifted to expose theembryo. Whenever the embryo remained attachedto the shell membrane or was not located directlyunder the opening, the egg was discarded. Sub-sequently, 0.1 ml. of antiserum was depositedover the embryo. The openings in the top and atboth ends were then closed with cellophane tape,sealed with paraffin, and the eggs incubated foran additional 50 to 70 hour period. After thisperiod, the embryos were removed from the eggsand examined under the dissecting microscopefor the presence of abnormalities. Deformed em-bryos were fixed in Bouin's fluid, sectioned, andstained with hematoxylin and eosin for histologicexamination.
Preparation of antibodies. A 25 per cent adultlens extract suspended in Freund's adjuvant in aratio of 3:2 was injected subcutaneously into anumber of rabbits in the amount of 1 ml. at eachof 5 widely separated sites. This procedure wasrepeated 4 to 10 times at weekly intervals. Theserum of the rabbits was obtained 14 days afterthe last injection.
Alpha, beta, and gamma crystallin were isolatedby running adult lens extract containing 15 mg.protein per milliliter in a Spinco Model CP ap-paratus for continuous flow electrophoresis. Theseparation was made at 4° C. in Veronal bufferat pH 8.6, ionic strength 0.02, and current 60Ma. The fractions were collected, dialyzed, con-centrated to a protein value of 35 mg. per cent,and tested for homogeneity in the ultracentrifugeand by the agar-diffusion method. The isolatedfractions were suspended in Freund's adjuvantand injected into rabbits as oudined above. Theantisera obtained are referred to as alpha, beta,and gamma crystallin antisera.
In some experiments the chick embryos weretreated with complete antiserum, whereas in
others only the gamma globulin fraction was used.This fraction was prepared by mixing the anti-serum with an equal amount of saturated am-monium sulfate. After leaving the mixture over-night in the cold, the precipitate obtained wassuspended in distilled water and dialyzed against0.9 per cent saline, until the dialysate was freeof ammonium sulfate. The gamma globulin solu-tion was then adjusted to a protein content of 20mg. per milliliter.
In the second experiment, young adult rabbitswere injected intravenously with 10 per cent chicklens extract 3 to 4 weeks before the onset of preg-nancy. When the amount of circulating lens anti-bodies was sufficientiy high (titer 1:10,000), therabbits were fertilized. A number of these animalswere operated on at the tenth day of pregnancy,the uterus exposed and quickly frozen with carbondioxide. In this manner, it was possible to peel offthe uterine wall and tissues surrounding the yolksac and to collect the yolk-sac fluid as a spheruleof ice. Subsequently, the concentration and num-ber of lens antibodies in the yolk-sac fluid weredetermined by means of Boyd's- precipitin tech-nique and OuchterlonyV5- 1G double diffusionmethod. In another series of rabbits, the fetuseswere removed at the twenty-eighth day of preg-nancy; fetal blood and amnionic fluid were col-lected and examined for the presence of anti-bodies. Simultaneously, the eyes of the fetuseswere removed, fixed in Bouin-Hollande, sectioned,stained, and examined for the presence of eyeabnormalities.
Results
The influence of lens antibodies on chickembryos. Although, sham operation, treat-ment with Earle's solution, or normal serumdid not affect the development of the 32hour chick embryos, treatment with lensantibodies either resulted in death of theembryo or caused degeneration of thebrain, retina, iris, and lens. In addition, itwas observed that a number of embryosshowed gross abnormalities such as anen-cephalus, rachiscliisis, cyclopia, and vaiiousdegrees of anophthalmia without any signof degeneration.
In the series of chicks treated with un-diluted antiserum, the cytotoxic effect wasusually of such a serious nature that theembryos died shortly after the beginningof the treatment. In those treated withdiluted antisera, it was noticed that thedegeneration was located more on the left
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018
398 hangman and MaiselInvestigative Ophthalmology
June 1962
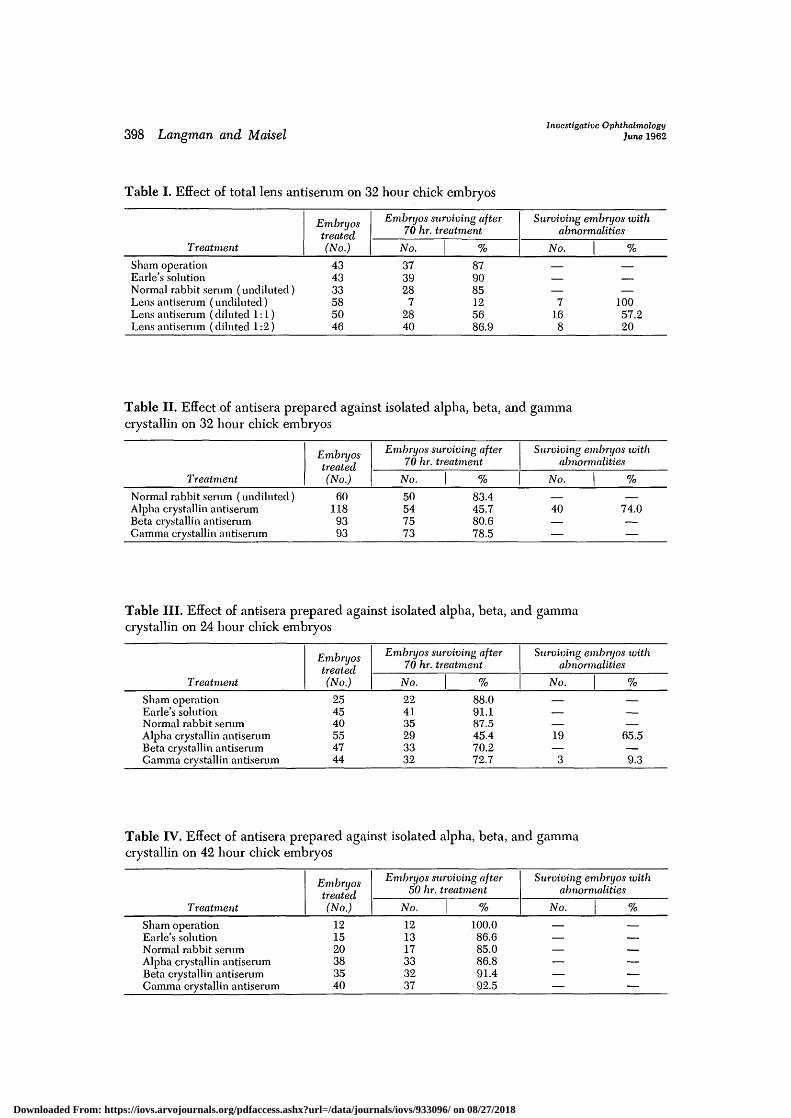
Table I. Effect of total lens antiserum on 32 hour chick
Treatment
Evibryostreated(No.)
embryos
Embryos surviving after70 hr. treatment
No. %
Surviving embryos withabnormalities
No. %
Sham operationEarle's solutionNormal rabbit serum (undiluted)Lens antiserum (undiluted)Lens antiserum (diluted 1:1)Lens antiserum (diluted 1:2)
434333585046
373928
72840
879085125686.9
—
—7
168
—
—10057.220
Table II. Effect of antisera prepared against isolated alpha, beta, and gammacrystallin on 32 hour chick embryos
Treatment
Normal rabbit serum (undiluted)Alpha crystallin antiserumBeta crystallin antiserumCamma crystallin antiserum
Embryostreated(No.)
601189393
Embryos surviving after70 hr. treatment
No.
50547573
%
83.445.780.678.5
Surviving embryos withabnormalities
No. %
40 74.0
Table III. Effect of antisera prepared against isolated alpha, beta, and gammacrystallin on 24 hour chick embryos
Treatment
Embryostreated(No.)
Embryos surviving after70 hr. treatment
No.
Surviving embryos withabnormalities
No.
Sham operationEarle's solutionNormal rabbit serumAlpha crystallin antiserumBeta crystallin antiserumGamma crystallin antiserum
254540554744
224135293332
88.091.187.545.470.272.7
——
19—
3
—
65.5—9.3
Table IV. Effect of antisera prepared against isolated alpha, beta, and gammacrystallin on 42 hour chick embryos
Treatment
Embryostreated(No.)
Embryos surviving after50 hr. treatment
No.
Surviving embryos xoithabnormalities
No.
Sham operationEarle's solutionNormal rabbit serumAlpha crystallin antiserumBeta crystallin antiserumGamma crystallin antiserum
121520383540
121317333237
100.086.685.086.891.492.5
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018
Volume 1Number 3 Lens antibodies and eye development 399
than on the right side of the head—a factattributed to the 90° anticlockwise rota-tion of the embryo shortly after the begin-ning of the treatment.
Since lens antiserum used in the firstexperiment contained antibodies against alllens proteins, in the following series theembryos were treated with antisera pre-pared against isolated alpha, beta, andgamma crystallin.
The results (Table II) clearly indicatethat alpha crystallin antiserum is respon-sible for the production of abnormalities,whereas beta and gamma crystallin anti-bodies do not interfere with embryonicdevelopment. The abnormalities found inthis series of experiments were similar tothose produced by total lens antiserum.
Table III shows the effect of antiseraprepared against isolated alpha, beta, andgamma crystallin on 24 hour embryos.
Despite the difference in age from thatof the embryos of the previous series, itis evident that, again, the alpha crystallinantibodies are responsible for the high rateof death and the resulting abnormalities inthe survivors. Since the abnormalities pro-duced in the 24 hour series were similarto those in the 32 hour series, it is sug-gested that the antibodies in both casesinterfere with the same developmentalprocesses.
The treatment of 42 hour embryos (TableIV) was ineffective. Despite the presenceof alpha and beta crystallin molecules invarious organs of the embryo during thetreatment period, no anomalies were seen.
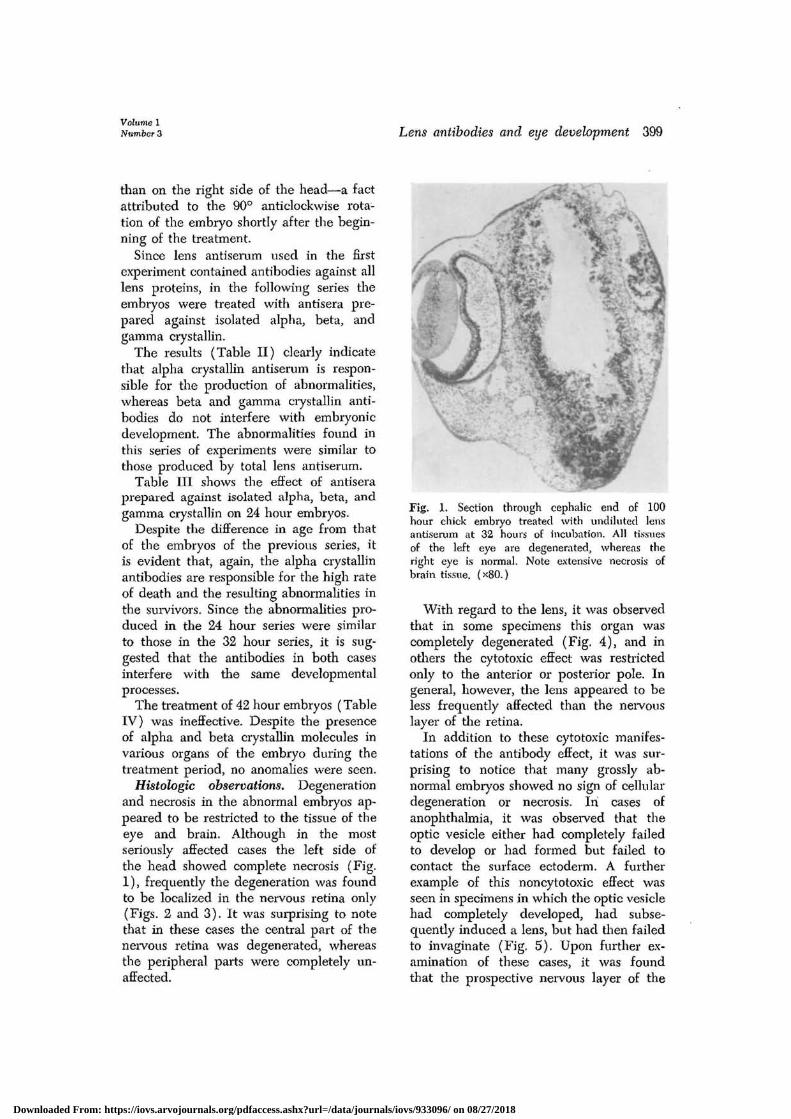
Histologic observations. Degenerationand necrosis in the abnormal embryos ap-peared to be restricted to the tissue of theeye and brain. Although in the mostseriously affected cases the left side ofthe head showed complete necrosis (Fig.1), frequently the degeneration was foundto be localized in the nervous retina only(Figs. 2 and 3). It was surprising to notethat in these cases the central part of thenervous retina was degenerated, whereasthe peripheral parts were completely un-affected.
Fig. 1. Section through cephalic end of 100hour chick embryo treated with undiluted lensantiserum at 32 hours of incubation. All tissuesof the left eye are degenerated, whereas theright eye is normal. Note extensive necrosis ofbrain tissue. (x80.)
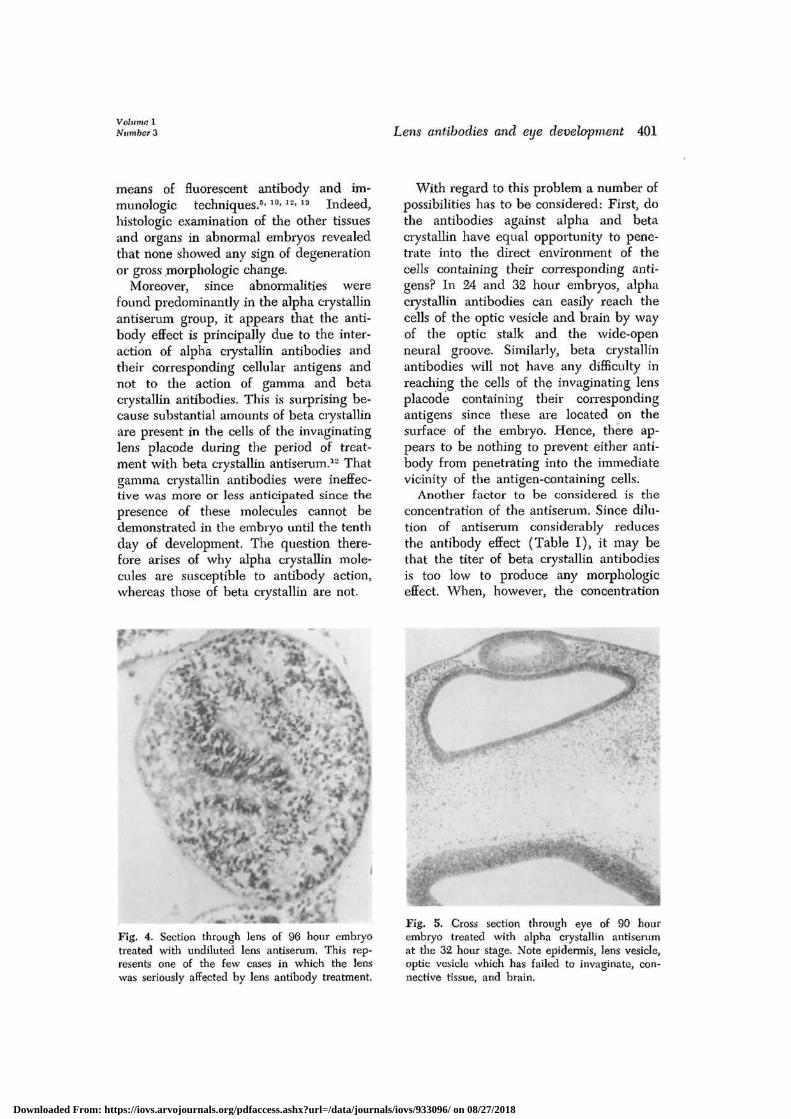
With regard to the lens, it was observedthat in some specimens this organ wascompletely degenerated (Fig. 4), and inothers the cytotoxic effect was restrictedonly to the anterior or posterior pole. Ingeneral, however, the lens appeared to beless frequently affected than the nervouslayer of the retina.
In addition to these cytotoxic manifes-tations of the antibody effect, it was sur-prising to notice that many grossly ab-normal embryos showed no sign of cellulardegeneration or necrosis. In cases ofanophthalmia, it was observed that theoptic vesicle either had completely failedto develop or had formed but failed tocontact the surface ectoderm. A furtherexample of this noncytotoxic effect wasseen in specimens in which the optic vesiclehad completely developed, had subse-quently induced a lens, but had then failedto invaginate (Fig. 5). Upon further ex-amination of these cases, it was foundthat the prospective nervous layer of the
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018

400 hangman and MaiselInvestigative Ophthalmology
June 1962
Fig. 2. Schematic representation of degenerationof nervous retina of 90 hour embryo treated withalpha crystallin antisemm at the 24 hour stage.Degeneration is restricted to central part ofnervous retina and has occurred shortly after lensinduction.
Fig. 3. Schematic representation of degenerationin nervous retina of 96 hour embryo, treated withalpha crystallin antiserum at the 32 hour stage.Degeneration is restricted to central part of nerv-ous retina and has occurred after complete in-vagination of optic vesicle.
retina did not show any mitotic activity,but had then formed pigment granules(Fig. 6).
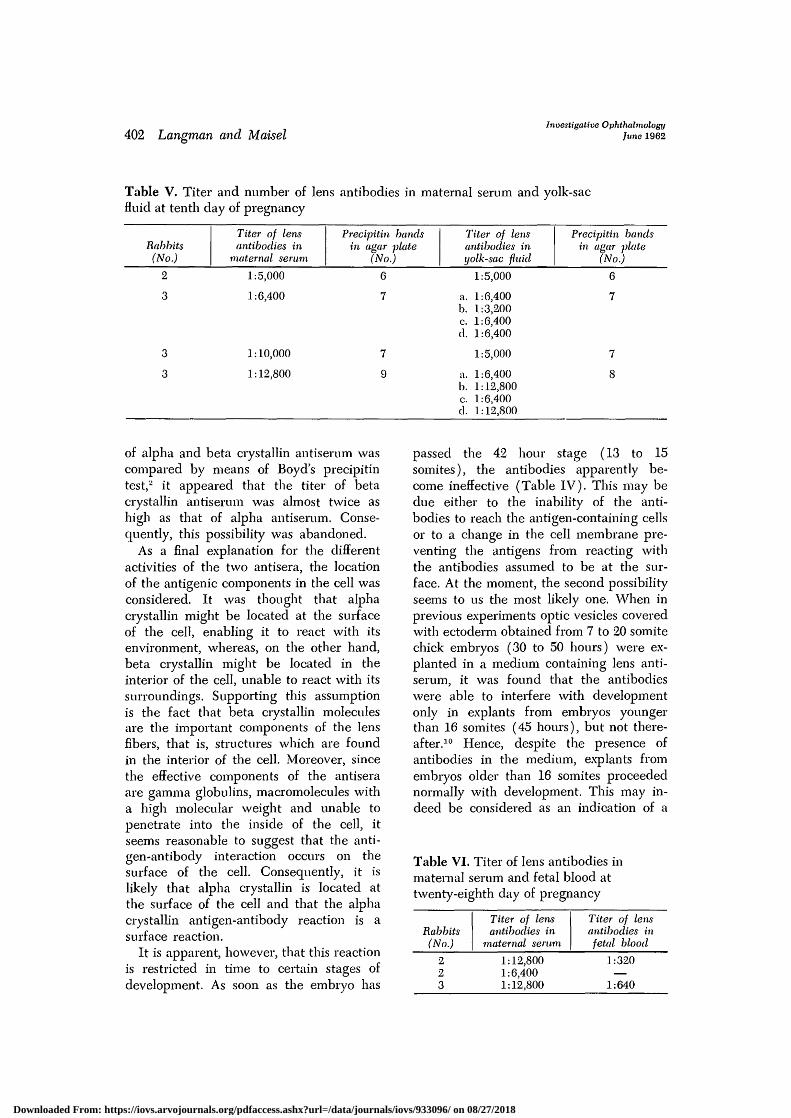
The influence of lens antibodies onrabbit embryos. Table V represents thedata found by testing yolk-sac fluid andmaternal serum for the presence of lensantibodies on the tenth day of pregnancy.
Although the titer of lens antibodies cir-culating in the rabbit blood stream variedfrom 1:5,000 to 1:12,800, the yolk-sac fluidshowed a titer varying from 1:3,200 to1:12,800—a fact which indicates that thelens antibodies had penetrated into theyolk-sac fluid in a concentration almostequal to that in the maternal serum. Bytesting yolk-sac fluid by the agar-diffusionmethod, 6 to 8 precipitin bands becamevisible, indicating that substantial amountsof 6 to 8 different lens antibodies werepresent in the yolk sac. Similarly, maternalserum showed the presence of 6 to 9 lensantibodies.
When, on the twenty-eighth day of
pregnancy, the titer of the lens antibodieswas determined, it was found to vary from1:6,400 to 1:12,800. In the fetal blood, how-ever, the concentration of lens antibodieswas low (1:640) or nonexistent. No lensantibodies were detected in the amnionicfluid.
Histologic examination of the fetal eyesdid not reveal any lens abnormalities. Itmay thus be concluded that despite thepresence of chick lens antibodies in thematernal blood stream throughout preg-nancy, in the yolk-sac fluid at the tenthday of embryonic development, and inthe fetal blood stream at the end ofpregnancy the antibodies did not interferewith lens development in the embryos.
Discussion
The abnormalities observed in the chickembryos are restricted to the tissues of theeye and brain, that is, those organs andtissues in which the presence of lens anti-gens has been demonstrated previously by
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018
Volume 1Number 3 Lens antibodies and eye development 401
means of fluorescent antibody and im-munologic techniques.5'10>]2-13 Indeed,histologic examination of the other tissuesand organs in abnormal embryos revealedthat none showed any sign of degenerationor gross morphologic change.
Moreover, since abnormalities werefound predominantly in the alpha crystallinantiserum group, it appears that the anti-body effect is principally due to the inter-action of alpha crystallin antibodies andtheir corresponding cellular antigens andnot to the action of gamma and betacrystallin antibodies. This is surprising be-cause substantial amounts of beta crystallinare present in the cells of the invaginatinglens placode during the period of treat-ment with beta crystallin antiserum.12 Thatgamma crystallin antibodies were ineffec-tive was more or less anticipated since thepresence of these molecules cannot bedemonstrated in the embryo until the tenthday of development. The question there-fore arises of why alpha crystallin mole-cules are susceptible to antibody action,whereas those of beta crystallin are not.
With regard to this problem a number ofpossibilities has to be considered: First, dothe antibodies against alpha and betacrystallin have equal opportunity to pene-trate into the direct environment of thecells containing their corresponding anti-gens? In 24 and 32 hour embryos, alphacrystallin antibodies can easily reach thecells of the optic vesicle and brain by wayof the optic stalk and the wide-openneural groove. Similarly, beta crystallinantibodies will not have any difficulty inreaching the cells of the invaginating lensplacode containing their correspondingantigens since these are located on thesurface of the embryo. Hence, there ap-pears to be nothing to prevent either anti-body from penetrating into the immediatevicinity of the antigen-containing cells.
Another factor to be considered is theconcentration of the antiserum. Since dilu-tion of antiserum considerably reducesthe antibody effect (Table I), it may bethat the titer of beta crystallin antibodiesis too low to produce any morphologiceffect. When, however, the concentration
• •f.
Fig. 4. Section through lens of 96 hour embryotreated with undiluted lens antiserum. This rep-resents one of the few cases in which the lenswas seriously affected by lens antibody treatment.
Fig. 5. Cross section through eye of 90 hourembryo treated with alpha crystallin antiserumat die 32 hour stage. Note epidermis, lens vesicle,optic vesicle which has failed to invaginate, con-nective tissue, and brain.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018
402 hangman and MaiselInvestigative Ophthalmology
June 1962
Table V. Titer and number of lens antibodies in maternal serum and yolk-sacfluid at tenth day of pregnancy
Rabbits(No.)
Titer of lensantibodies in
maternal serum
Precipitin bandsin agar •plate
(No.)
Titer of lensantibodies inyolk-sac fluid
Precipitin bandsin agar plate
(No.)
2
3
3
3
1:5,000
1:6,400
1:10,000
1:12,800
6
7
7
9
1:5,000
a. 1:6,400b. 1:3,200c. 1:6,400d. 1:6,400
1:5,000
a. 1:6,400b. 1:12,800c. 1:6,400d. 1:12,800
6
7
7
8
of alpha and beta crystallin antiserum wascompared by means of Boyd's precipitintest,2 it appeared that the titer of betacrystallin antiserum was almost twice ashigh as that of alpha antiserum. Conse-quently, this possibility was abandoned.
As a final explanation for the differentactivities of the two antisera, the locationof the antigenic components in the cell wasconsidered. It was thought that alphacrystallin might be located at the surfaceof the cell, enabling it to react with itsenvironment, whereas, on the other hand,beta crystallin might be located in theinterior of the cell, unable to react with itssurroundings. Supporting this assumptionis the fact that beta crystallin moleculesare the important components of the lensfibers, that is, structures which are foundin the interior of the cell. Moreover, sincethe effective components of the antiseraare gamma globulins, macromolecules witha high molecular weight and unable topenetrate into the inside of the cell, itseems reasonable to suggest that the anti-gen-antibody interaction occurs on thesurface of the cell. Consequently, it islikely that alpha crystallin is located atthe surface of the cell and that the alphacrystallin antigen-antibody reaction is asurface reaction.
It is apparent, however, that this reactionis restricted in time to certain stages ofdevelopment. As soon as the embiyo has
passed the 42 hour stage (13 to 15somites), the antibodies apparently be-come ineffective (Table IV). This may bedue either to the inability of the anti-bodies to reach the antigen-containing cellsor to a change in the cell membrane pre-venting the antigens from reacting withthe antibodies assumed to be at the sur-face. At the moment, the second possibilityseems to us the most likely one. When inprevious experiments optic vesicles coveredwith ectoderm obtained from 7 to 20 somitechick embryos (30 to 50 hours) were ex-planted in a medium containing lens anti-serum, it was found that the antibodieswere able to interfere with developmentonly in explants from embryos youngerthan 16 somites (45 hours), but not there-after.10 Hence, despite the presence ofantibodies in the medium, explants fromembryos older than 16 somites proceedednormally with development. This may in-deed be considered as an indication of a
Table VI. Titer of lens antibodies inmaternal serum and fetal blood attwenty-eighth day of pregnancy
Rabbits(No.)
Titer of lensantibodies in
maternal serum
Titer of lensantibodies infetal blood
1:12,8001:6,4001:12,800
1:320
1:640
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018

Volume 1Number 3 Lens antibodies and eye development 403
change in the cell membrane at approxi-mately the 16 somite stage, rendering thealpha crystallin molecules unable to reactwith the antibodies. Summarizing thus far,it may be concluded that the effects ob-served in our experiments result from anantigen-antibody interaction restricted toone particular molecule, occur only duringcertain stages of development, and in allprobability take place at the surface ofthe cell.
With regard to the degeneration andnecrosis observed, it was surprising thatmany of the 32 hour embryos showed theseeffects only on the left side of the head.Observing the embryos during the treat-ment, it was noticed that the antiserum,immediately after being administered,spread out toward the surrounding yolkand albumin. As the subcephalic pocketwith its elevated crescentic amnionic foldsforms a cavity under the head end of theembryo, some of the antiserum would col-lect in it and the head of the embryowould be partly or completely submergedin antiserum. Synchronously with theflexion of the embryo, rotation along itslength axis takes place. This rotation oc-curs in such a manner that the left side ofthe embryo becomes located deep in thesubcephalic pocket, and the right side isturned up toward the surface. This wouldimply that a few hours after the beginningof the treatment, the right side of the headwould turn up and become located abovethe antiserum level in the subcephalicpocket. It is thus thought that the rotationof the embryo may explain the frequentlyobserved left-sided necrosis of the head.
One of the surprising observations in ourexperiments is the fact that an antigen-antibody interaction apparently does notnecessarily lead to cellular degenerationand necrosis, but may interfere with celllife in a less damaging manner, such as byinhibition of growth or rechanneling of thenormal differentiation pattern. It is thoughtthat the essential feature of this differencemight be found in the concentration of theantibody molecules reacting with the cell.
Fig. 6. Top shows posterior pole of 96 hour lens.Some of the nuclei show moderate pyknosis. Indirect contact with posterior pole of lens isprospective nervous layer of the retina, which hasfailed to invaginate. This layer shows pigmentgranules.
If the antibodies present in the immediateenvironment of the cell are insufficient innumber to occupy all the available reactivesites of the antigen molecules, then thecell may still be able to survive but unableto continue with its normal morphologicdevelopment and function. Another possi-bility to be considered is the fact that theaffected antigen molecule is only one in agroup of different molecules, all buildingup the molecular population of the cell.Depending upon its "essentialness" for themetabolic processes of the cell and its im-portance in the interaction with other mole-cules, the antibody effect may then varyto a great extent. Indeed, in the eye ahighly diversified picture was seen as theresult of the antigen-antibody reaction.Although in some embryos the optic vesicledid not develop fully, resulting in anoph-thalmia, in other cases the optic vesicleinduced a lens but failed thereafter toproliferate and invaginate. In many of thesespecimens, however, the anterior part ofthe optic vesicle, destined to become thenervous retina, subsequently differentiated
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018

404 Lansman and MaiselInvestigative Ophthalmology
June 1962
in another direction, thereby forming pig-ment granules (Fig. 6). It is thought thatin these cases the affected antigen is unableto play its normal role in the interactionof molecules, thus permitting the cell tochannel its capacities in another direction.
When we started to inject our rabbitswith chick lens extract, it was thoughtthat these lens antigens were identical tothose of the rabbit lens. This was basedon the observation of Uhlenhuth,17 who,by testing bovine lens antiserum with lensextracts from animals of different species,noticed a cross reaction between this anti-serum and all lens extracts used. He there-fore concluded that the lens antigens invarious species are identical. This anti-genic identity has since been confirmedrepeatedly by other investigators, such asWoods and Burky,19 van Dold, Flossner,and Hutscher,18 and several others.
Recently developed immunologic proce-dures, such as the agar-diffusion techniqueof Ouchterlony, have provided us withnew tools to analyze the species specificityof organ and tissue antigens. To determinethe number of identical antigens in chickand rabbit lens, therefore, the latter wastested with chick lens antiserum, with theuse of the agar-diffusion test. When chicklens extract was tested with chick lensantiserum prepared in the rabbit, 6 to 9precipitin bands became visible. This indi-cates that 6 to 9 antibodies against thechick lens were circulating in the rabbitblood and even had penetrated into theyolk-sac fluid. However, to be effective,these antibodies must find correspondingantigens in the lens of the rabbit embryo.This apparently was not the case, because,when rabbit lens extract was tested withchick lens antiserum, only one precipitinband was formed,13 indicating that rabbitand chicken lens have only one antigenicfraction in common. Further experimentsshowed that this fraction was alpha crystal-lin and that it was only partially identicalto that of the chick. Hence, of all lens anti-bodies circulating in the rabbit, only onewill be able to interfere with lens forma-
tion in the rabbit, whereas the others,having no relation to rabbit lens antigens,are idle. Since the concentration of anti-bodies is of great importance to the pro-duction of abnormalities (Table I), andthe titer of the lens antibodies in the fetalcirculation was low (1:640), this mayimply that the concentration of the specificantibody was too low to effect formationof the embryonic lens in the treatedrabbits.
By comparison of the antibody treatmentin both series of experiments, it is evidentthat there is an important difference. Chickembiyos were treated with antiserum pre-pared by injecting chick lens extract intorabbits. The rabbit embryos, however, weretreated with chick lens extract likewiseprepared in the rabbit. In order to makeantibody treatment in both experimentssimilar, rabbit embryos should be treatedwith an antiserum prepared by injectionof rabbit lens extract into chicken. Sur-prisingly, this brings us back to the experi-ments of Guyer and Smith,7's who im-munized rabbits passively with an anti-serum prepared by injecting rabbit lensinto chicken and observed ocular mal-formations in the offspring. Huxley andSaunders,9 however, repeating this experi-ment, did not succeed in obtaining eyeanomalies in mammalian embryos by anti-body action. Recently, Miller14 immunizeda large series of rabbits with an antiserumprepared by injecting rabbit lens extractsinto chicken. When the eyes of 460 off-spring of the treated does were examined,only 4 had eye defects. The latter experi-ments, therefore, seem to indicate that cir-culating lens antibodies may interfere witheye development. In general terms, how-ever, considering the low percentage ofeye defects obtained in Miller's experiment,there does not seem to be much evidencethat lens antibodies can interfere with eyedevelopment in mammals.
We wish to thank Miss Jane Townsend for thepreparation of the histologic sections and technicalassistance in the experiments.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018

Volume 1Number 3 Lens antibodies and eye development 405
REFERENCES1. Barber, A. N., Willis, J., and Afeman, C :
Changes in the lens induced by maternalhypersensitivity in mice 51: 949, 1961.
2. Boyd, W. C : Fundamentals of immunol-ogy, London, 1956, Interscience Publishers,Ltd.
3. Brent, R. L., Averich, E., and Drapievvski,V. A.: Production of congenital malforma-tions using tissue antibodies. I. Kidney anti-sera, Proc. Soc. Exper. Biol. & Med. 106:523, 1961.
4. Clarke, W. M., and Fowler, I.: The inhibitionof lens-inducing capacity of the optic vesiclewith adult lens antisera, Develop. Biol. 2:155, 1960.
5. Fowler, I., and Clarke, W. M.: Developmentof anterior structures in the chick after directapplication of adult lens anitsera, Anat. Rec.136: 194, 1960.
6. Gluecksohn-Waelsch, S.: The effect of ma-ternal immunization against organ tissues onembryonic differentiation in the mouse, J.Embryol. & Exper. Morph. 5: 83, 1957.
7. Guyer, M. F., and Smith, E. A.: Studies oncytolysin. I. Some prenatal effects of lensantibodies, J. Exper. Zool. 26: 63, 1918.
8. Guyer, M. F., and Smith, E. A.: Furtherstudies on inheritance of eye defects inducedin rabbits, J. Exper. Zool. 38: 449, 1924.
9. Huxley, J. S., and Saunders, A. M. Carr: Ab-sence of prenatal effects of lens antibodiesin rabbits, J. Exper. Biol. 1: 216, 1923.
10. Langman, J.: The first appearance of specific
antigens during induction of the lens, J. Em-bryol. & Exper. Morph. 7: 193, 1959.
11. Langman, J.: The effect of lens antiserumon chick embryos, Anat. Rec. 137: 135,1960.
12. Maisel, H., and Langman, J.: An immuno-embryological study on the chick lens, J.Embryol. & Exper. Morph. 9: 191, 1961.
13. Maisel, H., and Langman, J.: Chick lens pro-teins in various tissues of the eye and in thelens of animals throughout the vertebrateseries, Anat. Rec. 140: 183, 1961.
14. Miller, W. J.: Antilens sera as a mutagen inrabbits, J. Exper. Zool. 137: 463, 1958.
15. Ouchterlony, O.: Antigen-antibody reactionsin gels, Acta path, et microbiol. scandinav.32: 231, 1953.
16. Ouchterlony, O.: Antigen-antibody reactionsin gels and the practical application of thisphenomenon in the laboratory diagnosis ofdiphtheria, Thesis, Karolinska Institutet,Stockholm, 1949.
17. Uhlenhuth, P. T.: Zu Lehre von der Unter-scheidung verschiedener Eiweissarten mitHilfe spezifischer Sera, Festschrift zum 60.Geburtstage v. Robert Kock, Jena, 1903,Gustave Fischer Verlag, pp. 48-74.
18. Van Dold, H., Flossner, D., and Hutscher,F.: Biologische Untersuchungen iiber dieLinsen-Eiweisskorper, Ztschr. Immunitatsforschung u. exper. Therap. 46: 36, 1926.
19. Woods, A. C , and Burky, E. L.: Lens proteinand its fractions. Preparations and imrauno-logical and chemical properties, J. A. M. A.89: 102, 1927.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933096/ on 08/27/2018