
Journal of Crustacean Biology Advance Access published 6 ...
13
The Crustacean Society © The Author(s) 2019. Published by Oxford University Press on behalf of The Crustacean Society. All rights reserved. For permissions, please e-mail: [email protected] Social behavior of the Pederson cleaner shrimp Ancylomenes pedersoni (Chace, 1958) (Decapoda: Caridea: Palemonidae) in the Caribbean Sea: size- and gender-based dominance hierarchy Jessica A. Gilpin and Nanette E. Chadwick Department of Biological Sciences, 101 Rouse Life Sciences Building, Auburn University, Auburn, AL 36849, USA Corresponding author: J.A. Gilpin; e-mail: [email protected] (Received 5 June 2019; accepted 10 October 2019) ABSTRACT Social interactions among Pederson cleaner shrimp Ancylomenes pedersoni (Chace, 1958), an as- sociate of sea anemones, likely affect cleaning services to reef fishes, but major aspects of their social group structure and behavior remain unknown. We hypothesized that Pederson shrimp form dominance hierarchies expressed through size- and gender-based behavioral inter- actions. Observation of 116 shrimp on coral reefs at St. Thomas, US Virgin Islands, revealed that they formed social groups of ~3 or 4 individuals (range 1–9) per host sea anemone, as a mix of large females, medium-size males, and/or small juveniles. Social groups were not rigidly size-structured, in that body size ratio between adjacent individuals did not differ sig- nificantly from random. Individuals, however, were spatially structured such that distance be- tween shrimp and the tentacles of the host anemone decreased significantly with shrimp body size; large individuals occupied the anemone tentacle crown, whereas smaller shrimp perched on reef or sand substrates adjacent to the anemone. During laboratory experiments, shrimp exhibited five types of exploitation competition behaviors toward client fish models (signal, approach, clean, no response, retreat), and four types of interference competition behaviors toward conspecific shrimp (approach, attack, no response, retreat). Large females more fre- quently signaled toward, approached, and cleaned fish models than did males and juveniles, which did not respond or retreated. Females also approached and attacked conspecifics more than did males and juveniles, which retreated at frequencies that increased with decreasing body size. We conclude that Pederson shrimp form gender and size-based hierarchies in which large females monopolize food resources (ectoparasites on client fishes), and interfere with re- source access by smaller males and juveniles. These behaviors cause them to spatially partition microhabitats on anemones, and hypothetically could lead to males and juveniles dispersing among anemones more frequently than females. These shrimp social interactions may thus enhance spatial and temporal variation among cleaning stations, with consequences for the health and mobility of reef fishes. Key words: anemone shrimps, aggression, behavior, cleaning symbiosis, coral reefs, coral reef fishes, habitat segregation, sea anemones INTRODUCTION Corkscrew sea anemones Bartholomea annulata (Le Sueur, 1817) are common, large-bodied Caribbean cnidarians that host a variety of associated organisms on, among, and beneath their tentacles (Mahnken, 1972; Brooker et al., 2019). The Pederson cleaner shrimp Ancylomenes (Periclimenes) pedersoni (Chace, 1958) are obligate associates of corkscrew anemones. The shrimp also associates with other Caribbean sea anemones, including rosetip anemones Condylactis gigantea Weinland, 1860, branching anemones Lebrunia danae (Duchassaing & Michelotti, 1861), and sun anemones Stichodactyla helianthus (Ellis, 1768). The Pederson shrimp perch on the oral discs and tentacles of corkscrew anemones (Mahnken, 1972) and occupy reef crevices where these anemones attach Journal of Crustacean Biology Advance Access published 6 November 2019 Journal of Crustacean Biology Journal of Crustacean Biology 40(1), 24–36, 2020. doi:10.1093/jcbiol/ruz081 Downloaded from https://academic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn University user on 13 January 2020
Transcript of Journal of Crustacean Biology Advance Access published 6 ...

The Crustacean Society
© The Author(s) 2019. Published by Oxford University Press on behalf of The Crustacean Society. All rights reserved. For permissions, please e-mail: [email protected]
Social behavior of the Pederson cleaner shrimp Ancylomenes pedersoni (Chace, 1958) (Decapoda:
Caridea: Palemonidae) in the Caribbean Sea: size- and gender-based dominance hierarchy
Jessica A. Gilpin and Nanette E. ChadwickDepartment of Biological Sciences, 101 Rouse Life Sciences Building, Auburn University, Auburn, AL 36849, USA
Corresponding author: J.A. Gilpin; e-mail: [email protected]
(Received 5 June 2019; accepted 10 October 2019)
ABSTRACT
Social interactions among Pederson cleaner shrimp Ancylomenes pedersoni (Chace, 1958), an as-sociate of sea anemones, likely affect cleaning services to reef fishes, but major aspects of their social group structure and behavior remain unknown. We hypothesized that Pederson shrimp form dominance hierarchies expressed through size- and gender-based behavioral inter-actions. Observation of 116 shrimp on coral reefs at St. Thomas, US Virgin Islands, revealed that they formed social groups of ~3 or 4 individuals (range 1–9) per host sea anemone, as a mix of large females, medium-size males, and/or small juveniles. Social groups were not rigidly size-structured, in that body size ratio between adjacent individuals did not differ sig-nificantly from random. Individuals, however, were spatially structured such that distance be-tween shrimp and the tentacles of the host anemone decreased significantly with shrimp body size; large individuals occupied the anemone tentacle crown, whereas smaller shrimp perched on reef or sand substrates adjacent to the anemone. During laboratory experiments, shrimp exhibited five types of exploitation competition behaviors toward client fish models (signal, approach, clean, no response, retreat), and four types of interference competition behaviors toward conspecific shrimp (approach, attack, no response, retreat). Large females more fre-quently signaled toward, approached, and cleaned fish models than did males and juveniles, which did not respond or retreated. Females also approached and attacked conspecifics more than did males and juveniles, which retreated at frequencies that increased with decreasing body size. We conclude that Pederson shrimp form gender and size-based hierarchies in which large females monopolize food resources (ectoparasites on client fishes), and interfere with re-source access by smaller males and juveniles. These behaviors cause them to spatially partition microhabitats on anemones, and hypothetically could lead to males and juveniles dispersing among anemones more frequently than females. These shrimp social interactions may thus enhance spatial and temporal variation among cleaning stations, with consequences for the health and mobility of reef fishes.
Key words: anemone shrimps, aggression, behavior, cleaning symbiosis, coral reefs, coral reef fishes, habitat segregation, sea anemones
INTRODUCTION
Corkscrew sea anemones Bartholomea annulata (Le Sueur, 1817) are common, large-bodied Caribbean cnidarians that host a variety of associated organisms on, among, and beneath their tentacles (Mahnken, 1972; Brooker et al., 2019). The Pederson cleaner shrimp Ancylomenes (Periclimenes) pedersoni (Chace, 1958) are obligate
associates of corkscrew anemones. The shrimp also associates with other Caribbean sea anemones, including rosetip anemones Condylactis gigantea Weinland, 1860, branching anemones Lebrunia danae (Duchassaing & Michelotti, 1861), and sun anemones Stichodactyla helianthus (Ellis, 1768). The Pederson shrimp perch on the oral discs and tentacles of corkscrew anemones (Mahnken, 1972) and occupy reef crevices where these anemones attach
applyparastyle "fig//caption/p[1]" parastyle "FigCapt"applyparastyle "fig" parastyle "Figure"
Journal of Crustacean Biology Advance Access published 6 November 2019
Journal of
Crustacean BiologyJournal of Crustacean Biology 40(1), 24–36, 2020. doi:10.1093/jcbiol/ruz081
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

SOCIAL BEHAVIOR OF CLEANER SHRIMP
25
their pedal disks (Limbaugh et al., 1961). These large anemones serve as visual cues that attract reef fishes, which then engage in cleaning interactions with the shrimp (Huebner & Chadwick, 2012a). Client fishes may in turn provide excreted nutrients that benefit the anemone and its endosymbiotic microalgae (Cantrell et al., 2015). On branching anemones L. danae, the shrimp typ-ically perch on reef substrate within a few centimeters of the diurnally-expanded fronds of host anemones, or rest atop the fronds (Herrnkind et al., 1976). The shrimp-anemone complex forms a cleaning station, where shrimp consume parasites and decayed tissue from the body surfaces of fishes that visit the anemone (Bunkley-Williams & Williams, 1998; Côté, 2000). Pederson shrimp aggregate in social groups (as defined by Costa & Fitzgerald, 2005) ranging from 1–12 shrimp per host anemone (Mahnken, 1972, Titus et al., 2015), with individuals in each so-cial group varying in body size (Mahnken, 1972; Mascaró et al., 2012). Researchers have speculated that the number of Pederson shrimp in each social group depends on habitat space availability on the anemone host, as well as on the abundance of the local shrimp population; space limitation (host anemone abundance and body size) is thus thought to be more important in con-straining the abundance of this species than in other non-group-forming shrimps associated with Caribbean anemones (Mahnken, 1972; Nizinski, 1989). Recent analyses have confirmed that larger corkscrew anemones, which provide more habitat space, host sig-nificantly more Pederson shrimp than do smaller anemones (Titus et al., 2017; Huebner et al., in press). In contrast, the number of shrimp per anemone does not appear to vary with their frequency of cleaning or cheating behaviors toward fish clients (Titus et al., 2015, 2019). A size-dependent dominance hierarchy has been pro-posed to structure social groups of Pederson shrimp (Mahnken, 1972), but no quantitative data support this idea.
Many studies have described the important functional role of cleaning stations in coral reef ecosystems, including how cleaner organisms enhance health and abundance of reef fishes (Bunkley-Williams & Williams, 1998; Bshary, 2003; Becker & Grutter, 2004, 2005; Bshary et al., 2007), and how fishes use cleaning stations (Chapuis & Bshary, 2009; Huebner & Chadwick, 2012b; Titus et al., 2015, 2017, 2019; Caves et al., 2018). Analyses of behavioral interactions between cleaner shrimps and their cnidarian hosts, and of cleaner shrimp life histories, are nevertheless lacking outside of host preference and acclimation studies (Levine & Blanchard, 1980; Guo et al., 1996; Silbiger & Childress, 2008; Mascaró et al., 2012) and one population study of non-cleaner shrimp Periclimenes anthophilus (Holthuis & Eibl-Eibesfeldt, 1964) which are closely re-lated to Pederson shrimp (Nizinski, 1989). The only comprehen-sive life history study of Pederson shrimp indicates that this species has separate sexes in which females are larger than males, and that individuals live for about one year under laboratory condi-tions (Gilpin & Chadwick, 2017). The population size structure of this species is dominated by small individuals, with the number of individuals in each size class decreasing exponentially with body size and age (Gilpin & Chadwick, 2017). No studies have quanti-fied social group composition in Pederson cleaner shrimp, or the spatial structure of groups in terms of how shrimp body size and gender vary among microhabitats on host anemones. Research on other Caribbean anemone shrimps that form social groups indi-cates that relatively large individuals occupy more central posi-tions on anemone hosts, whereas small conspecifics are relegated to the anemone margins (Colombara et al., 2017).
Interference competition may occur among Pederson shrimp in social groups (Mascaró et al., 2012), but aggressive behavior in this species has not yet been described. Information about the level of aggressiveness of each individual (willingness to fight with other shrimp) would contribute to understanding the extent to which interference competition structures social dominance hierarchies in the shrimp. Knowledge about aggressiveness is particularly im-portant in invertebrate social hierarchies, because responses to
the “aggressive state” of other group members may be a stronger determinant of social status than individual recognition (Winston & Jacobson, 1978). Aggressive interactions are known to struc-ture arthropod social hierarchies, including crickets (Alexander, 1961), hermit crabs (Courchesne & Barlow, 1971), and crayfishes (Bovbjerg, 1953; Rubenstein & Hazlett, 1974; Copp, 1986; Issa et al., 1999).
In addition to aggressive state, relative body size also contrib-utes to determination of the outcome of conflicts among conspe-cifics (Bovbjerg, 1953; Ranta & Lindström, 1992; Pavey & Fielder, 1996). Large Pederson shrimp may occupy more central positions on anemones than do small shrimp (JG, personal observation), potentially indicating a spatially structured social hierarchy based in part on relative body size, similar to the pattern observed for other anemone shrimps (Colombara et al., 2017). The prominent habitats occupied by large individuals could be due not only to interference with smaller shrimp, but also to exploitation competi-tion for access to client fishes and thus food resources in the form of parasites and decayed fish tissue (Mascaró et al., 2012). The mechanism of exploitation competition utilized by large shrimp is potentially via signaling to and approaching fish clients more fre-quently than do small shrimp, which would allow them to monop-olize food resources in the form of fish ectoparasites before small individuals can access them.
We hypothesized that Pederson shrimp form social dominance hierarchies based on the outcomes of size- and gender-based be-havioral interactions. Our objectives were to test this hypothesis by quantifying 1) the structure of social groups observed under field conditions, and 2) the behaviors of shrimp in experimental social groups under laboratory conditions.
MATERIALS AND METHODS
Field observations
Field observations were conducted during July 2015 and August 2016 on patch reefs in Brewers Bay, St. Thomas, U.S. Virgin Islands, at ~6 m depth (18°19′N, 64°59′W; detailed site descrip-tions in Gilpin & Chadwick, 2017; Huebner & Chadwick, 2012a, b; O’Reilly et al., 2018; Huebner et al., in press). We conducted preliminary observations to locate reef areas that were within or near those examined in the above studies, and which contained high abundances of corkscrew sea anemones and associated Pederson shrimp. Other species of large coral reef sea anem-ones were rare at the study site; spotted cleaner shrimp Periclimenes yucatanicus (Ives, 1891) also occupied the corkscrew anemones, but were rare (N = 1, 0.9% of anemones occupied in 2105; N = 13, 9.3% anemones occupied in 2016). Other crustacean associates of the sea anemones were not censused (but see Huebner et al., in press). We then deployed six 50 m transects (three each year) hap-hazardly within the selected reef areas. As such, the data reported here represent patterns in areas of potentially maximal abun-dance for Pederson shrimp in this bay. An area extending 1 m to the right and left of each transect line was examined, resulting in a 100 m2 area examined per 50 m belt transect (600 m2 total).
For each corkscrew sea anemone observed inside the tran-sects (N = 109, 40 with Pederson shrimp (2015); N = 140, 76 with Pederson shrimp (2016)), we approached close enough to detect the associated shrimp (N = 135 Pederson shrimp in 2015; N = 154 in 2016), but took care to not touch or otherwise disturb the shrimp or anemones. We recorded the number of shrimp on each anemone, and the type of microhabitat that each occupied on the anemone body as classified into five zones: 1) anemone column, 2) inner tentacle crown, 3) outer tentacle crown, 4) hard substrate adjacent to anemone, and 5) soft substrate adjacent to anemone after Khan et al., 2004; Colombara et al., 2017; Huebner et al., in press (Figs. 1, 2). We also recorded the distance to the nearest anemone tentacle tip for shrimp that occurred on the
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

J. A. GILPIN AND N. E. CHADWICK
26
substrate adjacent to the anemone using a Vernier caliper meas-ured to the nearest mm. Body size of anemones was determined using a Vernier caliper, without touching the anemone to avoid tentacle contraction. Tentacle crown length and width were measured in mm for calculation of tentacle crown surface area
(TCSA; after Hirose, 1985; Hattori, 2002; Huebner et al., 2012; O’Reilly & Chadwick, 2017; Dixon et al., 2017). We then col-lected all Pederson shrimp on each anemone using a small hand net, and transferred them underwater into a plastic bag filled with seawater, so that their body sizes could be accurately measured. We used a caliper to measure shrimp body size (total length (TL) as anterior tip of rostrum to posterior tip of telson), through the clear plastic of the bag and recorded the presence of any oocytes or developing embryos to indicate female status (after Gilpin & Chadwick, 2017). All individuals that lacked oocytes or embryos and were > 13.1 mm TL were classified as males, and those smaller were classified as juveniles, based on previous results (fe-males 13.1–31.85 mm, males 13.1–21.41 mm, and juveniles < 13.0 mm TL (described as PMTL in Gilpin & Chadwick, 2017). Carapace length (CL) was not measured because it was diffi-cult to recognize in the field. We manually measured body size rather than taking photographs for later analysis, because the shrimp were too small and translucent to easily visualize their body dimensions in photographs taken against reef and sand backgrounds. We released each group of shrimp back into their original host anemone after measurements. Each shrimp was in a plastic bag for < 5 min and immediately re-associated with its host anemone upon release.
A different reef area was examined in Brewers Bay, US Virgin Islands during each of the two survey years. Most corkscrew anemones (O’Reilly & Chadwick, 2017) and Pederson shrimp live for about one year (Gilpin & Chadwick, 2017), so the data from 2015 and 2016 were considered to be independent samples, resulting in total sample sizes of 116 anemones and 289 shrimp examined (see Gilpin & Chadwick, 2017 for details of field sam-pling methods).
Laboratory experiments
Laboratory behavioral experiments were conducted at Auburn University during August 2016 to May 2017. We used shrimp that had been cultured for a multi-year laboratory study on life his-tory patterns (see Gilpin & Chadwick, 2017 for details of shrimp collection and culture). We redistributed 28 individuals that re-mained from that study among six tanks (40 l each) to create social groups of 4–5 individuals per tank for behavioral studies (similar to social group sizes in the field; Mahnken, 1972). Groups of at least four individuals per tank were set up, because preliminary observations indicated that shrimp in smaller groups rarely en-countered each other in the tanks and did not engage in many social interactions. Less than six individuals were used per tank to facilitate individual identification based on relative body sizes. Carapace length (CL) was used as a body-size measure of labora-tory shrimp because it is possible to measure CL accurately in the laboratory, it correlates significantly with TL, and is a standard parameter for crustacean body size (Bauer, 1986, 2004). Each so-cial group included both females and males, but not always juven-iles depending on availability (similar to social group composition in the field): 1–4 females, one male, and 0–2 juveniles per group (reproductive status determined after Gilpin & Chadwick, 2017; females 3.4–6.1 mm, males 3.4–4.1 mm, juveniles < 3.3 mm CL). Shrimp were measured initially for carapace length (CL to mm) using a Vernier caliper, then all females were assigned to a body size class (large > 5.0 mm, medium 4.2–4.9 mm; small < 4.2 mm CL). In August 2016, we formed social groups by ran-domly selecting large, medium, and small females from the pool of individuals until we obtained three groups each with a clear size hierarchy of alpha, beta, gamma, and delta females (classi-fied as the largest, next largest, and next largest individuals in the group; after Lee & Fielder 1982), and one male (3.3–4.0 mm CL; see Gilpin & Chadwick, 2017). All individuals not used in these three initial groups (including juveniles) were then distributed among the three remaining tanks to create similar social groups
Figure 1. Microhabitats on the corkscrew sea anemone Bartholomea annulata occupied by associated crustaceans, classified as five zones on or near the host anemone body: 1, anemone column; 2, inner tentacle crown, 3, outer tentacle crown; 4, hard substrate adjacent to anemone; 5, soft substrate ad-jacent to anemone. Shown are typical zones occupied by four species of obligate crustacean associates: snapping shrimp Alpheus armatus (zone 1), anemone mysid shrimp Heteromysis actiniae (zone 2), spotted anemone shrimp Periclimenes yucatanicus (zone 3), and Pederson cleaner shrimp Ancylomenes pedersoni (zones 3, 4, and 5), on coral reefs at St. Thomas, U.S. Virgin Islands. A typical social group of five individuals that segregate microhabitat by relative body size is shown for A. pedersoni: largest individual at outer edge of zone 3, perched on the anemone tentacle tips; all other group members in zones 4 or 5, with second largest individual near but not touching the anemone, and smaller individuals at greater distances up to ~10 cm from the host. Diagram by L. Huebner and JAG.
Figure 2. Photograph of a social group of five Pederson cleaner shrimp Ancylomenes pedersoni associated with a corkscrew sea anemone Bartholomea annulata (in crevice at center) on a coral reef at Saba, Lesser Antilles. Note that the four large individuals (1–4) occupy habitat touching the outer anemone tentacles (zone 3; Fig. 1); smaller individual (5) occurs on hard substrate not contacting the anemone (~2.5 cm distant, zone 4). The four large shrimp are facing outwards (paired violet-banded chelae extending away from anemone center) and are spaced evenly around the periphery of the anemone tentacle crown, with their paired long white antennae ex-tending backwards and almost touching, indicating a regular distributional pattern potentially mediated through behavioral interactions (see Table 1). Photograph by JAG.
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020
SOCIAL BEHAVIOR OF CLEANER SHRIMP
27
(six social groups in total). During random assignment of females to social groups, individuals were assigned to the same group only if they differed in relative body size by at least 0.2 mm in carapace length between adjacent group members (similar to minimum dif-ference in body size between adjacent group members in the field: x ± SD = 0.58 + 0.53 mm CL (0.02–2.87 mm). The resulting social groups consisted of either 1) five individuals (N = 3 tanks of 15 shrimp total), four females (alpha, beta, gamma, and delta, in descending order of body size), and one male or 2) four individ-uals (N = 3 tanks of 12 shrimp total), either three females (alpha, beta, and gamma) and one male (one tank), two females (alpha and beta), one male and one juvenile (one tank), or one female, one male, and two juveniles (one tank).
Each tank was lit by ceiling lights in the culture room (~12:12 h light:dark cycle), and contained an external hanging filter driven by a small pump (AQUA-Tech® Power Filter 5-15, Tetra, Blacksburg, VA, USA). High-output tank lights were not used, in contrast to previous methods for anemones and shrimp cul-tured together in the laboratory, because high irradiance was not needed to support sea anemone photosynthesis (see Cantrell et al. 2015). A small unglazed clay pot (~10 × 10 × 10 cm) was placed in the middle of each tank to provide three-dimensional structure in the form of an anemone-sized shelter space. Individuals were fed every other day (~3× per week) with fish food pellets (Formula One and Formula Two Pellets, Ocean Nutrition, Newark, CA,
USA). The shrimp grew, produced eggs, and released larvae under these culture conditions, indicating healthy physiological condition (after Gilpin & Chadwick, 2017).
Social groups were allowed to acclimate for two weeks, after which we conducted the first set of behavioral observations during August-November 2016. Some shrimp mortality occurred during this 4 m period, mostly due to natural causes (senescence followed by end of natural lifespan; Gilpin & Chadwick, 2017), resulting in 14 individuals remaining by December. During January 2017, 10 of the remaining individuals were moved between two tanks to create new social groups, each consisting of five individuals: four females (alpha, beta, gamma, and delta) and one male. Groups for the second set of observations were selected to be similar in so-cial group size, gender composition, and relative body size in each group, as those formed during the first set, but to contain different combinations of individuals. The remaining four individuals were placed in a third tank and not included in the second set of obser-vations, because they did not form a social group representative of field patterns (all four individuals were male). As such, shrimp used during the second set of behavioral observations (January-April 2017) comprised a subset of those used during the first set (August-December 2016), but in different social group combin-ations: three shrimp remained in the same type of social group and the same relative size position within that group, with the re-maining seven shrimp in different relative body size positions in
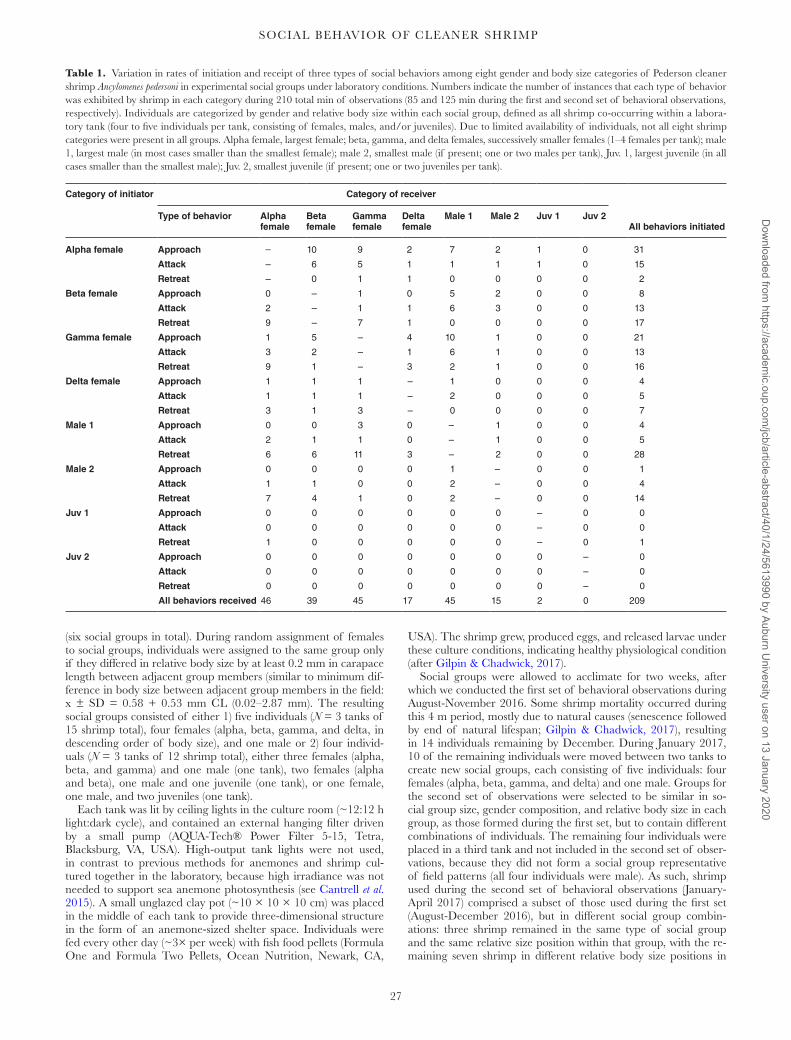
Table 1. Variation in rates of initiation and receipt of three types of social behaviors among eight gender and body size categories of Pederson cleaner shrimp Ancylomenes pedersoni in experimental social groups under laboratory conditions. Numbers indicate the number of instances that each type of behavior was exhibited by shrimp in each category during 210 total min of observations (85 and 125 min during the first and second set of behavioral observations, respectively). Individuals are categorized by gender and relative body size within each social group, defined as all shrimp co-occurring within a labora-tory tank (four to five individuals per tank, consisting of females, males, and/or juveniles). Due to limited availability of individuals, not all eight shrimp categories were present in all groups. Alpha female, largest female; beta, gamma, and delta females, successively smaller females (1–4 females per tank); male 1, largest male (in most cases smaller than the smallest female); male 2, smallest male (if present; one or two males per tank), Juv. 1, largest juvenile (in all cases smaller than the smallest male); Juv. 2, smallest juvenile (if present; one or two juveniles per tank).
Category of initiator Category of receiver
All behaviors initiatedType of behavior Alpha
femaleBeta female
Gamma female
Delta female
Male 1 Male 2 Juv 1 Juv 2
Alpha female Approach – 10 9 2 7 2 1 0 31
Attack – 6 5 1 1 1 1 0 15
Retreat – 0 1 1 0 0 0 0 2
Beta female Approach 0 – 1 0 5 2 0 0 8
Attack 2 – 1 1 6 3 0 0 13
Retreat 9 – 7 1 0 0 0 0 17
Gamma female Approach 1 5 – 4 10 1 0 0 21
Attack 3 2 – 1 6 1 0 0 13
Retreat 9 1 – 3 2 1 0 0 16
Delta female Approach 1 1 1 – 1 0 0 0 4
Attack 1 1 1 – 2 0 0 0 5
Retreat 3 1 3 – 0 0 0 0 7
Male 1 Approach 0 0 3 0 – 1 0 0 4
Attack 2 1 1 0 – 1 0 0 5
Retreat 6 6 11 3 – 2 0 0 28
Male 2 Approach 0 0 0 0 1 – 0 0 1
Attack 1 1 0 0 2 – 0 0 4
Retreat 7 4 1 0 2 – 0 0 14
Juv 1 Approach 0 0 0 0 0 0 – 0 0
Attack 0 0 0 0 0 0 – 0 0
Retreat 1 0 0 0 0 0 – 0 1
Juv 2 Approach 0 0 0 0 0 0 0 – 0
Attack 0 0 0 0 0 0 0 – 0
Retreat 0 0 0 0 0 0 0 – 0
All behaviors received 46 39 45 17 45 15 2 0 209
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

J. A. GILPIN AND N. E. CHADWICK
28
the newly-created groups. Social groups were created using the same random choice method as employed in the first set.
We categorized during the first and second sets of observa-tions five types of exploitation competition behaviors that shrimp exhibited toward a client fish model (see below), ordered from subordinate to dominant behaviors: 1) retreat from fish model (locomote away), 2) no response (no change in position), 3) signal toward model (slowly sway body from side to side, and/or rap-idly vibrate antennae; after Becker et al., 2005; Chapuis & Bshary, 2010), 4) approach (locomote toward), 5) clean (mount fish model and pick at it with chelae while climbing over the model surface; after Mascaró et al., 2012), and four types of interference com-petition behaviors that shrimp exhibited toward conspecifics, also ordered from subordinate to dominant behaviors: 1) retreat from conspecific (locomote away), 2) no response (no change in pos-ition), 3) approach (locomote toward), and 4) attack (make con-tact with conspecific, pinch with chelae (after Bovbjerg, 1956; Issa et al., 1999).
We created a fish model by downloading from the internet and printing onto paper a high resolution color photograph of a common Caribbean fish client (the blue tang, Acanthurus coeruleus Bloch & Schneider, 1801) (after Huebner & Chadwick 2012a). The photograph was printed at a small adult body size for this species (13.5 cm TL; Humann & DeLoach, 2002) then laminated with clear, stiff plastic to render it water resistant. A thin wire was attached to the top of the model so that it could be lowered into the tanks during behavioral trials. This type of two-dimensional photographic model has been used to evoke behavioral responses by cleaner fishes (Cheney et al., 2009), and was selected as likely to evoke natural responses by Pederson shrimp because the shrimp are color blind and have coarse spatial resolution (Caves et al., 2016), which may cause them to recognize two-dimensional photographic models as fish clients (Caves et al., 2018). Preliminary observations indicated that shrimp responded readily to the model and exhibited behaviors similar to their natural interactions with client fishes (Caves et al., 2018).
We observed all shrimp in each tank once per week for 17 weeks (August-November 2016) during the first set of behavioral obser-vations. Every week, we selected each of the six tanks randomly for observations either with the fish model present (four tanks) or absent (two tanks). We included a more limited number of obser-vations in the absence of the fish model (only two tanks per week), because we were primarily interested in social behaviors of the shrimp in the context of their role as cleaning organisms. Thus, observations were made weekly on each of six tanks for 17 weeks (total of 102 tank observations) and on each of the 28 shrimp in those tanks (~4.5 shrimp per tank) for 17× per shrimp (15× with the fish model present and 2× with the model absent), resulting in a total of 459 shrimp behavioral observations during the first set. We observed all shrimp in each tank ~2 or 3× each week for 15 weeks (January-April 2017) during the second set of observa-tions, because there were only two tanks to observe. We observed both tanks with the fish model present every week, and also during most weeks with the model absent. Observations were thus made ~2 or 3× per week on each of the two tanks for 15 weeks (50 tank observations total), and on each of the 10 shrimp in those tanks (five shrimp per tank) for 25 times per shrimp (15× with the fish model present, and 10× with the model absent), resulting in a total of 250 shrimp behavioral observations during the second set.
Prior to each behavioral observation, we starved the shrimp for 2 d to ensure they were hungry enough to potentially engage with the fish model, because cleaner shrimp motivation to signal and clean increases with hunger level (Chapuis & Bshary, 2010). Tanks were selected randomly for order of observation each day, with all observations occurring between 0800–1600. A cardboard blind with a viewing slot was set up around each tank before each observation began to remove visual stimuli from outside the tank (after Jonasson, 1987). The blind was left in place for 5 min of
acclimation, then behavioral data were recorded for 5 min (after Huebner & Chadwick, 2012b; Titus et al., 2015, who found that fish cleans by Pederson shrimp each last < 5 min). All instances of the nine focal types of behaviors (see above) were recorded for each of the 4–6 shrimp in the tank, using direct observation be-cause preliminary video recordings indicated that behavioral de-tails for all 4–6 shrimp were not clearly distinguishable in videos, in part due to the small size of some of the shrimp. If the tank had been selected for observation in the presence of a fish model, the model was inserted vertically at the start of the 5 min observa-tion period and rested motionless on the tank bottom for 2.5 min at ~10 cm from the nearest shrimp to observe initial shrimp re-actions to a motionless model. If there was no observed reaction, or if other shrimp in the group were not near the model, the model then was moved closer to the shrimp (within 2–3 cm) be-cause shrimp visual systems do not perceive fine scale resolution of stationary objects (Caves et al., 2016, 2018). Distance of the fish model to shrimp was ~2–10 cm, because shrimp appeared to per-ceive and react to the fish model when they were up to 10 cm away (closer than the 25 cm distance used by Caves et al., 2018). The fish model was moved inside the tank for 2–5× per observa-tion period (5 min) depending on the distance of shrimp from the model.
Data analysis
Variation in the body size of sea anemones occupied versus those unoccupied by Pederson shrimp during both years examined in the field was analyzed using two-way ANOVA (factors: occupation status and year). Variation in shrimp characteristics (body size, number of individuals per social group, and type of microhabitat zone occupied (Fig. 1); dependent variables) with environmental factors (independent variables: anemone body size, distance from anemone) was analyzed using linear regressions.
To analyze body size ratios among shrimp in the field, we ranked individuals (ranks 1–9) within each social group (i.e., all shrimp that co-occurred on each sea anemone), based on their relative body sizes (TL), with the largest shrimp in each group ranked number one. We then determined the ratio of TL of in-dividuals adjacent in rank within each group (TL rank N / TL rank N + 1; after Buston & Cant, 2006). Individuals with rank N were considered dominant to all individuals with ranks greater than N, based on preliminary observations. We also analyzed whether the body size ratios of individuals adjacent in rank were non-randomly distributed. This allowed us to determine whether individuals were evenly spaced in terms of their body size ratios, and whether there was a minimum size ratio between adjacent individuals, in which similarly sized individuals did not co-occur in a social group. We compared the observed distribution of size ratios with a random distribution of size ratios expected under a null model. We calculated 92 body size ratios of individuals that were adjacent in rank within each observed social group of shrimp during 2015, and 81 body size ratios in 2016, to obtain an observed distribution of ratios each year. We created null dis-tributions of 9200 (2015) and 8100 (2016) ratios using a Monte Carlo procedure in R (Buston & Cant, 2006). For this procedure, we randomly selected individuals from the pool of 119 available individuals (in the same 27 groups) in 2015 and 123 available indi-viduals (in the same 42 groups) in 2016 (not including shrimp that were alone on anemones), and combined them into social groups according to the observed distribution of social group sizes. We then calculated the body size ratios of individuals adjacent in rank from this null population. The procedure was iterated 100 times, generating an expecting random distribution of ratios to which the observed ratios were compared.
To analyze how shrimp behavior varied with shrimp type (six types: females: alpha, beta, gamma, delta; males; juveniles), we employed ANOVAs followed by Tukey multiple comparisons
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

SOCIAL BEHAVIOR OF CLEANER SHRIMP
29
tests, for four types of behaviors toward fish models (signal, ap-proach, clean, retreat; no response was not included in the analysis because it was a neutral behavior), and three types of behaviors toward conspecifics (approach, attack, retreat; no re-sponse not included).
RESULTS
Field observations
Corkscrew sea anemones exhibited stable population size structure during 2015 and 2016, co-consisting of many small individuals and few large ones (Fig. 3). Less than half the anemones in the smallest size classes contained shrimp during both years, whereas almost all anemones in the largest size classes did during 2016. Anemone body size did not differ significantly between the two years examined (two-way ANOVA, F = 0.98, P = 0.32), but varied significantly with the presence of shrimp (two-way ANOVA, F = 24.64, P = < 0.001), in that anemones occupied by shrimp were larger than those unoccupied by shrimp.
The number of Pederson shrimp that occupied each anemone (shrimp social group size) varied widely among anemones. Occupied anemones of all body sizes contained shrimp social groups, but small occupied anemones (< 25 cm2 TCSA) each contained a maximum of only four shrimp during both years, while larger anemones each hosted up to 6–9 shrimp. Shrimp social group size thus increased significantly with anemone body size during 2016 (F = 35.89, P < 0.001), but not during 2015 (F = 0.63, P = 0.43; Fig. 4). There appeared to be a maximum social group size for shrimp (~6–9 individuals per anemone) even in very large anemones, in that during 2016 as anemone body size reached ~100 cm2 TCSA, shrimp social group size reached an asymptote.
Most shrimp occupied near-by soft or hard substrate regardless of body size, except in contact with the host anemone (in micro-habitat zones 4 and 5; 86.7 % of N = 135 individuals in 2015, and 72.1 % of N = 154 individuals in 2016). The remainder shrimp perched on the anemone tentacle crown in zones 2 and 3. No shrimp occurred along the host anemone column in zone 1 (Fig. 6). The body size of shrimp decreased significantly with distance from the anemone in terms of absolute shrimp size and their size rank relative to other shrimp in each social group during the two years examined (Fig. 5). Large shrimp (> 15 mm PMLT) resided either among the tentacles of each anemone (zero distance from the anemone in zone 3 and rarely in zone 2; Fig. 1) or on adja-cent substrate < 2 cm from the anemone tentacle crown (zones 4 and 5). In a few cases, large shrimp occurred up to 4 cm from the anemone tentacle crown (Fig. 1). In contrast, small shrimp (< 10 mm TL) rarely contacted the anemone tentacles, and instead perched on adjacent substrate up to 8.5 cm from the anemone. During 2016, the type of microhabitat occupied by shrimp also depended on shrimp body size, in that large shrimp (> 15 mm TL) resided significantly more frequently in zones 2 and 3 (in contact with the anemone on the inner and outer tentacle crown; occupied by 20.9 and 45.0% of large individuals, N = 43 and 60 in 2015 and 2016, respectively) than did small shrimp (< 10 mm TL), which occurred more frequently in zones 4 and 5 (on soft or hard substratum near the anemone, 0.0–8.5 cm distance from the anemone tentacle tips; occupied by 87.1 and 86.5% of small individuals, N = 31 and 52 in 2015 and 2016, respectively; F = 20.91, P < 0.0001 in 2015; F = 3.69, P = 0.057 2016; Fig. 6).
During both years, the distribution of body size ratios between adjacent individuals within each social group did not differ sig-nificantly from random (Kolmogorov-Smirnov test; D[2] = 0.067, P = 0.81 in 2015; D[2] = 0.094, P = 0.48 in 2016; Fig. 7). Shrimp body size ratios, both those randomly-generated and observed, ranged 1:1 to 4:1 during both years, indicating that the individuals
in each social group exhibited a wide range of body sizes rela-tive to those of adjacent members. Shrimp thus co-occurred in random groupings of individuals in terms of their relative body sizes, and individuals very close in body size occurred together in some social groups.
Laboratory experiments
Behaviors toward the fish model. The percentage of positive inter-actions with the fish model varied significantly with the category of shrimp (ANOVA: F = 2.86; P < 0.05). Alpha female shrimp signaled toward, approached, and cleaned the fish model during most of their interactions with the model (i.e., 67.8% of inter-actions were positive; Fig. 8A). Other shrimp (beta, gamma, and delta females, males and juveniles) often ignored the model (no
Figure 3. Population size structure of the corkscrew sea anemone Bartholomea annulata on coral reefs at St. Thomas, U.S. Virgin Islands during July 2015 (N = 109) and August 2016 (N = 140), and proportion of anemones in each size class that were occupied by Pederson cleaner shrimp Ancylomenes pedersoni (N = 40 in 2015; N = 76 in 2016). Less than half of anemones in the smallest size class were occupied by Pederson shrimp, even though this was the most abundant size class; Almost all of the rare anemones in the largest size class were occupied. TCSA, tentacle crown surface area.
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

J. A. GILPIN AND N. E. CHADWICK
30
reaction) or retreated from it; members of each of these five groups exhibited < 60% positive interactions with the fish model. Pairwise comparisons revealed that alpha females positively inter-acted with the fish model significantly more frequently than did male shrimp (P < 0.05), with no significant differences among all other types of shrimp (Fig. 8A). Percent positive interaction with the fish model did not correlate significantly with shrimp body size (F = 3.67; P = 0.38).
Behaviors toward conspecifics. Shrimp frequently interacted with each other in the laboratory tanks (Table 1, Fig. 9A). As in their high frequencies of approach to the fish model, alpha female shrimp frequently exhibited social dominance behaviors toward con-specifics. The most frequent type of social behavior by alpha fe-males was to approach conspecifics (64.6% of all alpha female social interactions). Conspecific approach rate varied significantly with shrimp type (ANOVA: F = 9.38; P < 0.0001), with alpha fe-males exhibiting approaches at a significantly higher frequency
than did all other categories of shrimp (pairwise comparisons; P < 0.05). In contrast, each of the five other main types of shrimp (beta, gamma, and delta females, males, and juveniles) exhibited approach behavior in less than half of their social interactions (< 50% of conspecific interactions by each type of shrimp), and their frequencies of approach did not differ significantly from each other (pairwise comparisons; P = 0.42–1.00; Fig. 9A).
All four types of females often attacked conspecifics at similar frequencies of ~30% of their social interactions (alpha females 31.2% of all social interactions, beta females 34.2%, gamma fe-males 26.0%, delta females 31.2%). Attacks consisted mostly of individuals contacting conspecifics with their chelae, using the chelae to pinch the antennae and/or forelimbs of the receiving shrimp. In some cases, the receiver reciprocated by contacting the initiator with its own chelae, and pinching the antennae and/or forelimbs of the initiator. Males and juveniles rarely attacked conspecifics (23.6% and 0% of social interactions; Fig. 9). The percent of conspecific interactions involving attacks did not vary significantly with shrimp type (ANOVA F = 1.24; P = 0.32).
All four types of females rarely exhibited subordinate behaviors; they retreated from conspecifics at low frequencies (only 4.2% of social interactions in alpha females, 44.7% in beta females, 32.0% in gamma females, 43.8% in delta females). In contrast, both rela-tively large (male 1) and small male shrimp (male 2) frequently retreated during interactions with conspecifics (77.8% and 73.7% of social interactions, respectively), and juvenile shrimp retreated during all (100%) of their social interactions. As such, the fre-quency of retreat behavior varied significantly with shrimp type (ANOVA F = 2.82; P < 0.05). Pairwise comparisons revealed that large males (male 1) retreated significantly more frequently than did alpha females (P < 0.05), but retreat frequency did not differ significantly among any other categories of shrimp.
Behaviors after change in social group status. When shrimp were moved among the laboratory tanks between the first and second set of be-havioral observations, their behavioral patterns likewise changed, depending on their new body sizes relative to other shrimp in the tank, and thus their status within each social group. The only alpha female that remained in the alpha position, and the only beta female that became alpha, both increased their number of positive interactions with the fish model (alpha to alpha from 27 to 57 positive interactions per 85 and 125 min, respectively, in first and second set of observations; beta to alpha three to 18 per 85 and 125 min). In contrast, the two alpha females that became beta decreased from ten to six positive interactions per 85 and 125 mins, and 12 to four per 85 and 125 min. Positive interactions with the fish model increased for the two gamma female shrimp that remained gamma females, from one to 19 positive inter-actions per 85 and 125 min, and 11 to 30 per 85 and 125 min. Gamma females that reduced their social status by becoming delta females, decreased their number of positive interactions with the fish model, from eight to zero per 85 and 125 min. Likewise, males decreased their already-low number of positive interactions with the fish model, from three to one per 85 and 125 min, but males placed in a tank with a smaller male slightly increased their number of positive interactions with the fish model from two to three per 85 and 125 min. Juveniles that had matured into males greatly increased their number of positive interactions with the fish model from two to 15 per 85 and 125 min.
In terms of interactions with conspecifics, the one alpha female shrimp that remained in the alpha position exhibited no change, with all of its interactions remaining aggressive toward conspe-cifics (five and nine aggressive interactions exhibited per 85 and 125 min, respectively, in first and second set of observations). Of the two alpha females that were moved to the beta female pos-ition, both increased their number of aggressive interactions, from one to 11 interactions and zero to four interactions per 85 and 125 min. The only beta female that moved into the alpha
Figure 4. Variation in the number of associated Pederson shrimp Ancylomenes pedersoni in relation to the body size of the host, the corkscrew sea anemone Bartholomea annulata, during July 2015 and August 2016 on coral reefs at St. Thomas, U.S. Virgin Islands. The number of shrimp per anemone (size of shrimp social group) correlated significantly (P < 0.0001) with anemone body size during 2016; an exponential relationship yielded the highest r-squared value. TCSA, tentacle crown surface area.
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

SOCIAL BEHAVIOR OF CLEANER SHRIMP
31
position increased her aggressive interactions from four to seven occurrences per 85 and 125 min, representing an increase from 66.7% to 87.5% of her social interactions. The two gamma fe-males that remained in the gamma position also substantially in-creased their aggressive interactions, from zero to six and one to 20 occurrences per 85 and 125 min, indicating increases from 0% to 60% and 20% to 76.9% of their social interactions, respect-ively. The only gamma female that decreased in status to the delta position decreased her aggressive interactions from four to two occurrences per 85 and 125 min, representing a large drop from 80% to 28.6% of her social interactions. During the second set of behavioral observations, the two males that moved among social groups increased their aggressive interactions (zero to four and zero to five per 85 and 125 min) corresponding to increases of 0% to 28.6% and 0% to 31.3% of their social interactions, respectively. The juvenile that matured into a male increased his number of aggressive interactions from zero to five per 85 and 125 total min, representing an increase from 0% to 26.3% of his social interactions.
Construction of social dominance hierarchies. Based on the above behav-ioral interactions of shrimp with the fish model and with each other during both sets of observations, shrimp were classified into positions in a social dominance hierarchy based on gender and relative body size within gender. Alpha females (the largest female within each so-cial group) were placed at the top of the hierarchy and exhibited the highest frequencies of dominance behaviors (signal, approach, and clean fish model; approach and attack conspecifics) and the lowest frequencies of subordinate behaviors (no response, retreat; Table 1; Figs. 8B, 9B). Beta, gamma, and delta females (defined as smaller females in descending order of relative body size) were positioned at intermediate levels in the hierarchy, exhibiting intermediate rates of both dominant and subordinate behaviors, with some reversals in the directionality of their behaviors toward conspecifics of both higher and lower rank. Males and juveniles (which were in all cases smaller than females, with juveniles being smaller than males) were positioned at the bottom of the hierarchy and in almost all of their interactions exhibiting subordinate behaviors (retreat) and were the receivers of social dominance behaviors by females.
Figure 5. Variation in the distance of Pederson shrimp Ancylomenes pedersoni from corkscrew sea anemones Bartholomea annulata with absolute shrimp body size (A, B) and relative shrimp body size (C, D) (rank in each social group: 1, largest shrimp; 10, smallest shrimp), during July 2015 and August 2016 on coral reefs at St. Thomas, U.S. Virgin Islands. The distance of each shrimp from its host anemone varied widely, but large high-ranking shrimp occurred significantly closer (P < 0.001 in 2015 and 2016) to anemones than the small low-ranking shrimp during both years. Relative shrimp body size (size rank) correlated more tightly with distance from the host anemone than did absolute shrimp body size (TL).
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

J. A. GILPIN AND N. E. CHADWICK
32
Most of the 209 observed instances of social interactions oc-curred among female shrimp, or between females and the largest male in the social group (male 1; Table 1). Both the smallest male within each social group (male two) and the juveniles rarely inter-acted with any of the other shrimp.
DISCUSSION
We demonstrate that on Caribbean coral reefs, Pederson shrimp form social groups that vary widely in structure among host anemones in terms of the relative body sizes and sexual repro-ductive status of shrimp on each anemone. These social groups are spatially structured, with large females occupying more cen-tral microhabitats on each anemone than do all other shrimp (smaller females, males, and juveniles), which are relegated to
peripheral habitats on reef or sand substrate adjacent to the anemone. Shrimp exhibited a size- and gender-based behavioral dominance hierarchy under laboratory conditions, with large alpha females displaying dominance in terms of both exploitation and interference competition behaviors being more frequent than do smaller subordinate group members. Alpha females frequently signaled towards, approached, and cleaned a model fish (poten-tially exploiting a food source of in the form of fish ectoparasites), and also approached and attacked smaller group members, an ex-pression of interference competition, potentially for habitat space and access to food resources. Conversely, small females, males, and juveniles all exhibit more neutral or negative interactions with the fish model (no response or retreat from the model), and more sub-ordinate social behaviors in the form of retreats from conspecifics, than do large alpha females. These behavioral interactions result in large shrimp occupying more dominant social positions than do small shrimp in each social group, allowing the former to secure prominent microhabitat locations on the tips of sea anemone tent-acles. This spatial resource monopolization likely grants the largest shrimp in each social group more frequent access to the food re-sources provided by client fishes that visit anemones as cleaning stations for the removal of ectoparasites (Huebner & Chadwick, 2012b). These shrimp therefore can be considered to form true social groups, in that they exhibit “reciprocal communication of
Figure 6. Variation among three body size classes (small ≤ 10 mm total length (TL); medium 11–15 mm; large > 15 mm) of Pederson shrimp Ancylomenes pedersoni in the percent of individuals that occurred in each of five microhabitat zones (Fig. 1) on corkscrew sea anemones Bartholomea annulata at St. Thomas, U.S. Virgin Islands during July 2015 and August 2016. Sample sizes (number of shrimp in each size class) are shown at the top of each bar. A higher proportion of large shrimp occupied zones 2 and 3 than did small shrimp and a higher proportion of small shrimp were rele-gated to zone 5 (near anemone on sand) than medium and large shrimp; no individuals occupied zone 1 (see Fig. 1), so this zone was not included here.
Figure 7. Observed distributions of body size ratios between individuals adjacent in rank (dark grey and light gray) within social groups of Pederson shrimp Ancylomenes pedersoni on host sea anemones Bartholomea annulata, during July 2015 and August 2016 on coral reefs at St. Thomas, U.S. Virgin Islands. The random distributions of ratios expected under a null model generated using Monte Carlo procedure are shown in white and light gray. Light gray represents the area where the observed and random distributions overlap. Observed distributions of ratios were not significantly different (P = 0.81 in 2015; P = 0.48 in 2016) during both years from those generated randomly. Body sizes were measured as total length (TL).
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

SOCIAL BEHAVIOR OF CLEANER SHRIMP
33
a cooperative nature” (Costa & Fitzgerald, 2005) through social signals and responses that allow them to partition limited habitat space according to their gender and relative body size on each host anemone. Related types of social spacing behavior have been observed in schools of pelagic mysids (Clutter, 1969), and may be distinguished from the eusocial behavior of some snap-ping shrimps Synalpheus Spence Bate, 1888, which form groups of overlapping adult generations with division of labor and repro-ductive altruism (Duffy et al., 2000).
The randomly distributed size ratios that we observed between shrimp of adjacent social rank indicate that some individuals within social groups may be quite similar in body size, in con-trast to social groups of anemonefishes that maintain a minimum
1.2–1.3 body size ratio within each host sea anemone (Buston & Cant, 2006). High rates of cleaner shrimp dispersal among host anemones, in which shrimp move nocturnally among nearby anemones (Chadwick et al. 2008), could in part cause this pattern. The observed ability of Pederson shrimp to rapidly alter their behavioral interactions when their social position changes also
Figure 8. Variation in percent positive interactions with a fish model among six categories of Pederson shrimp Ancylomenes pedersoni based on gender and relative body size in social groups under laboratory conditions. Positive interactions with the fish model were calculated for each shrimp in each category as a percent of all that individual’s behavioral interactions with the model (including three types of positive interactions (signal, ap-proach, clean) and two types of neutral/negative interactions (no response, retreat) observed). Absolute percent positive interactions (A); relative per-cent positive interactions (line widths) (B). α, alpha female; β, beta female; γ, gamma female; δ, delta female; Juv, juvenile. Sample sizes above each bar indicate the number of shrimp observed in each category.
Figure 9. Variation in percent aggressive interactions toward conspecifics among seven categories of Pederson shrimp Ancylomenes pedersoni based on gender and relative body size in social groups under laboratory conditions. Aggressive interactions with conspecifics were calculated for each shrimp in each category as a percent of all behavioral interactions of individuals with conspecifics (including two types of aggressive interactions (approach, attack) and two types of neutral/negative interactions (no response, retreat) observed). Absolute percent aggressive interactions (A); relative number of aggressive interactions (line widths) (B). Relative numbers of observed be-haviors are shown rather than relative percent and males are separated into male 1 (larger) and male 2 (smaller) to more clearly distinguished rates of aggression toward different types of conspecifics. Rare occurrences of ag-gression (< 5 for each shrimp category) were also excluded from B to show only the most common types of interactions for clarity. α, alpha female; β, beta female; γ, gamma female; δ, delta female; Juv, juvenile. Sample sizes above each bar indicate the number of shrimp observed in each category.
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

J. A. GILPIN AND N. E. CHADWICK
34
contributes to these shrimp being able to adjust behaviorally to frequent changes in social status as they disperse among anem-ones. Examination of larger sample sizes in both field and labora-tory settings, and the collection of data at other field sites, may reveal somewhat different social patterns of Pederson shrimp, and should be explored in future studies.
Field observations
Our field observations on corkscrew sea anemones revealed popu-lations composed mostly of small individuals, indicating stable population structure with high recruitment. This pattern is similar to that observed for this anemone during previous studies at the same field site (O’Reilly et al., 2018), as well as in nearshore Florida habitats (O’Reilly & Chadwick, 2017). Many corkscrew anemones hosted Pederson shrimp at our study site, but the presence of un-occupied anemones indicates that these shrimp may not be space limited by the number of anemones in the local population. This pattern is similar to that observed for closely-related shrimps on sea anemones in Bermuda (Nizinski, 1989), in Belize (Brooker et al., 2019), and in the northern Red Sea (Chadwick et al., 2008).
The field pattern of increasing shrimp social group size with anemone body size we observed (also reported by Titus et al., 2017 and Huebner et al., in press) may occur because large anemones provide more habitat space for shrimp than small ones, and also visually attract more food resources in the form of client fishes laden with ectoparasites (Huebner & Chadwick 2012a; Titus et al., 2017). The large social groups of shrimp observed, of up to nine individuals per anemone, are unusual for crustacean associates of cnidarians, which occur usually as a pair of mated individuals per host (Duffy, 1996; Baeza et al., 2002; Hirose, 2012). The small body size of Pederson shrimp relative to their host, the cleaning habit, and other ecological factors may cause these shrimp to occur in mixed-gender social groups (reviewed in Gilpin & Chadwick, 2017), similar to the pattern observed for anemone-associated cleaner shrimps on Indo-West Pacific coral reefs (Chadwick et al., 2008). Likewise, individuals of the squat anemone shrimp Thor amboinensis (De Man, 1888) have small body sizes relative to host size, and are sequential hermaphrodites that live in groups of up to 11 individuals per host (Baeza & Piantoni, 2010). In contrast, other anemone shrimps on corkscrew sea anemones Bartholomea annulata, such as snapping shrimp Alpheus armatus (Rathbun, 1901) live in male-female pairs, reside along the host anemone column, and do not clean fishes (Knowlton, 1980).
The relationship reported here between shrimp body size (both absolute and relative to other social group members) and distance from the host anemone reveals behavioral habitat partitioning according to body size in this shrimp species. Occupation of anemone tentacle tips may allow alpha female shrimp to gain enhanced protection from predators relative to that provided fur-ther away from the anemone, as well as prime access to food re-sources through visual prominence on the host for signaling to, approaching, and cleaning reef fishes (Caves et al., 2018). Studies are needed to examine variation in the nutritional status and gut contents of cleaner shrimps relative to their body size and repro-ductive status, to further understand how these factors impact their diet and nutritional state. Alpha females may tolerate the presence of small conspecifics in social groups, in part because they utilize fewer resources (space, food) than do other large fe-males, as described for spider crabs Inachus phalangium (Fabricius, 1775) on corkscrew sea anemones (Wirtz & Diesel, 1983).
The broad range of shrimp sizes in each social group, and the randomly-distributed size ratios between adjacent ranking indi-viduals, both indicate that social groups in this species are only loosely organized by body size. Shrimp may colonize into and emigrate away from individual anemones frequently enough to preclude the formation of highly size-structured, temporally stable social groups, in contrast to anemonefishes (Huebner et al., 2012).
The behavioral interactions of Pederson shrimp within even tem-porary social groups, however, appear to create clear dominance hierarchies based on relative body size and gender. These shrimp have the ability to disperse frequently between anemones, as do some other cleaner shrimps (Chadwick et al., 2008; review in Gilpin & Chadwick, 2017), which could in part explain why they do not exhibit highly size-structured social groups.
Laboratory experiments
Pederson shrimp that occupy dominant social positions (alpha fe-males) in laboratory social groups exhibit the highest frequencies of positive interactions with model reef fish, indicating they are likely to signal toward, approach, and clean client fishes in the field. These behaviors appear to be mechanisms of exploitation competition that allow them to attract and access food resources pre-emptively before other shrimp in the social group. The two-way behavioral signaling (shrimp body swaying and antenna waving, fish posing and color change) that occurs between shrimp and fishes further may allow the most prominently positioned and largest shrimp on a given anemone to engage in the reciprocal interactions that culminate in a successful fish clean (Caves et al., 2018). Alpha females most frequently approach and attack sub-ordinate conspecifics, expressing behaviors that appear to func-tion as mechanisms of interference competition. These behaviors likely exclude infringing subordinate shrimp from access to space and food resources in that they cause the retreat of subordinates, thereby allowing alpha females to gain dominant social status and monopolize resources. High rates of beta females attacking sub-ordinate (smaller) individuals reveal that they also behave aggres-sively, which allows them to maintain their mid-level position in the hierarchy. Conversely, the high rate of retreat observed for small males indicates their subordinate status relative to larger more dominant individuals in the group. These interactions pro-vide behavioral mechanisms whereby shrimp may maintain their spatial partitioning of microhabitats on host sea anemones. This type of size-structured habitat partitioning, in which large individ-uals occupy more central habitat on sea anemones than do small individuals, also has been observed in the squat anemone shrimp Thor amboinensis on rosetip anemones Condylactis gigantea on the Caribbean coast of Mexico (Colombara et al., 2017) and for the anemone fish Amphiprion chrysopterus (Cuvier, 1830) on the anemone Heteractis magnifica (Quoy & Gaimard, 1833) in Moorea, French Polynesia (Holbrook & Schmidt, 2004). Anemonefishes interspe-cifically partition microhabitats on host anemones, with dominant species affecting the niche use of subordinate species on shared hosts and large individuals excluding small ones from the most sheltered habitat near the polyp center (Buston 2003; Hattori, 2002; Camp et al. 2016).
This type of spatial pattern has not been previously quan-tified for cleaner shrimps, and indicates that their social groups are highly structured in relation to reef microhabitats, leading to consequences for the dynamics of cleaning interactions. Small subordinate shrimp in a given social group may rarely engage in cleaning behavior, which could cause them to disperse among anemones until they find a host where they are able to occupy a more dominant social position; in contrast, alpha females may be less mobile and more consistent residents of a given cleaning sta-tion. Further studies are needed to track how mobility in the field varies with shrimp body size and gender, in this and other species of cleaner shrimps that form social groups.
Conclusions
The host-use patterns and social group interactions of the ob-served shrimp reveal a clear social hierarchy among individuals based on their reproductive status and body size, which is organ-ized through behavioral dominance interactions. Reproductively
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

SOCIAL BEHAVIOR OF CLEANER SHRIMP
35
active females are the most dominant individuals on each host anemone, with males and juveniles acting as subordinates. This social hierarchy may in part cause small individuals to occupy more peripheral habitats on anemones, and to be relatively mo-bile, in that small subordinates may disperse frequently among anemones in search of access to resources (mates, food, habitat space) that they are prevented from obtaining by the present of large dominant shrimp on a given anemone. This pattern is some-what similar to that known for coral-pit (Simon-Blecher & Achituv, 1997) and gall crabs (Cryptochiridae) (van der Meij, 2014), in which large females remain stationary on each host, while small males migrate among hosts and mate with multiple females. The complex and potentially dynamic composition of Pederson shrimp social groups at cleaning stations (sea anemones) may cause reef fishes to search for new cleaning stations to visit for parasite re-moval, thus enhancing fish movement both within and among reef areas. Further research is needed to elucidate the impacts of shrimp social dynamics on cleaning symbioses with reef fishes, es-pecially in terms of both cleaner shrimp and client fish mobility among sea anemones, and effects of variation in shrimp dispersal rates with social and reproductive status.
ACKNOWLEDGEMENTS
We thank members of our laboratory who assisted with daily care to support the required culture. We also thank Mark Stuart for serving as a dive buddy during fieldwork, Steve Ratchford for hosting us on St. Thomas, and diving officer Steve Prosterman (University of the Virgin Islands Center for Marine and Environmental Studies) for diving support. Courtney Andes and Ellen Reiber assisted with collecting behavioral data, Benjamin Staton with statistical analysis, and Grace Denniston with figure design. This research was funded in part by a Lerner Grey Grant from the American Museum of Natural History, an International Women’s Fishing Association Scholarship, a Women Divers Hall of Fame Honorable Mention Award, and a Graduate Research Grant from the Auburn University Graduate School, all to JAG. This work was submitted in partial fulfillment of the requirements for the doctoral degree to JAG at Auburn University. This is publication #189 of the Auburn University Marine Biology Program.
REFERENCESAlexander, R.D. 1961. Aggressiveness, territoriality, and sexual behavior in
field crickets (Orthoptera: Gryllidae). Behaviour, 17: 130–223.Baeza, J.A. & Piantoni, C. 2010. Sexual system, sex ratio, and group
living in the shrimp Thor amboinensis (De Man): Relevance to resource-monopolization and sex-allocation theories. Biological Bulletin, 219: 151–165.
Baeza, J.A., Stotz, W. & Thiel, M. 2002. Agonistic behaviour and devel-opment of territoriality during ontogeny of the sea anemone dwelling crab Allopetrolisthes spinifrons (H. Milne Edwards, 1837) (Decapoda: Anomura: Porcellanidae). Marine and Freshwater Behavior and Physiology, 35: 189–202.
Bauer, R.T. 1986. Sex change and life history pattern in the shrimp Thor manningi (Decapoda: Caridea): A novel case of partial protandric herm-aphroditism. Biological Bulletin, 170: 11–31.
Bauer, R.T. 2004. Remarkable shrimps: adaptations and natural history of the carideans. University of Oklahoma Press, Norman, OK, USA.
Becker, J.H.A. & Grutter, A.S. 2005. Client fish ectoparasite loads and cleaner shrimp Urocaridella sp. c hunger levels affect cleaning behaviour. Animal Behavior, 70: 991–996.
Becker, J.H.A., Curtis, L.M. & Grutter, A.S. 2005. Cleaner shrimp use a rocking dance to advertise cleaning service to clients. Current Biology, 15: 760–764.
Bovbjerg, R.V. 1953. Dominance order in the crayfish Orconectes virilis (Hagen). Physiological Zoology, 26: 173–178.
Bovbjerg, R.V. 1956. Some factors affecting aggressive behavior in cray-fish. Physiological Zoology, 29: 127–136.
Brooker, R.M., Feeney, W.E., Sih, T.L., Ferrari, M.C.C. & Chivers, D.P. 2019. Comparative diversity of anemone-associated fishes and decapod crustaceans in a Belizean coral reef and seagrass system. Marine Biodiversity [doi:10.1007/s12526-019-00993-5].
Bshary, R. 2003. The cleaner wrasse, Labroides dimidiatus, is a key organism for reef fish diversity at Ras Mohammed National Park, Egypt. Journal of Animal Ecology, 72: 169–176.
Bshary, R., Oliveira, R.F., Oliveira, T.S.F. & Canario, A.V.M. 2007. Do cleaning organisms reduce the stress response of client reef fish? Frontiers in Zoology, 4: 1–8.
Bunkley-Williams, L. & Williams, E.H. 1998. Ability of Pederson cleaner shrimp to remove juveniles of the parasitic cymothoid isopod, Anilocra haemuli, from the host. Crustaceana, 71: 862–869.
Buston, P.M. 2003. Size and growth modification in clownfish. Nature, 424: 145–146.
Buston, P.M. & Cant, M.A. 2006. A new perspective on size hier-archies in nature: Patterns, causes, and consequences. Oecologia, 149: 362–372.
Cantrell, C.E., Henry, R.P. & Chadwick, N.E. 2015. Nitrogen transfer in a Caribbean mutualistic network. Marine Biology, 162: 2327–2338.
Camp, E.F., Hobbs, J.A., De Brauwer, M., Dumbrell, A.J. & Smith, D.J. 2016. Cohabitation promotes high diversity of clowfishes in the Coral Triangle. Proceedings of the Royal Society B, 283: 20160277 [doi: 10.1098/rspb.2016.0277].
Caves, E.M., Frank, T.M. & Johnsen, S. 2016. Spectral sensitivity, spatial resolution and temporal resolution and their implications for conspe-cific signalling in cleaner shrimp. Journal of Experimental Biology, 219: 597–608.
Caves, E.M., Green, P.A. & Johnsen, S. 2018. Mutual visual signalling between the cleaner shrimp Ancylomenes pedersoni and its client fish. Proceedings of the Royal Society B 285: 20180800 [doi: 10.1098/rspb.2018.0800].
Chace, F.A. 1958. A new shrimp of the genus Periclimenes from the West Indies. Proceedings of the Biological Society of Washington, 71: 125–132.
Chadwick, N.E., Duriš, Z. & Horká, I. 2008. Biodiversity and behavior of shrimps and fishes symbiotic with sea anemones in the Gulf of Aqaba, northern Red Sea. In: The improbable gulf: History, biodiversity, and protec-tion of the Gulf of Aqaba-Eilat (F.D. Por, ed), pp. 209–233. Magnes Press, Hebrew University, Jerusalem.
Chapuis, L. & Bshary, R. 2009. Strategic adjustment of service quality to client identity in the cleaner shrimp, Periclimenes longicarpus. Animal Behavior, 78: 455–459.
Chapuis, L. & Bshary, R. 2010. Signalling by the cleaner shrimp Periclimenes longicarpus. Animal Behavior, 79: 645–647.
Cheney, K.L., Grutter, A.S., Blomberg, S.P. & Marshall, N.J. 2009. Blue and yellow signal cleaning behavior in coral reef fishes. Current Biology, 19: 1283–1287.
Clutter, R.I. 1969. The microdistribution and social behavior of some pe-lagic mysid shrimps. Journal of Experimental Marine Biology and Ecology, 3: 125–155.
Colombara, A.M., Quinn, D. & Chadwick, N.E. 2017. Habitat segrega-tion and population structure of Caribbean sea anemones and asso-ciated crustaceans on coral reefs at Akumal Bay, Mexico. Bulletin of Marine Science, 93: 1025–1047.
Copp, N.H. 1986. Dominance hierarchies in the crayfish Procambarus clarkii (Girard, 1852) and the question of learned individual recognition (Decapoda, Astacidea). Crustaceana, 51: 9–24.
Costa, J.T. & Fitzgerald, T.D. 2005. Social terminology revisited: Where are we ten years later? Annales Zoologici Fennici, 42: 559–564.
Côté, I.M. 2000. Evolution and ecology of cleaning symbioses in the sea. Oceanography and Marine Biology: An Annual Review, 38: 311–355.
Courchesne, E. & Barlow, G.W. 1971. Effect of isolation on components of aggressive and other behavior in the hermit crab, Pagurus samuelis. Journal of Comparative Physiology, A, 75: 32–48.
Dixon, A. K., McVay, M.J. & Chadwick, N.E. 2017. Demographic mod-elling of giant sea anemones: Population stability and effects of mu-tualistic anemonefish in the Jordanian Red Sea. Marine and Freshwater Research, 68: 2145–2155.
Duffy, J.E. 1996. Species boundaries, specialization, and the radiation of sponge-dwelling alpheid shrimp. Biological Journal of the Linnean Society, 58: 307–324.
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020

J. A. GILPIN AND N. E. CHADWICK
36
Duffy, J.E., Morrison, C.L. & Rios, R. 2000. Multiple origins of eusoci-ality among sponge-dwelling shrimps (Synalpheus). Evolution, 54: 503–516.
Fabricius, J. C., 1775. Systema Entomologiae, sistens Insectorum Classes, Ordines, Genera, Species, adjectis Synonymis, Locis, Descriptionibus, Observationibus. Officina Libraria Kortii, Flensburgi & Lipsiae [Leipzig].
Gilpin, J.A. & Chadwick, N.E. 2017. Life history traits and popula-tion structure of Pederson cleaner shrimps Ancylomenes pedersoni. Biological Bulletin, 233: 190–205.
Guo, C.C., Hwang, J.S. & Fautin, D.G. 1996. Host selection by shrimps symbiotic with sea anemones: A field survey and experimental labora-tory analysis. Journal of Experimental Marine Biology and Ecology, 202: 165–176.
Hattori, A. 2002. Small and large anemonefishes can coexist using the same patchy resources on a coral reef, before habitat destruction. Journal of Animal Ecology, 71: 824–831.
Herrnkind, W., Stanton, G. & Conklin, E. 1976. Initial characterization of the commensal complex associated with the anemone, Lebrunia danae, at Grand Bahama. Bulletin of Marine Science, 26: 65–71.
Hirose, G.L. 2012. New record and biological features of the commensal porcellanid crab Polyonyx gibbesi (Crustacea: Anomura) from the north-eastern Brazilian coast. Marine Biodiversity Records, 5: e43 [doi: org/10.1017/S1755267212000188].
Hirose, Y. 1985. Habitat, distribution and abundance of coral reef sea-anemones (Actiniidae and Stichodactylidae) in Sesoko Island, Okinawa, with notes of expansion and contraction behavior. Galaxea, 4: 113–127.
Holbrook, S.J. & Schmidt, R.J. 2004. Population dynamics of a damselfish: Effects of a competitor that also is an indirect mutualist. Ecology, 85: 979–985.
Holthuis, L.B. & Eibl-Eibesfeldt, I. 1964. A new species of the genus Periclimenes from Bermuda. Senckenbergiana biologica, 45: 185–192.
Huebner, L.K. & Chadwick, N.E. 2012a. Reef fishes use sea anemones as visual cues for cleaning interactions with shrimp. Journal of Experimental Marine Biology and Ecology, 416: 237–242.
Huebner, L.K. & Chadwick, N.E. 2012b. Patterns of cleaning behaviour on coral reef fish by the anemone shrimp Ancylomenes pedersoni. Journal of the Marine Biological Association of the United Kingdom, 92: 1557–1562.
Huebner, L.K., Dailey, B. Titus, B.M., Khalaf, M. & Chadwick, N.E. 2012. Host preference and habitat segregation among Red Sea anemonefish: Effects of sea anemone traits and fish life stages. Marine Ecology Progress Series, 464: 1–15.
Huebner, L.K., Shea, C.P., Schueller, P.M., Terrell, A.D., Ratchford, S.G. & Chadwick, N.E. In press. Crustacean associates of Caribbean sea anemones Bartholomea annulata: Occupancy modeling, habitat partitioning, and persistence. Marine Ecology Progress Series.
Humann, P. & DeLoach, N. 2002. Reef fish identification: Florida, Caribbean, Bahamas. New World Publications, Jacksonville, FL, USA.
Issa, F.A., Adamson, D.J. & Edwards, D.H. 1999. Dominance hierarchy formation in juvenile crayfish Procambarus clarkii. Journal of Experimental Biology, 202: 3497–3506.
Ives, J.E. 1891. Crustacea from the northern coast of Yucatan, the harbor of Vera Cruz, the west coast of Florida and the Bermuda Islands. Proceedings of the Academy of Natural Sciences of Philadelphia, 1891: 176–207.
Jonasson, M. 1987. Fish cleaning behaviour of shrimp. Journal of Zoology. 213: 117–131.
Khan, R.N., Becker, J.H., Crowther, A.L. & Lawn, I.D. 2004. Spatial dis-tribution of symbiotic shrimps (Periclimenes holthuisi, P. brevicarpalis, Thor amboinensis) on the sea anemone Stichodactyla haddoni. Journal of the Marine Biological Association of the United Kingdom, 84: 201–203.
Knowlton, N. 1980. Sexual selection and dimorphism in two demes of a symbiotic, pair-bonding snapping shrimp. Evolution, 34: 161–173.
Lee, C.L. & Fielder, D.R. 1982. Agonistic behaviour and the develop-ment of dominance hierarchies in the freshwater prawn, Macrobrachium australiense Holthuis, 1950 (Crustacea: Palaemonidae). Behaviour 83: 1–17.
Levine, D.M. & Blanchard, O.J. Jr. 1980. Acclimation of two shrimps of the genus Periclimenes to sea anemones. Bulletin of Marine Science, 30: 460–466.
Limbaugh, C., Pederson, H. & Chace, F.A. 1961. Shrimps that clean fishes. Bulletin of Marine Science, 11: 237–257.
Mahnken, C. 1972. Observations on cleaner shrimps of the genus Periclimenes. Bulletin of the Natural History Museum of Los Angeles County, 14: 71–83.
Man, J.G. de. 1888. Bericht über die von Herrn Dr. J. Brock im indischen Archipel gesammelten Decapoden und Stomatopoden. Archiv für Naturgeschichte, 53: 289–600.
Mascaró, M., Rodríguez-Pestaña, L., Chiappa-Carrara, X. & Simões, N. 2012. Host selection by the cleaner shrimp Ancylomenes pedersoni: Do anemone host species, prior experience or the presence of conspecific shrimp matter? Journal of Experimental Marine Biology and Ecology, 413: 87–93.
Nizinski, M.S. 1989. Ecological distribution, demography and behavioral observations on Periclemenes anthophilus, an atypical symbiotic cleaner shrimp. Bulletin of Marine Science, 45: 174–188.
O’Reilly, E.E. & Chadwick, N.E. 2017. Population dynamics of corkscrew sea anemones Bartholomea annulata in the Florida Keys. Marine Ecology Progress Series, 567: 109–123.
O’Reilly, E., Titus, B.M., Nelsen, M.W., Ratchford, S. & Chadwick, N.E. 2018. Giant ephemeral anemones? Rapid growth and high mortality of corkscrew sea anemones Bartholomea annulata (Le Sueur, 1817) under variable conditions. Journal of Experimental Marine Biology and Ecology, 509: 44–53.
Pavey, C.R. & Fielder, D.R. 1996. The influence of size differential on agonistic behaviour in the freshwater crayfish, Cherax cuspidatus (Decapoda: Parastacidae). Journal of Zoology, 238: 445–457.
Ranta, E. & Lindström, K. 1992. Power to hold sheltering burrows by juveniles of the signal crayfish, Pasifastacus leniusculus. Ethology, 92: 217–226.
Rathbun, M.J. 1901. The Brachyura and Macrura of Porto Rico. Bulletin of the United States Fish Commission, 200: 1–127.
Rubenstein, D.I. & Hazlett, B.A. 1974. Examination of the agonistic be-haviour of the crayfish Orconectes virilis by character analysis. Behaviour, 50: 193–215.
Silbiger, N.J. & Childress, M.J. 2008. Interspecific variation in anemone shrimp distribution and host selection in the Florida Keys (USA): Implications for marine conservation. Bulletin of Marine Science, 83: 329–345.
Simon-Blecher, N. & Achituv, Y. 1997. Relationship between the coral pit crab Cryptochirus coralliodytes Heller and its host coral. Journal of Experimental Marine Biology and Ecology, 215: 93–102.
Spence Bate, C. 1888. Report on the Crustacea Macrura collected by the Challenger during the years 1873–76. In: Report on the scientific results of the Voyage of H.M.S. “Challenger” during the years 1973-76, Vol. 24, pp. 1–942, pls. 1–157.
Titus, B.M., Daly, M. & Exton, D.A. 2015. Temporal patterns of Pederson shrimp (Ancylomenes pedersoni Chace 1958) cleaning interactions on Caribbean coral reefs. Marine Biology, 162: 1651–1664.
Titus, B.M., Vondriska, C. & Daly, M. 2017. Comparative behavioral ob-servations demonstrate the “cleaner shrimp” Periclimenes yucatanicus en-gages in true symbiotic cleaning interactions. Royal Society Open Science, 4: 170078 [do: org/10.1098/rsos.170078].
Titus, B.M., Daly, M., Vondriska, C., Hamilton, I. & Exton, D.A. 2019. Lack of strategic service provisioning by Pederson’s cleaner shrimp (Ancylomenes pedersoni) highlights independent evolution of cleaning behaviors between ocean basins. Scientific Reports, 9: 629 [doi: org/10.1038/s41598-018-37418-5].
van der Meij, S.E. 2014. Host species, range extensions, and an ob-servation of the mating system of Atlantic shallow-water gall crabs (Decapoda: Cryptochiridae). Bulletin of Marine Science, 90: 1001–1010.
Winston, M.L. & Jacobson, S. 1978. Dominance and effects of strange conspecifics on aggressive interactions in the hermit crab Pagurus longicarpus (Say). Animal Behavior, 26: 184–191.
Wirtz, P. & Diesel, R. 1983. The social structure of Inachus phalangium, a spider crab associated with the sea anemone. Ethology, 62: 209–234.
Dow
nloaded from https://academ
ic.oup.com/jcb/article-abstract/40/1/24/5613990 by Auburn U
niversity user on 13 January 2020


















