Journal of Biotechnology - deficitdao.org
11
Journal of Biotechnology 227 (2016) 120–130 Contents lists available at ScienceDirect Journal of Biotechnology j ourna l ho me pa ge: www.elsevier.com/locate/jbiotec Characterization of recombinant human diamine oxidase (rhDAO) produced in Chinese Hamster Ovary (CHO) cells Elisabeth Gludovacz a,b , Daniel Maresch c,1 , Maximilian Bonta d,1 , Helen Szöllösi b,1 , Paul G. Furtmüller c , Robert Weik e , Friedrich Altmann c , Andreas Limbeck d , Nicole Borth a , Bernd Jilma b,∗ , Thomas Boehm b a Department of Biotechnology, University of Natural Resources and Life Sciences, Muthgasse 18, 1190 Vienna, Austria b Department of Clinical Pharmacology, Medical University of Vienna, Waehringer Guertel 18-20, 1090 Vienna, Austria c Department of Chemistry, University of Natural Resources and Life Sciences, Muthgasse 18, 1190 Vienna, Austria d Institute of Chemical Technologies and Analytics, Vienna University of Technology, Getreidemarkt 9, 1060 Vienna, Austria e Polymun Scientific Immunbiologische Forschung GmbH, Donaustraße 99, 3400 Klosterneuburg, Austria a r t i c l e i n f o Article history: Received 11 December 2015 Received in revised form 16 March 2016 Accepted 4 April 2016 Available online 7 April 2016 Keywords: DAO Recombinant human diamine oxidase CHO Chinese Hamster Ovary Amiloride-sensitive amine oxidase AOC1 a b s t r a c t Human diamine oxidase (hDAO) efficiently degrades polyamines and histamine. Reduced enzyme activ- ities might cause complications during pregnancy and be involved in histamine intolerance. So far hDAO has been characterized after isolation from either native sources or the heterologous production in insect cells. Accessibility to human enzyme is limited and insect cells produce non-human glycosylation patterns that may alter its biochemical properties. We present the heterologous expression of hDAO in Chinese Hamster Ovary (CHO) cells and a three step purification protocol. Analysis of metal content using ICP-MS revealed that 93% of the active sites were occupied by copper. Topaquinone (TPQ) cofactor content was determined using phenylhydrazine titra- tion. Ninety-four percent of DAO molecules contained TPQ and therefore the copper content at the active site was indirectly confirmed. Mass spectrometric analysis was conducted to verify sequence integrity of the protein and to assess the glycosylation profile. Electronic circular dichroism and UV–vis spectra data were used to characterize structural properties. The substrate preference and kinetic parameters were in accordance with previous publications. The establishment of a recombinant production system for hDAO enables us to generate decent amounts of protein with negligible impurities to address new scientific questions. © 2016 Elsevier B.V. All rights reserved. 1. Introduction Human diamine oxidase (hDAO, E.C. 1.4.3.6), formerly named histaminase or amiloride-binding protein (ABP1), is encoded by the AOC1 gene and belongs to the group of copper-containing amine oxidases (McGrath et al., 2010b). CAOs are found in bacteria (Parsons et al., 1995; Wilce et al., 1997; Zeng and Spiro, 2013), yeast (Dove et al., 1996; Klema et al., 2013; Li et al., 1998), plants (Cona et al., 2014; Kumar et al., 1996), and mammals (Finney et al., 2014; Kaitaniemi et al., 2009; Salmi and Jalkanen, 2005). They oxida- tively deaminate primary amines to the corresponding aldehydes, ∗ Corresponding author. E-mail address: [email protected] (B. Jilma). 1 These authors contributed equally to this work. ammonia and hydrogen peroxide through a two-step ping-pong mechanism (Brazeau et al., 2004; Mure et al., 2002): RCH 2 NH 2 + O 2 + H 2 O → RCHO + NH 3 + H 2 O 2 Human DAO preferentially degrades histamine and polyamines such as putrescine, cadaverine, spermine and spermidine (Elmore et al., 2002; Schwelberger et al., 2013a). It is an important enzyme in the extracellular catabolism of histamine (Biega ´ nski et al., 1983). High levels of hDAO expression are found in the intestine, where they are predominantly associated with mature enterocytes in the villus tips (Wollin et al., 1998). Human DAO protein is also abun- dant in the kidney (Schwelberger et al., 2013b; Schwelberger et al., 1998) and placenta, where a decreased enzyme activity might cause complications during pregnancy (Maintz et al., 2008). About 1% of the general population suffers from histamine intolerance and reduced activity of DAO is implicated in the devel- opment of the typical symptoms, like nausea, vomiting, diarrhea, http://dx.doi.org/10.1016/j.jbiotec.2016.04.002 0168-1656/© 2016 Elsevier B.V. All rights reserved.
Transcript of Journal of Biotechnology - deficitdao.org

Cp
EPBa
b
c
d
e
a
ARRAA
KDRCCAA
1
hta((eKt
h0
Journal of Biotechnology 227 (2016) 120–130
Contents lists available at ScienceDirect
Journal of Biotechnology
j ourna l ho me pa ge: www.elsev ier .com/ locate / jb io tec
haracterization of recombinant human diamine oxidase (rhDAO)roduced in Chinese Hamster Ovary (CHO) cells
lisabeth Gludovacza,b, Daniel Mareschc,1, Maximilian Bontad,1, Helen Szöllösib,1,aul G. Furtmüllerc, Robert Weike, Friedrich Altmannc, Andreas Limbeckd, Nicole Bortha,ernd Jilmab,∗, Thomas Boehmb
Department of Biotechnology, University of Natural Resources and Life Sciences, Muthgasse 18, 1190 Vienna, AustriaDepartment of Clinical Pharmacology, Medical University of Vienna, Waehringer Guertel 18-20, 1090 Vienna, AustriaDepartment of Chemistry, University of Natural Resources and Life Sciences, Muthgasse 18, 1190 Vienna, AustriaInstitute of Chemical Technologies and Analytics, Vienna University of Technology, Getreidemarkt 9, 1060 Vienna, AustriaPolymun Scientific Immunbiologische Forschung GmbH, Donaustraße 99, 3400 Klosterneuburg, Austria
r t i c l e i n f o
rticle history:eceived 11 December 2015eceived in revised form 16 March 2016ccepted 4 April 2016vailable online 7 April 2016
eywords:AOecombinant human diamine oxidaseHOhinese Hamster Ovary
a b s t r a c t
Human diamine oxidase (hDAO) efficiently degrades polyamines and histamine. Reduced enzyme activ-ities might cause complications during pregnancy and be involved in histamine intolerance. So far hDAOhas been characterized after isolation from either native sources or the heterologous production in insectcells. Accessibility to human enzyme is limited and insect cells produce non-human glycosylation patternsthat may alter its biochemical properties.
We present the heterologous expression of hDAO in Chinese Hamster Ovary (CHO) cells and a three steppurification protocol. Analysis of metal content using ICP-MS revealed that 93% of the active sites wereoccupied by copper. Topaquinone (TPQ) cofactor content was determined using phenylhydrazine titra-tion. Ninety-four percent of DAO molecules contained TPQ and therefore the copper content at the activesite was indirectly confirmed. Mass spectrometric analysis was conducted to verify sequence integrity of
miloride-sensitive amine oxidaseOC1
the protein and to assess the glycosylation profile. Electronic circular dichroism and UV–vis spectra datawere used to characterize structural properties. The substrate preference and kinetic parameters werein accordance with previous publications.
The establishment of a recombinant production system for hDAO enables us to generate decentamounts of protein with negligible impurities to address new scientific questions.
© 2016 Elsevier B.V. All rights reserved.
. Introduction
Human diamine oxidase (hDAO, E.C. 1.4.3.6), formerly namedistaminase or amiloride-binding protein (ABP1), is encoded byhe AOC1 gene and belongs to the group of copper-containingmine oxidases (McGrath et al., 2010b). CAOs are found in bacteriaParsons et al., 1995; Wilce et al., 1997; Zeng and Spiro, 2013), yeastDove et al., 1996; Klema et al., 2013; Li et al., 1998), plants (Cona
t al., 2014; Kumar et al., 1996), and mammals (Finney et al., 2014;aitaniemi et al., 2009; Salmi and Jalkanen, 2005). They oxida-ively deaminate primary amines to the corresponding aldehydes,
∗ Corresponding author.E-mail address: [email protected] (B. Jilma).
1 These authors contributed equally to this work.
ttp://dx.doi.org/10.1016/j.jbiotec.2016.04.002168-1656/© 2016 Elsevier B.V. All rights reserved.
ammonia and hydrogen peroxide through a two-step ping-pongmechanism (Brazeau et al., 2004; Mure et al., 2002):
RCH2NH2 + O2 + H2O → RCHO + NH3 + H2O2
Human DAO preferentially degrades histamine and polyaminessuch as putrescine, cadaverine, spermine and spermidine (Elmoreet al., 2002; Schwelberger et al., 2013a). It is an important enzymein the extracellular catabolism of histamine (Bieganski et al., 1983).High levels of hDAO expression are found in the intestine, wherethey are predominantly associated with mature enterocytes in thevillus tips (Wollin et al., 1998). Human DAO protein is also abun-dant in the kidney (Schwelberger et al., 2013b; Schwelberger et al.,1998) and placenta, where a decreased enzyme activity might cause
complications during pregnancy (Maintz et al., 2008).About 1% of the general population suffers from histamineintolerance and reduced activity of DAO is implicated in the devel-opment of the typical symptoms, like nausea, vomiting, diarrhea,

Biotec
hao
iccmPc(
cCRdwemogteipc(2bacdmtuswbAt2Pclwla(dwaboiuEwldsca
sml
E. Gludovacz et al. / Journal of
eadache, rhinorrhea, urticaria, hypotension and in severe casesnaphylaxis (Maintz and Novak, 2007). Nevertheless, the exact rolef hDAO in the pathogenesis of histamine intolerance is not clear.
X-ray crystallographic analyses of recombinant hDAO fromnsect cells confirmed the typical homodimeric structure of copper-ontaining amine oxidases (McGrath et al., 2009). Each active siteontains one Cu(II) ion in a distorted square-pyramidal arrange-ent and the topaquinone (TPQ) cofactor (McGrath et al., 2009;
arsons et al., 1995). Each hDAO subunit contains two additionalations – presumably Ca2+ – binding sites with unknown functionMcGrath et al., 2009).
Native DAO has been characterized from various origins usingumbersome purification protocols (Baylin and Margolis, 1975;orda et al., 1984; Crabbe et al., 1976; Mizuguchi et al., 1994;inaldi et al., 1982; Tamura et al., 1989). Elmore et al. (2002) con-ucted the first heterologous expression of hDAO in insect cellsith a detailed biochemical characterization of the enzyme. How-
ver, insect cells produce non-human like glycosylation, whichay affect the enzyme’s properties. Most insect cell glycans are
f paucimannosidic type that may be substituted with immuno-enic �1-3fucose at the N-acetylglucosamine core, as opposed tohe complex type glycosylation of mammalian cell lines (Ghaderit al., 2012). In addition, the expression of complex proteins innsect cells might influence their solubility, physical stability andharmacokinetics (Hollister et al., 2002; Varki, 1993). Anotheropper-containing amine oxidase, vascular adhesion protein-1VAP-1 or AOC3) has been expressed in CHO cells (Ernberg et al.,010). In the following sections we describe the generation of sta-le CHO-K1 clones producing hDAO, the purification of the enzymend its subsequent characterization. Recombinant hDAO (rhDAO)an be used for example to screen chemical libraries of approvedrugs for DAO inhibitors. Sattler et al. (1985) published a list ofedications that interfered with DAO activity, but in the mean-
ime many new drugs have been approved. These investigatorssed partially purified DAO from human and canine intestinal tis-ue, which might contain other proteins and enzymes interferingith DAO activity or the activity assay. Using rhDAO in a defined
uffer system would circumvent these methodological limitations.ny substance that might interfere with DAO activity can be readily
ested. Ethanol is described as DAO inhibitor (Maintz and Novak,007), but proper biochemical studies have not been performed.urified rhDAO can be also used to immunize mice to obtain mono-lonal antibodies, which recognize native hDAO. No hDAO enzymeinked immunoassay (ELISA) is currently commercially available,
hich can accurately quantify the absolute amount of hDAO in bio-ogical samples like serum or plasma. We tested two commerciallyvailable ELISAs, but their performance was far from satisfyingdata not shown). One ELISA was not able to measure DAO levelsuring third trimester pregnancy and the standards of this ELISAere not recognized by the second ELISA. It was not further evalu-
ted. In the manual of the second ELISA the difference in the medianetween DAO levels in pregnant women and healthy controls isnly about 2-fold. Nevertheless, during pregnancy DAO activitiesncrease several hundredfold as measured by several investigatorssing radioactive putrescine (Maintz et al., 2008). The range of thisLISA using the lowest and highest standard is only 27-fold. Whene measured a pregnancy serum at different dilutions, the calcu-
ated DAO levels differed more than 10-fold. This lack of linearityisqualifies the second ELISA for exact DAO concentration mea-urements. The reasons might be the use of two polyclonal sera forapturing and detection. Purified rhDAO would be an ideal sources standard for ELISA development.
Purified rhDAO might also be used to test the efficacy of DAOubstitution in different animal models, where excess histamineight play an important pathophysiological role like in anaphy-
axis.
hnology 227 (2016) 120–130 121
The availability of large quantities of purified rhDAO producedin CHO cells will allow to investigate important research questions,which until now could not be addressed properly.
2. Materials and methods
2.1. Cloning of the rhDAO expression plasmid
All cloning techniques were conducted according to proto-cols described by Green and Sambrook (2012) and according tothe manufacturer’s instructions. The exon sequence of the humandiamine oxidase gene (Chassande et al., 1994), flanked by alter-native untranslated regions and signal peptides obtained fromUTR (UniTargeting Research AS, Bergen, Norway), was synthesizedby Eurofins MWG Operon to include the restriction sites HindIIIand ClaI. The construct was cloned into a vector containing aCMV promoter, SV40 polyadenylation signal, a blasticidin resis-tance gene and the two heterospecific FRT sites, namely FRT-wt(Flp-recombinase target, wild-type) and FRT-F5 (Flp-recombinasetarget, mutant F5) (Schlake and Bode, 1994). The plasmid included apUC origin of replication and an ampicillin resistance gene. Restric-tion digest of the hDAO gene and the vector was performed usingHindIII and ClaI FastDigest restriction enzymes (Thermo Scien-tific), followed by ligation (T4 DNA ligase, New England Biolabs)and amplification of the final plasmid. Correctness of sequenceof all constructs was verified by DNA sequencing (Eurofins MWGOperon).
2.2. Generation of a stable cell line and cultivation
For the generation of stable DAO-producing CHO cell linesRecombinase-Mediated Cassette Exchange (RMCE) was performed(Schlake and Bode, 1994). A target CHO-K1 cell line with a stablyintegrated CD4 reporter gene flanked by two heterospecific FRT-sites was cultivated in CD CHO medium (Gibco®), supplementedwith 8 mM l-glutamine (Merck Millipore) and 0.2% Anti-ClumpingAgent (Gibco®) in a shaker-incubator at 37 ◦C, 7% CO2 and 140 rpm.1 × 107 cells were transfected with 7 �g of the rhDAO expres-sion plasmid and 21 �g of the Flp-recombinase expression plasmidpPGKFLPobpA (Addgene) (Raymond and Soriano, 2007) usingAmaxa NucleofectorTM I/program H-14 and the Amaxa cell linenucleofector kit V (Lonza Group Ltd.). The growth medium wassupplied with 10 �M CuSO4 (Sigma Aldrich), based on the resultsof preliminary test batches with various CuSO4 concentrations(data not shown). Six days post transfection 10 �g/mL BlasticidinS HCl (Life Technologies) was added to the culture medium toselect for cells that have exchanged the CD4 reporter gene for thehDAO gene. After recovery of cell viability fluorescence activatedcell sorting was performed. 1 × 107 cells were stained with CD4(VIT4)-PE antibody (Miltenyi Biotec) according to the manufac-turer’s instructions. rhDAO secreting cells were stained with 1:20dilutions of anti-ABP1 antibody produced in rabbit (SAB1410491,Sigma Aldrich) and anti-rabbit IgG (whole molecule)-FITC antibodyproduced in goat (Sigma Aldrich), as described by Pichler et al.(2009). rhDAO secreting, CD4 negative cells were sorted on anAstrios cell sorter (Beckman Coulter). PE was detected with the561 nm laser and a 579/16 bandpass filter, FITC with the 488 nmlaser and a 526/52 bandpass filter. One hundred thousand of thetop 5% rhDAO producing CHO-K1 cells were sorted into a 6-well
plate and expanded.For batch cultivation the stable CHO-K1 rhDAO cells wereseeded at a viable cell density of 0.15 × 106 cells/mL and incubatedfor 7 days.

1 Biotec
2
aUw5mnrAa3p(nU(rH(5c157ubuw
tu2cP
2
CwLNb5pTRpS
PPmM2tbTritw(
22 E. Gludovacz et al. / Journal of
.3. Purification of rhDAO from culture supernatant
The CHO-K1 batch culture was centrifuged for 10 min at 170gnd the supernatant was sterile filtered through Stericup® Filternits (Merck Millipore). Ultra- and diafiltration was performedith the Labscale TFF System in combination with a Pellicon XL
0 Ultrafiltration Cassette – Biomax Polyethersulfon with a 100 kDaolecular weight cut-off (both Merck Millipore). The culture super-
atant was concentrated 15-fold and the culture medium waseplaced by a 50 mM Hepes buffer, pH 7.5 (Hepes buffer grade,ppliChem). Ammonium sulfate (for analysis, AppliChem) wasdded under continuous mixing over 45 min until a saturation of5% (w/v) was achieved. After two hours of mixing at room tem-erature, the sample was ultracentrifuged for 20 min at 25,000 rpmBeckman 45Ti rotor, Beckman L70 ultracentrifuge). The super-atant containing rhDAO was filtered through a Stericup® Filternit (Merck Millipore) and loaded onto three HiTrap Phenyl FF
low sub) prepacked 5 mL columns connected in series at a flowate of 2.5 mL/min using an Äkta purifier HPLC device (both GEealthcare). Stepwise elution was performed with 30% (w/v), 25%
w/v), 20% (w/v), 12.5% (w/v) and 0% (w/v) ammonium sulfate in0 mM Hepes buffer. The 12.5% (w/v) ammonium sulfate eluate wasoncentrated and desalted using Merck Millipore’s Amicon Ultra-5Centrifugal Filter Units (100 kDa molecular weight cut-off) and a0 mM Hepes buffer containing 150 mM KCl (Merck Millipore), pH.2. The eluate was loaded onto a Superdex 200 pg (26/600) col-mn (GE Healthcare) at a flow rate of 2.0 mL/min using the sameuffer. Finally the rhDAO containing fraction was concentratedsing Amicon Ultra-15Centrifugal Filter Units (100 kDa moleculareight cut-off).
Purified rhDAO was quantified using NanoDrop 1000 spec-rophotometer (Thermo Scientific), protein A280 mode: molec-lar weight = 170 kDa, molar extinction coefficient at 280 nm:89,000 M−1 cm−1. Molecular weight and molar extinction coeffi-ient were calculated from the amino acid sequence of hDAO usingrotParam (Gasteiger et al., 2005).
.4. SDS-PAGE and western blot
For non-reducing SDS-PAGE analyses a Mini-PROTEAN® Tetraell in combination with 4–15% Mini-PROTEAN® TGXTM Gelsith 10 wells (Bio-Rad) and a PageRuler Plus Prestained Protein
adder (Thermo Scientific) was used. Samples were mixed withuPAGE® LDS Sample Buffer (4×) (Life Technologies) and incu-ated for 30 min at room temperature. For reducing conditions% 2-mercaptoethanol (Sigma Aldrich) were added to the sam-le buffer. Gels were run in 1× Laemmli buffer containing 25 mMris Base (Ultrol® Grade, Calbiochem), 250 mM Glycine (Pufferan®,oth) and 0.1% SDS (VWR Prolabo) at 300 V for 15 min. Staining waserformed on a shaker for 1 h at room temperature using Acquatain (Acqua Science).
For western blotting the gel was not stained but blotted on aVDF membrane using Trans-Blot® TurboTM Mini PVDF Transferacks and the Trans-Blot® TurboTM Transfer System (BioRad). Theembrane was blocked for two hours with 1× PBS (all componentserck Millipore) containing 3% (w/v) milk powder and 0.1% Tween
0 (Merck Millipore) and incubated overnight at 4 ◦C in a 1:500 dilu-ion of anti-ABP1 antibody (HPA031032, Sigma Aldrich) in blockinguffer. The membrane was washed 3 × 5 min with 1× PBS + 0.1%ween 20 followed by the addition of the IRDye® 800CW goat anti-abbit IgG (H + L) secondary antibody (Li-Cor) in a 1:5000 dilution
n blocking buffer. After 90 min of incubation at room tempera-ure, washing was performed as described above. The membraneas scanned at 800 �m using the Odyssey Infrared Imaging SystemLi-Cor).
hnology 227 (2016) 120–130
2.5. Peptide and N-glycan analysis using LC–ESI-MS
Twenty �g of sample were divided into two parts of 10 �geach. Reduction with DTT (5 mM), S-alkylation with iodoac-etamide (25 mM) and digestion with sequencing grade modifiedtrypsin/chymotrypsin (Promega) (0.2 �g) were performed. Thepeptide mixture was analysed using a Dionex Ultimate 3000 sys-tem directly linked to a QTOF instrument (maXis 4 G ETD, Bruker)equipped with the standard ESI source in positive ion, DDA mode(=switching to MSMS mode for eluting peaks). MS-scans wererecorded (range: 150–2200 m/z) and the 6 highest peaks wereselected for fragmentation. Instrument calibration was performedusing ESIcalibration mixture (Agilent). For separation of the pep-tides a Thermo BioBasic C18 separation column (5 �m particle size,150 × 0.360 mm) was used. A gradient from 95% solvent A—65 mMAmmonium formate buffer, pH 3.0 (Formic acid supplied by CarlRoth, Ammonia supplied by VWR BDH Prolabo) – and 5% solvent B– 100% Acetonitrile (VWR BDH Prolabo) – to 32% B in 45 min wasapplied, followed by a 15 min gradient from 32% B to 75% B, at aflow rate of 6 �L/min.
The analysis files were converted using Data Analysis 4.0(Bruker) to XML files, which are suitable to perform MS/MS ionsearches with MASCOT (embedded in ProteinScape 3.0, Bruker)for protein identification. For the searches the SwissProt databasewas used. Peptide MS/MS data were evaluated against the targetsequence using X! Tandem (www.thegpm.org/tandem/) with thefollowing settings: reversed sequences no; check parent ions forcharges 1, 2 and 3 yes; models found with peptide log e lower−1 and proteins log e lower −1; residue modifications: oxidationM, W and deamidation N, Q; isotope error was considered; frag-ment type was set to monoisotopic; refinement was used withstandard parameters; fragment mass error of 0.1 Da and ±100 ppmparent mass error; fragment types b and y ions; maximum par-ent ion charge of 3; missed cleavage sites allowed was set to 2;semi-cleavage yes.
Glycan analyses were performed with purified rhDAO frombatches of three individual CHO-K1 clones. One clone produced therhDAO wild-type, one a fusion construct and one a point mutant ofrhDAO wild-type. The fusion construct and the point mutation wereunlikely to have an effect on glycosylation.
2.6. ICP-MS
Purified rhDAO samples of two individual batches were dialyzedagainst a 50 mM Hepes buffer, pH 7.2 (prepared from 1 M Hepesbuffer, Biochrom). The sample concentrations were 12.4 �M (Batch1) and 7.1 �M (Batch 2). For all sample dilutions 1% (v/v) HNO3was used, which was prepared from high purity water (resistivity18.2 M� cm) dispensed from a Barnstead EASYPURE II water sys-tem (ThermoFisher Scientific) and concentrated HNO3 (for analysis,Merck). Liquid standards for ICP-MS measurements (ICP-multi ele-mental solution VIII, 24 elements at 100 mg/L in diluted HNO3 andindium standard for ICP, 1 g/L in 5% HNO3) were purchased fromSigma-Aldrich. ‘Tune B’ solution for tuning of the ICP-MS instru-mentation was supplied by ThermoFisher Scientific.
The presented measurements were performed usingquadrupole ICP-MS instrumentation (iCAP Qc, ThermoFisherScientific). Sample introduction was established using a Peltiercooled spray chamber equipped with a concentric PFA nebulizer.Automated sample introduction was accomplished using an ESISC2-DX sample changer (Elemental Scientific, Inc.). Qtegra soft-ware provided by the manufacturer was used for data recording
and processing.A detailed description of the measurement parameters andsample preparation steps can be found in the Supplementary infor-mation section.

Biotec
2
1acp1hbptawam
2
CfltUp1asUMUr11uapwtbwsaaKfr2K
3r
2
daobfis
were also evaluated using linear plots according to Lineweaver-
E. Gludovacz et al. / Journal of
.7. Phenylhydrazine titration
Purified rhDAO from two individual batches was analyzed.00 �L purified rhDAO (Batch 1 = 12.4 �M, Batch 2 = 7.1 �M)nd 100 �L 50 mM Hepes with 150 mM KCl buffer as negativeontrol were transferred to a transparent flat-bottom 96-welllate (NuncTM Thermo Scientific). For the titration of Batch 1 a00 �M solution, for Batch 2 a 50 �M solution of phenylhydrazineydrochloride (Sigma Aldrich) in 50 mM Hepes with 150 mM KCluffer were freshly prepared before use. Aliquots of 2 �L of thehenylhydrazine solution were added to both sample and nega-ive control. The titration was stopped when no major changes inbsorbance after further addition of the reagent occurred. The plateas shaken and incubated for 10 min at room temperature, before
bsorbance was measured at 445 nm with Tecan’s Infinite 200 PROultimode reader.
.8. Electronic circular dichroism (ECD) and UV–vis spectroscopy
ECD spectra were recorded on Applied Photophysics’hirascanTM CD spectrometer at 25 ◦C. The instrument wasushed with nitrogen at a flow rate of 5 L/min and allowed simul-aneous UV–vis and ECD monitoring. Measurements in the nearV and visible region (300–800 nm) range were performed withurified rhDAO at a concentration of 82.4 �M in 50 mM Hepes with50 mM KCl buffer at pH 7.2. The instrument parameters were sets follows: path length: 10 mm; spectral bandwidth: 1 nm; stepize: 1 nm; scan time per point: 5.0 s. For measurements in the farV, the Hepes buffer was replaced by a 50 mM KH2PO4 (Merckillipore) buffer, pH 7.2, using Amicon Ultra-15Centrifugal Filternits (50 kDa molecular weight cut-off). The concentration of
ecombinant hDAO was diluted to 2.9 �M. The path length was at mm, spectral bandwidth 3 nm and scan time-per-point was set at0.0 s. For prediction of secondary structure from spectra datapointnits were converted from ellipticity to molar circular dichroismnd evaluated by K2D3 software (wavelength range 200–240 nm,rotein size 170 kDa) (Louis-Jeune et al., 2012). The instrumentas equipped with a Peltier element for temperature control and
emperature-mediated denaturation was monitored at 218 nmetween 25 ◦C and 94 ◦C. Temperature was increased stepwiseith 1.0 ◦C/min and time-per-point were set at 10.0 s. Baseline
pectra were subtracted from each spectrum. Thereafter at 94 ◦C spectrum in the far UV region was recorded as described abovend secondary structure prediction was again performed using2D3 software. Consequently, the temperature was decreased
rom 94 ◦C to 25 ◦C as described above to investigate whetherefolding of rhDAO occurs. Finally, a spectrum was recorded at5 ◦C and secondary structure distribution was calculated using2D3 software.
A UV–vis spectrum (300–700 nm) was recorded on a Hitachi U-900 spectrophotometer at a scan speed of 120 nm/min. PurifiedhDAO at a concentration of 82.4 �M was used.
.9. Substrate preference
Oxidation preference of rhDAO for various substrates wasetermined using a chemiluminescence-based enzyme activityssay according to Schwelberger and Feurle (2007). Diaminexidase released hydrogen peroxide is used to oxidize luminoly horseradish peroxidase. The generated light can be quanti-ed in a luminometer. Recombinant hDAO from CHO-K1 cultureupernatant in PBS (see above) with 0.1% HSA (Albunorm 20%,
hnology 227 (2016) 120–130 123
Octapharma)4 was used. The amount of rhDAO in the CHO super-natant was quantified with an in-house developed DAO ELISA usingpurified rhDAO as standard (data not shown). We always usedduplicates of 110 �L final volume in white luminescence plates(Porvair). The 110 �L were composed of 50 �L rhDAO and addi-tional substances in 0.1% HSA PBS and 50 �L luminol solutionfrom Amersham ECL Western Blotting Detection Kit and 2 �g/mLhorse radish peroxidase (HRP, P6782, Sigma Aldrich). Measure-ments were performed with an unadjusted (pH 9.2), as well asa pH 8.0 adjusted luminol solution. The reaction was startedwith the addition of 10 �L substrate solution at a final concentra-tion of 360 �M. We used putrescine (P5780), cadaverine (C8561),histamine (53300), benzylamine (185701), spermine (85605), sper-midine (85578) (all Sigma Aldrich) as substrates. The RLU (relativelight units) were measured for 120 min at 30 ◦C in 5 min intervals.We conducted all measurements in duplicates.
All measurements were performed in a Victor2TM 1420Microtiter Plate Reader (Perkin Elmer).
2.10. Steady-state kinetics
The kinetic parameters of rhDAO for putrescine were deter-mined using DAOABA activity assay. Its characteristics aredescribed by Holmstedt and Tham (1959).
The assay was performed in UV-compatible 96-well half-areaclear polystyrene flat bottom, untreated plates (Sigma Aldrich) ina total reaction volume of 170 �L corresponding to a light path of1 cm. PBS (without MgCl2 and CaCl2, Gibco) with 0.1% human serumalbumin (HSA, Human-Albumin 20%, Behring) was used as assaymatrix.
o-Aminobenzaldehyde (oABA) was purchased from SigmaAldrich (A9628), dissolved in ethanol absolute (25 mg/mL, 206 mM)and stored at −20 ◦C. A final concentration of 1 mM oABA (con-taining 83 mM ethanol) was used for the assay. Ethanol does notinfluence DAO activity at this concentration (data not shown). Puri-fied rhDAO was added at a final concentration of 1 �g/mL (6 nM).The assay was pre-incubated for 30 min at room temperature.Putrescine (P5780, Sigma Aldrich) concentrations of 0, 25, 50, 100,150, 200, 300, 400 �M were added to start the reaction.
We quantified 2,3-trimethylene-1,2-dihydroquinazoliniumaccumulation by absorbance measurements at 450 nm. Measure-ments were performed at 30 ◦C every 3 min in a Victor2TM 1420Microtiter Plate Reader (PerkinElmer). The means of triplicateabsorbance values were calculated and used for plotting thecumulative absorbance over time. The rate of 2,3-trimethylene-1,2-dihydroquinazolinium formation at 30 ◦C was calculated bylinear regression in the linear range of the ABS curves.
Using wells without DAO, oABA or putrescine the ABS changewas less than 1% compared to wells containing all necessaryingredients (data not shown). The rate of 2,3-trimethylene-1,2-dihydroquinazolinium formation was normalized to the amount of1 mg enzyme. In order to compensate signal loss due to the wave-length shift from 430 nm to 450 nm, the enzyme activity valueswere corrected by 12.5% see Fig. 1 in Holmstedt and Tham (1959).
Nonlinear regression was carried out with GraphPad Prism tocalculate Km and kcat. Parameters of Michaelis-Menten kinetics
Burke, Eadie-Hofstee and Hanes-Wolf.
4 a 20% human serum albumin (HSA) solution, is authorized for human use and96% of the protein content is HSA. Its further ingredients do not inhibit DAO activity(data not shown).

124 E. Gludovacz et al. / Journal of Biotechnology 227 (2016) 120–130
Fig. 1. SDS-PAGE and Western blot analysis of purification steps of rhDAO fromBatch 1.(A) Lane 1: PageRuler Plus Prestained Protein Ladder. Lane 2: Ultra- and diafil-trated culture supernatant of CHO-K1 hDAO from 7 days batch culture, 10.5 �g. Lane3: after 35% ammonium sulfate precipitation, 10.5 �g. Lane 4: after hydrophobicinteraction chromatography, 10.5 �g. Lane 5: after size exclusion chromatography,5.3 �g. (B) Western blot of an undiluted culture supernatant from 5 day batch cul-tu
3
3
ggdig
taiace7p
Table 1Metal ion and TPQ content of purified rhDAO from two individual batches deter-mined via ICP-MS analysis and phenylhydrazine titration.
Cu Zn Ca Mg Fe Mn Co Ni TPQ
Batch 1 1.73 1.12 3.05 0.17 0.06 ND ND ND 1.78Batch 2 1.99 0.23 1.53 0.13 0.09 0.003 0.0002 0.01 1.97
Metal ion and TPQ concentrations in the samples were normalized to molar equiv-
FHit
ure of transiently rhDAO producing CHO-S cells. Lane 1: reduced sample. Lane 2:nreduced sample.
. Results and discussion
.1. Stable expression and purification
Producer cell lines were established by transfecting the hDAOene into a preselected CHO-K1 cell line that contained the hCD4ene flanked by FRT target sites. The resulting CHO-K1 rhDAO pro-ucer cell lines showed very homogeneous rhDAO production. This
s expected for a cell line with a single defined chromosomal inte-ration site.
Using ammonium sulfate precipitation, hydrophobic interac-ion and size exclusion chromatography we obtained rhDAO with
high degree of purity. Size exclusion chromatography resultedn three individual peaks (see Fig. S1). Eluate 1 contained proteinsnd aggregates of higher molecular weight. Eluate 3 was mainly
omposed of low molecular weight impurities. Recombinant hDAOluted in a single peak with a retention time of approximately0 min. Fig. 1 depicts the enrichment of the enzyme with eachurification step. Comparison of purified rhDAO on the SDS-PAGEig. 2. LC–ESI-MS sequence analysis of purified rhDAO.ighlighted fragments were identified. (Green): fragments after tryptic digestion, i
dentified with GPM software. (Red): manual identification. (Yellow): N-glycosylhis figure legend, the reader is referred to the web version of this article.)
alents of purified rhDAO.ND. . . not detectable.
gel and an enzyme specific western blot of culture supernatantof a transient expression of rhDAO in CHO-S reveals a consistentband pattern. Besides the intense band of dimeric DAO around200 kDa other bands are visible. The second band with approxi-mately 130 kDa might be due to fragmentation of rhDAO. Becausethe interchain disulfide bridge is located on the very C-terminus(cysteine 736), it might be speculated that the N-terminus of oneDAO monomer is fragmented and the fragment is still linked tothe other monomer via the interchain disulfide bond. Upon reduc-tion the fragmented dimer is cleaved into an intact monomer and a55 kDa N-terminal fragment of DAO, which appears reproducibly asa very faint band on western blots (Fig. 1B). A reducing SDS-PAGE,followed by enzyme specific western blotting indicated that theband at approximately 100 kDa corresponds to the DAO monomer(Fig. 1). Solutions of purified rhDAO exhibit a pink coloration dueto the TPQ cofactor (Rinaldi et al., 1999). Densitometric analysis ofthe SDS-PAGE showed a purity of at least 95% (data not shown).
For Batch 2 we calculated a purification yield of 82%. The culturesupernatant with a total protein concentration of 1.85 g/L contained57.4 mg rhDAO at a concentration of 20 mg/L. DAO was quanti-fied based on putrescine oxidation using the luminescence-basedactivity assay. The purification resulted in 47.2 mg enzyme.
3.2. Peptide and N-glycan analysis using LC–ESI-MS
Mass spectrometric analysis of purified rhDAO verified theintegrity of the amino acid sequence, particularly of the N-andC-terminal ends (Fig. 2). GPM software identified fragments aftertryptic and chymotryptic digestion of the enzyme. Manual eval-uation identified two further fragments and four N-glycosylationsites. Sixty-one percent of the amino acid sequence was therebyverified including both N- and C-terminal regions. The correctexpression and processing of the intact enzyme in CHO-K1 cells was
confirmed and the purification protocol did not lead to any trun-cation of rhDAO. The signal peptide was accurately cleaved beforesecretion into the culture medium.dentified with GPM software. (Blue): fragments after chymotryptic digestion,ation sites, manual identification. (For interpretation of the references to colour in

E. Gludovacz et al. / Journal of Biotechnology 227 (2016) 120–130 125
Fig. 3. LC–ESI-MS glycosylation analysis of purified rhDAO from 3 individual batches.(A) N-Glycosylation site Asn110, (B) N-Glycosylation site Asn168, (C) N-Glycosylation site Asn538, (D) N-Glycosylation site Asn745. Relative abundance of glycans werecalculated for each N-glycosylation site from total peak areas. Mean values of three individual batches and standard errors of mean are depicted. Man, M. . . mannose, Na. . .N . . N-ao ected
a rding
s(gisesnmlr
-acetylneuraminic acid = sialic acid, Hex. . . hexose, A. . . galactose, F. . . fucose, Gn.n www.proglycan.com. (E) Overview of the most abundant N-glycan structures det
schematic illustration of possible isoforms. Monosaccharides are represented acco
Mass spectrometric glycan analysis revealed that all four pos-ible N-glycosylation sites Asn110, Asn168, Asn538 and Asn745numbering according to (McGrath et al., 2009)) of rhDAO werelycosylated (see Fig. 3). Previous reports on hDAO expressed innsect cells suggested glycosylation of only three N-glycosylationites (McGrath et al., 2009). N-sites Asn168, Asn538 and Asn745xhibit CHO-typical complex bi-, tri- and tetra-antennary glycantructures, with a high degree of sialylation. The addition of termi-al sialic acid residues is known to be crucial for product quality ofost protein therapeutics, since it influences their circulatory half-
ife (Hossler et al., 2009). Some glycans identified also carry fucoseesidues that play diverse biological roles (Becker and Lowe, 2003).
cetylglucosamine. A detailed explanation of the glycan nomenclature can be found(≥10% relative abundance). Since linkage analysis was not performed, this is merely
to the guidelines of the Consortium of Functional Glycomics.
The N-glycosylation site Asn110 exhibits a high degree of man-nosylation. Go et al. also found two N-glycosylation sites to bepopulated with high mannose glycans, when producing HIV-1gp120 envelope proteins in CHO cells and noted that the cell typeand the metabolic state of the cell line during protein expressionhave an influence on the overall ratio of high mannose to complex-type glycans (Go et al., 2013). We observed unique glycosylationpatterns at each glycosylation site. While mannose type glycanscan exclusively be found on N-glycosylation site Asn110, complextri- and tetra-antennary structures appear only on N-glycosylation
site Asn168. N-glycosylation sites Asn538 and Asn745 carry solelycomplex biantennary glycans.
126 E. Gludovacz et al. / Journal of Biotechnology 227 (2016) 120–130
Fig. 4. Phenylhydrazine titration of TPQ to quantify the percentage of active sites inpurified rhDAO.Batch 1: 100 �L of a 12.4 �M rhDAO solution (50 mM Hepes with 150 mM KCl, pH7.2) were titrated with 2 �L aliquots of a 100 �M phenylhydrazine solution. Batch2: 100 �L of a 7.1 �M rhDAO solution (50 mM Hepes with 150 mM KCl, pH 7.2) weretmv
rccefe
3
tm(
tcataI12TTaaBtt
dwp(cTdvTera
bt
Table 2Secondary structure distribution of purified rhDAO.
% �-helices % �-strands % random coils
K2D3 14.9 ± 0.5 24.9 ± 0.6 60.2 ± 0.1PDB (3HIG) 13.3 32.1 54.6Raptor X 11.0 32.0 56.0
The far UV ECD-spectra of Batch 1 and 2 were evaluated with K2D3 software and
itrated with 2 �L aliquots of a 50 �M phenylhydrazine solution. Absorbance waseasured at 445 nm, 10 min after the addition of phenylhydrazine. The absorbance
alues were plotted against the amount of phenylhydrazine added.
When comparing N-glycosylation patterns of purified wild-typehDAO, a fusion protein and a point mutant from individual CHO-K1lones, we observed a higher degree of fucosylation of certain gly-ans on Asn538 and Asn745 in case of the fusion protein, hence thelevated standard errors for the respective glycans in Fig. 3. Apartrom that, a high degree of consistency in glycosylation patterns,ven between individual CHO-K1 clones, could be demonstrated.
.3. Metal content and TPQ cofactor
ICP-MS analysis was performed to determine the metal con-ent of purified rhDAO. Concentrations of copper, zinc, calcium,
agnesium, iron, manganese, cobalt and nickel were determinedTable 1).
Studies of Hansenula polymorpha copper amine oxidase revealedhat substitution of copper by zinc leads to significant structuralhanges of the active site, predominantly at the metal binding sitend Tyr405, the precursor of TPQ cofactor, and thereby to inactiva-ion of the enzyme (Chen et al., 2000). It is therefore desirable tochieve a high Cu/Zn ratio when expressing hDAO heterologously.n case of Batch 1 21.4 �M copper was found in the sample using2.4 �M rhDAO. Considering that 1 mol of dimeric enzyme binds
mol of Cu, 86.7% of possible sites are occupied with this metal ion.his corresponds to 1.73 mol copper per mol of dimeric enzyme.he remaining 13.3% are substituted with other metal ions, presum-bly Zn. We hypothesize that the remaining metal ions detectedre bound to the four peripheral cation binding sites. In case ofatch 2, containing 7.1 �M purified rhDAO, a copper concentra-ion of 14.1 �M was determined. One mol of dimeric enzyme isherefore occupied with 1.99 mol copper.
The findings of ICP-MS analysis was supported by phenylhy-razine titration. Phenylhydrazine reacts stoichiometrically onlyith active copper-TPQ complex and forms a yellow coloredhenylhydrazone adduct, which can be measured at 445 nmElmore et al., 2002; Janes and Klinman, 1991). In case of Batch 1 aonstant absorbance value of 0.17, meaning equimolarity betweenPQ and phenylhydrazine, was reached at 2.2 nmol phenylhy-razine added to 1.24 nmol purified rhDAO (Fig. 4). Using thesealues we calculated that 89.0% of DAO molecules carry an activePQ cofactor. This corresponds to 1.78 mol TPQ per mol of dimericnzyme. For Batch 2 we calculated that 1 mol of dimeric rhDAO car-ied 1.97 mol of reactive TPQ. Thus, the results of ICP-MS analysis
nd phenylhydrazine titration are consistent.The variability in copper and TPQ content between the twoatches might be caused by variations in viable cell density andherefore product yield. While Batch 1 showed a mean viable cell
compared with 3D structure data of hDAO expressed in insect cells (PDB ID: 3HIG)and results from secondary structure prediction tool Raptor X. Mean values andstandard errors of the mean of the two rhDAO batches are shown.
density of 8.0 × 106 cells/mL on day 7, the corresponding value was5.8 × 106 cells/mL in Batch 2. The same amount of copper was addedto both batches on day 0. Continuous addition of copper duringbatch cultivation might be considered to ensure a constant highcopper and TPQ occupancy of rhDAO molecules.
3.4. Electronic circular dichroism (ECD) and UV–vis spectroscopy
The UV–vis spectrum of purified rhDAO shows an absorptionpeak around 480 nm (see Fig. 5A), which can be attributed to theTPQ cofactor. This result is consistent with our findings in ECD-spectroscopy and with previous analyses of hDAO (Elmore et al.,2002).
ECD-spectra of purified rhDAO were recorded at visible and farUV wavelengths. Fig. 5B depicts changes in the ellipticity between300 and 800 nm. The spectrum exhibits two significant minima.Since TPQ has a broad absorption peak between 480 and 500 nm(Mure and Klinman, 1993; Rinaldi et al., 1999), we assume that thenegative band at 480 nm can be assigned to the cofactor (Elmoreet al., 2002). The minimum around 780 nm stems from the Cu (II)in a d-d transition state (Akitsu et al., 2009; Elmore et al., 2002).
At 25 ◦C a spectrum was recorded between 180 and 260 nm tostudy secondary structures of rhDAO (Fig. 6A) and evaluated withK2D3 software. We found that random coils (60.2%) are the pre-dominant secondary structure elements of rhDAO. 14.9% could beassigned to �-helices and 24.9% to �-strands. These results are con-sistent with the DAO data found in the RCSB Protein Data Bank (PDBID: 3HIG) and with the results obtained from the secondary struc-ture prediction software Raptor X (Källberg et al., 2012) (Table 2).
Furthermore a melting curve of purified rhDAO was recordedat a wavelength of 218 nm (Fig. 6B). The melting temperature wascalculated to be 63.9 ◦C (inflection point in Fig. 6B). The enzymecan therefore be considered to be thermally stable. The increase ofellipticity between 70 ◦C and 85 ◦C might be due to precipitationof rhDAO. This finding indicates that unfolding of rhDAO is irre-versible. Nevertheless a spectrum at 94 ◦C was recorded. We found asecondary structure distribution of 2.1% �-helices, 33.1% �-strandsand 64.8% random coils. Apparently unfolding of rhDAO results in aloss of mainly �-helices and a shift towards �-strands and randomcoils. To see whether refolding of rhDAO occurs, the temperaturewas decreased stepwise (1.0 ◦C/min) from 94 ◦C to room tempera-ture. As expected for many proteins, rhDAO does not refold to theinitial conformation (Fig. 6A and B) which is also underlined by thesecondary structure distribution (20.6% �-helices, 28.4% �-strands,51.0% random coils) of the refolded protein.
3.5. Substrate preference
We performed a luminescence-based activity assay of CHO cul-ture supernatant to determine substrate oxidation preference ofrhDAO. Based on the signals for each substrate at pH 8.0 (Fig. 7)
we calculated the percentage activities on the different substratesusing the putrescine signal as 100%: spermidine – 71%, cadaverine– 70%, histamine – 48%, spermine – 42% and benzylamine – 29%.When comparing these results with the data from Elmore et al.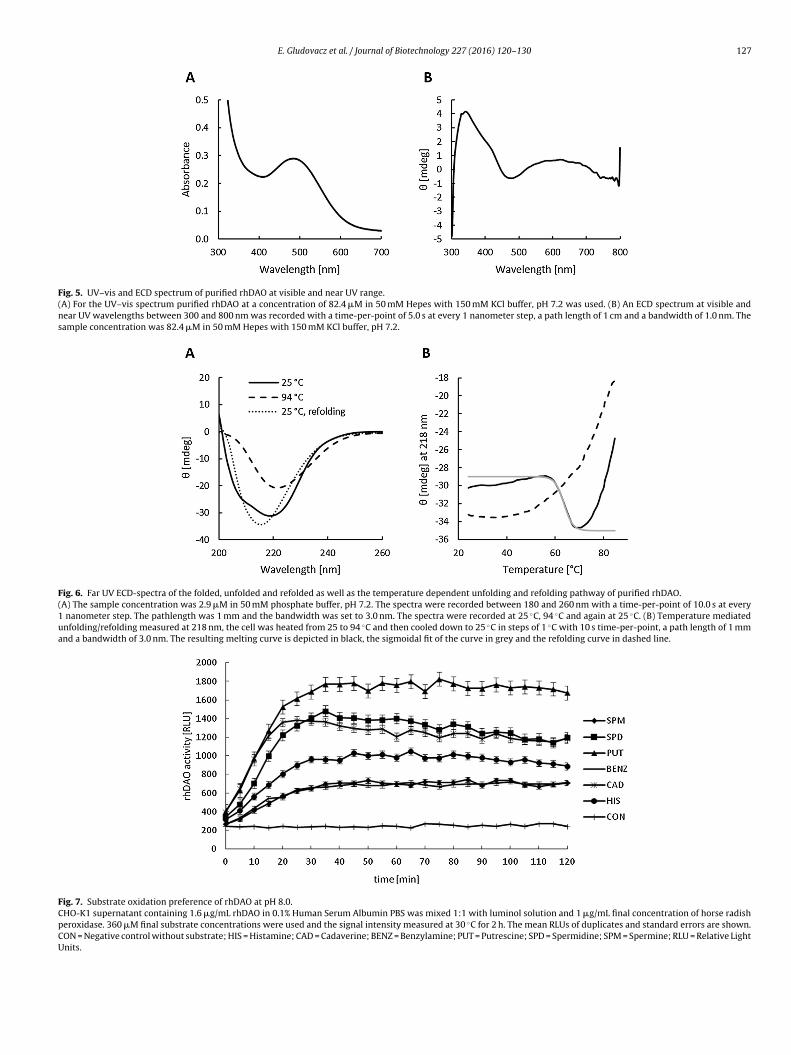
E. Gludovacz et al. / Journal of Biotechnology 227 (2016) 120–130 127
Fig. 5. UV–vis and ECD spectrum of purified rhDAO at visible and near UV range.(A) For the UV–vis spectrum purified rhDAO at a concentration of 82.4 �M in 50 mM Hepes with 150 mM KCl buffer, pH 7.2 was used. (B) An ECD spectrum at visible andnear UV wavelengths between 300 and 800 nm was recorded with a time-per-point of 5.0 s at every 1 nanometer step, a path length of 1 cm and a bandwidth of 1.0 nm. Thesample concentration was 82.4 �M in 50 mM Hepes with 150 mM KCl buffer, pH 7.2.
Fig. 6. Far UV ECD-spectra of the folded, unfolded and refolded as well as the temperature dependent unfolding and refolding pathway of purified rhDAO.(A) The sample concentration was 2.9 �M in 50 mM phosphate buffer, pH 7.2. The spectra were recorded between 180 and 260 nm with a time-per-point of 10.0 s at every1 nanometer step. The pathlength was 1 mm and the bandwidth was set to 3.0 nm. The spectra were recorded at 25 ◦C, 94 ◦C and again at 25 ◦C. (B) Temperature mediatedunfolding/refolding measured at 218 nm, the cell was heated from 25 to 94 ◦C and then cooled down to 25 ◦C in steps of 1 ◦C with 10 s time-per-point, a path length of 1 mmand a bandwidth of 3.0 nm. The resulting melting curve is depicted in black, the sigmoidal fit of the curve in grey and the refolding curve in dashed line.
Fig. 7. Substrate oxidation preference of rhDAO at pH 8.0.CHO-K1 supernatant containing 1.6 �g/mL rhDAO in 0.1% Human Serum Albumin PBS was mixed 1:1 with luminol solution and 1 �g/mL final concentration of horse radishperoxidase. 360 �M final substrate concentrations were used and the signal intensity measured at 30 ◦C for 2 h. The mean RLUs of duplicates and standard errors are shown.CON = Negative control without substrate; HIS = Histamine; CAD = Cadaverine; BENZ = Benzylamine; PUT = Putrescine; SPD = Spermidine; SPM = Spermine; RLU = Relative LightUnits.

128 E. Gludovacz et al. / Journal of Biotec
Fig. 8. Steady-state kinetics of 1 �g/mL purified rhDAO from Batch 2 usingputrescine as substrate.Using the DAOABA assay the mean putrescine oxidation rates of triplicates werecalculated using linear regression from the cumulative absorbance curves and cor-rected for wavelength. The R2 was at all concentrations except without addingany putrescine above 0.98. The oxidation rates were extrapolated to 1 mg purifiedrhDAO. The R2 of the fit is 99%. The calculated kcat and Km are 330 ± 11.7 min−1 and4Pm
(sta
(6lafvldr
3
maoeHEp
if8s2t
4
tots
2 ± 6.1 �M respectively. The curve was fitted using nonlinear regression in Graph-ad Prism. The values on the y-axis correspond to the specific enzyme activity perg rhDAO.
2002) and Schwelberger et al. (2013a), we found that the substratepecificity of rhDAO from CHO-K1 for diamines is consistent withhe previous reports. Although benzylamine is not a diamine, it islso oxidized.
Similar results were obtained performing the assay at pH 9.2see Fig. S2): putrescine – 100%, cadaverine – 94%, spermidine –4%, histamine – 40%, benzylamine – 38%, spermine – 28%. The
uminescence signal is strongly pH dependent. The signal strengtht pH 8.0, which is closer to the physiological pH of 7.4, was 7-old weaker, but the oxidation rates for the different substrates areery similar. The correlation coefficient is 80% between the two pHevels (data not shown). The differences might be explained by theifferent pKa values of the substrates and the different ionizationates at pH 8.0 versus 9.2.
.6. Steady-state kinetics
Kinetic parameters of purified rhDAO for putrescine were deter-ined using the DAOABA activity assay. Nonlinear regression
nalysis in GraphPad resulted in a Km of 42 ± 6,1 �M, a Vmax
f 2.0 �mol/min, and a kcat of 330 ± 11,7 min−1 (Fig. 8). Datavaluation using linear plots according to Lineweaver-Burke, Eadie-ofstee and Hanes-Wolf resulted in similar values (see Table S2).lmore et al. published a Km of 20 �M and a kcat of 475 min−1 forutrescine using insect cell derived DAO (Elmore et al., 2002).
Other kinetic studies of recombinant and native DAO from var-ous sources have been published and showed following Km valuesor putrescine: 26 �M (rhDAO, insect cells) (Juda et al., 2006),3 �M (human intestine) (Bieganski et al., 1983), 63 �M (humaneminal plasma) and 94 �M (human placenta) (Schwelberger et al.,013a). Our results are in good accordance with previous publica-ions.
. Conclusions
Since the first isolation of CHO cells in 1956, they have become
he most frequently used mammalian cell line in the productionf recombinant human biotherapeutics. Not only are they ableo rapidly grow to high cell densities in suspension cultures, toecrete proteins with high rates and to correctly fold them, buthnology 227 (2016) 120–130
most importantly to produce human-like posttranslational modi-fications (Jayapal et al., 2007). They are capable of adding complexglycans with terminal sialylation, while insect cells generate onlysimple glycosylation patterns with terminal mannose residues,which are known to potentially decrease the in vivo bioactivityand increase the allergenicity of proteins (Contreras-Gomez et al.,2014).
We generated a stable CHO-K1 cell line producing rhDAOwithin one month using recombinase-mediated cassette exchange.Classical cell line development procedures are time- and labor-consuming and can require more than six months. A three steppurification protocol was established that resulted in a final purityof >95% dimeric rhDAO.
At the beginning of each batch cultivation of the stable CHO-K1cell line 10 �M CuSO4 was added. ICP-MS analysis of purified rhDAOfrom two individual batches showed that 86.7% and 99.7% of theactive sites carried copper atoms. This result was substantiated byphenylhydrazine titration, which showed that 89.0% and 98.6% ofcopper-containing DAO molecules were occupied by an active TPQcofactor. This copper incorporation rates might be higher comparedto insect cell derived recombinant hDAO, which showed, includ-ing three batches, an average 0.95 mol per mol dimeric enzyme,implying that about 50% of the molecules in the dimer contain TPQ(Elmore et al., 2002). However, we cannot provide an obvious expla-nation, why CHO cells seem to be more efficient in incorporatingcopper into the active site of rhDAO.
Mass spectrometric analysis showed that the amino acidsequences of the N- and C-terminus of the purified enzyme wereas expected. Recombinant hDAO was not truncated during expres-sion and purification. We performed N-glycan analysis of rhDAO,which revealed that all four potential N-glycosylation sites wereindeed glycosylated. In X-ray crystallographic analysis of recom-binant DAO from insect cells only three glycosylated N-sites weredetected (McGrath et al., 2009). We found that three of them car-ried CHO typical complex 2–4 antennary glycans, most of themterminally sialylated, while one was oligomannosidic. Each sitecarried a unique pattern, as previously shown for other proteins(Go et al., 2013; Taschwer et al., 2012). Glycosylation of DAO doesnot seem to be important for enzymatic activity (Schwelberger andBodner, 1999). The influence of the different glycosylation sites onthe in vivo pharmacokinetic behavior is not known at the moment.The oligomannosidic site might be involved in the determinationof the half-life of DAO (Lee et al., 2002; Van Patten et al., 2007).
Electronic circular dichroism spectroscopy in the visible rangeresulted in a spectrum that indicated binding of copper and for-mation of the TPQ cofactor. It was consistent with the spectrumpresented by Elmore et al. (2002). The far UV spectrum showedthat random coils are the predominant secondary structure ele-ments of rhDAO. Our results are comparable with the results ofX-ray crystallographic analysis performed by McGrath et al. (2010a)and with the predictions of Raptor X software (Källberg et al., 2012).We confirmed that the purified enzyme is structurally intact.
Enzyme activity assays were used to determine substrate pref-erences and kinetic parameters of rhDAO. We found that our resultsfor the recombinant enzyme are consistent with previous publica-tions.
We were therefore able to obtain a source that enables us tocontinuously produce human diamine oxidase. The availability ofeasily accessible and active rhDAO enzyme allows us to addressresearch questions, which were until now not possible to inves-tigate. For example, recombinant human DAO could be used torapidly degrade histamine during anaphylactic episodes, where
histamine concentrations can reach several hundred nanogramsper milliliter (van der Linden et al., 1992). Nevertheless, neitherpharmacokinetic/pharmacodynamic nor toxicity studies in animals
Biotec
hi
A
BaaItdcGc
PlmScmSs
A
t0
R
A
B
B
B
B
B
C
C
C
C
C
C
D
E. Gludovacz et al. / Journal of
ave been published with recombinant human DAO and such stud-es might demand in total a few grams of purified active DAO.
cknowledgements
The study was initiated and supervised by Thomas Boehm,ernd Jilma and Nicole Borth. All experiments, except enzymectivity assays, LC–ESI-MS and ICP-MS, were performed by Elis-beth Gludovacz. LC–ESI-MS was conducted by Daniel Maresch,CP-MS by Maximilian Bonta and determination of kinetic parame-ers of rhDAO by Helen Szöllösi. Robert Weik provided his supporturing the establishment of the purification protocol. Electronic cir-ular dichroism and UV–vis spectroscopy were supervised by Paul. Furtmüller. The paper was written by Elisabeth Gludovacz withontributions from all authors.
We are grateful to Karin Petroczi (Department of Clinicalharmacology, Medical University of Vienna) for performing theuminescence-based activity assay and to Karin Kohlweiss (Depart-
ent of Biotechnology, University of Natural Resources and Lifeciences, Vienna) for performing FACSorting of DAO producing CHOells. Furthermore we thank Alexander Matlschweiger (Depart-ent of Biotechnology, University of Natural Resources and Life
ciences, Vienna) and Andrea Klima (Polymun Scientific) for theirupport during the establishment of the purification protocol.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at http://dx.doi.org/10.1016/j.jbiotec.2016.04.02.
eferences
kitsu, T., Yamaguchi, J., Uchida, N., Aritake, Y., 2009. The studies of conditions forinducing chirality to Cu(II) complexes by chiral Zn(II) and Ni(II) complexeswith schiff base. Res. Lett. Mater. Sci. 2009, 4.
aylin, S.B., Margolis, S., 1975. Purification of histaminase (diamine oxidase) fromhuman pregnancy plasma by affinity chromatography. Biochim. Biophys. Acta397, 294–306.
ecker, D.J., Lowe, J.B., 2003. Fucose: biosynthesis and biological function inmammals. Glycobiology 13, 41R–53R.
ieganski, T., Kusche, J., Lorenz, W., Hesterberg, R., Stahlknecht, C.D., Feussner, K.D.,1983. Distribution and properties of human intestinal diamine oxidase and itsrelevance for the histamine catabolism. Biochim. Biophys. Acta 756, 196–203.
ieganski, T., Kusche, J., Lorenz, W., Hesterberg, R., Stahlknecht, C.D., Feussner, K.D.,1983. Distribution and properties of human intestinal diamine oxidase and itsrelevance for the histamine catabolism. Biochim. Biophys. Acta 756, 196–203.
razeau, B.J., Johnson, B.J., Wilmot, C.M., 2004. Copper-containing amine oxidases:biogenesis and catalysis; a structural perspective. Arch. Biochem. Biophys. 428,22–31.
hassande, O., Renard, S., Barbry, P., Lazdunski, M., 1994. The human gene fordiamine oxidase, an amiloride binding protein. Molecular cloning, sequencing,and characterization of the promoter. J. Biol. Chem. 269, 14484–14489.
hen, Z., Schwartz, B., Williams, N.K., Li, R., Klinman, J.P., Mathews, F.S., 2000.Crystal structure at 2.5 A resolution of zinc-substituted copper amine oxidaseof Hansenula polymorpha expressed in Escherichia coli. Biochemistry 39,9709–9717.
ona, A., Tisi, A., Ghuge, S.A., Franchi, S., De Lorenzo, G., Angelini, R., 2014. Woundhealing response and xylem differentiation in tobacco plants over-expressing afungal endopolygalacturonase is mediated by copper amine oxidase activity.Plant Physiol. Biochem. 82, 54–65.
ontreras-Gomez, A., Sanchez-Miron, A., Garcia-Camacho, F., Molina-Grima, E.,Chisti, Y., 2014. Protein production using the baculovirus-insect cell expressionsystem. Biotechnol. Prog. 30, 1–18.
orda, M., Pellegrini, M., Rinaldi, A., 1984. Diamine oxidase from horse kidney:ionic strength dependence of stability and activity. Ital. J. Biochem. 33,303–310.
rabbe, M.J., Waight, R.D., Bardsley, W.G., Barker, R.W., Kelly, I.D., Knowles, P.F.,1976. Human placental diamine oxidase: improved purification and
characterization of a copper- and manganese-containing amine oxidase withnovel substrate specificity. Biochem. J 155, 679–687.ove, J.E., Smith, A.J., Kuchar, J., Brown, D.E., Dooley, D.M., Klinman, J.P., 1996.Identification of the quinone cofactor in a lysyl oxidase from Pichia pastoris.FEBS Lett. 398, 231–234.
hnology 227 (2016) 120–130 129
Elmore, B.O., Bollinger, J.A., Dooley, D.M., 2002. Human kidney diamine oxidase:heterologous expression, purification, and characterization. J. Biol. Inorg.Chem. 7, 565–579.
Ernberg, K., McGrath, A.P., Peat, T.S., Adams, T.E., Xiao, X., Pham, T., Newman, J.,McDonald, I.A., Collyer, C.A., Guss, J.M., 2010. A new crystal form of humanvascular adhesion protein 1. Acta Crystallogr. Sect. F 66, 1572–1578.
Finney, J., Moon, H.J., Ronnebaum, T., Lantz, M., Mure, M., 2014. Humancopper-dependent amine oxidases. Arch. Biochem. Biophys. 546, 19–32.
Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S., Wilkins, M.R., Appel, R.D.,Bairoch, A., 2005. Protein identification and analysis tools on the ExPASyserver. In: Walker, J.M. (Ed.), The Proteomics Protocol Handbook. HumanaPress, pp. 571––607.
Ghaderi, D., Zhang, M., Hurtado-Ziola, N., Varki, A., 2012. Production platforms forbiotherapeutic glycoproteins. Occurrence, impact, and challenges ofnon-human sialylation. Biotechnol. Genet. Eng. Rev. 28, 147–175.
Go, E.P., Liao, H.-X., Alam, S.M., Hua, D., Haynes, B.F., Desaire, H., 2013.Characterization of host-cell line specific glycosylation profiles of earlytransmitted/founder HIV-1 gp120 envelope proteins. J. Proteome Res. 12,1223–1234.
Green, M.R., Sambrook, J., 2012. Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
Hollister, J., Grabenhorst, E., Nimtz, M., Conradt, H., Jarvis, D.L., 2002. Engineeringthe protein N-glycosylation pathway in insect cells for production ofbiantennary, complex N-glycans. Biochemistry 41, 15093–15104.
Holmstedt, B., Tham, R., 1959. A spectrophotometric method for determination ofdiamine oxidase (DAO) activity. Acta Physiol. Scand. 45, 152–163.
Hossler, P., Khattak, S.F., Li, Z.J., 2009. Optimal and consistent protein glycosylationin mammalian cell culture. Glycobiology 19, 936–949.
Janes, S.M., Klinman, J.P., 1991. An investigation of bovine serum amine oxidaseactive site stoichiometry: evidence for an aminotransferase mechanisminvolving two carbonyl cofactors per enzyme dimer. Biochemistry 30,4599–4605.
Jayapal, K., Wlaschin, K., Hu, W.-S., Yap, M., 2007. Recombinant ProteinTherapeutics from Cho Cells – 20 Years and Counting. CHO Consortium: SBESpecial Edition, 40–47.
Juda, G.A., Shepard, E.M., Elmore, B.O., Dooley, D.M., 2006. A comparative study ofthe binding and inhibition of four copper-containing amine oxidases by azide:implications for the role of copper during the oxidative half-reaction.Biochemistry 45, 8788–8800.
Källberg, M., Wang, H., Wang, S., Peng, J., Wang, Z., Lu, H., Xu, J., 2012.Template-based protein structure modeling using the RaptorX web server. Nat.Protoc. 7, 1511–1522.
Kaitaniemi, S., Elovaara, H., Gron, K., Kidron, H., Liukkonen, J., Salminen, T., Salmi,M., Jalkanen, S., Elima, K., 2009. The unique substrate specificity of humanAOC2, a semicarbazide-sensitive amine oxidase. Cell. Mol. Life Sci. 66,2743–2757.
Klema, V.J., Solheid, C.J., Klinman, J.P., Wilmot, C.M., 2013. Structural analysis ofaliphatic versus aromatic substrate specificity in a copper amine oxidase fromHansenula polymorpha. Biochemistry 52, 2291–2301.
Kumar, V., Dooley, D.M., Freeman, H.C., Guss, J.M., Harvey, I., McGuirl, M.A., Wilce,M.C., Zubak, V.M., 1996. Crystal structure of a eukaryotic (pea seedling)copper-containing amine oxidase at 2.2 A resolution. Structure 4, 943–955.
Lee, S.J., Evers, S., Roeder, D., Parlow, A.F., Risteli, J., Risteli, L., Lee, Y.C., Feizi, T.,Langen, H., Nussenzweig, M.C., 2002. Mannose receptor-mediated regulationof serum glycoprotein homeostasis. Science 295, 1898–1901.
Li, R., Klinman, J.P., Mathews, F.S., 1998. Copper amine oxidase from Hansenulapolymorpha: the crystal structure determined at 2.4 A resolution reveals theactive conformation. Structure 6, 293–307.
Louis-Jeune, C., Andrade-Navarro, M.A., Perez-Iratxeta, C., 2012. Prediction ofprotein secondary structure from circular dichroism using theoreticallyderived spectra. Proteins 80, 374–381.
Maintz, L., Novak, N., 2007. Histamine and histamine intolerance. Am. J. Clin. Nutr.85, 1185–1196.
Maintz, L., Schwarzer, V., Bieber, T., van der Ven, K., Novak, N., 2008. Effects ofhistamine and diamine oxidase activities on pregnancy: a critical review. Hum.Reprod. Update 14, 485–495.
McGrath, A.P., Hilmer, K.M., Collyer, C.A., Shepard, E.M., Elmore, B.O., Brown, D.E.,Dooley, D.M., Guss, J.M., 2009. Structure and inhibition of human diamineoxidase. Biochemistry 48, 9810–9822.
McGrath, A.P., Caradoc-Davies, T., Collyer, C.A., Guss, J.M., 2010a. Correlation ofactive site metal content in human diamine oxidase withtrihydroxyphenylalanine quinone cofactor biogenesis. Biochemistry 49,8316–8324.
McGrath, A.P., Hilmer, K.M., Collyer, C.A., Dooley, D.M., Guss, J.M., 2010b. A newcrystal form of human diamine oxidase. Acta Crystallogr. Sect. F 66, 137–142.
Mizuguchi, H., Imamura, I., Takemura, M., Fukui, H., 1994. Purification andcharacterization of diamine oxidase (histaminase) from rat small intestine. J.Biochem. 116, 631–635.
Mure, M., Klinman, J.P., 1993. Synthesis and spectroscopic characterization ofmodel compounds for the active site cofactor in copper amine oxidases. J. Am.Chem. Soc. 115, 7117–7127.
Mure, M., Mills, S.A., Klinman, J.P., 2002. Catalytic mechanism of the topa quinonecontaining copper amine oxidases. Biochemistry 41, 9269–9278.
Parsons, M.R., Convery, M.A., Wilmot, C.M., Yadav, K.D., Blakeley, V., Corner, A.S.,Phillips, S.E., McPherson, M.J., Knowles, P.F., 1995. Crystal structure of a

1 Biotec
P
R
R
R
S
S
S
S
S
S
S
30 E. Gludovacz et al. / Journal of
quinoenzyme: copper amine oxidase of Escherichia coli at 2 A structure.Structure 3, 1171–1184.
ichler, J., Hesse, F., Wieser, M., Kunert, R., Galosy, S.S., Mott, J.E., Borth, N., 2009. Astudy on the temperature dependency and time course of the cold captureantibody secretion assay. J. Biotechnol. 141, 80–83.
aymond, C.S., Soriano, P., 2007. High-efficiency FLP and PhiC31 site-specificrecombination in mammalian cells. PLoS One 2, e162.
inaldi, A., Floris, G., Finazzi-Agro, A., 1982. Purification and properties of diamineoxidase from Euphorbia latex. Eur. J. Biochem. 127, 417–422.
inaldi, A.C., Rescigno, A., Rinaldi, A., Sanjust, E., 1999. Modeling novelquinocofactors: an overview. Bioorg. Chem. 27, 253–288.
almi, M., Jalkanen, S., 2005. Cell-surface enzymes in control of leukocytetrafficking. Nat. Rev. Immunol. 5, 760–771.
attler, J., Hesterberg, R., Lorenz, W., Schmidt, U., Crombach, M., Stahlknecht, C.D.,1985. Inhibition of human and canine diamine oxidase by drugs used in anintensive care unit: relevance for clinical side effects? Agents Actions 16,91–94.
chlake, T., Bode, J., 1994. Use of mutated FLP recognition target (FRT) sites for theexchange of expression cassettes at defined chromosomal loci. Biochemistry33, 12746–12751.
chwelberger, H.G., Bodner, E., 1999. Analysis of the glycosylation of porcinediamine oxidase. Inflamm. Res. 48 (Suppl. 1), S83–S84.
chwelberger, H.G., Feurle, J., 2007. Luminometric determination of amine oxidaseactivity. Inflamm. Res. 56 (Suppl. 1), S53–54.
chwelberger, H.G., Hittmair, A., Kohlwein, S.D., 1998. Analysis of tissue andsubcellular localization of mammalian diamine oxidase by confocal laserscanning fluorescence microscopy. Inflamm. Res. 47 (Suppl. 1), S60–61.
chwelberger, H.G., Feurle, J., Ahrens, F., 2013a. Characterization of diamineoxidase from human seminal plasma. J. Neural Transm. 120, 983–986.
hnology 227 (2016) 120–130
Schwelberger, H.G., Feurle, J., Houen, G., 2013b. New tools for studying oldquestions: antibodies for human diamine oxidase. J. Neural Transm. 120,1019–1026.
Tamura, H., Horiike, K., Fukuda, H., Watanabe, T., 1989. Kinetic studies on theinhibition mechanism of diamine oxidase from porcine kidney byaminoguanidine. J. Biochem. 105, 299–306.
Taschwer, M., Hackl, M., Hernandez Bort, J.A., Leitner, C., Kumar, N., Puc, U., Grass,J., Papst, M., Kunert, R., Altmann, F., Borth, N., 2012. Growth, productivity andprotein glycosylation in a CHO EpoFc producer cell line adapted toglutamine-free growth. J. Biotechnol. 157, 295–303.
Van Patten, S.M., Hughes, H., Huff, M.R., Piepenhagen, P.A., Waire, J., Qiu, H.,Ganesa, C., Reczek, D., Ward, P.V., Kutzko, J.P., Edmunds, T., 2007. Effect ofmannose chain length on targeting of glucocerebrosidase for enzymereplacement therapy of Gaucher disease. Glycobiology 17, 467–478.
Varki, A., 1993. Biological roles of oligosaccharides: all of the theories are correct.Glycobiology 3, 97–130.
Wilce, M.C., Dooley, D.M., Freeman, H.C., Guss, J.M., Matsunami, H., McIntire, W.S.,Ruggiero, C.E., Tanizawa, K., Yamaguchi, H., 1997. Crystal structures of thecopper-containing amine oxidase from Arthrobacter globiformis in the holo andapo forms: implications for the biogenesis of topaquinone. Biochemistry 36,16116–16133.
Wollin, A., Wang, X., Tso, P., 1998. Nutrients regulate diamine oxidase release fromintestinal mucosa. Am. J. Physiol. 275, R969–975.
Zeng, J., Spiro, S., 2013. Finely tuned regulation of the aromatic amine degradation
pathway in Escherichia coli. J. Bacteriol. 195, 5141–5150.van der Linden, P.W., Hack, C.E., Poortman, J., Vivie-Kipp, Y.C., Struyvenberg, A., vander Zwan, J.K., 1992. Insect-sting challenge in 138 patients: relation betweenclinical severity of anaphylaxis and mast cell activation. J. Allergy Clin.Immunol. 90, 110–118.