
Isozyme Characterization and Diversity Among the...
12
Philippine Journal of Science 135 (2): 93-104, December 2006 ISSN 0031 - 7683 93 Sancho G. Bon 1* , Teresita H. Borromeo 1 , Nestor C. Altoveros 2 and Avelino D. Raymundo 3 Key Words: wild rice, cluster analysis, zymogram, germplasm conservation *Corresponding author: [email protected] Isozyme Characterization and Diversity Among the Philippine Populations of Oryza officinalis Wall ex. Watt conserved ex situ 1 Crop Science Cluster, College of Agriculture University of the Philippines Los Baños, College, Laguna 2 Crop Science Cluster, College of Agriculture University of the Philippines Los Baños, College, Laguna 3 Crop Protection Cluster, College of Agriculture University of the Philippines Los Baños, College, Laguna INTRODUCTION It is well recognized that wild rice species are rich potential sources of resistance genes for various biotic and abiotic stresses (Jha et al. 1999). Some of the useful traits successfully introduced into the cultivated rice include: resistance to bacterial blight from Oryza longistaminata, Oryza officinalis, Oryza minuta, Oryza latifolia, Oryza Thirty-nine ex situ accessions of Philippine Oryza officinalis Wall ex Watt were characterized for isozyme electrophoretic profile and analyzed for diversity. Polymorphism was detected in phosphoglucoisomerase, arginine aminopeptidase, alanine aminopeptidase, esterase-9, and leucine aminopeptidase. Zymograms revealed the electrophoretic profiles of the populations across enzymes. Polymorphism was shown in the number of resulting bands, positions of bands, and in the general banding pattern. Zymogram patterns showed that the experimental populations can be grouped into two sets: seed collections and vegetative collections. Vegetative collections showed more variations relative to banding characteristics, hence, indicating comparative diversity. Seed collections exhibited very low variation in isozyme features. Cluster analyses confirmed separation of the two sets. Vegetative collections also showed some degree of geographic association. Reduced isozyme polymorphism indicated loss of allelic variation among the seed collections and could be ascribed to factors relating to collecting sample size, and procedural practices in handling germplasm for ex situ regeneration and conservation. On the other hand, technical instability of the isozyme system may have similarly contributed to the present results obtained. Further molecular investigation including studies on factors that influence genetic structure of populations under ex situ conservation are recommended to verify the results obtained. australiensis and Oryza brachyantha; blast from Oryza minuta; brown plant hopper from Oryza officinalis, Oryza minuta, Oryza latifolia, and Oryza australiensis; resistance to grassy stunt virus from Oryza nivara; and tolerance to acid sulfate soils from Oryza rufipogon (Borromeo 2000). Oryza longiglumis and Oryza meyeriana are potential gene sources for shade tolerance; Oryza meridionalis for drought tolerance and Oryza ridleyi for stem borer resistance (Borromeo 2000; Vaughan 1994).
Transcript of Isozyme Characterization and Diversity Among the...

Philippine Journal of Science135 (2): 93-104, December 2006ISSN 0031 - 7683
93
Sancho G. Bon1*, Teresita H. Borromeo1, Nestor C. Altoveros2 and Avelino D. Raymundo3
Key Words: wild rice, cluster analysis, zymogram, germplasm conservation
*Corresponding author: [email protected]
Isozyme Characterization and Diversity Amongthe Philippine Populations of Oryza officinalis
Wall ex. Watt conserved ex situ
1Crop Science Cluster, College of AgricultureUniversity of the Philippines Los Baños, College, Laguna
2Crop Science Cluster, College of AgricultureUniversity of the Philippines Los Baños, College, Laguna
3Crop Protection Cluster, College of AgricultureUniversity of the Philippines Los Baños, College, Laguna
INTRODUCTIONIt is well recognized that wild rice species are rich potential sources of resistance genes for various biotic and abiotic stresses (Jha et al. 1999). Some of the useful traits successfully introduced into the cultivated rice include: resistance to bacterial blight from Oryza longistaminata, Oryza officinalis, Oryza minuta, Oryza latifolia, Oryza
Thirty-nine ex situ accessions of Philippine Oryza officinalis Wall ex Watt were characterized for isozyme electrophoretic profile and analyzed for diversity. Polymorphism was detected in phosphoglucoisomerase, arginine aminopeptidase, alanine aminopeptidase, esterase-9, and leucine aminopeptidase. Zymograms revealed the electrophoretic profiles of the populations across enzymes. Polymorphism was shown in the number of resulting bands, positions of bands, and in the general banding pattern. Zymogram patterns showed that the experimental populations can be grouped into two sets: seed collections and vegetative collections. Vegetative collections showed more variations relative to banding characteristics, hence, indicating comparative diversity. Seed collections exhibited very low variation in isozyme features. Cluster analyses confirmed separation of the two sets. Vegetative collections also showed some degree of geographic association. Reduced isozyme polymorphism indicated loss of allelic variation among the seed collections and could be ascribed to factors relating to collecting sample size, and procedural practices in handling germplasm for ex situ regeneration and conservation. On the other hand, technical instability of the isozyme system may have similarly contributed to the present results obtained. Further molecular investigation including studies on factors that influence genetic structure of populations under ex situ conservation are recommended to verify the results obtained.
australiensis and Oryza brachyantha; blast from Oryza minuta; brown plant hopper from Oryza officinalis, Oryza minuta, Oryza latifolia, and Oryza australiensis; resistance to grassy stunt virus from Oryza nivara; and tolerance to acid sulfate soils from Oryza rufipogon (Borromeo 2000). Oryza longiglumis and Oryza meyeriana are potential gene sources for shade tolerance; Oryza meridionalis for drought tolerance and Oryza ridleyi for stem borer resistance (Borromeo 2000; Vaughan 1994).

94
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Of about 20 wild species under the genus Oryza, 4 can be found in the Philippines. These are O. meyeriana, O. rufipogon, O. minuta, and O. officinalis (Borromeo et al. 1994). Of the 4, O. officinalis has the widest geographic distribution. Early accounts reported its occurrence in the islands of Mindoro, Bohol, Samar, Leyte, Negros in central Philippines, and the central and western sections of Mindanao island (Tateoka & Pancho 1963). More recently, it was found in Victoria, Laguna (Evangelista et al. 2000) and in Kabankalan, Negros Occidental (Bon 2005). O. officinalis is normally found along or near water systems or at least in moist soils adjacent to cultivated rice (Vaughan 1994). O. officinalis is a potential gene source for resistance to several insect pests and for higher biomass production (Gao et al. 2001). Its resistance to white-backed plant hopper has been successfully introduced into the cultivated rice (Borromeo 2000). O. officinalis is a rhizomatous, hence, vegetatively propagating species.
Currently, there are 39 Philippine populations of O. officinalis being conserved as ex situ collection. These germplasm materials were assembled within the periods 1963-1991 and 1996-2000. The first set is maintained as seed accessions, and the second as live plant specimens. A morpho-agronomic study on the nature of genetic diversity of these materials consistently showed sub-grouping based on the length of period that materials have been under ex situ conservation, rather than their geographic origins. The grouping was shown across different sets of phenotypic markers (Bon et al. 2005). Hence, this particular study prompted a closer look at the genetic variability of the Philippine populations of O. officinalis at the biochemical level. Specifically, the objectives of the present study are:
– To characterize the isozyme electrophoretic profile of the 39 ex situ accessions of O. officinalis collected in the Philippines.
– To determine the distribution of variability in isozyme pattern among the 39 ex situ accessions of O. officinalis collected in the Philippines, and
– To compare isozyme variability with the known qualitative morpho-agronomic diversity, and determine their combined effect on the distribution of variation among the 39 ex situ accessions of O. officinalis found in the Philippines.
MATERIALS AND METHODS
Experimental PopulationsThirty-nine (39) Philippine populations of O. officinalis maintained ex situ were used in the study. O. officinalis found in the Philippines is known to be diploid with 2n=24 designated as CC. Thirty-two (32) accessions were obtained as seeds samples from the Genetic Resources Center (GRC) of the International Rice Research Institute (IRRI) and designated seed collections. The 7 populations were obtained from the University of the Philippines Los Baños – Philippine Rice Research Institute (UPLB-PhilRice) living plant collection and designated as vegetative collections. The vegetative collections represent populations that were recently found (1996-2000), while seed collections were populations collected mostly during the early 1960s and some in the early 1990s (1963 – 1991).
For live plant samples, a small clump of young tillers were sectioned from source pots and directly replanted into size 8 clay pots with fresh garden soil. For seed samples, seeds were first acclimatized to room temperature for 2 wks prior to dormancy breaking by oven dry heat for 2 wks at 54 °C, and germinated under sterile condition. Germinating seedlings were transferred to plastic trays with sterile garden soil. Trays were kept moist but not submerged for 2 wks inside the greenhouse, and then subjected to submerged condition for another 2 wks. Complete fertilizer was applied once while foliar fertilizer was applied at about 2-wk interval. About month-old seedlings (not tillering yet) were individually transplanted to size 8 clay pots.
Pots were placed under partial shade to partially simulate natural environment, and for easier maintenance and management of the set up. Commercial organic (coconut dust-based) fertilizer was applied about every 2 months. Populations were maintained under saturated condition by plugging the pot holes and maintaining water level. The plants were repeatedly ratooned prior to tissue collection to obtain uniform plant (tiller) age in both the seed-grown and vegetatively-grown plants. Each accession had 5 replicate pots (one seedling or tiller section per pot) arranged in a randomized complete block design.
Enzyme Electrophoresis Enzyme extraction and gel electrophoresis. Newly emerging shoots from ratooned tillers were collected from each pot (one shoot sample per replicate pot) to form the composite tissue samples from 5 replicate pots. The innermost youngest growing leaf was collected and cut into

95
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
small pieces for enzyme extraction. This is a modification from the use of plumules from young seedlings. On the spot plates, tissues were added with 2-3 drops of ice-cold 0.1%mercapto-ethanol: water solution and ground using a glass rod. Strips of filter paper (Whatman No. 3) used for enzyme electrophoresis were placed over the plates to absorb the crude extract, and lightly blotted dry and clean over sheet of tissue paper. Strips were then inserted vertically into a prepared starch gel slab. Two strips were inserted side-by-side per accession extract and subsequently subjected to electrophoresis. The starch gel electrophoresis protocol at the Molecular and Systematic Laboratory (GRC) based on Glaszman et al. (1988) was followed. No check genotype(s) or standard enzyme marker were available during the conduct of the study.
Enzyme Staining. Each cut gel slab was placed in plastic container with the particular enzyme stain solution. Eight available enzyme systems were used in the study: arginine aminopeptidase (ARAP), leucine aminopeptidase (LAP), phosphoglucose isomerase (PGI), esterase-9 (EST-9), alanine amonipeptidase (ALAP), alcohol dehydrogenase (ADH), malate dehydrogenase (MDH), and endopeptidase (ENP). The protocol of the Molecular and Systematics Laboratory of the GRC for enzyme stain composition and staining were followed. After staining, excess stain solution was washed off from the gel using distilled water. Gel slabs were mounted on a board and covered with thin plastic film, then dried in hot plate-vacuum gel drier (Biorad) for about 40 min. Dried gels were then scored and scanned for image presentation.
Scoring and equivalent descriptor-state determination. Considerations for enzyme scoring were clarity or band resolution and polymorphism. An isozyme is considered polymorphic when at least 1 accession showed a variant pattern. Pattern variation is based on the number and relative position of the bands. Intensity of banding, though significant in certain enzymes, was not considered. Positional consideration is based on the distinct visual
separation. Slight differences were not considered. Zymograms were diagrammatically interpreted for final scoring and analysis.
Scoring was based on individual band appearance along a particular position. Presence of the band is scored 1 and absence 0. Hence, accession with the most number of bands had the most score of 1s and the least, the most 0s.
Also, the resulting zymogram patterns (combination of bands e.g. 2-band bands is a pattern and a 3-band type is another) were arbitrarily considered as equivalent qualitative descriptor states (nominal type) of the character (isozymes). Each distinct pattern in an isozyme served as a type and correspondingly coded nominally similar to qualitative descriptor-state coding. Four types were considered for ARAP and EST-9 and three types for LAP, PGI, and ALAP. The data set was combined with the previously generated (Bon et al. 2005) 16 qualitative morpho-agronomic characterization data to form the combined 21 descriptors matrix.
Cluster AnalysesTwo sets of cluster analyses were performed. The first used only the isozyme data matrix based on the binary, 1 and 0 scoring. The second used the combined qualitative and isozyme data based on defined equivalent descriptors. Cluster analysis by sequential agglomerative hierarchical nested (SAHN) technique based on un-weighted pair group method, arithmetic average (UPGMA) were performed in Numerical Taxonomy and Multivariate Analysis System for pc, version 1.7 (NTSYS-pc v1.7) (Applied Biostatistics, Inc. 1992). Both analyses used the simple matching similarity coefficient. Dendrograms were generated to visualize results. To facilitate dendrogram interpretation, original accession numbers were re-coded based on place of collection (e.g. Accession IRGC 80073 was re-coded ‘zam1’ for Zamboanga, Accession IRGC 101142 was re-coded ‘boh’ for Bohol, etc., see Table 1).
Table 1. Code and collecting origin of the 39 experimental populations.
CODE ACCESSION NO. PROVINCE MUNICIPALITY
1 scot IRGC 80776 SOUTH COTABATO KORONADAL
2 zam1 IRGC 80773 ZAMBOANGA MORET
3 zds1 IRGC 80777 ZAMBOANGA DEL SUR MANIKAHAN
4 zds2 IRGC 80778 ZAMBOANGA DEL SUR MANIKAHAN
5 zds3 IRGC 80779 ZAMBOANGA DEL SUR MANIKAHAN
6 neg1 IRGC 100973 NEGROS OCCIDENTAL HINIGARAN
7 cot1 IRGC 101074 COTABATO TAMNAG
8 om1-i IRGC 101077 OCCIDENTAL MINDORO -
9 zam2 IRGC 101112 ZAMBOANGA STA. MARIAsee table continuation next page.....

96
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Philippine Journal of ScienceVol. 135 No. 2, December 2006
RESULTS AND DISCUSSION
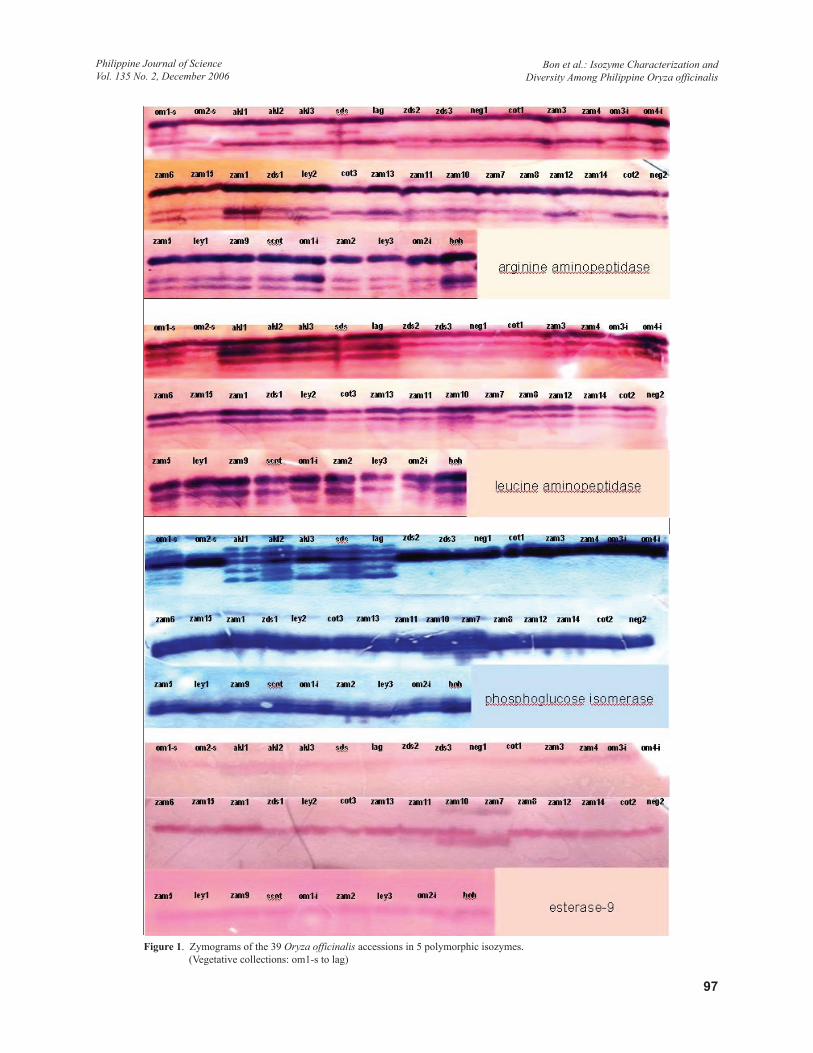
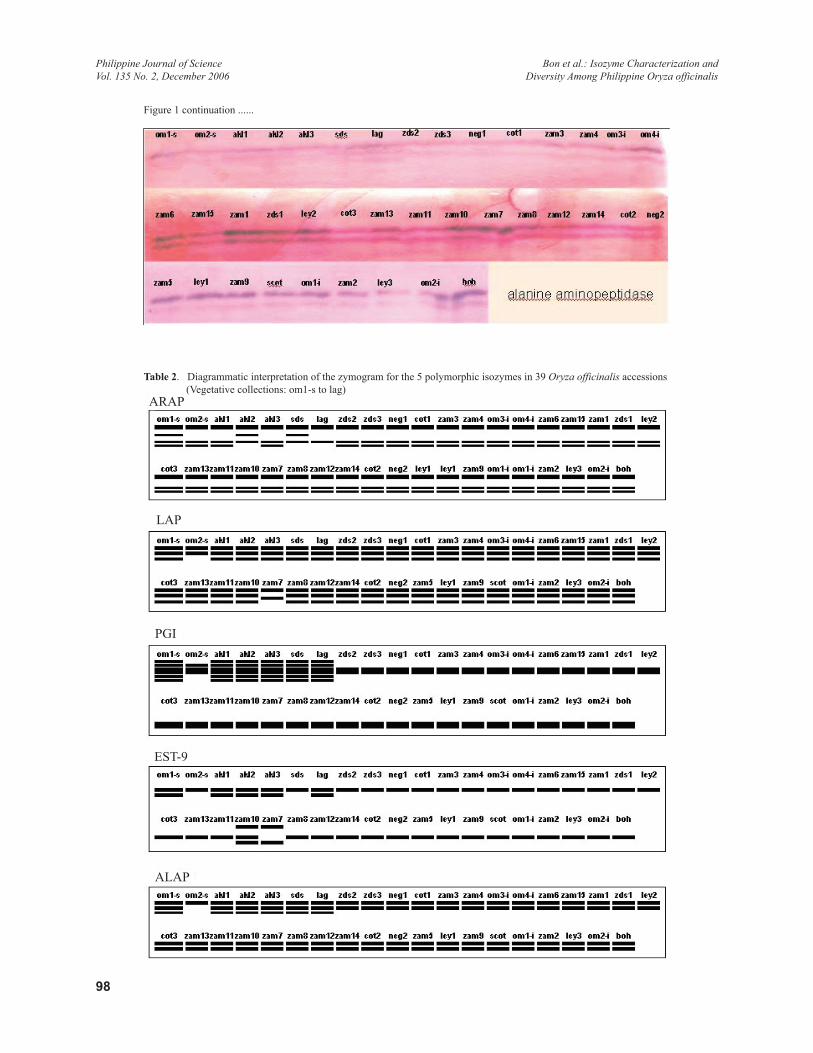
Isozyme CharacterizationFive enzyme systems out of 8 were found to produce clear polymorphic banding pattern namely, ARAP, LAP, PGI, EST-9, and ALAP. Except for EST-9 that showed generally a single relatively thin banding pattern, the 4 other resulting zymograms showed multiple banding patterns (Figure 1). Based on scoring of the diagrammatic interpretation of the zymograms, a total of 19 band positions were identified. Four bands were scored for ARAP and LAP, 3 for EST-9, and ALAP, and 5 for PGI (Table 2). In general, the vegetative collections produced more number of bands in all of the enzyme systems compared to the seed collections.
In ARAP, the general banding pattern was characterized by a 3-band zymogram with the topmost (first) showing the most intense staining and widely separated from the middle (third) band. The latter closely positioned with the lowest (fourth) band. As much as 5 alleles have been identified among the cultivated rice varieties (Kinoshita & Ishikawa 2002). Pattern variation among the populations was exhibited by an additional band (second) appearing between the first and the third bands, as shown by oms-1, akl2, and sds. On the other hand, lag had only the first and third bands. The rest of the accessions produced the characteristic 3-band pattern including om2-s, akl1, and akl3. Visually, vegetative collections showed a slightly lower third band compared to the same band in the seed collections. The observed variation was not considered for differential scoring. Also, the second band was observed
10 zam3 IRGC 101113 ZAMBOANGA TETUAN
11 zam4 IRGC 101115 ZAMBOANGA TALABAAN
12 zam5 IRGC 101116 ZAMBOANGA TALABAAN
13 om2-i IRGC 101121 OCCIDENTAL MINDORO -
14 ley1 IRGC 101139 LEYTE -
15 boh IRGC 101142 BOHOL LACAPAN SUR
16 zam6 IRGC 101166 ZAMBOANGA -
17 zam7 IRGC 105082 ZAMBOANGA -
18 zam8 IRGC 105083 ZAMBOANGA -
19 zam9 IRGC 105084 ZAMBOANGA -
20 zam10 IRGC 105085 ZAMBOANGA -
21 zam11 IRGC 105112 ZAMBOANGA STA. MARIA
22 zam12 IRGC 105113 ZAMBOANGA TETUAN
23 zam13 IRGC 105114 ZAMBOANGA -
24 zam14 IRGC 105115 ZAMBOANGA TALABAAN
25 cot2 IRGC105116 COTABATO TALIAN
26 cot3 IRGC 105117 COTABATO TAMNAG
27 neg2 IRGC 105118 NEGROS OCCIDENTAL CANDUMARAO
28 om3-i IRGC 105119 OCCIDENTAL MINDORO -
29 om4-i IRGC 105120 OCCIDENTAL MINDORO -
30 ley2 IRGC 105121 LEYTE -
31 zam15 IRGC 105175 ZAMBOANGA -
32 ley3 IRGC 106480 LEYTE TANAUAN
33 om1-s SDC-1 OCCIDENTAL MINDORO CALINTAAN
34 om2-s SDC-3 OCCIDENTAL MINDORO SABLAYAN
35 akl1 SDC-4 AKLAN LIBACAO
36 akl2 SDC-5 AKLAN LIBACAO
37 akl3 SDC-6 AKLAN KALIBO
38 sds SDC-8 SURIGAO DEL SUR TAGO
39 lag SDC-15 LAGUNA VICTORIA
Table 1 continuation.....
97
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Figure 1. Zymograms of the 39 Oryza officinalis accessions in 5 polymorphic isozymes. (Vegetative collections: om1-s to lag)
98
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Table 2. Diagrammatic interpretation of the zymogram for the 5 polymorphic isozymes in 39 Oryza officinalis accessions (Vegetative collections: om1-s to lag)
Figure 1 continuation ......
ARAP
LAP
ALAP
PGI
EST-9

99
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
only in the vegetative collections, and none among the seed collections. The seed collections produced the 3-band pattern with the fourth band appearing quite faint in cot3, zam7, and zam14.
In LAP, 4 bands were identified with clear resolution among the test populations. The profile is a 3-band pattern similar to ARAP. All 3 bands however appeared close to each other, with the 2 upper bands showing more staining intensity. The pattern was similar in most of the populations except in om2-s, and zam7, with the former lacks the lowest (3rd) band and the latter lacks both the second and third bands, but produced a band just between the second and third band positions. Also, om2-s had a fainter second band. The observed polymorphism in the isozyme was generally limited with only 2 populations showing variation from the basic pattern observed. A total of 5 alleles were reported in the rice varieties for this isozyme (Kinoshita & Ishikawa 2003).
In PGI, 2 banding types were observed: single thick darkly-stained band and multiple thin bands. The latter is composed of 5 bands. Thick darkly-stained band type was observed in all of the seed collections and in 1 of the vegetative populations, om2-s. In addition, om-2-s had an additional faint band on top of the thick band. Pattern variation was exhibited between the seed collections and vegetatively collected populations. The 5-band pattern was observed in 6 of the 7 vegetative populations except in om2-s. Based on the band pattern appearance, the vegetative collections showed some degree of allelic diversity compared to the seed collections. PGI is coded for by 2 genes with compatible products. Pgi1 has 4 alleles and Pgi2 has 5. Banding pattern can be as high as 10 bands for double heterozygotes (Kinoshita & Ishikawa 2003)
For EST-9, the general pattern was a single band that migrated towards the cathodal end, a similar migration pattern reported for O. sativa (Glaszmann et al. 1988). The band is comparatively well-stained. There were 3 variations observed. One type is the presence of a faint second band below. This pattern was observed in the 5 of the vegetative collections, such as om1-s, akl1, akl2, akl3, and sds. No similar pattern was observed among the seed collections. The second variation in the pattern was a 3-band type as produced by zam10, a seed collection. It included upper and lower bands. The third variation is the absence of the main (center) band but presence of both the lower and upper bands as shown by zam7. In relation to the dominant pattern types, the test populations can be grouped as the seed collections and the vegetative collections. Thirty of the 32 seed collections produced the single cathodal band type with the exception of the above-mentioned 2 accessions. On the other hand, majority of the
vegetative collections had the 2-band pattern, not observed among the seed collections.
For ALAP, a total of 3 bands were considered for scoring. The pattern is characterized by 2 prominent main bands appearing close to each other. The upper band stained comparatively darker. The third band appears below the second (middle) band. It is much fainter compared to the other 2. Its presence is limited to the vegetative collections only. It was scored in 6 of the populations except om2-s, the same accession that was scored differently in the other 4 isozymes. On the other hand, the seed collections showed 1 type of banding pattern, the 2-band type. No variation was observed among them. In this isozyme, the populations can be similarly grouped into 2 classes: the 3-band and the 2-band accessions. The former is composed of the vegetative collections except om-2s. The second group is composed of all the seed collections. Clearly, vegetative populations can be readily distinguished from the seed, with the former exhibiting a distinct pattern.
Practically in all of the 5 enzyme systems, differences in pattern distinguished the living plant collection from the seed collections. The two accession groups can be differentiated in relation to the period of collection. The latter were assembled between early 1960’s to 1991, and the former were collected from 1996 to 2000. In general, the vegetative accessions showed more number of bands compared to the seed accessions in all of the polymorphic enzymes systems. This is particularly notable in PGI, ARAP, EST-9, and ALAP. Such high number of bands appearing in the vegetative accessions indicates comparatively more alleles. The vegetative collections could also suggest possible composite or heterogeneous nature of the populations. Possibly, originally collected clusters where vegetative samples were obtained could have grown from different source plants. On the other hand, natural crossing among in situ populations may also be a factor in the heterogeneity observed among the vegetative populations. In wild rice, Breese (1989) cited as much as 50% outcrossing rate. Plant samples of the vegetative collections used in the study were still part of the original vegetative samples collected from the sites. In the case of om2-s, it was collected from a solitary hill with no other clumps growing in the immediate surrounding. Collecting account suggests that the small cluster could have germinated from seeds deposited from elsewhere through water run-offs. The population was found over an un-shaded dry irrigation ditch heavily silted with run-off sand, a condition where O. officinalis is not likely to be found naturally. Though the aims and strategy of field sampling have been suggested (Brown 1978) in few cases such as this, sampling is but limited to the immediately available plants. Further, Breese (1989) stressed that

100
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Philippine Journal of ScienceVol. 135 No. 2, December 2006
bottlenecks is effectively created by collecting small samples. In such situations, expanded exploration of the site could be relevant to locate other populations.
On the other hand, Brown (1978) noted that cultivated forms are markedly depauperate of alleles than the wild populations, and domestication encourages selfing resulting to reduction in diversity. This situation could be simulated in handling wild germplasm for ex situ conservation. To begin with, the genetic identity of each accession is defined by the nature of the original materials received at the genebank. In this study, many of the seed accessions were initially regenerated from the original 10-seed per accessions sent by the Japanese collectors (de Guzman personal communication). Detailed information on the nature of seeds samples received is not presently available. If these wild germplasm accessions were found in the Philippines and repatriated back to GRC from the Japanese collecting institution, then it is highly probable that they have already undergone seed regeneration cycles in Japan. During subsequent regenerations, differential germination and seedling environment may have further reduced the number of base populations for seed recovery and multiplication, resulting to limited number of plants being maintained. Moreover, the controlled regeneration environment inside screenhouse coupled with bagging of panicles or the whole plant significantly reduce wind movement within the plant’s immediate environment. Consequently, selfing may have been unconsciously encouraged and chances of outcrossing and/or pollen contamination were reduced. It has been concluded that restricted geneflow, among others, enhances inbreeding and could lead to genetic depletion (Kinoshita & Ishikawa 2002). O. officinalis, which generally grows in open areas have a generally high estimate of outcrossing rate at 31.6% (Vaughan et al. 2003). On the other hand, seed processing for storage may contribute to further changes in the structure of the resultant populations. Unconsciously, directed selection may have been favored during manual seed cleaning that selects high-seed set and well-developed grains resulting to near uniform seed type lots. Such practices may well serve as convergent selection pressure across germplasm accessions. To some extent, some of these factors were employed in handling wild germplasm accessions used in this study. The handling procedure, however, was carried out due to logistical management and environmental premises (Almazan personal communication). The extent to which each factor contributed to the results obtained in this study is difficult to quantify in the absence of complete and specific information relating to the nature of the original populations received, regeneration/processing records, such as number of regeneration cycles, number of source-plants maintained, regeneration batch composition,
and lay-out, and pollination controls. But as a net effect, Bon et al. (2005) noted that the seed collections have significantly higher mean fertility rate, heavier 10-grain weight, wider grain width and thicker grains compared to the vegetative collections. These could be related to selfing and/or selection effects.
Given the same procedure followed for all the 39 test populations, it is clear that seed accessions lack significant polymorphism across the 5 enzyme systems. In addition to the probable cause discussed above, factors such as isozyme and procedure including choice of tissue type and age, may have influenced the zymogram pattern with respect to the seed collections. Minute variation in the procedures could result to significant differences in mobility patterns.(Glaszmann et al. 1988). Zymograms may also vary according to growth stages and type of tissue, enzyme system, and the loci encoding for the enzyme. For instance, alcohol dehydrogenase is not expressed in plumules, a common source for enzyme extraction in rice when grown aerobically, whereas aminopeptidase-2 has stable expression within the plant and the calli, except in mature leaf (Kinoshita & Ishikawa 2003). It may also be noted that although the tissues harvested were of approximate age relative to emergence from the source-plants, the original source of propagating materials were different, i.e. seed and vegetative tissues. In the common Asian wild rice, the asexually propagating type has high intra-population variability, while the seed propagating type has high fixation index within populations (Kinoshita & Ishikawa 2002). Enzyme extraction in the present study was based on composite sample, hence, the vegetative collections indicate a more heterogeneous structure than the seed collections. While pattern differences in zymograms were obtained, the present study was limited by the unavailability of the check genotypes for the isozymes. Genetic constitutions of the populations used were unknown. Results largely showed the isozyme electrophoretic diversity observed among the 39 accessions. Further investigation at the molecular level is recommended to validate observations in the present study.
Cluster AnalysisFigure 2 shows the clustering pattern of the 39 populations based on zymogram band scoring. As expected 2 groups were formed, the seed accessions and the vegetative collections at about 0.74 similarity. Om2-s and zam7 were outliers. The presence of all 4 bands in ARAP differentiated om1-s from the other 3 populations in its subgroup. Cluster IB separated from IA, with the latter lacking the 4th band in ARAP and the presence of a second band. An intra-population variability is also apparent with

101
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Figure 2. Dendogram of the cluster analysis of the 39 Oryza officinalis accessions based on band presence scoring
Coefficient0.60 0.70 0.80 0.90 1.00
om1-sMW
om1-s akl1 akl3 lag akl2 sds om2-s zds2 zds3 boh ley3 zam2 om1-i scot zam9 ley1 zam5 neg2 cot2 zam14 zam12 zam8 om2-i neg1 zam11 zam13 cot3 ley2 zds1 zam15 zam1 zam6 om4-i om3-i zam4 zam3 cot1 zam10 zam7
IA
IBI
II
the separation of akl2 from akl1 and akl3 with respect to ARAP banding pattern. All Akl populations were collected from different sites within the province of Aklan. Variation among the 3 populations was earlier noted based on qualitative and quantitative agronomic characters (Bon et al. 2005).
On the other hand, Group II has the same band scoring, hence, clustering at maximum similarity. Zam10 was differentiated from the main group on EST-9 and zam7 on both EST-9 and LAP. Between the 2 main groups, PGI is a significant differentiating isozyme with the seed collections having only 1 band present out of 5. Differences between the 2 groups were likewise exhibited in ALAP
and EST-9. But the 2 groups shared essentially the same scoring in LAP and ARAP. Most of the vegetative collections were scored positively in 17 of the 19 bands, while most of the seed collections were positively scored only on 10 bands. The relative number of bands within the 2 clusters could indicate the nature of allelic diversity among the conserved populations. Vegetative collections showed more alleles compared to the seed accessions. The result obtained in the present study was consistent with an earlier report by Bon et al. (2005) on the same set of materials using morpho-agronomic characters.
Figure 3 shows the dendrogram for combined isozyme banding type scoring and secondary qualitative

102
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Figure 3. Dendogram of the cluster analysis of the 39 Oryza officinalis accessions based on combined data matrix
Coefficient0.60 0.70 0.80 0.90 1.00
om1-iMW
zam1 zds3 zam14 zam13 zam8 cot2 neg1 cot1 neg2 om3-i scot zds1 zds2 zam5 cot3 om1-i om2-i ley2 ley1 zam6 ley3 zam9 zam2 zam3 zam15 zam11 om4-i zam4 zam10 zam12 boh zam7 om2-s om1-s akl2 akl1 akl3 sds lag
II
I
IB
IA
characterization data. The populations can be grouped into 2 (I, seed collections and II, vegetative collections) at 0.66 similarity with om2-s as an outlier. Between the 2 main clusters, Group I showed a very similar clustering pattern earlier reported using qualitative morpho-agronomic characters alone (Bon et al. 2005). Zam7 and boh remained outliers from the group. Most of the seed collections can be subdivided into 2 subgroups, IA and IB at about 0.80 similarity. Sub-groupings were a mixture of accessions collected from different geographic locations. The distance of the outliers, boh and zam7 from Group I was due to the combined effects of variation in isozyme pattern and qualitative phenotypic markers. The rest of accessions in the cluster did not show distinctly clear
variation in the isozyme pattern. This could be due to the strict interpretation and scoring of the isozyme pattern. Very slight variation in band position and relative band height (thickness), though visually detectable, were not considered for differential scoring and diagrammatic interpretation. Conversely, enzymes used may not be effective in detecting minute variation within the given cluster of accessions. Stebbins (1990) noted that enzymes could differ in a number of ways other than electrophoretic mobility. Survey of more polymorphic enzymes or further refinement in the procedure such as tissue and growth stage optimization, could improve zymogram resolution and validate results herein obtained. However, molecular analysis could provide a better clarification between the

103
Philippine Journal of ScienceVol. 135 No. 2, December 2006
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
issues of technical isozyme protocol and the inherent genetic background of the populations studied relative to the reduced isozyme variability among the seed collections.
Group II was composed of the vegetative collections, except om2-s. The populations can be separated into 3 approximate geographic areas. IIA is composed of Aklan and Occidental Mindoro populations (om1-s, akl12, akl1 and akl3). The 2 island provinces lie approximately on the same general geographic location (west central Philippines) but classified as different bio-geographic zones (DENR-UNEP 1997). IIB (sds) represents the southern Philippines (Mindanao zone). IIC (lag) represents northern Philippines (north-south Luzon zone). Though geographic association is apparent, it is currently less conclusive given the limited number of available samples. However, geographic association of diversity was earlier noted among populations of O. officinalis collected in different locations in Malaysia based on RAPDs (Vaughan et al. 1998). Vaughan et al. (2003) further noted that Asian populations of O. officinalis also exhibit greater differences between regions. A similar observation was also reported in O. granulata populations (Qian et al. 2001).
Among the vegetative collections, all 5 isozymes were polymorphic in at least 1 of the population. IIA had the same band type in 4 except ARAP. Three types were observed with Aklan populations having 2 different band types. Om1-s was similarly distinct from the Aklan populations. IIB was differentiated from IIA on ARAP and EST-9, while IIC differ in ARAP and PGI. Except for om2-s LAP, and ALAP would be least sensitive in differentiating the populations. Results thus indicate that observed phenotypic variation in Group II had a similar parallel pattern of variation in the isozymes as exemplified by om2-s and lag. Absence of phenotypic variation can be detected through isozymes as in the case of sds. Results also indicate presence of intra-population diversity among the local populations of O. officinalis as shown by Aklan and Occidental Mindoro populations. Intra-population variability in wild Oryza is generally reported as low (Quian et al. 2001; Vaughan et al. 1998; Song et al. 1999). Present observation therefore may require sufficient sampling from their natural habitats.
SUMMARY AND CONCLUSIONThe 39 Oryza officinalis germplasm accessions produced polymorphic electrophoretic profiles in 5 of the 8 enzyme systems tested. Polymorphism is indicated by variation in the general pattern, number of bands, and band position.
A notable observed variation is a positive increase in the number of bands, which may correspond to allelic diversity. Diagrammatic interpretation of the zymograms yields 5 clear bands in PGI followed by ARAP and LAP with 4. ALAP and EST-9 have both 3 distinct positions scored. ARAP is characterized by a 3-band pattern with 4 banding types identified. Similarly, LAP showed a 3-band pattern varying in position and intensity. Three banding types were considered. PGI showed 2 general patterns, single thick darkly-stained band, and multiple thin bands. Three banding types were identified. EST-9 is characterized generally by a cathodal single or 2-bands types. Four banding types were identified. Polymorphism is shown by the presence of a lower band appearing very close (relative to the center band), absence of the main center band, and presence/absence of band and their relative positions. Lastly, ALAP showed 2 prominent main band appearing close to each other with variation shown by the appearance of a third faint band. Three banding types were identified.
Cluster analyses confirmed the visual distinction of the 2 germplasm sets. Vegetative accessions formed a distinct group separate from the seed collections. The lone vegetative collection with low electrophoretic count clustered along with the seed accessions. Among the vegetative collections, analysis of combined data indicated some degree of geographic association of variation and potential intra-population variability. Also, phenotypic variability among the vegetative collections had a similar parallel of polymorphism in the isozymes. The observations, however, were not shared among the seed collections.
By and large, the 39 wild rice accessions produced a range of electrophoretic profiles in 5 sensitive enzymes systems. Isozyme profiles vary in number of bands, band position and intensity. Based on these pattern features, the 39 accessions can be grouped according to the source of experimental populations i.e. seed and vegetative accessions. Seed populations were generally depauperate of isozyme bands while vegetative populations produce generally more number of bands. Observations and results obtained in the present study may be ascribed to 2 main factors. One relates to the route of wild germplasm accession under ex situ conservation. This includes sample size and genetic structure of the original base population collected/received; differential seed germination and seedling survival during subsequent regeneration cycles; regeneration environment controls and practices including population size maintained; and seed processing and storage practices serving as selection pressures. Their accumulated combined effects may lead to changes in the genetic structure of the resulting populations. In

104
Bon et al.: Isozyme Characterization and Diversity Among Philippine Oryza officinalis
Philippine Journal of ScienceVol. 135 No. 2, December 2006
the present study, results suggest a possible convergent homogenization effects leading to loss of allelic diversity among the seed populations. The second factor, on the other hand, relates to the inherent technical instability of isozyme systems. Electrophoretic profiles may be influenced by a number of factors relating to plant tissue, enzyme species, and procedure. Given the nature of the genetic structure of the seed populations, the enzymes systems used were generally insensitive in detecting variability among them.
In view of the results obtained, further investigation at the molecular level is recommended to verify present observations. It should also include studies on the population structure of both germplasm accession sets; tracing the specific route of germplasm flow under ex situ conservation, and possibly estimate/quantify the potential genetic effects of the factors involved vis a vis the goals of ex situ conservation.
REFERENCESAPPLIED BIOSTATISTICS, INC. 1992. NTSYS-pc ver.
1.7. Applied Biostatistics, Inc., New York.
BON SG, BORROMEO TH, ALTOVEROS NC, BRAR DS and RAYMUNDO AD. 2005. Morpho-agronomic diversity of thirty-nine Philippine populations of Oryza officinalis Wall ex. Watt. Philipp Agric Scientist 88 (3): 281-291.
BON SG. 2005. Rice Germplasm Conservation Project-UPLB. Collecting Trip Report: Negros Occidental, April, 2005
BORROMEO TH. 2000. Philippine wild rices: diverse and disappearing. Nueva Ecija, Philippines: Philipp Agric Scientist 83: 133-144.
BREESE EL. 1998. Regeneration and multiplication of germplasm resources in seed genebanks: the scientific background. Rome: International Board for Plant Genetic Resources [IBPGR] 69p.
BROWN AHD. 1978. Isozymes, plant populations genetic structure ands genetic conservation. Theor Appl Genet 52: 145- 157.
[DENR] Department of Environment and Natural Resources – [UNEP] United Nations Environment Program. 1997. Philippine Biodiversity: An Assessment and Action Plan. Makati City, Philippines: Bookmark, Inc. 298p.
ELLSTRAND NC and ELAM DR. 1993. Population genetic consequences of small population size:
Implications for plant conservation. Annu Rev Ecol Syst 24: 217-242.
EVANGELISTA EM, BORROMEO TH and BON SG. 2000. Rice Germplasm Conservation Project-UPLB. Collecting Trip Report: Masapang, Victoria, Laguna.
GAO LZ, GE S and HONG DY. 2001. High levels genetic differentiation of Oryza officinalis Wall. ex Watt. from China. The J Heredity 92: 511-516.
GLASZMAN JC, DE LOS REYES BG and KHUSH GS. 1988. Electrophoretic variation of isozymes in plumules of rice (Oryza sativa L.) – a key to the identification of 76 alleles at 24 loci. IRRI Research Paper Series No. 143. Manila: International Rice Research Institute (IRRI). 14pp.
JHA SK, AWASTHI LP and MAURYA DM. 1999. Genetic diversity in wild rice germplasm of eastern U.P., India. Oryza. 36: 157-158.
QIAN W, GE S and HONG D. 2001. Genetic variation within and among populations of a wild rice Oryza granulata from China detected by RAPD and ISSR markers. Theor Appl Genet 102: 440-449.
SONG GE, GIANCARLO C, OLIVEIRA X, SCHAAL B, GAO L and HONG D. 1999a. RAPD variation within and between natural populations of the wild rice Oryza rufipogon from China and Brazil. Heredity 82: 638 – 644.
STEBBINS GL. 1990. Introduction. In: SOLTIS, DE and SOLTIS, PS. (eds). Isozymes in Plant Biology. London: Chapman and Hall. p. 1-4.
TATEOKA T and PANCHO JV. 1963. Report on exploration for wild species of Oryza in the Philippines.
VAUGHAN DA, TAMOOKA N, KOBAYASHI N and SARI AO. 1998. In situ conservation of plant genetic resources: characterization and evaluation. In: Plant Genetic Resources: Characterization and Evaluation. Tsukuba, Japan: National Institute of Agrobiological Resources. 256p.
VAUGHAN DA. 1994. The Wild Relatives of Rice: A Genetic Resources Handbook. Manila: International Rice Research Institute (IRRI). 96p.


















