
Intraspecific usurpation of incipient fire ant colonies - Behavioral
5
Behavioral Ecology Vol. 8 No. 1: 99-103 Intraspecific usurpation of incipient fire ant colonies Michael T. Balas and Eldridge S. Adams Department of Biology, University of Rochester, Rochester, NY 14627, USA Brood raiding, or reciprocal stealing of brood, is common among incipient colonies of the Ere ant SoUnopsis invicta. Paradox- ically, workers from a colony that loses its brood during a raid often abandon their nest and join the winning colony. Queens abandoned by their workers may then migrate from their original nest site and attempt to forcefully usurp another incipient colony by displacing that colony's queen or queens. This study examined factors that influence the success of usurpation attempts. Queens attempting to usurp a nearby colony after laboratory brood raids were successful in less than 30% of trials. Usurpation attempts were more successful if workers familiar to the migrating queen were present in the nest, as would happen if a queen were to find the colony to which her workers had moved. Cross-fostering experiments showed that this effect was due to familiarity rather than relatedness. Usurpation attempts were less successful if they were delayed by 16 h. The probability of usurpation success was not reduced by doubling the number of defending workers in the invaded colony. However, colonies founded by three queens were almost always able to resist usurpation attempts. These results support the hypothesis that workers abandon their natal colony after losing a brood raid to increase the likelihood that their queen can usurp the colony to which they migrate. These results also provide the first evidence that colonies of ants founded by several queens are better able to resist usurpation attempts than colonies founded by a single queen. Key words brood raids, fire ants, kin recognition, SoUnopsis invicta, usurpation. [Bthav Ecol 8:99-103 (1997)1 I ntraspecific competition among incipient colonies of social insects is often intense, resulting in high mortality of queens before the colonies reach reproductive maturity (H61- ldobler and Wilson, 1990). In some ant species, much of this mortality is associated with brood raiding. During brood raids, incipient colony workers remove eggs, larvae, and pupae from neighboring colonies and take them to their home colony, thereby supplementing the worker force (Bartz and Holldob- ler, 1982; Rissing and Pollock, 1987; Stamps and Vinson, 1991; Tschinkel, 1992a,b). Brood raids by the fire ant SoUnopsis in- victa (Buren) cause a substantial reduction in the density of incipient colonies and queens (Adams and Tschinkel, 1995a). Queens whose colonies have been depleted of brood through these raids often attempt to usurp nearby colonies whose brood population has been increased through raiding (Adams and Tschinkel 1995a,b; Tschinkel, 1992b). Success in brood raids rises with the size of the worker pop- ulation (Rissing and Pollock, 1991; Stamps and Vinson, 1991; Tschinkel, 1992b). One way a colony can produce more work- ers in its first brood is through pleometrosis, or colony foun- dation by several queens (reviewed in Rissing and Pollock, 1988). In three species for which raiding has been described, Mymucocystus mimicus (Bartz and H&lldobler, 1982), Mtssor prrganda (Rissing and Pollock, 1987, 1991), and £ invicta (Adams and Tschinkel, 1995b; Tschinkel, 1992b), pleometrot- ic colonies are more likely to win brood raids. Tschinkel (1992b) demonstrated that this effect was due to increased worker number and not increased queen number. A peculiar behavior associated with brood raiding is the abandonment of an incipient colony by its workers after its brood has been removed. When abandonment occurs, work- ers leave their colony's queen or queens at the original nest Address correspondence to E. S. Adams. M. T. -Balai u now at St. John'i University, 300 Howard Ave., Staten Island, NY 10301, USA. Received 31 August 1995; revised 27 March 1996; accepted 29 March 1996. 1045-2249/97/15.00 O 1997 International Society for Behavioral Ecology and peacefully join the colony that successfully raided it (Bartz and HoUdobler, 1982; Rissing and Pollock. 1987; Tschinkel, 1992b). However, a queen from a losing colony will usually attempt usurpation shortly after her workers abandon her (Adams and Tschinkel, 1995a,b; Bartz and HoUdobler, 1982; Rissing and Pollock, 1987; Tschinkel, 1992b). It has been pro- posed that workers' relocation is adaptive if the workers' mother queen has a high probability of joining them in the new colony, a process termed "hopeful co-emigration" (Pol- lock and Rissing, 1989). Queens attempting to enter other incipient colonies may encounter resistance. Workers of fire ant incipient colonies recognize familiar queens and often expel or kill unfamiliar queens (Balas and Adams, 1996b). However, migrating queens occasionally succeed in displacing the resident queen or queens, which may be killed and discarded (Adams and Tschinkel, 1995a,b; Rissing and Pollock, 1987; Tschinkel, 1992a). In addition, Adams and Tschinkel (1995b) showed that if a queen attempts to usurp a colony that has won a raid, her probability of success is greater if the workers that joined the colony are her daughters. However, it is not known wheth- er this effect is due to the presence of familiar workers or simply to the smaller number of defending workers in die invaded colony. This study examines whether workers of S. invicta are able to assist their mothers in colony usurpations. First, we tested whether workers of S. invicta are better able to usurp colonies to which their daughters have moved. Second, we tested whether the effect of daughters on usurpation success is due to familiarity with the queen or to genetic relatedness. Ag- gressive behavior during these usurpation attempts was ob- served to determine if the presence of familiar or related workers influenced the aggressive response toward either the resident queen or the queen attempting usurpation. In addition, preliminary experiments suggested that a queen's probability of usurping the colony to which her daughters have moved may decline if she does not migrate immediately after a brood raid. This may be because queens rapidly lose familiarity to their former nestmates. We tested Downloaded from https://academic.oup.com/beheco/article/8/1/99/211854 by guest on 16 January 2022
Transcript of Intraspecific usurpation of incipient fire ant colonies - Behavioral

Behavioral Ecology Vol. 8 No. 1: 99-103
Intraspecific usurpation of incipient fire antcolonies
Michael T. Balas and Eldridge S. AdamsDepartment of Biology, University of Rochester, Rochester, NY 14627, USA
Brood raiding, or reciprocal stealing of brood, is common among incipient colonies of the Ere ant SoUnopsis invicta. Paradox-ically, workers from a colony that loses its brood during a raid often abandon their nest and join the winning colony. Queensabandoned by their workers may then migrate from their original nest site and attempt to forcefully usurp another incipientcolony by displacing that colony's queen or queens. This study examined factors that influence the success of usurpationattempts. Queens attempting to usurp a nearby colony after laboratory brood raids were successful in less than 30% of trials.Usurpation attempts were more successful if workers familiar to the migrating queen were present in the nest, as would happenif a queen were to find the colony to which her workers had moved. Cross-fostering experiments showed that this effect wasdue to familiarity rather than relatedness. Usurpation attempts were less successful if they were delayed by 16 h. The probabilityof usurpation success was not reduced by doubling the number of defending workers in the invaded colony. However, coloniesfounded by three queens were almost always able to resist usurpation attempts. These results support the hypothesis that workersabandon their natal colony after losing a brood raid to increase the likelihood that their queen can usurp the colony to whichthey migrate. These results also provide the first evidence that colonies of ants founded by several queens are better able toresist usurpation attempts than colonies founded by a single queen. Key words brood raids, fire ants, kin recognition, SoUnopsisinvicta, usurpation. [Bthav Ecol 8:99-103 (1997)1
Intraspecific competition among incipient colonies of socialinsects is often intense, resulting in high mortality of
queens before the colonies reach reproductive maturity (H61-ldobler and Wilson, 1990). In some ant species, much of thismortality is associated with brood raiding. During brood raids,incipient colony workers remove eggs, larvae, and pupae fromneighboring colonies and take them to their home colony,thereby supplementing the worker force (Bartz and Holldob-ler, 1982; Rissing and Pollock, 1987; Stamps and Vinson, 1991;Tschinkel, 1992a,b). Brood raids by the fire ant SoUnopsis in-victa (Buren) cause a substantial reduction in the density ofincipient colonies and queens (Adams and Tschinkel, 1995a).Queens whose colonies have been depleted of brood throughthese raids often attempt to usurp nearby colonies whosebrood population has been increased through raiding (Adamsand Tschinkel 1995a,b; Tschinkel, 1992b).
Success in brood raids rises with the size of the worker pop-ulation (Rissing and Pollock, 1991; Stamps and Vinson, 1991;Tschinkel, 1992b). One way a colony can produce more work-ers in its first brood is through pleometrosis, or colony foun-dation by several queens (reviewed in Rissing and Pollock,1988). In three species for which raiding has been described,Mymucocystus mimicus (Bartz and H&lldobler, 1982), Mtssorprrganda (Rissing and Pollock, 1987, 1991), and £ invicta(Adams and Tschinkel, 1995b; Tschinkel, 1992b), pleometrot-ic colonies are more likely to win brood raids. Tschinkel(1992b) demonstrated that this effect was due to increasedworker number and not increased queen number.
A peculiar behavior associated with brood raiding is theabandonment of an incipient colony by its workers after itsbrood has been removed. When abandonment occurs, work-ers leave their colony's queen or queens at the original nest
Address correspondence to E. S. Adams.M. T. -Balai u now at St. John'i University, 300 Howard Ave., Staten
Island, NY 10301, USA.Received 31 August 1995; revised 27 March 1996; accepted 29
March 1996.1045-2249/97/15.00 O 1997 International Society for Behavioral Ecology
and peacefully join the colony that successfully raided it (Bartzand HoUdobler, 1982; Rissing and Pollock. 1987; Tschinkel,1992b). However, a queen from a losing colony will usuallyattempt usurpation shortly after her workers abandon her(Adams and Tschinkel, 1995a,b; Bartz and HoUdobler, 1982;Rissing and Pollock, 1987; Tschinkel, 1992b). It has been pro-posed that workers' relocation is adaptive if the workers'mother queen has a high probability of joining them in thenew colony, a process termed "hopeful co-emigration" (Pol-lock and Rissing, 1989).
Queens attempting to enter other incipient colonies mayencounter resistance. Workers of fire ant incipient coloniesrecognize familiar queens and often expel or kill unfamiliarqueens (Balas and Adams, 1996b). However, migrating queensoccasionally succeed in displacing the resident queen orqueens, which may be killed and discarded (Adams andTschinkel, 1995a,b; Rissing and Pollock, 1987; Tschinkel,1992a). In addition, Adams and Tschinkel (1995b) showedthat if a queen attempts to usurp a colony that has won a raid,her probability of success is greater if the workers that joinedthe colony are her daughters. However, it is not known wheth-er this effect is due to the presence of familiar workers orsimply to the smaller number of defending workers in dieinvaded colony.
This study examines whether workers of S. invicta are ableto assist their mothers in colony usurpations. First, we testedwhether workers of S. invicta are better able to usurp coloniesto which their daughters have moved. Second, we testedwhether the effect of daughters on usurpation success is dueto familiarity with the queen or to genetic relatedness. Ag-gressive behavior during these usurpation attempts was ob-served to determine if the presence of familiar or relatedworkers influenced the aggressive response toward either theresident queen or the queen attempting usurpation.
In addition, preliminary experiments suggested that aqueen's probability of usurping the colony to which herdaughters have moved may decline if she does not migrateimmediately after a brood raid. This may be because queensrapidly lose familiarity to their former nestmates. We tested
Dow
nloaded from https://academ
ic.oup.com/beheco/article/8/1/99/211854 by guest on 16 January 2022

100 Behavioral Ecology Vol. 8 No. 1
this possibility by delaying usurpation attempts for 16 h afterthe conclusion of brood raids.
Finally, since colonies with more workers have an advantagein brood raids (Tschinkel, 1992b), we tested the possibilitythat increasing worker number in a colony increases that co-lony's ability to resist usurpation. Since worker number co-varies with queen number, we also tested whether incipientcolonies with several queens resist usurpation attempts moresuccessfully.
METHODS
We collected mated queens after nuptial flights in Tallahassee,Florida, USA, during April and May 1994. Queens were placedin glass nest-tubes 8-10 mm in diam and 75 mm long thatwere half-filled with water retained by a cotton plug and alsoplugged with cotton at the open end. These colonies weremaintained at 29°C during the experiment. As toon as threeworkers had edosed in a colony, the nest tube was openedand placed in a plastic box 10.5 X 10.5 X 3.4 cm, the wallsof which were coated with Fluon (Northern Products Inc.,Woonsocket, Rhode Island) to prevent escapes. Colonies werefed crickets and diluted honey immediately after the firstworker edosed and 1 day before the experimental trial. Toassure that the colonies were healthy, we used only coloniesthat had at least eight workers 5 days after the first workeredosed. This is approximately the average number of workerspresent per single-queen nest at the time brood raids beginin the field (Adams and Tschinkel, 1995a). We distinguishedqueens by unique paint marks on the queens' alitrunks. Thesemarks have no effect on aggression against familiar or unfa-miliar queens (Balas and Adams, 1996a).
Tests of usurpation «"«y»««
Brood raids were staged in the laboratory in two differentcontainers: the colonies' nest-boxes, in which the interior ofeach nest was visible for behavioral observations, and cupsfilled with sand, which more closely mimic natural conditions.In the nest-boxes, raids were initiated by removing a colony'snest tube from its box and placing it beside a second colony'snest tube. To prepare raids in cups of sand, 1 frounce cupswere filled with damp sand with a small hole about 7 cm deeppoked into the center to serve as an artificial nest site. Wethen gently tapped a colony's queen, workers, and brood outof the nest tube and onto the sand's surface. The workers andqueen quickly found the hole and moved their brood into itWe allowed the colony to settle into the hole overnight. Thenext morning, raids were initiated by placing a second colo-ny's nest tube on the sand's surface. With either nest design,raids were usually completed between 2 and 4 h after thecolonies were placed together. In a minority of trials (26 of478; 5.4%), raids had not begun after 4 h. We discarded thesetrials to avoid isolating the queen for longer than the exper-imental treatment indicated.
Umxunipulated raids
The unmanipulated raids experiment measured the successrate of queen migration and usurpation after raids betweentwo one-queen incipient colonies. We placed colonies in pairsin plastic nest-boxes and observed them at 20-min intervals.The colony that contained all the brood at the raid's conclu-sion was recorded, as well as any migrations by the losing co-lony's queen to the winning colony's tube. In some cases,queen migration was not observed directly but was inferred
from the queens' paint marks. Hereafter, any queens thatmoved to or were placed at the entrance of other colonies inorder to simulate a usurpation attempt (see below) are re-ferTed to as "migrating queens." We observed the colony 1 hafter queen migration for aggressive behavior (e.g., biting orstinging) by the workers against either queen. Each colonywas scanned once a day for 3 days after the raid, and anyevidence of queen injury, such as missing appendages or le-thargic movements, was recorded. We noted dead or expelledqueens as well. The experiment was terminated after 3 days,and the fates of both queens were determined.
Effects of *"""*"• or related workers on usurpation m w
We tested whether queens are better able to usurp a colonywhen workers familiar with her are present in that colony andwhether this effect depends on genetic relatedness betweenthe queens and workers. This was achieved using a two-factordesign that varied both workers' previous familiarity with andgenetic relatedness to a queen. Before raids were initiated inthis experiment, one queen from each pair of colonies waschosen at random and held aside in a dean test tube duringthe raid. Raids took place between die queenless colony andan unmanipulated one-queen colony. After the raid's conclu-sion, usurpation attempts were initiated by placing die isolat-ed queen at the opening of the consolidated colony.
We introduced queens either to colonies containing theirformer workers or to colonies in which all workers were un-familiar with her. Also, by cross-fostering queens between col-onies shortly before their workers edosed, replicates were cre-ated in which workers were not familiar with dieir biologicalmother as adults. The five experimental treatments were (1)the migrating queen was both related to and familiar to theraided workers (familiar kin treatment); (2) the queen wasneither related to nor familiar to the raided workers (unfa-miliar nonkin treatment); (3) the queen was familiar to theraided workers but was not related to them (familiar nonkintreatment). These were queens that were cross-fostered withunrelated pupae just before the workers edosed; (4) thequeen was related to die raided workers but was unfamiliarto diem (unfamiliar kin treatment). In this treatment, a queentried to usurp a colony that contained her daughters, but herdaughters had been cross-fostered with an unrelated queen;(5) the migrating queen was introduced mto a colony thathad not raided (no raid treatment).
One hour after the queens' introductions, half of the col-onies were observed for any incidents of aggression. The oth-er raids took place in sand as stated previously; therefore, thequeens were usually not visible. We determined die fates ofqueens after 3 days by observing dead or expelled queens orby excavating colonies in sand.
Influence of of queen migration on usurpation
We tested whether delaying a usurpation attempt by 16 h re-duced usurpation success. Raid trials were conducted as in theprevious experiment, except diat in this experiment die iso-lated queens were released into die enlarged colony about 16h after die raid's condusion. Only die familiar kin and un-familiar nonkin treatments were induded in diis experiment
Influences of resident queen and worker number
Finally, we tested die effects of increasing queen or workernumber in a resident colony on die success rate of usurpationattempts after raids. Raids were induced under diree experi-mental conditions: (1) die resident colony's worker number
Dow
nloaded from https://academ
ic.oup.com/beheco/article/8/1/99/211854 by guest on 16 January 2022
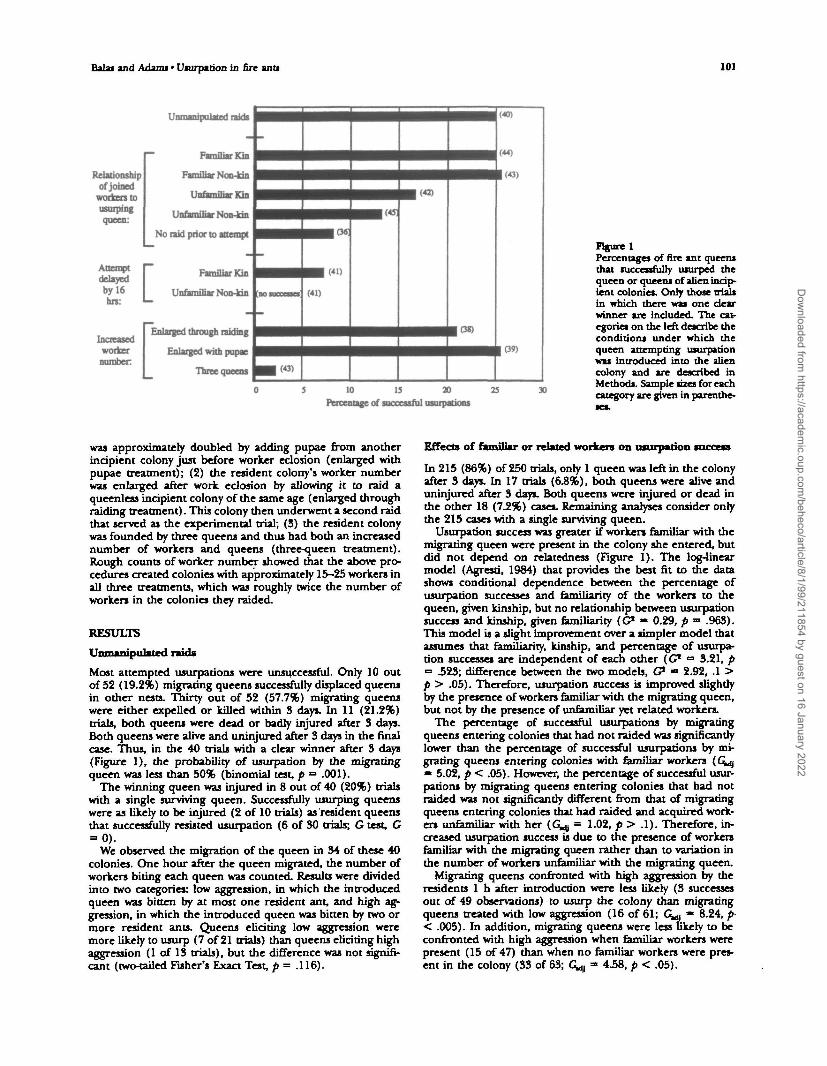
Balas and Adams • Usurpation in fire ants 101
Relationshipofjoined
workers tousurpingqueen:
Attemptdelayedby 16hrs:
Increasedworkernumber
Familiar Kin
Unfamiliar Kin
Unfamiliar Non-kin
No raid prior to attempt
Familiar Kin
Unfamiliar Non-kin | a o
Enlarged through raiding
Enlarged with pupae
Three queens
r
laoncseoo
• B (43)
(41)
• (43)
(44)
10 IS 20Percentage of successful usurpations
figure 1Percentages of fire ant queensthat successfully usurped thequeen or queens of alien incip-ient colonies. Only those trialsin which there was one dearwinner are included. The cat-egories on the left describe theconditions under which thequeen attempting usurpationwas introduced into the aliencolony and are described inMethods. Sample sizes for eachcategory are given in parenthe-
was approximately doubled by adding pupae from anotherincipient colony just before worker eclosion (enlarged withpupae treatment); (2) the resident colony's worker numberwas enlarged after work eclosion by allowing it to raid aqueenless incipient colony of the same age (enlarged throughraiding treatment). This colony then underwent a second raidthat served as the experimental trial; (3) the resident colonywas founded by three queens and thus had both an increasednumber of workers and queens (three-queen treatment).Rough counts of worker number showed that the above pro-cedures created colonies with approximately 15-25 workers inall three treatments, which was roughly twice the number ofworkers in the colonies they raided.
RESULTS
Unmsoiipulftted raids
Most attempted usurpations were unsuccessful. Only 10 outof 52 (19.2%) migrating queens successfully displaced queensin other nests. Thirty out of 52 (57.7%) migrating queenswere either expelled or killed within 3 days. In 11 (21.2%)trials, both queens were dead or badly injured after 3 days.Both queens were alive and uninjured after 3 days in the finalcase. Thus, in the 40 trials with a clear winner after 3 days(Figure 1), the probability of usurpation by the migratingqueen was less than 50% (binomial test, p =• .001).
The winning queen was injured in 8 out of 40 (20%) trialswith a single surviving queen. Successfully usurping queenswere as likely to be injured (2 of 10 trials) as resident queensthat successfully resisted usurpation (6 of 30 trials; G test, G= 0).
We observed the migration of the queen in 34 of these 40colonies. One hour after the queen migrated, the number ofworkers biting each queen was counted. Results were dividedinto two categories: low aggression, in which the introducedqueen was bitten by at most one resident ant, and high ag-gression, in which the introduced queen was bitten by two ormore resident ants. Queens eliciting low aggression weremore likely to usurp (7 of 21 trials) than queens eliciting highaggression (1 of 13 trials), but the difference was not signifi-cant (two-tailed Fisher's Exact Test, p = .116).
Effects of familiar or related workers on usurpation success
In 215 (86%) of 250 trials, only 1 queen was left in the colonyafter 3 days. In 17 trials (6.8%), both queens were alive anduninjured after 3 days. Both queens were injured or dead inthe other 18 (7.2%) cases. Remaining analyses consider onlythe 215 cases with a single surviving queen.
Usurpation success was greater if workers familiar with themigrating queen were present in the colony she entered, butdid not depend on relatedness (Figure 1). The log-linearmodel (Agresti, 1984) that provides the best fit to the datashows conditional dependence between the percentage ofusurpation successes and familiarity of the workers to thequeen, given kinship, but no relationship between usurpationsuccess and kinship, given familiarity (G* • 0.29, p =» .963).This model is a slight improvement over a simpler model thatassumes that familiarity, kinship, and percentage of usurpa-tion successes are independent of each other (C1 = 3.21, p= 323; difference between the two models, C - 2.92, .1 >p > .05). Therefore, usurpation success is improved slightlyby the presence of workers familiar with the migrating queen,but not by the presence of unfamiliar yet related workers.
The percentage of successful usurpations by migratingqueens entering colonies that had not raided was significantlylower than the percentage of successful usurpations by mi-grating queens entering colonies with familiar workers (G^m 5.02, p < .05). However, the percentage of successful usur-pations by migrating queens entering colonies that had notraided was not significantly different from that of migratingqueens entering colonies that had raided and acquired work-ers unfamiliar with her (G^ = 1.02, p > .1). Therefore, in-creased usurpation success is due to the presence of workersfamiliar with the migrating queen rather than to variation inthe number of workers unfamiliar with the migrating queen.
Migrating queens confronted with high aggression by theresidents 1 h after introduction were less likely (3 successesout of 49 observations) to usurp the colony than migratingqueens treated with low aggression (16 of 61; G^n m 8.24, p-< .005). In addition, migrating queens were less ukety to beconfronted with high aggression when familiar workers werepresent (IS of 47) than when no familiar workers were pres-ent in the colony (33 of 63; Gy, =• 4.58, p < .05).
Dow
nloaded from https://academ
ic.oup.com/beheco/article/8/1/99/211854 by guest on 16 January 2022

102 Behavioral Ecology Vol. 8 No. 1
Resident queens elicited high aggression in 7 out of 91(7.7%) trials when workers unfamiliar with the resident queenwere present in the colony before the migrating queen wasintroduced. High aggression against resident queens was notassociated with an increased likelihood that the residentqueen would be usurped (2 usurpations out of 7 cases of highaggression against resident queens versus 14 usurpations outof 84 cases of low aggression; G^ = 0.49, p > .1).
Influence of rimfatg of Queen migration on usurpation
When introductions of migrating queens were delayed by 16h after conclusions of brood raids, the percentage of success-ful usurpations declined, whether or not workers familiar withthe migrating queen were present (Figure 1). These percent-ages were significantly lower for both the familiar kin andunfamiliar nonkin treatments, compared with the similartreatments when migrating queens were introduced immedi-ately after a raid's conclusion (familiar kin, Gu = 4.92, p <.05; unfamiliar nonkin, G^ = 7.41, p < .01). As in the pre-vious experiment, migrating queens usurped colonies moreoften if workers familiar with the migrating queen were pres-ent than if no workers familiar with the migrating queen werepresent (Figure 1), but this difference was only weakly signif-icant (G^ = 3.60, .1 > p > .05).
Influences of resident queen and worker number
Migrating queens were just as likely to usurp colonies withartificially supplemented worker populations as they were tousurp a colony with an equal number of workers (Figure 1;unmanipulated raids versus enlarged through raiding, G^ =0.22, p > .30; unmanipulated raids versus enlarged with pu-pae, G^g = 0.004, p > .95). However, a migrating queen wassignificantly less likely to usurp a colony that contained threecofounding queens than colonies that contained one queen
10.38, p < .005).
DISCUSSION
Usurpation attempts by queens whose colonies have lostbrood raids are usually unsuccessful. However, evidence fromthis study and elsewhere (Adams and Tschinkel, 1995b) in-dicates that usurpation attempts are more successful if themigrating queen enters a colony containing familiar workers.The percentages of successful usurpations reported in the twostudies do not differ significantly (familiar kin: G^ = 0.96, p> .1; unfamiliar nonkin: G^ = 1.44, p> .1), although a weak-er effect was found in this study than by Adams and Tschinkel(1995b). When the data from these two studies are pooled,migrating queens usurped invaded colonies in 35 out of 124attempts (28.2%) when workers familiar with her were pres-ent, but in only 15 out of 124 attempts (12.1%) if no workersfamiliar with her were present. This difference is significant(Gkg ** 10.1, p < .001), indicating that a migrating queen is2.3 times as likely to usurp a colony if it contains familiarworkers.
The increase in usurpation success associated with the pres-ence of familiar workers supports the "hopeful co-emigra-tion" hypothesis proposed by Pollock and Rissing (1989). Thishypothesis suggests that the workers and queen of a colonythat has lest all of its brood attempt to make the best of a badsituation by usurping a surviving colony. In general, workerswill usually attempt to win a raid first by stealing another co-lony's brood. If a colony loses a raid, the workers defect tothe winning colony (Bartz and HoUdobler, 1982; -Kissing andPollock, 1987; Stamps and Vmson, 1991; Tschinkel, 1992b).
It has recently been suggested that queens move freelyamong incipient nests in S. mvicta (Pfennig, 1995) and thatbrood raiding may actually be a form of cooperative nest con-solidation (Nonacs, 1993). Yet this study and others (Adamsand Tschinkel, 1995a,b; Balas and Adams, 1996b) show thatworkers actively resist entry by unfamiliar queens, which theyoften kill. The phenomenon of "traitor" raiders that movebrood away from their home nest (Tschinkel, 1992b) may sug-gest that cooperative nest consolidation (Nonacs, 1993) ratherthan raiding is taking place. However, traitors have only beenreported for & mvicta and are less common than workers thatmaintain fidelity to their nest site (Tschinkel, 1992b). Fur-thermore, most attempted usurpations are unsuccessful (Fig-ure 1). Thus, although queens can occasionally usurp othernests after raids, usurpation appears to be an attempt to re-cover from what would otherwise be a competitive loss.
The presence of familiar workers improves migratingqueens' usurpation chances. Figure 1 shows that migratingqueens are better able to usurp colonies containing workersfamiliar with her than either colonies that have not raided orcolonies that acquired workers unfamiliar with the migratingqueen. The mechanism by which familiar workers confer thisadvantage is not fully understood. They may attack the resi-dent queen when their former nest mate queen arrives. How-ever, aggression against resident queens was uncommon andwas not associated, with their subsequent death or expulsion.
Alternatively, by joining surviving colonies, the joiningworkers may lower the aggressive response by the residentworkers toward their own invading queen. In support of thishypothesis, migrating queens elicited lower aggression in col-onies containing workers familiar with her, and low aggressionwas associated with greater usurpation success. One possiblemechanism for this effect is that the original residents of thecolony adjust their recognition template to include the colonyodor carried by the new workers. When the new workers'mother queen arrives shortly thereafter, they may be slightlyless likely to judge her as unfamiliar. However, the usurpationsuccess was lower if the attempt was delayed, and the windowof opportunity for this advantage is less than 16 h. This maybe because workers quickly acquire die odor of the new col-ony after their migration.
Doubling the worker number in a single-queen resident col-ony did not increase the chance that the colony would suc-cessfully resist a usurpation attempt, even though increasingworker number increases the likelihood that a colony may wina raid (Adams and Tschinkel, 1995b; Rissing and Pollock,1987; Stamps and Vinson, 1991; Tschinkel, 1992b). However,colonies founded by three queens successfully resisted usur-pation in all but one trial. This shows that one advantage tocooperative colony foundation in S. mvicta is the increasedability of multiple-queen colonies to resist usurpation at-tempts. This is die first report of this phenomenon in ants,although two cases of such an advantage gained by pleome-trotic associations are known within the wasp genus PoHstts(Gamboa, 1978; Klahn, 1988).
Usurpation success was not improved by the presence of amigrating queen's biological daughters if the daughters wereunfamiliar with her. Although heritable cues have been im-plicated in recognition by workers of mature & mvicta colo-nies, they are neither necessary nor sufficient for acceptanceof queens in incipient colonies (Balas and Adams, 1996b). hiaddition, environmentally correlated cues play a much largerrole than heritable cues in recognition in mature fire ant col-onies, and these cues are learned by the workers (Obin andVander Meer, 1988, 1989a,b). Nest mate recognition cues ap-pear to be learned by incipient colony workers as well, be-cause migrating queens had a usurpation advantage even iftheir former nest mate workers were unrelated to them. This
Dow
nloaded from https://academ
ic.oup.com/beheco/article/8/1/99/211854 by guest on 16 January 2022

Balas and Adams • Umrpadon in lire ants 103
adds to the large body of evidence that nest mate recognitionis based on cues learned after worker edosion (reviewed inBreed and Bennett, 1987; see also Alloway and Ryckman,1991; Breed and Julian, 1992; Errard, 1994; OWn and VinderMeer, 1988, 1989a,b; Stuart, 1988). Heritable cues may be in-volved in nest mate recognition (Carlin, 1989), but there islittle evidence that ants have the ability to recognize unfamil-iar kin (but see Provost, 1991).
We are grateful to Walter Twhinkcl and the Department of BiologicalSdence at Florida State University for their assistance during thecourse of this project Steven Kissing and Larry Wolf provided helpfulcomments on the manuscript. This work was supported by a Fellow-ship in Science and Engineering from the David and LucUe PackardFoundation to ZSJ
REFERENCES
Adams ES, Tschinkel WR, 1995a. Density-dependent competition inAre ants: effects on colony survivorship and size variation. J AnimEcol 64315-324.
Adams ES, Tschinkel WR, 1995b. Effects of foundress number onbrood raids, queen survival and population dynamics in the fire antSoUnopnt invicta. Behav Ecol Sodobiol 37:233-242.
Agresti A, 1984. Analysis of ordinal categorical data. New fork: JohnWiley and Sons.
Alloway TM, Ryckman D, 1991. Learned social attachment to queensin Ltptothomx embiguus Emery ant workers. Behaviour 118:235-243.
Balas MT, Adams ES, 1996a. The dissolution of cooperative groups:mechanisms of queen mortality in incipient fire ant colonies. BehavEcol Sodobiol 38 J91-399.
Balas MT, Adams ES, 1996b. Nestmate discrimination and competi-tion in incipient colonies of fire ants. Anim Behav 51:49-59.
Bam SH, HolldoWer B, 1982. Colony founding in Mynottocysttu mim-icus Wheeler (Hymenoptera: Formiddae) and the evolution offoundress associations. Behav Ecol Sodobiol 10:137-147.
Breed MD, Bennett B, 1987. Kin recognition in highly eusodal in-sects. In: Kin recognition in animals (Fletcher DJC, Mkhener CD,eds). New Yfark: John Wiley and Sons; 243-285.
Breed MD, Julian GE, 1992. Do simple rules apply in honey-bee nest-mate discrimination? Nature 357:685-686.
Carlin NF, 1989. Discrimination between and within colonies of socialinsects: two null hypotheses. Neth J Zool 39:86-100.
Errard C, 1994. Long-term memory involved in nestmate recognitionin ants. Anim Behav 48:263-271.
Gamboa GJ, 1978. Intraspecific defense: advantage of social cooper-ation among paper wasp foundresses. Science 199:1463-1465.
Holldobler B, Wilson EO, 1990. The ants. Cambridge, Massachusetts:Bdknap Press.
Klahn J, 1988. Intraspecific comb usurpation in the social wasp Pohitafiucotiu. Behav Ecol Sodobiol 23:1-8.
Nonacs P, 1993. The economics of brood raiding and nest consoli-dation during ant colony founding. Evol Ecol 7:625-633.
Obin MS, vander Meer RK, 1988. Sources of nestmate recognitioncues in the imported fire ant, Soimopsis invicta Buren (Hymenop-tera: Formiddae). Anim Behav 36:1361-1370.
Obin MS, Vinder Meer RK, 1989a. Mechanism of template-labelnrjtrhing in fir* ant, HrJttutfnij ^,,n»/r Riirw ni-OTniti- rfmgnitWi
Anim Behav 58:430-435.Obin MS, Vander Meer RK, 1989b. Nestmate recognition in fire ants
(SoUnopns invicta Buren). Do queens label workers? Ethology 80:255-264.
Pfennig DW, 1995. Absence of joint nesting advantage in desert seedharvester ante evidence from a field experiment. Anim Behav 49:567-575.
Pollock GB, Rissing SW, 1989. Intraspecific brood raiding, territori-ah'ty, and slavery in ants. Am Nat 133.-61-70.
Provost E, 1991. Nonnestmate kin recognition in the ant LtptotMoraxUckitnsUhu. evidence that genetic factors regulate colony recogni-tion. Behav Gen 21:151-167.
Rissing SW, Pollock GB, 1987. Queen aggression, pleometrotic advan-tage and brood raiding in the ant Varmmter ptrganda (Hymenop-tera: Formiddae)./nim Behav 35:975-981.
Rissing SW, Polkxk GB, 1988. Pleometrosis and polygyny in ants. In:Interindividual behavioral variation in social insects (Jeanne RL,ed). Boulder, Colorado: Westview Press; 179-222.
Rissing SW, Pollock GB, 1991. An experimental analysis of pleome-trotic advantage in the desert seed-harvester ant Mtaor ptrgmtUi(Hymenoptera: Formiddae). Insectes Soc 38:205-211.
Stamps WT, Vinson SB, 1991. Raiding in newly founded colonies ofSoUnopns invicta Buren (Hymenoptera: Formiddae). Environ En-tomol 20:1037-1041.
Stuart RJ, 1988. Collective cues u a basis for nrsrmatr recognition inpotygynous leptothoradne ants. Proc Nad Acad Sd USA 85:4572-4575.
Tschinkel WR, 1992a. Brood raiding in the fire ant, SoUnopns invicta(Hymenoptera: Formiddae): laboratory and field observations.Ann Entomol Soc Am 85:638-646.
Tschinkel WR, 1992b. Brood raiding and the population dynamics offounding and Indpient colonies of the fire ant, SoUnopns invicta.Ecol Entomol 17:179-188.
Dow
nloaded from https://academ
ic.oup.com/beheco/article/8/1/99/211854 by guest on 16 January 2022


















