
Intra- and Interspecies Regulation of Gene Expression …iai.asm.org/content/69/12/7625.full.pdf ·...
10
INFECTION AND IMMUNITY, 0019-9567/01/$04.000 DOI: 10.1128/IAI.69.12.7625–7634.2001 Dec. 2001, p. 7625–7634 Vol. 69, No. 12 Copyright © 2001, American Society for Microbiology. All Rights Reserved. Intra- and Interspecies Regulation of Gene Expression by Actinobacillus actinomycetemcomitans LuxS KAREN P. FONG, 1 WHASUN O. CHUNG, 2 RICHARD J. LAMONT, 2 AND DONALD R. DEMUTH 1 * Department of Biochemistry, School of Dental Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, 1 and Department of Oral Biology, University of Washington, Seattle, Washington 2 Received 6 July 2001/Returned for modification 28 August 2001/Accepted 17 September 2001 The cell density-dependent control of gene expression is employed by many bacteria for regulating a variety of physiological functions, including the generation of bioluminescence, sporulation, formation of biofilms, and the expression of virulence factors. Although periodontal organisms do not appear to secrete acyl-homoserine lactone signals, several species, e.g., Porphyromonas gingivalis, Prevotella intermedia, and Fusobacterium nuclea- tum, have recently been shown to secrete a signal related to the autoinducer II (AI-2) of the signal system 2 pathway in Vibrio harveyi. Here, we report that the periodontal pathogen Actinobacillus actinomycetemcomitans expresses a homolog of V. harveyi luxS and secretes an AI-2-like signal. Cell-free conditioned medium from A. actinomycetemcomitans or from a recombinant Escherichia coli strain (E. coli AIS) expressing A. actinomyce- temcomitans luxS induced luminescence in V. harveyi BB170 >200-fold over controls. AI-2 levels peaked in mid-exponential-phase cultures of A. actinomycetemcomitans and were significantly reduced in late-log- and stationary-phase cultures. Incubation of early-log-phase A. actinomycetemcomitans cells with conditioned me- dium from A. actinomycetemcomitans or from E. coli AIS resulted in a threefold induction of leukotoxic activity and a concomitant increase in leukotoxin polypeptide. In contrast, no increase in leukotoxin expression occurred when cells were exposed to sterile medium or to conditioned broth from E. coli AIS , a recombinant strain in which luxS was insertionally inactivated. A. actinomycetemcomitans AI-2 also induced expression of afuA, encoding a periplasmic iron transport protein, approximately eightfold, suggesting that LuxS-dependent signaling may play a role in the regulation of iron acquisition by A. actinomycetemcomitans. Finally, A. actinomycetemcomitans AI-2 added in trans complemented a luxS knockout mutation in P. gingivalis by modu- lating the expression of the luxS-regulated genes uvrB and hasF in this organism. Together, these results suggest that LuxS-dependent signaling may modulate aspects of virulence and the uptake of iron by A. actinomycetemcomitans and induce responses in other periodontal organisms in mixed-species oral biofilm. The regulation of bacterial gene expression in response to changes in cell density is known as quorum sensing. Quorum- sensing bacteria synthesize and secrete extracellular signaling molecules called autoinducers, which accumulate in the envi- ronment as the population increases. When a critical threshold concentration of autoinducer is attained, a signal transduction cascade is triggered, resulting in an alteration in gene expres- sion and a change in behavior of the organism (3, 14). For example, two quorum-sensing systems in Vibrio harveyi, signal systems 1 and 2, function in parallel to control the density- dependent expression of bioluminescence (4) and other cellu- lar processes (32). The autoinducer of signal system 1 (AI-1) has been identified as an acyl-homoserine lactone (acyl-HSL) (5, 10), whereas the structure of AI-2 has not yet been fully resolved. The synthesis of the acyl-HSL AI-1 requires two polypeptides encoded by luxL and luxM (4), whereas AI-2 is synthesized by the luxS gene (49). AI-1 and AI-2 are recog- nized by their cognate sensor kinase proteins, LuxN and LuxQ, respectively, although AI-2 interaction with the LuxQ sensor may also be mediated by a periplasmic-binding protein, desig- nated LuxP (5). At low cell density, and hence low autoinducer concentration, the sensors LuxN and LuxQ function as kinases and autophosphorylate (32). Phosphate is subsequently trans- ferred to a shared integrator protein, LuxU, which in turn donates phosphate to the response regulator protein, LuxO (6, 16, 17). Phosphorylated LuxO is active and presumably induces the expression or activity of an unidentified repressor of the luciferase structural operon, luxCDABE (32). In contrast, at high cell density, LuxN and LuxQ bind their cognate signals and function as phosphatases, which draws phosphate away from LuxO in a LuxU-dependent reaction. Thus, LuxO be- comes inactive and the downstream repression of luxCDABE is removed, resulting in the production of light. Acyl-HSL-dependent quorum-sensing systems exist in many gram-negative bacteria, e.g., Pseudomonas aeruginosa, Agrobacterium tumefaciens, and Ralstonia solanacearum, and control diverse cellular functions, including toxin and alginate production in P. aeruginosa (46), type IV secretion in A. tume- faciens (35), exoenzyme production in Burkholderia cepacia (31), and the expression of other virulence-associated factors (2, 3, 13, 19, 37, 48, 49). However, a recent survey of gram- negative periodontal organisms by Frias et al. (18) suggested that this group of bacteria do not possess acyl-HSL-dependent signaling circuits. Instead, several of these organisms, e.g., Fu- sobacterium nucleatum, Porphyromonas gingivalis, and Pre- votella intermedia, appeared to secrete a signal related to AI-2 of V. harveyi. Indeed, highly conserved homologs of V. harveyi luxS have recently been identified in both gram-negative and * Corresponding author. Mailing address: Room 540, Levy Re- search Building, Department of Biochemistry, School of Dental Med- icine, University of Pennsylvania, 4010 Locust St., Philadelphia, PA 19014-6002. Phone: (215) 898-2125. Fax: (215) 898-3695. E-mail: [email protected]. 7625 on August 27, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Intra- and Interspecies Regulation of Gene Expression …iai.asm.org/content/69/12/7625.full.pdf ·...

INFECTION AND IMMUNITY,0019-9567/01/$04.00�0 DOI: 10.1128/IAI.69.12.7625–7634.2001
Dec. 2001, p. 7625–7634 Vol. 69, No. 12
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Intra- and Interspecies Regulation of Gene Expression byActinobacillus actinomycetemcomitans LuxS
KAREN P. FONG,1 WHASUN O. CHUNG,2 RICHARD J. LAMONT,2 AND DONALD R. DEMUTH1*
Department of Biochemistry, School of Dental Medicine, University of Pennsylvania, Philadelphia, Pennsylvania,1
and Department of Oral Biology, University of Washington, Seattle, Washington2
Received 6 July 2001/Returned for modification 28 August 2001/Accepted 17 September 2001
The cell density-dependent control of gene expression is employed by many bacteria for regulating a varietyof physiological functions, including the generation of bioluminescence, sporulation, formation of biofilms, andthe expression of virulence factors. Although periodontal organisms do not appear to secrete acyl-homoserinelactone signals, several species, e.g., Porphyromonas gingivalis, Prevotella intermedia, and Fusobacterium nuclea-tum, have recently been shown to secrete a signal related to the autoinducer II (AI-2) of the signal system 2pathway in Vibrio harveyi. Here, we report that the periodontal pathogen Actinobacillus actinomycetemcomitansexpresses a homolog of V. harveyi luxS and secretes an AI-2-like signal. Cell-free conditioned medium from A.actinomycetemcomitans or from a recombinant Escherichia coli strain (E. coli AIS) expressing A. actinomyce-temcomitans luxS induced luminescence in V. harveyi BB170 >200-fold over controls. AI-2 levels peaked inmid-exponential-phase cultures of A. actinomycetemcomitans and were significantly reduced in late-log- andstationary-phase cultures. Incubation of early-log-phase A. actinomycetemcomitans cells with conditioned me-dium from A. actinomycetemcomitans or from E. coli AIS resulted in a threefold induction of leukotoxic activityand a concomitant increase in leukotoxin polypeptide. In contrast, no increase in leukotoxin expressionoccurred when cells were exposed to sterile medium or to conditioned broth from E. coli AIS�, a recombinantstrain in which luxS was insertionally inactivated. A. actinomycetemcomitans AI-2 also induced expression ofafuA, encoding a periplasmic iron transport protein, approximately eightfold, suggesting that LuxS-dependentsignaling may play a role in the regulation of iron acquisition by A. actinomycetemcomitans. Finally, A.actinomycetemcomitans AI-2 added in trans complemented a luxS knockout mutation in P. gingivalis by modu-lating the expression of the luxS-regulated genes uvrB and hasF in this organism. Together, these resultssuggest that LuxS-dependent signaling may modulate aspects of virulence and the uptake of iron by A.actinomycetemcomitans and induce responses in other periodontal organisms in mixed-species oral biofilm.
The regulation of bacterial gene expression in response tochanges in cell density is known as quorum sensing. Quorum-sensing bacteria synthesize and secrete extracellular signalingmolecules called autoinducers, which accumulate in the envi-ronment as the population increases. When a critical thresholdconcentration of autoinducer is attained, a signal transductioncascade is triggered, resulting in an alteration in gene expres-sion and a change in behavior of the organism (3, 14). Forexample, two quorum-sensing systems in Vibrio harveyi, signalsystems 1 and 2, function in parallel to control the density-dependent expression of bioluminescence (4) and other cellu-lar processes (32). The autoinducer of signal system 1 (AI-1)has been identified as an acyl-homoserine lactone (acyl-HSL)(5, 10), whereas the structure of AI-2 has not yet been fullyresolved. The synthesis of the acyl-HSL AI-1 requires twopolypeptides encoded by luxL and luxM (4), whereas AI-2 issynthesized by the luxS gene (49). AI-1 and AI-2 are recog-nized by their cognate sensor kinase proteins, LuxN and LuxQ,respectively, although AI-2 interaction with the LuxQ sensormay also be mediated by a periplasmic-binding protein, desig-nated LuxP (5). At low cell density, and hence low autoinducer
concentration, the sensors LuxN and LuxQ function as kinasesand autophosphorylate (32). Phosphate is subsequently trans-ferred to a shared integrator protein, LuxU, which in turndonates phosphate to the response regulator protein, LuxO (6,16, 17). Phosphorylated LuxO is active and presumably inducesthe expression or activity of an unidentified repressor of theluciferase structural operon, luxCDABE (32). In contrast, athigh cell density, LuxN and LuxQ bind their cognate signalsand function as phosphatases, which draws phosphate awayfrom LuxO in a LuxU-dependent reaction. Thus, LuxO be-comes inactive and the downstream repression of luxCDABE isremoved, resulting in the production of light.
Acyl-HSL-dependent quorum-sensing systems exist in manygram-negative bacteria, e.g., Pseudomonas aeruginosa,Agrobacterium tumefaciens, and Ralstonia solanacearum, andcontrol diverse cellular functions, including toxin and alginateproduction in P. aeruginosa (46), type IV secretion in A. tume-faciens (35), exoenzyme production in Burkholderia cepacia(31), and the expression of other virulence-associated factors(2, 3, 13, 19, 37, 48, 49). However, a recent survey of gram-negative periodontal organisms by Frias et al. (18) suggestedthat this group of bacteria do not possess acyl-HSL-dependentsignaling circuits. Instead, several of these organisms, e.g., Fu-sobacterium nucleatum, Porphyromonas gingivalis, and Pre-votella intermedia, appeared to secrete a signal related to AI-2of V. harveyi. Indeed, highly conserved homologs of V. harveyiluxS have recently been identified in both gram-negative and
* Corresponding author. Mailing address: Room 540, Levy Re-search Building, Department of Biochemistry, School of Dental Med-icine, University of Pennsylvania, 4010 Locust St., Philadelphia, PA19014-6002. Phone: (215) 898-2125. Fax: (215) 898-3695. E-mail:[email protected].
7625
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

gram-positive bacteria (49), including the periodontal patho-gen P. gingivalis (11), where inactivation of luxS influenced theexpression of several genes encoding virulence factors andproteins which may be involved in the uptake of iron. This isconsistent with recent evidence suggesting that LuxS-depen-dent quorum sensing controls aspects of virulence in Esche-richia coli, Salmonella enterica serovar Typhimurium, andVibrio cholerae (47–49). Interestingly, Frias et al. (18) found noevidence of AI-2 activity in the periodontal pathogen Actinoba-cillus actinomycetemcomitans, suggesting that this organismmay not possess luxS.
A. actinomycetemcomitans is associated with a variety ofinfectious disease processes, including endocarditis, brain ab-scesses, osteomyelitis, subcutaneous abscesses, and early-onsetperiodontal disease (7, 36, 43, 52, 53), but little is known aboutthe mechanisms of A. actinomycetemcomitans pathogenesis.However, the organism produces an array of potential viru-lence factors that may contribute to pathogenesis. For exam-ple, the breakdown of the extracellular matrix and the induc-tion of bone resorption that occur in periodontitis may befacilitated by expression of collagenase (38), lipopolysaccha-ride (28), and GroEL-like proteins (20). A. actinomycetem-comitans expresses various adherence factors (34) (includingfimbriae [39]), invades human epithelial cells (33, 45), pro-duces several toxins which target various components of theimmune system, and may play a role in modulating the hostresponse by killing cells of the lymphocytic and monomyelo-cytic lineages. The best-characterized toxin is leukotoxin (29),a member of the RTX (repeats in toxin) family of gram-neg-ative bacterial toxins (15, 50).
In this study, we report that A. actinomycetemcomitans se-cretes an AI-2-like signal that stimulates light production in V.harveyi and induces the expression of leukotoxin and aperiplasmic iron-transporting protein in early-log-phase A. ac-tinomycetemcomitans cells. In addition, we show that condi-tioned broth from a recombinant E. coli strain expressing A.actinomycetemcomitans luxS complements a luxS mutation inP. gingivalis. These results show that A. actinomycetemcomitanspossesses a LuxS-dependent signal circuit and suggest thatLuxS-dependent signaling may mediate intra- and interspeciesresponses among periodontal pathogens in the human oralcavity.
MATERIALS AND METHODS
Bacterial strains and culture conditions. The bacterial strains used in thisstudy are shown in Table 1. A. actinomycetemcomitans (strain JP2) was grown inbrain heart infusion (Difco, Detroit, Mich.) supplemented with 40 mg ofNaHCO3 per liter. Cultures were maintained at 37°C in an atmosphere of 5%CO2. E. coli strains were grown in Luria-Bertani (LB) medium (1% tryptone,0.5% yeast extract, 0.5% NaCl) with aeration at 37°C. V. harveyi BB170 (sensor1�, sensor 2�) was kindly provided by B. Bassler (Princeton University) andgrown in AB medium (47) overnight at 30°C. AB medium consists of 10 mMpotassium phosphate (pH 7.0), 0.3 M NaCl, 0.05 M MgSO4, 0.2% vitamin-freeCasamino Acids (Difco), 2% glycerol, 1 mM L-arginine, 1 �g of thiamine per ml,and 0.01 �g of riboflavin per ml. For recombinant strains carrying plasmids,antibiotic selection was carried out by supplementing the appropriate mediumwith ampicillin (100 �g/ml) or chloramphenicol (34 �g/ml). P. gingivalis strainswere grown anaerobically at 37°C in Trypticase soy broth (BBL) supplementedwith 1 mg of yeast extract per ml, 5 �g of hemin per ml, and 1 �g of menadioneper ml.
Cloning of the A. actinomycetemcomitans luxS gene. A 750-bp DNA fragmentcontaining the luxS gene from A. actinomycetemcomitans JP2 was PCR amplifiedfrom genomic DNA using the primers Aa_luxS5 (5�-TAAAGCCTGCGATTTTCCTG-3�) and Aa_luxS3 (5�-CTTATTGTTTTAATAAGCTTTCGTC-3�).Primer sequences were derived from the sequence of the A. actinomycetemcomi-tans HK1651 genome (B.A. Roe, F. Z. Najar, S. Clifton, T. Ducey, L. Lewis, andD. Dyer, Actinobacillus Genome Sequencing Project, University of Oklahoma).The resulting PCR product was cloned into pGEMT-Easy (Promega) to gener-ate plasmid pGEMT750, which was subsequently modified by insertion of achloramphenicol resistance marker at the unique ClaI site within the luxS openreading frame to generate pGEMT750C. The chloramphenicol resistance genewas PCR amplified from pACYC184 (New England Biolabs) using the primersCm-3 (5�-GGCGATCGATCACGGTCACA-3�) and Cm-4 (5�-CGCGATCGATTGAGACGTTG-3�), cloned into pGEMT750, and transformed into compe-tent E. coli DH5�. Plasmid was purified from the resulting recombinant organ-ism and cleaved with ClaI. The insert was purified and ligated with pGEMT750that had been cut with ClaI, and recombinant organisms were selected forresistance to ampicillin and chloramphenicol. Plasmids pGEMT750 andpGEMT750C were subsequently transformed into competent E. coli DH5�(Gibco BRL) to generate E. coli AIS and AIS�, respectively.
To construct pGEMT1.5K, a 1.5-kbp DNA fragment containing the entire luxSgene was amplified from genomic DNA using primers lux5-1 (5�-CCATCGAAGTTCAAAGTTTG-3�) and lux3-1 (5�-CGCTCCCATCAATTACGCCTGC-3�). Primer sequences were derived from the strain HK1651 genomic sequence asdescribed above. The resulting fragment was cloned into pGEMT to producepGEMT1.5, which was further modified by the addition of the kanamycin resis-tance determinant from pUC4K into the unique ClaI site within the luxS openreading frame. To generate an isogenic luxS-deficient strain of JP2, pGEMT1.5K(which does not replicate in A. actinomycetemcomitans) was introduced into A.actinomycetemcomitans JP2 by electroporation, and colonies were selected forresistance to 25-�g of kanamycin per ml, and subsequently counterselected onmedium containing kanamycin and 100 �g of ampicillin per ml. Two populationsof clones were identified: some clones were resistant to both antibiotics, whereasothers were sensitive to ampicillin and resistant to kanamycin. To conformintegration of pGEMT1.5K into the genome, genomic DNA was isolated and
TABLE 1. Bacterial strains
Strain Reference Comments
A. actinomycetemcomitans JP2 8 Highly leukotoxicA. actinomycetemcomitans 652 8 Minimally leukotoxicA. actinomycetemcomitans ATCC 29524 Minimally leukotoxicA. actinomycetemcomitans HK890 8 Minimally leukotoxicA. actinomycetemcomitans HK905 8 Minimally leukotoxicA. actinomycetemcomitans Fambo Clinical isolate; highly leukotoxicA. actinomycetemcomitans Emory Clinical isolate; highly leukotoxicE. coli AIS This study Carries A. actinomycetemcomitans luxS on pGEM-EasyE. coli AIS� This study luxS insertional mutant of E. coli AISP. gingivalis ATCC 33277 ATCC type strainP. gingivalis PLM1 11 Isogenic luxS insertional mutant of ATCC 33277
7626 FONG ET AL. INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

analyzed by PCR using primers Aa_luxS5 and Aa_luxS3, described above. Cloneswhich had undergone gene replacement were also analyzed for AI-2 productionas described below.
AI-2 assay. The V. harveyi luminescence bioassay was performed essentially asdescribed by Surette and Bassler (47). To obtain cell-free conditioned broth forthese assays, an overnight A. actinomycetemcomitans culture was diluted 1:20 intofresh medium and incubated for 2 h (early log phase) to 7 h (late log phase) at37°C as described above. Cells were removed by centrifugation, and the resultingsupernatant was filtered through 0.2-mm-pore-size filters and used immediatelyor stored at �70°C. For the determination of V. harveyi bioluminescence, anovernight culture of V. harveyi BB170 was diluted 1:5,000 into fresh AB medium,and 90 �l of the diluted cells was added to wells on a 96-well microtiter dish.Cell-free conditioned medium was added to the diluted V. harveyi culture at a10% (vol/vol) final concentration. Positive control wells contained 10 �l ofcell-free conditioned medium from V. harveyi BB170, while negative control wellscontained 10 �l of sterile AB growth medium. The microtiter dish was shaken ina rotary shaker at 500 rpm at 30°C. Light production was measured hourly usinga Wallac (Gaithersburg, Md.) model 1450 Microbeta Plus liquid scintillationcounter in the chemiluminescence mode. The data are reported as the increasein light emission by V. harveyi BB170 exposed to the various conditioned mediaover the level of luminescence obtained for the negative control containingsterile growth medium alone.
For some experiments, cell-free conditioned media from recombinant E. colistrains containing intact (strain AIS) or inactivated (strain AIS�) A. actinomy-cetemcomitans luxS were analyzed. Strains AIS and AIS� containing pGEMT750and pGEMT750C, respectively, were grown at 37°C to mid-logarithmic to latelogarithmic phase in LB medium supplemented with 0.5% glucose, while V.harveyi BB170 was grown as described above. Cell-free conditioned media wereprepared by centrifuging the bacterial cultures at 8,000 � g, and the supernatantwas filtered through 0.2-mm-pore-size filters and stored at �70°C.
Determination of leukotoxic activity. Leukotoxin-mediated cytolysis of humanHL-60 cells was determined by trypan blue exclusion as described previously (8).Previous results had shown that intact A. actinomycetemcomitans cells are leu-kotoxic and that whole-cell cytotoxicity correlates with the level of leukotoxinpolypeptide expressed by the bacterial cell (8). HL-60 cells (12) were cultured at37°C in an atmosphere of 5% CO2 in RPMI 1640 (Gibco Laboratories) contain-ing 10% heat-inactivated fetal calf serum, penicillin G (100 �g/ml), and strep-tomycin (100 �g/ml). Prior to use, the cells were washed with RPMI 1640 toremove the antibiotics and were suspended in RPMI 1640 without antibiotics ata density of 4 � 106 cells per ml. Early-log-phase A. actinomycetemcomitans cells(optical density � 0.1 to 0.15) were exposed to cell-free conditioned broth fromA. actinomycetemcomitans cultures or from E. coli AIS or AIS� for 15 to 90 minat 37°C. The bacterial cells were then harvested, washed in RPMI 1640, andsuspended in the same medium at a density of 108 cells per ml. Bacterial cells (50�l) were mixed with HL-60 cells (50 �l) in Eppendorf tubes and incubated at37°C for 15 min. A negative control consisting of HL-60 cells without bacteriawas run for each reaction. All reactions were terminated by the addition of 100�l of 0.4% trypan blue, and surviving cells were counted using a hemocytometer.At least four fields were counted for each sample, and percent lysis was calcu-lated by dividing the number of surviving cells by the number of cells in thenegative control. Values are the averages of triplicate assays.
Enzyme-linked immunosorbent assay analysis of A. actinomycetemcomitansleukotoxin. Early-log-phase A. actinomycetemcomitans cells were incubated at37°C for 15 to 90 min in cell-free conditioned medium from E. coli strain AIS orAIS� as described above. Cells were harvested, washed in phosphate-bufferedsaline (PBS; 50 mM sodium phosphate [pH 7.5]–150 mM NaCl), and suspendedin the same buffer at a density of 108 cells per ml. Serial twofold dilutions of thecell suspension were spotted onto a nitrocellulose membrane. The membranewas incubated for 1 h at room temperature with gentle agitation in PBS con-taining 1% bovine serum albumin and then for 1 h in the same buffer containingpolyclonal leukotoxin antibody (1:1,000 dilution). Filters were washed threetimes with PBS, reacted with goat anti-immunoglobulin G–peroxidase conjugate,and developed using diaminobenzidine (0.5 mg/ml in 50 mM Tris [pH 7.5]–0.03% H2O2) as the substrate. The developed filters were scanned on a Hewlett-Packard ScanJet 6100C, and the digital images were analyzed with a MolecularDynamics personal densitometer.
RNA isolation and RT-PCR. A. actinomycetemcomitans total RNA was iso-lated using the RNeasy mini kit (Qiagen) according to the manufacturer’s in-structions. Reverse transcriptase PCR (RT-PCR) of A. actinomycetemcomitansRNA was performed using the Platinum quantitative RT-PCR Thermoscriptone-step system (Gibco BRL) as described by the manufacturer. Reverse tran-scription was routinely performed at 60°C for 30 min using the afuA2 primer (seebelow) and 10 ng of total RNA as the template. The resulting cDNA was
amplified using the primers afuA1 (5�-CTTGCCGGTCAGTTAAAAGA-3�)and afuA2 (5�-TCCTGCCTGTTCAATCAATT-3�) derived from the publishedafuI sequence (53), under the following conditions: denaturation at 94°C for 45 s,annealing at 55°C for 45 s, and elongation at 68°C for 45 s for 40 cycles, followedby extension at 72°C for 4 min. Controls without RT were included in allexperiments. Products were visualized after electrophoresis in 1% agarose gels.
For complementation of the P. gingivalis luxS knockout mutant (strain PLM1),a fresh culture of P. gingivalis PLM1 was incubated overnight with 1% filteredsupernatants of E. coli AIS or with sterile LB medium in an anaerobic chamberat 37°C. Cells were harvested, washed, and suspended at 1010 cells/ml, and totalRNA was isolated using the Totally RNA isolation kit (Ambion). Reverse tran-scription was performed in the presence of 1 �g of total RNA, 50 ng of antisenseprimer (random hexamers), 50 U of RT (Ambion), 13 U of RNase inhibitor, 10mM deoxynucleoside triphosphate, and 1� RT buffer. Annealing of primer andtemplate was carried out at 72°C for 2 min and then at 48°C for 1 h. Controlswithout RT were included in all experiments. The resulting cDNA was amplified,with each 100 �l of PCR mixture containing 1� PCR buffer, 3 �l of cDNA, 1.5mM MgCl2, 10 mM deoxynucleoside triphosphate, 100 ng of each primer (seebelow), and 2.5 U of Taq DNA polymerase. The amplification conditions weredenaturation at 94°C for 30 s, annealing at 45°C for 30 s, and elongation at 72°Cfor 2 min for 35 cycles. Primers used in these reactions were 5�-TACAAGGAGCACGCAGACAG-3� and 5�-TCCCGTGGACGATATGTAGG-3�, specificfor P. gingivalis uvrB, and 5�-ATACGGAGGAGGTGAGCGTA-3� and 5�-AGTGATGCAATGCTCTGACG-3�, specific for P. gingivalis hasF. These primerswere previously described by Chung et al. (11).
RESULTS
Secretion of an AI-2-like signal by A. actinomycetemcomitans.To determine if A. actinomycetemcomitans secretes AI-2 activ-ity, we exposed the V. harveyi reporter strain BB170 (sensor 1�,sensor 2�; kindly supplied by B. Bassler) to cell-free condi-tioned medium from a mid- to late-log-phase A. actinomyce-temcomitans culture (see Materials and Methods) or to condi-tioned medium from an overnight V. harveyi culture. As shownin Fig. 1A, luminescence of V. harveyi BB170 increased approx-imately 1,200-fold when cells were exposed to conditionedbroth from the overnight V. harveyi culture and increased ap-proximately 250-fold when they were incubated with the A.actinomycetemcomitans broth. In contrast, conditioned me-dium from E. coli DH5�, which carries a mutation in luxSrendering it inactive (49), induced very little luminescence in V.harveyi BB170. To determine if AI-2 activity varied during thegrowth of A. actinomycetemcomitans, V. harveyi luminescencewas measured after exposure of cells to conditioned mediumfrom early-, mid-, and late-log-phase A. actinomycetemcomi-tans cultures. As shown in Fig. 1B, AI-2 activity was maximal inearly and mid-log phase and deceased significantly in late logphase.
Isolation of A. actinomycetemcomitans luxS. To identify A.actinomycetemcomitans luxS, the partially completed genomeof A. actinomycetemcomitans HK1651 (Actinobacillus GenomeSequencing Project) was searched using the E. coli luxS se-quence as a probe. These searches yielded an open readingframe that encoded a polypeptide exhibiting significant se-quence similarity to LuxS proteins from gram-negative andgram-positive bacteria (Table 2). The A. actinomycetemcomi-tans sequence was most similar to LuxS from Pasteurella mul-tocida, Neisseria meningitidis, Haemophilus influenzae, and V.harveyi and exhibited significantly lower similarity to the LuxSproteins of Borrelia burgdorferi, P. gingivalis, and several gram-positive organisms. PCR amplification of A. actinomycetem-comitans JP2 genomic DNA using primers designed from thesequence identified above yielded a 750-bp product which en-coded a LuxS protein capable of synthesizing AI-2 in E. coli
VOL. 69, 2001 A. ACTINOMYCETEMCOMITANS LuxS SIGNALING 7627
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

DH5� (see below). Similar 750-bp products were also obtainedfrom PCRs using genomic DNA of A. actinomycetemcomitans652, 29524, HK890, HK905, Emory, and Fambo (data notshown). The 750-bp product obtained from JP2 genomic DNA
was cloned into pGEMT and introduced into E. coli DH5� togenerate E. coli AIS. Sequencing of the plasmid insert from E.coli AIS confirmed that it possessed the luxS open readingframe and the putative luxS promoter. This plasmid was thenfurther modified by ligating the chloramphenicol resistancedeterminant of pACYC184 into a unique ClaI site within theluxS open reading frame to inactivate the gene. The resultingE. coli strain containing the inactivated luxS was designatedAIS�. As shown in Fig. 2, conditioned broth from E. coli AISinduced V. harveyi luminescence approximately 400-fold,whereas the induction of light by medium from strain AIS�
was minimal, similar to that in the host E. coli DH5�. To-gether, these results show that A. actinomycetemcomitans luxSis necessary and sufficient for the synthesis of an extracellularsignaling molecule that is capable of inducing light productionthrough the V. harveyi AI-2 signaling system.
A. actinomycetemcomitans homologs of V. harveyi LuxP,LuxQ, and LuxO. Signal transduction in V. harveyi requires theperiplasmic protein LuxP, the sensor kinase LuxQ, the phos-phorelay protein LuxU, and the response regulator LuxO. Todetermine if A. actinomycetemcomitans possesses homologs ofthese proteins, each sequence was used as a probe to searchthe available genomic sequence database. As shown in Table 3,proteins exhibiting sequence similarity to LuxP, LuxQ, andLuxO were identified in A. actinomycetemcomitans. Consistentwith the previously reported results of Bassler et al. (4, 5), A.actinomycetemcomitans LuxP is homologous to the periplasmicribose binding protein of E. coli and LuxO exhibits similarity tothe NtrC family of response regulators (32). No homolog ofLuxU was found in A. actinomycetemcomitans.
The LuxS-dependent signal influences leukotoxin and irontransport protein expression in A. actinomycetemcomitans. Sev-eral reports have suggested that LuxS-dependent signalingcontrols the expression of virulence determinants in E. coli(44) and P. gingivalis (11). To determine if the LuxS-dependentsignal influenced the expression of leukotoxin (a member ofthe RTX family of gram-negative toxins) in A. actinomycetem-comitans JP2, early-logarithmic-phase bacteria were exposedto cell-free conditioned medium from a mid- to late-log-phaseculture or with sterile medium for 15, 30, 60, and 90 min at37°C. In fresh growth medium, the doubling time for A. acti-nomycetemcomitans under these conditions is approximately
FIG. 1. AI-2 activity secreted by A. actinomycetemcomitans. (A)AI-2 activity in cell-free conditioned medium from an overnight V.harveyi BB170 culture (Vh) and from mid-exponential-phase A. acti-nomycetemcomitans (Aa) and E. coli DH5� (DH5) cultures was de-termined by monitoring the induction of V. harveyi luminescence asdescribed in Materials and Methods. V. harveyi luminescence was de-termined after incubation of cells with the appropriate conditionedmedium for 4 h. A negative control reaction (not shown) consisted ofV. harveyi cells incubated with sterile medium. The increase in induc-tion was calculated by dividing the light production of the experimentalsamples by that of the negative control. (B) AI-2 activity is maximal inmid-exponential-phase A. actinomycetemcomitans cultures. An over-night A. actinomycetemcomitans culture was diluted into fresh medium(1:20) and harvested after incubation at 37°C for 2, 3, 5, and 7 h.Cell-free conditioned medium was prepared from each sample andanalyzed for AI-2 activity as described above. (C) Growth curve of A.actinomycetemcomitans JP2. O.D., optical density.
TABLE 2. Similarity of selected bacterial LuxS proteins
Organism with LuxS % Identity and % similarity withA. actinomycetemcomitans LuxSa
P. multocida.................................................................. 83, 88H. influenzae ................................................................. 81, 89V. harveyi....................................................................... 72, 86S. enterica serovar Typhimurium ............................... 70, 82E. coli ............................................................................ 68, 83Campylobacter jejuni .................................................... 68, 83Clostridium perfringens ................................................ 50, 66Staphylococcus aureus.................................................. 43, 62H. pylori......................................................................... 42, 61S. mutans ...................................................................... 41, 61Streptococcus pyogenes................................................. 39, 61B. burgdorferi ................................................................ 30, 52P. gingivalis ................................................................... 31, 46
a Comparisons were conducted with BLAST2 and include conservative sub-stitutions using the Blosum62 matrix (1).
7628 FONG ET AL. INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
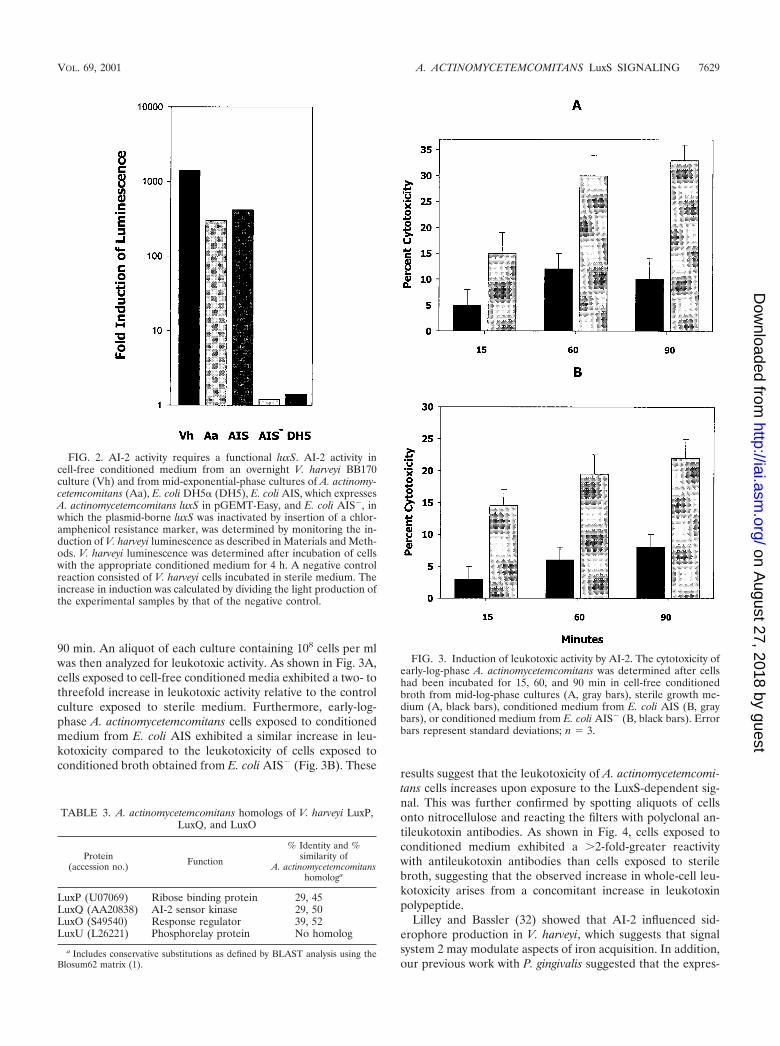
90 min. An aliquot of each culture containing 108 cells per mlwas then analyzed for leukotoxic activity. As shown in Fig. 3A,cells exposed to cell-free conditioned media exhibited a two- tothreefold increase in leukotoxic activity relative to the controlculture exposed to sterile medium. Furthermore, early-log-phase A. actinomycetemcomitans cells exposed to conditionedmedium from E. coli AIS exhibited a similar increase in leu-kotoxicity compared to the leukotoxicity of cells exposed toconditioned broth obtained from E. coli AIS� (Fig. 3B). These
results suggest that the leukotoxicity of A. actinomycetemcomi-tans cells increases upon exposure to the LuxS-dependent sig-nal. This was further confirmed by spotting aliquots of cellsonto nitrocellulose and reacting the filters with polyclonal an-tileukotoxin antibodies. As shown in Fig. 4, cells exposed toconditioned medium exhibited a �2-fold-greater reactivitywith antileukotoxin antibodies than cells exposed to sterilebroth, suggesting that the observed increase in whole-cell leu-kotoxicity arises from a concomitant increase in leukotoxinpolypeptide.
Lilley and Bassler (32) showed that AI-2 influenced sid-erophore production in V. harveyi, which suggests that signalsystem 2 may modulate aspects of iron acquisition. In addition,our previous work with P. gingivalis suggested that the expres-
FIG. 2. AI-2 activity requires a functional luxS. AI-2 activity incell-free conditioned medium from an overnight V. harveyi BB170culture (Vh) and from mid-exponential-phase cultures of A. actinomy-cetemcomitans (Aa), E. coli DH5� (DH5), E. coli AIS, which expressesA. actinomycetemcomitans luxS in pGEMT-Easy, and E. coli AIS�, inwhich the plasmid-borne luxS was inactivated by insertion of a chlor-amphenicol resistance marker, was determined by monitoring the in-duction of V. harveyi luminescence as described in Materials and Meth-ods. V. harveyi luminescence was determined after incubation of cellswith the appropriate conditioned medium for 4 h. A negative controlreaction consisted of V. harveyi cells incubated in sterile medium. Theincrease in induction was calculated by dividing the light production ofthe experimental samples by that of the negative control.
FIG. 3. Induction of leukotoxic activity by AI-2. The cytotoxicity ofearly-log-phase A. actinomycetemcomitans was determined after cellshad been incubated for 15, 60, and 90 min in cell-free conditionedbroth from mid-log-phase cultures (A, gray bars), sterile growth me-dium (A, black bars), conditioned medium from E. coli AIS (B, graybars), or conditioned medium from E. coli AIS� (B, black bars). Errorbars represent standard deviations; n � 3.
TABLE 3. A. actinomycetemcomitans homologs of V. harveyi LuxP,LuxQ, and LuxO
Protein(accession no.) Function
% Identity and %similarity of
A. actinomycetemcomitanshomologa
LuxP (U07069) Ribose binding protein 29, 45LuxQ (AA20838) AI-2 sensor kinase 29, 50LuxO (S49540) Response regulator 39, 52LuxU (L26221) Phosphorelay protein No homolog
a Includes conservative substitutions as defined by BLAST analysis using theBlosum62 matrix (1).
VOL. 69, 2001 A. ACTINOMYCETEMCOMITANS LuxS SIGNALING 7629
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

sion of genes involved in the acquisition of iron were modu-lated by LuxS-dependent signaling (11). To determine if luxSmay influence iron acquisition in A. actinomycetemcomitans,we examined the expression of AfuA, a periplasmic proteinwhich is highly related to HitA of H. influenzae (21, 51) and amajor component involved in the transport of iron by A. acti-nomycetemcomitans. As shown in Fig. 5A, the expression ofafuA, as determined by RT-PCR, was increased approximatelyeightfold when early-log-phase A. actinomycetemcomitans cellswere exposed to conditioned broth from E. coli AIS versusconditioned broth from E. coli AIS�. In contrast, the expres-sion of cdtB, encoding the cytolethal distending toxin protein Bof A. actinomycetemcomitans, was unaffected by exposure tothe conditioned broth (Fig. 5B). These results suggest thatLuxS-dependent signaling may increase the expression of leu-kotoxin, a virulence-associated protein, and afuA, involved iniron transport.
Complementation of luxS knockout mutation in P. gingivalisby A. actinomycetemcomitans AI-2. P. gingivalis and A. actino-mycetemcomitans reside in a complex oral microbial biofilm,and we have shown that both organisms possess luxS (11; thispaper). We also showed previously that inactivation of luxSinfluenced the expression of specific genes in P. gingivalis (11).To determine if AI-2 from A. actinomycetemcomitans is capa-ble of complementing the luxS mutation in P. gingivalis, weused RT-PCR to compare the expression of two luxS-regulated
genes, hasF and uvrB, in P. gingivalis cells exposed to condi-tioned medium from E. coli AIS or to sterile growth medium.These genes were chosen because they exhibited differentialbehavior in response to luxS inactivation (11), in that loss ofluxS function in P. gingivalis resulted in a decrease of uvrBexpression but increased hasF expression (Fig. 6A). Thus,complementation by A. actinomycetemcomitans AI-2 would re-quire that it reverse the opposing effects exhibited by thesegenes. As shown in Fig. 6B, exposing strain PLM1 to cell-freeconditioned medium from E. coli AIS resulted in an increase inuvrB expression relative to the control cells that were incu-bated with sterile broth (lane 1). Furthermore, the expressionof hasF was turned off in the presence of A. actinomycetem-comitans AI-2 (Fig. 6B, lane 2). Indeed, A. actinomycetemcomi-tans AI-2 appeared to influence hasF expression to a greaterdegree than the endogenous signal in wild-type P. gingivalis33277 cells (Fig. 6A). This may be due to increased signaldosage arising from the expression of luxS from a multicopyplasmid in E. coli AIS. Thus, A. actinomycetemcomitans luxSsynthesizes a signal that is capable of modulating the expres-sion of luxS-regulated genes in P. gingivalis, suggesting that thebasic structure of the signal molecule and the mechanism fortransducing signal information are conserved in these two peri-odontal organisms.
Generation of an isogenic luxS-deficient A. actinomycetem-comitans strain. To facilitate further study of the role of luxS-dependent signaling of A. actinomycetemcomitans, an isogenicluxS-deficient mutant was constructed by transforming strainJP2 with pGEMT1.5K and selecting for kanamycin-resistantcolonies. Five resistant clones were selected for further anal-ysis by PCR using the luxS primers Aa_luxS5 and Aa_luxS3. Asshown in Fig. 7, three clones exhibited amplification productsof 750 and 2,000 bp and were resistant to both kanamycin and
FIG. 4. Induction of leukotoxin protein by AI-2. Early-log-phase A.actinomycetemcomitans cells were incubated for 60 min in cell-freeconditioned medium from E. coli AIS (gray bars) or AIS� (black bars)and spotted onto nitrocellulose. The filter was washed with PBS con-taining 1% bovine serum albumin and reacted with polyclonal antileu-kotoxin antibodies. Immunoreactivity was determined by measuringthe relative intensity of spots on the developed filter using a MolecularDynamics personal densitometer.
FIG. 5. AI-2 stimulates the expression of afuA in A. actinomyce-temcomitans. RT-PCR was carried out with primers that were specificfor A. actinomycetemcomitans afuA (A) or cdtB (B) using 10 ng of totalRNA from early-log-phase A. actinomycetemcomitans cells that hadbeen exposed to conditioned medium from E. coli AIS (lane 1) orAIS� (lane 2). A 10-�l aliquot of the RT-PCR mixture was electro-phoresed in 1% agarose. Lanes M, DNA size markers.
7630 FONG ET AL. INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

ampicillin, suggesting that they represent strains arising fromsingle Campbell-type recombination events. The remainingtwo kanamycin-resistant clones were sensitive to ampicillin andexhibited only the 2,000-bp PCR product, indicating thatgenomic luxS was replaced by the inactivated copy of the genefrom pGEMT1.5K. These results were confirmed by Southernblotting of EcoRI-digested genomic DNA using the 750-bpluxS fragment as a probe (not shown). In addition, the induc-tion of V. harveyi BB170 luminescence by conditioned culturemedium from the knockout strains was reduced by 90% rela-tive to wild-type JP2 (Fig. 7B), suggesting that the mutant isincapable of synthesizing AI-2.
DISCUSSION
The LuxS-dependent quorum-sensing circuit was originallyidentified in the marine organism V. harveyi, but recent studieshave shown that many gram-negative and gram-positive bac-teria possess homologs of the luxS gene (3, 11, 18, 24, 44, 49).Indeed, a recent survey of periodontal pathogens by Frias et al.(18) showed that P. intermedia, F. nucleatum, and P. gingivalissecrete AI-2-like signals that are capable of inducing light in V.harveyi. The luxS gene has also been recently cloned from P.gingivalis and appears to be important for regulating aspects ofiron acquisition by this organism (11). Interestingly, Frias et al.(18) did not detect AI-2 in A. actinomycetemcomitans strains.Our results clearly show that A. actinomycetemcomitans pos-sesses luxS and secretes AI-2. Conditioned broth from early-
FIG. 6. A. actinomycetemcomitans AI-2 complements a luxS knock-out mutation in P. gingivalis. The wild-type P. gingivalis strain ATCC33277 and its isogenic mutant (PLM1) in which luxS has been inacti-vated have been described (11). (A) RT-PCR using RNA isolated fromP. gingivalis ATCC 33277 and PLM1 showed that inactivation of P.gingivalis luxS results in reduced expression of uvrB (lane 1) and in-creased expression of hasF (lane 2). (B) P. gingivalis PLM1 was sub-sequently incubated with cell-free conditioned broth from E. coli AISor with sterile growth medium (sM) as described in Materials andMethods, and RT-PCRs were carried out using the uvrB- and hasF-specific primers used for panel A. Exposure to A. actinomycetemcomi-tans AI-2 induced the expression of uvrB (lane 1) and turned off hasFexpression (lane 2). The fimA gene, which is not regulated by P.gingivalis luxS, was unaffected by exposure to A. actinomycetemcomi-tans AI-2 (lane 3).
FIG. 7. Isogenic luxS-deficient A. actinomycetemcomitans JP2. (A)Five kanamycin-resistant colonies of strain JP2 that had been trans-formed with pGEMT1.5K were analyzed by PCR using the primersAa_luxS5 and Aa_luxS3 as described in Materials and Methods. Threeclones (lanes 1, 2, and 5) contained the two amplification productspredicted to result from single recombination of genomic luxS withpGEMT1.5K. The remaining clones (lanes 3 and 4) generated thesingle PCR product predicted to arise from the replacement ofgenomic luxS with the inactivated gene from pGEMT1.5K. Lane M,DNA size markers. (B) Conditioned culture medium from one of theseclones (JP2lx�) was also analyzed and exhibited significantly reducedinduction of luminescence from the V. harveyi BB170 reporter straincompared to conditioned medium from strain JP2. The induction oflight by JP2 was arbitrarily assigned a value of 1.0.
VOL. 69, 2001 A. ACTINOMYCETEMCOMITANS LuxS SIGNALING 7631
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

and mid-exponential-phase A. actinomycetemcomitans culturescontained the greatest AI-2 activity, as determined by moni-toring the production of light by V. harveyi BB170. Activitydecreased significantly in late log and stationary phases. Thus,the expression of AI-2 by A. actinomycetemcomitans is similarto that reported for Helicobacter pylori (24) and S. entericaserovar Typhimurium (48, 49). Furthermore, luxS was presentin six additional A. actinomycetemcomitans strains. Thesestrains represented both highly leukotoxic and minimally leu-kotoxic organisms (Table 1) and were representative of sero-types b (JP2 and ATCC 29524) and c (652). This suggests thatthe presence of the signaling circuit is independent of leuko-toxic phenotype and occurs in at least two of six A. actinomy-cetemcomitans serotypes. However, in light of the inability ofFrias et al. (18) to detect AI-2 activity in a serotype c strain(ATCC 33384), a more extensive analysis will be necessary todetermine the distribution of the LuxS-dependent signal sys-tem in A. actinomycetemcomitans.
The A. actinomycetemcomitans LuxS polypeptide is similarin sequence to LuxS proteins from a variety of organisms (e.g.,P. multocida, H. influenzae, E. coli, and V. harveyi) but exhibitssignificantly lower similarity to LuxS of other oral organisms(e.g., Streptococcus mutans and P. gingivalis). In addition, ho-mologs of V. harveyi LuxP, LuxQ, and LuxO were identified inA. actinomycetemcomitans, but these proteins did not exhibitthe high degree of sequence similarity that was observed be-tween the respective LuxS proteins. Furthermore, no homologof LuxU was identified in searches of the A. actinomycetem-comitans genome. Although we cannot exclude the possibilitythat our inability to identify LuxU arises from the incompletenature of the A. actinomycetemcomitans genome sequence,searches of the completed E. coli, S. enterica serovar Typhi, andP. gingivalis genomes also failed to identify homologs of LuxUbut did identify proteins exhibiting similarity to LuxP, LuxQ,and LuxO. This suggests that mechanistic differences may oc-cur in the pathways of AI-2 signal transduction in V. harveyiand A. actinomycetemcomitans. Interestingly, the putative A.actinomycetemcomitans sensor protein corresponding to LuxQis similar to a family of tripartite sensor kinases which arecapable of catalyzing autophosphorylation and two intramo-lecular phosphotransfer reactions. Thus, the A. actinomycetem-comitans sensor kinase may encompass the function of bothLuxQ (autophosphorylation and first phosphotransfer) andLuxU (phosphorelay), raising the possibility that AI-2 signaltransduction in A. actinomycetemcomitans may not require anindependent phosphorelay protein corresponding to LuxU.Studies are under way to investigate the role of the LuxQ-homologous polypeptide in AI-2 signal transduction by A. ac-tinomycetemcomitans.
The extent of the cellular functions regulated by LuxS-de-pendent signaling are not known. In V. harveyi, AI-2 controlslight production by the lux operon, the expression of genesinvolved in siderophore production and regulates aspects ofcolony morphology (32). In addition, Sperandio et al. (44)recently showed that AI-2 regulates the expression of the locusof enterocyte effacement (LEE) operon in E. coli O157:H7,suggesting that LuxS may play a role in modulating virulence.Our results show that the expression of the A. actinomycetem-comitans leukotoxin is influenced by AI-2 and increases byseveral fold in early-log-phase cells after exposure to condi-
tioned medium from recombinant E. coli cultures expressingluxS. The leukotoxin is an RTX pore-forming toxin that in-duces apoptosis (at low concentration) or cell lysis (at highconcentration) in a defined set of human leukocytes (26, 29,30). In addition, several recent studies have shown that A.actinomycetemcomitans strains which express high levels of leu-kotoxin are associated with severe forms of early-onset peri-odontal diseases (9, 22, 23), suggesting that the toxin is impor-tant for A. actinomycetemcomitans pathogenesis. Since A.actinomycetemcomitans thrives in a complex biofilm that existsin the gingival pocket, it is conceivable that the LuxS-depen-dent induction of leukotoxin by AI-2 may play an importantrole in the expression of virulence in vivo. Experiments areunder way to determine if other potential virulence factors ofA. actinomycetemcomitans (e.g., cytolethal distending toxin [29,40, 41] and adherence factors [25, 34]) are regulated by LuxS-dependent signaling.
Signal system 2 may also control the acquisition of iron. Forexample, AI-2 regulates the expression of siderophore produc-tion in V. harveyi (32) and influences the expression of hemRand rgpA in P. gingivalis (11), both of which encode proteinsthat may be involved in the acquisition of hemin (27, 42). In A.actinomycetemcomitans, an important mode of iron acquisitionand transport involves afuA, which encodes a 35-kDa periplas-mic protein related to H. influenzae HitA that is coexpressedwith outer membrane proteins corresponding to HitBC (21,51). Together, these polypeptides function to transport ironacross the outer membrane and periplasm of A. actinomyce-temcomitans. Our results show that expression of afuA is dra-matically increased upon exposure to AI-2, suggesting thatLuxS-dependent signaling may stimulate iron acquisition in A.actinomycetemcomitans.
The widespread distribution of luxS and the observationsthat AI-2 from diverse organisms induce luminescence in V.harveyi has lead to the hypothesis that signal system 2 tran-scends species barriers and may function to report total bac-terial cell density and the metabolic potential of the environ-ment (3, 49). Indeed, such a role for LuxS-dependent signalingmay be particularly relevant for organisms in the oral cavity,where there exist many distinct ecological niches inhabited byspecific populations of bacteria and where populational shiftsin this complex community contribute to the onset and/orprogression of disease. Inherent to this hypothesis is that spe-cific luxS-regulated genes in a given species should respond toa heterologous AI-2 that is produced by another organism.Until now, cross-species signaling has been demonstrated onlyby the induction of luminescence in V. harveyi, and it is notknown whether interspecies signaling is widespread or whetherV. harveyi is simply promiscuous in responding to AI-2 signals.Our studies show that AI-2-mediated cross talk occurs betweenA. actinomycetemcomitans and P. gingivalis and that specificluxS-regulated genes involved in diverse physiologic processesin P. gingivalis respond to a heterologous signal generated by A.actinomycetemcomitans. These results suggest that the generalstructure of AI-2 may be conserved among these two organ-isms and they support the hypothesis that LuxS-dependentsignaling may function in interspecies communication. How-ever, since we have thus far examined relatively few targetgenes modulated by AI-2, we cannot exclude the possibilitythat LuxS-dependent signaling may also mediate species-spe-
7632 FONG ET AL. INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

cific responses and that some luxS-regulated genes of P. gingi-valis may not respond to a heterologous signal or may exhibita differential response to cognate and heterologous signals.The identification and analysis of additional luxS-regulatedtargets will address these issues, and such experiments arebeing carried out.
Our results also show that AI-2 signal concentration is max-imal during mid-exponential-phase growth of A. actinomyce-temcomitans and decreases significantly as cells approach sta-tionary phase. This is consistent with previous results reportedfor E. coli, S. enterica serovar Typhimurium, and H. pylori (25,48, 49). Thus, LuxS-dependent signaling may function at rela-tively low cell density in A. actinomycetemcomitans comparedto other quorum-sensing bacteria. The basis for this discrep-ancy among quorum-sensing bacteria has not been fully ex-plained. However, Surette and Bassler have suggested thatpathogenic E. coli and S. enterica serovar Typhimurium maynever reach stationary phase in vivo (48). A similar situationmay occur with A. actinomycetemcomitans in multispecies oralbiofilms, where host antimicrobial activities, the constant flowof saliva, and competition for nutrients among the variousorganisms may prevent A. actinomycetemcomitans from attain-ing high cell density. Under these adverse conditions, theLuxS-dependent signal system may be adapted to function atlower cell density.
In summary, A. actinomycetemcomitans expresses luxS andsecretes a signal related to AI-2 of V. harveyi. LuxS-dependentsignaling was shown to induce the expression of leukotoxin anda periplasmic transport protein that may be involved in theacquisition of iron by A. actinomycetemcomitans. The A. acti-nomycetemcomitans signal also complemented a luxS mutationin P. gingivalis, suggesting that the LuxS-dependent signal cir-cuit of A. actinomycetemcomitans may induce both intra- andinterspecies responses in the mixed-species microbial commu-nities that exist in the oral cavity.
ACKNOWLEDGMENTS
We thank Bonnie L. Bassler for kindly providing the V. harveyiBB170 reporter strain.
This work was supported by Public Health Service grants DE10729and DE12505 from the National Institutes of Health.
REFERENCES
1. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller,and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generationof protein database search programs. Nucleic Acids Res. 25:3389–3402.
2. Bainton, N. J., B. W. Bycroft, S. Chhabra, P. Stead, L. Gledhill, P. J. Hill,C. E. D. Rees, M. K. Winson, G. P. C. Salmond, G. S. A. B. Stewart, and P.Williams. 1992. A general role for the lux autoinducer in bacterial cellsignaling: control of antibiotic biosynthesis in Erwinia. Gene 116:87–91.
3. Bassler, B. L. 1999. How bacteria talk to each other: regulation of geneexpression by quorum sensing. Curr. Opin. Microbiol. 2:582–587.
4. Bassler, B. L., M. Wright, R. E. Showalter, and M. R. Silverman. 1993.Intercellular signalling in Vibrio harveyi: sequence and function of genesregulating expression of luminescence. Mol. Microbiol. 9:773–786.
5. Bassler, B. L., M. Wright, and M. R. Silverman. 1994. Multiple signallingsystems controlling expression of luminescence in Vibrio harveyi: sequenceand function of genes encoding a second sensory pathway. Mol. Microbiol.13:273–286.
6. Bassler, B. L., M. Wright, and M. R. Silverman. 1994. Sequence and functionof LuxO, a negative regulator of luminescence in Vibrio harveyi. Mol. Mi-crobiol. 12:403–412.
7. Block, P. J., A. C. Fox, C. Yoran, and A. J. Kaltman. 1973. Actinobacillusactinomycetemcomitans endocarditis: report of a case and review of theliterature. Am. J. Med. Sci. 276:387–392.
8. Brogan, J. M., E. T. Lally, K. Poulsen, M. Kilian, and D. R. Demuth. 1994.Regulation of Actinobacillus actinomycetemcomitans expression: analysis of
the promoter regions of leukotoxic and minimally leukotoxic strains. Infect.Immun. 62:501–508.
9. Bueno, L. C., M. P. Mayer, and J. M. DiRienzo. 1998. Relationship betweenconversion of localized juvenile periodontitis-susceptible children fromhealth to disease and Actinobacillus actinomycetemcomitans leukotoxin pro-moter structure. J. Periodontol. 69:998–1007.
10. Cao, J., and E. I. Meighen. 1989. Purification and structural identification ofan autoinducer for the luminescence system of Vibrio harveyi. J. Biol. Chem.264:21670–21676.
11. Chung, W., Y. Park, R. J. Lamont, R. McNab, B. Barbieri, and D. R.Demuth. 2001. A signaling system in Porphyromonas gingivalis based on aLuxS protein. J. Bacteriol. 183:3903–3909.
12. Collins, S., and R. C. Gallo. 1980. Human myeloid leukemic cells in vitro(HL-60): growth, differentiation and comparison to Friend cells, p. 477–501.In G. B. Rossi (ed.), In vivo and in vitro erythropoiesis, the Friend system.Elsevier/North Holland Biomedical Press, New York, N.Y.
13. DeKievit, T. R., and B. H. Iglewski. 2000. Bacterial quorum sensing inpathogenic relationships. Infect. Immun. 68:4839–4849.
14. Engebrecht, J., K. Nealson, and M. Silverman. 1983. Bacterial biolumines-cence: isolation and genetic analysis of functions from Vibrio fischeri. Cell32:773–781.
15. Fives-Taylor, P. M., D. H. Meyer, K. P. Mintz, and C. Brissette. 1999.Virulence factors of Actinobacillus actinomycetemcomitans. Periodontology20:136–167.
16. Freeman, J. A., and B. L. Bassler. 1999. A genetic analysis of the function ofLuxO, a two-component response regulator involved in quorum sensing inVibrio harveyi. Mol. Microbiol. 31:665–677.
17. Freeman, J. A., and B. L. Bassler. 1999. Sequence and function of LuxU: atwo-component phosphorelay protein that regulates quorum sensing inVibrio harveyi. J. Bacteriol. 181:899–906.
18. Frias, J., E. Olle, and M. Alsina. 2001. Periodontal pathogens producequorum sensing signal molecules. Infect. Immun. 69:3431–3434.
19. Fuqua, C., S. C. Winans, and, and E. P. Greenberg. 1996. Census andconsensus in bacterial ecosystems: the LuxR-LuxI family of quorum sensingtranscriptional regulators. Annu. Rev. Microbiol. 50:727–751.
20. Gouhlen, F., A. Hafezi, V.-J. Uitto, D. Hinode, R. Nakamura, D. Grenier, andD. Mayrand. 1998. Subcellular localization and cytotoxic activity of theGroEL-like protein isolated from Actinobacillus actinomycetemcomitans. In-fect. Immun. 66:5307–5313.
21. Graber, K. R., L. M. Smoot, and L. A. Actis. 1998. Expression of iron bindingproteins and hemin binding activity in the dental pathogen Actinobacillusactinomycetemcomitans. FEMS Microbiol. Lett. 163:135–142.
22. Guthmiller, J. M., E. T. Lally, and J. Korostoff. 2001. Beyond the specificplaque hypothesis: are highly leukotoxic strains of Actinobacillus actinomy-cetemcomitans a paradigm for periodontal pathogenesis? Crit. Rev. OralBiol. Med. 12:116–124.
23. Haraszthy, V. I., G. Hariharan, E. M. Tinoco, J. R. Cortelli, E. T. Lally, E.Davis, and J. J. Zambon. 2000. Evidence for the role of highly leukotoxicActinobacillus actinomycetemcomitans in the pathogenesis of localized juve-nile and other forms of early-onset periodontitis. J. Periodontol. 71:912–922.
24. Joyce, E. A., B. L. Bassler, and A. Wright. 2000. Evidence for a signalingsystem in Helicobacter pylori: detection of a luxS-encoded autoinducer. J.Bacteriol. 182:3638–3643.
25. Kachlaney, S. C., P. J. Planet, R. Desalle, D. H. Fine, D. H. Figurski, andJ. B. Kaplan. 2001. flp-1, the first representative of a new pilin gene sub-family, is required for non-specific adherence of Actinobacillus actinomyce-temcomitans. Mol. Microbiol. 40:542–554.
26. Karakelian, D., D. Lear, E. T. Lally, and J. C. Tanaka. 1998. Characteriza-tion of Actinobacillus actinomycetemcomitans leukotoxin pore formation inHL60 cells. Biochim. Biophys. Acta 1406:175–187.
27. Karunakaran, T., T. Madden, and H. Kuramitsu. 1997. Isolation and char-acterization of a hemin-regulated gene, hemR, from Porphyromonas gingiva-lis. J. Bacteriol. 179:1898–1908.
28. Kato, T., K. Honma, A. Yamanaka, T. Miura, and K. Okuda. 2000. Heter-ogeneity in the immune response to serotype b LPS of Actinobacillus acti-nomycetemcomitans in inbred strains of mice. FEMS Immunol. Med. Micro-biol. 28:67–70.
29. Lally, E. T., I. R. Kieba, A. Sato, C. L. Green, J. Rosenbloom, J. Korostoff,J. F. Wang, B. S. Shenker, S. Ortlepp, M. K. Robinson, and P. C. Billings.1997. RTX toxins recognize a 2 integrin on the surface of human targetcells. J. Biol. Chem. 272:30463–30469.
30. Lear, J. D., D. Karakelian, U. Furblur, E. T. Lally, and J. C. Tanaka. 2000.Conformational studies of Actinobacillus actinomycetemcomitans leukotoxin:partial denaturation enhances toxicity. Biochim. Biophys. Acta 1476:350–362.
31. Lewenza, S., B. Conway, E. P. Greenberg, and P. A. Sokol. 1999. Quorumsensing in Burkholderia cepacia: identification of the luxRI homologs cepRI.J. Bacteriol. 181:748–756.
32. Lilley, B. N., and B. L. Bassler. 2000. Regulation of quorum sensing in Vibrioharveyi by LuxO and sigma-54. Mol. Microbiol. 36:940–954.
33. Meyer, D. H., J. E. Lippman, and P. M. Fives-Taylor. 1994. Invasion ofepithelial cells by Actinobacillus actinomycetemcomitans: a dynamic multistep
VOL. 69, 2001 A. ACTINOMYCETEMCOMITANS LuxS SIGNALING 7633
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

process. Infect. Immun. 64:2988–2997.34. Meyer, D. H., and P. M. Fives-Taylor. 1994. Characteristics of adherence of
Actinobacillus actinomycetemcomitans to epithelial cells. Infect. Immun. 62:928–935.
35. Oger, P., K. S. Kim, R. L. Sackett, K. R. Piper, and S. K. Farrand. 1998.Octopine-type Ti plasmids code for mannopine-inducible dominant-negativeallele of TraR, the quorum-sensing activator that regulates Ti plasmid con-jugal transfer. Mol. Microbiol. 27:277–288.
36. Page, M. I., and E. O. King. 1966. Infection due to Actinobacillus actinomy-cetemcomitans and Haemophilus aphrophilus. N. Engl. J. Med. 275:181–188.
37. Passador, L., J. M. Cook, M. J. Gambello, L. Rust, and B. H. Iglewski. 1993.Expression of Pseudomonas aeruginosa virulence genes requires cell to cellcommunication. Science 260:1127–1130.
38. Robertson, P. B., M. Lantz, P. T. Marucha, K. S. Kornman, C. L. Trummel,and S. C. Holt. 1982. Collagenolytic activity associated with Bacteroidesspecies and Actinobacillus actinomycetemcomitans. J. Periodontal Res. 17:275–283.
39. Rosan, B., J. Slots, R. J. Lamont, M. A. Listgarten, and G. M. Nelson. 1988.Actinobacillus actinomycetemcomitans fimbriae. Oral Microbiol. Immunol.3:58–63.
40. Shenker, B. J., T. McKay, S. Datar, M. Miller, R. Chowden, and D. R.Demuth. 1999. Actinobacillus actinomycetemcomitans immunosuppressiveprotein is a member of the family of cytolethal distending toxins capable ofcausing G2 arrest in human T cells. J. Immunol. 162:4773–4780.
41. Shenker, B. S., R. H. Hoffmaster, T. McKay, and D. R. Demuth. 2000.Expression of cytolethal distending toxin (cdt) operon in Actinobacillus ac-tinomycetemcomitans: evidence that the CdtB protein is responsible for G2arrest of the cell cycle in human T cells. J. Immunol. 165:2612–2618.
42. Shi, Y., D. B. Ratnayake, K. Okamoto, N. Abe, K. Yamamoto, and K.Hakayama. 1999. Genetic analysis of proteolysis, hemoglobin binding, andhemagglutination of Porphyromonas gingivalis. Construction of mutants witha combination of rgpA, rgpB, kgp and hagA. J. Biol. Chem. 274:17955–17960.
43. Slots, J., H. S. Reynolds, and R. J. Genco. 1980. Actinobacillus actinomyce-
temcomitans in human periodontal disease: a cross-sectional microbiologicalinvestigation. Infect. Immun. 29:1013–1020.
44. Sperandio, V., J. L. Mellies, W. Nguyen, S. Shin, and J. B. Kaper. 1999.Quorum sensing controls expression of the type III secretion gene transcrip-tion and protein secretion on enterohemorrhagic and enteropathogenicEscherichia coli. Proc. Natl. Acad. Sci. USA 96:15196–15201.
45. Sreenivasan, P. K., D. H. Meyer, and P. M. Fives-Taylor. 1993. Require-ments for invasion of epithelial cells by Actinobacillus actinomycetemcomi-tans. Infect. Immun. 61:1239–1245.
46. Storey, D. G., E. E. Ujack, H. R. Rabin, and I. Mitchell. 1998. Pseudomonasaeruginosa lasR transcription correlates with the transcription of lasA, lasBand toxA in chronic lung infections associated with cystic fibrosis. Infect.Immun. 66:2521–2528.
47. Surette, M. G., and B. L. Bassler. 1998. Quorum sensing in Escherichia coliand Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 95:7046–7050.
48. Surette, M. G., and B. L. Bassler. 1999. Regulation of autoinducer produc-tion in Salmonella typhimurium. Mol. Microbiol. 31:585–595.
49. Surette, M. G., M. B. Miller, and B. L. Bassler. 1999. Quorum sensing inEscherichia coli, Salmonella typhimurium, and Vibrio harveyi: a new family ofgenes responsible for autoinducer production. Proc. Natl. Acad. Sci. USA96:1639–1644.
50. Tervahartiala, B., V.-J. Uitto, K. Kari, and T. Laasko. 1989. Outer mem-brane vesicles and leukotoxic activity of Actinobacillus actinomycetemcomi-tans from subjects with different periodontal status. J. Dent. Res. 97:33–42.
51. Willemsen, P. T., I. Vulto, M. Boxem, and J. de Graaff. 1997. Characteriza-tion of a periplasmic protein involved in iron utilization of Actinobacillusactinomycetemcomitans. J. Bacteriol. 179:4949–4952.
52. Zambon, J. J. 1985. Actinobacillus actinomycetemcomitans in human peri-odontal disease. J. Clin. Periodontol. 12:1–20.
53. Zambon, J. J., J. Slots, and R. J. Genco. 1983. Serology of oral Actinobacillusactinomycetemcomitans and serotype distribution in human periodontal dis-ease. Infect. Immun. 41:19–27.
Editor: E. I. Tuomanen
7634 FONG ET AL. INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from


















