
Integrative Biology Dynamic Article · PDF fileWith the advent of high-throughput biology, ......
9
724 Integr. Biol., 2011, 3, 724–732 This journal is c The Royal Society of Chemistry 2011 Cite this: Integr. Biol., 2011, 3, 724–732 Dynamic modeling and analysis of cancer cellular network motifsw Mathieu Cloutier a and Edwin Wang* ab Received 28th January 2011, Accepted 5th May 2011 DOI: 10.1039/c0ib00145g With the advent of high-throughput biology, we now routinely scan cells and organisms at practically all levels, from genome to protein, metabolism, signaling and other cellular functions. This methodology allowed biological studies to move from a reductionist approach, such as isolation of specific pathways and mechanisms, to a more integrative approach, where biological systems are seen as a network of interconnected components that provide specific outputs and functions in response to stimuli. Recent literature on biological networks demonstrates two important concepts that we will consider in this review: (i) cellular pathways are highly interconnected and should not be studied separately, but as a network; (ii) simple, recurrent feedback motifs within the network can produce very specific functions that favor their modular use. The first theme differs from the traditional approach in biology because it provides a framework (i.e., the network view) in which large datasets are analyzed with an unbiased view. The second theme (feedback motifs) shows the importance of locally analyzing the dynamic properties of biological networks in order to better understand their functionality. We will review these themes with examples from cell signaling networks, gene regulatory networks and metabolic pathways. The deregulation of cellular networks (metabolism, signaling etc.) is involved in cancer, but the size of the networks and resulting non-linear behavior do not allow for intuitive reasoning. In that context, we argue that the qualitative classification of the ‘building blocs’ of biological networks (i.e. the motifs) in terms of dynamics and functionality will be critical to improve our understanding of cancer biology and rationalize the wealth of information from high-throughput experiments. From the examples highlighted in this review, it is clear that dynamic feedback motifs can be used to provide a unified view of various cellular processes involved in cancer and this will be critical for future research on personalized and predictive cancer therapies. Introduction Cancer is a complex and multifactorial disease in which gene mutations in signaling, 1–3 metabolic 4–6 and gene regulation 7,8 networks accumulate in a dynamic process, leading to deregulation of cell death and/or proliferation. Most of the components of the underlying biological networks have been a Computational Chemistry and Bioinformatics Group, Biotechnology Research Institute, National Research Council Canada, 6100 Royalmount Avenue, Montreal, Quebec H4P 2R2, Canada. E-mail: [email protected]; Tel: (514) 496-0914 b Center for Bioinformatics, McGill University, Montreal, Canada w Electronic supplementary information (ESI) available. See DOI: 10.1039/c0ib00145g Insight, innovation, integration This article reviews feedback motifs, and their associated dynamic properties, in the context of cancer research. Feedback motifs provide a rational explanation to widespread concepts in cancer biology such as homeostasis, adaptation or bistability. However, we often observe a gap between the identification of a specific biological mechanism and the rigorous explanation of the underlying dynamical system. This review will thus provide some examples where the mechanisms are specific to cancer biological processes, but always keeping in mind that the underlying dynamical systems are generic and can be analyzed in order to retrieve their properties. We thus report that the importance of feedback motifs applies to multiple levels of biological organization, from gene expression to signalling and metabolism. Integrative Biology Dynamic Article Links www.rsc.org/ibiology CRITICAL REVIEW Downloaded by Universite de Montreal on 19 July 2011 Published on 15 June 2011 on http://pubs.rsc.org | doi:10.1039/C0IB00145G View Online
-
Upload
dinhnguyet -
Category
Documents
-
view
215 -
download
1
Transcript of Integrative Biology Dynamic Article · PDF fileWith the advent of high-throughput biology, ......

724 Integr. Biol., 2011, 3, 724–732 This journal is c The Royal Society of Chemistry 2011
Cite this: Integr. Biol., 2011, 3, 724–732
Dynamic modeling and analysis of cancer cellular network motifsw
Mathieu Cloutieraand Edwin Wang*
ab
Received 28th January 2011, Accepted 5th May 2011
DOI: 10.1039/c0ib00145g
With the advent of high-throughput biology, we now routinely scan cells and organisms at
practically all levels, from genome to protein, metabolism, signaling and other cellular functions.
This methodology allowed biological studies to move from a reductionist approach, such as
isolation of specific pathways and mechanisms, to a more integrative approach, where biological
systems are seen as a network of interconnected components that provide specific outputs and
functions in response to stimuli. Recent literature on biological networks demonstrates two
important concepts that we will consider in this review: (i) cellular pathways are highly
interconnected and should not be studied separately, but as a network; (ii) simple, recurrent
feedback motifs within the network can produce very specific functions that favor their modular
use. The first theme differs from the traditional approach in biology because it provides a
framework (i.e., the network view) in which large datasets are analyzed with an unbiased view.
The second theme (feedback motifs) shows the importance of locally analyzing the dynamic
properties of biological networks in order to better understand their functionality. We will review
these themes with examples from cell signaling networks, gene regulatory networks and metabolic
pathways. The deregulation of cellular networks (metabolism, signaling etc.) is involved in cancer,
but the size of the networks and resulting non-linear behavior do not allow for intuitive
reasoning. In that context, we argue that the qualitative classification of the ‘building blocs’ of
biological networks (i.e. the motifs) in terms of dynamics and functionality will be critical to
improve our understanding of cancer biology and rationalize the wealth of information from
high-throughput experiments. From the examples highlighted in this review, it is clear that
dynamic feedback motifs can be used to provide a unified view of various cellular processes
involved in cancer and this will be critical for future research on personalized and predictive
cancer therapies.
Introduction
Cancer is a complex and multifactorial disease in which gene
mutations in signaling,1–3 metabolic4–6 and gene regulation7,8
networks accumulate in a dynamic process, leading to
deregulation of cell death and/or proliferation. Most of the
components of the underlying biological networks have been
a Computational Chemistry and Bioinformatics Group,Biotechnology Research Institute, National Research CouncilCanada, 6100 Royalmount Avenue, Montreal, Quebec H4P 2R2,Canada. E-mail: [email protected];Tel: (514) 496-0914
bCenter for Bioinformatics, McGill University, Montreal, Canadaw Electronic supplementary information (ESI) available. See DOI:10.1039/c0ib00145g
Insight, innovation, integration
This article reviews feedback motifs, and their associated
dynamic properties, in the context of cancer research. Feedback
motifs provide a rational explanation to widespread concepts in
cancer biology such as homeostasis, adaptation or bistability.
However, we often observe a gap between the identification of a
specific biological mechanism and the rigorous explanation of
the underlying dynamical system. This review will thus provide
some examples where the mechanisms are specific to cancer
biological processes, but always keeping in mind that the
underlying dynamical systems are generic and can be analyzed
in order to retrieve their properties. We thus report that the
importance of feedback motifs applies to multiple levels of
biological organization, from gene expression to signalling and
metabolism.
Integrative Biology Dynamic Article Links
www.rsc.org/ibiology CRITICAL REVIEW
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online

This journal is c The Royal Society of Chemistry 2011 Integr. Biol., 2011, 3, 724–732 725
studied for decades using traditional approaches in biology
such as reductionism, linear logic and conceptual models
(i.e., a drawing or qualitative representation of the biological
mechanism). For the most studied biological networks,
including metabolic pathways, we now have access to complete
mapping of the system in online databases such as KEGG
(Kyoto Encyclopedia of Genes and Genomes: www.kegg.jp).
Similarly, signaling pathways relevant to cancer are well
determined and publicly available. The information gathered
in KEGG mostly comes from traditional biology studies; the
pathways were studied separately, and their topologies were
determined through careful experimental investigation of the
biochemistry of cell extracts. The advent of high-throughput
biology, starting with the sequencing of the human genome,
showed that reductionism gives a far from complete picture.
Biological function, and thus dysfunction in cancer, is based
on networks, nonlinearities and dynamic processes.9 Within
this context, the traditional approach is still relevant because it
provides valuable information on the components of the
system (i.e., the bottom up approach). However, it is now
recognized that cancer biology must be complemented with
networks10,11 and systems thinking.12–15
We will briefly present the network and systems biology
framework with emphasis on dynamical analysis of the network
motifs. The mathematical frameworks for such studies will be
briefly presented with focus on Ordinary Differential Equation
(ODE) models. With this perspective in mind, we will then
review works on biological networks relevant to cancer,
showing that the local properties induced by network and
feedback motifs are critical for cancer development. The
relevance of the network and systems approach for predictive
and personalized medicine will also be highlighted.
Dynamic networks for cancer biology
From pathways to dynamic network motifs
As presented in Fig. 1, we will review recent works in network
and systems biology, with emphasis on the dynamic properties
of such networks, and the emergence of functional properties
from simple feedback motifs.16,17 Interestingly, emergent features,
such as adaptation, can be observed in many biological
networks, ranging from metabolism18 to signaling,19 gene
regulation20 and physiology.21 Even more interesting is the
fact that the underlying principles behind such properties are
strikingly similar, even though they cover the complete range
of biological organization from gene expression to organ
physiology. A better understanding of how these functional
properties emerge from dynamic interactions is at the core of
the systems biology approach and will provide essential tools
for predictive modeling in medicine.22,23 Fig. 1 highlights the
changes in thinking that recently occurred in biology, whereas
functional properties are not linked specifically to one pathway
or gene, but to a network of dynamically interacting components
(Fig. 1B). However, due to the complexity of these networks,
we argue that reductionism is still relevant, mostly because the
local properties of network motifs are responsible for the
emergence of functionality from simple biochemical interactions.
Thus, from a complex and large network, it is possible to
extract ‘functional units’ composed of just a few nodes and to
analyze their dynamical properties and relevance to the
biological problems under scrutiny. Fig. 1C shows a simple
3-node network motif19 and possible functions, including
adaptation16,19 or bistability.16
Mathematical modeling of dynamic biological networks
Biological networks, such as signaling networks, metabolic
networks and gene regulatory networks are generally too large
and complex for their dynamic behavior to be determined by
intuitive reasoning. The same can be said for disease-related
pathways (such as the ones found in KEGG), as these are
composed of parts from the aforementioned networks. This
implies that even if we have computational and experimental
tools to estimate the local properties of biological systems,
such as the binding affinity of a drug for a target, this does not
lead to a comprehensive understanding of the effects of the
drug at the systems level.24 The most well-known examples we
can relate to this problem come from the field of metabolic
engineering, where micro-organisms are genetically manipulated
in order to improve the productivity of a specific molecule.
Many studies report failures in which the overexpression of a
locally sensitive enzyme (i.e., phosphofructokinase in glycolysis)
did not lead to increased flux in the pathway,25 an observation
that is reproduced by dynamic modeling at the systems level.26
It is now recognized that modeling and systems analysis is
necessary for full appreciation of the sensitivity of biological
systems to perturbations27 and that this analysis relates well to
drug development and novel therapies for complex diseases
such as cancer.28 Thus, an important goal in future medical
research will be to understand how large-scale networks
dynamically respond to local perturbations induced by drugs
and treatments.
Our review of the dynamic modeling of biological networks
and its relevance to cancer systems biology will be principally
centered on the mathematical modeling by ODEs and further
functional analyses. ODEs represent the most common frame-
work for the modeling of physical systems in science and
engineering and are also used widely in biological sciences.29,30
Depending on the specifics of the biological problem, other
modeling frameworks can be used; for example, Boolean
networks31 and Bayesian networks32 are widely used for fast
computation and data analysis of gene regulatory networks.
The ODE approach, despite a few drawbacks such as the
identification of kinetic parameters and computing time, can
be applied and scaled to a wide range of problems. This
facilitates the drawing of general conclusions based on the
observation of biological function and organization at
different levels. The approach has been used for practically
all cellular processes, including gene regulation networks,8,33
cancer-related signaling pathways,34–37 energy metabolism in
cancer38,39 and tumor physiology.40 The ODE approach
can also be complemented in various ways to account for
stochasticity41 or discontinuities42 (i.e., hybrid systems) when
the system contains a small number of molecules or discrete
events. Moreover, the ODE framework is also amenable to
integration of cellular processes that are of different natures.
For example, Kelly et al. built an ODE model to link tumor
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online

726 Integr. Biol., 2011, 3, 724–732 This journal is c The Royal Society of Chemistry 2011
glycolysis to HIF signaling;35 other modeling reports have also
linked metabolism to energy signaling.38 Some studies using
ODE models go even further and integrate cancer-related
processes at various levels, accounting for cell cycle, drug
penetration, tumor physiology and metabolism.41 As we want
to emphasize the recurrent dynamic properties in biological
networks of various scales (gene, signaling and metabolism) in
this review, we will concentrate on the ODE representations of
these network motifs.
Integrating networks and feedback motifs in cancer
cells
Biological features of cancer network motifs and their
implications
Cancer signaling network motifs have certain properties and
lead to implications in molecular mechanisms of tumorigenesis
and practical applications such as prediction of prognosis
and the clinical outcomes of cancer patients. A large-scale
characterization of network motifs in cancer signaling showed
that positive signaling regulatory network motifs are preferentially
used by cancer driving mutating oncogenes, in contrast,
negative signaling regulatory network motifs are preferentially
used by methylated genes and tumor suppressors in cancer
cells.1 These observations are in agreement with the fact that
oncogenes often get mutated and then increase their biochemical
activities. Such a mutated oncogene in a positive feedback loop
might amplify the output signal and therefore consistently trigger
the downstream signals in cancer cells. On the other hand,
methylated genes and tumor suppressor mutations cause the
genes to lose or decrease their biochemical activities. Such a
gene in a negative signaling regulatory loop might also amplify
the cellular signals and trigger the downstream cellular signals
in cancer cells.
Combining these insights with the features of the degradation
of mRNAs or proteins in network motifs led to more under-
standing of the molecular mechanisms of the tumorigenesis
and even practical applications in cancer. In general, in a gene
regulatory motif, the regulator has a short half-life.43 Such a
feature decreases the response time of the network motifs to
the signal cues from environment or neighbouring cells. This
insight triggered the further examination of mRNA or protein
degradation in these motifs. microRNA (miRNA), a short
RNA sequence, which binds to 30UTR region of the genes, has
a function of blocking mRNA transcription or inducing
mRNA/protein degradation. Interestingly, microRNAs are
preferentially regulating positive signaling regulatory network
motifs but avoid regulating negative signaling regulatory
network motifs.2 miRNA is processed by the miRNAmachinery.
Therefore, these observations suggest that low activity of
the miRNA machinery could produce less miRNAs, which
could reduce the degradation of the gene products in positive
regulatory network motifs, and finally make the corresponding
signals amplified. This result is similar to that of oncogene
mutations in the positive regulatory network motifs, and
then leads to cancer signal amplification. Low expression of
key genes of the miRNA processing machinery could thus
promote cancer progression and metastasis. By examining
the expression of Dicer, which is the key gene of the
miRNA processing machinery, in two breast cancer patient
cohorts containing 581 samples,44,45 we found that the
expression of Dicer is indeed lower in metastatic samples than
the non-metastatic samples. Moreover, low expression of
Dicer is able to predict poor outcomes for breast cancer
patients.
Fig. 1 Representations of cell signaling networks. (A) Traditional, pathway specific view with receptors activating linear signaling cascades
leading to TFs activation, gene expression and protein. (B) Network view where biological information from high-throughput experiments is used
to build an unbiased network where components are not necessarily linked to one pathway. Also shown in (B) is the possible feedback loops within
gene expression networks. C: Systems analysis of biological networks components. Here a 3 nodes network is analyzed with ODEs, showing
2 recurrent cases: adaptation and bistable switching (further analysis of these motifs is provided in Fig. 2).
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online

This journal is c The Royal Society of Chemistry 2011 Integr. Biol., 2011, 3, 724–732 727
Protein ubiquitination preferentially occurs in positive
signaling regulatory network motifs, however, these motifs have
functions of predominately inducing or positively regulating
apoptosis, a major component in cancer signaling.46 Therefore,
higher activities of protein degradation in these network
motifs could block apoptosis in cells, and then promote cancer
progression and metastasis. Indeed, the high expression level of
the genes for the 26S proteasome is significantly correlated with
tumor progression and metastasis.46 Furthermore, the expression
of the 26S proteasome gene set predicts the clinical outcome of
breast cancer patients.46 These observations suggest that the
molecular mechanisms of mRNA or protein degradation are in
general highly active in cancer cells. This insight has implications
for the development of cancer treatments and prognostic markers
could focus on the machinery of the molecular mechanisms of
mRNA or protein degradation.
Cancer network motifs tend to form clusters as hotspots
in the human signaling network47 and even form a larger
sub-network or a cancer signaling map. A cancer signaling
map has been constructed by mapping the cancer mutation
data derived from large-scale cancer genome sequencing onto
a human signaling network.1 Further network analysis identified a
network module containing B50 genes which are highly
interconnected and enriched with tumor suppressors and cell
cycle genes. This network module (common cancer signaling
module) is frequently used by many cancer samples regardless
of cancer types such that at least one gene from this network
module gets mutated cross B600 cancer samples studied.1
This result highlights that the common cancer signaling
network module is critical to cancer signaling for many
different cancer types. Furthermore, this insight led to the
development of an algorithm for finding high-quality cancer
Fig. 2 Feedback principles and their application within biological network motifs. The network motifs are modeled by ODE according to
principles presented in recent works.18,19,59,74
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online

728 Integr. Biol., 2011, 3, 724–732 This journal is c The Royal Society of Chemistry 2011
biomarkers, focusing on cancer hallmarks’ gene expression
profiles.48 Moreover, each identified network prognostic
module contains a set of cancer mutating genes and their
interacting protein partners, both of which represent the same
biological process for a cancer hallmark.48 These modules
have high predicting accuracy (B90%) and high robustness
as validated in more than 1300 breast cancer patients.
Taken together, cancer network motifs and modules exhibit
special network properties and have mechanistic and practical
implications in cancer. Therefore, it will be critical to further
investigate how functionality arises (and is lost) in these
cancer-related motifs and modules and the following text will
provide some thought on how to use ODE models to that end.
Importance of feedback interactions
As shown above, positive and negative network motifs are
prevalent in cancer cells. The notion that feedback is important in
biology has been put forward in control theory49 and can be
traced back to the origins of cybernetics.50 The two major
concepts in this regard are positive and negative feedback.
Positive feedback, in which a molecule, cell or organism
promotes its own production or replication, is a fundamental
principle of biological evolution and organization, i.e., all
forms of life are self-replicating systems. Positive feedback
principles have many implications for cancer, the first being
that cancer cells are better at self-replication and are thus
favored in a selective environment.51 However, positive feed-
back can also be traced to the internal functioning of cancer
networks,52,53 and we will return to this point later. Negative
feedback is an important mechanism to organize and control
biological processes so that exponential growth is avoided16,18
or noise is reduced.54 Feedback in biological systems is able to
induce complex, dynamic behavior;16,55,56 this will not be
observed from the static, steady-state response of the system,
but rather from the temporal response to a perturbation of the
system. This has implication for experimental design, as the
typical dose-response experiments in biology do not account
for the dynamical aspects of the problem, and so experiments
should be designed accordingly.29,36,37,53
Functionality arising from simple, recurrent motifs and feedback
rules
The analysis of feedback motifs in biology allows for
identification of simple, yet rigorous, explanations for complex
dynamic behavior. In this regard, many observations can be
drawn on the functional properties of these network motifs
and have been reviewed elsewhere.55–57,16,20,24 In this review,
we will touch upon some of the possible functional properties
that can arise from feedback interactions in a 3-node network
motif (see Fig. 2); these motifs were built with current
knowledge and frameworks.19,20 Previous studies have only
highlighted specific aspects of feedback motifs, such as
adaptation.19 In Fig. 2, we want to emphasize that 3-node
networks are able to summarize simple and recurrent feedback
concepts in biology. Some common topologies are shown and
the recurrent mechanisms we show can be summarized as
follows.
� Node A (blue) mediates the input signal (cyan) and
communicates information to the rest of the network, e.g., a
membrane receptor or signaling kinase.
� Node C (red) is the output node from which a certain
dynamic response is expected, e.g., a TF in gene regulation, a
specific gene expression level in signaling or the production of
a molecule in a metabolic pathway.
� Node B (green) is the ‘buffer node’,19,20 which, depending
on the topology, corrects, delays or amplifies the response of
node C.
From this perspective, the question we ask is how the
topology and dynamics of the 3-node motif produces a certain
shape in the output node (C) for a specific input signal (I). In
Fig. 2, we highlight some of most common examples drawn
from various cellular processes. Careful dynamical analysis of
these networks motifs will be extremely important if we are
interested in understanding the behavior of normal and cancer
cells at the systems level. Thus, we will review these network
motifs, their properties and their relevance to cancer biological
networks. Note that Fig. 2 shows generic examples of feedback
motifs while many specific examples will be slightly more
complicated. For example, we show negative and positive
feedback loops containing 3 nodes, while the actual feedback
loops in a specific cellular process might be hidden in more
complex cascades consisting of more than 3 nodes. Apart from
the obvious delay that would result from the longer paths, the
basic principles remain essentially the same. The following
subsections will concentrate on the 6 network motifs presented
in Fig. 2 and their implications for cancer biology at various
levels, including gene regulation, signaling and metabolic
networks. One recurring theme in the subsequent analysis will
be the response of the motifs after a certain input ‘I’ is applied.
In terms of experimental investigations, this input is generally
an extracellular molecule (ligand, drug etc.) for which we can
decide the concentration profile.
A complete description of the network motifs and
computational methods is presented in Supplementary
Material 1 (SM1). The objective of this review is not to
highlight all the available computational methods to build
and calibrate ODE models. The examples shown in Fig. 2 are
thus intentionally generic and will be analyzed with emphasis
on their recurrence (even if only qualitatively) at various
levels of biological organization. The issue of parameter
calibration and network determination is also briefly discussed
in SM1 and interesting examples can be found in the
literature.1,14,22,36,37,58,59 The ODEs models of the network
motifs were implemented in the Matlab computing environment
(The MathWorks Inc., Natick, MA, USA) using the Systems
Biology toolbox.60
Negative feedback: adaptive response
The first row in Fig. 2A shows the adaptive response that
occurs for the integral negative feedback motif. In this study,
the concept of ‘integral feedback’ refers to the well-known
control theory principle of integrating the output error over
time and using this information to correct the process. More
specifically in Fig. 2A, the output is the level of node C and its
‘error’ is the deviation from 0. As seen in Fig. 2A (and from
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online

This journal is c The Royal Society of Chemistry 2011 Integr. Biol., 2011, 3, 724–732 729
the equations in SM1), node B accumulates at a rate propor-
tional to signal C, the result being that the level of B is,
mathematically speaking, the integral of C with respect to
time. The interested reader is referred to previous works
on the subject,18 where a more elaborate development on this
equivalence between control theory and biological regulation
was presented.
The concept of integral negative feedback is observed at
practically all levels of biological organization and is an important
part of the adaptive response in biological systems.16,18–21,57,61 As
shown in Fig. 2A, if the dynamic response of node B is slower than
A–C, some time is available for C to accumulate. However, any
change in C is eventually corrected by the accumulation of B, and
the system returns to its original value. The result is that a
continuous signal (I) is transformed to a transient signal (C),
which has been observed in signaling cascades57,61 and gene
regulation.20 Recently, this configuration has been studied
extensively and has been shown to be one of two possible
ways to achieve adaptation.19 As this sort of transient
behavior is observed in signaling cascades related to cancer,57,61
its deregulation would have oncogenic properties, especially if
a growth signal that should be temporal (i.e., shaped as the C
node response) instead became continuous. In that regard,
oncogenic potential might not lie within a specific gene, but
rather in the disruption of a negative feedback motif in the
network. Interestingly, it should be noted that integral feedback
by a buffer molecule has also been identified in glycolysis,18
which has implications for the adaptive response of glycolysis
to perturbations. Because glycolysis and energy metabolism
are deregulated in many cancers,4,5 a more complete under-
standing of the underlying regulation structures will be crucial
in establishing which enzymes are the most promising
therapeutic targets.
Negative feedforward loop motif: robust response
Recently, there has been substantial interest in the structure
and function of the ‘incoherent feedforward loop’ motif in
biological networks.20 The term ‘incoherent’ comes from the
fact that the buffer node B (Fig. 2A, second motif) seems to
work in opposition to the signal flow between A and C (i.e., it
tries to shut down the signal). However, as has been reported
in many studies, this motif is recurrent57 and produces specific
responses, such as fold-change detection62 and noise attenuation.63
If node B is relatively slow to accumulate, adaptation will be
observed19 in a similar vein as the negative feedback loop.
However, an interesting property of the negative feedforward
loop is its potential to act faster because the signal from A
arrives to B and C at the same time. Shown in Fig. 2A (second
motif) is a situation in which node B accumulates in the same
dynamic range as A. Thus, the changes in C, which are
induced by A, are rapidly compensated by B, and the output
node C is limited to a narrow range of values (i.e., the motif
filters out noise from the input).
Interestingly, because a rapid response and negative regulation
is required from node B, we can expect miRNAs to be good
candidates to perform such a function in gene regulation
networks; this has been verified.64 Considering that miRNAs
mutations are involved in many cancers2 we can safely assume
that a better delineation of the regulation motifs in which
these mutations occur will be critical. Additionally, a negative
feedforward mechanism has been reported in a modeling
study of brain energy regulation;18 energy demand in
neurons was signaled to nearby astrocytes, which reacted by
providing energy substrates to neurons, thereby potentially
reducing noise in energy levels. Mutation of enzymes involved
in energy metabolism are found in a significant proportion
of gliomas;4 thus, a good understanding of the underlying
regulation motifs in brain metabolism will be important,
especially as many drugs also target energy metabolism in
cancer.5,6
Positive feedback motifs: the robust switch
Fig. 2B shows the positive feedback motif in which node C,
once activated by A, can keep itself active through a loop with
node B. If the feedback is strong enough, the loop between B
and C can remain active even after the input signal is
completely removed, thus making the switch irreversible. This
will transform the transient input signal into a continuous one.
Based on the same principle, the switch can also be reversible,16
only operate for a specific input range and shut down when the
input is removed. Accordingly, it is important to distinguish
between exclusive and non-exclusive switches. The first case
presented in Fig. 2B is the non-exclusive loop, in which nodes
B and C are activated together. Replacing the double positive
feedback with double negative feedback (Fig. 2B, second
motif) produces exclusive switching. More specifically, the
motif shown here is the toggle switch,16 whereas the system
can be switched ‘on’ (i.e. higher concentration of C) or ‘off’
(lower concentration) by transient pulses of a signal around an
‘operating point’ (shown here with I = 0.5).
Many examples of such mechanism have been described for
cell signaling65,66 and gene regulation.67 Cell division and cell
cycle are also regulated by reversible positive feedback loops,68
which allow switching between critical steps with more
robustness (i.e. the ‘all-or-none’ response) compared to, for
example, a direct response to a growth signal.16,24 However,
this robust response from the positive feedback motif comes at
a price; a reversible switch for normal cellular growth might
become deregulated by oncogenic mutations and then easily
become irreversible, leading to continuous growth and
uncontrolled proliferation. Recent examples of uncontrolled
cell proliferation due to positive feedback gone wrong can be
found52,53 and it is clear that the concept of positive feedback
will become more important as we unravel the complexities of
cancer-related networks. The experimental and modeling
approaches used to identify such structures53,68 should thus
be carefully studied and applied to other cellular processes.
At the metabolic level, examples of positive feedback are
much less common, but some examples can be found in oxidative
metabolism, such as reactive oxygen species can increase their own
production in aging cells through positive feedback,69 which could
potentially have implications for cancer biology. Again, instead of
focusing on specific genes, the identification of positive feedback
loops and their components in biological networks will improve
the process of mining the wealth of experimental data on
cancer-related mutations.1,14,34,52,68
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online

730 Integr. Biol., 2011, 3, 724–732 This journal is c The Royal Society of Chemistry 2011
Delays and cascades
Fig. 2C shows a slightly different case: reactions from A to C
now include a cascade. If the cascade is direct (A - B - C)
and the transition between states is sharp,16,20 the response
node C can be a temporal transition of the input, which has
implications mostly for differentiation and developmental
processes.56 If we add negative feedback in the cascade
(C - B - A - C, second motif in Fig. 2C), the structure
becomes prone to oscillations, although the parameters of the
system could be such that the loop is fast enough to be stable
(as shown in the first panel in Fig. 2). Oscillations caused by
negative feedback with delay are quite common in biological
systems and are observed in many processes, including cell
cycle,70 metabolism,18,71 signaling72 and gene expression.58
This seemingly unstable behavior might be seen as an undesirable
effect of a feedback loop gone wrong (i.e., we intuitively expect
the feedback to work as shown in first panel in Fig. 2).
However, the oscillations can be desirable for circadian
rhythms, the synchronization of populations or the transient
expression of certain genes. With regard to cancer, many
examples are found, with the most important being the
negative feedback regulation of p53 via Mdm2, which can
generate pulsed responses (i.e. damped oscillations) to radiation
stress.58,73,74 The advantage of generating oscillations in p53
concentration is not clear yet, but it could help to repair DNA
damage if there is not a continuous exposure of the cellular
environment to this protein. Again, a better understanding of
the dynamic regulation of p53 and its oscillations will be
crucial for cancer research.75 A very interesting observation
concerning p53 oscillations is that the system seems to be
tunable depending on the amount of damage imposed. Moreover,
in this system, cancer cells react differently compared to
normal cells,73–75 with a critical difference being the development
of continuous oscillations (as shown in Fig. 2) instead of
damped oscillations. Although p53 and its interacting neighbors
are well-known therapeutic targets in cancer therapy, it might
be impossible to design therapies without a better understanding
of the dynamics of the system, and, in that regard, it is clear
that the analysis of the underlying dynamic network motifs is
critical.75 Finally, it is also known that metabolic pathways
can oscillate due to negative feedback, with glycolysis (under
anaerobic conditions) being one of the many examples. The
implication of glycolytic deregulation in cancer has been
understood since the pioneering work of Warburg,76 and
many studies now integrate energy metabolism into the global
picture of cancer biology,77 which has direct consequences for
drug development.78 The analysis of energy metabolism in
terms of feedback motifs has been performed in a theoretical
study;16 it will be important to assess the stability of these
metabolic feedback motifs in the presence of cancer mutations.
Implications for future research
As we can see from these recent works on biological feedback
motifs and their implications for cancer, the ‘local’ dynamical
properties in networks are the functional units on which larger
networks are built. In that context, it is clear that systems
biology and network approaches are not just about large-scale
data, but also about careful integration of feedback functions.
For most biological processes involved in cancer, we now have
access to a wealth of data, to a point where it is hard to draw
clear conclusions. A major problem is that the data are
context-dependent22,36,53 and do not always take into account
the dynamic behavior of the system. Therefore, we emphasized
that dynamic network motifs analysis provides a framework,
with which it becomes possible to relate seemingly unrelated
observations. We thus emphasized the possible links between
signaling, gene regulation and metabolic networks in the sense
that they share common functional units that are deregulated
in various cancers.
Thus, a major endeavor for future research will be the
integration of ‘omics’ data within the framework of dynamical
systems.79 However, the size of biological networks, often in
the range of a few hundred to a few thousand components,
makes it difficult to rapidly identify interesting targets,
especially when experimental studies are required. Therefore,
when studying a certain biological process related to cancer, a
first step should be to look for specific functional motifs, such
as the ones highlighted in this review, and to determine if they
are significantly involved in the specific process that is under
scrutiny. From this basic, preliminary understanding of the
system, it becomes easier to develop a rigorous experimental
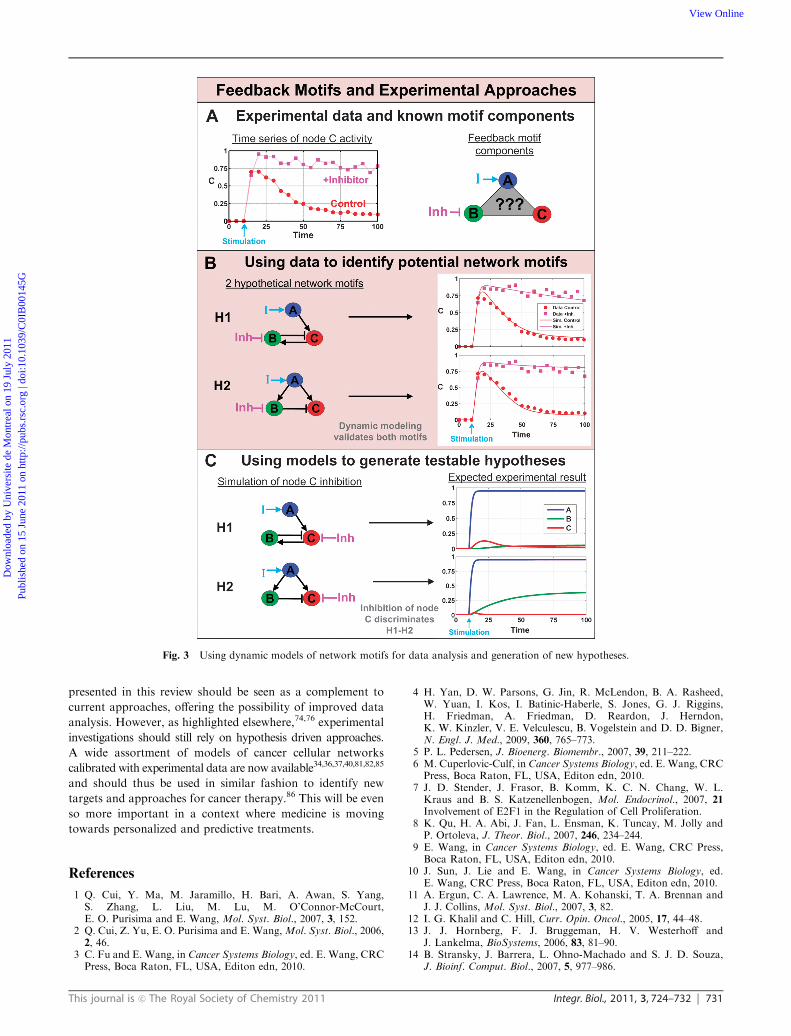
design.59,80 As a synthetic example of this, Fig. 3 shows how
time series data can be used to construct and analyze dynamic
network motifs. First, we consider a case where the dynamic
response of a system (i.e. node C in our example) is measured
in normal conditions (red dots in Fig. 3A) and in the presence
of an inhibitor (purple dots in Fig. 3A), whereas in this specific
case, we know that the inhibitor affects node B. The major
observation is a loss of adaptive capacity for node C in the
presence of the inhibitor. Even though the profile shown here
is a hypothetical example, it does follow along with known
examples of adaptive response in signaling pathways.34,81,82
From this information, we can hypothesize that a negative
feedback (H1 in Fig. 3B) or feedforward loop (H2) is present
in the system, with node B acting as the buffer.19 As shown in
Fig. 3B, models for both motifs can be calibrated to reproduce
the experimental data (see parameters values in SM1). At this
point, it would thus be impossible to further discriminate
between the two motifs. However, using the predictive capacity
of ODE models, we can simulate the effect of an inhibitor of
node C, which reveals different responses for the two feedback
motifs (see Fig. 3C). More specifically, the major difference to
be expected will be in the response of node B in the presence of
the inhibitor, whereas it is active in H2 and inactive in H1.
This type of approach, where models are constructed from
experimental results and then used for hypotheses generation,
will speed up the experimental investigations with minimal
assumptions. The analysis of an ‘atlas’ of feedback motifs was
recently shown to be very effective for the study of develop-
mental processes83 and we argue that cancer research will
benefit from the use of this framework as well. A recent study
also showed how Bayesian modeling can be used to infer
pathway topology from time profile data.84 In that context,
and as shown in Fig. 3, ODE modeling could produce
additional insights and predictions. It is thus important to
highlight that the study of the dynamics in network motifs as
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online
This journal is c The Royal Society of Chemistry 2011 Integr. Biol., 2011, 3, 724–732 731
presented in this review should be seen as a complement to
current approaches, offering the possibility of improved data
analysis. However, as highlighted elsewhere,74,76 experimental
investigations should still rely on hypothesis driven approaches.
A wide assortment of models of cancer cellular networks
calibrated with experimental data are now available34,36,37,40,81,82,85
and should thus be used in similar fashion to identify new
targets and approaches for cancer therapy.86 This will be even
so more important in a context where medicine is moving
towards personalized and predictive treatments.
References
1 Q. Cui, Y. Ma, M. Jaramillo, H. Bari, A. Awan, S. Yang,S. Zhang, L. Liu, M. Lu, M. O’Connor-McCourt,E. O. Purisima and E. Wang, Mol. Syst. Biol., 2007, 3, 152.
2 Q. Cui, Z. Yu, E. O. Purisima and E. Wang,Mol. Syst. Biol., 2006,2, 46.
3 C. Fu and E. Wang, in Cancer Systems Biology, ed. E. Wang, CRCPress, Boca Raton, FL, USA, Editon edn, 2010.
4 H. Yan, D. W. Parsons, G. Jin, R. McLendon, B. A. Rasheed,W. Yuan, I. Kos, I. Batinic-Haberle, S. Jones, G. J. Riggins,H. Friedman, A. Friedman, D. Reardon, J. Herndon,K. W. Kinzler, V. E. Velculescu, B. Vogelstein and D. D. Bigner,N. Engl. J. Med., 2009, 360, 765–773.
5 P. L. Pedersen, J. Bioenerg. Biomembr., 2007, 39, 211–222.6 M. Cuperlovic-Culf, in Cancer Systems Biology, ed. E. Wang, CRCPress, Boca Raton, FL, USA, Editon edn, 2010.
7 J. D. Stender, J. Frasor, B. Komm, K. C. N. Chang, W. L.Kraus and B. S. Katzenellenbogen, Mol. Endocrinol., 2007, 21
Involvement of E2F1 in the Regulation of Cell Proliferation.8 K. Qu, H. A. Abi, J. Fan, L. Ensman, K. Tuncay, M. Jolly andP. Ortoleva, J. Theor. Biol., 2007, 246, 234–244.
9 E. Wang, in Cancer Systems Biology, ed. E. Wang, CRC Press,Boca Raton, FL, USA, Editon edn, 2010.
10 J. Sun, J. Lie and E. Wang, in Cancer Systems Biology, ed.E. Wang, CRC Press, Boca Raton, FL, USA, Editon edn, 2010.
11 A. Ergun, C. A. Lawrence, M. A. Kohanski, T. A. Brennan andJ. J. Collins, Mol. Syst. Biol., 2007, 3, 82.
12 I. G. Khalil and C. Hill, Curr. Opin. Oncol., 2005, 17, 44–48.13 J. J. Hornberg, F. J. Bruggeman, H. V. Westerhoff and
J. Lankelma, BioSystems, 2006, 83, 81–90.14 B. Stransky, J. Barrera, L. Ohno-Machado and S. J. D. Souza,
J. Bioinf. Comput. Biol., 2007, 5, 977–986.
Fig. 3 Using dynamic models of network motifs for data analysis and generation of new hypotheses.
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online

732 Integr. Biol., 2011, 3, 724–732 This journal is c The Royal Society of Chemistry 2011
15 E. Wang, A. E. Lenferink and M. O’Connor-McCourt, Cell. Mol.Life Sci., 2007, 64, 1752–1762.
16 J. J. Tyson, K. C. Chen and B. Novak, Curr. Opin. Cell Biol., 2003,15, 221–231.
17 L. H. Hartwell, J. J. Hopfield, S. Leibler and A. W. Murray,Nature, 1999, 402, C47–C52.
18 M. Cloutier and P. Wellstead, J. R. Soc. Interface, 2010, 7, 651–665.19 W. Ma, A. Trusina, H. El-samad, W. A. Lim and C. Tang, Cell,
2009, 138, 760–773.20 S. Mangan and U. Alon, Proc. Natl. Acad. Sci. U. S. A., 2003, 100,
11980–11985.21 H. El-Samad, J. P. Goff and M. Khammash, J. Theor. Biol., 2002,
214.22 K. A. Janes, J. G. Albeck, S. Gaudet, P. D. Sorger,
D. A. Lauffenburger and M. B. Yaffe, Science, 2005, 310, 1646–1653.23 B. Ribba, T. Colin and S. Schnell, Theor. Biol. Med. Modell., 2006,
3, 7.24 J. J. Tyson, K. C. Chen and B. Novak, Nat. Rev. Mol. Cell Biol.,
2001, 2, 908–916.25 S. Thomas and D. A. Fell, Eur. J. Biochem., 1998, 258, 956–967.26 M. Cascante, L. Boros and J. Boren, in Handbook of Neuro-
chemistry and Molecular Neurobiology, ed. A. Aea, Springer,Orangeburg, NY, USA, 2007, Editon edn, pp. 861–875.
27 P. Morandini, Plant Sci., 2009, 176, 441–451.28 M. Cascante, L. Beros, B. Comin-Anduix, P. Atauri, J. J. Centelles
and P. Lee, Nat. Biotechnol., 2002, 20, 243–249.29 J. W. Haefner, Modelling Biological Systems: Principles and
Applications, Chapman & Hall, New York, 1996.30 R. Heinrich and S. Schuster, The Regulation of Cellular Systems,
Chapman & Hall, New York, 1996.31 S. A. Kauffman, J. Theor. Biol., 1974, 22, 437–467.32 M. Zou and S. Conzen, Bioinformatics, 2005, 21, 71–79.33 A. Polynikis, S. J. Hogan and B. M. di, Journal of Theoretical
Biology, 2009.34 R. J. Orton, M. E. Adriaens, A. Gormand, O. E. Sturm, W. Kolch
and D. R. Gilbert, BMC Syst. Biol., 2009, 3, 100.35 C. Kelly, K. Smallbone and M. Brady, J. Theor. Biol., 2008, 254,
508–513.36 W. W. Chen, B. Schoeberl, P. J. Jasper, M. Niepel, U. B. Nielsen,
D. A. Lauffenburger and P. D. Sorger,Molecular Systems Biology,2008, 5, 239.
37 M. R. Birtwistle, M. Hatakeyama, N. Yumoto, B. A. Ogunnaike,J. B. Hoek and B. N. Kholodenko, Mol. Syst. Biol., 2007, 3, 144.
38 M. Cloutier, in Cancer Systems Biology, ed. E. Wang, CRC Press,Boca Raton, FL, USA, Editon edn, 2010.
39 R. Venkatasubramanian, M. A. Henson and N. S. Forbes,J. Theor. Biol., 2006, 242, 440–453.
40 R. Venkatasubramanian, M. A. Henson and N. S. Forbes,J. Theor. Biol., 2008, 253, 98–117.
41 D. T. Gillespie, J. Phys. Chem., 1977, 81, 2340–2361.42 B. Karasozen, H. Oktem and M. Kahraman, J. Process Control,
2009, 19, 1257–1264.43 E. Wang and E. O. Purisima, Trends Genet., 2005, 21, 492–495.44 H. Y. Chang, D. S. A. Nuyten, J. B. Sneddon, T. Hastie,
R. Tibshirani, T. SArlie, H. Dai, Y. D. He, L. J. van’t Veer,H. Bartelink, M. van de Rijn, P. O. Brown and M. J. van de Vijver,Proc. Natl. Acad. Sci. U. S. A., 2005, 102, 3738–3743.
45 Y. Wang, J. G. M. Klijn, Y. Zhang, A. M. Sieuwerts, M. P. Look,F. Yang, D. Talantov, M. Timmermans, M. E. Meijer-van Gelder,J. Yu, T. Jatkoe, E. M. J. J. Berns, D. Atkins and J. A. Foekens,The Lancet, 2005, 365, 671–679.
46 C. Fu, J. Li and E. Wang, Mol. BioSyst., 2009, 5, 1809–1816.47 A. Awan, H. Bari, F. Yan, S. Mokin, S. Yang, S. Chowdhury,
Q. Cui, Z. Yu, E. O. Purisima and E. Wang, IET Syst. Biol., 2007,1, 292–297.
48 J. Li, A. E. Lenferink, Y. Deng, C. Cantin, Q. Cui, E. O. Purisima,M. D. O’Connor-McCourt and E. Wang, Nature Communications,2010, 1, 34.
49 D. S. Riggs, Control Theory and Physiological Feedback Mechanisms,R.E. Krieger Pub. Co., 1976.
50 N. Weiner, Cybernetics, or Control and Communication in theAnimal and the Machine, John Wiley and Sons Inc., New York,1948.
51 T. S. Deisboeck and I. D. Couzing, BioEssays, 2009, 31, 190–197.52 B. D. Aguda, Y. Kim, M. G. Piper-Hunter, A. Friedman and
C. B. Marsh, Proc. Natl. Acad. Sci. U. S. A., 2009, 105,19678–19683.
53 D. Kim, O. Rath, W. Kolch and K. H. Cho, Oncogene, 2007, 26,4571–4579.
54 H. H. McAdams and A. Arkin, Trends Genet., 1999, 15, 65–69.55 O. Brandmann and T. Meyer, Science, 2008, 322, 390–395.56 B. N. Kholodenko, Nat. Rev. Mol. Cell Biol., 2006, 7, 165–176.57 R. Milo, S. Schen-Orr, S. Itzkovitz, N. Kashtan, D. Chklovski and
U. Alon, Science, 2002, 298, 824–827.58 R. L. Bar-Or, R. Maya, L. A. Segel, U. Alon, A. J. Levine and
M. Oren, Proceedings of the National Academy of Science, 2000, 97,11250–11255.
59 U. Alon, Nat. Rev. Genet., 2007, 8, 450–461.60 H. Schmidt and M. Jirstrand, Bioinformatics, 2006, 22, 514–515.61 M. Behar, N. Hao, H. G. Dohlman and T. C. Elston, Biophys. J.,
2007, 93, 806–821.62 L. Goentoro, O. Shoval, M. W. Kirschner and U. Alon, Mol. Cell,
2009, 36, 894–899.63 B. S. Chen and Y. C. Wang, BMC Bioinformatics, 2006, 7, 52.64 J. Tsang, J. Zhu and v. Oudenaarden,Mol. Cell, 2008, 26, 753–767.65 J. E. Ferrell and W. Xiong, Chaos, 2001, 11, How to make
continuous processes discontinuous and reversible processesirreversible.
66 N. I. Markevich, J. B. Hoek and B. N. Kholodenko, J. Cell Biol.,2004, 164, 353–359.
67 A. Becskei, B. Seraphin and L. Serrano, EMBO J., 2001, 20, celldifferentiation by graded to binary response conversion.
68 G. Yao, T. J. Lee, S. Mori, J. R. Nevins and L. You, Nat. CellBiol., 2008, 10, 476–482.
69 A. Kriete, W. J. Bosl and G. Booker, PLoS Comput. Biol., 2010, 6,e1000820.
70 A. Sveiczer, A. Csikasz-Nagy, B. Gyorffy, J. J. Tyson andB. Novak, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 7865–7870.
71 C. Liu, S. Li, T. Liu, J. Borjigin and J. D. Lin, Nature, 2007, 447,477–481.
72 B. N. Kholodenko, Eur. J. Biochem., 2000, 267, 1583–1588.73 N. Geva-Zatorsky, N. Rosenfeld, S. Itzkovitz, R. Milo, A. Sigal,
E. Dekel, T. Yarnitzky, Y. Liron, P. Polak, G. Lahav and U. Alon,Mol. Syst. Biol., 2006, 2, 33.
74 K. B. Wee, U. Surana and B. D. Aguda, PLoS One, 2009, 4, e4407.75 J. J. Tyson, Mol. Syst. Biol., 2006, 2, 32.76 O. Warburg, The Metabolism of Tumours, London Constable Co.
Ltd., London, 1930.77 M. G. Vander Haider, J. W. Locasale, K. D. Swanson, H. Sharfi,
G. J. Heffron, D. Amador-Noguez, H. R. Christofk, G. Wagner,J. D. Rabinowitz, J. M. Asara and L. C. Cantley, Science, 2010,329, 1492–1499.
78 K. Garber, Nat. Biotechnol., 2010, 28, 888–891.79 B. Stransky, J. Barrera, L. Ohno-Machado and S. J. D. Souza,
J. Bioinf. Comput. Biol., 2007, 5, 977–986.80 U. Alon, An Introduction to Systems Biology: Design Principles of
Biological Circuits, Chapman & Hall/CRC/Taylor & Francis, BocaRaton, FL, USA, 2006.
81 S. W. Chung, F. L. Miles, R. A. Sikes, C. R. Cooper,M. C. Farach-Carson and B. A. Ogunnaike, Biophys. J., 2004,96, 1733–1750.
82 J. Witt, S. Barisic, E. Schumann, F. Allgower, O. Sawodny,T. Sauter and D. Kulms, BMC Syst. Biol., 2009, 3, 71.
83 J. Cotterell and J. Sharpe, Mol. Syst. Biol., 2010, 6, 425.84 T.-R. Xu, V. Vyshemirsky, A. Gormand, A. von Kriegsheim,
M. Girolami, G. S. Baillie, D. Ketley, A. J. Dunlop, G. Milligan,M. D. Houslay and W. Kolch, Sci. Signaling, 2010, 3, ra20.
85 X. Cai and Z. M. Yuan, J. Comput. Biol., 2009, 16, 917–933.86 A. M. Del Rosario and F. M. White, Curr. Opin. Genet. Dev., 20,
23–30.
Dow
nloa
ded
by U
nive
rsite
de
Mon
trea
l on
19 J
uly
2011
Publ
ishe
d on
15
June
201
1 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
0IB
0014
5GView Online


















