
Intake Food
25
SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE Integration of Metabolism and Intake Regulation: A Review Focusing on Periparturient Animals K. L. Ingvartsen and J. B. Andersen Danish Institute of Agricultural Sciences Department of Animal Health and Welfare Research Centre Foulum P.O. Box 50 DK-8830 Tjele, Denmark ABSTRACT There has been great interest in dry matter intake regulation in lactating dairy cattle to enhance perfor- mance and improve animal health and welfare. Pre- dicting voluntary dry matter intake (VDMI) is complex and influenced by numerous factors relating to the diet, management, housing, environment and the animal. The objective of this review is to identify and discuss important metabolic factors involved in the regulation of VDMI and their integration with metabolism. We have described the adaptations of intake and metabo- lism and discussed mechanisms of intake regulation. Furthermore we have reviewed selected metabolic sig- nals involved in intake regulation. A substantial dip in VDMI is initiated in late preg- nancy and continues into early lactation. This dip has traditionally been interpreted as caused by physical constraints, but this role is most likely overemphasized. The dip in intake coincides with changes in reproduc- tive status, fat mass, and metabolic changes in support of lactation, and we have described metabolic signals that may play an equally important role in intake regu- lation. These signals include nutrients, metabolites, re- productive hormones, stress hormones, leptin, insulin, gut peptides, cytokines, and neuropeptides such as neu- ropeptide Y, galanin, and corticotrophin-releasing fac- tor. The involvement of these signals in the peripartur- ient dip in intake is discussed, and evidence supporting the integration of the regulation of intake and metabo- lism is presented. Still, much research is needed to clarify the complex regulation of VDMI in lactating dairy cows, particularly in the periparturient animal. (Key words: intake regulation, metabolism, peripart- urient, peptides) Received July 28, 1999. Accepted January 10, 2000. Corresponding author: Klaus L. Ingvartsen; e-mail: KlausL. [email protected]. 2000 J Dairy Sci 83:1573–1597 1573 Abbreviation key: APR = acute phase inflammatory response, ARN = arcuate nucleus, BBB = blood-brain barrier, CCK = cholecystokinin, CNS = central nervous system, CRF = corticotrophin-releasing factor, GLP-1 = glucagon-like peptide 1, GLP-2 = glucagon-like pep- tide 2, IL = interleukin, IV = intravenous, ICV = intrace- rebroventricular, LH = lateral hypothalamus, LPS = lipopolysaccharide, NPY = neuropeptide Y, PVN = paraventricular nucleus, SS = somatostatin, TNF-α = tumor necrosis factor-α, VDMI = voluntary DMI, VLH = ventrolateral hypothalamus, VMH = ventromedial hypothalamus. INTRODUCTION The factors affecting voluntary DMI (VDMI) of lactat- ing dairy cattle have received much attention for many decades. The traditional motivation for studying factors affecting VDMI has been, and still is, the scope for increasing intake and hence production, efficiency, and profitability. An alternative motivation for studying factors affecting VDMI and feed intake regulation is to better understand the role of, and ultimately manipu- late, intake in relation to animal health. Most of the health problems in the highly selected dairy cow, of both metabolic and infectious nature, occur in early lactation and have been related to relatively low intake just prior to parturition (342). Furthermore, infectious diseases are also known to down-regulate appetite via cytokines in the immune system (172, 334). To under- stand and possibly prevent hypophagia, we need to bet- ter understand and be able to quantify the linkages between mechanisms of intake regulation, metabolism, and the immune system in the dairy cow. Numerous models for predicting VDMI have been developed as shown in reviews by Ingvartsen (161) and Mertens (225). Critical evaluation of some of the models currently in use indicates that these models are inaccu- rate since they exclude important factors known to in- fluence VDMI (161, 225). In particular, nearly all mod- els are incapable of predicting intake in the peripartur- ient animal (161, 265) and late (> 40 wk) lactation (265). The few recent models developed for periparturient
-
Upload
septian-widyantoro -
Category
Documents
-
view
49 -
download
2
description
food
Transcript of Intake Food

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE
Integration of Metabolism and Intake Regulation:A Review Focusing on Periparturient Animals
K. L. Ingvartsen and J. B. AndersenDanish Institute of Agricultural Sciences
Department of Animal Health and WelfareResearch Centre Foulum
P.O. Box 50DK-8830 Tjele, Denmark
ABSTRACT
There has been great interest in dry matter intakeregulation in lactating dairy cattle to enhance perfor-mance and improve animal health and welfare. Pre-dicting voluntary dry matter intake (VDMI) is complexand influenced by numerous factors relating to the diet,management, housing, environment and the animal.The objective of this review is to identify and discussimportant metabolic factors involved in the regulationof VDMI and their integration with metabolism. Wehave described the adaptations of intake and metabo-lism and discussed mechanisms of intake regulation.Furthermore we have reviewed selected metabolic sig-nals involved in intake regulation.
A substantial dip in VDMI is initiated in late preg-nancy and continues into early lactation. This dip hastraditionally been interpreted as caused by physicalconstraints, but this role is most likely overemphasized.The dip in intake coincides with changes in reproduc-tive status, fat mass, and metabolic changes in supportof lactation, and we have described metabolic signalsthat may play an equally important role in intake regu-lation. These signals include nutrients, metabolites, re-productive hormones, stress hormones, leptin, insulin,gut peptides, cytokines, and neuropeptides such as neu-ropeptide Y, galanin, and corticotrophin-releasing fac-tor. The involvement of these signals in the peripartur-ient dip in intake is discussed, and evidence supportingthe integration of the regulation of intake and metabo-lism is presented. Still, much research is needed toclarify the complex regulation of VDMI in lactatingdairy cows, particularly in the periparturient animal.(Key words: intake regulation, metabolism, peripart-urient, peptides)
Received July 28, 1999.Accepted January 10, 2000.Corresponding author: Klaus L. Ingvartsen; e-mail: KlausL.
2000 J Dairy Sci 83:1573–1597 1573
Abbreviation key: APR = acute phase inflammatoryresponse, ARN = arcuate nucleus, BBB = blood-brainbarrier, CCK = cholecystokinin, CNS = central nervoussystem, CRF = corticotrophin-releasing factor, GLP-1= glucagon-like peptide 1, GLP-2 = glucagon-like pep-tide 2, IL = interleukin, IV = intravenous, ICV = intrace-rebroventricular, LH = lateral hypothalamus, LPS =lipopolysaccharide, NPY = neuropeptide Y, PVN =paraventricular nucleus, SS = somatostatin, TNF-α =tumor necrosis factor-α, VDMI = voluntary DMI, VLH= ventrolateral hypothalamus, VMH = ventromedialhypothalamus.
INTRODUCTION
The factors affecting voluntary DMI (VDMI) of lactat-ing dairy cattle have received much attention for manydecades. The traditional motivation for studying factorsaffecting VDMI has been, and still is, the scope forincreasing intake and hence production, efficiency, andprofitability. An alternative motivation for studyingfactors affecting VDMI and feed intake regulation is tobetter understand the role of, and ultimately manipu-late, intake in relation to animal health. Most of thehealth problems in the highly selected dairy cow, ofboth metabolic and infectious nature, occur in earlylactation and have been related to relatively low intakejust prior to parturition (342). Furthermore, infectiousdiseases are also known to down-regulate appetite viacytokines in the immune system (172, 334). To under-stand and possibly prevent hypophagia, we need to bet-ter understand and be able to quantify the linkagesbetween mechanisms of intake regulation, metabolism,and the immune system in the dairy cow.
Numerous models for predicting VDMI have beendeveloped as shown in reviews by Ingvartsen (161) andMertens (225). Critical evaluation of some of the modelscurrently in use indicates that these models are inaccu-rate since they exclude important factors known to in-fluence VDMI (161, 225). In particular, nearly all mod-els are incapable of predicting intake in the peripartur-ient animal (161, 265) and late (> 40 wk) lactation (265).The few recent models developed for periparturient
INGVARTSEN AND ANDERSEN1574
cows (264) are probably not universal since they aredeveloped to fit a certain type of cow and diet. We believethe metabolic component in intake regulation has beenunderemphasized in most previous attempts to developmodels for predicting intake. Original attempts havebeen made by Forbes (112) and Fisher et al. (104) tostudy the interactions between metabolic control andphysical limitations of feed intake in ruminants. How-ever, intake regulation is a very complex biological phe-nomenon. Eating behavior and food intake are the re-sults of neural integration of numerous signals relatingto the environment, feed, and the physiological state ofthe animal. We have some insight into the nature ofthese signals in ruminants (12, 100, 114, 236), but muchstill has to be learned about which factors are involvedin intake regulation and how these factors are inte-grated with the physiological state and metabolism ofthe animal.
More research is needed on intake regulation andhow intake is integrated with metabolism, particularlyin periparturient animals. A better understanding ofintake regulation and its integration with metabolismwill most likely improve concepts on intake regulationand, thereby, the accuracy of models for prediction ofintake, production, and energy balance, and ultimatelymake it possible to further improve animal health andproductivity in the future. Consequently, the objectiveof the present review is to identify and discuss im-portant metabolic factors that may regulate VDMI andtheir integration with metabolism of dairy cattle. Inparticular, we will examine the nature of the dip inintake in periparturient animals.
We do not attempt to give a complete review of thephysiology of intake regulation, but rather to focus onpossible signals involved in the homeorhetic regulationof intake. We have chosen to describe the naturallyoccurring adaptations of intake and metabolism in peri-parturient animals and briefly describe the regulationof the metabolic adaptations. Evidence is given of meta-bolic regulation of intake, the link between body re-serves and intake, and humoral signals involved in in-tegrating appetite and BW to show that metabolic fac-tors are equally important as physical factors inregulating intake. Subsequently, we review the role ofselected signals involved in intake regulation. We havechosen to primarily focus on signals involved in thelong-term regulation of intake and include nutrient,metabolite, hormonal, gut peptide, and cytokine sig-nals. Finally, we briefly discuss central integration ofmetabolism and intake regulation and the role of se-lected neuropeptides. Much of the literature reviewedis on laboratory animals and humans because informa-tion on ruminants is still lacking or limited.
Journal of Dairy Science Vol. 83, No. 7, 2000
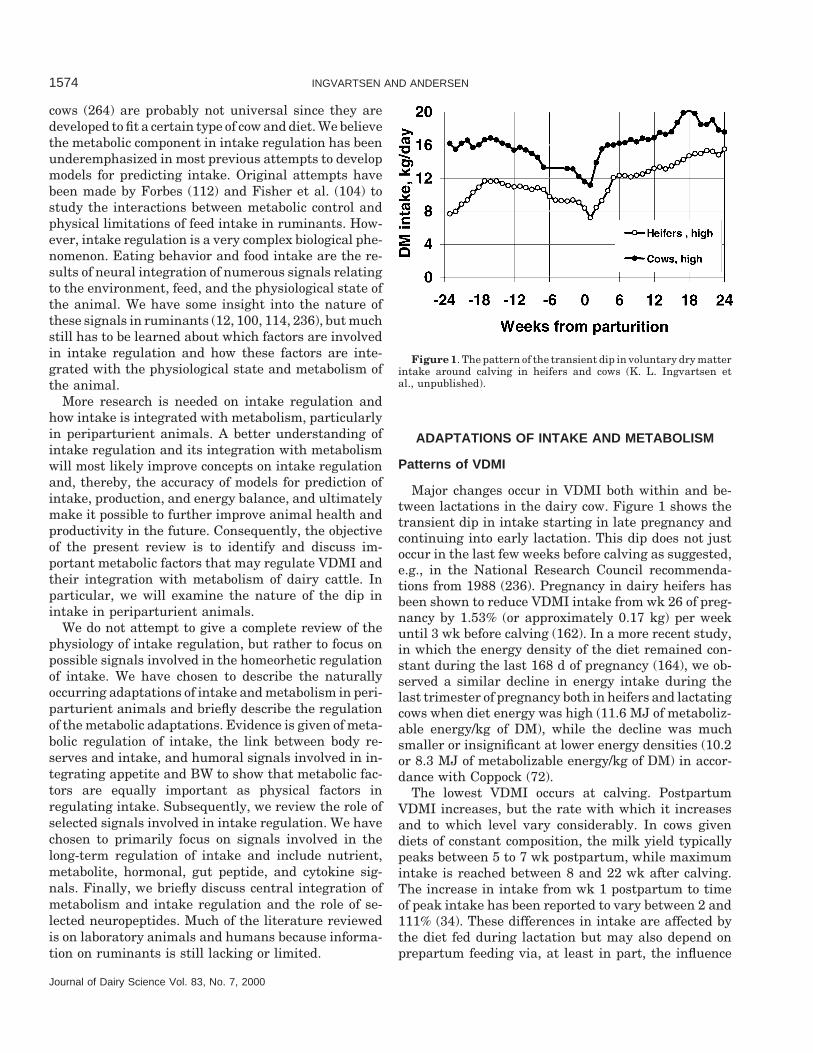
Figure 1. The pattern of the transient dip in voluntary dry matterintake around calving in heifers and cows (K. L. Ingvartsen etal., unpublished).
ADAPTATIONS OF INTAKE AND METABOLISM
Patterns of VDMI
Major changes occur in VDMI both within and be-tween lactations in the dairy cow. Figure 1 shows thetransient dip in intake starting in late pregnancy andcontinuing into early lactation. This dip does not justoccur in the last few weeks before calving as suggested,e.g., in the National Research Council recommenda-tions from 1988 (236). Pregnancy in dairy heifers hasbeen shown to reduce VDMI intake from wk 26 of preg-nancy by 1.53% (or approximately 0.17 kg) per weekuntil 3 wk before calving (162). In a more recent study,in which the energy density of the diet remained con-stant during the last 168 d of pregnancy (164), we ob-served a similar decline in energy intake during thelast trimester of pregnancy both in heifers and lactatingcows when diet energy was high (11.6 MJ of metaboliz-able energy/kg of DM), while the decline was muchsmaller or insignificant at lower energy densities (10.2or 8.3 MJ of metabolizable energy/kg of DM) in accor-dance with Coppock (72).
The lowest VDMI occurs at calving. PostpartumVDMI increases, but the rate with which it increasesand to which level vary considerably. In cows givendiets of constant composition, the milk yield typicallypeaks between 5 to 7 wk postpartum, while maximumintake is reached between 8 and 22 wk after calving.The increase in intake from wk 1 postpartum to timeof peak intake has been reported to vary between 2 and111% (34). These differences in intake are affected bythe diet fed during lactation but may also depend onprepartum feeding via, at least in part, the influence
SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1575
Table 1. A partial list of metabolic changes associated with onset of lactation in ruminants (23).
Physiological function Metabolic change Tissue involved
Milk synthesis ↑ synthetic capacity Mammary↑ blood flow↑ nutrient uptake and use
Lipid metabolism ↑ lipolysis Adipose↓ lipogenesis↑ use of lipid as energy Other body tissues
Glucose metabolism ↑ gluconeogenesis Liver↓ use of glucose Other body tissues
Protein metabolism ↑ protein mobilization Muscle and other bodytissues
Mineral metabolism ↑ absorption Gut↑ mobilization Bones
Intake ↑ food consumption Central nervous systemDigestion ↑ Hypertrophy of digestive tract Digestive tract (incl. Liver)
↑ capacity for nutrient absorption
↓ : decreasing; ↑ : increasing.
on the degree of fatness or BCS of periparturient ani-mals (45, 124, 163).
Voluntary DMI is considerably higher in multiparouscows compared with primiparous cows as illustratedin Figure 1. The intake capacity of primiparous cowscalving at an age of 2 yr is only around 80% of that ofmultiparous cows in the first part of lactation (170, 188).
The normal pattern of intake may be severely influ-enced by disease states. Both clinical and subclinicalinfections are known to substantially reduce appetiteand performance. On the surface, the occurrence of hy-pophagia due to infectious and noninfectious sub-stances, particularly in the periparturient cow, seemssomewhat paradoxical. Febrile animals not only needenergy to fuel elevated body temperature and nutrientsfor repair of tissue breakdown but hypophagia mayalso lead to metabolic diseases such as fatty liver andketosis, at least in the periparturient dairy cow. How-ever, as pointed out by Hart (143), the behavior of sickanimals is not only maladaptive response, but ratheran organized strategy that facilitates recovery andsurvival.
Adaptation and Patterns of Metabolism
During late pregnancy and lactation, energy require-ments increase considerably. Fetal energy require-ments on d 250 of pregnancy have been calculated to 2.3Mcal/d for Holstein cows (27). During lactation, energyrequirement is increased to 26 Mcal net energy in cowsproducing 30 kg of milk per day (27). Major changes inmetabolism occur to cope with this increase in nutrientrequirements (Table 1) and comprehensive reviews onthis topic exist (23, 27, 223, 327). The changes outlinedin Table 1 certainly indicate that lactation is not just
Journal of Dairy Science Vol. 83, No. 7, 2000
a function of the mammary gland but involves manyphysiological processes and body tissues.
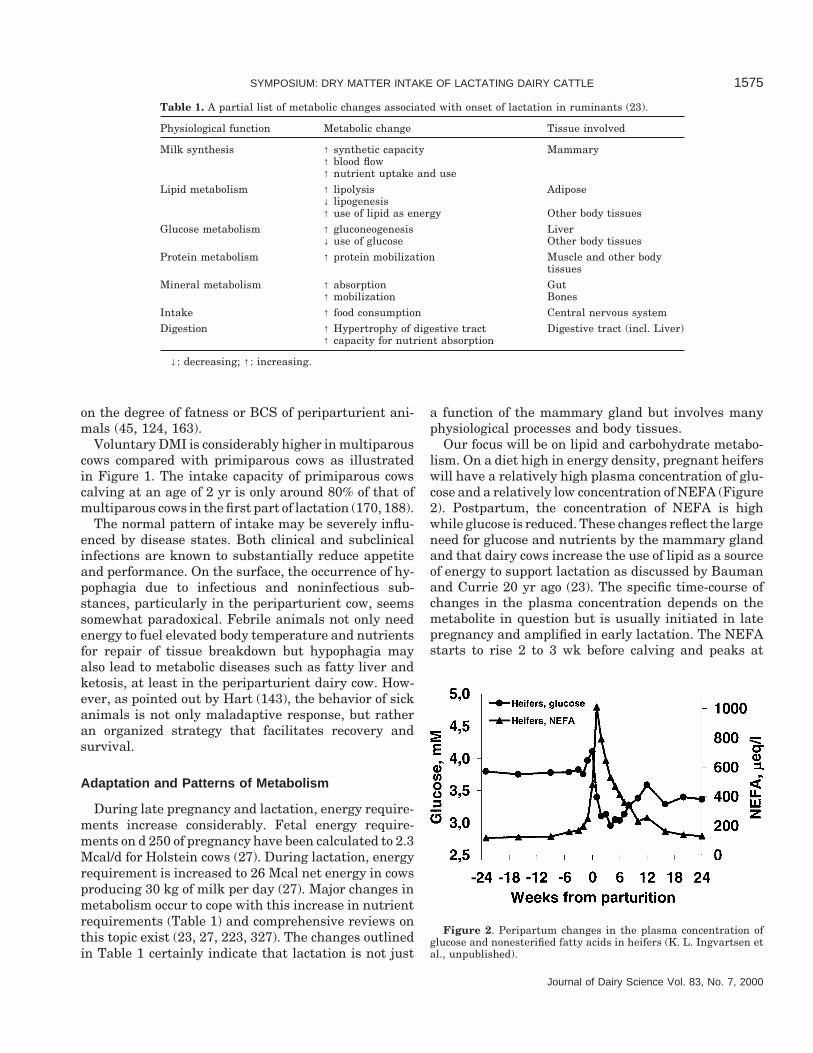
Our focus will be on lipid and carbohydrate metabo-lism. On a diet high in energy density, pregnant heiferswill have a relatively high plasma concentration of glu-cose and a relatively low concentration of NEFA (Figure2). Postpartum, the concentration of NEFA is highwhile glucose is reduced. These changes reflect the largeneed for glucose and nutrients by the mammary glandand that dairy cows increase the use of lipid as a sourceof energy to support lactation as discussed by Baumanand Currie 20 yr ago (23). The specific time-course ofchanges in the plasma concentration depends on themetabolite in question but is usually initiated in latepregnancy and amplified in early lactation. The NEFAstarts to rise 2 to 3 wk before calving and peaks at
Figure 2. Peripartum changes in the plasma concentration ofglucose and nonesterified fatty acids in heifers (K. L. Ingvartsen etal., unpublished).

INGVARTSEN AND ANDERSEN1576
Table 2. Changes in some homeorhetic and homeostatic hormones, tissue sensitivity, and responsivenessand effect in selected tissues in pregnancy and lactation.
LactogenesisMid-pregnancy Late pregnancy Early lactation
Potential homeorhetic hormones1
Progesterone ↑ (↓ ) ↓Placental lactogen ↑ ↓Estrogens ↑ ↓Prolactin - (↓ ) ↑Somatotropin - (↓ ) ↑Leptin ? ? ?
Homeostatic hormones1
Insulin ↑ ↓Glucagon - - -CCK and somatostatin ? ? ?
Tissue sensitivityInsulin ↑ ↓ ↓Catecolamines ↑ ↑
Tissue responsivenessInsulin ↓ ↓Catecolamines ↓ ↑ ↑
Liver2
Gluconeogenesis ↑Ketogenesis ↑
Adipose tissue2
Lipogenesis ↑ ↓ ↓FA esterification ↑ ↓ ↓Lipolysis ↑ ↑Glucose utilization ↓ ↓
Skeletal muscle2
Protein synthesis ↓ ↓Protein degradation ↑ ↑Glucose utilization ↓ ↓
↓ : decreasing; ↑ : increasing; ?: unknown in ruminants; -: no significant changes.1Plasma hormone concentration changes.2Changes in rate of metabolic processes.
calving or during the first week of lactation. Glucoseincreases during the last week before calving and dropsabruptly postpartum to reach a minimum 1 to 3 wkinto lactation. Postpartum changes in the plasma con-centration of BHBA are generally opposite those ofglucose.
Regulation of Metabolic Adaptations inPeriparturient Animals
The endocrine system is playing a pivotal role, butthe nervous system and the immune system are alsoinvolved in the regulation of metabolism and nutrientpartitioning. Bauman and Currie (23) applied the con-cept of homeorhesis to the regulation of nutrient parti-tioning in lactation and defined homeorhesis as “theorchestrated or coordinated changes in the metabolismof body tissues necessary to support a physiologicalstate.” In Table 2, an outline of changes in homeorhetichormones and their assumed consequences on tissuesensitivity and responsiveness is presented for differentphysiological states. The reader is referred to recent
Journal of Dairy Science Vol. 83, No. 7, 2000
reviews for further details on the regulation of meta-bolic adaptations to lactation (23, 27, 28, 327).
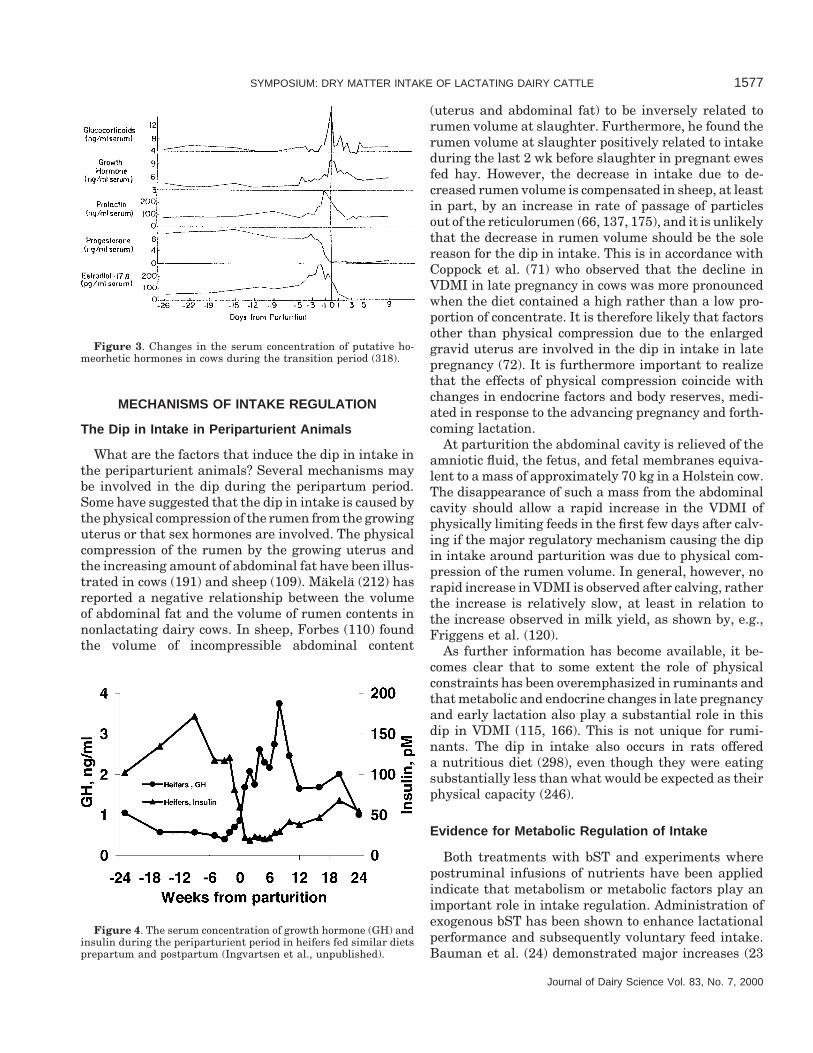
The time-course of changes of the different homeor-hetic hormones in periparturient cattle varies consider-ably as shown in Figures 3 and 4. Tucker (318) hasshown that plasma concentrations of sex hormones, glu-cocorticoids, as well as other hormones, change in arelatively narrow time period around parturition. Ap-proximately 5 d before parturition, major increases oc-cur in the plasma concentrations of estrogen and corti-sol. Estrogen and cortisol have their highest concentra-tion at 3 and 0 d before parturition, respectively, afterwhich the concentrations decrease to their normal post-partum levels within few days. In Figure 4, changes insomatotropin and insulin are shown to illustrate thenegative correlation between a homeorhetic hormone(somatotropin) and a homeostatic hormone (insulin).Clearly, a number of hormones are potentially involvedin the adaptation of metabolism and some of these arealso, directly or indirectly, involved in the majorchanges in intake, as will be discussed in later sections.
SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1577
Figure 3. Changes in the serum concentration of putative ho-meorhetic hormones in cows during the transition period (318).
MECHANISMS OF INTAKE REGULATION
The Dip in Intake in Periparturient Animals
What are the factors that induce the dip in intake inthe periparturient animals? Several mechanisms maybe involved in the dip during the peripartum period.Some have suggested that the dip in intake is caused bythe physical compression of the rumen from the growinguterus or that sex hormones are involved. The physicalcompression of the rumen by the growing uterus andthe increasing amount of abdominal fat have been illus-trated in cows (191) and sheep (109). Makela (212) hasreported a negative relationship between the volumeof abdominal fat and the volume of rumen contents innonlactating dairy cows. In sheep, Forbes (110) foundthe volume of incompressible abdominal content
Figure 4. The serum concentration of growth hormone (GH) andinsulin during the periparturient period in heifers fed similar dietsprepartum and postpartum (Ingvartsen et al., unpublished).
Journal of Dairy Science Vol. 83, No. 7, 2000
(uterus and abdominal fat) to be inversely related torumen volume at slaughter. Furthermore, he found therumen volume at slaughter positively related to intakeduring the last 2 wk before slaughter in pregnant ewesfed hay. However, the decrease in intake due to de-creased rumen volume is compensated in sheep, at leastin part, by an increase in rate of passage of particlesout of the reticulorumen (66, 137, 175), and it is unlikelythat the decrease in rumen volume should be the solereason for the dip in intake. This is in accordance withCoppock et al. (71) who observed that the decline inVDMI in late pregnancy in cows was more pronouncedwhen the diet contained a high rather than a low pro-portion of concentrate. It is therefore likely that factorsother than physical compression due to the enlargedgravid uterus are involved in the dip in intake in latepregnancy (72). It is furthermore important to realizethat the effects of physical compression coincide withchanges in endocrine factors and body reserves, medi-ated in response to the advancing pregnancy and forth-coming lactation.
At parturition the abdominal cavity is relieved of theamniotic fluid, the fetus, and fetal membranes equiva-lent to a mass of approximately 70 kg in a Holstein cow.The disappearance of such a mass from the abdominalcavity should allow a rapid increase in the VDMI ofphysically limiting feeds in the first few days after calv-ing if the major regulatory mechanism causing the dipin intake around parturition was due to physical com-pression of the rumen volume. In general, however, norapid increase in VDMI is observed after calving, ratherthe increase is relatively slow, at least in relation tothe increase observed in milk yield, as shown by, e.g.,Friggens et al. (120).
As further information has become available, it be-comes clear that to some extent the role of physicalconstraints has been overemphasized in ruminants andthat metabolic and endocrine changes in late pregnancyand early lactation also play a substantial role in thisdip in VDMI (115, 166). This is not unique for rumi-nants. The dip in intake also occurs in rats offereda nutritious diet (298), even though they were eatingsubstantially less than what would be expected as theirphysical capacity (246).
Evidence for Metabolic Regulation of Intake
Both treatments with bST and experiments wherepostruminal infusions of nutrients have been appliedindicate that metabolism or metabolic factors play animportant role in intake regulation. Administration ofexogenous bST has been shown to enhance lactationalperformance and subsequently voluntary feed intake.Bauman et al. (24) demonstrated major increases (23

INGVARTSEN AND ANDERSEN1578
Figure 5. The pattern of average daily food intake in rats offereda diet high in nutrient density in pregnancy and lactation (298).
to 41%) in milk yield and a subsequent increase inVDMI when dairy cows were treated with varying dailydoses of bST. Since then several studies, particularlystudies using prolonged-release formulations, have con-firmed the increased milk yield, which is typically about10 to 15% (22, 47, 58, 95). The general pattern of re-sponse in milk yield is a rapid increase over the firstfew days, rising to a maximum usually within the firstweek of treatment. Voluntary feed intake does not in-crease until several weeks after the increase in milkyield has occurred (22, 58). The increase in intake de-pends not only on the increase in milk yield but is alsoinfluenced by the quality or density of the diet (58). Theabove findings indicate that energy requirements formilk production are “pulling” VDMI up and that intakeregulation therefore is influenced by metabolism. How-ever, physical limitations clearly also play a role, andgood nutrition and management are prerequisites forimproved performance (22).
Other examples showing that metabolic factors in-fluence VDMI via postruminal mechanisms may befound in experiments using abomasal or duodenal infu-sions of nutrients. Abomasal infusions of fat have re-duced VDMI in some trials or treatments (41, 60, 61,64, 122) but not all (41, 64, 86, 123, 238). The intakeresponse depends on the amount of fat infused but otherfactors such as degree of saturation and the fatty acidchain length may also influence intake. The mecha-nisms involved in the depression of VDMI caused byfat infusion have been little investigated in cattle, butgastrointestinal hormones (62, 242) and fatty acid oxi-dation (276) are reported to be involved and will bediscussed later.
Journal of Dairy Science Vol. 83, No. 7, 2000
Body Reserves and Voluntary Intake
Voluntary DMI in ruminants is negatively correlatedwith body reserves at a given physiological state. Bineset al. (36) and Bines and Morant (35) studied the effectof body fatness on the food intake of nonlactating cowsin a changeover design. They fed cows to become fat orthin and compared ad libitum intake of straw, hay, andconcentrates. No difference was seen in straw intake,but daily intake of hay and concentrate was reducedby approximately 23%, when cows were fat comparedto thin. They suggested that there was a more rapidutilization of lipogenic substrates by thin cows than byfat cows and this would prevent the excessive accumula-tion in the blood of lipogenic precursors and therebyenhance their absorption from the rumen and stimulatefeed intake in thin cows. The only difference betweenfat and thin cows in rumen and blood metabolites thateven approached significance (P < 0.1) was a lower pro-pionate and a higher glucose postfeeding in the fatcows (35).
Prepartum feeding level may influence the amount ofenergy reserves or BCS and thereby peripartum healthand postpartum performance (44, 124, 133, 163). In-gvartsen et al. (163) reviewed the effect of prepartumfeeding and weight gain on peripartum metabolism andfeed intake based on 24 studies (6 in heifers, 10 in drycows, and 8 in cows in late lactation and the dry period).They argued that the inconsistent postpartum re-sponses to prepartum treatments could be explained,at least in part, by the large differences in prepartuminitial BW and BW gain between treatments. They ob-served a positive relationship between prepartumweight gain and the extent of postpartum mobilizationof body tissues and argued that differences in prepar-tum feeding causing excessive mobilization, more thanapproximately 40 kg of BW, appeared to depress feedintake.
Increased fatness or body reserves also down-regu-lates VDMI in sheep (108, 324). If lean adult sheep arefed a high-energy diet ad libitum they gain weight untilthey weigh approximately twice their initial weight andthey reduce intake during the latter part of the fat-tening period (216, 324). Fat sheep at their static BWreduced energy intake per kilogram of BW to a levelsimilar to that of thin control ewes fed a maintainingdiet (216, 324). Carcass fat increased from between 9and 25% to between 35 and 49% (216, 324). This indi-cates that sheep try to reach certain body fatness andmaintain this body fatness by adjusting intake. Follow-ing a period of force feeding, rats adjusted voluntaryintake to return to the BW prior to the imposed feeding(87). The above clearly indicates that the regulationof body reserves and food intake is coordinated and

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1579
supports the lipostatic theory for appetite regulationproposed by Kennedy in 1953 (180). Coincident withthe changes in food intake and body stores, plasmaconcentrations of insulin, thyroid hormones, and NEFAoccurred (216). During the first phases of fattening, alinear increase in plasma insulin concentration oc-curred until around 20 to 25 wk of fattening. Thereafterplasma insulin concentration seemed to increase rap-idly to a sustained very high concentration of around249 pM (216, 324). Furthermore, these changes werecoincidental with a substantial increase in mean fatcell weight that occurred after 20 to 24 wk of fattening(324). However, the above relations do not provide proofof a humoral link between the adipose tissue and appe-tite nor of the signals involved.
Evidence of Humoral Signals CoordinatingAppetite and Body Reserves
Parabiosis is the union of two living individuals. Thisunion may occur spontaneously as in joined twins ormay be produced in experimental animals by surgicaltechniques. The parabiosis technique has been used todemonstrate the involvement of circulating factors orhormones in the regulation of physiological systems(103) and, consequently, whether or not humoral sig-nals are involved in coordinating BW and intake. Her-vey (148) was the first to report that obesity inducedby lesion of the ventromedial hypothalamus (VMH) inone member of a parabiotic rat pair led to hypophagiaand weight loss in the unlesioned rat, results that havebeen confirmed by Nishizawa and Bray (237). WhenParameswaran et al. (243) instead stimulated the lat-eral hypothalamus (LH) of one parabiotic rat electri-cally to obtain a marked increase in intake and weight,the unstimulated parabiont ate progressively less andbecame increasingly thin in accordance with the find-ings of Hervey (148). At death the stimulated parabionthad a major increase in the adipose tissue mass, whilethe unstimulated rat was essentially devoid of fat. Ga-vage overfeeding of one parabiotic rat to induce obesitylikewise caused hypophagia and loss of weight and bodyfat in its partner (140, 237). Reversal of the hypophagiaby tube-feeding the lean parabiont with 100% of theenergy intake of the control rats prevented the loss ofBW and, when gavage overfeeding was stopped, bothanimals returned to normal BW (141). Circulating con-centrations of insulin, glucagon, somatotropin, cortico-sterone, glucose, NEFA, and BHBA (140, 243) were notable to explain the observed changes in intake and lossof weight. These experiments indicated that an un-known humoral satiety factor was acting and was re-lated to elevated body fatness.
Journal of Dairy Science Vol. 83, No. 7, 2000
There appears to be a genetic basis for this factor aslean mice in parabiosis with genetically obese (db/db)mice quickly lost interest in food and rapidly lost weightand died of apparent starvation (68). Furthermore,weight gain was markedly depressed in obese ob/obmice by parabiosis with lean littermates (67). Coleman(67) observed that obese mice ate less when paired withlean mice compared with other obese mice. He further-more observed that when obese ob/ob mice were pairedwith obese db/db mice they experienced hypophagia andfat loss and had a poor survival rate. The above resultsformed the basis of work which shows that obesity inanimals can occur with a deficiency of an adipose tissue-related satiety factor (ob/ob) or a deficiency in its recep-tor (db/db). We now know that the factor alluded to byKennedy’s lipostatic theory (180) is the hormone leptinthat will be discussed in detail in a later section.
NUTRIENTS AND METABOLITES AS SIGNALSIN INTAKE REGULATION
We have so far discussed aspects of adaptations ofintake and given evidence for the involvement of hor-monal and metabolic factors in intake regulation. Inthe following we will review the involvement of selectedhormonal and metabolic signals in the homeorhetic reg-ulation of intake.
There are numerous nutrients and metabolites thatcould potentially be considered if we were to discussaspects of satiety in relation to meal patterns. In theirpaper, Koopmans et al. (184) have substantiated therole of endogenous signals from the gut or absorbednutrients and their metabolic consequences in intakeregulation. Studies with intravenous (IV) infusion ofnutrients indicate that rats are able to sense plasmanutrients and that they will reduce intake, at leastpartially (239). The sensing system responds to differ-ent nutrients with different degrees of behavioral com-pensation, suggesting that each of the plasma nutrientshas its own sensing system. Because our focus is onthe long-term rather than the short-term regulation ofintake, and is furthermore focused on the peripartur-ient animal, we will limit this discussion to fat mobi-lized from adipose tissue.
Oxidation of Metabolic Fuels
As discussed in previous sections, large amounts ofadipose tissue are mobilized during late pregnancy andearly lactation in support of lactation, causing a rise inthe circulating concentrations and use of NEFA, glyc-erol, and ketone bodies. During this period, a negativerelationship between the plasma concentration ofNEFA and VDMI is observed in cattle (133, 165). In

INGVARTSEN AND ANDERSEN1580
rats recovering from insulin-induced obesity, negativecorrelations were observed between food intake andplasma levels of NEFA, glycerol, and ketone bodies (52).Therefore, it is reasonable to speculate that NEFA, glyc-erol, and ketone bodies act as potential signals in intakeregulation and that they may play a role in the dip inintake in the periparturient animal.
NEFA. Continuous long-term intravenous infusionof long-chain fatty acids has been shown to cause hypo-phagia in rats (51) and in adult sheep (325). In dairycows, a short-term (4 h) intravenous intralipid infusionresulted in a slightly reduced DM intake in two trials(16). The infusion provided 16.7 MJ of NEL, but themean decrease in intake in the two experiments wasonly 3.8 and 7.1 MJ of NEL.
Fatty acid oxidation in the brain and the liver hasbeen suggested as signals in intake regulation. Kasseret al. (176) suggested that changes in fatty acid utiliza-tion in the ventral hypothalamus reflected changes inthe peripheral energy balance. They found that changesin the rate of fatty acid oxidation in the ventrolateralhypothalamus (VLH) are inversely proportional tochanges in peripheral fat storage and, therefore, mayparticipate in feeding responses to changes in periph-eral fat stores (177, 178). However, testing this hypothe-sis by inducing chronic (14 d) changes in VLH fattyacid oxidation caused no effect on food intake, BW, andplasma glucose, insulin, and NEFA (30). It is thereforeunlikely that fatty acid oxidation in the VLH changesfood intake or peripheral metabolism.
The β-oxidation of fatty acids may be inhibited bymercaptoacetate, which depresses long-chain acyl-CoAdehydrogenase activity (21) or methyl palmoxirate,which depresses the mitochondrial carnitine palmitoyl-transferase I concentration (320). Inhibiting fatty acidoxidation with intraperitoneal injections of mercap-toacetate (276, 292) or methyl palmoxirate (119, 153)increased cumulative food intake in rats but only whenthe rate of fatty acid oxidation was relatively high (276,291). Interestingly, the feeding response to mercap-toacetate is lower in S5B/PL rats that are resistant todietary fat-induced obesity, compared with sensitiveOsborne-Mendel rats (293). The hypophagia is mostlikely mediated by hepatic receptors, because the feed-ing response in rats fed a high fat diet to an inhibitionof fatty acid oxidation by mercaptoacetate diet was par-tially blocked by hepatic branch vagotomy (198). Thehypophagia is probably linked to mitochondrial oxida-tion of NEFA providing satiety signals mediated byvagal afferents (277). Satiety may be caused by mito-chondrial oxidation of fatty acids that may affect thehepatocyte membrane potential through cytosolic ATPand Na-pump activity (193). This theory is supportedby the finding that methyl palmoxirate-induced in-
Journal of Dairy Science Vol. 83, No. 7, 2000
creases in food intake were negatively related to liverATP content, ATP-to-ADP ratio, and phosphorylationpotential, indicating that a decrease in fatty acid oxida-tion can stimulate feeding behavior by reducing hepaticenergy production (118).
Choi et al. (63) tested the hypothesis that inhibitionof fatty acid oxidation by sodium mercaptoacetateblocks the decrease in feed intake by dairy heifers whenthe fat content of their diet is increased. Instead of theexpected increase in VDMI, they observed a substantialdecrease in intake during the first 4 h postinjection.The reason for the depressed VDMI remains unex-plained but may relate to mercaptoacetate-inducedchanges in hormones and metabolite changes (63, 322).More research is needed to clarify the role of β-oxidationof fatty acids in feed intake regulation and the lowintake of the periparturient cow with high lipid mobili-zation.
Glycerol. The effect of glycerol on intake in rats hasrecently been reviewed (278). Glycerol may influencefeeding through a central nervous system (CNS) mech-anism since intracerebroventricular infusions of glyc-erol in rats have been shown to suppress feeding (78).Subcutaneous administration of low levels of glycerolto rats, however, has given variable results and onlydoses of glycerol increasing plasma glycerol to nonphys-iological levels reliably reduce food intake (51, 105,259). Glycerol infused into the portal vein of castratedmale sheep at a rate of 0.3 mmol min−1 for 3 h did notincrease the peripheral concentration of glycerol andhad no short-term effect on intake (114). Glycerol doesnot seem to be a strong candidate for a signal mediatingreduced intake in periparturient cattle.
Ketone bodies. Increased mobilization of body tis-sues in early lactation is generally associated with anincreased production and concentration of ketones. Inrats, subcutaneous injections of BHBA have beenshown to cause hypophagia (105), whereas its metabolicproduct, acetoacetate did not (196). Because section ofthe hepatic vagus branch eliminated the hypophagiafollowing subcutaneous injections of BHBA (194, 195),it seems reasonable to assume that the liver is involved(278). However, Stricker et al. (307), who also postu-lated a peripheral effect of BHBA on satiety, did notbelieve in a hepatic effect since the liver cannot oxidizeBHBA beyond acetoacetate. Possibly BHBA may affectintake via central mechanisms since chronic intracere-broventricular (ICV) infusion reduces intake in rats(8). Recently, Sun et al. (309) found that chronic (28 d)ICV infusions of BHBA reduced BW gain but not foodintake. The physiological role of BHBA in intake regula-tion is still not clear.

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1581
Table 3. Hormones and peptides that increase or to decrease feed intake (3, 39, 173, 267, 272, 335, 337,340).
Increase intake Decrease intake
β-Endorphin Agouti-related protein InsulinDynorphin Anorectin LeptinGalanin Amylin NeurotensinGrowth hormone-releasing hormone Bombesin Norepinephrine (β)Neuropeptide Y Caerulin OxytocinNorepinephrine (α2) CCK 8 and 33 SatietinMelanin-concentrating hormone Corticotropin-releasing factor SerotoninMelanocyte stimulating hormone α Cyclo-His-Pro SomatostatinOpoids Dopamine Substance POrexin A and Orexin B Enterostatin Thyrotropin-Progesterone Estrogen Releasing hormonePeptide YY Gastrin-releasing peptide Vasopressin
Glucagon XeninGlucagon-like peptide-1
HORMONAL SIGNALS IN INTAKE REGULATION
As outlined in previous sections, the reproductivehormones, stress hormones, leptin, hormones involvedin nutrient partitioning, and gut peptides are all poten-tially involved in the regulation of intake. The lengthof the list of potential regulatory hormones or peptideshas increased rapidly in the last 25 yr. A large numberof these hormones or peptides have been shown to playa role in the regulation of intake (Table 3). Clearly, itis beyond our scope to review all of the hormones andpeptides listed in Table 3. We will focus on the reproduc-tive hormones, hormones of the stress-axis, leptin, andinsulin that we believe are involved in the regulationof intake in periparturient animals as previously dis-cussed. Furthermore, we will briefly summarize theinformation on the gut peptides glucagon, glucagon-like peptide-1 (GLP-1), cholecystokinin (CCK) and so-matostatin (SS). For most of the other hormones orpeptides mentioned in Table 3, limited information isavailable for ruminants.
Reproductive Hormones
Reproductive hormones play an important role in theregulation of appetite and energy metabolism in rats(331) and ruminants (114). Ovariectomy in rats re-sulted in a transient increase in intake for 3 to 4 wkand a sustained elevation of BW (229, 314). The effectsof ovariectomy were reversed by replacement therapywith physiological doses of estrogen—a transient hypo-phagia and a sustained reduction of BW were observedas long as the estrogen treatment continued (229, 314).In general, progesterone reverses nearly all effects ofestrogen on energy balance, including an increase infeed intake (266, 329). The reason for the transienteffect of ovariectomy on intake is not clear but may, atleast in part, be due to simultaneous changes in the
Journal of Dairy Science Vol. 83, No. 7, 2000
degree of fatness and concurrent changes in leptin con-centration.
Intravenous infusion of 17β-estradiol in amountssimilar to those secreted at estrus and in late pregnancycaused a dose-dependent decrease in food intake in cas-trated male sheep and in goats (113). In cows, intrave-nous injections of 17β-estradiol decreased both milkyield and feed intake (134). Intraventricular injectionsof estradiol-benzoate in castrated male sheep increasedintake at low doses but depressed intake at doses higherthan 60 µg (111), but this may be above the physiologi-cal range. Studies in rats also indicate that the effectof estrogen on food intake is due, at least in part, todirect actions in the brain. The paraventricular nucleus(PVN) of the hypothalamus rather than the ventrome-dial hypothalamus (VMH) is the principal site of actionfor estrogen on food intake (48). Progesterone has notbeen reported to have a direct effect on intake but hasbeen reported to block the effects of estrogen both insheep (111) and in cows (233).
Stress Hormones
Acute stress reactions are coordinated and regulatedby the CNS, partly via increased secretion of hormonesof which corticotrophin-releasing factor (CRF) plays acentral role. The CRF is secreted in the brain, primarilyfrom the PVN of the hypothalamus that has also beenshown to mediate an effect on feeding (187). The CRFhas also been located in the gastrointestinal tract (247).CRF decreases food intake by central mechanisms. Cen-trally administered, CRF reduces feed intake in rodents(9, 154, 261), monkeys (129), and cattle (268). Krahnet al. (186) demonstrated a blocking of stress-inducedhypophagia and partial reversal of CRF-induced satietyafter central administration of a CRF antagonist. Be-cause CRF released into the pituitary portal system

INGVARTSEN AND ANDERSEN1582
stimulates ACTH, and consequently cortisol, these hor-mones could mediate the decreased appetite. However,hypophysectomy has no effect on feeding or on the ac-tions of CRF on feeding (202, 231). This agrees withthe finding that exogenous cortisol administration doesnot influence intake in sheep (11) and cattle (146). Ifwe assume that the increased cortisol around calving(318) is mediated primarily by CRF and ACTH, thenCRF may play a role in the dip in intake around parturi-tion. Because the concentration of cortisol is reducedto normal postpartum levels within few days of parturi-tion, it is unlikely that CRF plays any major role inintake regulation beyond this time in early lactationunless the animal is stressed.
Leptin
Recently, positional cloning has been used to success-fully identify the ob gene (344) and its receptor (313),the db gene (56), in mice. The product secreted by the obgene was named leptin (from the Greek ‘leptos’ meaningthin) and was isolated in 1994 by Zhang et al. (344).Since the cloning of the gene for leptin and its receptorin mice, the gene has also been cloned in the bovine(171), the pig (31, 260), and chicken (312).
Leptin and its receptor. Leptin is produced primar-ily by adipose cells (210, 215). It circulates both free ofand bound to leptin-binding proteins in rodents andhumans (157, 294). No difference is evident in the pro-portion of free or bound leptin either pre- or postpartumor between normal and insulin-dependent diabetic sub-jects (204). The leptin-binding proteins have not yetbeen characterized in farm animals. Plasma leptinshows a circadian rhythm (296) and appears to have apulsatile secretion (297). Following the identificationof the leptin receptor (Ob-R) by Tartaglia et al. (313),several isoforms have been located. The long isoform,Ob-Rl, is expressed at higher levels in the hypothala-mus and at a lower level in other tissues, while shortisoforms (Ob-Rs) are found in almost all tissues (128,201).
Leptin concentration is affected by body re-serves, BW changes and pregnancy. The plasma lep-tin concentration is positively correlated with body re-serves in rodents (117, 245) and man (50, 70, 211).A diet-induced weight loss causes the plasma leptinconcentration to be markedly reduced in humans androdents (70, 89, 211). A weight gain in humans due tooverfeeding is associated with increased basal leptinconcentrations (211) as much as 300% (181). The levelof plasma leptin drops markedly (approximately 70%)with short-term fasting (24 h) (38, 183) and is, therefore,not solely affected by body fatness. During fasting,plasma leptin concentrations are positively correlated
Journal of Dairy Science Vol. 83, No. 7, 2000
with glucose and insulin concentrations, while therewas no significant relationship between leptin andBHBA concentrations (38). Fasting under euglycemicconditions did not cause changes in leptin concentra-tions during a 72-h fast (181).
Leptin levels in rats are increased 1.8-fold duringpregnancy, followed by a decrease just before parturi-tion (57, 179). Leptin receptor mRNA levels in theuterus also increase about 2.7-fold during pregnancy,whereas no changes were observed in other tissues (57).Chien et al. (57) suggest that, apart from having effectson appetite, leptin may be involved in the partitioningof nutrients from the mother to the fetus. In humans,Schubring et al. (281) observed that leptin levels in-creased continuously during pregnancy and reached amaximum at 38 to 40 wk. At birth, leptin concentrationsdid not change significantly (25.8 to 23.5 ng/ml) but 3d after delivery, a dramatic decrease in leptin levels to10.6 ng/ml was observed. Six weeks after delivery, lep-tin levels were comparable to the values measured atthe beginning of pregnancy. Other studies have re-ported similar changes in pregnancy or around parturi-tion in humans (310) and rats (4, 179). Terada (316)observed no increase in leptin during pregnancy butobserved a much lower leptin concentration in lactatingcompared with nonlactating rats. The increase in theplasma leptin concentration in early pregnancy, beforeany major changes in body fat and resting metabolicrate, suggests that pregnancy represents a leptin-resis-tant state (149). Leptin levels during pregnancy did notcorrelate with maternal sex steroids (281), but humanchorionic gonadotropin and estrogen significantly in-crease leptin secretion by cultures of 3T3-L1 adipocytes.In early lactation, plasma leptin concentration is re-duced to levels observed in virgin control rats (57, 248)which may contribute to the increase in intake in earlylactation. Pickavance et al. (248) observed that the linkbetween nocturnal food intake and increased serumleptin was broken during lactation and speculated thatthe hypoleptinemia may be an important factor promot-ing the hyperphagia of lactation.
Plasma concentrations of leptin are influenced by anumber of hormones (69, 295). Cortisol within the phys-iological range increases plasma leptin concentrationsubstantially in humans (91, 182). Insulin increasesthe ob-gene expression in rats (273), but short-term(<6 h) or acute hyperinsulinemia does not affect leptinconcentration (76, 183). However, long-term (64 to 72h) increases in insulin, using a hyperinsulinemic eugly-cemic clamp technique, have been shown to increaseboth the peripheral and portal concentration of leptinin rats by up to 50% (183, 185). The basal concentrationsof leptin and IGF-I are not correlated but chronic ad-

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1583
ministration of recombinant IGF-I causes a sustaineddecrease in leptin levels in humans (77).
Food intake. Daily intraperitoneal injections of re-combinant leptin reduce food intake and BW and in-crease energy expenditure in ob/ob mice (18, 138, 244,245). In parabiotic studies with mice, recombinant lep-tin has been shown to cross between animals (142) andto reduce food intake, serum glucose and insulin, andBW in ob/ob partners of db/db mice (139). Using genetherapy to induce hyperleptinemia (6- to 10-fold in-crease in plasma) in normal mice caused a 30 to 50%decrease in food intake. These mice gained only 22 g,while control animals gained 124 g during the 24-dexperimental period (56). The site of action at whichleptin exerts its major effect is the hypothalamus. In-deed, ICV injections of leptin cause a rapid decrease inintake and body reserves in ob/ob mice (49, 228, 306),rats (75, 107, 269, 285, 288, 289), and monkeys (311).In the rat, the arcuate nucleus (ARN) appears to bedecisive in controlling appetite (275). However, theVMH also seems to be involved in leptin-induced hypo-phagia since VMH lesions caused hyperphagia despitevery high leptin concentrations (275). Recently, evi-dence for leptin regulation of food intake in humanshas been given (200). In pigs, ICV injections of porcineleptin decreased intake substantially and in a dose-dependent manner (14). Barb et al. (14) found that lep-tin modulates growth hormone secretion—leptin in-creased growth hormone when it was given in supra-physiological levels, but reduced growth hormone re-sponse to growth-hormone releasing factor. Serum IGF-I, insulin, thyroxin, glucose, and NEFA were unaffectedby leptin treatment.
Ruminants. Little is known about the biology of lep-tin in ruminants. In sheep, leptin primarily binds tothe dorso- and ventromedial nuclei of the hypothalamusand to the ARN (90). In ovariectomized ewes, ICV ad-ministration of recombinant human leptin over 3 d de-creased the VDMI significantly (147). Long-term (24 h)but not short-term (0.5 h) incubations of ovine anteriorpituitary tissue with leptin caused an increase ingrowth hormone secretion but reduced the response togrowth hormone-releasing factor (263), which agreeswith results in pigs (14). Leptin treatment elevatedplasma NEFA and lactate but did not affect glucoseor insulin levels, indicating a state of negative energybalance (147). Chilliard et al. (59) cite French studiesthat show a positive correlation between body fatnessand plasma leptin in sheep and cattle. Further, theyreport plasma immunoreactive leptin concentrations insheep and cattle and adipose leptin mRNA in sheepto be decreased by undernutrition and increased byoverfeeding as in rodents and humans.
Journal of Dairy Science Vol. 83, No. 7, 2000
Based on data from rodents and humans, plasmaleptin concentration is expected to increase in dairycows during the concurrent fattening that usually oc-curs during pregnancy. In contrast, plasma leptin con-centration is expected to decrease in the immediateprepartum period and early lactation when dairy cowsmobilize adipose tissue in support of lactation. Thisfall in leptin should be associated with an enhancedappetite. In the immediate prepartum period, such anincreased appetite does not occur, perhaps because in-take in this period is affected significantly by otherfactors. A reduced leptin concentration in early lacta-tion is potentially involved in intake regulation in earlylactation but clearly more research in cattle is needed.
Insulin
The short-term effects of insulin on satiety in rumi-nants have recently been reviewed by Grovum (132),who focused on its relationship with short chain fattyacids and post-prandial release. Thus, we will focuson the role of insulin as a signal of energy status tothe brain.
Insulin clearly exerts biological effects on brain tis-sue. Insulin acts as a neuromodulator within the ner-vous system (19, 20), and has been implicated in theautonomic function (271) and the growth and develop-ment of the CNS (20). Twenty-five years ago, insulinwas actually the first peptide shown to be able to de-crease intake when injected short-term into the intra-ventromedial hypothalamus of rats (144). Since then,insulin has been shown to be involved in the centralregulation of both food intake and BW (258). Woods etal. (339) suggested that insulin might be a sensor ofthe peripheral metabolic status in monogastrics. Thistheory is supported by the positive correlation betweenplasma insulin concentration and degree of fatness innonreproductive mature monogastrics (10, 257, 284)and ruminants (216, 323). Insulin from the peripheryis actively transported into the CNS via a saturablemechanism consistent with insulin binding to blood-brain barrier (BBB) insulin receptors and subsequenttranscytosis through microvessel endothelial cells (25,26, 286). Within the CNS, insulin binds to specific insu-lin receptors located on neurons and glial cells (19, 145),and some of the highest concentrations of insulin recep-tor-expressing neurons are found in the areas of thebrain which are important for the control of food intakeand energy metabolism (19).
The effect of insulin on the regulation of food intakemay depend on chronic infusions and acute injectionsmay have no effect (255), although acute effects havebeen reported (144). When insulin is chronically in-jected ICV in low doses, intake and BW are reduced

INGVARTSEN AND ANDERSEN1584
Figure 6. The changes in daily food intake in dairy cows duringa long-term hyperinsulinemic-euglycemic clamp. The studies in-cluded are: 1 McGuire et al. (218), 2 Griinari et al. (130), 3 Marcleet al. (212), 4 Annen et al. (6), 5 Giesy and McGuire, preliminarydata, 6 preliminary data from authors lab. The daily intakes givenare as DM except in study four, which are given in kilograms as-fed.
in rats (42, 54, 160, 217, 253, 262, 326), baboons (338),and sheep (116). Insulin antibodies administered ICVincreased intake and weight gain (218, 308). Strubbeand Mein (308) observed that injection of insulin anti-bodies into the ventromedial nucleus stimulated noc-turnal feeding in rats, whereas injection in the lateralhypothalamus had no effect, suggesting that endoge-nous insulin at the ventromedial nucleus acts to inhibitfeeding. The responses to ICV infusions of insulin onintake and BW in rats seem dependent on the nutrientcontent of the diet (55). The largest effect on intake wasobserved when rats were given a high carbohydratediet. A minor response was observed on fat-based diets.Insulin can bind to insulin-like growth factor I (IGF-I)receptors and mimic the response of IGF-I. However,food intake is most likely not affected via IGF-I-recep-tors since Foster et al. (116) found the level of intake tobe depressed when infusing ICV insulin but not IGF-I.
Acute peripheral infusion of insulin causing hypogly-cemia decreased food intake in monogastrics (131) andruminants (5, 79). The decreased intake was most likelydue to hypoglycemia since glucose infusion preventsinsulin-induced hypophagia in man (169) and in rumi-nants (155). Chronic infusion of insulin in situations ofnormo- or hyperglycemia reduces intake in rats (326).In dairy cows hyperinsulinemic euglycemic clamp tech-niques applied for long-term infusions (4 d) generallydepress intake as shown in Figure 6 while short-term (4h) infusions under euglycemic conditions did not affectintake (17).
Journal of Dairy Science Vol. 83, No. 7, 2000
In summary, insulin may play a role in long-termintake and weight regulation in ruminants like in thestudies of Vandermeerschen-Doize et al. (323) andMcCann et al. (216). In the dairy cow, insulin may playa role in the dip in intake in late pregnancy, but itis unlikely that insulin plays any significant role indepressing intake in dairy cows in early lactation whenthe insulin concentration is low.
Glucagon and GLP-1
Proglucagon is expressed primarily in the α-cells ofthe pancreatic islands and in the endocrine cells of thegastrointestinal mucosa. In the human pancreas, pro-glucagon is the precursor of glucagon and a number ofproglucagon fragments, while proglucagon in the smallintestine is the precursor of glicentin, GLP-1, GLP-2and other fragments (2, 152). Here we will only addressglucagon and GLP-1 because these are the biologicallyactive peptides (152).
An early report (282) demonstrated that glucagoncould decrease food intake in humans. Later studieshave confirmed that glucagon reduces food intake inrats when administered peripherally (127) or centrally(168). But glucagon is most likely not acting directlyon the brain but rather on the liver since the depressiveeffect of glucagon could be prevented by hepatic vagot-omy (126, 214). Furthermore, glucagon is more potentwhen administered into the hepatic portal vein thaninto the inferior vena cava (125). When antibodiesagainst glucagon are given intraperitoneally to ratsprior to feeding, meal size and duration were enhanced,indicating that glucagon may act as satiety factor (199).
The sequence of GLP-1 (7-36) is completely conservedin all mammalian species studied, implying that it playsa critical physiological role (136). In 1996, Turton et al.(319) showed that GLP-1 powerfully inhibits feeding infasted rats and suggested that central GLP-1 is a newphysiological mediator of satiety via the paraventricu-lar nucleus of the hypothalamus and central nucleusof the amygdala. These centers are probably stimulatedvia peripheral GLP-1 passing through BBB leaks (240)or vagal pathways (2). Subsequent studies have con-firmed the profound inhibitory effect of GLP-1 on foodintake in rats through ICV administration of GLP-1 oran antagonist to GLP-1, exendin (159, 224, 317, 332).
Only few investigations are available in ruminantson the influence of glucagon and GLP-1 on food intake.Glucagon administered intravenously at physiologicalconcentrations does reduce intake in sheep (80). Circu-lating concentrations of GLP-1 are significantly higherin lactating compared to dry sheep (99). The increasedcirculating concentration during lactation is most likelydue to increased secretion, probably resulting from the

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1585
increased feed intake during lactation, since the half-life of GLP-1 does not change due to lactation. Clearly,more research is needed in ruminants on the effect ofglucagon and GLP-1 to unravel their importance inintake regulation.
Cholecystokinin
Cholecystokinin was first isolated from the porcinegastrointestinal tract (235). The CCK is perhaps thebest-studied putative endogenous satiety signal andhas been considered “the prototypic peripheral satietyhormone” (230). Therefore, it is not surprising that alarge number of reviews on CCK are available (12, 13,74, 206, 207, 279, 290, 300) in which the reader mayfind detailed information on the biology of CCK. Conse-quently, we will only give a very brief summary inthe following.
The CCK is primarily secreted from the duodenumand jejunum, but CCK-secreting cells have been local-ized throughout the gut and in the CNS, including sev-eral brain areas (13). Many different molecular formsof CCK, molecules of 58, 39, 33, 25, 22, 18, 8, 7, 5, and4 AA, have been isolated in the intestine, brain, andcirculation of multiple species. These different formsmay have different biological activities (207). Ingestionof food and particularly the digestion products of fat andprotein are potent stimuli of CCK secretion, althougheffects of these stimulators differ among species (43).Also, there is increasing evidence for a CCK-releasingfactor of intestinal origin (206). In ruminants, Choi andPalmquist (62) tested the effect of fat feeding on plasmaCCK concentrations in cows and observed a dose-depen-dent increase in CCK 3 h postfeeding. Furuse et al.(121), however, did not observe any differences inplasma CCK concentration around meals of concen-trates and hay in cows, which might reflect the continu-ous flow of digesta from the rumen or just the limitednumber of cows (n = 3) in this study. The numerousbiological actions of CCK (74) are now known to actthrough two different receptors, a peripheral type(CCK-A receptor) and a brain type (CCK-B receptor)(207).
The CCK may cause satiety via various peripheralmechanisms such as smooth muscle contractile effects,decreased gastric emptying, and direct vagal stimula-tion (290). The direct vagal effect on satiety suggeststhat food in the intestine causes the release of CCK,which acts on CCK-A receptors in the vagus nerve toprovide sensory information to the VMH and PVN. Thesatiety effect of CCK varies with species, age, feeding,and experimental conditions (13, 74, 114, 279, 290, 300).In sheep, ICV injection of different doses of CCK-8,believed to be within the physiological range, caused a
Journal of Dairy Science Vol. 83, No. 7, 2000
dose-dependent hypophagia (81, 82) and decreasedmeal size (83). The ICV injections of CCK-8 also de-creased the rate and amplitude of rumen contractions(46, 83). Larger forms of the CCK molecule (CCK-33)likewise cause satiety when administered ICV in sheep(84). The most convincing evidence for a satiety role ofCCK is the ICV injection of specific CCK-antiserumthat caused a significant hyperphagia in sheep (85).
Somatostatin
Brazeau et al. (40) isolated and characterized a hypo-thalamic peptide with a potent inhibitory effect on so-matotropin release. Somatostatin (SS), a tetradecapep-tide, is not only secreted in the brain but also in thegut (341). Via neuroendocrine, exocrine, paracrine, andendocrine mechanisms, SS plays numerous regulatoryroles in the organism, including regulation of feed in-take. Manipulation of the SS concentration in the brainindicates a central depressing effect of SS on intakein most experiments with rodents (7, 151, 208) andruminants (303, 304), although not in all (203, 209).Peripherally administered SS has likewise been shownto reduce feeding in rats and baboons (203, 209) by avagal mediated mechanism indicating a link betweenthe gastrointestinal tract and the central regulation offeed intake in the brain. However, SS is effective onlyin animals with mild degrees of hunger (203). Immuni-zation against SS in cattle also indicates that circulat-ing SS reduces VDMI. In a meta-analytic evaluation of11 studies, Ingvartsen and Sejrsen (167) found thatgrowing cattle immunized against SS consumed moreDM (4.2 ± 1.4%), had a higher daily gain (11.4 ± 2.3%),and improved feed conversion ratio (3.3 ± 2.0%) com-pared to controls.
SIGNALS FROM THE IMMUNE SYSTEMREGULATING INTAKE
Recently, Weingarten (334) argued for the impor-tance of the immune system in ingestive behavior. Theimmune system can cause hypophagia via cytokines,which are polypeptide mediators produced by a varietyof immune cells. The cytokines that most profoundlyaffect intake are those involved in the acute phase in-flammatory responses (APR), tumor necrosis factor-α(TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6) (172, 192), but cytokines such as interferons, IL-8and other chemokines or intercrines may also inducehypophagia (250). A commonly used model for studyingthe effects of APR cytokines is to use lipopolysaccharide(LPS) administration as a tool to provoke APR. Indeed,LPS-administration decreases food intake and BW inhamsters (135), rats (251), mice (97, 98), and cattle (92).

INGVARTSEN AND ANDERSEN1586
Tumor Necrosis Factor-α
The hypophagic effect of cytokines has been demon-strated by peripheral or central injection of cytokinesor by blocking their action. The TNF-α reduces intakein rodents when administered intraperitoneally (29).The administration of antibodies against TNF-α en-hanced food intake in tumor-bearing rats, although theeffect was not dramatic and was only statistically sig-nificant on d 2 after injection (299). Both acute (254)and chronic (256) ICV infusion of TNF-α reduce intake,but not to the extent of IL-1β (256). In pigs, both LPS-administration and ICV infusion of TNF-α reduce in-take (333). In cattle, low doses (<0.5 mg/100 kg) of re-combinantly derived bovine TNF-α caused no effect,while prolonged treatment with higher doses (>0.5 mg/100 kg) caused hypophagia, depression, cachexia, anddiarrhea (32). The hypophagic effect of TNF-α on intakein ruminants may, in part, be due to a TNF-mediatedinhibition of rumen motility (321).
Interleukin-1-β
The IL-1 reduces intake when acutely administeredto rats peripherally (197, 221) or ICV (249, 254). Simi-larly, chronic administration of IL-1β ICV to rats for 3to 7 d reduces intake considerably (101, 150). The ICVtreatment with IL-1 receptor antagonist concomitantlywith IL-1β (252) or in relation to colitis (220) has alsoshown the role of IL-1β in hypophagia and that it ismediated by a direct action in the CNS. The hypophagiafollowing an IL-1β administration is due to a reductionin both meal numbers and their size (249). Very littleinformation is available on IL-6 and food intake, butICV administration of IL-6 and a soluble IL-6 receptorin rats indicates a role of IL-6 in intake regulation (280).
Acute phase inflammatory response cytokines proba-bly act adaptively or synergistically on intake (302)and may also interact with leptin. An increased leptinexpression is observed after an LPS administration(135, 274) probably caused by TNF-α that increasesleptin secretion (102). Leptin is also increased by exper-imentally induced infections in rats (ileitis) (15) andmice (peritonitis) (232). This suggests that leptin maybe involved in the APR and hypophagia in relation toinflammation. It is possible that leptin participates inthe host response to inflammation by modulating thehost immune and cytokine responses (96), but leptinper se is not essential in the hypophagia caused byinfections since LPS-treatment of leptin-receptor defi-cient (db/db) mice also caused hypophagia (98).
Not only are the cytokines, such as TNF-α, IL-1-β,and IL-6 affecting intake, they are also having a majorimpact on metabolism. It is beyond the scope of this
Journal of Dairy Science Vol. 83, No. 7, 2000
Figure 7. A simplified diagram showing putative peripheral sig-nals involved in intake regulation in dairy cattle. Humoral signals(full lines) due to pregnancy, estrus, adipose tissue mass, pancreatichormones, and cytokines from the immune system may down-regu-late intake. Progesterone has no direct effect on intake but blocksthe depressive effects of estrogen on intake. The gut peptides chole-cystokinin (CCK), somatostatin (SS) and glucagon-like peptide-1(GLP-1) is secreted from the small intestine into the circulationwhere they act on specific receptors of vagal efferents (broken lines).Stimulation of these afferents activates central neural pathwaysresulting in down-regulation of intake. Gut peptides, at least GLP-1, may also cross the blood-brain barrier and act via receptors in thebrain. The liver senses metabolic processes like β-oxidation (filleddiamond) providing satiety signals mediated by vagal afferents (bro-ken lines).
review to address this interesting field here and readersare referred to recent reviews (93, 94).
INTEGRATION OF METABOLISM ANDINTAKE REGULATION
In Figure 7, a simplified diagram showing putativeperipheral signals involved in intake regulation in dairycattle is given. In the following, we will briefly touchon neuropeptides that may be involved in the regulationof intake and their role in integrating peripheral signalsand metabolism.
Central Mechanisms and Integration
Although much remains unknown, some progress hasbeen made in understanding how the brain processesperipheral information via neuropeptides. We will notattempt to review all the neuropeptides potentially in-volved in intake regulation (see Table 3) but will focuson some intake-stimulating neuropeptides such as neu-ropeptide Y (NPY) and galanin believed to be importantin mediating the effects of peripheral signals to thebrain. We have already dealt with some intake de-

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1587
pressing gut peptides (GLP-1, CCK, SS) and the neuro-peptide CRF and will only touch on these in order toexplain aspects of integration.
The NPY is found in cell bodies and nerve terminalsof many areas of the brain, particularly those involvedin intake regulation and energy balance. Acute injec-tions of NPY ICV cause hyperphagia in satiated rats(33, 65, 222) and sheep (226, 227). Chronically adminis-tered NPY over 7 d (53) results in sustained hyperpha-gia and increased body fat accumulation in rats (343),while chronic ICV administration of NPY-antibodiescauses hypophagia (88). When administered centrally,NPY is one of the most potent inducers of food intakeand is able to increase intake and daily gain twofoldand sixfold, respectively (305). Like NPY, galanin iswidespread in the brain, but with particularly high con-centrations in the PVN. In rats, ICV injection of galanincauses hyperphagia (189), often with a preference forfat (190, 315). We have found no information on galan-in’s effect on feeding in ruminants.
There is now evidence for an activation of the ARN-PVN NPY pathway in response to signals associatedwith negative energy balance. Negative energy balancedue to feed restriction (1, 174, 270), intense exercise(205), or lactation (301) is accompanied by increasedlevels of NPY mRNA or rise in concentration of thepeptide itself in different brain regions. It is speculatedthat NPY plays an important role in the increasedVDMI in early lactation.
The response to negative energy balance is, at leastin part, mediated by reduced negative feedback fromleptin and insulin. Leptin administered centrally re-duces the NPY expression and synthesis in the hypo-thalamus of rodents (75, 269, 283, 285) and ewes (147).However, leptin does not only mediate its effect on in-take via NPY. In studies with ob/ob mice and wild-typemice in which NPY was knocked out, the ob/ob micedid not completely normalize their weight and the wild-type lacking the NPY still controlled intake and BWnormally (241). Other mediators of leptin action suchas proopiomelanocortin, melanocortin stimulating hor-mone, and aguti-related peptide are likely to be in-volved as indicated in recent reviews (106, 241). Indeed,Sahu (269) has shown that central administration ofleptin decreased food intake and weight gain in associa-tion with a decrease in hypothalamic galanin, melano-cortin-stimulating hormone, proopiomelanocortin, andNPY gene expression and an increase in neurotensinegene expression. Furthermore, CRF seems to be in-volved since leptin administered to rodents peripherally(73) or centrally (269, 285) increases CRF expressionor secretion. The intake depressive effects of insulinmay also be mediated, at least in part, via NPY since
Journal of Dairy Science Vol. 83, No. 7, 2000
ICV infusion of insulin can inhibit the rise in NPYmRNA levels that usually occurs during fasting (287).
Leptin and Neuropeptides as Integratorsof Intake and Metabolism
In previous sections, we have discussed the role ofboth peripheral and central signals in intake regula-tion. Clearly, all the peripheral signals discussed inprevious sections exert a variety of other biological func-tions apart from being involved in intake regulation.As an example, leptin also affects energy expenditure,glucose metabolism, insulin secretion and action, theadrenal axis and hormones of the growth hormone axisas discussed in recent reviews (37, 156, 158, 328). Mostof these metabolic processes may not necessarily bedirectly affected by leptin since the effect could be medi-ated via leptin-induced changes in NPY and CRF (37,336).
There is little doubt that animal intake is highlyintegrated with its metabolism and environment andthat this integration takes place in the brain. However,the majority of the information available is on rodentsor at least monogastrics. Differences among species cannot be ruled out and much work has yet to be done toimprove our understanding of intake regulation and itsintegration with metabolism, particularly in cattle.
CONCLUDING REMARKS AND PERSPECTIVES
In the present review we have focused on the dip inintake that occurs around calving and the mechanismsand signals that may mediate this dip. The dip in intakein periparturient cattle coincides with adaptations inmetabolism to the advancing pregnancy and forthcom-ing lactation. This dip in intake occurs not only in rumi-nants but also in monogastric animals such as the ratand seems to be a normal adaptive event in peripartur-ient animals. However, in some circumstances the mag-nitude and duration of the dip can increase consider-ably, possibly influencing the health and well-being ofthe periparturient cow.
Physical constraints play an important role in intakeregulation, but we believe that its role has been overem-phasized and have described metabolic factors that mayplay an equally important role in intake regulation inthe periparturient cow. These factors include nutrients,metabolites, reproductive hormones, stress hormones,leptin, insulin, gut peptides, cytokines, and neuropep-tides such as NPY, galanin, and CRF. The dip in intakein late pregnancy may be mediated via body reservesand endocrine factors in response to the advancingpregnancy. Leptin and insulin are likely to play anincreasing role in the concurrent increase in body fat-

INGVARTSEN AND ANDERSEN1588
ness. Also, estrogen reduces intake during the latestages of pregnancy and the first days of lactation. Themechanisms involved in intake regulation in early lac-tation are not very well understood. The general in-crease in intake in early lactation is speculated to bedue to an increase in neuropeptides, such as NPY andgalanin. Furthermore, a reduction in intake depressingsignals from hormones such as leptin and insulin maylikewise stimulate and increase intake in early lacta-tion. Nutrients and their metabolism may also influenceintake. Particularly, feedback signals from the oxida-tion of NEFA in the liver are speculated to down-regu-late intake in late pregnancy and early lactation whenmobilization is high. This mechanism may also, at leastin part, explain the lower intake in fat cows mobilizingmore body reserves than thin cows in early lactation.Little is known in ruminants on how specific nutrientsaffect the release of satiety peptides from the gut. How-ever, it seems likely that the large variation in intakereduction observed when different fat sources are in-fused into the abomasum may be due to differencesin their ability to stimulate, e.g., CCK secretion. Theimmune system may also affect intake during infection.During the APR, cytokines that may severely reduceintake are released.
The integration of signals involved in intake regula-tion takes place in the brain. The effect of several ofthe peripheral signals is mediated via central neuropep-tides. The neuropeptides are not only involved in thecentral regulation and integration of intake but alsoexert multiple effects on metabolism, which indicatestheir role in integrating metabolism and intake regu-lation.
Whether factors affecting intake in monogastricshave similar effects in the dairy cow needs to be tested.Furthermore, there is a need for long-term studies giv-ing detailed information on intake, performance, andphysiological parameters relevant for forming conceptson the desire of the dairy cow to store and mobilize bodyreserves at different physiological states. Physiologicalmeasures reflecting, e.g., body reserves and mobiliza-tion in early lactation have the potential to provideuseful physiological indicators of abnormally low intakeand subsequent metabolic stress. A better understand-ing of the mechanisms in intake regulation, includingits integration with metabolism, should result in im-proved animal selection and feed management and,thereby, better performance, health, and animal well-being.
Before trying to improve feed management, weshould try to understand the desire of the animal. Forexample, is the cytokine-mediated hypophagia associ-ated with infection beneficial to the animal? Hypopha-gia may seem somewhat paradoxical since it may lead
Journal of Dairy Science Vol. 83, No. 7, 2000
to metabolic diseases such as fatty liver and ketosis.However, the behavior of sick animals may be an orga-nized strategy that facilitates recovery and survivalof the animal and not a maladaptive response to thedisease. This has been exemplified in the study of Mur-ray and Murray (234) who studied the role of hypopha-gia as a mechanism of host defense mechanisms. Theydid so by comparing mortality in Listeria-infected micewith that of Listeria-infected mice force-fed to the sameenergy intake as in control mice. Ad libitum fed infectedmice ate 58% of controls. Force-feeding of infected miceincreased mortality (93 vs. 43%) and shortened survivaltime (3.9. vs. 8.7 d), suggesting that hypophagia playsa significant role in the early host defense and survival.
A better understanding of the mechanisms regulat-ing intake and metabolism, and particularly their inte-gration, is important for the development of better con-cepts in predictive models for intake, production, energybalance and health—not implying, though, that it isnecessary to make models more complicated by addingnumerous physiological parameters. To advance ourunderstanding of intake regulation, and ability to ma-nipulate intake in the periparturient cow, we need tofurther characterize the adaptive changes occurring inmetabolism around parturition and the desire of cowsto mobilize body tissue in support of lactation. Informa-tion is lacking on a number of important areas of intakeregulation in dairy cattle, as indicated in this review.We hope that some of the issues and assumptions putforward in this paper will stimulate further critical ex-periments which in turn will lead to a better under-standing of intake regulation in early lactation and itsintegration with metabolism.
ACKNOWLDGEMENTS
We wish to thank Dale Bauman, Mark McGuire,Miko Griinari, and Tim Marcle for giving us access tofeed intake data from their studies on insulin using thehyperinsulinemic-euglycemic clamp technique. Fur-thermore, we thank Karin V. Østergaard for her skilledassistance in preparing the manuscript and the staffat our library for providing us with the large numberof articles used for this review. The Danish Ministry ofFood, Agriculture and Fisheries is acknowledged forthe funding of a previous project (GRU-SH-2) importantfor this work and support to the projects HUS97-2 andHUS97-3.
REFERENCES
1 Ahima, R. S., D. Prabakaran, C. Mantzoros, D. Qu, B. B. Lowell,E. Maratos-Flier, and J. S. Flier. 1996. Role of leptin in theneuroendocrine response to fasting. Nature 382:250–252.

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1589
2 Ahren, B. 1998. Glucagon-like peptide-1 (GLP-1): a gut hor-mone of potential interest in the treatment of diabetes. BioEs-says 20:642–651.
3 Alexiou, C., J. P. Zimmermann, R. R. Schick, and V. Schusdzi-arra. 1998. Xenin—a novel suppressor of food intake in rats.Brain Res. 800:294–299.
4 Amico, J. A., A. Thomas, R. S. Crowley, and L. A. Burmeister.1998. Concentrations of leptin in the serum of pregnant, lactat-ing, and cycling rats and of leptin messenger ribonucleic acidin rat placental tissue. Life Sci. 63:1387–1395.
5 Anil, M. H., and J. M. Forbes. 1980. Feeding in sheep duringintraportal infusions of short-chain fatty acids and the effectof liver denervation. J. Physiol. 298:407–414.
6 Annen, E. L., M. A. McGuire, T. W. Hanson, and D. E. Bauman.1998. Milk protein production in cows subjected to abomasalinfusion of branched-chain amino acids (BCAA) and hyperin-sulinemic-euglycemic clamp. J. Dairy Sci. 81:354(Abstr.).
7 Aponte, G., P. Leung, D. Gross, and T. Yamada. 1984. Effectsof somatostatin on food intake in rats. Life Sci. 35:741–746.
8 Arase, K., J. S. Fisler, N. Shargill, D. A. York, and G. A. Bray.1988. Intraventricular infusions of 3-hydroxybutyrate and in-sulin in a rat model of dietary obesity. Am. J. Physiol.255:R974–R981.
9 Arase, K., D. A. York, H. Shimizu, N. Shargill, and G. Bray.1988. Effect of corticotropin releasing factor on food intake andbrown adipose tissue thermogenesis in rats. Am. J. Physiol.255:E255–E259.
10 Bagdade, J. D., E. L. Bierman, and D. Porte, Jr. 1967. Thesignificance of basal insulin levels in the evaluation of the insu-lin response to glucose in diabetic and non-diabetic subjects.J. Clin. Invest. 46:1549–1557.
11 Baile, C. A., and H. F. Martin. 1971. Hormones and aminoacids as possible factors in the control of hunger and satietyin sheep. J. Physiol. 54:897–905.
12 Baile, C. A., and C. L. McLaughlin. 1987. Mechanisms control-ling feed intake in ruminants: a review. J. Anim. Sci.64:915–922.
13 Baile, C. A., C. L. McLaughlin, and M. A. Della-Fera. 1986.Role of cholecystokinin and opioid peptides in control of foodintake. Physiol. Rev. 66:172–234.
14 Barb, C. R., X. Yan, M. J. Azain, R. R. Kraeling, G. B. Rampacek,and T. G. Ramsay. 1998. Recombinant porcine leptin reducesfeed intake and stimulates growth hormone secretion in swine.Domest. Anim. Endocrinol. 15:77–86.
15 Barbier, M., C. Cherbut, A. C. Aube, H. M. Blottiere, and J. P.Galmiche. 1998. Elevated plasma leptin concentrations in earlystages of experimental intestinal inflammation in rats. Gut43:783–790.
16 Bareille, N., and P. Faverdin. 1996. Lipid metabolism and in-take behavior of dairy cows: effects of intravenous lipid andbeta-adrenergic supplementation. J. Dairy Sci. 79:1209–1220.
17 Bareille, N., and P. Faverdin. 1996. Modulation of the feedingresponse of lactating dairy cows to peripheral insulin adminis-tration with or without a glucose supply. Reprod. Nutr. Dev.36:83–93.
18 Barrachina, M. D., V. Martinez, J. Y. Wei, and Y. Tache. 1997.Leptin-induced decrease in food intake is not associated withchanges in gastric emptying in lean mice. Am. J. Physiol.272:R1007–R1011.
19 Baskin, D. G., D. P. Figlewicz, S. C. Woods, D. Porte, Jr., andD. M. Dorsa. 1987. Insulin in the brain. Annu. Rev. Physiol.49:335–347.
20 Baskin, D. G., B. J. Wilcox, D. P. Figlewicz, and D. M. Dorsa.1988. Insulin and insulin-like growth factors in the CNS.Trends Neurosci. 11:107–111.
21 Bauche, F., D. Sabourault, Y. Giudicelli, J. Nordmann, and R.Nordmann. 1981. 2–Mercaptoacetate administration depressesthe β-oxidation pathway through an inhibition of long-chainacyl-CoA dehydrogenase activity. Biochem. J. 196:803–809.
22 Bauman, D. E. 1992. Bovine somatotropin: review of an emerg-ing animal technology. J. Dairy Sci. 75:3432–3451.
Journal of Dairy Science Vol. 83, No. 7, 2000
23 Bauman, D. E., and W. B. Currie. 1980. Partitioning of nutri-ents during pregnancy and lactation: a review of mechanismsinvolving homeostasis and homeorhesis. J. Dairy Sci.63:1514–1529.
24 Bauman, D. E., P. J. Eppard, M. J. DeGeeter, and G. M. Lanza.1985. Responses of high-producing dairy cows to long-termtreatment with pituitary somatotropin and recombinant soma-totropin. J. Dairy Sci. 68:1352–1362.
25 Baura, G. D., D. M. Foster, K. Kaiyala, D. Porte, S. E. Kahn,and M. W. Schwartz. 1996. Insulin transport from plasma intothe central nervous system is inhibited by dexamethasone indogs. Diabetes 45:86–90.
26 Baura, G. D., M. W. Schwartz, D. M. Foster, D. Porte, Jr.,S. E. Kahn, R. Bergman, and C. Cobelli. 1993. Saturable trans-port of insulin from plasma into the central nervous system indogs in vivo. A mechanism for regulating insulin delivery tothe brain. J. Clin. Invest. 92:1824–1830.
27 Bell, A. W. 1995. Regulation of organic nutrient metabolismduring transition from late pregnancy to early lactation. J.Anim. Sci. 73:2804–2819.
28 Bell, A. W., and D. E. Bauman. 1997. Adaptations of glucosemetabolism during pregnancy and lactation. J. Mamm. GlandBiol. Neopl. 2:265–278.
29 Bernstein, I. L., E. M. Taylor, and K. L. Bentson. 1991. TNF-Induced anorexia and learned food aversions are attenuatedby area postrema lesions. Am. J. Physiol. 260:R906–R910.
30 Beverly, J. L., and R. J. Martin. 1991. Influence of fatty acidoxidation in lateral hypothalamus on food intake and bodycomposition. Am. J. Physiol. 261:R339–R343.
31 Bidwell, C. A., S. Ji, G. R. Frank, S. G. Cornelius, G. M. Willis,and M. E. Spurlock. 1997. Cloning and expression of the porcineobese gene. Anim. Biotech. 8:191–206.
32 Bielefeldt, O. H., M. Campos, M. Snider, N. Rapin, T. Beskor-wayne, Y. Popowych, M. J. Lawman, A. Rossi, and L. A. Babiuk.1989. Effect of chronic administration of recombinant bovinetumor necrosis factor to cattle. Vet. Pathol. 26:462–472.
33 Billington, C. J., J. E. Briggs, M. Grace, and A. S. Levine. 1991.Effects of intracerebroventricular injection of neuropeptide Yon energy metabolism. Am. J. Physiol. 260:R321–R327.
34 Bines, J. A. 1979. Voluntary food intake. Pages 23–48 in Feed-ing Strategy for the High Yielding Dairy Cow. W. H. Brosterand H. Swan, ed. Granada Publishing, London, UK.
35 Bines, J. A., and S. V. Morant. 1983. The effect of body conditionon metabolic changes associated with intake of food by the cow.Br. J. Nutr. 50:81–89.
36 Bines, J. A., S. Suzuki, and C. C. Balch. 1969. The quantitativesignificance of long-term regulation of food intake in the cow.Br. J. Nutr. 23:695–704.
37 Blum, W. F., P. Englaro, A. M. Attanasio, W. Kiess, and W.Rascher. 1998. Human and clinical perspectives on leptin. Proc.Nutr. Soc. 57:477–485.
38 Boden, G., G. Chen, M. Mozzoli, and I. Rayan. 1996. Effect offasting on serum leptin in normal human subjects. J. Clin.Endocrinol. Metab. 81:3419–3423.
39 Bray, G. A., and D. A. York. 1998. The MONA LISA hypothesisin the time of leptin. Recent Progr. Horm. Res. 53:95–117.
40 Brazeau, P., W. Vale, R. Burgus, N. Ling, M. Butcher, J. Rivier,and R. Guillemin. 1973. Hypothalamic polypeptide that inhibitsthe secretion of immunoreactive pituitary growth hormone. Sci-ence 179:77–79.
41 Bremmer, D. R., L. D. Ruppert, J. H. Clark, and J. K. Drackley.1998. Effects of chain length and unsaturation of fatty acidmixtures infused into the abomasum of lactating dairy cows.J. Dairy Sci. 81:176–188.
42 Brief, D. J., and J. D. Davis. 1984. Reduction of food intakeand body weight by chronic intraventricular insulin infusion.Brain Res. Bull. 12:571–575.
43 Brodish, R. J., B. W. Kuvshinoff, A. S. Fink, J. Turkelson,D. W. McFadden, and T. E. Solomon. 1994. Intraduodenal acidaugments oleic acid (C18)-induced cholecystokinin release.Ann. N.Y. Acad. Sci. 713:390.

INGVARTSEN AND ANDERSEN1590
44 Broster, W. H., and V. J. Broster. 1984. Long term effects ofplane of nutrition on the performance of the dairy cow. J. DairyRes. 51:149–196.
45 Broster, W. H., and V. J. Broster. 1998. Body score of dairycows. J. Dairy Res. 65:155–173.
46 Bueno, L., A. Duranton, and Y. Ruckebusch. 1983. Antagonisticeffects of naloxone on CCK-octapeptide induced satiety andrumino-reticular hypomotility in sheep. Life Sci. 32:855–863.
47 Burton, J. L., B. W. McBride, E. Block, D. R. Glimm, and J. J.Kennelly. 1994. A review of bovine growth hormone. Can. J.Anim. Sci. 74:167–201.
48 Butera, P. C., and R. J. Beikirch. 1989. Central implants ofdiluted estradiol: Independent effects on ingestive and repro-ductive behaviours of ovariectomized rats. Brain Res.491:266–273.
49 Campfield, L. A., F. J. Smith, Y. Guisez, R. Devos, and P. Burn.1995. Recombinant mouse OB protein: evidence for a peripheralsignal linking adposity and central neural networks. Science269:546–549.
50 Caro, J. F., M. K. Sinha, J. W. Kolaczynski, P. L. Zhang, and R.V. Considine. 1996. Leptin: the tale of an obesity gene. Diabetes45:1455–1462.
51 Carpenter, R. G., and S. P. Grossman. 1983. Plasma fat metabo-lites and hunger. Physiol. Behav. 30:57–63.
52 Carpenter, R. G., and S. P. Grossman. 1983. Reversible obesityand plasma fat metabolites. Physiol. Behav. 30:51–55.
53 Catzeflis, C., D. D. Pierroz, F. Rohner-Jeanrenaud, J. E. Rivier,P. C. Sizonenko, and M. L. Aubert. 1993. Neuropeptide Y ad-ministered chronically into the lateral ventricle profoundly in-hibits both the gonadotropic and the somatotropic axis in intactadult female rats. Endocrinology 132:224–234.
54 Chavez, M., K. Kaiyala, L. J. Madden, M. W. Schwartz, andS. C. Woods. 1995. Intraventricular insulin and the level ofmaintained body weight in rats. Behav. Neurosci. 109:528–531.
55 Chavez, M., C. A. Riedy, G. Van Dijk, and S. C. Woods. 1997.Central insulin and macronutrient intake in the rat. Am. J.Physiol. 271:R727–R731.
56 Chen, H., O. Charlat, L. A. Tartaglia, E. A. Woolf, X. Weng,S. J. Ellis, N. D. Lakey, J. Culpepper, K. J. Moore, R. E. Breitb-art, G. M. Duyk, R. I. Tepper, and J. P. Morgensten. 1996.Evidence that the diabetis gene encodes the leptin receptor:Identification of a mutation in the leptin receptor gene in db/db mice. Cell 84:491–495.
57 Chien, E. K., M. Hara, M. Rouard, H. Yano, M. Philippe, K. S.Polonsky, and G. I. Bell. 1997. Increase in serum leptin anduterine leptin receptor messenger RNA levels during pregnancyin rats. Biochem. Biophys. Res. Commun. 2237:476–480.
58 Chilliard, Y. 1989. Long-term effects of recombinant bovinesomatotropin (rBST) on dairy cow performance: a review. Pages61–87 in Use of Somatotropin in Livestock Production. K. Sejr-sen, M. Vestergaard, and A. Neimann-Sørensen, ed. ElsevierAppl. Sci., New York, NY.
59 Chilliard, Y., F. Bocquier, C. Delavaud, M. Guerre-Millo, M.Bonnet, P. Martin, Y. Faulconnier, and A. Ferlay. 1998. Leptinin ruminants: effects of species, breed, adiposity, photoperiod,beta-agonists and nutritional status. Pages 65–74 in CornellNutrition Conference for Feed Manufacturers, Syracuse, NY.Cornell Univ., Ithaca, NY.
60 Chilliard, Y., and M. Doreau. 1997. Influence of supplementaryfish oil and rumen-protected methionine on milk yield and com-position in dairy cows. J. Dairy Sci. 64:173–179.
61 Chilliard, Y., G. Gagliostro, A. Ollier, and D. Bauchart. 1990.Effects of rapeseed oil duodenal infusion on dairy cow perfor-mances and blood metabolites during early lactation. Reprod.Nutr Dev. Suppl. 2:229(Abstr.).
62 Choi, B. R., and D. L. Palmquist. 1996. High fat diets increaseplasma cholecystokinin and pancreatic polypeptide, and de-crease plasma insulin and feed intake in lactating cows. J.Nutr. 126:2913–2919.
63 Choi, B. R., D. L. Palmquist, and M. S. Allen. 1997. Sodiummercaptoacetate is not a useful probe to study the role of fat inregulation of feed intake in dairy cattle. J. Nutr. 127:171–176.
Journal of Dairy Science Vol. 83, No. 7, 2000
64 Christensen, R. A., J. K. Drackley, D. W. LaCount, and J. H.Clark. 1994. Infusion of four long-chain fatty acid mixturesinto the abomasum of lactating dairy cows. J. Dairy Sci.77:1052–1069.
65 Clark, J. T., P. S. Kalra, W. R. Crowley, and S. P. Kalra. 1984.Neuropeptide Y and human pancreatic polypeptide stimulatefeeding behavior in rats. Endocrinology 115:427–429.
66 Coffey, K. P., J. A. Paterson, C. S. Saul, L. S. Coffey, K. E.Turner, and J. G. Bowman. 1989. The influence of pregnancyand source of supplemental protein on intake, digestive kineticsand amino acid absorption by ewes. J. Anim. Sci. 67:1805–1814.
67 Coleman, D. L. 1973. Effects of parabiosis of obese with diabetesand normal mice. Diabetologia 9:294–298.
68 Coleman, D. L., and K. P. Hummel. 1969. Effects of parabiosisof normal with genetically diabetic mice. Am. J. Physiol.217:1298–1304.
69 Considine, R. V., and J. F. Caro. 1997. Leptin and the regulationof body weight. Int. J. Biochem. Cell Biol. 29:1255–1272.
70 Considine, R. V., M. K. Sinha, M. L. Heiman, A. Kriauciunas,T. W. Stephens, M. R. Nyce, J. P. Ohannesian, C. C. Marko,L. J. McKee, T. L. Bauer, and J. F. Caro. 1996. Serum immuno-reactive-leptin concentrations in normal-weight and obese hu-mans. New Engl. J. Med. 334:292–295.
71 Coppock, C. E., C. H. Noller, and S. A. Wolfe. 1974. Effect offorage-concentrate ratio in complete feeds fed ad libitum onenergy intake in relation to requirements by dairy cows. J.Dairy Sci. 57:1371–1380.
72 Coppock, C. E., C. H. Noller, S. A. Wolfe, C. J. Callahan, andJ. S. Baker. 1972. Effect of forage-concentrate ratio in completefeeds fed ad libitum on feed intake prepartum and the occur-rence of abomasal displacement in dairy cows. J. Dairy Sci.55:783–789.
73 Costa, A., A. Poma, E. Martignoni, G. Nappi, E. Ur, and A.Grossman. 1997. Stimulation of corticotrophin-releasing hor-mone release by the obese (ob) gene product, leptin, from hypo-thalamic explants. NeuroReport 8:1131–1134.
74 Crawley, J. N., and R. L. Corwin. 1994. Biological actions ofcholecystokinin. Peptides 15:731–755.
75 Cusin, I., F. Rohner-Jeanrenaud, A. Stricker-Krongrad, and B.Jeanrenaud. 1996. The weight-reducing effect of an intracere-broventricular bolus injection of leptin in genetically obese fa/fa rats. Reduced sensitivity compared with lean animals. Diabe-tes 45:1446–1450.
76 Dagogo, J. S., C. Fanelli, D. Paramore, J. Brothers, and M.Landt. 1996. Plasma leptin and insulin relationships in obeseand nonobese humans. Diabetes 45:695–698.
77 Dagogo, J. S., S. C. Franklin, A. Vijayan, J. Liu, H. Askari,and S. B. Miller. 1998. Recombinant human insulin-like growthfactor-I (IGF-I) therapy decreases plasma leptin concentrationin patients with chronic renal insufficiency. Int. J. Obes. Relat.Metab. Disord. 22:1110–1115.
78 Davis, J. D., D. Wirtshafter, K. E. Asin, and D. Brief. 1981.Sustained intracerebroventricular infusions of brain fuels re-duces body weight and food intake in rats. Science 212:81–82.
79 Deetz, L. E., and P. J. Wangsness. 1980. Effect of intrajugularadministration of insulin on feed intake, plasma glucose andplasma insulin of sheep. J. Nutr. 110:1976–1982.
80 Deetz, L. E., and P. J. Wangsness. 1981. Influence of intrajugu-lar administration of insulin, glucagon and propionate on vol-untary feed intake of sheep. J. Anim. Sci. 53:427–433.
81 Della-Fera, M. A., and C. A. Baile. 1979. Cholecystokinin octa-peptide: continuous picomole injections into the cerebral ventri-cles of sheep suppress feeding. Science 206:471–473.
82 Della-Fera, M. A., and C. A. Baile. 1980. CCK-Octapeptideinjected in CSF and changes in feed intake and rumen motility.Physiol. Behav. 24:943–950.
83 Della-Fera, M. A., and C. A. Baile. 1980. CCK-Octapeptideinjected in CSF decreases meal size and daily food intake insheep. Peptides 1:51–54.
84 Della-Fera, M. A., and C. A. Baile. 1981. Peptides with CCK-likeactivity administration intracranially elicit satiety in sheep.Physiol. Behav. 26:979–983.

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1591
85 Della-Fera, M. A., and C. A. Baile. 1984. Control of feed intakein sheep. J. Anim. Sci. 59:1362–1368.
86 Drackley, J. K., T. H. Klusmeyer, A. M. Trusk, and J. H. Clark.1992. Infusion of long-chain fatty acids varying in saturationand chain length into the abomasum of lactating dairy cows.J. Dairy Sci. 75:1517–1526.
87 Drewry, M. M., R.B.S. Harris, and R. J. Martin. 1989. Theeffect of increased adiposity on food intake of juvinile rats.Physiol. Behav. 45:381–386.
88 Dube, M. G., B. Xu, W. R. Crowley, P. S. Kalra, and S. P. Kalra.1994. Evidence that neuropeptide Y is a physiological signalfor normal food intake. Brain Res. 646:341–344.
89 Dubuc, G. R., S. D. Phinney, J. S. Stern, and P. J. Havel.1998. Changes of serum leptin and endocrine and metabolicparameters after 7 days of energy restriction in men andwomen. Metab. Clin. Exp. 47:429–434.
90 Dyer, C. J., J. M. Simmons, R. L. Matteri, and D. H. Keisler.1997. Leptin receptor mRNA is expressed in ewe anterior pitu-itary and adipose tissues and is differentially expressed in hy-pothalamic regions of well-fed and feed-restricted ewes.Domest. Anim. Endocrinol. 14:119–128.
91 Elimam, A., U. Knutsson, M. Bronnegard, P. Stierna, W. K.Albertsson, and C. Marcus. 1998. Variations in glucocorticoidlevels within the physiological range affect plasma leptin levels.Eur. J. Endocrinol. 139:615–620.
92 Elsasser, T. H., T. J. Caperna, and T. S. Rumsey. 1995. Endo-toxin administration decreases plasma insulin-like growth fac-tor (IGF)-1 and IGF-binding protein-2 in Angus × Herefordsteers independent of changes in nutritional intake. J. Endocri-nol. 144:109–117.
93 Elsasser, T. H., S. Kahl, N. C. Steele, and T. S. Rumsey. 1997.Nutritional modulation of somatotropic axis-cytokine relation-ships in cattle: a brief review. Comp. Biochem. Physiol.116A:209–221.
94 Elsasser, T. H., N. C. Steele, and R. Fayer. 1995. Cytokines,stress, and growth modulation. Pages 261–290 in Cytokines inAnimal Health and Disease. M. J. Myers and M. P. Murtaugh,ed. Marcel Dekker, Inc., NY.
95 Etherton, T. D., and D. E. Bauman. 1998. Biology of somato-tropin in growth and lactation of domestic animals. Physiol.Rev. 78:745–761.
96 Faggioni, R., G. Fantuzzi, C. Gabay, A. Moser, C. A. Dinarello,K. R. Feingold, and C. Grunfeld. 1999. Leptin deficiency en-hances sensitivity to endotoxin-induced lethality. Am. J. Phys-iol. 276:R136–R142.
97 Faggioni, R., G. Fantuzzi, P. Villa, W. Buurman, T.L.J. van,and P. Ghezzi. 1995. Independent down-regulation of centraland peripheral tumor necrosis factor production as a result oflipopolysaccharide tolerance in mice. Infect. Immun.63:1473–1477.
98 Faggioni, R., J. Fuller, A. Moster, K. R. Feingold, and C. Grun-feld. 1997. LPS-Induced anorexia in leptin-deficient (ob/ob)and leptin receptor-deficient (db/db) mice. Am. J. Physiol.273:R181–R186.
99 Faulkner, A., and P. A. Martin. 1997. The concentrations ofsome gut polypeptides are elevated during lactation in rumi-nants. Comp. Biochem. Physiol. B. Biochem. Mol. Biol.118:563–568.
100 Faverdin, P., R. Baumont, and K. L. Ingvartsen. 1995. Controland prediction of feed intake in ruminants. Pages 95–120 inRecent Developments in the Nutrition of Herbivores. M. Jour-net, E. Grenet, M. H. Farce, M. Theriez, and C. Demarquilly,ed. INRA Editions, Paris.
101 Finck, B. N., and R. W. Johnson. 1997. Anorexia, weight lossand increased plasma interleukin-6 caused by chronic intrace-rebroventricular infusion of interleukin-1beta in the rat. BrainRes. 761:333–337.
102 Finck, B. N., K. W. Kelley, R. Dantzer, and R. W. Johnson.1998. In vivo and in vitro evidence for the involvement of tumornecrosis factor-alpha in the induction of leptin by lipopolysac-charide. Endocrinology 139:2278–2283.
Journal of Dairy Science Vol. 83, No. 7, 2000
103 Finerty, J. 1952. Parabiosis in physiological studies. Physiol.Rev. 32:277–302.
104 Fisher, D. S., J. C. Burns, and K. R. Pond. 1987. Modeling adlibitum DM intake by ruminants as regulated by distensionand chemostatic feedbacks. J. Theor. Biol. 126:407–418.
105 Fisler, J. S., M. Egawa, and G. A. Bray. 1995. Peripheral 3-hydroxybutyrate and food intake in a model of dietary-fat in-duced obesity: effect of vagotomy. Physiol. Behav. 58:1–7.
106 Flier, J. S., and F. E. Maratos. 1998. Obesity and the hypothala-mus: novel peptides for new pathways. Cell 92:437–440.
107 Flynn, M. C., T. R. Scott, T. C. Pritchard, and C. R. Plata-Salaman. 1998. Mode of action of OB protein (leptin) on feeding.Am. J. Physiol. 275:R174–R179.
108 Foot, J. Z. 1972. A note on the effect of body condition on thevoluntary food intake of dried grass wafers by Scottish Black-face ewes. Anim. Prod. 14:131–134.
109 Forbes, J. M. 1968. The physical relationship of the abdominalorgans in the pregnant ewe. J. Agric. Sci. 70:171–177.
110 Forbes, J. M. 1969. The effect of pregnancy and fatness on thevolume of rumen content in the ewe. J. Agric. Sci. 72:119–121.
111 Forbes, J. M. 1974. Feeding in sheep modified by intraventricu-lar estradiol and progesterone. Physiol. Behav. 12:741–747.
112 Forbes, J. M. 1980. A model of the short-term control of feedingin the ruminant: effects of changing animal or feed characteris-tics. Appetite 1:21–41.
113 Forbes, J. M. 1986. The effects of sex hormones, pregnancy, andlactation on digestion, metabolism, and voluntary food intake.Pages 420–435 in Control of Digestion and Metabolism in Rumi-nants. L. P. Milligan, W. L. Grovum, and A. Dobson, ed. Pren-tice-Hall, Englewood Cliffs, NJ.
114 Forbes, J. M. 1995. Voluntary food intake and diet selection infarm animals. CAB International, Wallingford. 532 pp.
115 Forbes, J. M. 1995. Voluntary intake: a limiting factor to pro-duction in high yielding dairy cows? Br. Soc. Anim. Sci. Occ.Publ. 19:13–19.
116 Foster, L. A., N. K. Ames, and R. S. Emery. 1991. Food intakeand serum insulin responses to intraventricular infusions ofinsulin and IGF-I. Physiol. Behav. 50:745–749.
117 Frederich, R. C., A. Hamann, S. Anderson, B. Lollmann, B. B.Lowell, and J. S. Flier. 1995. Leptin levels reflect body lipidcontent in mice: evidence for diet-induced resistance to leptinaction. Natl. Med. 1:1311–1314.
118 Friedman, M. I., R.B.S. Harris, H. Ji, I. Ramirez, and M. G.Tordoff. 1999. Fatty acid oxidation affects food intake by alter-ing hepatic energy status. Am. J. Physiol. 276:R1046–R1053.
119 Friedman, M. I., M. G. Tordoff, and I. Ramirez. 1986. Integratedmetabolic control of food intake. Brain Res. Bull. 17:855–859.
120 Friggens, N. C., G. C. Emmans, I. Kyriazakis, J. D. Oldham,and M. Lewis. 1998. Feed intake relative to stage of lactationfor dairy cows consuming total mixed diets with a high or lowratio of concentrate to forage. J. Dairy Sci. 81:2228–2239.
121 Furuse, A., S. I. Yang, Y. H. Choi, N. Kawamura, A. Takahashi,and J. Okumura. 1991. A note on plasma cholecystokinin con-centration in dairy cows. Anim. Prod. 53:123–125.
122 Gagliostro, G., and Y. Chilliard. 1991. Duodenal rapeseed oilinfusion in early and midlactation cows. 2. Voluntary intake,milk production, and composition. J. Dairy Sci. 74:499–509.
123 Gagliostro, G., Y. Chilliard, and M. J. Davicco. 1991. Duodenalrapeseed oil infusion in early and midlactation cows. 3. Plasmahormones and mammary apparent uptake of metabolites. J.Dairy Sci. 74:1893–1903.
124 Garnsworthy, P. C. 1988. The effect of energy reserves at calv-ing on performance of dairy cows. Pages 157–170 in Nutritionand Lactation in the Dairy Cow. P. C. Garnsworthy, ed. Butter-worths, London, UK.
125 Geary, N., J. Le-Sauter, and U. Noh. 1993. Glucagon acts in theliver to control spontaneous meal size in rats. Am. J. Physiol.264:R116–R122.
126 Geary, N., and G. P. Smith. 1983. Selective hepatic vagotomyblocks pancreatic glucagon’s satiety effect. Physiol. Behav.31:391–394.

INGVARTSEN AND ANDERSEN1592
127 Geary, N. W., and G. P. Smidt. 1982. Pancreatic glucagon andpostprandial satiety in the rat. Physiol. Behav. 28:313–322.
128 Ghilardi, N., S. Ziegler, A. Wiestner, R. Stoffel, M. H. Heim,and R. C. Skoda. 1996. Defective STAT signalling by the leptinreceptor in diabetic mice. Proc. Natl. Acad. Sci. 93:6231–6235.
129 Glowa, J. R., and P. W. Gold. 1991. Corticotropin releasinghormone produces profound anorexigenic effects in rhesus mon-key. Neuropeptides 18:55–61.
130 Griinari, J. M., M. A. McGuire, D. A. Dwyer, D. E. Bauman,and D. L. Palmquist. 1997. Role of insulin in the regulation ofmilk fat synthesis in dairy cows. J. Dairy Sci. 80:1076–1084.
131 Grossman, S. P. 1986. The role of glucose, insulin and glucagonin the regulation of food intake and body weight. Neurosci.Biobehav. Rev. 10:295–315.
132 Grovum, W. L. 1995. Mechanisms explaining the effect of shortchain fatty acids on feed intake in ruminants—osmotic pres-sure, insulin and glucagon. Pages 173–198 in Ruminant Physi-ology: Digestion, Metabolism, Growth and Reproduction. W. v.Engelhardt, S. Leonhard-Marek, G. Breves, and D. Giesecke,ed. Ferdinand Enke Verlag, Stuttgart.
133 Grummer, R. R. 1993. Etiology of lipid-related metabolic disor-ders in periparturient dairy cows. J. Dairy Sci. 76:3882–3896.
134 Grummer, R. R., S. J. Bertics, D. W. LaCount, J. A. Snow, M.R. Dentine, and R. H. Stauffacher. 1990. Estrogen inductionof fatty liver in dairy cattle. J. Dairy Sci. 73:1537–1543.
135 Grunfeld, C., C. Zhao, J. Fuller, A. Pollack, A. Moser, J. Fried-man, and K. R. Feingold. 1996. Endotoxin and cytokines induceexpression of leptin, the ob gene product, in hamsters. J. Clin.Invest. 97:2152–2157.
136 Gunn, I., D. O’Shea, and S. R. Bloom. 1997. Control of appe-tite—the role of glucagon-like peptide-1 (7-36) amide. J. Mam-mary Gland Biol. Neopl. 155:197–200.
137 Gunter, S. A., M. B. Judkins, L. J. Krysl, J. T. Broesder, R. K.Barton, B. R. Rueder, D. M. Hallford, and D. W. Holcombe.1990. Digesta kinetics, ruminal fermentation characteristicsand serum metabolites of pregnant and lactating ewes fedchopped alfalfa hay. J. Anim. Sci. 68:3821–3831.
138 Halaas, J. L., K. S. Gajiwala, M. Maffei, S. L. Cohen, B. T.Chait, D. Rabinowitz, R. L. Lallone, S. K. Burley, and J. M.Friedman. 1995. Weight-reducing effects of the plasma proteinencoded by the obese gene. Science 269:543–546.
139 Harris, R. B. 1999. Parabiosis between db/db and ob/ob or db/+ mice. Endocrinology 140:138–145.
140 Harris, R.B.S., and R. J. Martin. 1984. Specific depletion ofbody fat in parabiotic partners of tube-fed obese rats. Am. J.Physiol. 247:R380–R386.
141 Harris, R.B.S., and R. J. Martin. 1990. Site of action of putativelipostatic factor: food intake and peripheral pentose shunt ac-tivity. Am. J. Physiol. 259:R45–R52.
142 Harris, R.B.S., J. Zhou, D. S. Weigle, and J. L. Kuijper. 1997.Recombinant leptin exchanges between parabiosed mice butdoes not reach equilibrium. Am. J. Physiol. 272:R1800–R1808.
143 Hart, B. J. 1988. Biological basis of the behavior of sick animals.Neurosci. Biobehav. Rev. 12:123–137.
144 Hatfield, J. S., J. Millard, and C.J.V. Smith. 1974. Short terminfluence of intra-ventromedial hypothalamic administrationof insulin on feeding in normal and diabetic rats. Pharmacol.Biochem. Behav. 2:223–330.
145 Havrankova, J., J. Roth, and M. Brownstein. 1978. Insulinreceptors are widely distributed in the central nervous systemof the rat. Nature 272:827–829.
146 Head, H. H., W. W. Thatcher, C. J. Wilcox, and K. C. Bachmann.1976. Effect of a synthetic corticoid on milk yield and composi-tion and on blood metabolites and hormones in dairy cows. J.Dairy Sci. 59:880–888.
147 Henry, B. A., J. W. Goding, W. S. Alexander, A. J. Tilbrook, B.J. Canny, F. R. Dunshea, A. Rao, A. Mansell, and I. J. Clarke.1999. Central administration of leptin to ovariectomized ewesinhibits food intake without affecting the secretion of hormonesfrom the pituitary gland: evidence for a dissociation of effectson appetite and neuroendocrine function. Endocrinology140:1175–1182.
Journal of Dairy Science Vol. 83, No. 7, 2000
148 Hervey, G. R. 1959. The effects of lesions in the hypothalamusin parabiotic rats. J. Physiol. 145:336–352.
149 Highman, T. J., J. E. Friedman, L. P. Huston, W. W. Wong,and P. M. Catalano. 1998. Longitudinal changes in maternalserum leptin concentrations, body composition, and restingmetabolic rate in pregnancy. Am. J. Obstet. Gynecol.178:1010–1015.
150 Hill, A. G., L. Jacobson, J. Gonzalez, J. Rounds, J. A. Majzoub,and D. W. Wilmore. 1996. Chronic central nervous system expo-sure to interleukin-beta causes catabolism in the rat. Am. J.Physiol. 271:R1142–R1148.
151 Ho, L. T., Y. F. Chern, and M. T. Lin. 1989. The hypothalamic-somatostatinergic pathways mediate feeding behavior in therat. Experientia 45:161–162.
152 Holst, J. J. 1997. Enteroglucagon. Annu. Rev. Physiol.59:257–271.
153 Horn, C. C., A. Addis, and M. I. Friedman. 1999. Neural sub-strate for integrated metabolic control of feeding behavior. Am.J. Physiol. 276:R113–R119.
154 Hotta, M., T. Shibasaki, N. Yamauchi, H. Ohno, R. Benoit,N. Ling, and H. Demura. 1991. The effects of chronic centraladministration of corticotropin-pituitary-adrenocortical hor-mones. Life Sci. 48:1491.
155 Houpt, T. R. 1974. Stimulation of food intake in ruminants by2-deoxy-D-glucose and insulin. Am. J. Physiol. 227:161–167.
156 Houseknecht, K. L., C. A. Baile, R. L. Matteri, and M. E. Spur-lock. 1998. The biology of leptin: a review. J. Anim. Sci.76:1405–1420.
157 Houseknecht, K. L., C. S. Mantzoros, R. Kuliawat, E. Hadro,J. S. Flier, and B. B. Kahn. 1996. Evidence for leptin bindingto proteins in serum of rodents and humans: modulation withobesity. Diabetes 45:1638–1643.
158 Houseknecht, K. L., and C. P. Portocarrero. 1998. Leptin andits receptors: regulators of whole-body energy homeostasis.Domest. Anim. Endocrinol. 15:457–475.
159 Hwa, J. J., L. Ghibaudi, P. Williams, M. B. Witten, R. Tedesco,and C. D. Strader. 1998. Differential effects of intracerebroven-tricular glucagon-like peptide-1 on feeding and energy expendi-ture regulation. Peptides 19:869–875.
160 Ikeda, H., D. B. West, J. J. Pustek, D. P. Figlewicz, M.R.C.Greenwood, D. Porte, and S. C. Woods. 1986. Intracerobroven-tricular insulin reduces food intake and body weight of leangut not obese sugar rats. Appetite 7:381–386.
161 Ingvartsen, K. L. 1994. Models of voluntary food intake incattle. Livest. Prod. Sci. 39:19–38.
162 Ingvartsen, K. L., H. R. Andersen, and J. Foldager. 1992. Effectof sex and pregnancy on feed intake capacity of growing cattle.Acta Agric. Scand., Sect. A, Anim. Sci. 42:40–46.
163 Ingvartsen, K. L., A. Danfær, P. H. Andersen, and J. Foldager.1995. Prepartun feeding of dairy cattle: a review of the effecton prepartum metabolism, feed intake, production and health.Pages 83 in Book of Abstracts of the 46th Annual Meeting ofthe EAAP, Pragh. J.A.M. Arendonk, ed. Wageningen Press, Wa-geningen.
164 Ingvartsen, K. L., J. Foldager, and O. Aaes. 1997. Effect ofprepartum TMR energy concentration on feed intake, milkyield, and energy balance in dairy heifers and cows. J. DairySci. 80:211(Abstr.).
165 Ingvartsen, K. L., J. Foldager, O. Aaes, and P. H. Andersen.1995. Effect of feeding level the last 24 weeks prepartum onfeed intake, production and metabolism in heifers and cows.Internal Report no. 47: 60–74. Research Centre Foulum, DanishInstitute of Animal Science. Transition to Lactation. Feedingand Physiology of the Dairy Cow During Pregnancy and AroundParturition. (In Danish)
166 Ingvartsen, K. L., N. C. Friggens, and F. Faverdin. 1999. Feedintake regulation in late pregnancy and early lactation. Br.Soc. Anim. Sci. Occ. Publ. 24:37–54.
167 Ingvartsen, K. L., and K. Sejrsen. 1995. Effect of immunizationagainst somatostatin (SS) in cattle: a review of performance,carcass composition and possible mode of action. Acta Agric.Scand., Sect. A, Anim. Sci. 45:124–131.

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1593
168 Inokuchi, A., Y. Oomura, and H. Nishimura. 1984. Effect ofintracerebroventricularly infused glucagon on feeding behav-ior. Physiol. Behav. 33:397–400.
169 Janowitz, H. D., and A. C. Ivy. 1948. Role of blood sugar levelsin spontaneous and insulin-induced hunger in man. J. Appl.Physiol. 1:643–645.
170 Jarrige, R. 1986. Voluntary intake in dairy cows and its predic-tion. Int. Dairy Fed. Bull. 196:4–16.
171 Ji, S., G. M. Willis, R. R. Scott, and M. E. Spurlock. 1998.Partial cloning and expression of the bovine leptin gene. Anim.Biotech. 9:1–14.
172 Johnson, R. W. 1998. Immune and endocrine regulation of foodintake in sick animals. Domest. Anim. Endocrinol. 15:309–319.
173 Kaiyala, K. J., S. C. Woods, and M. W. Schwartz. 1995. Newmodel for the regulation of energy balance and adiposity by thecentral nervous system. Am. J. Clin. Nutr. 62:1123S–1134S.
174 Kalra, S. P., M. G. Dube, A. Sahu, C. P. Phelps, and P. S. Kalra.1991. Neuropeptide Y secretion increases in the paraventricu-lar nucleus in association with increased appetite for food. Proc.Natl. Acad. Sci. U.S.A. 88:10931–10935.
175 Kaske, M., and A. Groth. 1997. Changes in factors affectingthe rate of digesta passage during pregnancy and lactation insheep fed on hay. Reprod. Nutr. Dev. 37:573–588.
176 Kasser, T. R., R.B.S. Harris, and R. J. Martin. 1985. Level ofsatiety: fatty acid and glucose metabolism in three brain sitesassociated with feeding. Am. J. Physiol. 248:R447–R452.
177 Kasser, T. R., R.B.S. Harris, and R. J. Martin. 1989b. Level ofsatiety: in vitro energy metabolism in brain during hypophagicand hyperphagic body weight recovery. Am. J. Physiol.257:R1322–R1327.
178 Kasser, T. R., R.B.S. Harris, and R. J. Martin. 1989a. Site ofaction of putative lipostatic factor: hypothalamic metabolismof parabiotic rats. Am. J. Physiol. 257:R224–R228.
179 Kawai, M., M. Yamaguchi, T. Murakami, K. Shima, Y. Murata,and K. Kishi. 1997. The placenta is not the main source ofleptin production in pregnant rats: gestational profile of leptinin plasma and adipose tissues. Biochem. Biophys. Res. Com-mun. 240:798–802.
180 Kennedy, G. C. 1953. The role of depot fat in the hypothalamiccontrol of food intake in the rat. Proc. Royal Soc., Ser. B139:578–592.
181 Kolaczynski, J. W., R. V. Considine, J. Ohannesian, C. Marco,I. Opentanova, M. R. Nyce, M. Myint, and J. F. Caro. 1996.Responses of leptin to short-term fasting and refeeding in hu-mans. A link with ketogenesis but not ketones themselves.Diabetes 45:1511–1515.
182 Kolaczynski, J. W., B. J. Goldstein, and R. V. Considine. 1997.Dexamethasone, OB gene, and leptin in humans; effect of exoge-nous hyperinsulinemia [see comments]. J. Clin. Endocrinol.Metab. 82:3895–3897.
183 Kolaczynski, J. W., M. R. Nyce, R. V. Considine, G. Boden, J.J. Nolan, R. Henry, S. R. Mudaliar, J. Olefsky, and J. F. Caro.1996. Acute and chronic effect of insulin on leptin productionin humans. Diabetes 45:701.
184 Koopmans, H. S., E. K. Walls, and A. E. Willing. 1991. Doesthe integrated level of all plasma nutrients control daily foodintake? Brain Res. Bull. 27:429–434.
185 Koopmans, S. J., M. Frolich, E. H. Gribnau, R. G. Westendorp,and R. A. DeFronzo. 1998. Effect of hyperinsulinemia on plasmaleptin concentrations and food intake in rats. Am. J. Physiol.274:E998–E1001.
186 Krahn, D. D., B. A. Gosnell, M. Grace, and A. S. Levine. 1986.CRF antagonist partially reverses CRF- and stress-inducedeffects on feeding. Brain Res. Bull. 17:285–289.
187 Krahn, D. D., B. A. Gosnell, A. S. Levine, and J. E. Morley.1984. Localization of the effects of corticotropin releasing factoron feeding. Proc. Soc. Neurosci. 10:302(Abstr.).
188 Kristensen, V. F., and K. L. Ingvartsen. 1985. Variations involuntary feed intake in cattle related to animal factors. Dairycows. Pages 302–303 in 36th Annual Meeting of the EAAP,Summaries, Kallithea, Greece.
Journal of Dairy Science Vol. 83, No. 7, 2000
189 Kyrkouli, S. E., B. G. Stanley, and S. F. Leibowitz. 1986. Ga-lanin: stimulation of feeding induced by medial hypothalamicinjection of this novel peptide. Eur. J. Pharmacol. 122:159–160.
190 Kyrkouli, S. E., B. G. Stanley, R. D. Seirafi, and S. F. Leibowitz.1990. Stimulation of feeding by galanin: anatomical localizationand behavioral specificity of this peptide’s effects in the brain.Peptides 11:995–1001.
191 Lagerlof, N. 1929. Investigations of the topography of the ab-dominal organs in cattle, and some clinical observations andremarks in connection with the subject. Skand. Veterinærtids19:253–365.
192 Langhans, W. 1996. Bacterial products and the control of in-gestive behavior: clinical implications. Nutrition 12:303–315.
193 Langhans, W. 1996. Metabolic and glucostatic control of feed-ing. Proc. Nutr. Soc. 55:497–515.
194 Langhans, W., G. Egli, and E. Scharrer. 1985b. Regulation offood intake by hepatic oxidative metabolism. Brain Res. Bull.15:425–428.
195 Langhans, W., G. Egli, and E. Scharrer. 1985. Selective hepaticvagotomy eliminates the hypophagic effect of different metabo-lites. J. Auton. Nervous Sys. 13:255–262.
196 Langhans, W., K. Pantel, and E. Scharrer. 1985. Ketone kinet-ics and D-(-)-3-hydroxybutyrate induced inhibition of feedingin rats. Physiol. Behav. 34:579–582.
197 Langhans, W., D. Savoldelli, and S. Weingarten. 1993. Compar-ison of the feeding responses to bacterial lipopolysaccharideand interleukin-1b. Physiol. Behav. 53:643–649.
198 Langhans, W., and E. Scharrer. 1987. Evidence for a vagallymediated satiety signal derived from hepatic fatty acid oxida-tion. J. Auton. Nervous Sys. 18:13–18.
199 Langhans, W., U. Ziegler, E. Scharrer, and N. W. Geary. 1982.Stimulation of feeding rats by intraperitoneal injections of anti-bodies to glucagon. Science 218:894–896.
200 Larsson, H., S. Elmstahl, G. Berglund, and B. Ahren. 1998.Evidence for leptin regulation of food intake in humans. J. Clin.Endocrinol. Metab. 83:4382–4385.
201 Lee, G. H., R. Proenca, J. A. Montez, K. M. Caroll, J. G. Darvish-zadeh, J. I. Lee, and J. M. Friedman. 1996. Abnormal splicingof the leptin receptor in diabetic mice. Nature 379:632–635.
202 Levine, A. S., B. Rogers, J. Kneip, M. Grace, and J. E. Morley.1983. Effect of centrally administered corticotropin releasingfactor (CRF) on multiple feeding paradigms. Neuropharmacol-ogy 22:337–339.
203 Levine, A. S., and J. E. Morley. 1982. Peripherally administeredsomatostatin reduces feeding by a vagal mediated mechanism.Pharmacol. Biochem. Behav. 16:897–902.
204 Lewandowski, K., R. Horn, C. J. O’Callaghan, D. Dunlop,G. F. Medley, P. O’Hare, and G. Brabant. 1999. Free leptin,bound leptin, and soluble leptin receptor in normal and diabeticpregnancies. J. Clin. Endocrinol. Metab. 84:300–306.
205 Lewis, D. E., L. Shellard, D. G. Koeslag, D. E. Boer, H. D.McCarthy, P. E. McKibbin, J. C. Russell, and G. Williams. 1993.Intense exercise and food restriction cause similar hypothala-mic neuropeptide Y increases in rats. Am. J. Physiol.264:E279–E284.
206 Liddle, R. A. 1995. Regulation of cholecystokinin secretion byintraluminal releasing factors. Am. J. Physiol. 269:G319–G327.
207 Liddle, R. A. 1997. Cholecystokinin cells. Annu. Rev. Physiol.59:221–242.
208 Lin, M. T., Y. F. Chen, and L. T. Ho. 1987. Hypothalamicinvolvement in the hyperglycemia and satiety actions of so-matostatin in rats. Neuroendocrinology 45:62–67.
209 Lotter, E., R. Krinsky, J. M. McKay, C. Treneer, D. Porte, andS. Woods. 1981. Somatostatin decreases food intake of rats andbaboons. J. Comp. Physiol. Psych. 95:278–287.
210 Maffei, M., H. Fei, G. H. Lee, C. Dani, P. Leroy, Y. Zhang, R.Proenca, R. Negrel, G. Ailhaud, and J. M. Friedman. 1995.Increased expression in adipocytes of ob RNA in mice lesionsof the hypothalamus and with mutations at the db locus. Proc.Natl. Acad. Sci. 92:6957–6960.
211 Maffei, M., J. Halaas, E. Ravussin, R. E. Pratley, G. H. Lee,Y. Zhang, H. Fei, S. Kim, R. Lallone, S. Ranganathan, P. A.

INGVARTSEN AND ANDERSEN1594
Kern, and J. M. Friedman. 1995. Leptin levels in human androdent: measurement of plasma leptin and ob RNA in obeseand weight-reduced subjects. Nature Med. 1:1155–1161.
212 Makela, A. 1956. Studies on the question of bulk in the nutritionof farm animals with animals with special reference to cattle.Acta Agralia Fennica 85:130.
213 Marcle, T. R., D. A. Dwyer, K. L. Ingvartsen, P. Y. Chouinard,and D. E. Bauman. 1999. Effect of insulin and amino acids onmilk protein concentration and yield from dairy cows. J. DairySci. 82:1512–1524.
214 Martin, J. R., D. Novin, and D. A. Vanderweele. 1978. Loss ofglucagon suppression of feeding following vagotomy in rats.Am. J. Physiol. 234:E314–E318.
215 Masuzaki, H., Y. Ogawa, N. Isse, N. Satoh, T. Okazaki, M.Shigemoto, K. Mori, N. Tamura, K. Hosoda, Y. Yoshimasa, H.Jingami, T. Kawada, and K. Nakao. 1995. Adipocyte-specificexpression and regional differences in the adipose tissue. Dia-betes 44:855–858.
216 McCann, J. P., E. N. Bergman, and D. H. Beermann. 1992.Dynamic and static phases of severe dietary obesity in sheep:food intakes, endocrinology and carcass and organ chemicalcomposition. J. Nutr. 122:496–505.
217 McGowan, M. K., K. M. Andrews, D. Fenner, and S. P. Gross-man. 1993. Chronic intrahypothalamic insulin infusion in therat: behavioral specificity. Physiol. Behav. 54:1031–1034.
218 McGowan, M. K., K. M. Andrews, and S. P. Grossman. 1992.Chronic intrahypothalamic infusions of insulin or insulin anti-bodies alter body weight and food intake in the rat. Physiol.Behav. 51:753–766.
219 McGuire, M. A., J. M. Griinari, D. A. Dwyer, and D. E. Bauman.1995. Role of insulin in the regulation of mammary synthesisof fat and protein. J. Dairy Sci. 78:816–824.
220 McHugh, K. J., S. M. Collins, and H. P. Weingarten. 1994.Central interleukin-1 receptor contributes to suppression offeeding after acute colitis in the rat. Am. J. Physiol.266:R1659–R1663.
221 McLaughlin, C. L., G. J. Rogan, J. Tou, C. A. Baile, and W. D.Joy. 1992. Food intake and body temperature responses of ratsto recombinant human interleukin-1β and a tripeptide interleu-kin-1β antagonist. Physiol. Behav. 52:1155–1160.
222 McMinn, J. E., R. J. Seeley, C. W. Wilkinson, P. J. Havel, S.C. Woods, and M. W. Schwartz. 1998. NPY-induced overfeedingsuppresses hypothalamic NPY mRNA expression: potentialroles of plasma insulin and leptin. Reg. Peptides 75–76:425–431.
223 McNamara, J. 1995. Role and regulation of metabolism in adi-pose tissue during lactation. J. Nutr. Biochem. 6:120–129.
224 Meeran, K., D. O’Shea, C. M. Edwards, M. D. Turton, M. M.Heath, I. Gunn, S. Abusnana, M. Rossi, C. J. Small, A. P.Goldstone, G. M. Taylor, D. Sunter, J. Steere, S. J. Choi, M.A. Ghatei, and S. R. Bloom. 1999. Repeated intracerebroventric-ular administration of glucagon-like peptide-1-(7-36) amide orexendin-(9-39) alters body weight in the rat. Endocrinology140:244–250.
225 Mertens, D. R. 1995. Methods in modelling feeding behaviourand intake in herbivores. Pages 1–17 in IVth Int. Symp. Nutr.Herbivores. Inst. Natl. Agron. Paris-Grignon and INRA, Paris.
226 Miner, J. L., M. A. Della-Fera, J. A. Paterson, and C. A. Baile.1989. Lateral cerebroventricular injection of neuropeptide Ystimulates feeding in sheep. Am. J. Physiol. 257:R383–R387.
227 Miner, J. L., F. M. Della, and J. A. Paterson. 1990. Lateralcerebroventricular injection of neuropeptide Y stimulates feed-ing in sheep. J. Anim. Sci. 68:3805–3811.
228 Mistry, A. M., A. G. Swick, and D. R. Romsos. 1997. Leptinrapidly lowers food intake and elevates metabolic rates in leanand ob/ob mice. J. Nutr. 127:2065–2072.
229 Mook, D. G., N. J. Kenny, S. Roberts, A. I. Nussbaum, and W.I. Rodier. 1972. Ovarian-adrenal interaction in regulation ofbody weight by female rats. J. Comp. Physiol. Psych.81:198–211.
230 Morley, J. E. 1990. Appetite regulation by gut peptides. Annu.Rev. Nutr. 10:383–395.
Journal of Dairy Science Vol. 83, No. 7, 2000
231 Morley, J. E., and A. S. Levine. 1982. Corticotropin-releasingfactor, grooming and ingestive behavior. Life Sci. 31:1459–1464.
232 Moshyedi, A. K., M. D. Josephs, E. K. Abdalla, S. L. Mackay,C. K. Edwards, E. M. Copeland, and L. L. Moldawer. 1998.Increased leptin expression in mice with bacterial peritonitisis partially regulated by tumor necrosis factor alpha. Infect.Immun. 66:1800–1802.
233 Muir, L. A., J. W. Hibbs, H. R. Conrad, and K. L. Smith. 1972.Effect of estrogen and progesterone on feed intake and hydroxy-proline excretion following induced hypocalcemia in dairy cows.J. Dairy Sci. 31:145–148.
234 Murray, M. J., and A. B. Murray. 1979. Anorexia of infectionas a mechanism of host defence. Am. J. Clin. Nutr. 32:593–596.
235 Mutt, V., and J. E. Jorpes. 1968. Structure of porcine cholecysto-kinin-pancreozymin. Cleavage with thrombin and with trypsin.Eur. J. Biochem. 6:156–162.
236 National Research Council. 1988. Nutrient Requirements ofDairy Cattle, 6th revised edn. National Acadamy Press, Wash-ington, D.C.
237 Nishizawa, Y., and G. A. Bray. 1980. Evidence for a circulatingergostatic factor: studies on parabiotic rats. Am. J. Physiol.239:R344–R351.
238 Oldick, B. S., C. R. Staples, W. W. Thatcher, and P. Gyawu.1997. Abomasal infusion of glucose and fat—effect on digestion,production, and ovarian and uterine functions of cows. J. DairySci. 80:1315–1328.
239 Opara, E. I., M. M. Meguid, Z.-J. Yang, and W. G. Hammond.1996. Studies on the regulation of food intake using rat totalparenteral nutrition as a model. Neurosci. Biobehav. Rev.20:413–443.
240 Orskov, C., S. S. Poulsen, M. Moller, and J. J. Holst. 1996.Glucagon-like peptide I receptors in the subfornical organ andthe area postrema are accessible to circulating glucagon-likepeptide I. Diabetes 45:832–835.
241 Palmiter, R. D., J. C. Erickson, G. Hollopeter, S. C. Baraban,and M. W. Schwartz. 1998. Life without neuropeptide Y. Re-cent. Progress Horm. Res. 53:163–199.
242 Palmquist, D. L. 1994. The role of dietary fats in efficiency ofruminants. J. Nutr. 124:1377S–1382S.
243 Parameswaran, S. V., A. B. Steffens, G. R. Hervey, and L. DeRuiter. 1977. Involvement of a humoral factor in the regulationof body weight in parabiotic rats. Am. J. Physiol. 232:R150–R157.
244 Pelleymounter, M. A., M. J. Cullen, M. B. Baker, R. Hecht, D.Winters, T. Boone, and F. Collins. 1995. Effects of the obesegene product on body weight regulation in ob/ob mice. Science269:540–543.
245 Pelleymounter, M. A., M. J. Cullen, D. Healy, R. Hecht, D.Winters, and M. McCaleb. 1998. Efficacy of exogenous recombi-nant murine leptin in lean and obese 10- to 12-mo-old femaleCD-1 mice. Am. J. Physiol. 275:R950–R959.
246 Peterson, A. D., and B. R. Baumgardt. 1976. Influence of levelof energy demand on the ability of rats to compensate for dietdilution. J. Nutr. 101:1069–1074.
247 Petrusz, P., and I. Merchenthaler. 1992. The corticotropin-re-leasing factor system. Pages 129-183 in Neuroendocrinology.C. B. Nemeroff, ed. CRC Press, Boca Raton.
248 Pickavance, L., M. Tadayyon, G. Williams, and R. G. Vernon.1998. Lactation suppresses diurnal rhythm of serum leptin.Biochem. Biophys. Res. Commun. 248:196–199.
249 Plata-Salaman, C. R. 1994. Meal patterns in response to theintracerebroventricular administration of interleukin-1 beta inrats. Physiol. Behav. 55:727–733.
250 Plata-Salaman, C. R. 1998. Cytokine-induced anorexia. Behav-ioral, cellular, and molecular mechanisms. Ann. N.Y. Acad. Sci.856:160–170.
251 Plata-Salaman, C. R., and J. P. Borkoski. 1993. Centrally ad-ministered bacterial lipopolysaccharide depresses feeding inrats. Pharmacol. Biochem. Behav. 46:787–791.
252 Plata-Salaman, C. R., and J.M.H. French-Mullen. 1992. Intra-cerebroventricular administration of a specific IL-1 receptor

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1595
antagonist blocks food and water intake suppression inducedby interleukin-1 beta. Physiol. Behav. 51:1277–1279.
253 Plata-Salaman, C. R., and Y. Oomura. 1986. Effect of intra-third ventricular administration of insulin on food intake afterfood deprivation. Physiol. Behav. 37:735–739.
254 Plata-Salaman, C. R., Y. Oomura, and Y. Kai. 1988. Tumornecrosis factor and interleukin-1 beta: suppression of food in-take by direct action in the central nervous system. Brain Res.448:106–114.
255 Plata-Salaman, C. R., Y. Oomura, and H. Shimizu. 1986. De-pendence of food intake on acute and chronic ventricular admin-istration of insulin. Physiol. Behav. 37:735–739.
256 Plata-Salaman, C. R., G. Sonti, J. P. Borkoski, C. D. Wilson,and J. M. French-Mullen. 1996. Anorexia induced by chroniccentral administration of cytokines at estimated pathophysio-logical concentrations. Physiol. Behav. 60:867–875.
257 Polonsky, K. S., B. D. Given, and E. Van Cauter. 1988. Twenty-four-hour profiles and pulsatile patterns of insulin secretion innormal and obese subjects. J. Clin. Invest. 81:442–448.
258 Porte, D., Jr., and S. C. Woods. 1981. Regulation of food intakeand body weight by insulin. Diabetologia 20:274–280.
259 Ramirez, I., and M. I. Friedman. 1982. Glycerol is not a physi-ologi signal in the control of food intake in rats. Physiol. Behav.29:921–925.
260 Ramsay, T. G., X. Yan, and C. Morrison. 1998. The obesity genein swine: sequence and expression of porcine leptin. J. Anim.Sci. 76:484–490.
261 Richard, D. 1998. Involvement of corticotropin-releasing factorin the control of food intake and energy expenditure. Ann. N.Y.Acad. Sci. 697:155–172.
262 Riedy, C. A., M. Chavez, and S. C. Woods. 1995. Central insulinenhances sensitivity to cholecystokinin. Physiol. Behav.58:755–760.
263 Roh, S., I. J. Clarke, R. Xu, J. W. Goding, K. Loneragan, andC. Chen. 1998. The in vitro effect of leptin on basal and growthhormone-releasing hormone-stimulated growth hormone secre-tion from the ovine pituitary gland. Neuroendocrinology68:361–364.
264 Roseler, D. K., D. G. Fox, L. E. Chase, A. N. Pell, andW. C. Stone. 1997. Development and evaluation of equationsfor prediction of feed intake for lactating Holstein dairy cows.J. Dairy Sci. 80:878–893.
265 Roseler, D. K., D. G. Fox, A. N. Pell, and L. E. Chase. 1997.Evaluation of alternative equations for prediction of intake forHolstein dairy cows. J. Dairy Sci. 80:864–877.
266 Ross, G. E., and I. Zucker. 1974. Progesterone and the ovarian-adrenal modulation of energy balance in rats. Horm. Behav.5:43–62.
267 Rowland, N. E., A. Morien, and B. H. Li. 1996. The physiologyand brain mechanisms of feeding. Nutrition 12:626–639.
268 Ruckebusch, Y., and C. H. Malbert. 1986. Stimulation and inhi-bition of food intake in sheep by centrally-administered hypo-thalamic releasing factors. Life Sci. 38:929–934.
269 Sahu, A. 1998. Evidence suggesting that galanin (GAL), mela-nin-concentrating hormone (MHC), neurotensin (NT), proopio-melanocortin (POMC) and neuropeptide Y (NPY) are targetsof leptin signaling in the hypothalamus. Endocrinology139:795–798.
270 Sahu, A., P. S. Kalra, and S. P. Kalra. 1988. Food deprivationand ingestion induce reciprocal changes in neuropeptide Y con-centrations in the paraventricular nucleus. Peptides 9:83–86.
271 Sakaguchi, T., M. Takahshi, and G. A. Bray. 1999. Diurnalchanges in sympathetic activity. Relation to food intake andto insulin injected into the ventromedial or suprachiasmaticnucleus. J. Clin. Invest. 82:282–286.
272 Sakurai, T., A. Amemiya, M. Ishii, I. Matsuzaki, R. M. Chemelli,H. Tanaka, S. C. Williams, J. A. Richardson, G. P. Kozlowski,S. Wilson, J. R. Arch, R. E. Buckingham, A. C. Haynes, S. A.Carr, R. S. Annan, D. E. McNulty, W. S. Liu, J. A. Terrett, N.A. Elshourbagy, D. J. Bergsma, and M. Yanagisawa. 1998.Orexins and orexin receptors: a family of hypothalamic neuro-
Journal of Dairy Science Vol. 83, No. 7, 2000
peptides and G protein-coupled receptors that regulate feedingbehavior. Cell 92:573–585.
273 Saladin, R., V. P. De, M. M. Guerre, A. Leturque, J. Girard, B.Staels, and J. Auwerx. 1995. Transient increase in obese geneexpression after food intake or insulin administration. Nature377:527–529.
274 Sarraf, P., R. C. Frederich, E. M. Turner, G. Ma, N. T. Jaskow-iak, D. J. Rivet III, J. S. Flier, B. B. Lowell, D. L. Fraker, andH. R. Alexander. 1997. Multiple cytokines and acute inflamma-tion raise mouse leptin levels: potential role in inflammatoryanorexia. J. Exp. Med. 185:171–175.
275 Satoh, N., Y. Ogawa, G. Katsuura, T. Tsuji, H. Masuzaki, J.Hiraoka, T. Okazaki, M. Tamaki, M. Hayase, Y. Yoshimasa,S. Nishi, K. Hosoda, and K. Nakao. 1997. Pathophysiologicalsignificance of the obese gene product, leptin, in ventromedialhypothalamus (VMH)-lesioned rats: evidence for loss of its sati-ety effect in VMH-lesioned rats. Endocrinology 138:947–954.
276 Scharrer, E., and W. Langhans. 1986. Control of food intakeby fatty acid oxidation. Am. J. Physiol. 250:R1003–R1006.
277 Scharrer, E., and W. Langhans. 1988. Metabolic and hormonalfactors controlling food intake. Int. J. Vit. Nutr. Res. 58:249–261.
278 Scharrer, E., and W. Langhans. 1990. Mechanisms for the effectof body fat on food intake. Pages 63–86 in The Control of BodyFat Content. J. M. Forbes and G. R. Hervey, ed. Smith-Gor-don, London.
279 Schick, R. R., V. Schusdziarra, T. L. Yaksh, and V. L. Go. 1994.Brain regions where cholecystokinin exerts its effect on satiety.Ann. N.Y. Acad. Sci. 713:242–254.
280 Schobitz, B., G. Pezeshki, T. Pohl, U. Hemmann, P. C. Heinrich,F. Holsboer, and J. M. Reul. 1995. Soluble interleukin-6 (IL-6) receptor augments central effects of IL-6 in vivo. FASEB J.9:659–664.
281 Schubring, C., P. Englaro, T. Siebler, W. F. Blum, T. Demirakca,J. Kratzsch, and W. Kiess. 1998. Longitudinal analysis of ma-ternal serum leptin levels during pregnancy, at birth and upto six weeks after birth: relation to body mass index, skinfolds,sex steroids and umbilical cord blood leptin levels. Horm. Res.50:276–283.
282 Schulman, J. L., J. L. Carleton, G. Whitney, and J. C.Whitehorn. 1957. Effect of glucagon on food intake and bodyweight in man. J. Appl. Physiol. 11:419–421.
283 Schwartz, M. W., D. G. Baskin, T. R. Bukowski, J. L. Kuijper,D. Foster, G. Lasser, D. E. Prunkard, D. Porte, S. C. Woods,R. J. Seeley, and D. S. Weigle. 1996. Specificity of leptin actionon elevated blood glucose levels and hypothalamic neuropepticeY gene expression in ob/ob mice. Diabetes 45:531–535.
284 Schwartz, M. W., J. L. Marks, A. J. Sipols, D. G. Baskin,S. C. Woods, S. E. Kahn, and D. Porte. 1991. Central insulinadministration reduces neuropeptide Y mRNA expression inthe arcuate nucleus of food-deprived lean (Fa/Fa) but not obese(fa/fa) Zucker rats. Endocrinology 128:2645–2647.
285 Schwartz, M. W., R. J. Seeley, L. A. Campfield, P. Burn, andD. G. Baskin. 1996. Identification of targets of leptin action inrat hypothalamus. J. Clin. Invest. 98:1101–1106.
286 Schwartz, M. W., A. J. Sipol, S. E. Kahn, D. E. Latlemann,G. J. Taborsky, Jr., R. N. Bergman, S. C. Woods, and D. Porte,Jr. 1990. Kinetics and specificity of insulin uptake from plasmainto cerebrospinal fluid. Am. J. Physiol. 259:E378–E383.
287 Schwartz, M. W., A. J. Sipols, J. L. Marks, G. Sanacora, J. D.White, A. Scheurink, S. E. Kahn, D. G. Baskin, S. C. Woods,D. P. Figlewicz, and D. Porte. 1992. Inhibition of hypothalamicneuropeptide Y gene expression by insulin. Endocrinology130:3608–3616.
288 Seeley, R. J., G. van Dijk, L. A. Campfield, F. J. Smith, P. Burn,J. A. Nelligan, S. M. Bell, D. G. Baskin, S. C. Woods, andM. W. Schwartz. 1996. Intraventricular leptin reduces foodintake and body weight of lean rats but not obese zucker rats.Horm. Metab. Res. 28:664–668.
289 Shi, Z. Q., A. Nelson, L. Whitcomb, J. Wang, and A. M. Cohen.1998. Intracerebroventricular administration of leptin mark-

INGVARTSEN AND ANDERSEN1596
edly enhances insulin sensitivity and systemic glucose utiliza-tion in conscious rats. Metabolism 47:1274–1280.
290 Silver, A. J., and J. E. Morley. 1991. Role of CCK in regulationof food intake. Prog. Neurobiol. 36:23–34.
291 Singer-Koegler, L. K., P. Magluyan, and S. Ritter. 1996. Theeffects of low-, medium-, and high-fat diets on 2-deoxy-D-glu-cose- and mercaptoacetate-induced feeding. Physiol. Behav.60:321–323.
292 Singer, L. K., and S. Ritter. 1993. Differential effects of infusednutrients on 2-deoxy-D-glucose- and 2-mercaptoacetate-in-duced feeding. Physiol. Behav. 56:193–196.
293 Singer, L. K., D. A. York, and G. Bray. 1997. Feeding responseto mercaptoacetate in Osborne-Mendel and S5B/PL rats. Obes.Res. 5:587–594.
294 Sinha, M. K., I. Opentanova, J. P. Ohannesian, J. W. Kolaczyn-ski, M. Heiman, J. Hale, G. W. Becker, R. R. Bowsher, T. W.Stephens, and J. F. Caro. 1996. Evidence of free and boundleptin in human circulation. J. Clin. Invest. 98:1277–1282.
295 Sinha, M. K., and J. F. Caro. 1998. Clinical aspects of leptin.Vit. Horm. 54:1–30.
296 Sinha, M. K., J. P. Ohannesian, M. L. Heiman, A. Kriauciunas,T. W. Stephens, S. Magosin, C. Marco, and J. F. Caro. 1996.Nocturnal rise of leptin in lean, obese, and non-insulin-depen-dent diabetes mellitus subjects. J. Clin. Invest. 97:1344–1347.
297 Sinha, M. K., J. Sturis, J. P. Ohannesian, S. Magosin, T. W.Stephens, M. L. Heiman, K. S. Polonsky, and J. F. Caro. 1996.Ultradian oscillations of leptin secretion in humans. Biochem.Biophys. Res. Commun. 228:733–738.
298 Slonaker, J. R. 1924. The effect of copulation, pregnancy, pseu-dopregnancy and lactation on the voluntary activity and foodconsumption of the albino rat. Am. J. Physiol. 71:362–394.
299 Smith, B. K., and M. J. Kluger. 1993. Anti TNF-antibodiesnormalized body temperature and enhanced food intake in tu-mor-bearing rats. Am. J. Physiol. 248:R615–R619.
300 Smith, G. P., and J. Gibbs. 1994. Satiating effect of cholecystoki-nin. Ann. N.Y. Acad. Sci. 713:236–241.
301 Smith, M. S. 1993. Lactation alters neuropeptide-Y and proopi-omelanocortin gene expression in the arcuate nucleus of therat. Endocrinology 133:1258–1283.
302 Sonti, G., S. E. Ilyin, and C. R. Plata-Salaman. 1996. Anorexiainduced by cytokine interactions at pathophysiological concen-trations. Am. J. Physiol. 270:R1394–R1402.
303 Spencer, G.S.G. 1986. Immuno-neutralization of somatostatinand its effect on animal production. Domest. Anim. Endocrinol.3:55–68.
304 Spencer, G.S.G., and A. M. Fadlalla. 1989. Effect of intracere-broventricular administration of anti-somatostatin serum onthe regulation of appetite in sheep. Pages 51–54 in Endocrinol-ogy of Farm Animals. K. Boda, ed. Kosice, Prague, Czech Re-public.
305 Stanley, B. G., S. E. Kyrkouli, S. Lampert, and S. F. Leibowitz.1986. Neuropeptide Y chronically injected into the hypothala-mus: a powerful neurochemical inducer of hyperphagia andobesity. Peptides 7:1189–1192.
306 Stephens, T. W., M. Basinski, P. K. Bristow, J. Bue-Valleskey,S. G. Burgett, L. Craft, J. Hale, J. Hoffmann, H. M. Hsiung,A. Kriauciunas, W. MacKellar, P. R. Rosteck, Jr., B. Schoner,D. Smith, F. C. Tinsley, X.-Y. Zhang, and M. Heiman. 1995.The role of neuropeptide Y in the antiobesity action of the obesegene product. Nature 377:530–532.
307 Stricker, E. M., N. Rowland, C. F. Saller, and M. I. Friedman.1977. Homeostasis during hypoglycemia: central control of ad-renal secretion and pheriferal control of feeding. Science196:79–81.
308 Strubbe, J. H., and C. G. Mein. 1977. Increased feeding inresponse to bilateral injections of insulin antibodies in theVMH. Physiol. Behav. 19:309–313.
309 Sun, M., R. J. Martin, and G. L. Edwards. 1997. ICV beta-hydroxybutyrate: effects on food intake, body composition, andbody weight in rats. Physiol. Behav. 61:433–436.
Journal of Dairy Science Vol. 83, No. 7, 2000
310 Tamas, P., E. Sulyok, I. Szabo, M. Vizer, T. Ertl, W. Rascher,and W. F. Blum. 1998. Changes of maternal serum leptin levelsduring pregnancy. Gynecol. Obstet. Invest. 46:169–171.
311 Tang-Christensen M., P. J. Havel, R. R. Jacobs, P. J. Larsen,and J. L. Cameron. 1999. Central administration of leptin in-hibits food intake and activates the sympathetic nervous sys-tem in rhesus macaques. J. Clin. Endocrinol. Metab. 84:711–717.
312 Taouis, M., J. W. Chen, C. Daviaud, J. Dupont, M. Derouet,and J. Simon. 1998. Cloning the chicken leptin gene. Gene208:239–242.
313 Tartaglia, L. A., M. Dembski, X. Weng, N. Deng, J. Culpepper,R. Devos, G. J. Richards, L. A. Campfield, F. T. Clark, J. Deeds,C. Muir, S. Sanker, A. Moriarty, K. J. Moore, J. S. Smutko, G.G. Mays, E. A. Woolf, C. A. Monroe, and R. I. Tepper. 1995.Identification and expression cloning of a leptin receptor, OB-R.Cell 83:1263–1271.
314 Tarttelin, M. F., and R. A. Gorski. 1973. The effect of ovariansteroids on food and water intake and body weight in the femalerat. Acta Endocrinol. 73:551–568.
315 Tempel, D. L., K. J. Leibowitz, and S. F. Leibowitz. 1988. Effectsof PVN galanin on macronutrient selection. Peptides 9:309–314.
316 Terada, Y., K. Yamakawa, A. Sugaya, and N. Toyoda. 1998.Serum leptin levels do not rise during pregnancy in age-matched rats. Biochem. Biophys. Res. Commun. 253:841–844.
317 Thiele, T. E., D. van Dijk, L. A. Campfield, F. J. Smith, P. Burn,S. C. Woods, I. L. Bernstein, and R. J. Seeley. 1997. Centralinfusion of GLP-1, but not leptin, produces conditioned tasteaversions in rats. Am. J. Physiol. 272:R726–R730.
318 Tucker, H. A. 1985. Endocrine and neural control of the mam-mary gland. Pages 39–79 in Lactation. B. L. Larson, ed. TheIowa State University Press, Iowa.
319 Turton, M. D., D. O’Shea, I. Gunn, S. A. Beak, C. M. Edwards,K. Meeran, S. J. Choi, G. M. Taylor, M. M. Heath, P. D. Lambert,J. P. Wilding, D. M. Smith, M. A. Ghatei, J. Herbert, and S.R. Bloom. 1996. A role for glucagon-like peptide-1 in the centralregulation of feeding. Nature 379:69–72.
320 Tutwiler, G. F., H. J. Brentzel, and T. C. Kiorpes. 1985. Inhibi-tion of mitochondrial carnitine palmitoyl transferase A in vivowith methyl 2-tetradecylglycidate (methyl palmoxirate) and itsrelationship to ketonemia and glycemia. Proc. Soc. Exp. Biol.Med. 178:288–296.
321 Van-Miert, A. S., C. T. van-Duin, and T. Wensing. 1992. Feverand acute phase response induced in dwarf goats by endotoxinand bovine and human recombinant tumour necrosis factoralpha. J. Vet. Pharmacol. Ther. 15:332–342.
322 Van Dijk, G., A. Scheurink, S. Ritter, and A. Steffens. 1993.Glucose homeostasis and sympathoadrenal activity in mercap-toacetate-treated rats. Physiol. Behav. 57:759–764.
323 Vandermeerschen-Doize, F., J. C. Bouchat, M.-A. Bouckoms-Vandermeir, and R. Paquay. 1983. Effects of long-term ad libi-tum feeding on plasma lipid components and blood glucose, b-hydroxybutyrate and insulin concentrations in lean adultsheep. Reprod. Nutr. Dev. 23:51–63.
324 Vandermeerschen-Doize, F., M.-A. Bouckoms-Vandermeir, andR. Paquay. 1982. Effects of long-term ad libitum feeding onthe voluntary food intake, body weight, body composition andadipose tissue morphology of lean adult sheep. Reprod. Nutr.Dev. 22:1049–1060.
325 Vandermeerschen-Doize, F., and R. Paquay. 1984. Effects ofcontinuous long-term intravenous infusion of long-chain fattyacids on feeding behaviour and blood components of adultsheep. Appetite 5:137–146.
326 Vanderweele, D. A., P. F. Xavier, D. Novin, and M. J. Bush.1980. Chronic insulin infusion suppresses food ingestion andbody weight gain in rats. Brain Res. Bull. 5:7–11.
327 Vernon, R. G., and C. M. Pond. 1997. Adaptations of maternaladipose tissue to lactation. J. Mammary Gland Biol. Neopl.2:231–241.
328 Vuagnat, B. A., D. D. Pierroz, M. Lalaoui, P. Englaro, F. P.Pralong, W. F. Blum, and M. L. Aubert. 1998. Evidence for a

SYMPOSIUM: DRY MATTER INTAKE OF LACTATING DAIRY CATTLE 1597
leptin-neuropeptide Y axis for the regulation of growth hor-mone secretion in the rat. Neuroendocrinology 67:291–300.
329 Wade, G. N. 1975. Some effects of ovarian hormones on foodintake and body weight in female rats. J. Comp. Physiol. Psych.88:183–193.
330 Wade, G. N., and J. M. Gray. 1979. Gonodal effects on foodintake and adiposity: a metabolic hypothesis. Physiol. Behav.22:583–593.
331 Wade, G. N., and J. E. Schneider. 1992. Metabolic fuels andreproduction in female mammals. Neurosci. Biobehav. Rev.16:235–272.
332 Wang, T. L., G. L. Edwards, C. A. Baile, and T. L. Wang. 1998.Glucagon-like peptide-1 (7–36) amide administered into thethird cerebroventricle inhibits water intake in rats. Proc. Soc.Exp. Biol. Med. 219:85–91.
333 Warren, E. J., B. N. Finck, S. Arkins, K. W. Kelley, R. W.Scamurra, M. P. Murtaugh, and R. W. Johnson. 1997. Coinci-dental changes in behavior and plasma cortisol in unrestrainedpigs after intracerebroventricular injection of tumor necrosisfactor-alpha. Endocrinology 138:2365–2371.
334 Weingarten, H. P. 1995. Cytokine and food intake: The rele-vance of the immune system to student of ingestive behavior.Neurosci. Biobehav. Rev. 20:163–170.
335 White, B. D., B. He, M. H. Porter, and R. J. Martin. 1996.Nutritional aspects of neuropeptide gene expression. Pages 51–82 in Nutrients and Gene Expression. Clinical Aspects. C. D.Berdanier, ed. CRC Press, Boca Raton.
Journal of Dairy Science Vol. 83, No. 7, 2000
336 White, B. D., and R. J. Martin. 1997. Evidence for a centralmechanism of obesity in the zucker rat: role of neuropeptide Yand leptin. Proc. Soc. Exp. Biol. Med. 214:232.
337 Wilding, J., P. Widdowson, and G. Williams. 1997. Neurobiol-ogy. Br. Med. Bull. 53:286–306.
338 Woods, S. C., E. C. Lotter, L. D. McKay, and D. Porte. 1979.Chronic intracerebroventricular infusion of insulin reducesfood intake and body weight of baboons. Nature 282:503–505.
339 Woods, S. C., D. Porte, Jr., E. Bobbioni, E. Ionescu, J. F. Sauter,F. Rohner-Jeanrenaud, and B. Jeanrenaud. 1985. Insulin: itsrelationship to the central nervous system and to the control offood intake and body weight. Am. J. Clin. Nutr. 42:1063–1071.
340 Woods, S. C., R. J. Seeley, D. J. Porte, and M. W. Schwartz.1998. Signals that regulate food intake and energy homeostasis.Science 280:1378–1383.
341 Yamada, T. 1986. Gut somatostatin. Pages 221–228 in Somato-statin. Basic and Clinical Status. S. Reichlin, ed. Plenum Press,New York.
342 Zamet, C. N., V. F. Colenbrander, C. J. Callahan, B. P. Chew,R. E. Erb, and N. J. Moeller. 1979. Variables associated withperipartum traits in dairy cows. I. Effect of dietary forages anddisorders on voluntary intake of feed, body weight and milkyield. Theriogenology. 11:229–244.
343 Zarjevski, N., I. Cusin, R. Vettor, J. F. Rohner, and B. Jeanren-aud. 1993. Chronic intracerebroventricular neuropeptide-Y ad-ministration to normal rats mimics hormonal and metabolicchanges of obesity. Endocrinology 133:1753–1758.
344 Zhang, Y., R. Proenca, M. Maffei, M. Barone, L. Leopold, andJ. M. Friedman. 1994. Positional cloning of the mouse obesegene and its human homologue. Nature 372:425–432.


















