
INSTITUTO MATERNO INFANTIL PROFESSOR FERNANDO … Brand;;20081125.pdf · PCR aninhada NK Natural...
68
PROGRAMA DE MESTRADO EM SAÚDE MATERNO INFANTIL ESTUDO DO VÍRUS LINFOTRÓPICO DE CÉLULAS T HUMANO (HTLV-1) NA MEDULA ÓSSEA DE PACIENTES PEDIÁTRICOS COM LEUCEMIA LINFOBLÁSTICA AGUDA DE CÉLULAS T (LLA-T) HEIKE ERNA BRAND RECIFE 2008 INSTITUTO MATERNO INFANTIL PROFESSOR FERNANDO FIGUEIRA
Transcript of INSTITUTO MATERNO INFANTIL PROFESSOR FERNANDO … Brand;;20081125.pdf · PCR aninhada NK Natural...

PROGRAMA DE MESTRADO EM SAÚDE MATERNO INFAN TIL
ESTUDO DO VÍRUS LINFOTRÓPICO DE CÉLULAS T HUMANO (H TLV-1)
NA MEDULA ÓSSEA DE PACIENTES PEDIÁTRICOS COM LEUCEM IA
LINFOBLÁSTICA AGUDA DE CÉLULAS T (LLA-T)
HEIKE ERNA BRAND
RECIFE 2008
INSTITUTO MATERNO INFANTIL
PROFESSOR FERNANDO FIGUEIRA

HEIKE ERNA BRAND
ESTUDO DO VÍRUS LINFOTRÓPICO DE CÉLULAS T HUMANO (H TLV-1)
NA MEDULA ÓSSEA DE PACIENTES PEDIÁTRICOS COM LEUCEM IA
LINFOBLÁSTICA AGUDA DE CÉLULAS T (LLA-T)
Linha de pesquisa: Saúde da criança
Dissertação apresentada ao Colegiado do Mestrado em
Saúde Materno Infantil do IMIP como parte dos requisitos
para obtenção do grau de Mestre em saúde Materno-
Infantil.
Orientador: Prof. João Guilherme Bezerra Alves
Co-Orientadora: Dra. Norma Lucena-Silva
RECIFE 2008

Aluna: Heike Erna Brand
Biomédica
Tel. (81) 8778-8069 ou (81) 3231-1589
e-mail. [email protected]
Orientador: Prof . João Guilherme Bezerra Alves
Coordenador do programa de Pós-Graduação do IMIP
Professor Adjunto da UPE
Doutor em Medicina – UFPE
Instituto Materno Infantil Prof. Fernando Figueira – IMIP
Rua dos Coelhos, 300 Boa Vista
Recife – PE – Brasil CEP 50.070-550
Tel. (81) 2122-4122
e-mail. [email protected]
Co-Orientadora: Dr. Norma Lucena-Silva
Pesquisadora Sênior do Centro de Pesquisas Aggeu Magalhães/Fiocruz
Responsável pelo Laboratório de Biologia Molecular do Serviço de Oncologia do
Instituto Materno Infantil Prof. Fernando Figueira – IMIP
Rua dos Coelhos, 300 Boa Vista
Recife – PE – Brasil CEP 50.070-550
Tel. (81) 2122-4764
e-mail. [email protected]

AGRADECIMENTOS
À Diretória do IMIP pelo apoio financeiro através da bolsa de Mestrado no periodo
de Mar 2007-Jun 2008. E ao CNPq pela bolsa de Mestrado a partir de julho 2008.
Ao Serviço de Oncologia Pediátrica do Instituto Materno Infantil, em especial a Dr.
Francisco Pedrosa, pela oportunidade em desenvolver esse projeto.
A Dra Norma Lucena pela excelente orientação, apoio, estímulo, aprendizagem e
confiança em fazer esse trabalho.
Ao Prof. João Guilherme pelo apoio de todo o momento e no direcionamento dos
meus passos.
Aos Professores do Mestrado e a 13. turma do Mestrado, pelo aprendizado, ajuda e
paciência em entender o meu português.
Aos colegas do Laboratório de Biologia Molecular, pela receptibilidade a minha
pessoa e as minhas perguntas.
Às minhas amigas alemãs, Iris, Michaela e Konny, que sempre estiveram presentes
em momentos difíceis, mesmo à distância.
Às minhas amigas brasileiras, Graça, Germannya, Celina e a Família de Purna
Ananda Ashram que me ajudaram em várias formas e me mostraram a beleza do
Brasil.
E, aos meus pais que sempre apoiaram as minhas decisões de vida.

Dedico aos pais
que sofreram a perda de um filho por câncer

Nesta vida, não podemos fazer grandes coisas.
Podemos apenas fazer pequenas coisas com
grande amor.
Madre Tereza de Calcutá

RESUMO
Introdução. A leucemia de células T (LLA-T) ocorre tanto no adulto quanto na criança. O vírus linfotrópico de células T (HTLV-1) é o agente etiológico de uma fração das LLA-T que ocorrem no adulto conhecida por leucemia de células T do adulto (ATL). Mundialmente são 10-20 milhões pessoas infectadas pelo HTLV-1. Contudo, apenas 2-5% dos indivíduos infectados desenvolvem a ATL após 40-60 anos de infecção. A medula óssea tem sido considerada um reservatório para o vírus pela incapacidade das células precursoras hematopoéticas, que albergam o vírus, serem incapazes de expressarem antígenos virais e, por conseguinte, de serem reconhecidas pelo sistema imune, sendo esse um dos mecanismos propostos de evasão da resposta imune em portadores assintomáticos durante latência clínica. A via de transmissão do vírus pela amamentação e durante o parto é um fator de risco para adquirir a ATL, enquanto a transmissão sexual ou endovenosa parece estar relacionada com o desenvolvimento da paraparesia espástica tropical. A criança pode, portanto, ser portadora do vírus já ao nascimento. Contudo, a associação do vírus na leucemogenesis da leucemia linfoblástica aguda de células T (LLA-T) pediátrica é pouco estudada. Objetivo. Determinar da freqüência do HTLV-1 e a expressão de genes virais na medula óssea de pacientes pediátricos com LLA-T e descrever as alterações celulares e características clínicas nos pacientes infectados pelo vírus. Metodologia. Amostras de medula óssea, armazenadas em banco de células, de 24 pacientes pediátricos diagnosticados com LLA-T entre 2004-2007 no Serviço de Oncologia Pediátrica do Instituto Materno Infantil Professor Fernando Figueira (IMIP) foram submetidas à reação em cadeia da polimerase (PCR) específica para detecção do genoma do HTLV-1 e análise de expressão de genes virais e celulares. Resultados. O genoma de HTLV-1 foi encontrado em 6 das 24 (25%) amostras de medula óssea. Mas também, quatro dos pacientes negativos mostraram expressão de genes virais. Considerando ambos resultados, do genoma e de RNA do HTLV-1, a presença do HTLV-1 apresentou uma freqüência de 33%. O grupo de 10 crianças com LLA-T e infecção pelo HTLV-1 apresentou uma menor contagem de leucócitos periféricos (p<0,02) e uma menor expressão celular do fator de transcrição c-fos (p<0,01) em relação ao grupo de 14 crianças com LLA-T sem infecção. Conclusão. A frequência do HTLV-1 e a expressão dos genes virais na medula óssea de crianças com LLA-T foi relatada pela primeira vez. Houve uma menor contagem de leucócitos e expressão celular do fator de transcrição c-fos nas crianças portadores de LLA-T e infectadas pelo HTLV-1. Os dados apontam para o início de atividade viral no portador jovem. A presença do HTLV-1 na medula óssea sugere o envolvimento na leucemogênese suscitando mais estudos.
Palavras-chave : HTLV-1, medula óssea, LLA-T, criança.

ABSTRACT
Introduction. The acute T-cell lymphoblastic leukemia (T-ALL) is a disease which shows manifestation in the adult as well as the child. The Human T-cell linfoma vírus-1 (HTLV-1) is known to be the etiologic agent of a subpopulation of T-ALL which manifests only in the adult, known as adult T-cell leukemia (ATL). Worldwide, 10-20 million people are infected with HTLV-1. However, only 2-5% of infected people develop ATL and only 40-60 years after infection. The bone marrow was thought to be a deposit for virus, due to lacking capacity of hematopoetic stem cells, which are infected by virus, to express viral antigens. This will result in the loss of detection through the immune response in asymptomatic carriers during clinical latency. One risk factor to acquire ATL is the route of transmission through breast milk, whereas sexual or intravenous transmission shows relation in the development of the tropical spastic Paraparesis (HAM/TSP). Therefore, children can already be considered to be carriers of the virus. However the association between the virus and pediatric acute T-cell lymphoblastic leukemia (T-ALL) is rarely studied. Objective. Determination of frequency of HTLV-1 and expression of viral genes in bone marrow of pediatric patients with T-ALL and description of cellular alterations and clinical characteristics of patients infected by the virus. Material and Methods. Bone Marrow (BM) samples, stored at the tumor bank, of 24 pediatric patients diagnosed with T-ALL between 2004-2007 at the pediatric Oncology Service of the Instituto Materno Infantil Prof Fernando Figueira (IMIP) were submitted to polimerase chain reaction (PCR) specific for detection of HTLV-1 provirus and viral gene expression. Results. Proviral DNA of HTLV-1 was found in 6 out of 24 (25%) BM samples. Four patients, negative for detection of provirus showed viral gene expression. Considering both results of provirus and mRNA expression, the presence of HTLV-1 give a frequency of 33%. Those 10 children with T-ALL and HTLV-1 infection showed lower leucocytes counts in blood (p<0,02) and a lower expression level of cellular transcription factor c-fos in relation to the 14 children with T-ALL and without viral infection. Conclusion. The frequency of HTLV-1 and the expression of viral genes in bone marrow in children with T-ALL were reported for the first time. Smaller leukocyte counts and lower expression levels of the cellular transcription factor c-fos in children carriers of T-ALL infected with HTLV-1 was observed. The data indicate a viral activity early in life. The presence of HTLV-1 in bone marrow suggests the involvement of leukemogenesis in T-ALL and recommends more studies. Keywords: HTLV-1, Bone Marrow, T-ALL, Leukemia, pediatric

LISTA DE ABREVIATURAS
ATL leucemia de célula T no adulto
APC célula apresentadora de antígeno
BLV vírus linfotrópico bovino
BM Bone Marrow (Medula óssea)
bp pares de bases
CD cluster of differentiation
(marcador na superfície de células hematopoéticas)
cDNA DNA complementar
CD4+ CD característico para linfócito auxilar
CD8+ CD característico para linfócito citotóxico
CD34+ CD característico para célula tronco hematopoética
CFU Colony forming Unit (unidade formadora de colônia)
CNPq Conselho Nacional de Desenvolvimento científico e tecnológico
CNS Central nervous system (sistema nervoso central)
CTL linfócito T citotóxico
DHL lactato dehidrogenase
DNA ácido desoxiribonucléico
EGIL Grupo Europeu de Estudos de Leucemias
ELISA Enzima linked imunoabsorband assay
Env gene viral codificando proteínas de “envelope”
FAB Grupo cooperativo Franco-Americano-Britânico
Gag gene de antigene viral
HAM/TSP mielopatia associada ao HTLV-1/paraparesia tropical espástica
HIV vírus da imunodeficiência humana
HTLV-1 vírus linfotrópico de células T tipo 1

IDH dermatite infecciosa associada ao HTLV-1
IL Interleucina
IL-R receptor de interleucina
IMIP Instituto Materno Infantil Professor Fernando Figueira
KDa Kilo Dalton
LLA leucemia linfoblástica aguda
LTR terminais longos de repetição
MHC Major Histocompatibility Complex
(complexo de histocompatibilidade maior)
MIC Critérios morfológicos, imunológicos e citogenéticos
MO Medula óssea
MW Molecular Weight (Peso Molecular)
Nested-PCR
PCR aninhada
NK Natural killer cell (célula assasina natural)
ORF Open Reading Frame
PB peripheral blood (sangue periférico)
PBMC células mononucleares do sangue periférico
PCR reação em cadeia da polimerase
Pol gene viral codificando a polimerase viral
primer oligonucleotídeo (iniciador)
Pro gene viral codificando a protease viral
mRNA ácido ribonucléico menssageiro
RT transcrição reversa
SP Sangue periférico
STLV vírus linfotrópico de células T de simio
WHO World Health Organization (Organização Mundial de Saúde)

LISTA DE FIGURAS
Figura 1 Hematopoiese ………………………………………………………….. 1
Figura 2 Mapa geográfico do Brasil mostrando a soroprevalência do
HTLV-1 nos estados…………………………………………………… 5
Figura 3 Mecanismo da infecção das células pelo HTLV-1 ………………… 7
Figura 4 Genoma do HTLV-1 …………………………………………….… 9
Figura 5 Esquema dos diferentes processamentos e Isoformas de mRNAs
virais ……………………………………………………………………... 10
Figura 6 Interação da proteína viral TAX nas vias metabolicas ……………... 13
Figura 7 Modelo da evolução de infecção pelo HTLV-1 até o
desenvolvimento de ATL …………………………………….………... 15
Figura 8 Modelo de latência do HTLV-1 na doença HAM/TSP ……………… 17

SUMÁRIO
1 INTRODUÇÃO .................................................................................. 1
2 OBJETIVOS ...................................................................................... 19
2.1 Objetivo geral .................................................................................. 19
2.2 Objetivos específicos ..................................................................... 19
3 REFERÊNCIAS ................................................................................. 20
4 ARTIGO PRINCIPAL ........................................................................ 28
SEGUNDO ARTIGO 51
5 ANEXOS e APÊNDICES ................................................................. 53
Apêndices I – Termo de Consentimento livre e escla recido
Anexo I – Aprovação Comitê de Ética
Anexo II – Carta de submissão à resvista - segundo artigo

1. INTRODUÇÃO
As leucemias são neoplasias hematológicas malignas e heterogêneas, que
têm a sua origem em células da medula óssea. Os primeiros casos foram relatados
no século XIX com a observação de alteração da medula óssea em pacientes
falecidos1. Desde esta época, o conhecimento sobre a procedência e o
desenvolvimento, tanto dos métodos diagnósticos como das formas de tratamento,
avançaram e de acordo com esses critérios, hoje as leucemias são classificadas em
vários tipos e subtipos.
A hematopoiese consiste no processo de maturação em cascata da linhagem
hematopoiética, iniciada pelas células pluripotentes que residem na medula óssea1
(fig.1).
fig1.
http://images.google.com.br
Dependendo do tipo da linhagem com desenvolvimento comprometido, dar-se
origem a um tipo de leucemia específico, como a leucemia mielóide ou linfóide. A
forma linfóide pode ser dividida segundo as células afetadas em: de células B ou de

células T. Em geral, as leucemias se distinguem em agudas e crônicas. As formas
agudas são caracterizadas pelo acúmulo das células-troncos (blastos) e a perda da
capacidade de diferenciação para células maduras. Modelos mais complexos foram
desenvolvidos para classificar as leucemias, com o objetivo de estratificá-las quanto
ao risco e melhor adequar o protocolo de tratamento. Os modelos variam quanto aos
métodos diagnósticos e se dividem em classificação FAB (Grupo cooperativo
Franco-Americano-Britânico, 1976, critérios morfológicos-citoquímicos), MIC (1986,
critérios morfológicos, imunológicos e citogenéticos), EGIL (Grupo Europeu de
Estudos, 1995, critérios imunofenotípicos) e WHO (World Health Organization, 2001,
critérios citogenéticos/moleculares, história de terapia prévia ou aspectos
mielodisplásicos)2-5 .
O diagnóstico laboratorial da leucemia inclui a hemograma e a mielograma
para a análise morfológica com contagem e testes citoquímicos das células. O
diagnóstico da leucemia aguda é dado pelo aumento de blastos na medula óssea e,
portanto, depende do modelo usado, ou seja, 20% para a WHO ou 30% para o
grupo FAB. A imunofenotipagem é uma técnica importante no diagnóstico, que se
baseia na determinação dos marcadores celulares (Cluster of differentiation, CD),
usando a citometria de fluxo. Com esse método, mudanças quantitativas e
qualitativas na hematopoiese podem ser identificadas, desta forma, permite a
identificação do estágio de maturação do clone leucêmico6. A análise molecular é
outra abordagem metodológica na caracterização do clone leucêmico que visa à
identificação da alteração genética associada a gênesis da leucemia7.
Mundialmente, o câncer é a primeira causa de morte. De acordo com a WHO,
em 2005, mais de 70% de todas as mortes causadas pelo câncer ocorreram em
países de renda per capita média ou baixa8. O câncer pediátrico representa 0,5% a

3% de todos os tumores, na maioria das populações. Os tumores predominantes no
paciente infantil são as leucemias. No Brasil, as leucemias abrangem 15 a 45% entre
todos os tumores infantis e no Recife, 22,57% de casos novos de tumores, são
devido a essa doença hematológica9,10. A leucemia linfoblástica aguda (LLA)
representa 80% de todos os casos de leucemia na criança e 15% desses são devido
às células T7, 11.
A LLA-T se encontra tanto no adulto quanto na criança. A etiologia não é bem
determinada mas fatores de risco ambientais (radiação durante gravidez, fumo) ou
fatores genéticos foram discutidos. Queixas e manifestações clínicas inespecíficas
envolvem febre, anemia, leucócitos baixo ou >100.000/µl, fadiga, dor óssea e
possível envolvimento do sistema nervoso central (SNC). As formas de tratamento
incluem quimioterapia, radioterapia, uso de esteróides e transplantes de medula
óssea. Na criança, a taxa de remissão é alta, chegando a atingir 100%, embora a
sobrevida livre de eventos em 10 anos, é apenas cerca de 63% e no adulto 25-35%7.
A leucemia de célula T do adulto (ATL) representa mais uma entidade entre
as leucemias linfóides e não deveria ser confundida com a LLA-T do adulto. Na
criança foram relatados casos esporádicos, com sintomas parecidos de ATL12, mas
em geral, a doença se refere só ao adulto.
A ATL foi descoberta no Japão em 1977 e existem quatro subtipos distintos.
Estes estão relacionados às características clínicas e se dividem em agudo, crônico,
smoldering e tipo linfoma. Manifestações clínicas típicas são hipercalcemia, lesões
de pele, crescimento de linfonodo, elevação de lactato desidrogenase (DHL),
números de leucócitos aumentados e massa de mediastino revelado ao raio X. Além
disso, na forma aguda também se encontram no esfregaço de sangue periférico,
linfócitos com uma morfologia característica, as chamadas flower cells13. As formas

agudas e tipo linfoma são mais agressivos e, o prognóstico é reservado. O tempo de
sobrevida médio, relacionado a essas formas, é de um ano. Ao lado da
quimioterapia combinada e transplantes de células-troncos alogênicas como as de
cordão umbilical, novas formas de tratamento incluindo drogas alvo de estruturas
moleculares e anticorpos monoclonais têm surgido14, 15.
HTLV-1
De acordo com a WHO, o HTLV-1 é reconhecido como agente etiológico da
ATL. Ele é um retrovírus delta, que faz parte da família Retroviridae de gênero de
HTLV-BLV16. Ele foi o primeiro retrovírus associado ao câncer no ser humano e
descoberto em 198017. Além da ATL, o vírus também é responsável por outras
doenças, como a mielopatia associada ao HTLV-1/paraparesia espástica tropical
(HAM/TSP), uma doença neurodegenerativa; a uveite, uma doença oftalmológica; a
dermatite infecciosa (IDH) e a artropatia inflamatoria crônica associada ao HTLV-1
(HAAP)18, 12,19. Outros tipos de HTLV foram descorbertos. O HTLV-2, apesar de não
estar relacionado ao desenvolvimento de leucemia, foi isolado primeiro num paciente
com a leucemia de células pilosas20. Recentemente, os HTLV-3 e HTLV-4 foram
isolados em indivíduos na África21.
Mundialmente 10-20 milhões pessoas são infectadas com HTLV-1. Áreas
endêmicas com mais de 1% da população infectada são: Japão, África Sub-
Saariano, América do Sul e as Ilhas do Caribe22. De acordo com dados mais
recentes obtidos em estudos com doadores de sangue, o Brasil tem o maior número
absoluto de pessoas infectadas pelo HTLV-1 no mundo. Estimativas calculam 2,5
milhões de Brasileiros infectados23. Um estudo feito nas capitais dos 27 estados no
Brasil mostrou a maior prevalência na região Norte e Nordeste. Com 10,0/1000 em

São Luís (Nordeste) comparados com 0,4/1000 em Florianópolis (Sul). Pernambuco
mostrou uma prevalência de 7,5/1000 pessoas infectadas24 (fig.2).
fig2. Catalan-Soares B et al. 2005
O vírus é transmitido sexualmente, através de sangue contaminado,
verticalmente pela amamentação ou durante o parto12,25. Mas, apenas 2-5% dos
portadores do HTLV-1 desenvolvem uma das doenças relacionadas ao vírus e
geralmente só 40 a 60 anos pós-infecção, sendo, portanto, a maioria dos indivíduos
infectados portadores assintomáticos15.
O diagnóstico de infecção pelo HTLV-1 é feito através da determinação de
anticorpos no soro pelo ELISA que são produzidos contra proteínas estruturais do
vírus26. Contudo, se mostrou que este teste pode subestimar a prevalência da
infecção. Um estudo de prevalência, realizado em Salvador/Brasil, com 1385
indivíduos não conseguiu detectar anticorpos em menores de 13 anos de idade27.

Testes confirmatórios são o Western Blot, que detecta proteínas virais e a PCR, que
identifica DNA proviral nas células mononucleares do sangue periférico26.
Uma característica do retrovírus é o genoma viral constituído de duas fitas
simples de RNA. O RNA viral, após a entrada na célula, é transcrito reversamente
em DNA viral, chamado provírus e assim pode se integrar no genoma da célula do
hospedeiro. A integração do HTLV-1 no genoma ocorre de forma aleatória e é
específica para cada célula infectada16.
Para a disseminação do vírus, existem várias possibilidades. Em contraste
com outros retrovírus, como o HIV, infecções de células pelo virion de HTLV-1 no
sobrenadante (livre de células) não se mostraram eficientes, todavia essa
possibilidade foi documentada em testes in vitro28. O HTLV-1 infecta uma célula
principalmente pelo contato célula infectada com célula não infectada. Na conjunção
entre essas células se forma um centro organizado de microtúbulos (MTOC) que é
estabilizado pelas várias moléculas celulares. No MTOC se cria uma sinapse viral
através da qual o RNA viral consegue entrar na célula ainda não infectada (fig.3)29.
Uma outra possibilidade para a disseminação do vírus e a forma mais
importante, inclui a proliferação da célula infectada. Assim o provírus integrado no
genoma do hospedeiro é entregue à célula filha durante o processo da divisão
celular (mitose)15.

fig.3 Matsuoka et al. Nature Reviews 2007
O HTLV-1, após a infecção, preferencialmente se integra no genoma
de células linfóide T (CD4+), aonde as proteínas virais conduzem as alterações
intracelulares15. Assim, durante muito tempo, pensou-se que o vírus tinha um
tropismo para esse tipo de célula30. Entretanto, outras células também mostraram
suscetibilidade ao vírus, como linfócitos T supressores29, linfócitos T citotóxico,
monócitos do sangue periférico, macrofágos, células dendríticas, células NK,
linfócitos B, astrócitos presentes no SNC, células gliais, fibroblastos, células
endoteliais e epiteliais31. Concomitantemente, as células CD34+ mostraram
suscetibilidade à infecção com HTLV-1. Análises com CD34+ humanas em
camundongos transgênicos mostraram a presença do provírus em todas as
linhagens hematopoéticas (incluindo linfócitos T, monócitos/macrófagos), depois da
diferenciação in vitro e in vivo. Se a infecção pelo HTLV-1 também altera o padrão
da diferenciação para uma linhagem específica conduzido, por exemplo, pela
ativação ou repressão de genes diferentes, ainda não está esclarecido32.
Em humanos, a suscetibilidade das células CD34+ ao vírus tem mostrado
resultados controversos. Um estudo utilizando a metodologia da PCR-Hibridização in

situ mostrou que em pacientes com HAM/TSP até 95% das células da medula óssea
(MO) estavam infectadas com provírus, apesar do mRNA viral não estar expresso31.
Outro estudo em pacientes com ATL usando a PCR mostrou a presença do vírus em
células mononucleares isolados da MO, contudo, células CD34+ purificadas através
da citometria de fluxo mostraram-se não infectadas32.
A defesa celular envolve os linfócitos T como principal mediador. A população
de linfócitos T se divide em células T auxiliares (CD4+), em células citotóxicas
(CD8+) e em células T supressoras (CD4-, CD8+). Os linfócitos T auxiliares e
citotóxicos circulam entre os órgãos linfóides ou não linfóides e o sangue periférico
(SP), procurando complexos de histocompatibilidade maior (MHC) na superfície das
células. Esses linfócitos interagem através dos seus receptores de células T (TCR)
com os MHCs das células, enquanto os linfócitos T supressores modulam a resposta
imunológica negativamente33. As alterações sofridas pelos linfócitos infectados
resultam em imortalidade das células e depois em transformações que prejudicam a
sua funcionalidade comprometendo a defesa, permitindo infecções oportunistas,
especialmente por Strongyloides stercoralis, Pneumocystis jiroveci e Mycobacterium,
que são frequentes em pacientes com ATL. Na infecção pelo HTLV-1, os CTLs
destroem as células CD4+ infectadas controlando a carga proviral sendo, portanto,
um dos principais reguladores da infecção15. Todavia, foi mostrado que durante a
infecção natural com HTLV-1, um número significante de CTLs também podem ser
infectadas; em conseqüência, essas CTLs se programam para a auto-destruição e
assim limitam a defesa antiviral31.
O genoma viral tem um tamanho de nove mil pares de bases (9kb) e é
dividido em várias regiões, a saber: LTR, gag, pol, env e pX (fig. 4). Em ambos as
extremidades (5´e 3´) se localizam os longos terminais de repetição (LTR), que têm

seqüências idênticas e servem como promotores para o vírus e a regulação da
transcrição dos RNA’s virais. As seqüências gag, pol e env representam genes que
codificam as proteínas estruturais e a região pX codifica as proteínas reguladoras do
vírus34. A ausência de um local de integração específica do HTLV-1 no genoma do
hospedeiro e a falta de um oncogene no genoma viral, sugeriu que um produto viral
deveria conduzir a imortalização das células infectadas. Várias proteínas virais são
codificadas na região pX, cuja atividade de imortalização em células primárias CD4+
tem sido atribuída35.
fig.4 Matsuoka et al. Nature Reviews 2007
Quando o provírus é integrado ao genoma e a célula T é ativada, um conjunto
de fatores de transcrição celular e co-fatores intracelulares se ligam ao LTR e iniciam
um nível basal de transcrição de mRNAs virais. Este é um processo complexo e a
etapa de iniciação ainda não está completamente entendida31. O acúmulo celular da
proteína viral TAX estabiliza e fortalece a ligação de fatores celulares ao LTR,
aumentando a transcrição dos genes virais16.
A primeira fita de RNA transcrita, representa uma fita de ribonucleotídeos,
completa da seqüência do provírus. Esse RNA primário (unspliced) será submetido a
um processamento alternativo que remove os íntrons e recombina os éxons em
seqüências determinadas. O provírus possui três éxons e os transcritos produzidos
podem ser distinguidos como unspliced, single spliced ou double spliced,
dependendo dos éxons incluídos36 (fig. 5). Assim, o vírus consegue criar uma
variedade de fitas de mRNA com um genoma viral relativamente pequeno.

Recentemente, um transcrito adicional foi descoberto codificando a proteína HTLV-1-
bZip (HBZ). Esse transcrito é produzido pela ação do promotor 3´LTR e está
codificado na fita negativa do provírus37. A transcrição de HBZ pode ser feita apartir
de dois sítios de iniciação e por isto resulta em dois transcritos de mRNA
diferentes38.
fig.5 Matsuoka et al. Nature Reviews 2007
Quantificações de mRNAs virais em vários tipos de células infectadas,
mostraram variabilidade na expressão, entre os tipos das células. Entretanto, os
níveis de expressão dos mRNAs entre si, foram iguais em cada célula39. Após a
transcrição dos genes virais, os mRNAs são traduzidos no citoplasma em proteínas,
usando a maquinaria da célula do hospedeiro.
A proteína REXp27 é uma proteína de tamanho 27 kDa que regula a
expressão dos genes virais pós-transcricional. Ela é responsável pelo transporte dos
mRNA’s virais unspliced ou single spliced, do núcleo para o citoplasma16. Uma
proteína REX truncada, de 21 kDa (p21Rex), também foi observada. Esta proteína é
traduzida a partir de um transcrito entre éxon 1 e 3 (single spliced) que é

preferencialmente produzido de provírus defeituosos (com deleções nas seqüências
gag, pol e env).
A função de p21 não é completamente esclarecida, mas uma ação
antagonista ao REXp27 foi sugerida16,40. A p12 possivelmente participa bloqueando a
apoptose da célula T dependente de interleucina 2 (IL-2). Adicionalmente, uma
redução da expressão das proteínas MHC classe I na superfície da célula foi
observada em células que expressam p1231. A proteína viral de 30kDa (p30) age
como modulador negativo pós-transcricional na expressão das proteínas virais41. A
proteína p13 é feita por um processamento simples do mRNA entre éxon 1 e éxon 3.
Na proteína p13 existe uma seqüência sinal para localização mitocondrial
(mitochondrial targeting signal, MTS) onde a p13 conduz alterações morfologicas à
mitocondria31. O interesse pela proteína HBZ surgiu mais recentemente a partir de
experimentos com camundongos transgênicos de TAX. Esses camundongos
desenvolveram uma série de tumores, contudo, nenhuma relação entre TAX e o
desenvolvimento de leucemia foi relatada, sugerindo a participação de uma outra
proteína viral no desenvolvimento da ATL e a possibilidade de HBZ estar envolvida
neste processo42. A HBZ mostrou uma função bimodal dependendo da forma em
que se apresenta, mRNA ou como proteína43. Além disso, a HBZ interage com
fatores de transcrição inibindo a interação entre eles e a TAX ou as seqüências em
promotores44. A expressão de mRNA HBZ é de 20-50x menor do que o mRNA Tax.
Todavia, ao contrário do mRNA de Tax, o mRNA de HBZ sempre é expresso em
células primárias de ATL. Ao lado da função supressora da proteína HBZ, o mRNA
de HBZ figura como estimulador na proliferação das células T42.
A TAX é uma fosfoproteína de 40kDa preferencialmente localizada no núcleo
na maioria das células e é resultado de um processamento duplo (double splicing)

de mRNA primário, entre éxon 1, 2 e 3. Esta proteína tem um papel pleiotrópico e é
a mais estudada entre as proteínas virais. As suas principais funções são: o
reconhecimento pelo sistema imunológico, a modificação da transcrição dos mRNAs
virais, a imortalização, transformação e oncogenêses das células infectadas pelo
HTLV-1. Esse processo ocorre nos vários níveis que envolve a modulação na
transcrição de genes celulares, repressão de apoptose, progressão do ciclo celular,
inativação de genes de supressores de tumores, amplificação de centrosomos e
dano de DNA estrutural31,16,45-47.
A modulação na transcrição exercida por TAX geralmente leva a um aumento
na transcrição de genes celulares incluindo os fatores de crescimento, citocinas,
receptores de fatores de crescimento, moléculas de adesão celular, transmissores
de sinais citoplasmáticos e fatores de transcrição nuclear15. A única repressão da
transcrição induzida pela TAX é a da enzima responsável pelo reparo de DNA, a
DNA polimerase beta16. A proteína não se liga diretamente ao promotor, mas
interage fisicamente pelos seus domínios com fatores de transcrição. Entre eles,
Nuclear Factor-kappa B (NFκB), Serum Response Factor (SRF) , Sp1/CBP, AP-1 e
c-Amp responsive element binding protein (CREB)16 (fig.6).
A seqüência sinal para exportação da TAX permite que a TAX produzida no
citoplasma seja secretada pela célula48. A estimulação de genes endógenos pela
TAX extracelular foi comprovada49. Contudo, a TAX no citoplasma mostrou um papel
importante na modulação de algumas vias de sinalização, interagindo com uma
variedade de proteínas e resultando em alterações downstream na via, como a via
de NFκB que também está envolvida em vários processos celulares50,31.

fig. 6 Yao et al. Frontiers in Bioscience
2000
HTLV-1 e ATL
As células envolvidas na ATL, encontradas no sangue periférico, são
predominantemente CD4+ e em 90-99% dessas células foram achados HTLV-1
positivos30. Para o desenvolvimento da doença, a célula infectada tem de passar por
várias etapas. A primeira etapa após a infecção é a imortalização, que resulta em
proliferação das células e é caracterizada pelo crescimento dependente de IL-2.
Esta proliferação é resumida a expressão constitutiva da cadeia α do Receptor IL-2
(IL-2R), que forma junto com as cadeias β e γ, um receptor funcionalmente ativo.
Porém, isto pode não ser suficiente para desenvolver a ATL sendo necessário mais
alterações ao nível genético ou epigenético e, provavalmente, uma alteração na
defesa imunológica do hospedeiro51. Após a imortalização, a célula sofre um
processo de transformação. Nesse processo, a célula desenvolve a possibilidade da
proliferação independente da ação de IL-2, mesmo na presença de uma expressão
aumentada da cadeia α de IL-2R52,16.

O processo inicial da transformação, a razão da latência clínica prolongada e
o porquê apenas 2-5% das pessoas infectadas com HTLV-1 desenvolvem a ATL,
ainda não são conhecidos. Vários fatores de riscos foram descritos, entre eles, o
sexo31, a hereditariedade15, aumento do número de linfócitos atípicos associado à
carga proviral53,54, baixo nível de anticorpos anti-TAX no soro54 e a transmissão
vertical de HTLV-115.
As mulheres geralmente apresentam uma maior prevalência de infecção pelo
HTLV-1, por serem mais vulneráveis à transmissão do vírus por via sexual. Todavia,
os homens têm uma probabilidade 40% maior em adoecer com ATL31. Um maior
número de casos de ATL foi descrito em famílias sugerindo uma predisposição
genética para desenvolver esse tipo de leucemia15. Pacientes com IDH durante a
infância, geralmente manifestam ATL55.
A mutagêneses na formação de tumores, pela inserção do retrovírus no
genoma, é bem documentada em animais56. O local da integração do provírus do
HTLV-1 no genoma do hospedeiro varia entre os pacientes com ATL, mas mostrou
uma integração preferencial em regiões ricas em nucleotídeos AT57 e, 53% das
integrações ocorrem em regiões de genes58. Adicionalmente, em cerca de 30% dos
pacientes existe uma correlação alta entre o número de cópias integradas por célula
e a manifestação clínica da doença, tendo 50,6% mostrado uma única cópia do
provírus enquanto em 20,6% dos casos foram encontrados múltiplos locais de
integração em células mononucleares de sangue periférico16.
Uma observação feita em células leucêmicas isoladas de paciente com ATL
em contraste com células infectadas em HAM/TSP é a ausência ou expressão baixa
da proteína viral TAX. Transcritos foram detectados apenas em aproximadamente
40% de células de ATL. A TAX causa alterações intracelulares, mas também induz a

resposta imunológica. Dessas observações, resultou a idéia de que a maioria das
células infectadas seria destruída logo pelas CTLs. Assim, apenas a célula infectada
que desenvolveu um mecanismo de baixar o nível da expressão das proteínas virais,
não seria atingida pela destruição e sobreviveria. Esta célula continuaria a se
proliferar, aumentando os números de células infectadas no corpo e se tornaria
leucêmica. Esta idéia explica, como as células infectadas na ATL conseguem se
expandir, mesmo com o sistema imunológico não afetado (fig. 7)15.
fig.7 Yasunaga J-I et al. Cancer Control 2007
Vários mecanismos para escapar da defesa imunológica foram propostos.
Provírus defeituosos, com deleções de seqüências gag-pol59, mutações nas
seqüências de TAX no local de ligação com HLA-A60 foram relatados. A perda da
região LTR no genoma viral em 39% dos casos com ATL, que é necessário para a
expressão dos mRNAs virais, também foi descrito61. Outro mecanismo de escape
imune proposto a partir da observação de mais de 50 regiões hipermetiladas no
genoma celular encontradas em células de ATL foi a ocorrência de modificações
epigenéticas, que interferem no processo da transcrição de genes62.
Contudo, o maior fator de risco em adquirir a ATL em comparação com outras
doenças causadas por HTLV-1, é a via da transmissão do vírus. A amamentação foi

relacionada a ATL, enquanto a infecção intravenosa ou por via sexual, mostrou uma
maior ocorrência em HAM/TSP63. A hipótese é, que a via da primeira infecção dê ao
vírus acesso para um conjunto de células específicas influenciando de forma
diferente o curso da doença. A maioria das células dentríticas e macrófagos se
encontram em membranas de mucosa. Assim pensou-se, que essas seriam as
células alvos do vírus, durante a infecção inicial pela amamentação31. As células
dendríticas são células apresentadoras de antígenos (APC) que eficientemente
capturam antígenos na periferia, deslocando-se aos órgãos linfáticos secundários
onde estimulam células nativas T e B ou células de memória31. Ratos infectados
oralmente com HTLV-1 mostraram uma resposta imunológica baixa em comparação
com as vias de transmissão intraperitoneal ou intravenosa, todavia, a carga proviral
era alta64. Isto pode ser atribuído às células dendríticas e macrófagos que
permanecem na fase pós-mitótica no ciclo vital, e como conseqüência promovem
uma expressão baixa de proteínas virais, que são necessários para o
reconhecimento pelas CTLs31.
A latência clínica é prolongada nas ATL e HAM/TSP e caracterizada como o
estado que envolve uma resposta imunológica efetiva e a intervenção na expansão
do vírus. A evasão da resposta imunológica pelos mecanismos que envolvem a
perda da expressão das proteínas virais já foi mencionada. Experimentos feitos em
macacos mostraram mais uma alternativa: a medula óssea como reservatório do
vírus. Logo após a infecção dos macacos via intravenosa, o provírus foi localizado
em células da MO e do sangue periférico31. As células CD4+ e CD8+ normalmente
migram entre os tecidos linfóides incluindo a MO e o sangue periférico, desta forma
os linfócitos T infectados conseguem entrar em contato com as células-troncos
hematopoéticas (CD34+)31 (fig. 8).
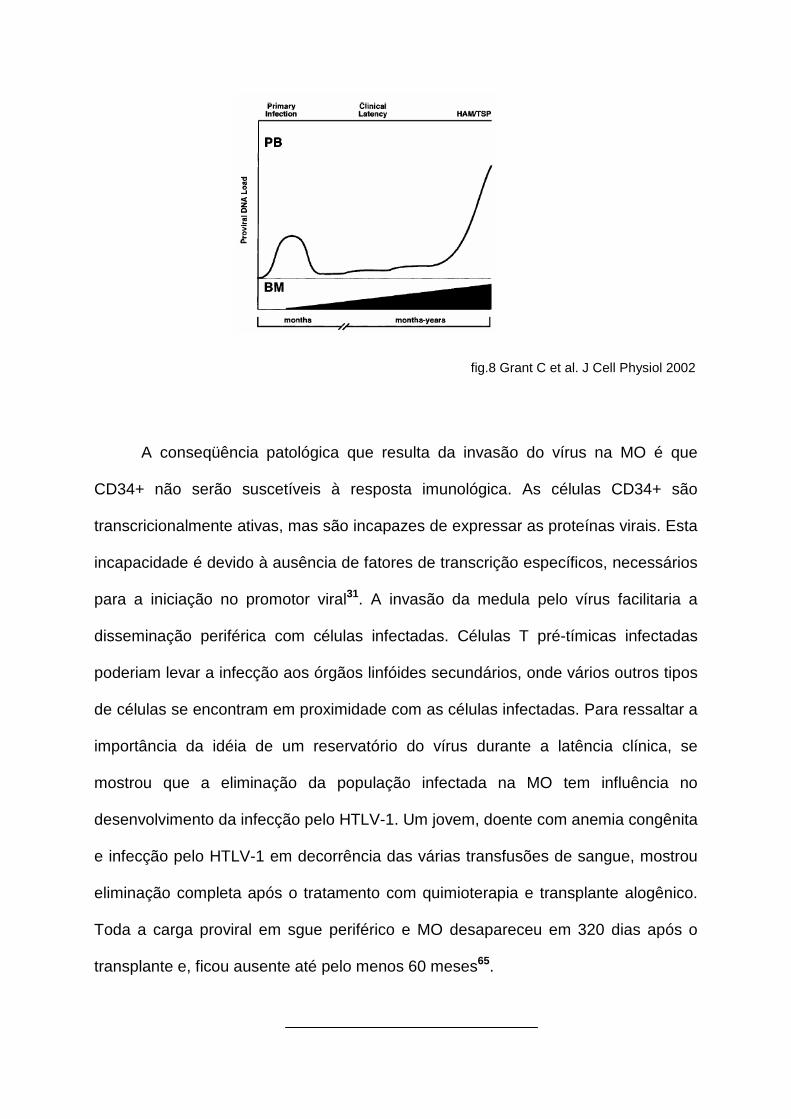
fig.8 Grant C et al. J Cell Physiol 2002
A conseqüência patológica que resulta da invasão do vírus na MO é que
CD34+ não serão suscetíveis à resposta imunológica. As células CD34+ são
transcricionalmente ativas, mas são incapazes de expressar as proteínas virais. Esta
incapacidade é devido à ausência de fatores de transcrição específicos, necessários
para a iniciação no promotor viral31. A invasão da medula pelo vírus facilitaria a
disseminação periférica com células infectadas. Células T pré-tímicas infectadas
poderiam levar a infecção aos órgãos linfóides secundários, onde vários outros tipos
de células se encontram em proximidade com as células infectadas. Para ressaltar a
importância da idéia de um reservatório do vírus durante a latência clínica, se
mostrou que a eliminação da população infectada na MO tem influência no
desenvolvimento da infecção pelo HTLV-1. Um jovem, doente com anemia congênita
e infecção pelo HTLV-1 em decorrência das várias transfusões de sangue, mostrou
eliminação completa após o tratamento com quimioterapia e transplante alogênico.
Toda a carga proviral em sgue periférico e MO desapareceu em 320 dias após o
transplante e, ficou ausente até pelo menos 60 meses65.

A partir de um estudo feito na Europa, que mostrou uma soroprevalência de
HTLV-1 seis vezes maior em gestantes do que em doadores de sangue, é esperado
que as soroprevalências encontradas no Brasil estejam subestimadas66. Embora a
via de transmissão pela amamentação e a possibilidade da criança nascer portadora
do vírus, a associação da LLA-T ao vírus em pacientes pediátricos é pouco
investigada. Estudos na criança encontrados na literatura científica principalmente
envolvem a determinação de anticorpos, diagnóstico molecular em células
mononucleares do sangue periférico ou focam a manifestações clínicas12,67. A LLA-T
é caracterizada pela proliferação de células-troncos CD34+ da medula óssea com
uma leucocitose elevada e tem um prognóstico per si mais reservado. Investigar a
participação do vírus na génesis da LLA-T e o possível controle que ele exerce na
célula regulando a expressão de genes celulares poderá contribuir para uma melhor
estratificação dos casos e abordagem terapêutica.
Nesse estudo, determinamos a presença de HTLV-1 na medula óssea de
pacientes infanto-juvenis com LLA-T e a expressão de isoformas de mRNA virais.
Também foram investigadas as alterações intracelulares induzidas pelo vírus e a
influência da presença viral nas características clínicas e celulares dos pacientes.

2 OBJETIVOS
2.1. OBJETIVO GERAL
Determinar a freqüência do HTLV-1 na medula óssea de pacientes pediátricos
portadores de leucemia de células T, diagnosticados entre 2004-2007 no Serviço de
Oncologia Pediátrica do Instituto Materno Infantil Professor Fernando Figueira (IMIP)
no Recife, Brasil.
2.2. OBJETIVOS ESPECÍFICOS
Em pacientes pediátricos de LLA-T diagnosticados entre 2004-2007 no Serviço de
Oncologia Pediátrica do Instituto Materno Infantil Professor Fernando Figueira (IMIP)
no Recife, Brasil:
1. Determinar a freqüência do provírus de HTLV-1 na medula óssea.
2. Determinar a expressão de genes virais na medula óssea.
3. Identificar possíveis alterações induzidas pelo vírus no nível da expressão de
protooncogenes.
4. Descrever a associação entre a presença viral e as variáveis biológicas,
clínicas e laboratoriais.

3 REFERÊNCIAS
1. Oliveira, RAG e Poli Neto, A. Anemias e Leucemias. São Paulo: ROCA, 2004
2. Bennett JM, Catovsky D, Daniel MT et al. Proposals for the classification of the acute
leukemias. Br J Haematol. 1976;33:451-458
3. Bennett J.M, Juliusson G, Mecucei C. Meeting Report Morphologic, immunologic and
cytogenetic classification of the chronic (mature) B and T lymphoid leukemias.
Fourth Meeting of the MIC Cooperative Study Group. Cancer Res 1990;50:2212
4. Bene MC, Castoldi G, Knapp W. Proposals for the immunological classification of
acute leukemias. European Group for the Immunologic al Characterization of
Leukemias (EGIL). Leukemia 1995;9:1783-1786
5. Jaffe ES, Harris NL, Stein H. Pathology and Genetics of Tumours of Haematopoietic
and Lymphoid Tissues . World Health Organization Classification of Tumours .
Lyon, France: IARC Press, 2001
6. Craig FE et al. Flow cytometric immunophenotyping for hematologic n eoplasms.
Blood 2008
7. Redaelli A, Laskin BL, Stephens JM, Botteman MF, Pashos CL A systematic literature
review of the clinical and epidemiological burden o f acute lymphoblastic
leukaemia (ALL). Eur J Cancer Care 2005;14(1):53-62
8. WHO - Cancer , fact sheet No. 297, February 2006. Disponível em:
http://www.who.int/mediacentre/factsheets/fs297/en/index.html Acesso em: Jan 2008
9. WHO – Global Info Base ; Disponível em:
http://www.who.int/infobase/report.aspx?rid=126 Acesso em: 10. Jan 2008
10. de Souza Reis R, de Oliveira Santos M, Santos Thuler MC. Incidência de tumores
pediátricos no brasil. Rev Brasil Cancerolog 2007;53(1):5-15
11. Uckun FM, Sensel MG, Sun L, Steinherz PG, Trigg ME, Heerema NA, Sather HN,
Reaman GH, Gaynon PS. Biology and treatment of childhood T-lineage acute
lymphoblastic leukemia. Blood. 1998 Feb 1;91(3):735-46

12. Bittencort A.L, Primo J, de Fatima Paim de Oliveira M. Manifestations of the human T-
cell lymphotropic virus type 1 infection in childho od and adolescence. J Pediatr
2006;82(6):411-20
13. Takatsuki K. Adult T-cell Leukemia. Internal Medicine 1995;34:947-952 Review
14. Takizawa J, Aoki S, Kurasaki T, Higashimura M, Honma K, Kitajima T, Momoi A,
Takahashi H, Nakamura N, Furukawa T, Aizawa Y. Succesful treatment of adult T-
cell leukemia with unrelated cord blood transplanta tion. Am J Hematol. 2007
Dec;82(12):1113-5
15. Yasunaga J-I, Matsuoka M. Human T-Cell Leukemia Virus Type I induces Adult T-
Cell Leukemia: From clinical aspects to molecular m echanisms. Cancer Control
2007;14:133-140
16. Yao J, Wigdahl B Human T Cell Lymphotropic Virus Type 1 Genomic expr ession
and impact on intracellular signaling pathways duri ng neurodegenerative
disease and Leukemia. Frontiers in Bioscience 2000;5:138-168
17. Poiesz BJ, Ruscetti FW, Gazdar AF, Bunnn PA, Minna JD, Gallo RC. Detection and
isolation of type C retrovirus particles from fresh and cultured lymphocytes of a
patient with cutaneous T-cell lymphoma. PNAS 1980; 77(12):7415-9
18. Gessain A, Barin F, Vernant JC et al. Antibodies to human T-lymphotropic virus type-
1 in patients with tropical spastic paraparesis. Lancet. 1985; 2:407-410
19. Nishioka K, Maruyama I,, Sato K, Kitajima I, Nakajima Y, Osame M. Chronic
inflammatory arthorpathy associated with HTLV-1. Lancet 1989;1:441
20. Kalyanaraman V.S, Sarngadharan M.G, Robert-Guroff M, Miyoshi M, Golde D, Gallo
R.C. A new subtype of human T-cellleukemia virus (HTLV-2 ) associated with a
T-cell variant of hairy cell leukemia. Science 1982;218:571
21. Mahieux R, Gessain A. New human retroviruses: HTLV-3 and HTLV-4. Med Trop
(Mars) 2005; 65(6):525-8
22. Proietti FA, Carneiro-Proietti AB, Catalan-Soares BC, Murphy EL. Global epidemiology
of HTLV-1 infection and associated diseases. Oncogene 2005;24:6058-6068

23. Carneiro-Proietti AB, Ribas JG, Catalan-Soares BC, Martins ML, Brito-Melo GE, Martins-
Filho OA. Infecção e doença pelos vírus linfotrópicos humanos de célula T
(HTLV1/2) no Brasil. Rev Soc Bras Med Trop 2002;35:499-508
24. Catalan-Soares B, Carneiro-Proietti F, Proietti F.A. Heterogeneous geographic
distribution of human T-cell lymphotropic viruses 1 and 2 (HTLV1/2):
serological screening prevalence rates in blood don ors from large ruban areas
in Brazil. Cad. Saúde Pública 2005;21(3):926-931
25. Kashiwagi K, Furusyo N, Nakashima H, Kubo N, et al. A decrease in mother-to-child
transmission of human T lymphotropic virus type 1 ( HTLV-1) in Okinawa,
Japan. Am J Trop Hyg 2004;70(2):158-163
26. Thorstensson R, Albert J, Andersson S. Strategies for diagnosis of HTLV-1 and –2.
Transfusion 2002;42(6):780-91
27. Dourado I, Alcantara LC, Barreto ML, da Gloria Teixeira M, Galvão-Castro B. HTLV-1 in
the general population of Salvador, Brazil: a city with African ethnic and
sociodemographic characteristics. J Acquir Immune Defic Syndr. 2003;34(5):527-
31
28. Seto A, Isono T, Ogawa K. Infection of inbred rabbits with cell-free HTLV-I. Leuk Res
1991;15:105-10
29. Matsuoka M, Jeang K-T. Human T-cell leukemia virus type 1 (HTLV-1) infecti vity and
cellular transformation. Nature Review Cancer 2007;7:270-280
30. Richardson JH, Edwards AJ, Cruickshank JK, Rudge P, Dalgleish AG. In vivo cellular
tropism of human T-cell leukemai virus type I. J Virol. 1990;64(11):5682-5687
31. Grant C, Barmak K, Alefantis T, Yao J, Jacobson S, Wigdahl B. Human T cell leukemia
Virus Type 1 and neurologic disease: Events in Bone Marrow, Peripheral Blood,
and Central Nervous System During Normal immune sur veillance and
Neuroinflammation. J Cellular Physiology 2002; 190:133-159
32. Nagafuji K, Harada M, Teshima T, Eto T, Takamatsu Y, Okamura T, Murakawa M,

Akashi K, Niho Y. Hematopoetic Progenitor Cells from patients with ad ult T-cell
leukemia-lymphoma are not infected with human T-cel l leukemia virus Type I.
Blood 1993;82:2823-2828
33. Abbas A.K, Lichtman A.H, Pober J.S. Imunologia cellular e molecular. Rio de Janeiro:
Revinter, 2002
34. Seiki M, Hattori S, Hirayama Y, Yoshida M. Human adult T-cell leukemia virus:
complete nucleotide sequence of the provirus genome integrated in leukemia
cell DNA. PNAS 1983;80:3618-3622
35. Grassmann R, Dengler C, Müller-Fleckenstein I, Fleckenstein B, McGuire K, Dokhelar M-
C, Sodroski J.G, Haseltine W.A. Transformation to continuous growth of primary
human T-lymphocytes by human T-cell leukemia virus type I X-region gene
transduced by a herpesvirus saimiri vector. PNAS 1989;86:3351-3355
36. Ciminale V, Pavlakis G.N, Derse D, Cunningham C.P, Felber B.K. Complex splicing in
the human T-cell leukemia virus (HTLV) family of re troviruses: novel mRNAs
and proteins produced by HTLV type 1. J Virol. 1992;66(3):1737-1745
37. Cavanagh MH, Landry S, Audet B, Arpin-André C, Hivin P, Paré ME, Thête J, Wattel E,
Marriott SJ, Mesnard JM, Barbeau B.
HTLV-1 antisense transcripts initiating in the 3´LT R are alternatively spliced and
polyadenylated. Retrovirology 2006;3:15
38. Murata K, Hayashibara T, Sugahara K, Uemura A, Yamaguchi T, Harasawa H,
Hasegawa H, Tsuruda K, Okazaki T, Koji T, Miyanishi T, Yamada Y, Kamihira S. A
novel alternative splicing isoform of human T-cell leukemia virus type 1 bZIP
factor (HBZ-SI) targets distinct subnuclear localiz ation. J Virol. 2006;80:2495-
2505
39. Li M, Green P. Detection and Quantification of HTLV-1 and HTLV-2 m RNA Species
by Real time PCR. J Virol Mehtods. 2007;142(1-2):159-168
40. Orita S, Kobayashi H, Aono Y, Saiga A, Maeda M, Igarashi H. p21X mRNA is

expressed as a singly spliced pX transcript from de fective provirus genomes
having a partial deletion of the pol-env region in human T-cell leukemia virus
type 1 infected cells. NAR 1993;21:3799-3807
41. Nicot C, Harrod RL, Ciminale V, Franchini G. Human T-cell leukemia7lymphoma virus
type I nonstructural genes and their functions. Review. Oncogene 2005;24:6026-
6034
42. Mesnard J-M, Barbeau B, Devaux C. HBZ, a new important player in the mystery of
adult T-cell leukemia. Blood 2006; 108:3979-3982
43. Satou Y, Yasunaga J, Yoshida M, Matsuoka M. HTLV-1 basic leucine zipper factor
gene mRNA supports proliferation of adult T cell le ukemia cells. PNAS.
2006;103:720-725
44. Lemasson I, Lewis MR, Polakowski N, Hivin P, Cavanagh MH, Thébault S, Barbeau B,
Nyborg JK, Mesnard JM. Human T-cell leukemia virus type 1 (HTLV-1) bZIP
interacts with the cellular transcription factor CR EB to inhibit HTLV-1
transcription. J Virol 2007;81:1543-1553
45. Tabakin-Fix Y, Azran I, Schavinky-Khrapunsky Y, Levy O, Aboud M. Functional
Inactivation of p53 by human T-cell leukemia virus type 1 Tax protein:
mechanism and clinical implication. Carcinogenesis 2006;27:673-681
46. Nitta T, Kanai M, Sugihara E, Tanaka M, Sun B, Nagasawa T, Sonoda S, Saya H, Miwa
M. Centrosome amplification in adult-T cell leukemia a nd human T-cell leukemia
virus type 1 Tax induced human T cells. Cancer Sci 2006;97:836-841
47. Jin D.Y, Spencer F, Jeang K.T. Human T cell leukemia virus type 1 oncoprotein Tax
targets the human mitotic checkpoint protein MAD1. Cell 1998;93:81-91
48. Jain P, Mostoller K, Flaig KE, Ahuja J, Lepoutre V, Alefantis T, Khan ZK, Wigdahl B.
Identification of Human T-cell leukemia virus type 1 TAX amino acid signals and
cellular factors involved in secretion of the viral onocprotein. JBC
2007;282:34581-34593
49. Aono H, Fujisawa K, Hasunuma T, Marriott SJ, Nishioka K. Extracellular Human T cell

Leukemia virus type 1 Tax protein stimulates the pr oliferation of Human
synovial cells. Arth & Rheum 1998;41:1995-2003
50. Dejardin E. The alternative NF-kB pathway from biochemistry to biology:pitfalls and
promises for future drug development. Biochem Pharmacol 2006;72:1161-1179
51. Giam CZ, Jeang KT. HTLV-1 Tax and adult T-cell leukemia Front. Biosci
2007;12:1496-1507
52. Franchini G. Molecular Mechanisms of Human T-Cell Leukemia/Lymph otropic virus
type 1 Infection. Blood 1995;86:3619-3639
53. Hisada M, Okayama A, Shioiri S, Spiegelman D, Stuver SO, Mueller NE. Risk factors
for adult T-cell leukemia among carriers of human T -lymphotropic virus type 1.
Blood 1998;92:3557-3561
54. Hisada M, Okayama A, Tachibana N, Stuver SO, Spiegelman DL, Tsubouchi H, Mueller
NE. Predictors of level of circulating abnormal lymphoc ytes among human T-
lymphotropic virus type 1 carriers in Japan. Int J Cancer 1998;77:188-192
55. Hanchard B, LaGrenade L, Carberry C, Fletcher V, Williams E, Cranston B, Blattner WA,
Manns A. Childhood infective dermatitis evolving into adult T-cell leukemia after
17 years. Lancet 1991;338:1593-4
56. Hayward WS, Neel BG, Astrin SM. Activation of a cellular onc gene by promotor
insertion in ALV-induced lymphoid leucosis. Nature 1981;290:475-80
57. Leclercq I, Mortreux F, Cavrois M, Leroy A, Gessain A, Wain-Hobson S, Wattel E. Host
sequences flanking the human T-cell leukemia virus type 1 provirus in vivo. J
Virol 2000;74:2305-12
58. Hanai S, Nitta T, Shoda M, Tanaka M, Iso N, Mizoguchi I, Yashiki S, Sonoda S,
Hasegawa Y, Nagasawa T, Miwa M. Integration of the HTLV-1 in genes of
leukemia cells of patients with adult T-cell leukem ia. Cancer Sci 2004;95:306-310
59. Shu M, Hill S.A, Derse D. Defective and Wild-Type Human T-cell Leukemia Virus
type I Proviruses: Characterization of Gene Product s and trans-Interactions
between Proviruses. Virology 1999;262:442-451

60. Furukawa Y, Kubota R, Tara M, Izumo S, Osame M. Existence of escape mutant in
HTLV-1 tax during the development of adult T-cell l eukemia. Blood. 2001;97:987-
993
61. Takeda S, Maeda M, Morikawa S, Taniguchi Y, Yasunaga J, Nosaka K, Tanaka Y,
Matsuoka M. Genetic and epigenetic inactivation of tax gene in adult T-cell
leukemia cells. Int J Cancer. 2004;109:559-567
62.Taniguchi Y, Nosaka K, Yasunaga J, Maeda M, Mueller N, Okayama A, Matsuoka M.
Silencing of human T-cell leukemia virus type 1 gen e transcription by
epigenetic mechanisms . Retrovirology. 2005;2:64
63. Barmak K, Harhaj E, Grant C, Alefantis T, Wigdahl B. Human T cell leukemia virus
type I-induced diseas: pathways to cancer and neuro degeneration. Virology
2003; 308(1):1-12
64. Hasegawa A, Ohashi T, Hanabuchi S, Kato H, Takemura F, Masuda T, Kannagi M.
Expansion of human T-cell leukemia virus type 1 (HT LV-1) reservoir in orally
infected rats: inverse correlation with HTLV-1-spec ific cellular immune
response. J Virol. 2003;77:2956-63
65. Kawa K, Nishiuchi R, Okamura T, Igarashi H. Eradication of human T-lymphotropic
virus type 1 by allogeneic bone-marrow transplantat ion. Lancet 1998;352:1034-
1035
66. Taylor GP, Bodéus M, Courtois F, Pauli G, Del Mistro A, Machuca A, Padua E,
Andersson S, Goubau P, Chieco-Bianchi L, Soriano V, Coste J, Ades AE, Weber
JN.The seroepidemiology of human T-lymphotropic viruse s: types 1 and 2 in
Europe: a prospective study of pregnant woman. J Acquir Immune Defic Syndr
2005;38(1):104-9
67. Pombo-de-Oliveira MS, Dobbin JA, Loureiro P, Borducchi D, Maia RC, Fernandes MA,
Cavalcanti GB Jr, Takemoto S, Franchini G. Genetic mutation and early onset of T-
cell leukemia in pediatric patients infected at bir th with HTLV-I. Leuk Res.
2002;26:155-61


4 ARTIGO PRINCIPAL
baseado nas normas da Revista international:
Leukemia Research

Detection of Human T-Cell Leukemia Virus Type I (HT LV-1) Provirus
and viral mRNA Isoforms, in Bone Marrow of Pediatri c Acute T-cell
Leukemia patients
AUTHORS
Brand, Heike1
Neves, Lídia1
Alves, João Guilherme Bezerra1
Pedrosa, Francisco1
Lucena-Silva, Norma1,2
The Pediatric Oncology Service, IMIP1 and Aggeu Magalhães Research
Center/Fiocruz2.
Corresponding author: Dr. Norma Lucena-Silva, The Pediatric Oncology
Service/Instituto Materno Infantil Prof. Fernando Figueira; Rua dos Coelhos, n.300,
Quinto andar, Boa Vista, cep:50070-550, Recife, Pernambuco, Brazil. Tel.: 55 81
21224764. e-mail: [email protected]
JOURNAL SUBMISSION
Leukemia Research [Elsevier]

Abstract
To investigate the association of HTLV-1 in pediatric T-cell leukemia we used
patient’s bone marrow for determining the virus presence and the gene activity of
proto-oncogenes normally upregulated by viral protein TAX. Proviral DNA was
detected in 6 out of 24 (25%) patients. Interestingly, four provirus-negative samples
showed viral gene expression. The expression of protooncogene c-fos is suggestive
of downregulated, depending on proviral load or viral mRNA expression. Overall,
these data indicate HTLV-1 infiltration of BM early in life and show the possibility of
viral gene expression in a cellular compartment, so far known as being transiently
silent.
Keywords: HTLV-1, T-cell leukemia, Bone Marrow, pediatric, Provirus, mRNA

I. Introduction
Acute Leukemias represent the most frequent type of leukemia in children and 15%
of those have T-cell origin. The remission rate is high, around 80%, but the event free
survival for more than 10 years lies at aprox. 63% [1].
The Human T-Cell Leukemia Virus Type I (HTLV-1) is known to be the etiologic
agent of adult T-cell leukemia (ATL) and other HTLV-1 related diseases, like HTLV-1
associated myelopathy/tropical spastic paraparesis (HAM/TSP) [2]. One risk factor to
acquire ATL is the route of transmission of virus, via breast milk [3]. However, only 2-
5% of infected people develop ATL and only after a long latent period of 40-60 years
[4].
The cells mainly affected by HTLV-1 infection, are CD4+ T-lymphocytes. These cells
have the capacity to shuttle into the Bone Marrow (BM), where they thought to be
able to infect CD34+ hematopoetic stem cells. BM was proposed to be a viral
reservoir, since asymptomatic carriers need to develop a mechanism to escape host
immune surveillance during clinical latency. The lack of expression of specific cellular
transcription factors in hematopoetic stem cells, normally necessary for viral gene
transcription, would enable CD34+ cells to escape from host immune defence.
During differentiation into more mature cells, the expression of those factors will
recuperate [5] and transcriptional active HTLV-1 infected cells could than be
constantly be delivered to peripheral blood (PB). In vitro and In vivo studies have
already shown susceptibility of CD34+ cells to HTLV-1 and detection of viral
sequences in all hematopoetic lineages, after differentiation [6]. In situ-Hibridization
technique used in HAM/TSP patients, showed 90-95% of cells in BM positive for
proviral load, although the virus was not found to be transcriptional active in these
cellular compartment [7]. These data was confirmed in BM of ATL patients, although
the infection of CD34+ progenitor cells was not detectable [8].
In the present study, we used the nested-PCR technique to detect provirus of HTLV-
1 in BM samples of pediatric patients with T-ALL. In order to demonstrate the
possibility of active viral gene transcription in this cellular compartment, we measured
the expression of viral mRNA isoforms. In an attempt to study the influencies of virus
on the cellular environment, we did a relative quantification of the c-fos and c-myc
mRNA. Finally, we investigated wether the presence of the HTLV-1 in BM is related
to differences on clinical and laboratorial variables.


II. Material and Methods
2.1. Patients
BM samples collected for diagnosis purpose in the Pediatric Oncology department at
the Instituto Materno Infantil Prof. Fernando Figueira (IMIP) Recife/Brazil are
afterwards stored at -80oC in a tumor bank with the authorization of the child legally
responsable. From the 31 patients diagnosed with T-ALL in the Oncology
department, between 2004 and 2007, 24 which still had BM stored were included in
our study. Clinical and Imunophenotyping data were obtained from clinical records.
Approval for the study was obtained from the institutional review board and informed
consent was provided, according to the declaration of Helsinki.
2.2. Detection of Provirus
5x106 cells were used for Isolation of genomic DNA, using DNAzol BD Reagent
(Invitrogen) according to the manufacturer’s instruction. DNA was purified afterwards,
using Phenol and precipitated with Ethanol under standard protocols. 100ng DNA
was used as template for the 1.PCR and 1µl out of the 1.PCR was used as template
in the Nested PCR. Conditions used in both PCR’s were, 1x PCR buffer, 0.75mM
MgCl2, 300µM dNTP’s, 1U Taq (Biotools) and 25pmol of each forward and reverse
primer (Figure 1). GAPDH primers were used in a separated reaction. PCR program
used for both Provirus PCR’s were: 95ºC for 3 min, 40 cycles at 95ºC 30 sec, 60ºC
30 sec, 72ºC 30 sec, and 72ºC for 7 min. For GAPDH 30 cycles 95ºC 3 min, 55ºC 1
min, 72ºC 2 min, and 72ºC for 7 min. Amplified products were separated on 2%
agarose gel, bands cut and purified using PURE Link Quick Gel extraction Kit
(Invitrogen).
2.3. Sequencing
10ng of purified PCR product was used for sequencing including 3.2pmol/µl specific
forward or reverse primer, 0.5µl Big Dye, 1µl Save Money Buffer in a total volume of
10µl. Conditions used for reaction were 92ºC 2min, 40 cycles 92ºC 15 sec, 55ºC 15
sec, 60ºC 4min. Samples were afterwards precipitated with Isopropanol, washed,
resuspended in 10µl High Dye Formamide and submitted for reading in the DNA
Sequencer 3100 (Applied/Hitashi).

2.4. Detection of viral mRNA Isoforms
5x106 cells were used for Isolation of total RNA using TRIzol Reagent (Invitrogen),
according to the manufacturer’s instruction. The RNA was resuspended in 20µl
H20/Depc and 5 µl used for treatment with 10U DNase I for 30min at 37ºC. DNase I
was inactivated by storing samples over night at -80ºC. Following day, c-DNA
synthesis was made using Superscript II RT (Invitrogen) and 0.2µg/µl Random
Hexamer primers in total volume of 20µl, following instructions of manufacturer. A
DNase I Control was set up (data not shown) for sample No. 20, by using the same
RT conditions, without adding Superscript Enzyme. Following, 1µl was used as
template for the 1.RT-PCR and for the Nested-PCR, 1µl out of the 1.RT-PCR. Primer
combinations for each HTLV-1 RNA reaction see Figure 1b. Reaction conditions for
PCRs were the same as mentioned above. Program conditions for the 1.PCR were
94ºC 3min, 40cycles 94ºC 1min, 58ºC 1min, 72ºC 1min and 72ºC 7 min. For the
Nested PCR 30 sec were used for denature, annealing and extension. GAPDH
program conditions were as used for genomic DNA. c-myc and c-fos RT-PCR
conditions were 94ºC 3min, 30 cycles 94ºC30sec, 58ºC 30sec, 72ºC 30sec and 72ºC
7 min. Amplified PCR products were separated on 2% Agarose Gel, cut and Gel
purified as mentioned above and submitted to sequencing. Protooncogene
expression was quantified and calibrated with GPDH expression levels using the
Kodak 1 D3.5 software.
2.5. Statistical analysis
The Epi-info program, Version 6.04B was used for the statistical analysis. The CDs
counts above 20% were considered positive. The continuous variables LDH, c-fos
and c-myc were segregated in two groups using the median value as cut off. Age and
leucocytes groups were choosen based on the literature. The qualitative and discrete
variables were represented by absolute frequency (n) and relative frequency (%) and
their significance analyzed using the chi-square test or Fisher’s exact test. The level
of rejection for the hypothesis of nullity (having HTLV-1) was always equal to or less
than 0.05% (α=5%).
III. Results

3.1. Characteristics of patient sample
All 24 patients included in this study were from Pernambuco, a state in the Northeast
of Brazil, which shows one of the highest rates of HTLV-1 prevalence (7.5/1000)
nation wide. Age at diagnosis lied between 1 to 15 years, 18 patients where male
and 6 female. High blastic infiltration rates (56 to 98%) were observed in all patient
BM samples, and the imunofenotyping using cytoCD3, CD3, CD45, CD34, CD8,
CD4, CD7 antibodies confirmed the diagnosis of T-cell leukemia. Leucocytes counts
and serum lactate dehydrogenase varied widely. Four patients at time of study had
already died. Mediastinal Mass was present in 14 (58%) out of 24 cases (Table 1).
3.2. Detection of Provirus in BM
Six patients (No. 3, 6, 9, 10, 18, 20) where tested positive for HTLV-1 in Bone
Marrow, showing a specific PCR product of 180bp in size (Figure 2). Unspecific PCR
products were present in Sample No. 11 and No. 3. Bands from sample No. 3, 6 and
9 were cut, gel purified and submitted to sequencing which confirmed specific
sequence for HTLV-1. The observed result corresponds to a frequency of 25% of
HTLV-1 infected BM in pediatric patients.
3.3. Detection of viral gene expression
Splicing is used in HTLV-1, to generate a variety of mRNA transcripts. Examining
different viral mRNA Isoforms four samples where shown to be transcriptionally
active, albeit not belonging to the same samples shown to be positive for provirus
(Figure 3).
Sample No. 5 was tested positive by using primer of Exon3 in the pX region.
Sequencing of the expected 180bp band confirmed HTLV-1 sequence. Due to primer
localization it was not possible to distinguish between unspliced mRNA, single
spliced env mRNA Isoforms or even viral genomic RNA, but other possible viral
mRNA variants like p30II or p12I could be excluded, due to no observed product in
the same sample, when using other primer sets.
Sample No. 25 was the unique positive for the Isoform HBZ. Expected size was
203bp.
Sample No.2, 26 and 27 showed different sized bands by using primers constructed
to detect mainly single spliced (Exon1 to Exon3) or some of the double spliced

mRNA Isoforms. Expected band sizes under PCR conditions chosen were 328bp
(Tax/Rex), 213bp (p13II), 138bp (p21Rex). Sample No. 2 showed an unusual band
size of aprox. 260bp. Sequencing of the band revealed a 88% homology with human
pyridoxal (pyridoxine, vitamin B6) kinase (PDXK). No. 26 showed 3 bands at aprox.
260bp, 220 and 140bp and No. 27 showed two bands at aprox. 320 and 140bp.
Using primers for detection of double spliced (Exon2-Exon3) mRNA Isoforms,
expected band sizes under condition chosen were 589bp (p12I), 529bp (p30I) and
57bp (Tax/Rex). The 1.PCR showed a band of aprox. 180bp in samples No. 1, 8, 9,
10, 11, 12, 17, 18, 19, 20, 22 and 27. The Nested-PCR showed the same band size,
although the bands were much fainter and disappeared in sample No. 8, 20 and 22.
Bands from the 1.PCR of sample No. 1 and 10 were cut, gel purified and sequenced.
Sequencing revealed a 95% identity with 5/131 Gaps, for MAD1-like protein in both
samples.
3.4. Expression profile of c-myc and c-fos
All samples were tested in RT-PCR for proto-oncogene c-myc and c-fos gene
expression and quantified and calibrated with GAPDH expression levels. The
samples showed a large variable expression level, with median values of 35 for c-
myc and 117 for c-fos (Table 1).
3.5. Analysis of HTLV-1-infected T-ALL
Considering HTLV-1 positive-samples that were positive for provirus DNA or for
expression of unspliced or HBZ mRNA, the frequency of viral detection even
increases to 33%. HTLV-1 infected patients showed leucocytes counts and c-fos
expression levels statistically different from the non-infected patients. c-fos showed
expression levels lower than its median value in 87.5% (p=0.02) and leucocytes
counts lower than 100.00/mm3 in 75% (p=0.01) of the HTLV-1 positive cases.
Nevertheless, the differences between HTLV-1 positive and negative samples were
seen with precaution, because of the small sampling (Table 2).
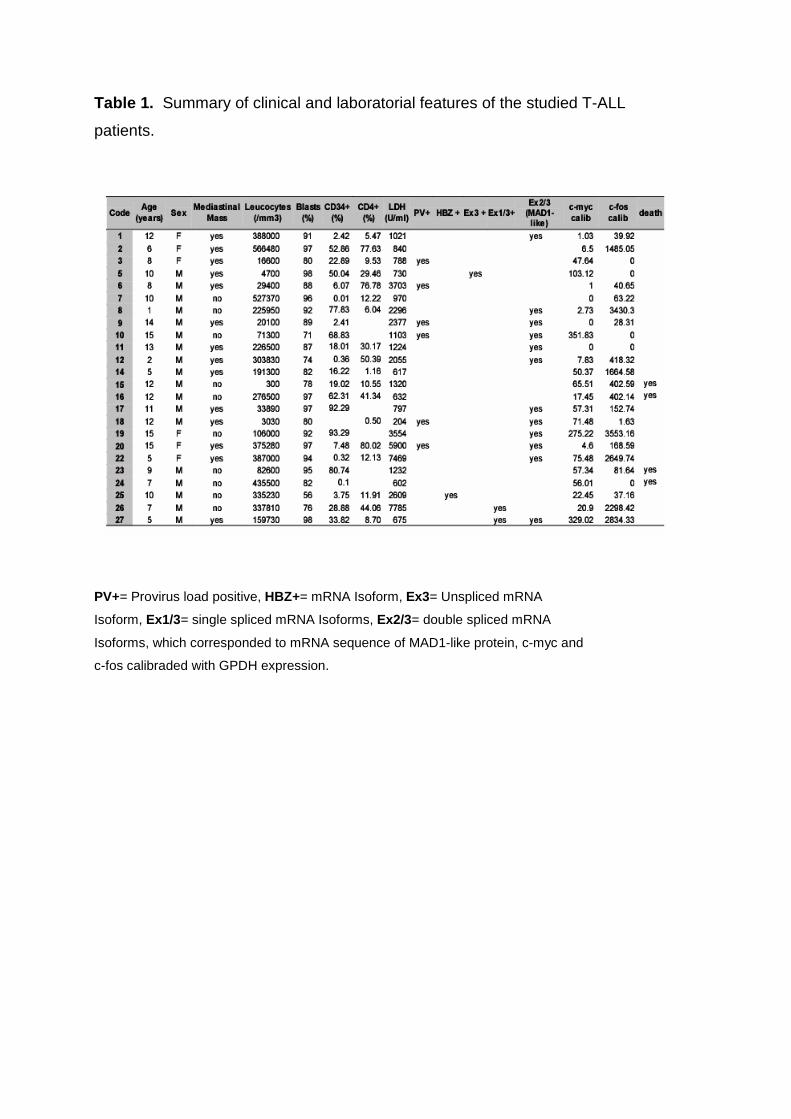
Table 1. Summary of clinical and laboratorial features of the studied T-ALL
patients.
PV+= Provirus load positive, HBZ+= mRNA Isoform, Ex3= Unspliced mRNA
Isoform, Ex1/3= single spliced mRNA Isoforms, Ex2/3= double spliced mRNA
Isoforms, which corresponded to mRNA sequence of MAD1-like protein, c-myc and
c-fos calibraded with GPDH expression.
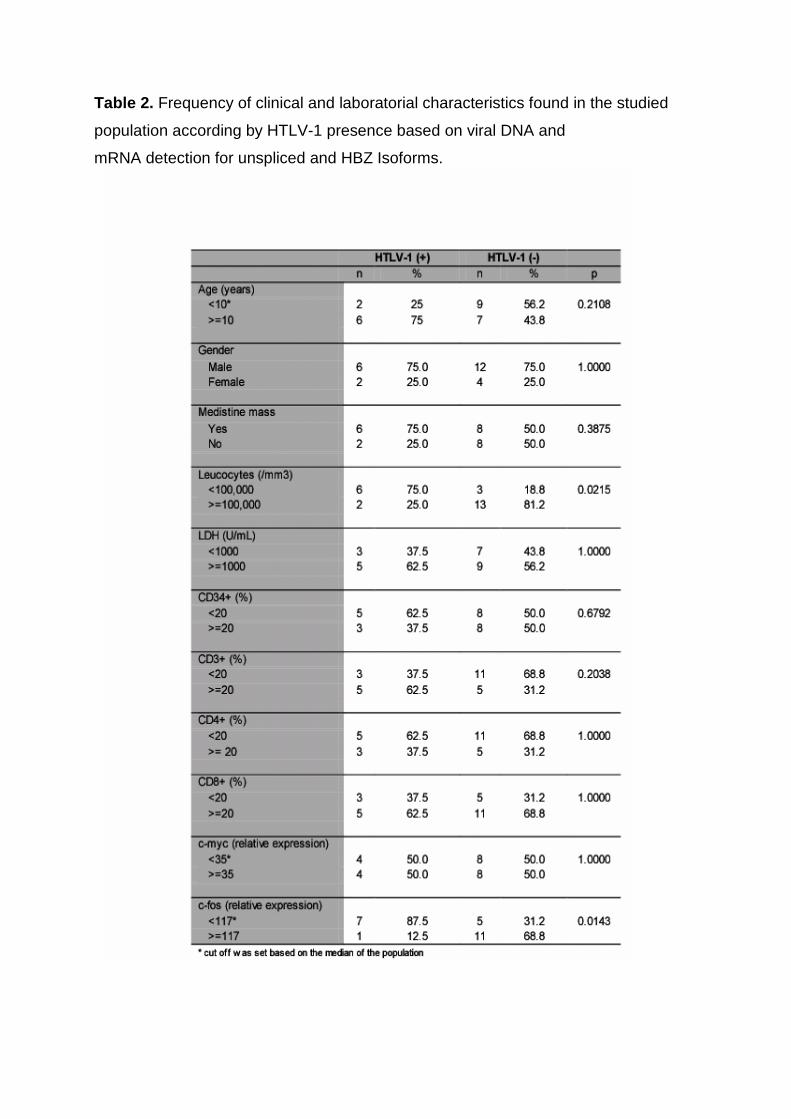
Table 2. Frequency of clinical and laboratorial characteristics found in the studied
population according by HTLV-1 presence based on viral DNA and
mRNA detection for unspliced and HBZ Isoforms.
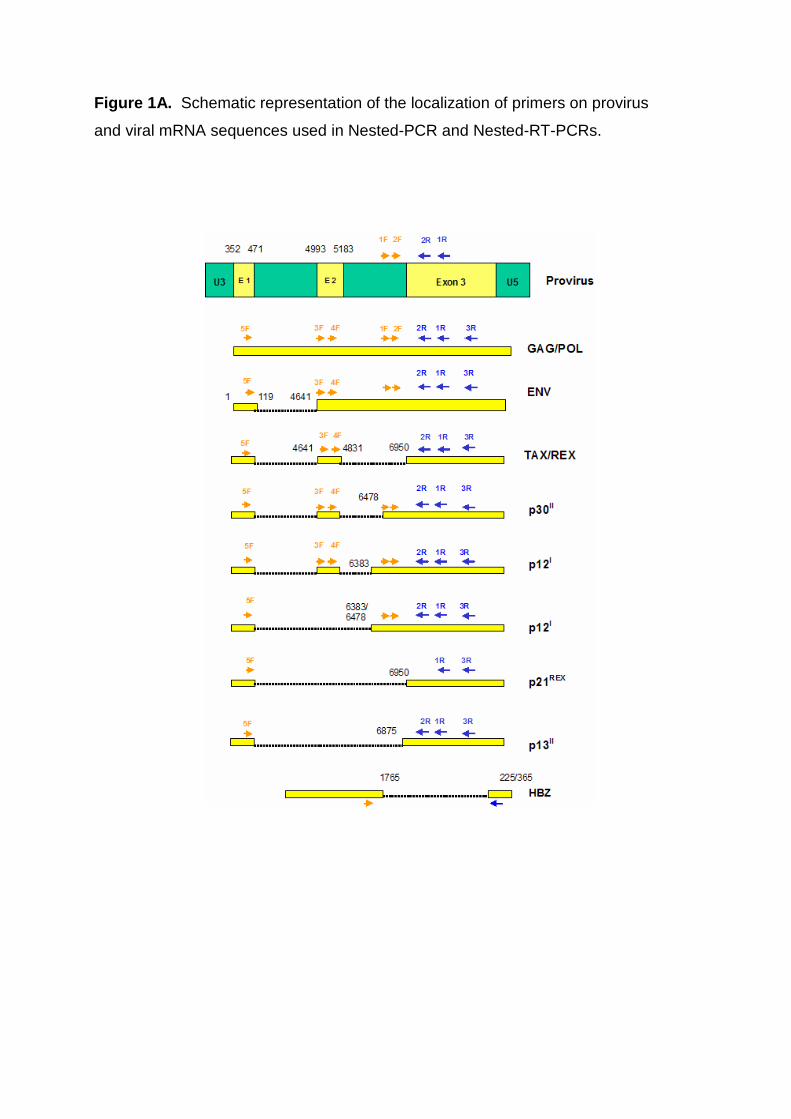
Figure 1A. Schematic representation of the localization of primers on provirus
and viral mRNA sequences used in Nested-PCR and Nested-RT-PCRs.

1B. Sequence description of primers used.
Figure 2. Detection of HTLV-1 provirus load using Nested-PCR.

PV= Provirus GAPDH= internal control
Sample No. 3, 6, 9, 10, 18 and 20 show specific amplified bands with expected size of 180bp. Band
pattern above observed in sample No. 3 and No. 11 were unspecific. GPDH was used as an internal
control.
PV GAPDH

Figure 3. Detection of viral mRNA Isoforms using different sets of primers in
Nested-RT-PCR.
Ex3= unspliced, expected size of 2.PCR is 180bp, No.5 positive; Ex2/3= double spliced, expected
size of 1.PCR is 241bp, No. 1, 8, 9, 10, 11, 12, 17, 18, 19, 20, 22, 27 showing smaller band size
corresponding to mRNA for MAD1-like protein; Ex1/3= single spliced, expected size of 2.PCR are
fragments of 328, 213, 138bp in size, No. 26 and 27 positive (although not sequenced yet), No.2
showed smaller product size sequence corresponding to human pyridoxal (pyridoxine, vitamin B6)
kinase (PDXK); HBZ expected size of 2.PCR for is 203bp, No. 25 positive. GAPDH was used as
internal control.
Ex3 Ex2/3 Ex1/3 HBZ GAPDH

4. Discussion
An estimated 10-20 million people worldwide are infected by HTLV-1. Endemic
regions with more then 1% of population infected are South America, Japan, Africa
Sub-Saharan and the Caribbean Islands [9, 10]. The association of HTLV-1 in
pediatric T-cell leukemia is rarely described and usually focuses on PB samples or
clinical features [11-17]. HTLV-1 infection of BM was reported to be a mechanism for
stable viral infection within a host with intact immune system and thought to be a
source for constant viral delivery to peripheral blood and lymphatic organs [5].
Furthermore, complete elimination of proviral load in PB was reported after treatment
with chemotherapy and allogenic BM transplantation [18]. In our study 25% of BM
samples from T-ALL diagnosed children were tested positive for proviral load.
Primers for this study were located in the pX region of the viral genome where have
been shown deletions or mutations [19] and therefore the chance of even higher
rates of BM samples infected by HTLV-1, can not be excluded.
A study which determined the seroprevalence in over 1300 individuals in a high
endemic region in the Northeast of Brazil was not able to detect antibodies in humans
younger than 13 years old [20]. Also considering the hypothesis made by Grant et al.,
that cells in BM of HAM/TSP patients are getting saturated by HTLV-1 infection over
a longer period of time [6], would indicate loss of detection in younger patients, due
to a small amount of proviral template in the sample. In our study, 4 out of 6 patients
positive for proviral load belonged to an elderly population (12-15 years old), which
would corroborate to this idea mainly if the viral transmission was due to breast
feeding. Vertical transmission of virus via breast milk was also reported to be time
dependent, showing infection rates of 5.1% for <3 month and 38.5% ≥ 12 month of
breastfeeding [3]. Unfortunately, we were not able to confirm the HTLV-1 exposure to
the patient’s mother.
Due to the choice of technical procedure, discrimination between cell-types present in
the BM compartment was not possible and therefore it cannot be postulated that
CD34+ cells were infected. However, cell to cell contact between uninfected and
infected cells and a cellular receptor ubiquitinately expressed on a variety of cell
types was thought to be necessary for infection with HTLV-1 [21]. This would suggest
the possibility of CD34+ stem cell infection on later time points, transmitted through
other cell types already infected with HTLV-1 and also present in the BM.

Acute Leukemia is characterized by increasing blast numbers in the BM
compartment. In order to investigate the possibility of involvement of viral proteins on
regulating the increase of blast cell, it was studied viral gene expression in our patient
samples. Interestingly, four samples showed viral gene expression, although proviral
load was not detectable in these samples. Single and Multiple insertion of provirus
into one genome have been reported and showed correlation with manifestation of
disease in 30% of patients. Single copy integration was shown in 50.6% whereas
multiple insertions were observed in only 20.6% of those patients [21]. The lack of
detection of provirus in samples showing viral gene expression could, as mentioned
above, be due to limitations in sensitivity of the technique used or failed primer
binding. Moreover, the high expression level of viral mRNA in cells with undetectable
integration of HTLV-1 genome suggests an active infection. Also defective provirus
were observed in 25-40% of all provirus integrated in lymphocytes, which could
explain why no expression level was observed in those 6 samples positive for proviral
load (22,23).
Gag and env are structural proteins, necessary for packaging of viral nucleocapsid
and made out of unspliced or single spliced mRNA transcript, respectively [24].
cDNA of sample No.5 has been shown to be positive using primers located in intron
and exon3 regions of the virus. These primers enable detection of those transcripts
and viral genomic RNA, suggesting active virus particle in patient sample No. 5.
Proviral DNA contamination could be excluded, since RNA samples were digested
with DNase I prior to c-DNA synthesis.
Recent studies focus on the importance of a new viral protein HBZ, coded on the
minus strand of the HTLV-1 genome. Although protein levels were not determined in
this study, our finding of mRNA expression in sample No. 25 can be of importance,
since HBZ was shown to have bimodal function. The HBZ protein was first reported
to suppress transactivation of viral genes [25]. Satou et al., using the same set of
primers, reported HBZ mRNA expression in fresh ATL cells as well as in peripheral
blood mononuclear cells of asymptomatic carriers and showed a decrease of cell
proliferation when HBZ transcripts were suppressed [26]. The patient harboring the
HBZ-expressing bone marrow cells showed high leucocytes counts (more then
300,000 cells) as do samples without HBZ expression and the lowest (56%) number
of blastic cells in bone marrow, however, what this might mean is unclear.

The detection of MAD-1-like protein sequence in our samples using exon 2 and exon
3 primers was unexpected mainly because it happened in about half of the samples.
It might be possible that this product was formed due to primer homology within the
gene sequence in the 1.PCR, since the Nested primers were not able to reamplify the
product. Further studies using primers designed specifically for MAD1-like protein
sequence would be necessary to determine the real expression pattern in those
patient samples.
Since Shuh et al. [27] was using the same set of primers we used in the 1.PCR, for
detection of single spliced viral transcripts, the finding of PDXK sequence in sample
No. 2 was unexplainable. One possibility is primer homology to that sequence, albeit
due to the same biological material used in all patient samples, a PCR product would
than be expected in more samples.
Although predicted band sizes obtained in sample No. 26 and 27, corresponding to
p13II, p21Rex and TAX/REX mRNA transcripts, we tried to reassure sequence
specificity of those bands. Due to technical limitations, we were not able to determine
the splice sites, which are specific for each mRNA isoform. Sequencing showed
mixture of DNA sequences which was due to isolation problems of small PCR
products with similar length, out of the agarose gel. Splice site specific primers could
be used in an extended study to overcome this problem.
The viral protein TAX is known to be responsible for immortalization and cellular
transformation processes in infected cells. CD4+ cells were shown to be infected in
up to 99 % in peripheral blood of ATL patients [28], although associated to a low or
no expression level of TAX. This is caused by a variety of mechanisms, like genetic
or epigenetic modifications [29] which protect the infected cell from destruction
through CTLs [30]. In the present study there was not found any difference on the
percentage of CD4+ or CD34+ cell population between the positive and negative
proviral load bone marrow cells. It is known, however, that TAX can be secreted from
an infected cell population inducing a proinflammatory cytokine response on
surrounding cells [31], suggesting possible effects on CD34+ hematopoetic stem
cells. However, significant lower leucocytes counts were observed in patients
grouped as HTLV-1 positive. This could be interesting, since T-ALL is a malignancy
with reserved prognostic and usually shows leukocyte counts higher than
100.000/mm3 . If the lower count occurred due to immune response against virally

infected leucocytes, could explain one possibility, but must be investigated in future
studies.
Viral protein TAX was also shown to upregulate c-myc and c-fos expression, due to
interaction with NFκB/IκB and Serum Response Factor (SRF), respectively. The
mechanism which underlies the lower expression level of c-fos observed in our study
would need further investigation. However this observation seems to be interesting,
since c-fos is known to act as a transcriptional factor on a variety of cellular genes.
Overall, our study could clearly demonstrate early infiltration of BM in pediatric
patients with T-ALL and the transcription of viral genes in that cellular compartment,
so far not reported to our knowledge. Since the mechanism of long clinical latency of
ATL and the transformation process of infected cells, which is thought to involve a
variety of transformational steps [32], is not clearly understood, our data could
indicate an early onset of viral activity in life. If our observation of proviral load and
viral mRNAs expression is involved in leukemogenesis of T-ALL in children can not
be clearly stated yet, but should keep in consideration. Furthermore, if HTLV-1
causes pediatric T-ALL it would be through a different biological mechanism of viral
infectivity due to its short latency and therefore need further investigation.

Acknowledgments
We are thankful to Gilvan Mariano da Silva for the photographic assistance. Financial
support: Oswaldo Cruz Foundation. The authors declare that there is not any conflict
of interest.

References
[1] Redaelli A, Laskin BL, Stephens JM, Botteman MF, Pashos CL. A systematic
literatrure review of the clinical and epidemiology burden of acute lymphoblastic
leukaemia (ALL). Europ J Cancer Care 2005;14:53-62
[2] Gessain A, Barin F, Vernant JC et al.. Antibodies to human T-lymphotropic virus
type-1 in patients with tropical spastic paraparesis. Lancet. 1985; 2:407-410
[3] Kashiwagi K, Furusyo N, Nakashima H, Kubo N, et al. A decrease in mother-to-
child transmission of human T lymphotropic virus type 1 (HTLV-1) in Okinawa, Japan.
Am J Trop Hyg 2004;70(2):158-163
[4] Barmak K, Harhaj E, Grant C, Alefantis T, Wigdahl B. Human T cell leukemia virus
type I-induced diseas: pathways to cancer and neurodegeneration. Virology 2003;
308(1):1-12
[5] Grant C, Barmak K, Alefantis T, Yao J, Jacobson S, Wigdahl B. Human T Cell
Leukemia Virus Type 1 and Neurologic Disease: Events in Bone Marrow, Peripheral
Blood, and Central Nervous System during normal Immune Surveillance and
Neuroinflammation. J Cell Physiol. 2002;190:133-159
[6] Feuer G., Fraser J, Zack J, Lee F, Feuer R, Chen I. Human T-cell leukemai virus
infection of human hematopoietic progenitor cells: maintenance during differentiation
in vitro and in vivo. J Virol. 1996;70:4038-4044
[7] Levin MC, Krichavsky M, Fox RJ, Lehky T, Jacobson S, Fox C, Kleghorn F,
White J, Young N, Edwards RJ, Jack NE, Bartholomew C. Extensive latent retroviral
infection in bone marrow of patients with HTLV-1-associated neurologic disease.
Blood 1997;89:346-348
[8] Nagafuji K, Harada M, Teshima T, Eto T, Takamatsu Y, Okamura T, Murakawa M,
Akashi K, Niho Y. Hematopoetic Progenitor Cells from patients with adult T-cell
leukemia-lymphoma are not infected with human T-cell leukemia virus Type I. Blood
1993;82:2823-2828
[9] Proietti FA, Carneiro-Proietti AB, Catalan-Soares BC, Murphy EL. Global
epidemiology of HTLV-1 infection and associated diseases. Oncogene
2005;24:6058-6068
[10] Catalan-Soares B, Carneiro-Proietti F, Proietti F.A. Heterogeneous geographic
distribution of human T-cell lymphotropic viruses 1 and 2 (HTLV1/2): serological
screening prevalence rates in blood donors from large ruban areas in Brazil. Cad.
Saúde Pública 2005;21(3):926-931

[11] Farre L, de Oliveira Mde F, Primo J, Vandamme AM, Van Weyenbergh J,
Bittencourt AL. Early sequential development of infective dermatitis, human T cell
lymphotropic virus type 1-associated myelopathy, and adult T cell
leukemia/lymphoma. Clin Infect Dis 2008;46(3):440-2
[12] Pombo-de-Oliveira MS, Dobbin JA, Loureiro P, Borducchi D, Maia RC,
Fernandes MA, Cavalcanti GB Jr, Takemoto S, Franchini G. Genetic mutation and
early onset of T-cell leukemia in pediatric patients infected at birth with HTLV-I. Leuk
Res. 2002;26:155-61
[13] Lin BT-Y, Musset M, Szekely AM et al. Human T-cell lymphotropic virus-I
positive T-cell leukemia/lymphoma in a child. Report of a case and review of the
literature. Arch Pathol Lab Med 1997;121:1282-6
[14] Broniscer A, Ribeiro RC, Srinivas RV, Behem FG, Head DR, Raimondi SC,
Mandrell B, Gill P. An adolescent with HTLV-1 associated adult T-cell leukemia
treated with interferon alpha and zidovudine. Leukemia 1996;10:1244-54
[15] Blank A, Yamaguchi K, Blank M et al. Six Columbian patients with adult T-cell
leukemia/lymphoma. Leuk Lymphoma 1993;9:407-12
[16] Fort JA, Graham-Pole J, Mottshaw G. Adult-type T-cell lymphoma in an
adolescent with human T-lymphotropic virus type I seropositivity. Med Pediatr Oncol
1989;17:236-8
[17] Vilmer E, le Deist F, Fisher A, Griscelli C, Nezelof C, Prost Y, Prieur M.
Smouldering T lymphoma related to HTLV-1 in a Sicilian child. Lancet 1985;ii:1302
[18] Kawa K, Nishiuchi R, Okamura T, Igarashi H. Eradication of human T-
lymphotropic virus type 1 by allogeneic bone-marrow transplantation. Lancet
1998;352:1034-1035
[19] Furukawa Y, Kubota R, Tara M, et al. Existence of escape mutant in HTLV-1 tax
during the development of adult T-cell leukemia. Blood. 2001;97:987-993
[20] Dourado I, Alcantara LC, Barreto ML, da Gloria Teixeira M, Galvão-Castro B.
HTLV-1 in the general population of Salvador, Brazil: a city with African ethnic and
sociodemographic characteristics. J Acquir Immune Defic Syndr. 2003;34(5):527-31.
[21] Yao J, Wigdahl B. Human T cell lymphotropic virus type 1 genomic expression
and impact on intracellular signaling pathways during neurodgenerative disease and
leukemia. Frontiers in Bioscience 2000;5:d138-168

[22] Hiramatsu K, Yoshikura H. Frequent partial deletion of human T-cell leukemia
virus type 1 proviruses in experimental transmission: Pattern and possible
implication. J Virol. 1986;58:508-512
[23] Tamiya S, Matsuoka M, Etoh K, Watanabe T, Kamihira S, Yamaguchi K,
Takatsuki K. Two types of defective human T-lymphotoropic virus type 1 provirus in
adult T-cell leukemia. Blood 1996;88:3065-3073
[24] Matsuoka M, Jeang K-T. Human T-cell leukemia virus type 1 (HTLV-1) infectivity
and cellular transformation. Nature Review Cancer 2007;7:270-280
[25] Gaudray G, Gachon F, Basbous J et al. The complementary strand of the human
T-cell leukemia virus type 1 RNA genome encodes a bZIP transcription factor that
down-regulates viral transcription. J Virol. 2002;76:12813-12822
[26] Satou Y, Yasunaga J, Yoshida M, et al. HTLV-1 basic leucine zipper factor gene
mRNA supports proliferation of adult T cell leukemia cells. PNAS. 2006;103:720-725
[27] Shu M, Hill S.A, Derse D. Defective and Wild-Type Human T-cell Leukemia Virus
type I Proviruses: Characterization of Gene Products and trans-Interactions between
Proviruses. Virology 1999;262:442-451
[28] Richardson JH, Edwards AJ, Cruickshank JK, Rudge P, Dalgleish AG. In vivo
cellular tropism of human T-cell leukemai virus type I. J Virol. 1990;64(11):5682-5687
[29] Takeda S, Maeda M, Morikawa S, et al. Genetic and epigenetic inactivation of
tax gene in adult T-cell leukemia cells. Int J Cancer. 2004;109:559-567
[30] Hanon E, Hall S, Taylor G.P, Saito M, Davis R, Tanaka Y, Usuku K, Osame M,
Weber J.N, Bangham C.R.M. Abundant Tax protein expression in CD4+ T cells
infected with human T-cell lymphotropic virus type 1 (HTLV-1) is prevented by
cytotoxic T lymphocytes. Blood 2000;95:1386-1392
[31] Aono H, Fujisawa K et al. Extracellular Human T cell Leukemia virus type 1 Tax
protein stimulates the proliferation of Human synovial cells. Arth & Rheum
1998;41:1995-2003
[32] Giam CZ, Jeang KT. HTLV-1 Tax and adult T-cell leukemia Front. Biosci
2007;12:1496-1507

SEGUNDO ARTIGO:
baseado nas normas da
Revista Brasileira de Hemato logia e Hemoterapia
Conteúdo equal da introdução da tese

Leucemia de Células T
T-Cell Leukemia
Autores
Heike Brand1
João Guilherme Bezerra Alves1
Francisco de Paula Ramos Pedrosa1
Norma Lucena-Silva1,2
1Serviço de Oncologia Pediátrica/IMIP, Rua dos Coelhos, n.300, Quinto andar, Boa
Vista, cep:50070-550, Recife, Pernambuco, Brasil.
2Centro de Pesquisas Aggeu Magalhães/Fiocruz, Av. Moraes Rego, s/n, campus da
UFPE, Cidade Universitária, cep:50670-420, Recife, Pernambuco, Brasil.
Autor de Correspondência: Dr. Norma Lucena-Silva, Serviço de Oncologia
Pediátrica/IMIP; Rua dos Coelhos, n.300, Quinto andar, Boa Vista, cep:50070-550,
Recife, Pernambuco, Brazil.
Tel.: 55 81 21224764.
e-mail: [email protected]

ANEXOS E APÊNDICES

TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO
Nome da Pesquisa: Banco de Tumores do Recife
Coordenadores responsáveis:
Norma Lucena Cavalcanti Licínio da Silva (Fundação Oswaldo Cruz / Centro de Pesquisas Aggeu Magalhães)
Francisco de Paula Ramos Pedrosa (Serviço de Oncologia Pediátrica do Instituto Materno Infantil Professor Fernando Figueira)
Prezado Sr(a).
O câncer é uma doença onde as células crescem sem controle podendo ocorrer em diferentes locais no nosso corpo. Alguns tipos de câncer já têm cura, outros são possíveis prevenir. Precisamos conhecer mais sobre esta doença, saber o que causa a doença para podermos nos proteger e melhor tratar. Para isso estamos: 1) criando um arquivo eletrônico (no computador) com informações importantes sobre a doença e fatores que possivelmente podem ter ajudado ao tumor desenvolver, além dos resultados de laboratório, o tratamento e evolução da doença. As informações obtidas serão analisadas com o objetivo de encontrar fatores que predispõem a tipos específicos de câncer. 2) Pretendemos coletar e armazenar amostras biológicas, incluindo sangue, secreção, produtos de excreção, exsudatos e fragmento do tumor, para podermos estudar quando possível as alterações genéticas e bioquímicas dos tumores para compreendermos a origem e evolução deste crescimento alterado. 3) Pretendemos também coletar amostras de sangue periférico para investigar o risco (a predisposição genética) do indivíduo ao câncer e estudo das alterações bioquímicas e imunes do tumor em busca de marcadores moleculares de diagnóstico e prognóstico.
As informações clínico-epidemiológicas obtidas na entrevista, ou do seu prontuário eletrônico terão caráter confidencial. O fragmento do tumor será coletado durante biópsia e/ou cirurgia quando houver indicação médica restrita, a punção venosa será feita por profissional experiente, com seringas e agulhas descartáveis. Estudos técnico-científicos, utilizando o fragmento de tumor e sangue armazenados no banco de tumores, só serão realizados após a aprovação do estudo no Comitê de Ética em Pesquisa.
Caso todas as dúvidas tenham sido esclarecidas e tendo entendido o conteúdo deste consentimento informado, declaro que concordo de livre e espontânea vontade em participar neste estudo.
________________________________ Data: ___/___/___
Assinatura do paciente
________________________________ Data: ___/___/___
Testemunha
________________________________ Data: ___/___/___
Assinatura de representante
da equipe médica responsável

--------------------------- Mensagem Original ---------------------------- Assunto: Parecer CEP-IMIP Proj. 725 De: [email protected] Data: Qua, Maio 21, 2008 8:00 am Para: [email protected] -------------------------------------------------------------------------- De: Comitê de Ética em Pesquisa em Seres Humanos do IMIP Para: Dra. Norma Lucena Cavalcanti Licinio da Silva Nº de Entrada no CEP-IMIP: 725 Ref: Projeto intitulado "Freqüência do vírus linfotrópico de célula T humana-1 (HTLV-1) em medula óssea de pacientes infanto-juvenis diagnosticados com leucemia e linfoma de célula T no período de 2003-2007, no serviço de oncologia do Instituto Materno Infantil Prof. Fernando Figueira" Prezado(a) Pesquisador(a), O Comitê de Ética em Pesquisa (CEP) do IMIP reunido em 08/05/2008 decidiu por aprovar com recomendação o protocolo de pesquisa acima mencionado. O parecer sobre o protocolo faz a seguinte solicitação: · Apresentar um Termo de Consentimento Livre e Esclarecido (TCLE) para ser utilizado com os participantes da pesquisa de acordo com a Resolução 196/96 e 347/05 CNS. (Ver parecer em anexo) O pesquisador deverá atender as solicitações através de carta assinada, juntamente com as devidas alterações. OBS: A declaração de aprovação só será liberada depois de atendidas as solicitações. Atenciosamente, Comitê de Ética em Pesquisa em Seres Humanos do Instituto Materno Infantil Professor Fernando Figueira Rua dos Coelhos, 300 Boa Vista - Recife-PE - Brasil Cep: 50070-550 Fone/Fax: (81) 2122-4756

From: [email protected]
Subject: Artigo Submetido SGP/ RBHH
Date: Thu, 2 Oct 2008 16:18:08 -0300
Revista Brasileira de Hematologia e Hemoterapia
Av. Nossa Sra. de Copacabana, 1.059 sala 1.201 -
Copacabana
Rio de Janeiro - RJ - Brazil
CEP 22060-001
Tel/Fax: (21) 2521-6905
email: [email protected]
Rio de Janeiro, quinta-feira, 2 de outubro de 2008
Ilmo(a) Sr.(a)
Prof(a), Dr(a) Norma Lucena Cavalcanti Licinio da Silva
Referente ao código de fluxo: 263
Classificação: Artigo de Revisão
Informamos que recebemos o manuscrito Leucemia de Células T será enviado para apreciação dos
revisores para possível publicação/participação na(o) Revista Brasileira de Hematologia e
Hemoterapia. Por favor, para qualquer comunicação futura sobre o referido manuscrito cite o número
de referência apresentado acima.
Obrigado por submeter seu trabalho a(o) Revista Brasileira de Hematologia e Hemoterapia.
Atenciosamente,
Milton Artur Ruiz
Editor


















