Instability in human forearm movements studied with feed-back ...
22
Journal of Physiology (1988), 402, pp. 421-442 421 Wfith 11 text-figures Printed in Great Britain INSTABILITY IN HUMAN FOREARM MOVEMENTS STUDIED WITH FEED-BACK-CONTROLLED MUSCLE VIBRATION BY A. PROCHAZKA* AND P. ST. J. TREND From the Department of Physiology, St Thomas's Hospital Medical School, London SEI 7EH (Received 28 May 1987) SUMMARY 1. Frequency-modulated vibration was applied to the elbow flexor and extensor tendons to produce reflex movements of the forearm in normal subjects. The modulating (command) signal caused equal and opposite deviations from the 40 Hz carrier frequency so that when flexor vibratory frequency increased, extensor frequency decreased, and vice versa. 2. It is argued that the movements resulted largely from the reflex action of muscle spindle primary afferents whose firing frequency had been 'taken over' and modulated by the vibration. 3. Bode plots relating forearm movements to command signal (modulating) frequency showed the transfer function of the I a afferent-CNS-muscle-load system to have a low-pass filter characteristic. The phase lag of movement on command increased progressively with command signal frequency, exceeding 180 deg at 3-4 Hz. 4. The transduced forearm movements were fed back to provide the command signal to the vibrators (and thus indirectly to the spindle afferents) via a filter mimicking the dynamic responsiveness of muscle spindle primary endings. Our aim was to 'break into' the reflex arc, and re-route it so that we could artificially vary the gain without significantly altering the dynamics of the pathway. 5. Nearly all subjects developed forearm oscillations (tremor) when the gain exceeded a threshold value. Subjects varied widely in their threshold, though for a given subject the threshold remained fairly constant from day to day. The results suggest that reflexly active individuals may not have a large safety margin with respect to forearm instability. 6. The frequency range of the oscillations observed in seven subjects was 3-8 Hz. The frequencies depended upon the level of flexor-extensor co-contraction, and increased from 3 to 5 Hz at 10 % co-contraction to 5-8 Hz at 100 % co-contraction. An analysis of the mechanical impedance of the arm provided estimates of tremor frequencies consistent with these results. 7. These unexpectedly low tremor frequencies led us to propose that it may be erroneous to expect stretch reflexes to contribute to forearm tremor in the range * Present address: Department of Physiology, University of Alberta, School of Medicine, Edmonton, Alberta T6G 2E1, Canada.
Transcript of Instability in human forearm movements studied with feed-back ...

Journal of Physiology (1988), 402, pp. 421-442 421Wfith 11 text-figuresPrinted in Great Britain
INSTABILITY IN HUMAN FOREARM MOVEMENTS STUDIED WITHFEED-BACK-CONTROLLED MUSCLE VIBRATION
BY A. PROCHAZKA* AND P. ST. J. TRENDFrom the Department of Physiology, St Thomas's Hospital Medical School,
London SEI 7EH
(Received 28 May 1987)
SUMMARY
1. Frequency-modulated vibration was applied to the elbow flexor and extensortendons to produce reflex movements of the forearm in normal subjects. Themodulating (command) signal caused equal and opposite deviations from the 40 Hzcarrier frequency so that when flexor vibratory frequency increased, extensorfrequency decreased, and vice versa.
2. It is argued that the movements resulted largely from the reflex action ofmuscle spindle primary afferents whose firing frequency had been 'taken over' andmodulated by the vibration.
3. Bode plots relating forearm movements to command signal (modulating)frequency showed the transfer function of the I a afferent-CNS-muscle-load systemto have a low-pass filter characteristic. The phase lag of movement on commandincreased progressively with command signal frequency, exceeding 180 deg at3-4 Hz.
4. The transduced forearm movements were fed back to provide the commandsignal to the vibrators (and thus indirectly to the spindle afferents) via a filtermimicking the dynamic responsiveness of muscle spindle primary endings. Our aimwas to 'break into' the reflex arc, and re-route it so that we could artificially varythe gain without significantly altering the dynamics of the pathway.
5. Nearly all subjects developed forearm oscillations (tremor) when the gainexceeded a threshold value. Subjects varied widely in their threshold, though for agiven subject the threshold remained fairly constant from day to day. The resultssuggest that reflexly active individuals may not have a large safety margin withrespect to forearm instability.
6. The frequency range of the oscillations observed in seven subjects was 3-8 Hz.The frequencies depended upon the level of flexor-extensor co-contraction, andincreased from 3 to 5 Hz at 10% co-contraction to 5-8 Hz at 100% co-contraction.An analysis of the mechanical impedance of the arm provided estimates of tremorfrequencies consistent with these results.
7. These unexpectedly low tremor frequencies led us to propose that it may beerroneous to expect stretch reflexes to contribute to forearm tremor in the range
* Present address: Department of Physiology, University of Alberta, School of Medicine,Edmonton, Alberta T6G 2E1, Canada.

A. PROCHAZKA AND P. ST. J. TREND
8-12 Hz (e.g. physiological and 'enhanced' physiological tremors). Rather, theircontributions should be sought in the range 3-8 Hz (e.g. pathological tremors suchas those of Parkinson's and cerebellar disease).
INTRODUCTION
In the control of limb movements, the CNS utilizes information continuously fedback from various groups of mechanoreceptors. A common feature of any feed-backloop is that if the (open) loop gain is increased beyond a certain threshold level, thesystem becomes unstable and sinusoidal oscillations tend to develop. Yet a high loopgain is often desirable, because it is associated with improvements in linearity,frequency response and resistance to external perturbations (Stein & Oguztoreli,1976). There is in fact no general agreement on the extent to which the stretch reflexarc in humans contributes to the control of movement. On the one hand, it has beenargued that the gain around the loop is low (for review see Matthews, 1972) and thatlimbs can be moved accurately even after deafferentation (Bizzi, Dev, Morasso &Polit, 1978; Rothwell, Traub, Day, Obeso, Thomas & Marsden, 1982). On the otherhand, it has been suggested that tremors in normal individuals ('physiological' and'enhanced physiological' tremor) may reflect limited instability in the spinal stretchreflex arc (Joyce & Rack, 1974). This would imply that the loop gain in such caseswas as high as it could safely be.
Involuntary oscillations of the extremities occur in association with various motorcontrol disorders. However, whereas physiological and enhanced physiologicaltremors occur in the frequency range 8-12 Hz (Marsden, 1984), the pathologicaltremors of Parkinson's disease, cerebellar disease, spastic clonus and essential tremoroccur in the 2-8 Hz range. For various reasons, it has become the prevailing viewthat tremors at these lower frequencies are usually generated largely from within theCNS (but cf. Rack & Ross, 1986), and that for stretch reflexes to support them, orindeed be solely responsible for them, they would have to be mediated by long-loop,long-latency CNS pathways (Dimitrijevic, Sherwood & Nathan, 1978; Freund,Hefter, Homberg & Reiners, 1984; Llina's, 1984). Joyce, Rack & Ross (1974) arguedthat in oscillatory movements about the elbow involving powerful co-contraction ofthe flexors and extensors, spinal stretch reflexes would be timed so as to assist10-12 Hz tremor, but to resist tremor at 4-6 Hz. Similarly, on the basis of atheoretical analysis of the stretch reflex arc in man, Stein & Oguztbreli (1976)suggested that overactive reflexes of spinal latency (20-50 ms) would give rise totremor in the 8-12 Hz range, whereas long-loop reflexes (50-120 ms) would result in4-6 Hz tremor.
In the present experiments, frequency-modulated (FM) muscle vibration was usedto 'take over control' of muscle spindle endings in the elbow flexor and extensormuscles. Sinusoidal command signals thus applied to the spindles were found reflexlyto evoke sinusoidal forearm movements. The movements showed phase lags on thecommand signals which increased with increasing frequency, reaching 180 deg in therange 2-5-4 Hz at low levels of background co-contraction. The frequency (fi80) atwhich there is a 180 deg phase lag around a feed-back loop has a special significance,in that when the loop is closed and the gain is sufficiently increased, instability
422

VIBRATION-EVOKED TREMOR
results, and oscillations develop at this frequency. In closed-loop trials, in whichforearm movements were fed back to control vibration frequency, oscillations didindeed develop in this frequency range. This suggests that low-frequency tremorabout the elbow could well be assisted, rather than resisted, by stretch reflexes.Subjects varied widely in their threshold for instability. As the threshold was simplydetermined by a single potentiometric adjustment, and was read off as an absolutevalue, namely frequency change per degree of elbow displacement, this techniquemay be useful in quantifying central reflex excitability in different individuals.
METHODS
Seven normal adult subjects in the age range 21-39 were studied in detail, two on severaldifferent occasions, but over fifty normal subjects were examined in all. A typical experiment lasted3 h. The subject was seated comfortably and the right shoulder was supported against a verticalbar (Fig. 1). The right upper limb was abducted 90 deg at the shoulder. The forearm was firmlystrapped to a horizontal arm rest in a position mid-way between pronation and supination. thesubject holding a vertical rod attached to the arm rest. The arm rest was pivoted at the elbow, thusallowing pure flexion and extension movements about the joint. The moment of inertia of the armrest was calculated from its physical dimensions to be 17-3 g mi2, and that of the vertical rod rangedfrom 4-7 to 10-3 g M2, depending upon the subject's arm length. The arm rest was further supportedat the distal end by a 1-5 m length of stout Nylon cord suspended from an overhanging part of thechair frame. Two heavy-duty vibrators (Ling Dynamic Systems Ltd, Type 101) were suspendedabout the arm and were held in light contact with biceps and triceps muscles, approximately overthe musculo-tendinous junctions, by a compliant coupling. Angular displacement of the elbow wasmeasured using a sensitive cantilever transducer (Kulite UHP 5000-060 semiconductor straingauge bonded to a 4 mm beryllium copper strip), one end of which was attached to the moving armrest, the other end being deflected by a lubricated Teflon post fixed to the vertical support. Theelectromyographic activity of biceps and triceps muscles was recorded using surface electrodes, wasamplified and then full-wave rectified, and in some cases smoothed (40 dB/decade, Butterworthlow-pass, 50 Hz).The subject was asked to co-contract biceps and triceps muscles with a force corresponding to
a specified percentage (usually 10%) of the maximum voluntary triceps EMG activity. To this end,the subject was provided with an oscilloscope display in which rectified, smoothed triceps EMGactivity provided the vertical deflection of a point. Forearm displacement provided the point'shorizontal deflection. The subject's task was to keep the point centred within a small target zonewhich corresponded to a predetermined tonic level of EMG activity and an elbow angle ofapproximately 90 deg. Thus if for example the subject flexed too far, the point moved to the leftof the target, and if there was too much co-contraction, the point moved above the target.The responses of the forearm were examined both in open- and in closed-loop modes as follows.
In open-loop mode, a sinusoidal test signal ('external input' in Fig. 1) and its inverse were used todrive reciprocally two voltage-controlled oscillators whose output in turn frequency modulated thetwo vibrators about a centre frequency of 40 Hz. Peak-to-peak movement of the vibrator headswas approximately 1 mm. In closed-loop mode the forearm displacement signal was fed backthrough an electronic filter which closely mimicked the transducing properties of cat muscle spindleIa afferents in the linear range (Poppele & Bowman, 1970). The transfer function we used is shownin the legend of Fig. 1, and in Fig. 2 the measured amplitude and phase responses of the filter areshown as dots superimposed on the experimental data of Poppele & Bowman (1970) and Matthews& Stein (1969). It should be noted that Poppele & Kennedy (1974) found that biopsied humanspindle primary endings studied in vitro had virtually identical frequency-response curves to thoseof the cat. A simple potentiometric adjustment ('Gain+ offset' in Fig. 1) allowed the gain of thespindle model to be altered without altering the dynamics of the transfer function. The filteredsignal and its inverse were used to drive reciprocally the two voltage-controlled oscillators whichin turn drove the vibrators. In this way forearm extension simultaneously caused biceps vibratorfrequency to increase and that of triceps to decrease, and vice versa during flexion. Instabilityusually occurred at some point as the loop gain was progressively increased.
423
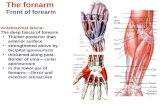
A. PROCHAZKA AND P. ST. J. TREND
Fig. 1. The experimental arrangement. The subject's elbow flexors and extensors were
independently vibrated at frequencies controlled by two voltage-controlled oscillators(VCO). The VCOs in turn received their inputs (one of which was inverted) either from an
external signal source or from a filtered version of the arm-movement signal. The filter(Op. amp. array) was designed to mimic the transducing properties of muscle spindleprimary endings. The transfer function used was that of Poppele & Bowman (1970):
K(s+044) (8+11-3) (8+44)
(s8+0816)
With this arrangement, the afferent limb of the stretch reflex arc was in effect'externalized', as the spindle endings were made to respond to elbow displacement in a
manner determined by the filter and gain settings.
co
0c
20
15
10
5
o
-5
-10
-15
0.2 0-4 1 2 4Modulation frequency (Hz)
_
135
90AM
0
10
10 20
Fig. 2. Spindle model frequency-response curves. Gain (dB) and phase (deg) of spindlemodel (@) as a function of modulation frequency (Hz), compared with data of Poppele& Bowman (1970) (continuous line) and those of Matthews & Stein (1969) (interruptedline).
424
Adbm----Nm-o imi
.adF*11.4.4.= -
-
I I

VIBRATION-EVOKED TREMOR
Recordings of forearm displacement, instantaneous vibrator frequencies and EMG activitieswere sampled and displayed with the use of a BBC microcomputer linked to a Unilab interface(O'Brien, Prochazka & Vincent, 1985). Single trials or averages of several trials were printed outwith a dot-matrix printer, or stored on magnetic disc for later analysis. In closed-loop experimentsresults were analysed at the threshold gain at which the arm first showed established tremor. Inother experiments results were obtained at higher levels of background co-contraction. Theresulting tremor was spectrally analysed with the use of a laboratory interface (CambridgeElectronic Design 1401) operating a Fast Fourier Transform algorithm. Bode plots wereconstructed showing the amplitude and phase of the displacement in response to the vibratorfrequency.
Mechanical impedance analysi8In one set of trials, carried out in the laboratory of Drs P. M. H. Rack and H. F. Ross in
Birmingham, frequency analyses of the mechanical impedance of the forearm were performed infour normal subjects. The object of these trials was to see the extent to which vibratorystimulation, as outlined above, altered the resistance of the elbow joint to imposed sinusoidaldisplacements. The frequency range over which negative viscosities were observed was comparedwith the frequencies of tremor obtained in separate trials in which the forearm was free to move.The technique and method of analysis have been described elsewhere (Joyce et al. 1974; Brown,Rack & Ross, 1982).
RESULTS
Closed-loop trialsThe original aim of our study was to mimic hyperreflexia by artificially increasing
the gain, without altering the dynamics, of the transfer function of the stretch reflexarc. Indeed our simplest and most reproducible finding was that with the closed-looparrangement (described in Methods), as the loop gain was progressively increased, atsome point the forearm developed sustained and fairly stable oscillations. In only onesubject out of more than fifty tested did we find it impossible to evoke tremor in thisway.An example of such tremor is shown in Fig. 3. It can be seen that each time the
forearm moved into flexion the triceps vibrator frequency increased, reaching a peakapproximately 90 deg ahead of peak flexion. The frequency of tremor generated inthis way was consistently in the range 3-5 Hz for background contractions of 10%maximum voluntary triceps EMG activity. In Fig. 3 the frequency was just under4 Hz. We found that each subject had a fairly fixed gain threshold at whichestablished tremor first occurred, and that this was fairly constant from hour to hourand from day to day. However, such thresholds varied widely between subjects. Thesubjects' ages ranged from 19 to 58, and qualitatively there was no apparentrelationship between age and the threshold gain for instability, though this was notinvestigated systematically.
Expressed as the depth of modulation of vibrator frequency per percentage musclelength modulation, thresholds ranged from 1500 to 9000 Hz/% rest length (assuming5 deg elbow rotation corresponds to 1 % rest length: see Discussion). It is interestingto note that cat soleus I a afferents respond to sinusoidal stretching at 4 Hz with asensitivity of approximately 1000 s-1 (% rest length)-1 (inferred from Goodwin,Hulliger & Matthews (1975), assuming a soleus rest length of 100 mm origin toinsertion). This comparison will be taken up again in the Discussion.
425

A. PROCHAZKA AND P. ST. J. TREND
Extension
eForearm
1 lxideg 0 \/movementF lexion I
Biceps EMG
NU Biceps100 * , ** * . .4 . % * /'. vibrator frequency
1 00F 0pe *,:* , , -" ** Tricepsvibrator frequency
Triceps EMG
0 0 5 1-0 1-5Time (s)
Fig. 3. Oscillation evoked by feed-back-controlled vibration (closed-loop trial). The toptrace shows forearm displacement (extension upwards). In the centre of the Figure theinstantaneous frequency of the biceps vibrator (increasing above the line) and the tricepsvibrator (increasing below the line) may be seen. The signal modulating vibratoryfrequency was the transduced displacement, filtered according to the transfer function ofFig. 2, to mimic transmission via spindle primary endings. The rectified, smoothed EMGactivity of biceps and triceps muscles is also shown. Note that the frequency of tremor wasjust under 4 Hz (10% maximal co-contraction). Subject, A. P.
loI
80 0
0.
E 60 -/\fb
x m~~~~~()40 -
20 -
2 4 6 8Frequency (Hz)
Fig. 4. The effect of flexor-extensor co-contraction on oscillatory frequency in six subjects(closed-loop trials). As the level of co-contraction increased, tremor frequencies alsoincreased, reaching 6-8 Hz at maximal voluntary co-contraction. Each frequencymeasurement was made on an averaged record of sixteen or thirty-two phase-alignedcycles.
426

VIBRATION-EVOKED TREMOR
Increased background co-contractionIt was noticed that the frequency of tremor increased when subjects tensed their
forearm muscles so as to exceed the target of a 10% maximum triceps EMG level.This effect was studied systematically in six subjects. Figure 4 shows that in eachsubject there was a clear increase in the frequency of tremor with increasing levelsof co-contraction, as quantified by the level of triceps EMG. At maximal levels of co-contraction, tremor frequencies ranged from 5-5 to 7-7 Hz.From earlier work (Joyce et al. 1974) we had expected that the frequency of tremor
generated by CNS reflexes would be in the range 8-12 Hz. We were surprisedconsistently to observe frequencies in the range 3-5 Hz at 10% co-contraction (Fig.4), and in order to understand why the frequency of oscillation was so low, we studiedthe open-loop response of the forearm to sinusoidally frequency-modulated vibration.
Open-loop experimentsIf we assume for the sake of argument that each vibratory cycle evokes a single
action potential in each of the vibrated muscle's spindle Ia afferents, any drivingsignal which modulated the frequency of muscle vibration (and thus Ia firing) couldbe viewed as causing movement after first undergoing (a) a conduction delay in theafferent limb of the reflex arc, (b) filtering by, and further delays in, the CNSpathways mediating I a activity, (c) efferent conduction delay and (d) filtering by themuscle-load system. The extent to which the above assumption is in error is takenup in detail in the Discussion.
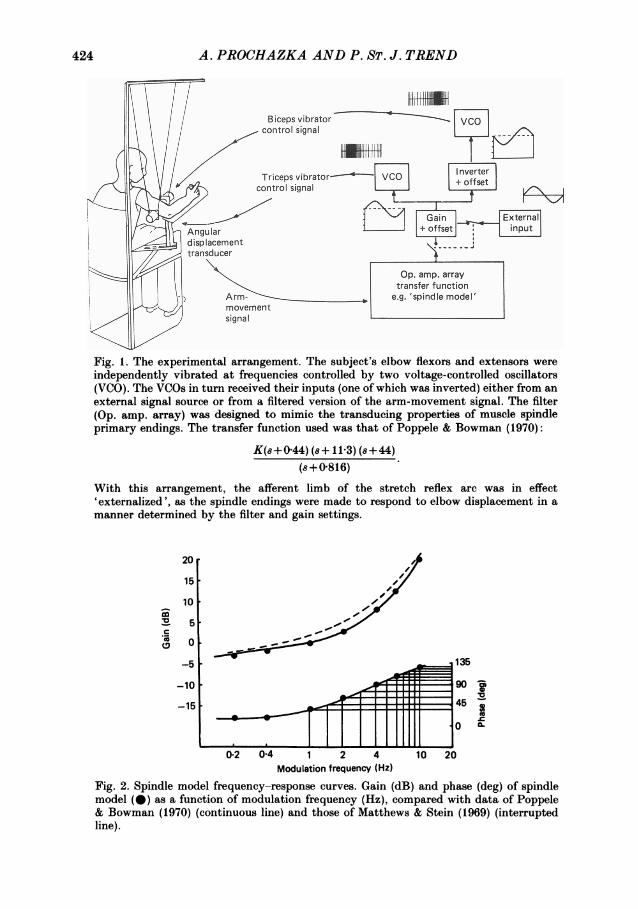
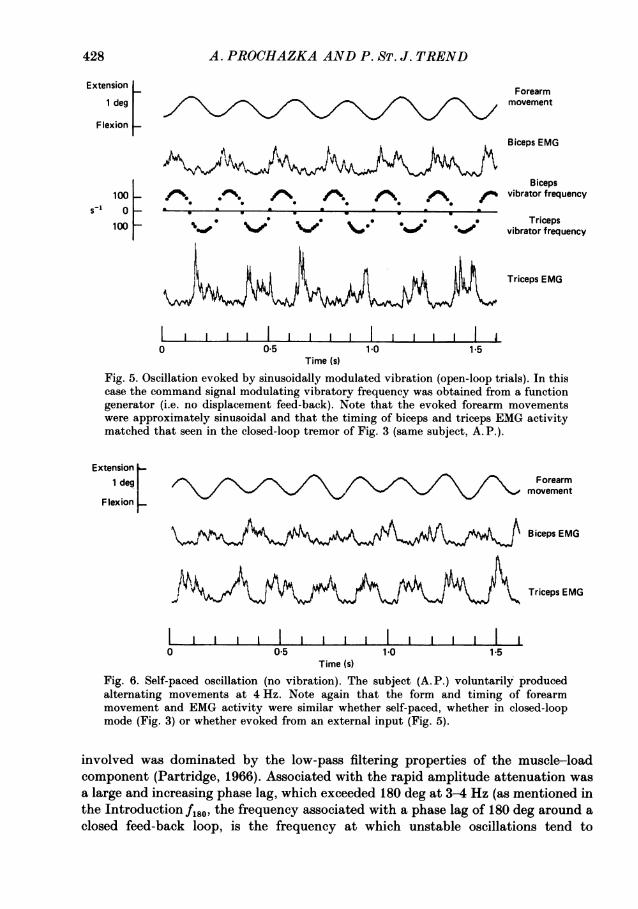
Figure 5 shows a sequence of oscillatory forearm movements evoked by vibrationwhose frequency was modulated by a 4 Hz sinusoid. The subjects' task was again tomaintain a background co-contraction of biceps and triceps equivalent to 10%maximum triceps EMG. The frequencies of the biceps and triceps vibrators werecontrolled reciprocally, as in the closed-loop experiments above, but the drivingsignal was obtained from an external function generator, and movements of the armhad no influence on the vibration. The movements were approximately sinusoidal,and the timing of the biceps and triceps EMG bursts is seen to be closely comparableto that in the closed-loop oscillations of Fig. 3, and of self-paced oscillatorymovements performed in the absence of any external inputs as shown in Fig. 6.
Initially we attempted to obtain frequency-response curves (Bode plots) of theopen-loop system by linearly increasing with time the frequency of the driving signalin the range 1-10 Hz, and plotting the amplitudes of the resultant oscillatorymovements. This technique is commonly employed in electronic harmonic analysers(and indeed in the mechanical impedance analysis described in the Appendix).However, the responses to the driving signals tended to be superimposed on slow,random shifts of forearm position, which made amplitude measurements of singlecycles unreliable. This difficulty was overcome by averaging the displacementresponses to thirty-two or sixty-four cycles at set frequencies of the driving signal(O'Brien et al. 1985), presented in a series of randomized order.
Figure 7 shows some typical averaged responses at driving signal frequencies of 1,2, 4 and 6 Hz. The responses at 4 and 6 Hz were only a fraction of those at 1 and2 Hz, indicating that the transfer function of the part of the neuromuscular system
427
A. PROCHAZKA AND P. ST. J. TREND
Extensionde
Forearm1 deg _ V movement
Flexion
Biceps EMG
Biceps100 /*0 *_. /% f\ \ f_ vibrator frequency
5-1 0
0 ~~~~~~~0Triceps100 - ._ 1 vibrator frequency
XAAAVGA2\M4& _Triceps EMG
I i Ii Ii I I Ii I I I0 0 5 1-0 1-5
Time (s)Fig. 5. Oscillation evoked by sinusoidally modulated vibration (open-loop trials). In thiscase the command signal modulating vibratory frequency was obtained from a functiongenerator (i.e. no displacement feed-back). Note that the evoked forearm movementswere approximately sinusoidal and that the timing of biceps and triceps EMG activitymatched that seen in the closed-loop tremor of Fig. 3 (same subject, A. P.).
Extension _1 deg Forearm
movementF lexion _
\AJY'vAR/\~p~4&_Biceps EMG
Triceps EMG
0 05 10 1-5Time (s)
Fig. 6. Self-paced oscillation (no vibration). The subject (A. P.) voluntarily producedalternating movements at 4 Hz. Note again that the form and timing of forearmmovement and EMG activity were similar whether self-paced, whether in closed-loopmode (Fig. 3) or whether evoked from an external input (Fig. 5).
involved was dominated by the low-pass filtering properties of the muscle-loadcomponent (Partridge, 1966). Associated with the rapid amplitude attenuation wasa large and increasing phase lag, which exceeded 180 deg at 3-4 Hz (as mentioned inthe IntroductionfI86, the frequency associated with a phase lag of 180 deg around aclosed feed-back loop, is the frequency at which unstable oscillations tend to
428

VIBRATION-EVOKED TREMOR
develop). At frequencies of 0 5 and 1 Hz it was found that although the movementresponses were essentially sinusoidal, they were contaminated to a small extent bywhat appeared to be harmonics at 2-2-5 Hz (e.g. Fig. 7, 1 Hz). In these cases, apopulation of sinusoids of different amplitudes photocopied onto a transparent sheetwere used as overlays to estimate by eye the amplitude of the fundamentalcomponent of the response.
Command signal87 deg'~~~~~~~~~ dg
Flexion
Forearm angular Tdisplacement /j/1deg
1 Hz 2 Hz Extension
is 0-5s
Command signal
277 deg i 329 deg Flexion
Forearm angular I Idisplacement 1 deg
4 Hz 6 Hz Extension
025s 0-25sFig. 7. Forearm displacement at four different modulating frequencies (open-loop trials).Each pair of traces shows averaged command signals (top) and resultant reflex-evokedforearm movements (bottom) at modulating frequencies of 1, 2, 4 and 6 Hz. Responseamplitudes decreased and phase lag increased with increasing frequency of the commandsignal. Already at 4 Hz the phase lag greatly exceeded 180 deg.
Figure 8 shows open-loop frequency-response curves (Bode plots) for two subjects.Small resonances, seen as peaks in the amplitude plots, were observed in five of theseven subjects for which these measurements were made. The frequencies at whichthe resonances occurred varied from subject to subject over the range 2-5-4-5 Hz.This frequency range is similar to that previously identified as encompassing thenatural frequency about the elbow due purely to the mechanical properties of theforearm and its supporting tissues in the relaxed state (Fox & Randall, 1970). It istherefore worth emphasizing that in these trials the subjects' task was to maintainan isometric co-contraction of biceps and triceps such that the rectified, low-passfiltered triceps EMG activity was kept at 10% of its maximum level.
Figure 9 shows the superimposed Bode plots for all seven subjects. fi80 ranged from2-4 to 3-8 Hz, and at 10 Hz the phase lag was a full 360 deg.
429

A. PROCHAZKA AND P. ST. J. TREND
0
m -10
-20 i/E O~~\_\\
-30
1 2 3 4 5 6 7 8 100 Modulation frequency (Hz)40
80,
120
X 160
200 \
9l 240
0- 280
320
360
Fig. 8. Amplitude and phase plots of arm movement vs. vibrator modulation frequency(open-loop trials). Only two subjects' results are shown for the sake of clarity. Smallresonant peaks are recognizable in the amplitude plots, between 2-5 and 4-5 Hz. Subjectswere asked to maintain 10% maximum voluntary co-contractions of elbow flexors andextensors.
0
-o0
-20*
< -30
0 1 2 3 4 5 6 7 8 9 10111240 -Modulation frequency (Hz)
120
280
320
360 _
400
440Fig. 9. Similar plots to those in Fig. 8 but here results for seven subjects are superimposed.The cross-over points at which 180 deg phase lags occurred ranged from 2-4 to 3-8 Hz.
430

VIBRATION-EVOKED TREMOR
Mechanical impedance of the forearmIn four subjects (authors A.P. and P.St.J.T., and volunteers P.M.H.R. and
S. J. F.), the mechanical impedances of the forearm were analysed under variousconditions, including concomitant open- and closed-loop vibration. The experimentaland analytical techniques have been detailed elsewhere (Joyce et al. 1974; Brownet al. 1982). Briefly, the subject was seated in the chair illustrated in Fig. 1, but witha rigid, close-fitting cast enclosing and supporting the forearm and hand. By meansof a flywheel-crank mechanism, the elbow joint was subjected to sinusoidalflexion-extension movements of slowly increasing or decreasing frequency. Forces inphase with and in quadrature to the movement were computed for each cycle. Theseresults were then collected together, and averages were computed for the in-phaseand quadrature components of stiffness over each 0 5 Hz increment in frequency.Flywheel speed changed at a fairly constant rate, so the higher the frequency, thelarger the number of cycles averaged within a group (e.g. thirty cycles at 10 Hz cf.five cycles at 2 Hz).The plots which make up Fig. 10 show the resistance to movements in the
frequency range 2-10 Hz. At each frequency the resistance to movements may berepresented as a vector, and in each part of the Figure these vectors are displayed asa polar plot. As the frequency of movement increased, the mass of the forearm cameto dominate the responses, and the vectors moved towards the left. Figure lOA andB may be regarded as controls: for Fig. IOA the subject performed a steady 40% co-contraction of the elbow muscles, and for Fig. lOB, a constant-frequency (40 Hz)vibration was also applied to the biceps and triceps tendons. For Fig. 1OC, thevibrator frequencies were modulated by the movement in accordance with thetransfer function already described, and with a depth of modulation which hadsupported oscillation of the freely moving limb.
The effect of limb ma88. Although the results of these experiments were affected by the mass of themould which encased the forearm, these effects could easily be measured and subtracted (Evans,Fellows, Rack, Ross & Walters, 1983). For the experiments of Fig. 10, a relatively strong and heavymould was used, but in drawing the Figure, the effects of this extra mass have been subtracted toleave the components of stiffness as they would have been with the lighter mould which was usedfor the tremor measurements (moment of inertia calculated as 27-4 g M2. Forearm moment ofinertia of subject A.P.: 106 g m2, estimated from the. passive trials).
In Fig. IOA, as frequency increased the viscous component of stiffness reduced byabout one-half in the range 54-9-0 Hz. Previous comparisons with electricallytetanized muscle have shown that reductions of this sort can have both reflex andnon-reflex origins (Brown et al. 1982). Constant-frequency vibration (40 Hz, appliedsimultaneously to the elbow flexors and extensors) caused a small further decrease inviscous stiffness (Fig. 1OB). We take this as evidence that the reflexes were slightlyenhanced, and that therefore the vibratory amplitudes in these particular trials,which were performed with improvised fixation of the vibrator bodies, may not havebeen adequate to ensure complete 1: 1 driving of the I a afferents (Cussons et al.1980).Application of closed-loop vibration at a gain just above the threshold for
instability led in this subject to large, reproducible changes in the stiffness loci
431

432 A. PROCHAZKA AND P. ST. J. TREND
(Fig. IOC and D). In a trial involving 40% co-contraction (Fig. IOC), the frequency atwhich the locus crossed the vertical axis was 6-8 Hz. At this point the forearmpresented a pure negatively viscous impedance, assisting rather than resisting theimposed oscillations (Brown et al. 1982). The 6-8 Hz stiffness null compares well withthe 6-7 Hz spectral peak of tremor measured in the same subject, under the same
Viscous stiffness (N m/rad)
A 200 -c 200-
2 Hz ~~~~~~~~~~~~2 Hz10z10 Hz
5 Hz 10 Hz
400 200 200 400 400 200 400Elastic stiffness (N m/rad) 5 Hz
200 200
B 200 D 200
10Hz 2HHz 10 Hz2 Hz
400 200 200 400 400 200 5 Hz 200 400
200 200Fig. 10. Nyquist plots of averaged viscous v8. elastic stiffnesses about the elbow forimposed sinusoidal displacements increasing from 2 to 10 Hz. Each point on the C-shapedcurves may be viewed as a vector, whose distance from the origin corresponds to themodulus of stiffness, while its inclination to the horizontal axis indicates the angle bywhich the measured force sinusoid leads (above horizontal axis) or lags behind (belowaxis) the applied displacement. A, control trial, no vibration, subject performing 40%maximal voluntary contraction (MVC). B, 40% MVC, constant-frequency vibration(40 Hz to both flexors and extensors). C, 40% MVC closed-loop vibration. Notedevelopment of negative viscosity, and close approach of the locus to the origin, withcross-over of vertical axis at 6-8 Hz. This is the frequency at which the component ofstiffness in phase with the movement approaches zero, and is predictive of the tremorfrequency of the unrestricted forearm. D, 10% MVC closed-loop vibration. Locus crossesvertical axis at 4-3 Hz.
experimental conditions on a different day (Fig. 4, curve furthest to right). At 10%co-contraction the frequency of tremor predicted from the locus in Fig. lOD is4*3 Hz, which again agrees quite well with the 5-3 Hz tremor actually observed in thissubject at 10% co-contraction on a different occasion (Fig. 4). Even largerdestabilizing effects of closed-loop vibration were seen in one of the other subjects(S. J. F.). Subject P. St. J. T. showed clear but erratic destabilization, but in subjectP. M. H. R. the effects were minimal, and negative viscosities were not elicitable.The results of the impedance analysis are consistent with the frequency-response
data presented earlier in this paper. It is interesting to note the broadness of thefrequency range over which viscosities were negative (e.g. Fig. 1OC). This indicates

VIBRATION-EVOKED TREMOR
that the reflexes are not critically tuned, and could support independently generatedtremor over a fairly wide frequency range. Furthermore, when different values offorearm inertia were used to generate Nyquist plots (not illustrated here), it was clearthat the potential for altering the expected tremor frequencies was significant.
DISCUSSION
The simplest and most reproducible finding in this study was that forearm tremorcould readily be elicited in the large majority of subjects by artificially increasing theresponsiveness of muscle afferents to movement. This increase in responsiveness wasachieved by using the transduced forearm movement to frequency modulate thevibration of biceps and triceps brachii muscles. The advantage of the approach overlinear mathematical modelling of the reflex arc is that only one element of the loop,the proprioceptive afferent pathway, is 'externalized' and any non-linearities ofspinal cord and neuromuscular transduction are preserved within the rest of theloop.
Afferents contributing to the instabilityVibration is known to excite different types of mechanoreceptors, some more
effectively than others. In our experiments, the vibrator heads were pressed lightlyagainst the skin overlying the distal tendons of the forearm flexors and extensors. Ithas been argued that when vibration is applied in this way, the receptors mostactivated are the muscle spindle primary endings and that it is these receptors whichare largely responsible for the resulting reflexly evoked electromyographic activity(Matthews, 1984). That cutaneous receptors play at most a minor role was supportedby our observation that instability could not be elicited by vibrating the skin onceit had been pulled well clear of the underlying tendons. Indeed we found that thisform of stimulation had a negligible reflex effect on the muscles acting about theelbow. Muscle spindle secondary endings and tendon organs respond to longitudinalmuscle vibration, though usually much less sensitively than primary endings(Brown, Engberg & Matthews, 1967). When vibration is applied laterally to thetendon or muscle belly, the difference between spindle primary and secondarysensitivity may be reduced (Bianconi & van der Meulen, 1963). Moreover, in anactively contracting muscle, a small number of its tendon organs can be quiteresponsive to localized vibration (personal observations in conscious cats). Thereforewhile it seems reasonable to conclude that the vibration in our experiments evokedreflex effects largely via spindle primary afferents, a small contribution from spindlesecondary endings and tendon organs cannot be ruled out (see also Jacks, Prochazka& Trend, 1988).
Frequencies of the tremorsWe had expected reflex action to resist, rather than to contribute to, resonance
about the elbow at frequencies as low as 3-8 Hz. It had previously seemed that atthese frequencies, the force resulting from reflex EMG activity in a muscleundergoing stretch would occur early enough to oppose the stretch, reduce itsamplitude and thus damp oscillation in consecutive cycles (Joyce et al. 1974).
433

A. PROCHAZKA AND P. ST. J. TREND
However, Fig. 6 shows that when a subject voluntarily produced self-pacedalternating movements at around 4 Hz, the EMG bursts in both the flexors and theextensors occurred mainly during the phases of muscle lengthening in each cycle. TheEMG was typically 280 deg phase advanced on muscle length (maximal EMG cf.maximal muscle shortening, as shown in Fig. 7). Assuming that the forearm presentsa largely inertial load to the muscles, force would be expected to lead displacementby 180 deg. The 100 deg additional phase advance of the EMG represents a delaybetween neural input and peak force of about 70ms, which is consistent with theknown properties of load-moving muscle (Partridge, 1966). An inspection of Figs 3,4 and 6 shows that the form and relative timing of forearm movement and EMGactivities were similar whether recorded in closed-loop mode (Fig. 3), whether evokedfrom an external input (Fig. 4) or whether self-paced (Fig. 6). This implies that thesame reflex pathway was being tested in both closed- and open-loop modes.The open-loop trials illustrated in Figs 7-9 showed that at frequencies as low as
2-5-3-5 Hz, the phase lag of movement on presumed spindle afferent input exceeded180 deg. As mentioned above, in a closed-feed-back loop, at high gain instabilitydevelops atf180, the frequency at which there is a 180 deg phase lag around the loop.However, in order to predict from Fig. 9 the frequency of tremor in the closed reflexloop, it is first necessary to take into account the phase-advancing effects of themuscle spindle primary endings. Note that these were by-passed in the open-looptrials by the direct control of the spindle afferent firing rates by frequency-modulatedvibration. In Fig. 11, the spindle phase advances shown in Fig. 2 are added to anaveraged phase plot derived from Fig. 9. The resultant curve crosses 180 deg at about5 Hz, which is therefore the predicted mean frequency of tremor in the closed loopat 10% co-contraction. This agrees reasonably well with the experimentallyobserved mean tremor frequency of 4-3 Hz (Fig. 4; 10% maximum EMG).
Joyce et al. (1974) attributed negative viscosities in the mechanical impedance ofthe forearm at 8-12 Hz to reflex action. Negative viscosities, not encountered inpassive systems, represent active assistance of imposed movements. If, in addition toa negative viscosity, the in-phase stiffness approached zero (e.g. if the contractingmuscles and their load behaved like a spring-mass system with an 8-12 Hz naturalfrequency) the unloaded forearm would be expected to oscillate. Measurements atlow contraction strengths indicated a natural frequency about the elbow of 5 Hz, andJoyce et al. (1974) concluded that 8-12 Hz tremor would only occur if an externalload brought the natural frequency up into the 8-12 Hz range.
These results made it seem unlikely that reflexes could contribute to tremor at thelower frequencies. With hindsight, however, it is evident that the viscosities onlyshowed sharp dips at 9-10 Hz in trials involving strong (ca. 50% maximal)contractions, conditions under which we also observed resonances at somewhatincreased frequencies (5-7-1 Hz; Fig. 4). At lower contraction strengths (ca. 15%maximal; Joyce et al. 1974, Fig. 5, 35 N force), the viscosity was less sharply tuned,and in fact remained close to zero over the frequency range 4-11 Hz. Thecomputational technique for subtracting inertial forces due to the apparatus (Brownet al. 1982) had not been developed at this stage, and so zero in-phase stiffness of theforearm alone could not be detected.
Finally, it should be noted that at very low levels of co-contraction, the natural
434

VIBRATION-EVOKED TREMOR
frequency of the forearm due purely to its mechanical properties (in the absence ofdetectable EMG modulation) is about 3 Hz (Fox & Randall, 1970). Taken togetherwith the present results, this indicates that at 10% co-contraction, reflexes andpurely mechanical resonances are mutually supportive. With increasing co-contraction, increased muscle stiffness would presumably raise the mechanical
0
40
80
120
160cn 200
a 240
280
320
360
400
440
1 2 3 4 5 6 8 10 12
Frequency (Hz)Fig. 11. Reconstruction of the phase lag around the reflex arc. The lower curve is theaveraged phase plot of the curves in Fig. 9. The upper curve was obtained by adding ateach frequency the phase advance of muscle spindles from the phase curve in Fig. 2. Theeffect of including the spindle phase advance is to shift the 180 deg cross-over frequencyfrom ca. 3 to ca. 5 Hz.
resonant frequency. It is not immediately obvious why reflexes should continue tosupport tremor up to 6-8 Hz as the results in Fig. 4 imply. With increasing co-contraction, higher-threshold, fast-twitch motor units are brought into play (Burke,1973). This could well have the effect of increasing the frequency at which a 180 degphase lag is reached in the reflex loop. The twitch time-to-peak of type FF motorunits is about half that of type S motor units (Burke, Levine, Zajac, Tsairis & Engel,1971), and so it is quite conceivable that in going from a 10% to a 100% maximalco-contraction, the increasing proportion of fast motor units would gradually shiftthe 180 deg cross-over frequency from around 5 Hz, as in Fig. 11, into the 6-8 Hzrange.
Inaccuracies in spindle activationThe transfer function used to model the dynamic response of muscle spindles was
originally obtained in deefferented spindle endings in cat tenuissimus and soleusmuscles (Matthews & Stein, 1969; Poppele & Bowman, 1970). Three questions arisewhich are crucial to the interpretation of our results. First, does the spindle modelwe chose accurately mimic the responses of human spindles under normal fusimotoraction and during phasic extrafusal muscle activity? Second, as it is unlikely that allspindle primary endings fired one-to-one with each vibratory cycle, to what extent
435

A. PROCHAZKA AND P. ST. J. TREND
did we actually 'take over control' of the afferent input? Third, if the phasecharacteristics of the spindle model were in error, and/or if the afferents did notfollow exactly the vibratory signals, how much would this affect our mainconclusions regarding threshold and tremor frequency ?
Accuracy of the modelThere is very little direct information on spindle afferent firing during isotonic
tremor in normal individuals. Indeed the only relevant single-unit recording is thatof Hagbarth, Wallin & Lofstedt (1975, Fig. 4A), which shows a finger flexor Iaafferent unit firing during the muscle-lengthening phases of voluntary alternatingmovements at about 2 Hz. Poppele & Kennedy (1974) showed that biopsied,deefferented human spindles had dynamic properties which were closely similar tothose of the cat, phase advances being within 10 deg of each other over the wholefrequency range (0-1-20 Hz). Therefore from the available evidence, there is noreason to doubt that the Poppele & Bowman (1970) model is accurate for humanspindles in the linear range. The linear range in cat soleus spindles (Matthews &Stein, 1969) is exceeded by stretches larger than 200 ,tm peak-to-peak (ca. 0-2% oforigin-to-insertion rest length). From our own measurements of X-ray images (Amis,Prochazka, Short, Trend & Ward, 1987), 1 deg of elbow displacement corresponds toabout 800 ,cim origin-to-insertion displacement of the long head of biceps brachii (ca.0-2% of rest length). Thus tremor such as that in Fig. 3 involves muscle displacementswhich are probably within the linear range of the muscle spindles. The effect onspindle dynamics of fusimotor action is probably minimal within this range,involving at most a 20 deg reduction in the phase lead of spindle firing on length from1 to 10 Hz (Goodwin et al. 1975; Chen & Poppele, 1978). On the other hand fusimotoraction can substantially alter spindle gain. While this has little bearing on our use ofthe Poppele and Bowman model to establish the frequencies of tremor to whichreflexes would contribute, it does complicate our attempts to estimate safety marginswith respect to instability in normal individuals (see later).The deeply modulated extrafusal force which gives rise to tremor could cause
internal displacements in the muscle which might be significantly phase shifted withrespect to the origin-to-insertion displacements. This in turn would result in phaseshifts in spindle firing not taken into account in the passive model. Indeed it recentlyseemed that tendon compliance could be high enough for a complete phase reversalto occur between intramuscular and elbow joint movements at a frequency of only2-2 Hz (Fellows & Rack, 1986).
However, two lines of evidence militate against these arguments. First, in awakecats the firing profiles of spindle primary afferents during paw shakes were verysimilar to, and closely in phase with, the origin-to-insertion muscle length signal afterit had been filtered by an analog version of Poppele & Bowman's (1970) spindlemodel (Diirmiiller, Hulliger, Prochazka & Trend, 1987). This was true even when theinertia of the foot was doubled and tripled by attaching external weights. Paw shakesinvolve some of the fastest movements of which mammals are capable: accelerationsof the paw in excess of 25 G have been reported (Hoy, Zernicke & Smith, 1985). Ifextrafusal contractions and tendon compliance caused significant discrepanciesbetween the length variations recorded between origin and insertion of the muscle
436

VIBRATION-EVOKED TREMOR
and those 'seen' by the spindles, this should have shown up clearly in these extremecircumstances. It could still be argued that the elbow flexors in humans might haverelatively longer and more compliant tendons than do cat soleus muscles, that theyact upon a relatively larger inertial load, and therefore that they are more likely todevelop intramuscular phase reversals. Frame-by-frame analysis of X-ray films ofelbow tremor has, however, failed to reveal phase reversals in the frequency range0-7 Hz (Prochazka & Trend, 1986; Amis et al. 1987). Similar results were obtainedwith the use of a different technique by Fellows & Rack (1987).We conclude that the Poppele & Bowman (1970) spindle model transduced the
forearm movements in much the same way as the subjects' own muscle spindleprimary afferents would have done, with a maximum phase discrepancy of about20 deg.
Extent of take-over of spindle firingConstant-frequency vibration applied laterally to the tendon of a muscle at rest
produces 1: 1 firing in most of the muscle's spindle primary endings provided that itis of adequate amplitude and is transferred efficiently to the tendon (cat: Bianconi& van der Meulen, 1963; personal observations; man: Burke, Hagbarth, Lofstedt &Wallin, 1976; Roll & Vedel, 1982). However, extrafusal contraction (Brown et al.1967) and muscle shortening (Matthews & Watson, 1981 a, but cf. Burke et al. 1976)can reduce the security of 1: 1 driving. Indeed Matthews & Watson (1981 a) couldadjust vibratory amplitude so as to produce 1: 1 driving only during the lengtheningphases of concomitant sinusoidal stretching (1-8 Hz) of soleus muscles in the cat.
It is impossible to be sure of the appropriate vibratory amplitude for securedriving under the conditions of our experiments. We found that for displacements ofthe vibrator heads in the range 0-5-1-0 mm peak-to-peak (40 Hz), the open-loop gainrequired to produce instability in the closed-loop experiments remained fairlyconstant in a given subject. Until threshold was reached, the forearm exhibited onlyminute, irregular movements associated with the subject maintaining the required10% co-contraction. These movements were of course fed back to modulatevibratory frequency in the closed loop, but because they were so small the depth ofmodulation about the 40 Hz carrier frequency was usually minimal. As tremor set in,the initial cycles were often at a very small amplitude, yet their frequency was closeto that of established tremor at much larger amplitudes. This suggests that anyinsecurity of driving caused by extrafusal activation had little effect on the transferfunction relating movement to afferent firing. As it happens, the phase of thedeveloped tremor cycle in which most of the vibration of a muscle group occurredcorresponded to the phase in which the security of 1: 1 driving was presumably thegreatest, namely during muscle lengthening (Matthews & Watson, 1981 a, b). Thiswas also true of the open-loop trials of Figs 8, 9 and 11, in the frequency range3-6 Hz, where the 180 deg phase lag cross-overs occurred.Though the above factors support 1: 1 driving in many, and possibly most, of the
relevant spindles, it does seem likely that at least some endings, particularly thosein the deeper elbow flexor muscles, brachialis and brachioradialis, did not have theirdischarge locked 1: 1 with the vibratory cycles. In the Results section the thresholdfor instability in our subjects (1500-9000 Hz/1 % rest length) was contrasted with
437

A. PROCHAZKA AND P. ST. J. TREND
the sensitivity of cat soleus Ia afferents (1000 s-1 (1% rest length)-'). On thesimplified assumptions of 1:1 driving and identical sensitivities of cat and humanspindles (cf. Newsom Davis, 1975), we would have concluded that the subjects withthe lowest thresholds needed only a small (10%) increase in their normal spindlesensitivities to go into instability. In fact a lack of secure driving in a proportion ofthe spindles in our trials would have led us to overestimate the feed-back sensitivityneeded to produce instability and thus to overestimate the safety margin withrespect to instability of our most reflexly active subjects under normal (unvibrated)conditions. Some of these subjects did indeed exhibit episodes of spontaneous tremorwhilst attempting to maintain a constant elbow angle at 10% maximum co-contraction.Fusimotor activation in normal subjects adds another interesting dimension to
this discussion. As mentioned above, though fusimotor action has little effect onspindle dynamics, it does influence gain. Strong static fusimotor action substantiallyreduces spindle primary responsiveness to small-amplitude stretches. In our view,this would reduce the tendency of stretch reflexes to generate or assist tremor.Dynamic fusimotor action slightly decreases spindle gain at long muscle lengths, butcan increase it at shorter lengths, particularly for the larger sinusoidal stretches(Goodwin et al. 1975; Chen & Poppele, 1978). Furthermore, low-level static fusimotoraction (single y-fibres stimulated at 20-30 Hz) paradoxically also increases spindleprimary gain at lower muscle lengths (Hulliger, Emonet-Denand & Baumann, 1985).This raises the interesting possibility that different combinations of fusimotor actionor 'set' could either increase or decrease the likelihood of tremor.
It must be conceded that the uncertainty surrounding the assumption of 1: 1driving, and the lack of hard data on the sensitivities of human forearm musclespindles in awake subjects, does limit the reliability of the above estimates of safetymargin. On the other hand, relative instability thresholds quantified using ourapproach might prove useful in detecting changes in the reflex excitability of a givensubject under different conditions (e.g. before and after medication), or incomparisons between different individuals. The advantage over existing methods isthat only a single observation is required, namely the onset of tremor, and a singlereading need be taken, namely the gain of the feed-back pathway.
Effect of errors in spindle activation on our main conclusionsThough we have argued above that the Poppele & Bowman (1970) spindle model
probably mimics human spindle responses very closely, the question arises whethereven quite small inaccuracies of the model, or inaccuracies in 1: 1 driving of spindleendings, might have substantially affected our observations and conclusions.To test the first possibility, we altered the transfer function of the model so that
at one extreme a flat response characteristic was used (i.e. no poles or zeros: 0 degphase advance at all frequencies) and at the other, a zero was introduced whichproduced an additional 30 deg phase advance at 4 Hz. The tremor frequencies thenobserved in closed-loop trials at these extremes were approximately 3 0 and 5-5 Hzrespectively. These values are in fact quite close to the frequencies at which 180 degphase lags around the reflex loop would be expected on the basis of the phase curvesof Fig. 11. Our conclusions regarding tremor frequencies at the elbow therefore seem
438

VIBRATION-EVOKED TREMOR
unlikely to be seriously affected by inaccuracies in the dynamics of the spindle modelwe used.
It is conceivable that stretch-dependent security of 1: 1 driving, apart frominfluencing gain, could also introduce hidden phase shifts between the vibratorcommand signal and the spindle response. To assess this effect, we deliberatelyreduced the amount of travel of the vibrator heads. We found that for vibratoryamplitudes below 0-5 mm peak-to-peak, the feed-back sensitivities required toproduce instability increased. However, the frequencies of the observed tremors werelittle affected, and at 10% co-contraction remained in the range 3-5.5 Hz.
Relationship to pathological tremorsOur results indicate that stretch reflexes mediated by spindle primary afferents
facilitate 3-8 Hz oscillations at the elbow. In normal individuals, the loop gainaround the reflex arc is apparently usually not high enough to result in instability,though the safety margin may be small in some subjects. There may be situationsinvolving apprehension, fear or unfamiliarity with a difficult motor task where thethreshold is exceeded. Some skilled rhythmical movements such as musicians'vibrato might depend upon a learned ability to deliberately 'wind up' reflex gain(Schlapp, 1973) or fusimotor action (Prochazka, Hulliger, Zangger & Appenteng,1985).
Pathological tremors in the 3-8 Hz range at the elbow are most often seen inParkinson's disease, cerebellar disorders and to a lesser extent in spasticity. It isgenerally believed that Parkinsonian and cerebellar tremors often depend uponrhythmic activity generated within the CNS (for review see Marsden, 1984), thoughan interaction with spinally mediated stretch reflexes is frequently mooted (e.g. Lee& Stein, 1981; Marsden, 1984; Rack & Ross, 1986). Our data show that in principlea very active stretch reflex loop could oscillate autonomously in the frequency range3-8 Hz. However, there is convincing evidence (Marsden, 1984) that this is not theusual mechanism underlying pathological tremors (with the possible exception ofspastic clonus: Rack, Ross & Thilmann (1984)). On the other hand, stretch reflexesmay contribute significantly to established tremors, and thus the more generalimplication of our findings is that reflex contributions should be sought in thefrequency range 3-8 Hz rather than, as hitherto, in the range 8-12 Hz.A central neural oscillator entirely unaffected by peripheral input would effectively
'see' the (closed) peripheral reflex loop as a neural-mechanical transducer with atransfer function exhibiting a resonant peak 'tuned' by flexor and extensor co-contraction to some frequency between 3 and 8 Hz. In such cases, the existence of thetremor would be independent of changes in reflex transmission though its amplitudemight be influenced to a moderate extent. If on the other hand the central oscillatorwere itself dependent upon long-loop feed-back from muscle afferents, the closed-loop transfer function of the peripheral reflex arc would effectively be interposed inthe forward pathway of the long loop. As such it would contribute to the overalltransfer function of the long loop, and could significantly influence stability. Indeedin marginal cases it could 'tip the balance' from stability to oscillation. Fluctuatinglevels of co-contraction could be quite important in this respect, and might in somecases underlie the characteristic waxing and waning of Parkinsonian tremor.
439

A. PROCHAZKA AND P. ST. J. TREND
Conclusion
We conclude that at moderate levels of co-contraction about the elbow, stretchreflexes contribute to tremor at 3-8 Hz rather than at 8-12 Hz, as previouslythought. It should be emphasized that this applies to unloaded, self-generatedmovements of the forearm, and is by no means generally true for all joints and allloading conditions. When external loads such as stiff springs are added (e.g. Joyce &Rack, 1974; Cussons, Matthews & Muir, 1980; Matthews & Muir, 1980) the closed-loop transfer function is altered not only by the impedance of the spring, but also bynon-linear changes in the signal-handling properties of the muscles (Partridge, 1966).This would be expected to affect appreciably the frequency at which spontaneoustremor would occur, and the extent to which stretch reflexes would contribute to orresist the instability.
We thank Miss K. M. Silvester for help in the early stages of this project and Mr S. Vincent fortechnical assistance. Drs P. M. H. Rack and R. B. Stein are thanked for their critical reading of themanuscript. Dr Rack and Dr H. Ross of the Department of Physiology, University of Birminghamare further thanked for kindly allowing us to use their facilities and helping us with the experimentsillustrated in Fig. 10. Dr P. St. J. Trend is supported by the British MRC as a Training Fellow. Theproject was funded by the National Fund for Crippling Diseases (Action Research for the CrippledChild) and the St Thomas's Hospital (Research) Endowments Committee.
REFERENCES
AMIS, A., PROCHAZKA, A., SHORT, D., TREND, P. & WARD. A. (1987). Relative displacements inmuscle and tendon during human arm movements. Journal of Physiology 389, 37-44.
BIANCONI, R. & VAN DER MEULEN, J. P. (1963). The responses to vibration of the end-organs ofmammalian muscle spindles. Journal of Neurophysiology 26, 177-190.
Bizzi, E., DEV, P., MORASSO, P. & POLIT, A. (1978). Effect of load disturbances during centrallyinitiated movements. Journal of Neurophysiology 41, 542-556.
BROWN, M. C., ENGBERG, I. & MATTHEWS, P. B. C. (1967). The relative sensitivity to vibration ofmuscle receptors of the cat. Journal of Physiology 192, 773-800.
BROWN, T. I. H., RACK, P. M. H. & Ross, H. F. (1982). Forces generated at the thumbinterphalangeal joint during imposed sinusoidal movements. Journal of Physiology 332,69-86.
BURKE, D., HAGBARTH, K.-E., LOFSTEDT, L. & WALLIN, B. G. (1976). The responses of humanmuscle spindle endings to vibration during isometric contraction. Journal of Physiology 261,695-711.
BURKE, R. E. (1973). On the central nervous system control of fast and slow twitch motor units.In New Developments in Electromyography and Clinical Neurophysiology, ed. DESMEDT, J. E.,pp. 69-94. Basel: Karger.
BURKE, R. E., LEVINE, D. N., ZAJAC, F. C., TSAIRIS, P. & ENGEL, W. K. (1971). Mammalian motorunits: physiological-histochemical correlation in three types in cat gastrocnemius. Science 174,709-712.
CHEN, W. J. & POPPELE, R. E. (1978). Small-signal analysis of response of mammalian musclespindles with fusimotor stimulation and a comparison with large-signal properties. Journal ofNeurophysiology 41, 15-27.
CUSSONS, P. D., MATTHEWS, P. B. C. & MUIR, R. B. (1980). Enhancement by agonist or antagonistmuscle vibration of tremor at the elastically loaded human elbow. Journal of Physiology 302,443-461.
DIMITRIJEVIC, M. R., SHERWOOD, A. M. & NATHAN, P. (1978). Clonus: peripheral and centralmechanisms. In Progress in Clinical Neurophysiology, vol. 5, ed. DESMEDT, J. E., pp. 173-182.Basel: Karger.
440

VIBRATION-EVOKED TREMOR
DijRMfULLER, N., HULLIGER, M., PROCHAZKA, A. & TREND, P. ST. J. (1987). Muscle receptor firingduring paw shakes in cats. Journal of Physiology 392, 62P.
EVANS, C. M., FELLOWS, S. J., RACK, P. M. H., Ross, H. F. & WALTERS, D. K. W. (1983).Response of the normal human ankle joint to imposed sinusoidal movements. Journal ofPhysiology 344, 483-502.
FELLOWS, S. J. & RACK, P. M. H. (1986). Relation of the length of an electrically stimulated humanbiceps to elbow movement. Journal of Physiology 376, 58P.
FELLOWS, S. J. & RACK, P. M. H. (1987). Changes in the length of the human biceps brachii muscleduring elbow movements. Journal of Physiology 383, 405-412.
Fox, J. R. & RANDALL, J. E. (1970). Relationship between forearm tremor and the bicepselectromyogram. Journal of Applied Physiology 29, 103-108.
FREUND, H.-J., HEFTER, H., H6MBERG, V. & REINERS, K. (1984). Determinants of tremor rate. InMovement Disorders: Tremor, ed. FINDLEY, L. J. & CAPILDEO, R., pp. 195-204. London:Macmillan.
GOODWIN, G., HULLIGER, M. & MATTHEWS, P. B. C. (1975). The effects of fusimotor stimulationduring small-amplitude stretching on the frequency-response of the primary ending of themammalian muscle spindle. Journal of Physiology 253, 175-206.
HAGBARTH, K.-E., WALLIN, G. & L6FSTEDT, L. (1975). Muscle spindle activity in man duringvoluntary fast alternating movements. Journal of Neurology, Neurosurgery and Psychiatry 38,625-635.
Hoy, M. G., ZERNICKE, R. F. & SMITH, J. L. (1985). Contrasting roles of inertial and musclemoments at knee and ankle during paw-shake response. Journal of Neurophysiology 5,1282-1294.
HULLIGER, M., EMONET-DE1NAND, F. & BAUMANN, T. K. (1985). Enhancement of stretch sensitivityof cat primary spindle afferents by low-rate static gamma action. In The Muscle Spindle, ed.BOYD, I. A. & GLADDEN, M. H., pp. 189-193. London: Macmillan.
JACKS, A., PROCHAZKA, A. & TREND, P. ST. J. (1988). Instability in human forearm movementsstudied with feedback-controlled electrical stimulation of muscles. Journal of Physiology 402,443-461
JOYCE, G. C. & RACK, P. M. H. (1974). The effects of load and force on tremor at the normal humanelbow joint. Journal of Physiology 240, 375-396.
JOYCE, G. C., RACK, P. M. & Ross, H. F. (1974). The forces generated at the human elbow jointin response to imposed sinusoidal movements of the forearm. Journal of Physiology 240,351-374.
LEE, R. G. & STEIN, R. B. (1981). Resetting of tremor by mechanical perturbations: a comparisonof essential tremor and Parkinsonian tremor. Annals of Neurology 10, 523-531.
LLINA'S, R. L. (1984). Rebound excitation as the physiological basis for tremor: a biophysical studyof the oscillatory properties of mammalian central neurones in vitro. In Movement Disorders:Tremor, ed. FINDLEY, L. J. & CAPILDEO, R., pp. 165-182. London: Macmillan.
MARSDEN, C. D. (1984). Origins of normal and pathological tremor. In Movement Disorders:Tremor, ed. FINDLEY, L. J. & CAPILDEO, R., pp. 37-84. London: Macmillan.
MATTHEWS, P. B. C. (1972). Mammalian Muscle Receptors and their Central Actions, pp. 591-597.London: Arnold.
MATTHEWS, P. B. C. (1984). Evidence from the use of vibration that the human long-latencystretch reflex depends upon spindle secondary afferents. Journal of Physiology 348, 383-415.
MATTHEWS, P. B. C. & MUIR, R. B. (1980). Comparison of electromyogram spectra with forcespectra during human elbow tremor. Journal of Physiology 302, 427-441.
MATTHEWS, P. B. C. & STEIN, R. B. (1969). The sensitivity of muscle spindle afferents to smallsinusoidal changes of length. Journal of Physiology 200, 723-743.
MATTHEWS, P. B. C. & WATSON, J. D. G. (1981 a). Action of vibration on the responses of catmuscle spindle Ia afferents to low frequency sinusoidal stretching. Journal of Physiology 317,365-382.
MATTHEWS, P. B. C. & WATSON, J. D. G. (1981 b). Effect of vibrating agonist or antagonist mliscleon the reflex response to sinusoidal displacement of the human forearm. Journal of Physiology321, 297-316.
NEWSOM DAVIS, J. (1975). The response to stretch of human intercostal muscle spindles studied invitro. Journal of Physiology 249, 561-579.
441

A. PROCHAZKA AND P. ST. J. TREND
O'BRIEN. R., PROCHAZKA, A. & VINCENT, S. (1985). Two versatile laboratory interfaces for multi-channel data acquisition and analysis, using the B3BC microcomputer. Journal ofPhysiology 372,3P.
PARTRIDGE, L. D. (1966). Signal-handling characteristics of load-moving skeletal muscle. AmericanJournal of Physiology 210, 1178-1191.
POPPELLE, R. E. & BOWMAN, R. J. (1970). Quantitative description of linear behaviour ofmammalian muscle spindles. Journal of Neurophysiology 33, 59-72.
POPPELE, R. E. & KENNEDY, W. R. (1974). Comparison between behavior of human and catmuscle spindles recorded in vitro. Brain Research 75, 316-319.
PROCHAZKA, A., HULLIGER, M., ZANGGER, P. & APPENTENG, K. (1985). 'Fusimotor set': newevidence for a-independent control of y-motoneurones during movement in the awake cat. BrainResearch 339, 136-140.
PROCHAZKA, A. & TREND, P. (1986). X-ray imaging of humaii muscle contractioll. Journal ofPhysiology 337, 15P.
RACK, P. M. H. & Ross, H. F. (1986). The role of reflexes in the resting tremor of Parkinson'sDisease. Brain 109, 115- 141.
RACK, P. M. H., Ross, H. F. & THILMANN, A. L. (1984). The ankle stretch reflexes in normal andspastic subjects. Brain 107, 637-654.
ROLL, J. P. & VEDEL, J. P. (1982). Kinaesthetic role of muscle afferents in man, studied by tendonvibration and microneurography. Experimental Brain Research 47, 177-190.
ROTHWELL, J. C., TRAUB, M. M., DAY, B. L., OBESO, J. A., THOMAS, P. K. & MARSDEN, C. D.(1982). Manual motor performance in deafferented man. Brain 105, 515-542.
SCHLAPP, M. (1973). Observations on a voluntary tremor - violinist's vibrato. Quarterly Journal ofExperimental Physiology 58, 357-368.
STEIN, R. B. & OGUZT6RELI, M. N. (1976). Does the velocity sensitivity of muscle spindles stabilizethe stretch reflex ? Biological Cybernetics 23, 219-228.
442