
Initiation of general recombination catalyzed in vitro by the recA
5
Proc. Natl. Acad. Sci. USA Vol. 76, No. 6, pp. 2615-2619, June 1979 Biochemistry Initiation of general recombination catalyzed in vitro by the recA protein of Escherichia coli (single-strand assimilation/D-loops/cooperative duplex DNA binding/DNA-dependent ATPase/ATP-dependent renaturation) K. MCENTEE, G. M. WEINSTOCK, AND I. R. LEHMAN Department of Biochemistry, Stanford University School of Medicine, Stanford, California 94305 Contributed by I. Robert Lehman, March 12, 1979 ABSTRACT Homogeneous recA protein catalyzes the hy- bridization of single-stranded DNA to homologous regions in duplex DNA. The products are D-loops, which are formed with equal efficiency in linear and supercoiled molecules. This as- similation reaction can be separated into two partial reactions. In the first, recA protein binds to duplex DNA and produces a recA protein-DNA complex. The binding shows a sigmoidal dependence on recA protein concentration, requires ATP, GTP or the 'y-thio analog of ATP, and Mg2+, but does not require hydrolysis of the nucleoside triphosphate. In the second reac- tion, single-stranded regions of the recA protein-ATP-duplex DNA intermediate hybridize with free complementary single strands to produce D-loop structures. This reaction is coupled to ATP hydrolysis and is analogous to the renaturation of sin- gle-stranded DNA catalyzed by the recA protein [Weinstock, G. M., McEntee, K. & Lehman, L R. (1979) Proc. Nat. Acad. Sci. USA 76,128-130]. Hydrolysis of ATP appears to be required in these reactions for dissociation of recA protein from the DNA. General recombination in Escherichia coli proceeds through the formation of a duplex DNA intermediate containing a complementary strand from each parental molecule. The for- mation of this heteroduplex region occurs at an early step in recombination and is thought to be initiated by the transfer of a single strand from one duplex to another (1). A product of this initiation reaction may be a locally triple-stranded structure called a D-loop containing one single-stranded and one dou- ble-stranded branch. The invading single strand may be derived from another duplex molecule or from free single-stranded DNA. The assimilation into intact duplex DNA must involve local denaturation of the recipient duplex and hybridization of the invading single strand to the denatured region. Holloman and coworkers have provided evidence that implicates D-loops in recombination and have demonstrated a requirement for the recA gene in assimilation in vivo (2, 3). Recently we have demonstrated that purified recA protein catalyzes the rena- turation of complementary single strands to form duplex DNA in a reaction that is coupled to the hydrolysis of ATP (4). The recA protein purified from a cold-sensitive recA mutant strain is cold labile in its capacity to catalyze DNA renaturation, a finding that correlates with the recombination defect in mvo. It is likely that this renaturation reaction reflects the hybrid- ization step in the assimilation process. These observations prompted us to investigate the interaction of recA protein with duplex DNA, the other substrate in the assimilation process. In this paper we demonstrate that recA protein catalyzes assimilation of single-stranded DNA into a homologous duplex region. The assimilation reaction has been The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "ad- vertisement" in accordance with 18 U. S. C. §1734 solely to indicate this fact. 2615 separated into two partial reactions consisting of the binding of recA protein to duplex DNA, followed by hybridization of complementary single-stranded regions. These results indicate that recA protein catalyzes initiation of strand exchange during genetic recombination. While this work was in -progress, Shibata et al. (5) commu- nicated to us their results demonstrating that recA protein catalyzes single-strand assimilation into superhelical DNA. MATERIALS AND METHODS The recA protein was purified from E. coli strain KM1842 as described (4). Only a single protein band with a molecular weight of approximately 40,000 was detected on polyacryl- amide gels containing sodium dodecyl sulfate (NaDodSO4) when 10,gg of protein was electrophoresed and stained with Coomassie blue. The purified material contained no detectable endo- or exonuclease, DNA or RNA polymerase, or polynu- cleotide phosphorylase activities. The enzyme was stable at 00C for several weeks. 3H-Labeled phage P22 DNA was prepared as described (6). 3H-Labeled phage M13 replicative form I (RFI) and unlabeled M13 viral single-strand DNA were generous gifts of J. Ko- bori. Single-stranded DNA was prepared by heating unlabeled P22 DNA at 1000C for 3 min followed by quick cooling on ice. Viral M13 DNA was heated at 100°C for 10 min to prepare linear single-stranded fragments. Assimilation of single strands into 3H-labeled P22 DNA was measured essentially as described by Beattie et al. (7). Incu- bations were performed as described below for binding of recA protein to duplex DNA except that 650 pmol of single-stranded DNA was included in the reaction. After incubation at 37°C, 1/10th vol of 10% Sarkosyl or 10% NaDodSO4 was added to remove protein from the DNA and the samples were placed on ice for 3-5 min. One milliliter of cold 1.5 M NaCl/150 mM sodium citrate was added and the samples were filtered through nitrocellulose filters (Millipore, type HA). Each filter was washed with an additional 3 ml of the 1.5 M NaCl/150 mM sodium citrate, dried, and assayed for radioactivity. In this assay, retention of 3H-labeled DNA is completely dependent upon added homologous single-stranded DNA. Binding of recA protein to duplex DNA was measured by retention of DNA on nitrocellulose filters. Reaction mixtures (200 ,tl) contained B buffer [20 mM Tris-HCI (pH 7.5)/10 mM MgCl2/20 mM NaCl/0.1 mM EDTA/0. 1 mM dithiothreitol/15 gg of bovine serum albumin per ml), 1.45 nmol of 3H-labeled Abbreviations: NaDodSO4, sodium dodecyl sulfate; RFI, supercoiled double-stranded DNA (replicative form I); ATPIy-SJ, adenosine 5'- [y-thioltriphosphate; ADP[NHJP, adenyl-5'-yl imidodiphosphate; ADP(CH21P, adenyl-5'-yl methylenediphosphate.
Transcript of Initiation of general recombination catalyzed in vitro by the recA

Proc. Natl. Acad. Sci. USAVol. 76, No. 6, pp. 2615-2619, June 1979Biochemistry
Initiation of general recombination catalyzed in vitro by the recAprotein of Escherichia coli
(single-strand assimilation/D-loops/cooperative duplex DNA binding/DNA-dependent ATPase/ATP-dependent renaturation)
K. MCENTEE, G. M. WEINSTOCK, AND I. R. LEHMANDepartment of Biochemistry, Stanford University School of Medicine, Stanford, California 94305
Contributed by I. Robert Lehman, March 12, 1979
ABSTRACT Homogeneous recA protein catalyzes the hy-bridization of single-stranded DNA to homologous regions induplex DNA. The products are D-loops, which are formed withequal efficiency in linear and supercoiled molecules. This as-similation reaction can be separated into two partial reactions.In the first, recA protein binds to duplex DNA and produces arecA protein-DNA complex. The binding shows a sigmoidaldependence on recA protein concentration, requires ATP, GTPor the 'y-thio analog of ATP, and Mg2+, but does not requirehydrolysis of the nucleoside triphosphate. In the second reac-tion, single-stranded regions of the recA protein-ATP-duplexDNA intermediate hybridize with free complementary singlestrands to produce D-loop structures. This reaction is coupledto ATP hydrolysis and is analogous to the renaturation of sin-gle-stranded DNA catalyzed by the recA protein [Weinstock,G. M., McEntee, K. & Lehman, L R. (1979) Proc. Nat. Acad. Sci.USA 76,128-130]. Hydrolysis of ATP appears to be required inthese reactions for dissociation of recA protein from theDNA.
General recombination in Escherichia coli proceeds throughthe formation of a duplex DNA intermediate containing acomplementary strand from each parental molecule. The for-mation of this heteroduplex region occurs at an early step inrecombination and is thought to be initiated by the transfer ofa single strand from one duplex to another (1). A product of thisinitiation reaction may be a locally triple-stranded structurecalled a D-loop containing one single-stranded and one dou-ble-stranded branch. The invading single strand may be derivedfrom another duplex molecule or from free single-strandedDNA. The assimilation into intact duplex DNA must involvelocal denaturation of the recipient duplex and hybridizationof the invading single strand to the denatured region. Hollomanand coworkers have provided evidence that implicates D-loopsin recombination and have demonstrated a requirement for therecA gene in assimilation in vivo (2, 3). Recently we havedemonstrated that purified recA protein catalyzes the rena-turation of complementary single strands to form duplex DNAin a reaction that is coupled to the hydrolysis of ATP (4). TherecA protein purified from a cold-sensitive recA mutant strainis cold labile in its capacity to catalyze DNA renaturation, afinding that correlates with the recombination defect in mvo.It is likely that this renaturation reaction reflects the hybrid-ization step in the assimilation process.
These observations prompted us to investigate the interactionof recA protein with duplex DNA, the other substrate in theassimilation process. In this paper we demonstrate that recAprotein catalyzes assimilation of single-stranded DNA into ahomologous duplex region. The assimilation reaction has been
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.
2615
separated into two partial reactions consisting of the bindingof recA protein to duplex DNA, followed by hybridization ofcomplementary single-stranded regions. These results indicatethat recA protein catalyzes initiation of strand exchange duringgenetic recombination.
While this work was in-progress, Shibata et al. (5) commu-nicated to us their results demonstrating that recA proteincatalyzes single-strand assimilation into superhelical DNA.
MATERIALS AND METHODSThe recA protein was purified from E. coli strain KM1842 asdescribed (4). Only a single protein band with a molecularweight of approximately 40,000 was detected on polyacryl-amide gels containing sodium dodecyl sulfate (NaDodSO4)when 10,gg of protein was electrophoresed and stained withCoomassie blue. The purified material contained no detectableendo- or exonuclease, DNA or RNA polymerase, or polynu-cleotide phosphorylase activities. The enzyme was stable at 00Cfor several weeks.
3H-Labeled phage P22 DNA was prepared as described (6).3H-Labeled phage M13 replicative form I (RFI) and unlabeledM13 viral single-strand DNA were generous gifts of J. Ko-bori.
Single-stranded DNA was prepared by heating unlabeledP22 DNA at 1000C for 3 min followed by quick cooling on ice.Viral M13 DNA was heated at 100°C for 10 min to preparelinear single-stranded fragments.
Assimilation of single strands into 3H-labeled P22 DNA wasmeasured essentially as described by Beattie et al. (7). Incu-bations were performed as described below for binding of recAprotein to duplex DNA except that 650 pmol of single-strandedDNA was included in the reaction. After incubation at 37°C,1/10th vol of 10% Sarkosyl or 10% NaDodSO4 was added toremove protein from the DNA and the samples were placed onice for 3-5 min. One milliliter of cold 1.5 M NaCl/150 mMsodium citrate was added and the samples were filtered throughnitrocellulose filters (Millipore, type HA). Each filter waswashed with an additional 3 ml of the 1.5 M NaCl/150 mMsodium citrate, dried, and assayed for radioactivity. In this assay,retention of 3H-labeled DNA is completely dependent uponadded homologous single-stranded DNA.
Binding of recA protein to duplex DNA was measured byretention of DNA on nitrocellulose filters. Reaction mixtures(200 ,tl) contained B buffer [20mM Tris-HCI (pH 7.5)/10mMMgCl2/20mM NaCl/0.1 mM EDTA/0. 1 mM dithiothreitol/15gg of bovine serum albumin per ml), 1.45 nmol of 3H-labeled
Abbreviations: NaDodSO4, sodium dodecyl sulfate; RFI, supercoileddouble-stranded DNA (replicative form I); ATPIy-SJ, adenosine 5'-[y-thioltriphosphate; ADP[NHJP, adenyl-5'-yl imidodiphosphate;ADP(CH21P, adenyl-5'-yl methylenediphosphate.

2616 Biochemistry: McEntee et al.
P22 DNA (3 X 104 cpm/nmol), recA protein, and nucleosidetriphosphate (ATP or GTP) as indicated. After incubation at370C, aliquots were applied directly to filters presoaked withB buffer. The samples were washed once with 0.5 ml of B buffercontaining 1.0 M NaCl and then washed with an additional 0.5ml of B buffer, dried, and assayed for radioactivity. In the ab-sence of recA protein less than 2% of the duplex DNA was re-
tained.Reaction products were visualized by electron microscopy
by means of the formamide spreading method of Davis et al.(8).
Hydrolysis of ATP was determined as described (4) exceptthat the reaction mixture (25 ttl) contained 225 pmol of duplexDNA, recA protein, and 500 IAM ATP.
3H-Labeled nucleoside triphosphates were obtained fromAmersham/Searle or New England Nuclear. Adenosine 5'-['y-thio]triphosphate (ATP['y-SJ), adenyl-5'-yl imidodiphos-phate (ADP[NH]P), and adenyl-5'-yl methylenediphosphate(ADP[CH2]P) were obtained from Boehringer Mannheim. SInuclease was purchased from Sigma and proteinase K was ob-tained from EM Laboratories (Elmsford, NY).
RESULTSrecA protein catalyzes assimilation of homologoussingle strands into duplex DNAIn the presence of ATP, Mg2+, and complementary (unlabeled)single-stranded P22 DNA, th~iiecA protein converts 3H-labeledduplex P22 DNA into a form that is retained on nitrocellulosefilters. Under optimal conditions,- approximately 50% of theDNA was retained on nitrocellulose filters. Retention of thesecomplexes was eliminated by Si nuclease treatment (less than5% retention) but was resistant to detergents (1% Sarkosyl or
1% NaDodSO4) or proteinase K treatment (data not shown).Retention of the labeled DNA was reduced more than 50%when the reaction products were heated to 550C for 5 min (seeDiscussion). These results suggest that recA protein promotesthe assimilation of single strands into the labeled duplex to formD-loops that are retained on nitrocellulose filters by virtue ofthe single-stranded regions that they possess.
When the products of the assimilation reaction were exam-
ined by electron microscopy, D-loop-containing molecules suchas those shown in Fig. 1 A and B were detected. D-loops werealso observed when recA protein was incubated with super-
coiled M13 RFI DNA and homologous single-stranded frag-ments (Fig. IC).
Single-strand assimilation required recA protein, ATP, andhomologous single-stranded DNA (Table 1). Heterologous calfthymus or ColEl single strands did not substitute for P22 DNA,although single-stranded phage X DNA produced a small butsignificant amount of binding. The interaction of X DNA withthe P22 duplex may reflect the limited homology between theseDNAs (9). The ATP requirement was not satisfied by any otherribonucleoside triphosphate, including the ATP-y-S derivative,which inhibits both recA protein-catalyzed DNA renaturationand ATP hydrolysis (4). The assimilation reaction, like the re-
naturation reaction, was sensitive to N-ethylmaleimide. Therequirements for uptake of single strands into superhelical DNAwere identical to those for linear duplex DNA.The kinetics of single-strand assimilation into linear P22
duplex DNA are shown in Fig. 2. After a lag of about 2 minafter addition of enzyme, the reaction was complete in 15 min,with nearly 50% of the DNA being retained. The extent was
unaffected by addition of more recA protein or ATP after 20min of incubation (data not shown). Similar kinetics were ob-served for assimilation of single-stranded M13 DNA fragmentsinto supercoiled M13 RFI DNA (data not shown).
'9t.t.,,; t\- A t
tsr~~~~~~~~~~~~~r
* 4e_.' +^>
~rw
it' X *<t'f' e 4'
f * *~~~~t- i,t
B
6~~, ,.,,:, 'w w.'*w-saJ6;t'
:w% si. ;, ) \f
Ao~~~~r
\_f-~' 'd.hI . ,.Y:
X _ S b ?, s ;Nlb 47
~ ~ ~ ~ ~ ~,*'- .
FIG 1 recA protein-catalyzed D-loop formation in linear P22DNA (A, B) and M13 RFI DNA (C ). D-loops are indicated by arrows.Reaction conditions in A and B are described in Materials andMethods. In C the reaction contained 2 nmol of M1:3 RFI [:3H]DNAand 650 pmol of M13 single-stranded DNA fragments instead of linearduplex P22 DNA and was incubated as in A and B. In C a super-twisted M13 molecule can be seen.
Binding of recA protein to duplex DNA requires apurine nucleoside triphosphate cofactorThe assimilation reaction described above implies that recAprotein interacts directly with duplex DNA. To test this point,binding of recA protein to 3H-labeled duplex P22 DNA wasmeasured by filter binding. Retention of duplex P22 DNA onnitrocellulose filters required recA protein but not single-stranded DNA and was completely eliminated by treatmentwith detergents (1% Sarkosyl or 1% NaDodSO4) or proteinaseK (data not shown). Efficient retention of these complexes wasobserved with alkali-treated nitrocellulose filters (10), whichdo not bind single-stranded DNA. Therefore, we conclude thatduplex DNA is retained on filters as a consequence of the in-teraction between bound recA protein and the nitrocellu-lose.
Proc. Natl. Acad. Sci. USA 76 (1979)
Proc. Natl. Acad. Sci. USA 76 (1979) 2617
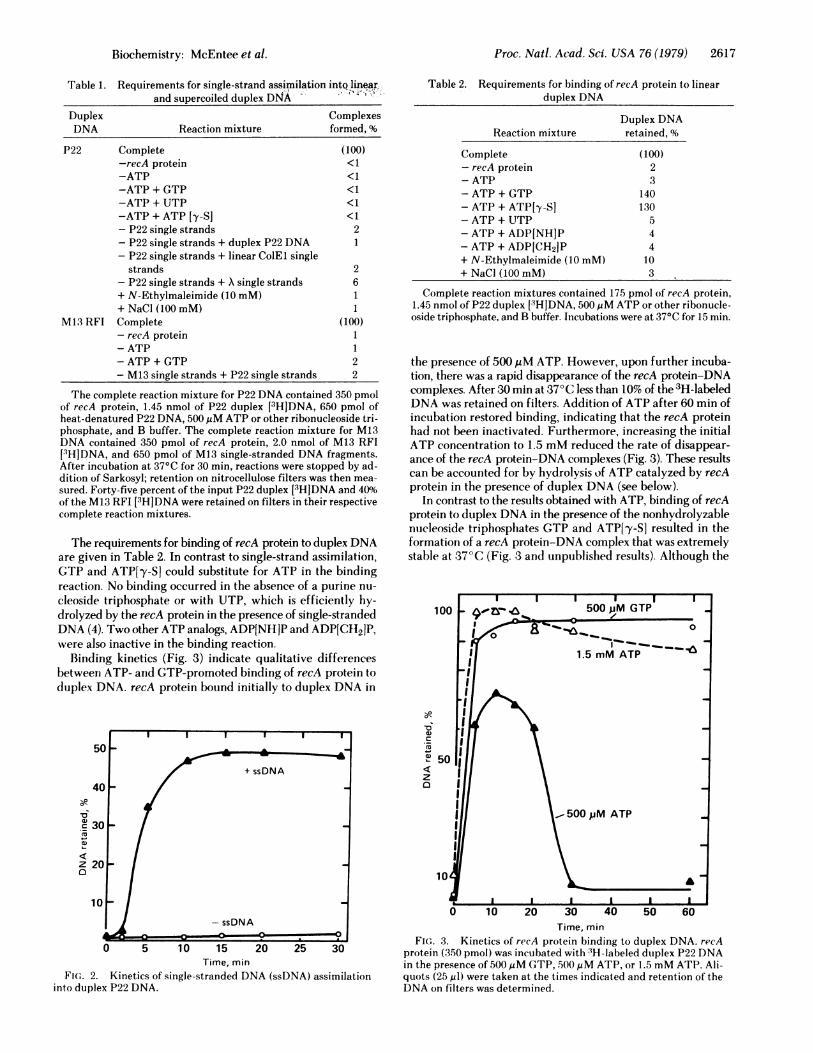
Table 1. Requirements for single-strand assimilation intQ linearrand supercoiled duplex DNA
Duplex ComplexesDNA Reaction mixture formed, %
P22 Complete (100)-recA protein <1-ATP <1-ATP + GTP <1-ATP + UTP <1-ATP + ATP [y-S] <1- P22 single strands 2- P22 single strands + duplex P22 DNA 1- P22 single strands + linear ColEl single
strands 2- P22 single strands + X single strands 6+ N-Ethylmaleimide (10 mM) 1+ NaCl (100 mM) 1
M13 RFI Complete (100)- recA protein 1- ATP 1-ATP + GTP 2- M13 single strands + P22 single strands 2
The complete reaction mixture for P22 DNA contained 350 pmolof recA protein, 1.45 nmol of P22 duplex [3H]DNA, 650 pmol ofheat-denatured P22 DNA, 500MM ATP or other ribonucleoside tri-phosphate, and B buffer. The complete reaction mixture for M13DNA contained 350 pmol of recA protein, 2.0 nmol of M13 RFI[3HJDNA, and 650 pmol of M13 single-stranded DNA fragments.After incubation at 37°C for 30 min, reactions were stopped by ad-dition of Sarkosyl; retention on nitrocellulose filters was then mea-
sured. Forty-rive percent of the input P22 duplex [3H]DNA and 40%of the M13 RFI [3H]DNA were retained on filters in their respectivecomplete reaction mixtures.
The requirements for binding of recA protein to duplex DNAare given in Table 2. In contrast to single-strand assimilation,GTP and ATPLy-SI could substitute for ATP in the bindingreaction. No binding occurred in the absence of a purine nu-
cleoside triphosphate or with UTP, which is efficiently hy-drolyzed by the recA protein in the presence of single-strandedDNA (4). Two other ATP analogs, ADP[NHlP and ADP[CHI21P,were also inactive in the binding reaction.
Binding kinetics (Fig. 3) indicate qualitative differencesbetween ATP- and GTP-promoted binding of recA protein toduplex DNA. recA protein bound initially to duplex DNA in
50
r ~~+ssDNA40
c 30
20
10
- ssDNA
0 5 10 15 20 25 30Time, min
FIG. 2. Kinetics of single-stranded DNA (ssDNA) assimilationinto duplex P22 DNA.
Table 2. Requirements for binding of recA protein to linearduplex DNA
Duplex DNAReaction mixture retained, %
Complete (100)- recA protein 2-ATP 3- ATP + GTP 140-ATP + ATP[y-S] 130-ATP + UTP 5- ATP + ADP[NH]P 4- ATP + ADP[CH2JP 4+ N-Ethylmaleimide (10 mM) 10+ NaCl (100 mM) 3
Complete reaction mixtures contained 175 pmol of recA protein,1.45 nmol of P22 duplex [3HJDNA, 500MM ATP or other ribonucle-oside triphosphate, and B buffer. Incubations were at 37°C for 15 min.
the presence of 500 AM ATP. However, upon further incuba-tion, there was a rapid disappearance of the recA protein-DNAcomplexes. After 30 min at 37°C less than 10% of the 3H-labeledDNA was retained on filters. Addition of ATP after 60 min ofincubation restored binding, indicating that the recA proteinhad not been inactivated. Furthermore, increasing the initialATP concentration to 1.5 mM reduced the rate of disappear-ance of the recA protein-DNA complexes (Fig. 3). These resultscan be accounted for by hydrolysis of ATP catalyzed by recAprotein in the presence of duplex DNA (see below).
In contrast to the results obtained with ATP, binding of recAprotein to duplex DNA in the presence of the nonhydrolyzablenucleoside triphosphates GTP and ATP[y-S] resulted in theformation of a recA protein-DNA complex that was extremelystable at 37°C (Fig. 3 and unpublished results). Although the
I I I I I100 -6500 PiM GTP10~~~~~0t500/ ~~~1.5 mM ATP
C
10 \~0 10 20 30 40 50 60
Time, minFIG. 3. Kinetics of recA protein binding to duplex DNA. recA
protein (350 pmol) was incubated with 3H-labeled duplex P22 DNAin the presence of 500 MM GTP, 500MM ATP, or 1.5 mM ATP. Ali-quots (25 ,l) were taken at the times indicated and retention of theDNA on filters was determined.
Biochemistry: McEntee et al.

2618 Biochemistry: McEntee et al.
initial formation of complexes was inhibited by 0.1 M NaCI(Table 2), once formed in the presence of CTP, the recA pro-
tein-DNA complex was not dissociated by incubation in 0.5 MNaCil at 370C for 60 miin (data not shown).
rem A protein hydrolyzes ATP in the presence of duplexDNAUnder conditions that promote binding of recA protein to du-plex DNA, recA protein catalyzes the hydrolysis of ATP toADP. After 20 min of incubation with duplex P22 DNA and500 jiM ATP, approximately 55% of the ATP was hydrolyzed.Incubation with the same amount of supercoiled ColEI DNAresulted in the hydrolysis of 63% of the ATP. The kinetics ofhydrolysis were linear for 10 min at 370 C. The failure to detecthydrolysis of ATP by recA protein in the presence of duplexDNA ill earlier experiments (4), is explained by the requirementfor high concentrations of recA protein (see below).
Although UTP is hydrolyzed by recA protein in the presence
of single-stranded DNA (4), 1o UTP hydrolysis (less than 3%)was detected when recA protein was incubated with duplexDNA under conditions in which ATP was extensively hydro-lyzed. This result is consistent with the inability of UTP topromote binding of recA protein to duplex DNA (Table 2). Asin the case of single-stranded DNA, recA protein did not hy-Jrolyze GTP in the presence of duplex DNA (unpublished re-
stilts).
Binding of recA protein to duplex DNA and ATPhydrolysis are cooperative
The binding of recA protein to duplex DNA in the presence ofATP or GTP is cooperative (Fig. 4). Cooperativity is also ob-served for the hydrolysis of ATP catalyzed by recA protein inthe presence of duplex DNA (Fig. 4). This is in contrast to thelinear dependence of ATP hydrolysis on recA protein con-
centration in the presence of single-stranded DNA (4). Theassimilation reaction shows a similar cooperative dependenceupon recA protein concentration (Fig. 5), which presumablyreflects the cooperativity of recA protein binding to duplexDNA. Assimilation of single-stranded M13 DNA into super-
100
.75
-6-w
cX 50
z2
0 10 20 30 40 50 60 70recA protein, pmol
80
60
50
40
10'00
30E0
20 cL
10
Flo;. 4. Effect of recA protein concentration on A'rP hydrolysisand duplex DNA binding. The indicated amounts of recA proteinwere incubated with 500 ,uM G'IP for 15 min at 37'C. A 25-fUl aliquotfr-omn each reaction was taken for the filter retention assay. Hydrolysisof ATlI was determined after 30 min of incubation at 37'C with theindicated amounts of re(cA protein.
40
30
-o0
cX 20
z0
10
0 50 100 150 200 250 300 350recA protein, pmol
Fic;. 5. Effect of recA protein concentration on single-strandassimilation. recA protein was incubated with either 1.45 ninol of':'H-labeled P22 1)NA and 6(50 pmol of heat-denatured 1I:22 DNA or
2.0 nmol ofM 1:3 RFI I)NA and 650 pmol of M 1:3 single-stranded DNAfragments for :30 min at. 37 C. The reaction mixtures were treated withSarkosyl and binding to filters was measured.
coiled M 13 RFI also exhibited a cooperative dependence uponrecA protein concentration (Fig. 5).
DISCUSSIONThe purified recA protein promotes assimilation of single-stranded DNA into homologous duplex DNA regions in vitro.The products of this reaction are D-loops in which one strandof the dut)lex is hydrogen bonded to an exogenous single strand.We have detected efficient D-loop formation in both linear andsupercoiled duplex DNA, suggesting that superhelicity is notrequired for strand uptake promoted by the recA protein. Thestability of D-loops in linear DNA. in contrast to superhelicalDNA. is extrenmels sensitive to ionic strength and temperature.At high temperature and low salt, the single strands are rapidlylost due to branch migration (11). In order to detect single-strand assimilation into linear duplex P22 DNA we have usedfiltration conditions (high salt and low temperature) that retardbranch migration. vivo. D-loops might be stabilized in re-
laxed DNA by other binding proteins or nucleases that couldcleave the displaced single strand and block branch tnigra-tion.A model for the single-strand assimilation reaction and its
relationship to single-stranded DNA renaturation catalyzed byrecA protein is depicted in Fig. 6. Binding of recA protein toduplex DNA is Cooperative and reqluires a purine nucleosidetriphosphate (ATP. CTP. ATP[Y-Sl) but does not require itshydrolvsis (A). We presume that the recA protein-DNA in-termetliate that is formed contains partially denatured regions.recA protein might enwind the lduplex directly; alternatively.it might bind to single-stranded regions formed lturing"breathing' of the duplex. destabilize adjacent duplex regions.and promote further binding of recA protein. thus accountingfor the observed oosperativity. ATP. when present as a cofactor
I.I I I I I I
P22 DNA
10,-e M13 RFI DNA
I I ]11//
IIII
I~~~~~~I
I
I I I I I I--
Proc. Natl. Acad. Sci. USA 76 (1979)
I I I I I I I I
ATPase,",..00 M.M.Mm.
I 0Duplex DNA
I binding (GTP)
IIII
- III10II
#0
I a a I a a I a I

Proc. Natl. Acad. Sci. USA 76 (1979) 2619
ADP + Pi
-ADP + Pi*-
ADP + Pi
Feet.. 6. Model for hvbridization reactions catalyzed by recAprotein. recA protein binding to duplex DNA or single-stranded DNAyields protein-DNA complexes I or II, respectively. Binding to duplexDNA results in a partially denatured intermediate (1) that can eitherrenature (B) or hybridize with homologous single-stranded DNA forrecA protein single-stranded DNA complexes (11)1 as in F to produceD)-loops. rec.A protein bound to single-stranded DNA (11) can promoterenaturation of free complementary strands (E) or dissociate froma single strand (D). Reactions E and D require ATP hydrolysis.
in the binding reaction, can be hydrolyzed by the recA pro-
tein-DNA complex (presumably because single-stranded DNAis generated after recA protein binding), leading to loss of thecomplex and release of the recA protein (B).
Binding of recA protein to single-stranded DNA is not co-
operative and does not require a purine nucleoside triphosphate(C) (unpublished results). ATP is hydrolyzed by recA proteinin the absence (D) or presence of a complementary single strand(E). In the latter reaction, recA protein promotes the renatur-ation of the DNA (4). When homologous duplex and single-stranded DNAs are incubated together with recA protein andATP, strand assimilation occurs (F). The hvbridization reactionsin single-strand assimilation (F) and between free single strands(F) are probably due to the same activity of the recA protein,because the recA protein purified from a cold-sensitive recAmutant strain is cold labile for the hybridization of free singlestrands (4) as well as for assimilation of single strands into duplexDNA in vitro (unpublished results).We propose that purine nucleoside triphosphates induce a
conformational change in the recA protein and thus increase
its affinity for DNA. In fact, recA protein-DNA complexesformed in the presence of GTP and ATPky-SJ are extremelystable and are not dissociated at a NaCl concentration (0.5 M)that prevents the initial binding step.When ATP is present in the binding reaction, the formation
of recA protein-DNA complexes is followed by dissociation ofrecA protein and concomitant hydrolysis of ATP. These resultssuggest that ATP hydrolysis restores recA protein to its originalconformation, leading to its release from the DNA due to itslower binding affinity. Sugino et al. (12) have recently sug-gested a similar role for ATP hydrolysis during formation ofsuperhelical DNA by DNA gyrase.The formation of D-loops in vitro catalyzed by recA protein
is consistent with the role of this protein in recombination invivo. recA function is required for general recombination invivo (13), a fact that is most easily explained if the role of therecA gene product is to promote the initial strand crossoverevent required by all recombination pathways. Differences inthe processing of this early intermediate may distinguish thedifferent recombination pathways.
Although single-strand assimilation in vitro requires recAprotein levels that probably exceed the intracellular recA pro-tein concentration in uninduced cells, other proteins might assistin duplex DNA unwinding in vivo. Alternatively, induction ofthe recA protein (14) may be required for single-strand as-similation in vivo.
We are grateful to Janice Chien and Laurien Polder for technicalassistance and to Fran Payne for help in preparing this manuscript.This work was supported by grants from the National Institutes ofHealth (GM06196) and the National Science Foundation (PCM 74-00865). K.M. and G.W. are Fellows of the Jane Coffin Childs MemorialFund for Medical Research.
1. Radding, C. M. (1978) Annu. Rev. Biochem. 47, 847-880.2. Holloman, W. K., Wiegand, R. C., Hoessli, C. & Radding, C. M.
(1975) Proc. Natl. Acad. Sci. lUSA 72,2394-2398.3. Holloman, W. K. & Radding, C. M. (1976) Proc Nat!. Acad. Sci.
USA 73, 3910-3914.4. Weinstock, G. M., McEntee, K. & Lehman, 1. R. (1979) Proc.
Natl. Acad. Sci. USA 76, 126-130.5. Shibata, T., DasGupta, C., Cunningham, R. P. & Radding. C. M.
(1979) Proc. Natl. Acad. Sci. USA 76,1638-1642.6. Botstein, D. (1968) J. Mol. Biol. 34, 621-641.7. Beattie, K. L., Wiegand, R. C. & Radding, C. M. (1977) J. Mol.
Biol. 116,783-803.8. Davis, R. W., Simon, M. & Davidson, N. (1971) Methods Enzy-
mol. 21, 413-428.9. Shalka, A. & Hansen, P. (1972) J. Virol. 9,583--.593.
10. Smolarsky, M. & Tal, M. (1970) Biochimn. Biophys. Acta 199,447-452.
11. Radding, C. M., Beattie, K. L., Holloman, W. K. & Wiegand. R.C. (1977) J. Mol. Biol. 116, 825-839.
12. Suigino, A., Higgins, N. P., Brown, P. 0.. Peebles, C. L. & Coz-zarelli, N. R. (1978) Proc. Natl. Acad. Sci. USA 75, 4838-4842.
1:3. Clark, A. J. (1973) Annu. Rev. Genet. 7, 67-86.14. McEntee, K. (1977) Proc. Natl. Acad. Sci. USA 74, 5275-
5279.
Biochemistry McEntee et al.


















