
Inhibitory effect of plant-originated glycoprotein (27 kDa) on expression of matrix...
8
Journal of Trace Elements in Medicine and Biology 25 (2011) 239–246 Contents lists available at SciVerse ScienceDirect Journal of Trace Elements in Medicine and Biology j ourna l homepage: www.elsevier.de/jtemb Toxicology Inhibitory effect of plant-originated glycoprotein (27 kDa) on expression of matrix metalloproteinase-9 in cadmium chloride-induced BNL CL.2 cells Jin Lee a , Kye-Taek Lim b,∗ a Molecular Biochemistry Laboratory, Biotechnology Research Institute & Center for the Control of Animal Hazards Using Biotechnology (BK21), Chonnam National University, 300 Yongbong-Dong, Gwang-ju 500-757, South Korea b Molecular Biochemistry Laboratory, Biotechnology Research Institute, Chonnam National University, 300 Yongbong-Dong, Gwang-ju 500-757, South Korea a r t i c l e i n f o Article history: Received 10 January 2011 Accepted 16 August 2011 Keywords: Gardenia jasminoides Ellis glycoprotein (27 kDa) Cadmium Arachidonic acid COX-2 MMP-9 a b s t r a c t Cadmium is very harmful to the environment and to human beings because of its long lifetime. The tox- icity of cadmium as an industrial pollutant and a food contaminant, and as one of the major components in cigarette smoke is well known. Cadmium can cause a number of lesions in many organs, such as the kidney, the lung, the liver, the brain, the blood system. However, the mechanism of toxicity of cadmium is not yet clear. Also, it has been well known as human carcinogen which is indirectly caused inflammation- mediated hepatocarcinoma. In the present study it was demonstrated that glycoprotein (27 kDa) isolated from Gardenia jasminoides Ellis (GJE) protects BNL CL.2 cells from expression of inflammation-related fac- tors stimulated by cadmium chloride (10 M). Intracellular ROS and intracellular Ca 2+ using fluorescence, activities of activator protein (AP)-1, cyclooxygenase (COX)-2, matrix metalloproteinase (MMP)-9, and arachidonic acid (AA) using immunoblot analysis or radioactivity were evaluated. The results obtained from this experiment indicated that GJE glycoprotein (100 g/mL) inhibits the production of intracellular ROS, and intracellular Ca 2+ mobilization. Also, it significantly suppressed inflammatory factors [expres- sion of AP-1 (c-Jun and c-Fos), arachidonic acid, COX-2, and MMP-9]. Taken together, these findings suggest that GJE glycoprotein might be used for protection of inflammation caused by cadmium ion as one of natural compounds. © 2011 Elsevier GmbH. All rights reserved. Introduction One of the current burning issues related to human health is that heavy metal contamination exerts serious impact on the liv- ing environment due to discharge of industrial wastes, agricultural and urban sewage. It has been reported that humans and animals are easily exposed to highly toxic metals which are accumulated by organisms through a variety of pathways, including respiration, adsorption and ingestion. Subsequently, they cause much differ- ent kind of diseases such as cancer and immunodysfunction [1]. Cadmium is very harmful toxic metal to the environment and to human beings, and has been classified by the International Agency for Research on Cancer as carcinogen [2]. It has been reported that major sources of cadmium are from food, cigarette smoke, polyvinyl chloride (PVC), and re-charged nickel–cadmium batteries [3]. Cad- mium can cause a number of lesions in many organs, including the kidney, the testis, the lung, the liver, the brain, the bone and the blood. In particular, liver is the main target organ to expose cadmium in chronic and acute period [4]. It has reported that ∗ Corresponding author. Tel.: +82 62 530 2115; fax: +82 62 530 2129. E-mail address: [email protected] (K.-T. Lim). inflammation, adenomas and carcinoma in the liver were caused by at the exposure to cadmium [5,6]. Initiation stage of cancer is closely related to inflammation of certain organs and then generally developed in promo- tion and to transient stage. Hepatocellular carcinoma (HCC) is usually developed in activation or expression of inflamma- tory mediators (including cytokines and chemokines) and other carcinogenensis-related signals in the intracellular level [7]. One possible mechanism of HCC is generation of reactive oxygen species (ROS) and lipid peroxidation damage, alteration of intra- cellular calcium homeostasis caused by cadmium in liver [8]. It has been reported that activator protein (AP)-1 in nucleus is influenced by the cellular redox status and plays a critical role in pro-inflammatory and carcinogenic events [9,10]. Recent finding has suggested that AP-1 is closely related to induction or activation of oxide synthase (iNOS), cyclooxygenase (COX)- 2, and matrix metalloproteinases (MMPs). COX-2 is an enzyme involved in inflammatory processes and rate limiting enzyme in prostaglandin biosynthesis from arachidonic acid (AA). Inappro- priate up-regulation of COX-2 has been frequently observed in various premalignant and malignant tissues [11]. iNOS is another inducible enzyme that causes the overproduction of nitric oxide during inflammation and tumor development [12]. Nitric oxide 0946-672X/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.jtemb.2011.08.142
Transcript of Inhibitory effect of plant-originated glycoprotein (27 kDa) on expression of matrix...

T
Im
Ja
Yb
a
ARA
KG(CACM
I
tiaabaeChfmcmttc
0d
Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246
Contents lists available at SciVerse ScienceDirect
Journal of Trace Elements in Medicine and Biology
j ourna l homepage: www.elsev ier .de / j temb
oxicology
nhibitory effect of plant-originated glycoprotein (27 kDa) on expression ofatrix metalloproteinase-9 in cadmium chloride-induced BNL CL.2 cells
in Leea, Kye-Taek Limb,∗
Molecular Biochemistry Laboratory, Biotechnology Research Institute & Center for the Control of Animal Hazards Using Biotechnology (BK21), Chonnam National University, 300ongbong-Dong, Gwang-ju 500-757, South KoreaMolecular Biochemistry Laboratory, Biotechnology Research Institute, Chonnam National University, 300 Yongbong-Dong, Gwang-ju 500-757, South Korea
r t i c l e i n f o
rticle history:eceived 10 January 2011ccepted 16 August 2011
eywords:ardenia jasminoides Ellis glycoprotein
27 kDa)admiumrachidonic acidOX-2
a b s t r a c t
Cadmium is very harmful to the environment and to human beings because of its long lifetime. The tox-icity of cadmium as an industrial pollutant and a food contaminant, and as one of the major componentsin cigarette smoke is well known. Cadmium can cause a number of lesions in many organs, such as thekidney, the lung, the liver, the brain, the blood system. However, the mechanism of toxicity of cadmium isnot yet clear. Also, it has been well known as human carcinogen which is indirectly caused inflammation-mediated hepatocarcinoma. In the present study it was demonstrated that glycoprotein (27 kDa) isolatedfrom Gardenia jasminoides Ellis (GJE) protects BNL CL.2 cells from expression of inflammation-related fac-tors stimulated by cadmium chloride (10 �M). Intracellular ROS and intracellular Ca2+ using fluorescence,activities of activator protein (AP)-1, cyclooxygenase (COX)-2, matrix metalloproteinase (MMP)-9, and
MP-9 arachidonic acid (AA) using immunoblot analysis or radioactivity were evaluated. The results obtainedfrom this experiment indicated that GJE glycoprotein (100 �g/mL) inhibits the production of intracellularROS, and intracellular Ca2+ mobilization. Also, it significantly suppressed inflammatory factors [expres-sion of AP-1 (c-Jun and c-Fos), arachidonic acid, COX-2, and MMP-9]. Taken together, these findingssuggest that GJE glycoprotein might be used for protection of inflammation caused by cadmium ion asone of natural compounds.
© 2011 Elsevier GmbH. All rights reserved.
ntroduction
One of the current burning issues related to human health ishat heavy metal contamination exerts serious impact on the liv-ng environment due to discharge of industrial wastes, agriculturalnd urban sewage. It has been reported that humans and animalsre easily exposed to highly toxic metals which are accumulatedy organisms through a variety of pathways, including respiration,dsorption and ingestion. Subsequently, they cause much differ-nt kind of diseases such as cancer and immunodysfunction [1].admium is very harmful toxic metal to the environment and touman beings, and has been classified by the International Agency
or Research on Cancer as carcinogen [2]. It has been reported thatajor sources of cadmium are from food, cigarette smoke, polyvinyl
hloride (PVC), and re-charged nickel–cadmium batteries [3]. Cad-ium can cause a number of lesions in many organs, including
he kidney, the testis, the lung, the liver, the brain, the bone andhe blood. In particular, liver is the main target organ to exposeadmium in chronic and acute period [4]. It has reported that
∗ Corresponding author. Tel.: +82 62 530 2115; fax: +82 62 530 2129.E-mail address: [email protected] (K.-T. Lim).
inflammation, adenomas and carcinoma in the liver were causedby at the exposure to cadmium [5,6].
Initiation stage of cancer is closely related to inflammationof certain organs and then generally developed in promo-tion and to transient stage. Hepatocellular carcinoma (HCC)is usually developed in activation or expression of inflamma-tory mediators (including cytokines and chemokines) and othercarcinogenensis-related signals in the intracellular level [7]. Onepossible mechanism of HCC is generation of reactive oxygenspecies (ROS) and lipid peroxidation damage, alteration of intra-cellular calcium homeostasis caused by cadmium in liver [8].It has been reported that activator protein (AP)-1 in nucleusis influenced by the cellular redox status and plays a criticalrole in pro-inflammatory and carcinogenic events [9,10]. Recentfinding has suggested that AP-1 is closely related to inductionor activation of oxide synthase (iNOS), cyclooxygenase (COX)-2, and matrix metalloproteinases (MMPs). COX-2 is an enzymeinvolved in inflammatory processes and rate limiting enzyme inprostaglandin biosynthesis from arachidonic acid (AA). Inappro-priate up-regulation of COX-2 has been frequently observed invarious premalignant and malignant tissues [11]. iNOS is anotherinducible enzyme that causes the overproduction of nitric oxideduring inflammation and tumor development [12]. Nitric oxide
946-672X/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.jtemb.2011.08.142

240 J. Lee, K.-T. Lim / Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246
has been implicated in initiation, promotional stage of neoplas-mic transformation [13]. Among the MMPs, MMP-9 well knownas gelatinase B, 92 kDa gelatinase/type IV collagenase has a broadrange of substrate specificity for native collagens including type IV,V VII, and X as well as gelatin, proteoglycans and elastin. MMP-9 is secreted as a 92 kDa proenzyme and can be activated in vitroby organomercurials, trypsin, and �-chymotrypsin and in vivo bycatepsin G. MMP-9 is closely related to hepatocellular carcinomas.
It is a good strategy to use medical plants for the preven-tion or healing of several different immune dysfunctional diseasescaused by toxic heavy metal like cadmium, because most natu-ral compounds (like phytoglycoprotein) have less cytotoxicity thanartificial synthetic compounds such as drugs. Gardenia jasminoidesEllis (GJE) has been used for a long time to cure febrile diseases,jaundice, acute conjunctivitis, epistaxis, haematemesis, pyogenicinfections and ulcers of the skin as traditional medicine in Korea[14]. Recently, we isolated a glycoprotein with an approximatemolecular mass of 27 kDa from GJE. The glycoprotein has a car-bohydrate and protein content of 57.65% and 42.35%, respectively.In our previous study, the GJE glycoprotein was shown to possessa strong anti-oxidant-activity against, anti-inflammatory activity,and hepatoprotective activity [15,16]. Thus, we assume that GJEglycoprotein can probably prevent the liver cell damage cause bycadmium ion. However, it has not been reported that GJE glyco-protein prevents hepatocytes from the inflammation occurred bycadmium chloride.
Therefore, the aim of this study was to know whether GJE gly-coprotein prevents BNL CL.2 cells from inflammation caused bycadmium chloride. For this purpose, intracellular ROS, intracellu-lar Ca2+ mobilization, AP-1 activity, arachidonic acid, COX-2 andMMP-9 were evaluated in cadmium chloride-treated BNL CL.2 cells.
Materials and methods
Chemicals
All the plastic materials were purchased from Falcon Labware(Becton-Dickinson, Franklin Lakes, NJ). Penicillin G and strepto-mycin were obtained from Sigma (St Louis, MO, USA). Dulbecco’smodified Eagle medium (DMEM) and fetal bovine serum (FBS) werepurchased from Gibco BRL (Grand Island, NY, USA). Other chemicalsand reagents were of the highest analytical grade available.
Preparation of GJE glycoprotein
GJE glycoprotein was isolated and purified from Gardenia jasmi-noides Ellis (GJE) with slight Modifications, essentially as describedpreviously [16]. Samples of GJE were obtained in October 2003from the Naju traditional market in the Chonnam province, SouthKorea. They were identified by Dr. H.T. Lim (Chonnam National Uni-versity) and the fruits (8.20 kg) were chopped into small piecesand soaked in ethanol (7 l) for several months in a dark base-ment. The ethanol extracts were subsequently filtered throughWhatman filter paper (No. 2) and centrifuged at 8000 × g for10 min. The collected supernatants were concentrated with a rotaryevaporator (B465; Buchi, Switzerland) and the dried matter was380.05 g. The dried-crude extracts (5 g) were resolved in distilledand subjected to concanavalin sepharose 4B affinity chromatogra-phy (24–45 mm, Sigma, C9017). It was eluted with 0.5 M methylá-d-glucopyranoside containing 0.5 M NaCl at pH 7.4. The elutedsolution was dialyzed with a dialysis membrane (Spectra/por,MWCO 6000-8000, CAL., USA) against 20 mM Tris–Cl (pH 7.4) at4 ◦C overnight and lyophilized to yield 2.5 g (1.49%). The final purityof GJE glycoprotein was over 98 ± 2% and consisted of carbohy-drate (57.65%) and protein moiety (42.35%). The quantity of GJE
glycoprotein yielded was 5.68 mg (0.07%) after lyophilization andstored at 4 ◦C until use.
Cell culture
BNL CL.2 cells (murine embryonic liver cell) were incu-bated in DMEM containing 10% FBS, 100 U/mL of penicillin, and100 �g/mL of streptomycin at 37 ◦C and atmosphere containing5% CO2. The medium was renewed two times per week. The cells(1 × 106 cells/ml) were dispended into 96-well flat bottom platesor 6-well plates.
Measurement of intracellular ROS
The intracellular ROS were measured using a non-fluorescentdye, 2′,7′-dichlorodihydrofluorescein (H2DCF-DA), which is a mem-brane permeable fluorigenic tracer that is oxidized by variousspecies of ROS [17]. The dye is deacetylated by intracellu-lar esterases to non-fluorescent 2′,7′-dichlorohydrofluorescein(DCFH), which is oxidized to the fluorescent compound 2′,7′-dichlorofluorescein (DCF) by ROS. The cells were pre-incubatedwith 10 �M H2DCF-DA for 30 min at 37 ◦C, and then washed twicewith PBS to remove the extracellular H2DCF-DA. After that, thecells were treated with cadmium chloride (10 �M) for the indicatedtime, or co-treated with GJE glycoprotein (25, 50 and 100 �g/ml) inthe presence of 10 �M cadmium chloride for 6 h. Finally, the inten-sity of fluorescence was measured at an excitation of 485 nm andan emission of 530 nm using a fluorescence microplate reader (DualScanning SPECTRAmax, Molecular Devices Corporation, Sunnyvale,CA, USA). The values were calculated as the relative intensity of DCFfluorescence, compared with the control.
Detection of intracellular Ca2+ mobilization
Determination of intracellular Ca2+ was performed as describedpreviously [18]. Briefly, cells were rinsed with bath solution(140 mM NaCl, 5 mM KCl, 1 mM CaCl2, 0.5 mM MgCl2, 10 mM glu-cose, 5.5 mM HEPES, pH 7.5) and then preincubated in bath solutioncontaining 1 �M Fura-2/AM (Molecular probes, Eugene, OR, USA)with 5% CO2 at 37 ◦C for 45 min with gentle shaking. After shaking,cells were treated with GJE glycoprotein (25, 50 and 100 �g/ml),cadmium chloride (10 �M) and A23187 (1 �M) as positive control.The amount of intracellular Ca2+ was measured by using fluo-rescence device. The intensity of fluorescence was measured atan emission wavelength of 510 nm and excitation wavelength of340 nm and 380 nm using a fluoresencent microplate reader (DualScanning SPECTRAmax, Molecular Devices Corporation, Sunnyvale,CA, USA). The fluorescence ratio (340/380) was calculated as anindicator of intracellular Ca2+ intensity.
Assay of [3H] arachidonic acid release
The [3H] arachidonic acid (AA) release experiments were per-formed using a modification of the method reported by accordingto the method of Xing et al. [19]. BNL CL.2 cells were incubatedfor 24 h in DMEM containing 0.5 �Ci [3H]AA/ml. The cells werethen washed three times with DMEM and incubated at 37 ◦C for0–24 h in incubation medium. At the end of the incubation period,the culture medium was transferred to ice-cold tubes containing100 �l of 55 mM EGTA and EDTA (final concentration, 5 mM each).The culture medium was then centrifuged at 12,000 × g to elimi-nate the cell debris. The level of radioactivity in the supernatantwas determined by placing samples in scintillation vials contain-ing scintillation fluid and measuring the radioactivity using a liquidscintillation counter (LS 6500, Beckman Instruments, Fullerton, CA).

J. Lee, K.-T. Lim / Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246 241
Preparation of cell extracts
Preparation of cell extracts was basically as described else-where [20]. Briefly, the cells were treated with cadmium chloride(10 �M) in the presence of GJE glycoprotein (25, 50 and 100 �g/ml),then cells were washed in 1 ml of ice-cold PBS after the mediumwas removed, and scraped into 300 �l of ice-cold buffer C (20 mMHEPES, pH 7.9, 0.4 M NaCl, 0.2 mM EDTA, 1 mM DTT, 1 mM PMSF,1.5 mM MgCl2, 0.5% NP-40, 25% glycerol) containing a phosphataseinhibitor cocktail. For lysis, the cells were freeze-thawed, shaken for30 min at 4 ◦C, and centrifuged at 14,000 × g for 30 min. The super-natant was designated as a whole extract for immunoblotting ofCOX-2, iNOS and MMP-9.
To prepare nuclear extracts for immunoblotting of AP-1 (c-Jun and c-Fos) after cell activation for the times indicated, cellswere resuspended in 400 �l of Ice-cold hypotonic buffer D (10 mMHEPES, pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mMDTT, 0.5 mM PMSF, 1.5 mM MgCl2, 0.5% NP-40), left on 15 minand then centrifuged at 3000 × g for 5 min at 4 ◦C. Pelleted nucleiwere gently resuspended in 150 �L of NE buffer E (20 mM HEPES,pH 7.5, 0.4 M NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 1 mMPMSF, 1.5 mM MgCl2, 0.5% NP-40, 25% glycerol) containing a pro-tease inhibitor cocktail (Boehringer, Mannheim, Germany) andcentrifuged at 12,000 × g for 15 min at 4 ◦C. The amount of proteinwas measured by the method of Lowry et al. [21], and the cellularproteins were stored at −70 ◦C prior to use.
Western blotting
Cellular proteins were separated in a 10% polyacrylamide mini-gel at 100 V for 2 h at room temperature using a mini-protein IIelectrophoresis cell (Bio-Rad). After electrophoresis, the proteinswere transferred onto nitrocellulose membranes (Millipore, Bed-ford, MA, USA). The membranes were incubated for 1 h at roomtemperature in TBS-T solution (10 mM Tris–HCl, pH 7.6, 150 mMNaCl, and 0.1% (v/v) Tween-20) containing 5% non-fat dry milk. Themembranes were subsequently incubated for 2 h at room tempera-ture with primary antibodies (COX-2, iNOS, AP-1 c-Jun, AP-1 c-Fosand MMP-9) in TBS-T solution. After three times washes with TBS-T, the membranes were incubated for 1 h at room temperature withhorseradish peroxidase-conjugated goat anti-mouse IgG and anti-rabbit IgG (1:10,000; Cell Signaling Technology, MA, USA) in TBS-Tsolution. The resulting protein bands were visualized by enhancedchemiluminescence (Amersham Pharmacia Biotech, England, UK)The results of Western blot assay were calculated as relative inten-sity using Scion imaging software (Scion Image Beta 4.02, Frederick,MD, USA).
Gelatin zymography
BNL CL.2 cells were incubated in serum-free DMEM in the pres-ence of cadmium chloride with or without GJE glycoprotein for agiven time, and the conditioned medium was collected in samples.The unboiled samples were separated via electrophoresis on 10%sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) containing 0.1% gelatin. After electrophoresis, the gels werewashed twice in washing buffer (2.5% Triton X-100 in dH2O) atroom temperature for 30 min to remove SDS, and then incubatedin a reaction buffer [40 mM Tris–HCl (pH 8.0), 10 mM CaCl2, 0.01%NaN3, and 1 mM ZnCl2] at 37 ◦C for 16 h to allow for proteoly-sis of the gelatin substrate. Bands corresponding to the activitywere visualized by negative staining using Coomassie Brilliant blueR-250 (Bio-Rad Laboratories, Richmond, CA) and the molecularweight was estimated by reference to pre-stained SDS-PAGE mark-ers.
Fig. 1. Inhibitory effect of GJE glycoprotein on amount of ROS in cadmium chloride-exposed BNL CL.2 cells. BNL CL.2 cells were treated with cadmium chloride (10 �M)in the presence or absence of GJE glycoprotein (25, 50 and 100 �g/ml). The results arepresented as relative intensity of fluorescence based on monitoring of dichlorodi-hydrofluorescein (DCF) using a fluorescence microplate reader (Dual ScanningSPECTRAmax, Molecular Devices Corporation, Sunnyvale, CA, USA). All data wererepresented the means ± S.D. from experiments run in triplicate (n = 3). # Signifi-cant difference between control cells and those exposed to cadmium chloride alone
p < 0.05. * Significant difference between cells exposed to cadmium chloride aloneand those treated with GJE glycoprotein treatment in the presence of cadmiumchloride, P < 0.05.Invasion assay
Cell invasion was evaluated using 24-well BD biocoat Matrigelinvasion chambers according to method of Pan et al. [22]. A sus-pension of cells (2 × 104 cells/100 �l) in non-serum medium wasplaced in the matrigel-coated filters. The wells below were filledwith 0.75 ml medium supplemented with 5% FBS. After 22 h incu-bation in a humidified atmosphere with 5% CO2 in air at 37 ◦C,the non-migrated cells were removed by cotton swab and themigrated cells on the lower side of the filters were fixed andstained with Diff-Quick staining kit. Invaded cells were quantifiedby counting the stained cells in five random fields per membrane at200× magnification.
Statistical analysis
All experiments were carried out in triplicates (n = 3), anddata were expressed as means ± S.D. One-way analysis of variance(ANOVA) and Duncan test were carried out to determine signifi-cant differences of multiple comparisons (Statistical Package of theSocial Sciences program, version 11.0).
Results
Inhibitory effect of GJE glycoprotein on amount of ROS incadmium chloride-exposed BNL CL.2 cells
As shown in Fig. 1, when the BNL CL.2 cells were exposed tocadmium chloride (10 �M) for 6 h, the relative amounts of intra-cellular ROS were increased, compared to the control. However,when the BNL CL.2 cells were treated with GJE glycoprotein (25, 50and 100 �g/ml) in the presence of cadmium chloride, the intracellu-lar ROS production was significantly reduced in a dose dependentmanner, compared to the value obtained in the presence of cad-mium chloride alone. For example, the value of intracellular ROSwas reduced by 0.26-, 0.39- and 0.41-fold at 25, 50 and 100 �g/mlGJE glycoprotein in presence of cadmium chloride (10 �M) respec-tively.
Inhibitory effect of GJE glycoprotein on mobilization of
intracellular Ca2+ in cadmium chloride-exposed BNL CL.2 cellsWith reference to Ca2+ mobilization in the cells, when the BNLCL.2 cells were exposed to cadmium chloride (10 �M) alone, the
242 J. Lee, K.-T. Lim / Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246
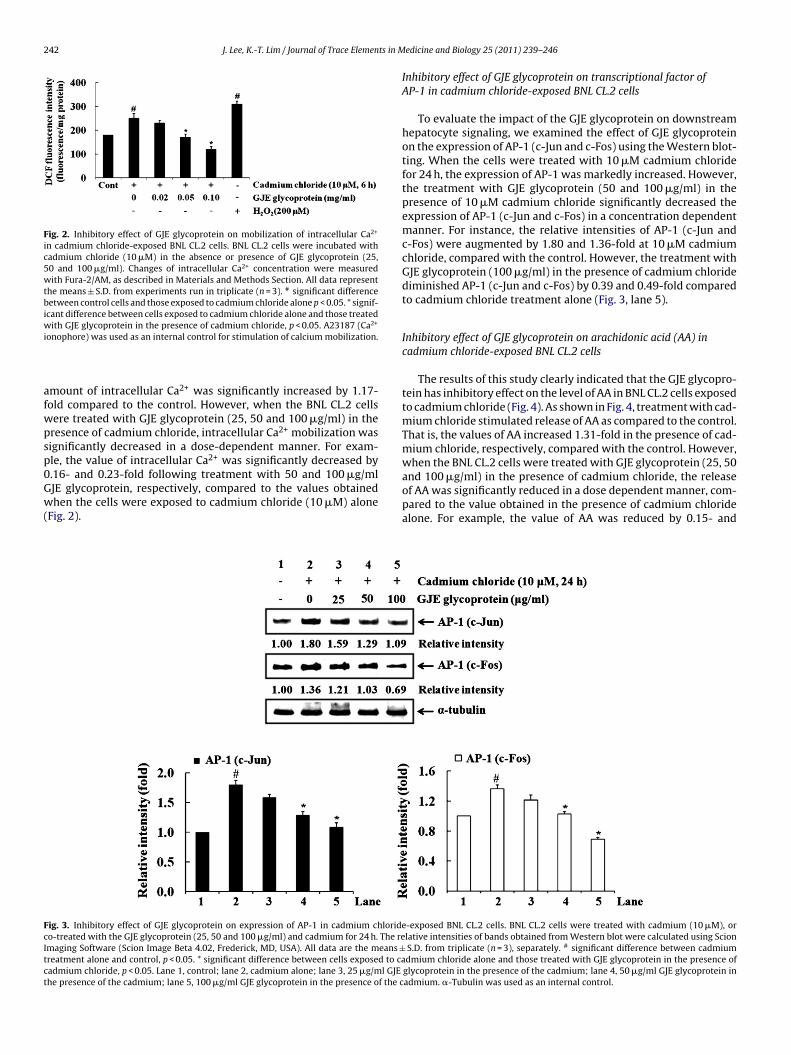
Fig. 2. Inhibitory effect of GJE glycoprotein on mobilization of intracellular Ca2+
in cadmium chloride-exposed BNL CL.2 cells. BNL CL.2 cells were incubated withcadmium chloride (10 �M) in the absence or presence of GJE glycoprotein (25,50 and 100 �g/ml). Changes of intracellular Ca2+ concentration were measuredwith Fura-2/AM, as described in Materials and Methods Section. All data representthe means ± S.D. from experiments run in triplicate (n = 3). # significant differencebetween control cells and those exposed to cadmium chloride alone p < 0.05. * signif-icant difference between cells exposed to cadmium chloride alone and those treatedwith GJE glycoprotein in the presence of cadmium chloride, p < 0.05. A23187 (Ca2+
i
afwpsp0Gw(
Inhibitory effect of GJE glycoprotein on transcriptional factor ofAP-1 in cadmium chloride-exposed BNL CL.2 cells
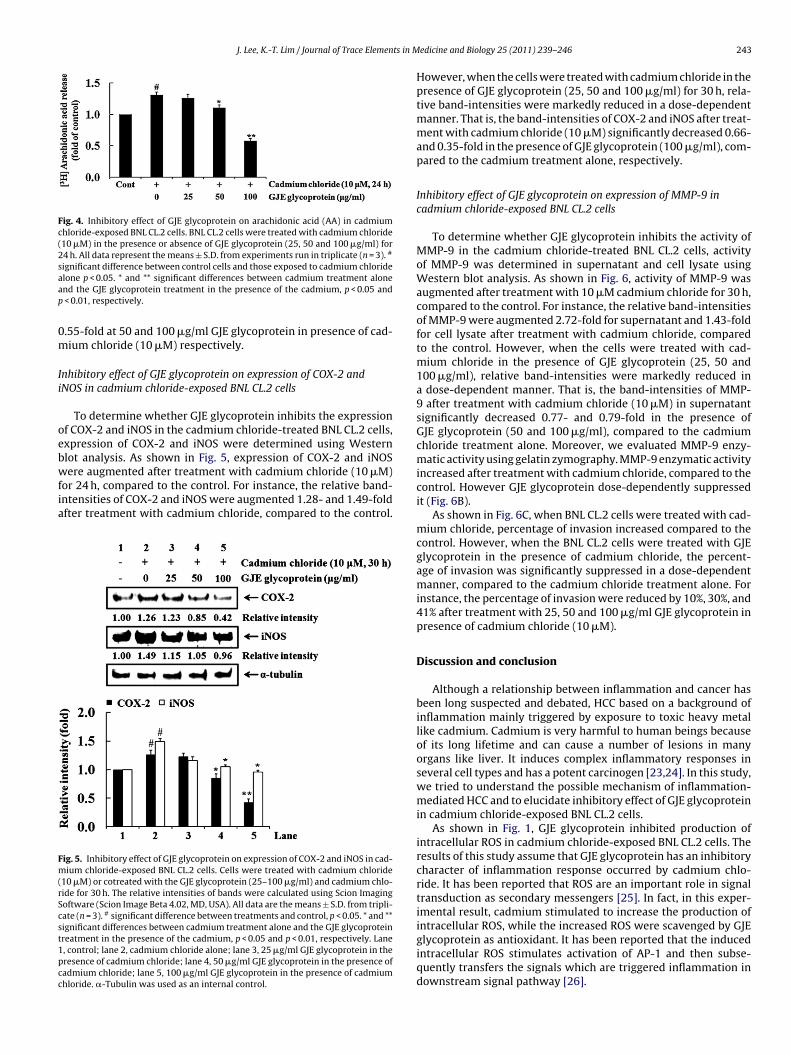
To evaluate the impact of the GJE glycoprotein on downstreamhepatocyte signaling, we examined the effect of GJE glycoproteinon the expression of AP-1 (c-Jun and c-Fos) using the Western blot-ting. When the cells were treated with 10 �M cadmium chloridefor 24 h, the expression of AP-1 was markedly increased. However,the treatment with GJE glycoprotein (50 and 100 �g/ml) in thepresence of 10 �M cadmium chloride significantly decreased theexpression of AP-1 (c-Jun and c-Fos) in a concentration dependentmanner. For instance, the relative intensities of AP-1 (c-Jun andc-Fos) were augmented by 1.80 and 1.36-fold at 10 �M cadmiumchloride, compared with the control. However, the treatment withGJE glycoprotein (100 �g/ml) in the presence of cadmium chloridediminished AP-1 (c-Jun and c-Fos) by 0.39 and 0.49-fold comparedto cadmium chloride treatment alone (Fig. 3, lane 5).
Inhibitory effect of GJE glycoprotein on arachidonic acid (AA) incadmium chloride-exposed BNL CL.2 cells
The results of this study clearly indicated that the GJE glycopro-tein has inhibitory effect on the level of AA in BNL CL.2 cells exposedto cadmium chloride (Fig. 4). As shown in Fig. 4, treatment with cad-mium chloride stimulated release of AA as compared to the control.That is, the values of AA increased 1.31-fold in the presence of cad-mium chloride, respectively, compared with the control. However,when the BNL CL.2 cells were treated with GJE glycoprotein (25, 50and 100 �g/ml) in the presence of cadmium chloride, the releaseof AA was significantly reduced in a dose dependent manner, com-
FcItct
onophore) was used as an internal control for stimulation of calcium mobilization.
mount of intracellular Ca2+ was significantly increased by 1.17-old compared to the control. However, when the BNL CL.2 cellsere treated with GJE glycoprotein (25, 50 and 100 �g/ml) in theresence of cadmium chloride, intracellular Ca2+ mobilization wasignificantly decreased in a dose-dependent manner. For exam-le, the value of intracellular Ca2+ was significantly decreased by.16- and 0.23-fold following treatment with 50 and 100 �g/mlJE glycoprotein, respectively, compared to the values obtained
hen the cells were exposed to cadmium chloride (10 �M) aloneFig. 2).pared to the value obtained in the presence of cadmium chloridealone. For example, the value of AA was reduced by 0.15- and
ig. 3. Inhibitory effect of GJE glycoprotein on expression of AP-1 in cadmium chloride-exposed BNL CL.2 cells. BNL CL.2 cells were treated with cadmium (10 �M), oro-treated with the GJE glycoprotein (25, 50 and 100 �g/ml) and cadmium for 24 h. The relative intensities of bands obtained from Western blot were calculated using Scionmaging Software (Scion Image Beta 4.02, Frederick, MD, USA). All data are the means ± S.D. from triplicate (n = 3), separately. # significant difference between cadmiumreatment alone and control, p < 0.05. * significant difference between cells exposed to cadmium chloride alone and those treated with GJE glycoprotein in the presence ofadmium chloride, p < 0.05. Lane 1, control; lane 2, cadmium alone; lane 3, 25 �g/ml GJE glycoprotein in the presence of the cadmium; lane 4, 50 �g/ml GJE glycoprotein inhe presence of the cadmium; lane 5, 100 �g/ml GJE glycoprotein in the presence of the cadmium. �-Tubulin was used as an internal control.
J. Lee, K.-T. Lim / Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246 243
Fig. 4. Inhibitory effect of GJE glycoprotein on arachidonic acid (AA) in cadmiumchloride-exposed BNL CL.2 cells. BNL CL.2 cells were treated with cadmium chloride(10 �M) in the presence or absence of GJE glycoprotein (25, 50 and 100 �g/ml) for24 h. All data represent the means ± S.D. from experiments run in triplicate (n = 3). #
significant difference between control cells and those exposed to cadmium chlorideaap
0m
Ii
oebwfia
Fm(rScst1pcc
However, when the cells were treated with cadmium chloride in thepresence of GJE glycoprotein (25, 50 and 100 �g/ml) for 30 h, rela-tive band-intensities were markedly reduced in a dose-dependentmanner. That is, the band-intensities of COX-2 and iNOS after treat-ment with cadmium chloride (10 �M) significantly decreased 0.66-and 0.35-fold in the presence of GJE glycoprotein (100 �g/ml), com-pared to the cadmium treatment alone, respectively.
Inhibitory effect of GJE glycoprotein on expression of MMP-9 incadmium chloride-exposed BNL CL.2 cells
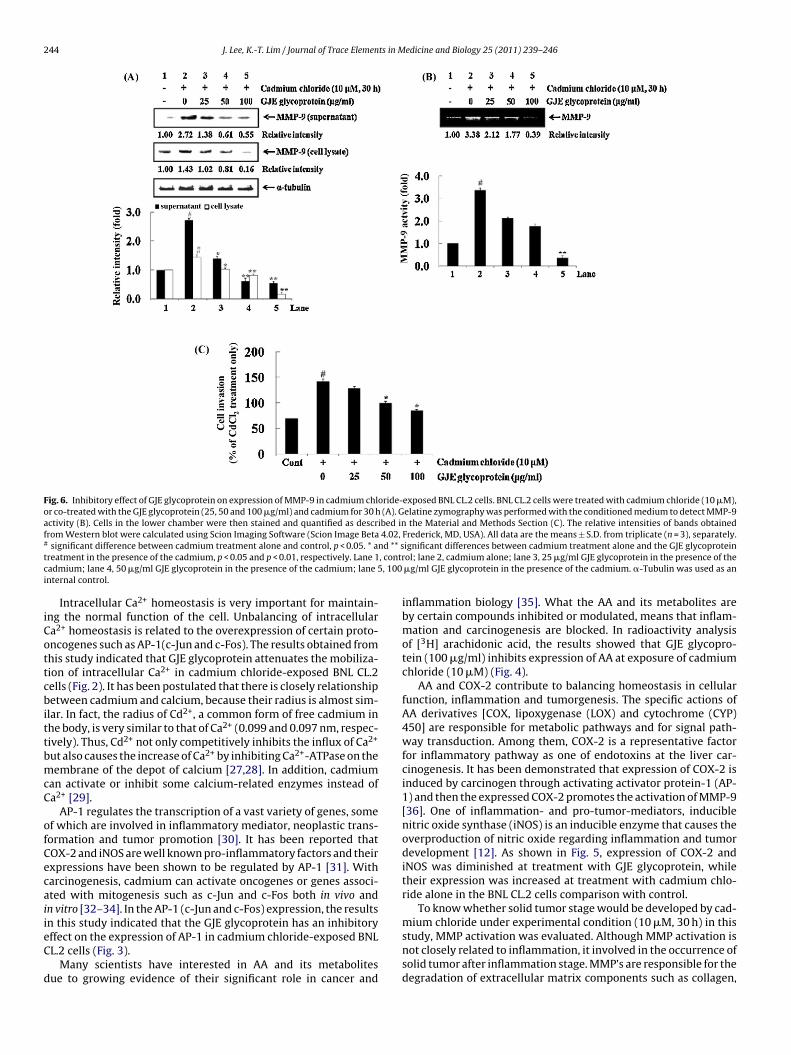
To determine whether GJE glycoprotein inhibits the activity ofMMP-9 in the cadmium chloride-treated BNL CL.2 cells, activityof MMP-9 was determined in supernatant and cell lysate usingWestern blot analysis. As shown in Fig. 6, activity of MMP-9 wasaugmented after treatment with 10 �M cadmium chloride for 30 h,compared to the control. For instance, the relative band-intensitiesof MMP-9 were augmented 2.72-fold for supernatant and 1.43-foldfor cell lysate after treatment with cadmium chloride, comparedto the control. However, when the cells were treated with cad-mium chloride in the presence of GJE glycoprotein (25, 50 and100 �g/ml), relative band-intensities were markedly reduced ina dose-dependent manner. That is, the band-intensities of MMP-9 after treatment with cadmium chloride (10 �M) in supernatantsignificantly decreased 0.77- and 0.79-fold in the presence ofGJE glycoprotein (50 and 100 �g/ml), compared to the cadmiumchloride treatment alone. Moreover, we evaluated MMP-9 enzy-matic activity using gelatin zymography. MMP-9 enzymatic activity
lone p < 0.05. * and ** significant differences between cadmium treatment alonend the GJE glycoprotein treatment in the presence of the cadmium, p < 0.05 and
< 0.01, respectively.
.55-fold at 50 and 100 �g/ml GJE glycoprotein in presence of cad-ium chloride (10 �M) respectively.
nhibitory effect of GJE glycoprotein on expression of COX-2 andNOS in cadmium chloride-exposed BNL CL.2 cells
To determine whether GJE glycoprotein inhibits the expressionf COX-2 and iNOS in the cadmium chloride-treated BNL CL.2 cells,xpression of COX-2 and iNOS were determined using Westernlot analysis. As shown in Fig. 5, expression of COX-2 and iNOS
ere augmented after treatment with cadmium chloride (10 �M)or 24 h, compared to the control. For instance, the relative band-ntensities of COX-2 and iNOS were augmented 1.28- and 1.49-foldfter treatment with cadmium chloride, compared to the control.
ig. 5. Inhibitory effect of GJE glycoprotein on expression of COX-2 and iNOS in cad-ium chloride-exposed BNL CL.2 cells. Cells were treated with cadmium chloride
10 �M) or cotreated with the GJE glycoprotein (25–100 �g/ml) and cadmium chlo-ide for 30 h. The relative intensities of bands were calculated using Scion Imagingoftware (Scion Image Beta 4.02, MD, USA). All data are the means ± S.D. from tripli-ate (n = 3). # significant difference between treatments and control, p < 0.05. * and **ignificant differences between cadmium treatment alone and the GJE glycoproteinreatment in the presence of the cadmium, p < 0.05 and p < 0.01, respectively. Lane, control; lane 2, cadmium chloride alone; lane 3, 25 �g/ml GJE glycoprotein in theresence of cadmium chloride; lane 4, 50 �g/ml GJE glycoprotein in the presence ofadmium chloride; lane 5, 100 �g/ml GJE glycoprotein in the presence of cadmiumhloride. �-Tubulin was used as an internal control.
increased after treatment with cadmium chloride, compared to thecontrol. However GJE glycoprotein dose-dependently suppressedit (Fig. 6B).
As shown in Fig. 6C, when BNL CL.2 cells were treated with cad-mium chloride, percentage of invasion increased compared to thecontrol. However, when the BNL CL.2 cells were treated with GJEglycoprotein in the presence of cadmium chloride, the percent-age of invasion was significantly suppressed in a dose-dependentmanner, compared to the cadmium chloride treatment alone. Forinstance, the percentage of invasion were reduced by 10%, 30%, and41% after treatment with 25, 50 and 100 �g/ml GJE glycoprotein inpresence of cadmium chloride (10 �M).
Discussion and conclusion
Although a relationship between inflammation and cancer hasbeen long suspected and debated, HCC based on a background ofinflammation mainly triggered by exposure to toxic heavy metallike cadmium. Cadmium is very harmful to human beings becauseof its long lifetime and can cause a number of lesions in manyorgans like liver. It induces complex inflammatory responses inseveral cell types and has a potent carcinogen [23,24]. In this study,we tried to understand the possible mechanism of inflammation-mediated HCC and to elucidate inhibitory effect of GJE glycoproteinin cadmium chloride-exposed BNL CL.2 cells.
As shown in Fig. 1, GJE glycoprotein inhibited production ofintracellular ROS in cadmium chloride-exposed BNL CL.2 cells. Theresults of this study assume that GJE glycoprotein has an inhibitorycharacter of inflammation response occurred by cadmium chlo-ride. It has been reported that ROS are an important role in signaltransduction as secondary messengers [25]. In fact, in this exper-imental result, cadmium stimulated to increase the production ofintracellular ROS, while the increased ROS were scavenged by GJEglycoprotein as antioxidant. It has been reported that the inducedintracellular ROS stimulates activation of AP-1 and then subse-quently transfers the signals which are triggered inflammation indownstream signal pathway [26].
244 J. Lee, K.-T. Lim / Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246
Fig. 6. Inhibitory effect of GJE glycoprotein on expression of MMP-9 in cadmium chloride-exposed BNL CL.2 cells. BNL CL.2 cells were treated with cadmium chloride (10 �M),or co-treated with the GJE glycoprotein (25, 50 and 100 �g/ml) and cadmium for 30 h (A). Gelatine zymography was performed with the conditioned medium to detect MMP-9activity (B). Cells in the lower chamber were then stained and quantified as described in the Material and Methods Section (C). The relative intensities of bands obtainedfrom Western blot were calculated using Scion Imaging Software (Scion Image Beta 4.02, Frederick, MD, USA). All data are the means ± S.D. from triplicate (n = 3), separately.# significant difference between cadmium treatment alone and control, p < 0.05. * and ** significant differences between cadmium treatment alone and the GJE glycoproteintreatment in the presence of the cadmium, p < 0.05 and p < 0.01, respectively. Lane 1, control; lane 2, cadmium alone; lane 3, 25 �g/ml GJE glycoprotein in the presence of thecadmium; lane 4, 50 �g/ml GJE glycoprotein in the presence of the cadmium; lane 5, 100 �g/ml GJE glycoprotein in the presence of the cadmium. �-Tubulin was used as aninternal control.
Intracellular Ca2+ homeostasis is very important for maintain-ing the normal function of the cell. Unbalancing of intracellularCa2+ homeostasis is related to the overexpression of certain proto-oncogenes such as AP-1(c-Jun and c-Fos). The results obtained fromthis study indicated that GJE glycoprotein attenuates the mobiliza-tion of intracellular Ca2+ in cadmium chloride-exposed BNL CL.2cells (Fig. 2). It has been postulated that there is closely relationshipbetween cadmium and calcium, because their radius is almost sim-ilar. In fact, the radius of Cd2+, a common form of free cadmium inthe body, is very similar to that of Ca2+ (0.099 and 0.097 nm, respec-tively). Thus, Cd2+ not only competitively inhibits the influx of Ca2+
but also causes the increase of Ca2+ by inhibiting Ca2+-ATPase on themembrane of the depot of calcium [27,28]. In addition, cadmiumcan activate or inhibit some calcium-related enzymes instead ofCa2+ [29].
AP-1 regulates the transcription of a vast variety of genes, someof which are involved in inflammatory mediator, neoplastic trans-formation and tumor promotion [30]. It has been reported thatCOX-2 and iNOS are well known pro-inflammatory factors and theirexpressions have been shown to be regulated by AP-1 [31]. Withcarcinogenesis, cadmium can activate oncogenes or genes associ-ated with mitogenesis such as c-Jun and c-Fos both in vivo andin vitro [32–34]. In the AP-1 (c-Jun and c-Fos) expression, the resultsin this study indicated that the GJE glycoprotein has an inhibitoryeffect on the expression of AP-1 in cadmium chloride-exposed BNLCL.2 cells (Fig. 3).
Many scientists have interested in AA and its metabolitesdue to growing evidence of their significant role in cancer and
inflammation biology [35]. What the AA and its metabolites areby certain compounds inhibited or modulated, means that inflam-mation and carcinogenesis are blocked. In radioactivity analysisof [3H] arachidonic acid, the results showed that GJE glycopro-tein (100 �g/ml) inhibits expression of AA at exposure of cadmiumchloride (10 �M) (Fig. 4).
AA and COX-2 contribute to balancing homeostasis in cellularfunction, inflammation and tumorgenesis. The specific actions ofAA derivatives [COX, lipoxygenase (LOX) and cytochrome (CYP)450] are responsible for metabolic pathways and for signal path-way transduction. Among them, COX-2 is a representative factorfor inflammatory pathway as one of endotoxins at the liver car-cinogenesis. It has been demonstrated that expression of COX-2 isinduced by carcinogen through activating activator protein-1 (AP-1) and then the expressed COX-2 promotes the activation of MMP-9[36]. One of inflammation- and pro-tumor-mediators, induciblenitric oxide synthase (iNOS) is an inducible enzyme that causes theoverproduction of nitric oxide regarding inflammation and tumordevelopment [12]. As shown in Fig. 5, expression of COX-2 andiNOS was diminished at treatment with GJE glycoprotein, whiletheir expression was increased at treatment with cadmium chlo-ride alone in the BNL CL.2 cells comparison with control.
To know whether solid tumor stage would be developed by cad-mium chloride under experimental condition (10 �M, 30 h) in thisstudy, MMP activation was evaluated. Although MMP activation isnot closely related to inflammation, it involved in the occurrence ofsolid tumor after inflammation stage. MMP’s are responsible for thedegradation of extracellular matrix components such as collagen,

J. Lee, K.-T. Lim / Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246 245
laminin and proteoglycans. The MMP’s are activated by removalof an approximately 10 kDa segment from the N-terminus in thepresence of zinc ion, while they are inhibited by tissue inhibitorof metalloproteinases (TIMP) [37]. It means that overexpression ofMMPs is involved in tissue remodeling, tumor invasion, metasta-sis, and angiogenesis [38]. Particularly, MMP-9 has been knownspecifically to cleave type IV collagen which can degrade mostECM components forming the basal membrane [39]. It has beenreported that MMP-9 promoter has several transcription factor-binding motifs, including AP-1 in liver cells [40]. As shown in Fig. 6,MMP-9 expression is significantly decreased by GJE glycoprotein(100 �g/ml) in the cadmium chloride-induced BNL CL.2 cells. More-over, we evaluated that whether SJSZ glycoprotein prevents cellinvasion or not. Several studies research cell invasion by cadmiumchloride using Material invasion chamber [41]. As shown in Fig. 6,quantitative data derived from three independent experimentssupported GJE glycoprotein effectively inhibited the invasion.
There are two possible mechanisms that GJE glycoprotein pro-tected BNL CL.2 cells from the carcinogenesis caused by treatmentwith cadmium chloride. One of them is that GJE glycoprotein scav-enges intracellular ROS induced by cadmium chloride, because ithas a strong antioxidative character. Also, the other is that GJEglycoprotein interacts with cadmium with positive divalent cationof cadmium and then builds a complex compound before acrossthe cell membrane. This means that GJE glycoprotein likely playsas a chelate agent to inactive cadmium. Thus, cadmium chlorideconsequently loses its cytotoxicity. Taken together, GJE glycopro-tein prevents the BNL CL.2 cells from the carcinogenesis caused bytreatment with cadmium chloride.
In conclusion, the results in this study showed that GJE glyco-protein (100 �g/ml) inhibits the production of intracellular ROS,mobilization of intracellular Ca2+, expression of AP-1 (c-Jun andc-Fos), arachidonic acid, COX-2, and MMP-9 in the cadmiumchloride-treated BNL CL.2 cells. Taken together, these findingssuggest that GJE glycoprotein might be used for protection ofinflammation caused by cadmium ion as one of natural compounds.Still, it remains problems to elucidate whether GJE glycoproteinalso blocks cancer developing initiative stage after inflammationstage caused by cadmium ion in vivo.
Acknowledgements
This work was financially supported by Chonnam National Uni-versity in 2011.
References
[1] Zhou JL, Salvador SM, Liu YP, Sequeria M. Heavy metals in the tissues of dol-phins (Delphinus delphis) stranded on the Portuguese coast. Sci Total Environ2001;273:61–76.
[2] IARC Monographs on the Evaluation of the Carcinogenic Risks to Humans. Beryl-lium, Cadmium, Mercury, and Exposures in the Glass Manufacturing Industry,vol. 58. International Agency for Research on Cancer, Lyon, France, 1993, pp.119–238.
[3] Järup L. Hazards of heavy metal contamination. Br Med Bull 2003;68:167–82.[4] Kataranovski M, Jankovic S, Kataranovski D, Stosic J, Bogojevic D. Gender dif-
ferences in acute cadmium-induced systemic inflammation in rats. BiomedEnviron Sci 2009;22:1–7.
[5] Yamano T, DeCicco LA, Rikans LE. Attenuation of cadmium-induced liver injuryin senescent male fischer 344 rats: role of Kupffer cells and inflammatorycytokines. Toxicol Appl Pharmacol 2000;162:68–75.
[6] Kuester RK, Waalkes MP, Goering PL, Fisher BL, McCuskey RS, Sipes IG. Differ-ential hepatotoxicity induced by cadmium in Fischer 344 and Sprague-Dawleyrats. Toxicol Sci 2002;65:151–9.
[7] Farombi EO, Shrotriya S, Surh YJ. Kolaviron inhibits dimethyl nitrosamine-induced liver injury by suppressing COX-2 and iNOS expression via NF-kappaBand AP-1. Life Sci 2009;84:149–55.
[8] Stacey NH, Cantilena Jr LR, Klaassen CD. Cadmium toxicity and lipid peroxida-tion in isolated rat hepatocytes. Toxicol Appl Pharmacol 1980;53:470–80.
[9] Surh YJ, Kundu JK, Na HK, Lee JS. Redox-sensitive transcription factors as primetargets for chemoprevention with anti-inflammatory and antioxidative phyto-chemicals. J Nutr 2005;135:2993S–3001S.
[10] Eferl R, Wagner EF. AP-1: a double-edged sword in tumorigenesis. Nat RevCancer 2003;3:859–68.
[11] Mohan S, Epstein JB. Carcinogenesis and cyclooxygenase: the potentialrole of COX-2 inhibition in upper aerodigestive tract cancer. Oral Oncol2003;39:537–46.
[12] Chung WY, Park JH, Kim MJ, Kim HO, Hwang JK, Lee SK, Park KK. Xanthorrhizolinhibits 12-O-tetradecanoylphorbol-13-acetate-induced acute inflammationand two-stage mouse skin carcinogenesis by blocking the expression ofornithine decarboxylase, cyclooxygenase-2 and inducible nitric oxide synthasethrough mitogen-activated protein kinases and/or the nuclear factor-kappa B.Carcinogenesis 2007;28:1224–31.
[13] Mordan LJ, Burnett TS, Zhang LX, Tom J, Cooney RV. Inhibitors of endogenousnitrogen oxide formation block the promotion of neoplastic transformation inC3H 10T1/2 fibroblasts. Carcinogenesis 1993;14:1555–9.
[14] Chang HM, But PPH. Pharmacology and applications of Chinese Materia Medica,vol. 2. Singapore: World Scientific Publishing; 1987.
[15] Oh PS, Lim KT. Plant originated glycoprotein has anti-oxidative and anti-inflammatory effects on dextran sulfate sodium-induced colitis in mouse. JBiomed Sci 2006;13:549–60.
[16] Lee SJ, Oh PS, Lim KT. Hepatoprotective and hypolipidaemic effects of glyco-protein isolated from Gardenia jasminoides ellis in mice. Clin Exp PharmacolPhysiol 2006;33:925–33.
[17] Lee J, Lim KT. Plant-originated glycoprotein (36 kDa) suppresses interleukin-4and -10 in bisphenol A-stimulated primary cultured mouse lymphocytes. DrugChem Toxicol 2010;33:421–9.
[18] Kimata M, Shichijo M, Miura T, Serizawa I, Inagaki N, Nagai H. Ca2+ and proteinkinase C signaling for histamine and sulfidoleukotrienes released from humancultured mast cells. Biochem Biophys Res Commun 1999;257:895–900.
[19] Xing M, Tao L, Insel PA. Role of extracellular signal-regulated kinase and PKCalpha in cytosolic PLA2 activation by bradykinin in MDCK-D1 cells. Am J Physiol1997;272:C1380–7.
[20] Oh PS, Lim KT. Plant glycoprotein modulates the expression of interleukin-1beta via inhibition of MAP kinase in HMC-1 cells. Biosci Biotechnol Biochem2008;72:2133–40.
[21] Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with theFolin phenol reagent. J Biol Chem 1951;193:265–75.
[22] Pan MH, Chiou YS, Chen WJ, Wang JM, Badmaev V, Ho CT. Pterostilbene inhib-ited tumor invasion via suppressing multiple signal transduction pathways inhuman hepatocellular carcinoma cells. Carcinogenesis 2009;30:1234–42.
[23] Souza V, Escobar Md Mdel C, Gómez-Quiroz L, Bucio L, Hernández E, CossioEC, Gutiérrez-Ruiz MC. Acute cadmium exposure enhances AP-1 DNA bindingand induces cytokines expression and heat shock protein 70 in HepG2 cells.Toxicology 2004;197:213–28.
[24] Waalkes MP. Cadmium carcinogenesis. Mutat Res 2003;533:107–20.[25] Kamata H, Hirata H. Redox regulation of cellular signalling. Cell Signal
1999;11:1–14.[26] Klaunig JE, Kamendulis LM, Hocevar BA. Oxidative stress and oxidative damage
in carcinogenesis. Toxicol Pathol 2010;38:96–109.[27] Swandulla D, Armstrong CM. Calcium channel block by cadmium in chicken
sensory neurons. Proc Natl Acad Sci USA 1989;86:1736–40.[28] Visser GJ, Peters PH, Theuvenet AP. Cadmium ion is a non-competitive inhibitor
of red cell Ca(2+)-ATPase activity. Biochim Biophys Acta 1993;1152:26–34.[29] Beyersmann D, Hechtenberg S. Cadmium, gene regulation, and cellular sig-
nalling in mammalian cells. Toxicol Appl Pharmacol 1997;144:247–61.[30] Dong Z, Birrer MJ, Watts RG, Matrisian LM, Colburn NH. Blocking of tumor
promoter-induced AP-1 activity inhibits induced transformation in JB6 mouseepidermal cells. Proc Natl Acad Sci USA 1994;91:609–13.
[31] Nanji AA, Jokelainen K, Tipoe GL, Rahemtulla A, Thomas P, Dannenberg AJ.Curcumin prevents alcohol-induced liver disease in rats by inhibiting theexpression of NF-kappa B-dependent genes. Am J Physiol Gastrointest LiverPhysiol 2003;284:G321–7.
[32] Zheng H, Liu J, Choo KH, Michalska AE, Klaassen CD. Metallothionein-I and -IIknock-out mice are sensitive to cadmium-induced liver mRNA expression ofc-jun and p53. Toxicol Appl Pharmacol 1996;136:229–35.
[33] Joseph P, Muchnok TK, Klishis ML, Roberts JR, Antonini JM, Whong WZ, Ong T.Cadmium-induced cell transformation and tumorigenesis are associated withtranscriptional activation of c-fos, c-jun, and c-myc proto-oncogenes: role ofcellular calcium and reactive oxygen species. Toxicol Sci 2001;61:295–303.
[34] Liu J, Qu W, Kadiiska MB. Role of oxidative stress in cadmium toxicity andcarcinogenesis. Toxicol Appl Pharmacol 2009;238:209–14.
[35] Hyde CA, Missailidis S. Inhibition of arachidonic acid metabolism and its impli-cation on cell proliferation and tumour-angiogenesis. Int Immunopharmacol2009;9:701–15.
[36] Grau R, Punzón C, Fresno M, Iniguez MA. Peroxisome-proliferator-activatedreceptor alpha agonists inhibit cyclo-oxygenase 2 and vascular endothelialgrowth factor transcriptional activation in human colorectal carcinoma cellsvia inhibition of activator protein-1. Biochem J 2006;395:81–8.
[37] Naglich JG, Jure-Kunkel M, Gupta E, Fargnoli J, Henderson AJ, Lewin AC, TalbottR, Baxter A, Bird J, Savopoulos R, Wills R, Kramer RA, Trail PA. Inhibition of angio-genesis and metastasis in two murine models by the matrix metalloproteinaseinhibitor, BMS-275291. Cancer Res 2001;61:8480–5.
[38] Peng PL, Hsieh YS, Wang CJ, Hsu JL, Chou FP. Inhibitory effect of berber-ine on the invasion of human lung cancer cells via decreased productions

246 J. Lee, K.-T. Lim / Journal of Trace Elements in Medicine and Biology 25 (2011) 239– 246
of urokinase-plasminogen activator and matrix metalloproteinase-2. ToxicolAppl Pharmacol 2006;214:8–15.
[39] Chantrain CF, Henriet P, Jodele S, Emonard H, Feron O, Courtoy PJ,DeClerck YA, Marbaix E. Mechanisms of pericyte recruitment in tumourangiogenesis: a new role for metalloproteinases. Eur J Cancer 2006;42:308–10.
[40] Kim S, Kim Y, Lee Y, Cho KH, Kim KH, Chung JH. Cholesterol inhibits MMP-9 expression in human epidermal keratinocytes and HaCaT cells. FEBS Lett2007;581:3869–74.
[41] Rose P, Huang Q, Ong CN, Whiteman M. Broccoli and watercress suppressmatrix metalloproteinase-9 activity and invasiveness of human MDA-MB-231breast cancer cells. Toxicol Appl Pharmacol 2005;209:105–13.


![Mapping of the binding sites of human diamine oxidase (DAO) … · catalysed by diamine oxidase (DAO, EC 1.4.3.22) [4 –6]. Human DAO is a homodimeric glycoprotein of ca. 200 kDa](https://static.fdocuments.us/doc/165x107/5ea831c2f5501e308b0e243b/mapping-of-the-binding-sites-of-human-diamine-oxidase-dao-catalysed-by-diamine.jpg)















