Influence of Some Physicochemical Parameters on Artemia ... · o Copyrisbt by tbc Wodd AquacuI...
5
JOURNAL OF TUE WORLD AQUACULTURE SOCIETY Vol. 35, No. 2 June, 2004 Influence of Some Physicochemical Parameters on Artemia Biomass and Cyst Production in Some Thalassohaline Aquatic Environments in the Colombian Caribbean WILLIAM N. CAMARGO Fisheries and l/Iinois Aquaculture Center. Southem l/Iinois University Carbondale. Carbondale. l/Iinois 62901 USA JOE S. ELY Departmentof Plant Biology. Southem l/Iinois Universityof Carbondale.Carbondale. l/linois 62901 USA GABRIEL M. DURAN-COBO Eseuela Superior Politeeniea del Utoral (ESPOL). Guayaquil. Eeuador PATRICK SORGELOOS Artemia Referenee Center and Laboratory of Aquaculture. Ghent University. Rover 44. Ghent B-9000 Belgium Abstraet.-From July 1998 to June 2000 four thal- assohaline aquaûc environments along the Colombian Caribbean coast (Manaure. Galerazamba, Salina Cero, and Tayrona) wcre surveycd monthly to determine the influence of salinity, percent 0, saturaÛon. pH. tem- perature, and nutrients (NO, -, NO, - and PO. ") on Ar- temia (Cmstacean, Anosuacan) biomass producûon and cyst producûon potenûal. The effects of the reg- ularly measurcd physicocbemical parameters on bic- mass and cyst producûon potenûal were anaIyzed us- ing univariate anaIysis of variance (SPSS VIO.O).The infIuence of physicochemical parameters on biomass producûon was not significant (P > O.05). In contrast, there was a significant interacûon (P < O.05)of salin- ity, percent 0, saturaûon. and nitrate (uscd as a proxy for chlorophyll a) on cyst producûon potenûal. In ad- diûon. for all four locaûons nitrate levels were directly propotûonal to salinity. Tbis might be exp1ainedby the fact that in saltWorks numerous organisms are trapped and slowly die as salinity increases progressively in the evaporaÛDgbasins; thus, organic matter accumu- lates and decomposes. Consequently, the concentration of the nitrogenous compounds, first nitrite and later nitrate, increases through time as salinity increases. Moreover, decreasing nitrate levels seem toîiierease cyst producûon potenûa1; thus supporûng the noûon that when insufficient food is available cyst producûon increases. Increasing sampling efforts in Latin America for potential Artemia (Crostacean, Anostracan) sites during the mid 1990s have resu1ted in numerous reports of Artem- ia populations and the characterization of Artemia habitats (Camargo et al. 2000; Ca- margo 2002; Camargo et al. 2002a, 2003), particu1ar1y with respect to remote thalas- sohaline aquatic environments. Lenz (1987) observed that zooplankton population dynamics are influenced by abi- otic factors (salinity, temperature, and nu- trients concentration) and by bio10gical in- teractions (predation, competition, and grazers). Further, Por (1980) and Lenz (1987) showed that bio10gical interactions are limited in hypersaline communities where species diversity is 10w, and abiotic parameters, particular1y those that regu1ate seasonal characteristics, are dominant. Lenz (1987) identified tWo critical factors that determine the population dynamics of Ar- temia and they are: 1) habitat conditions al- 10wing the survival of the animals through- ou~~ar; and 2)-p'~dicta'pi1ity of the seasonality of the environment. Bisexual and parthenogenetic Artemia species are characterized by an extensive genetic variation resu1ting in difference in their life history traits (Lenz and Browne 1991). Under laboratory conditions, 10n- gevity, fecundity, and reproductive period vary one order of magnitude among Artem- ia species and populations (Browne 1980, o Copyrisbt by tbc Wodd AquacuI Sacicty 2004 274 ARTEMIA BIOMASS AND CYST PRODucnON IN COLOMBIA 275 ., ~1. 1988; Lenz 1984, 1987; Dana and Lenz 1986; Browne and Hoopes 1990). Browne et al. (2002) measured reproductive and life span traits for tWo obligate parthenogenetic types and three sexual (tWo A. franciscana and one A. sinica) brine shrimp populations and determined that, for aU traits studied, the environmental component was greater than the genetic component measured. However, every trait has a genetic compo- nent that can potential1y be acted upon by selection. The average genetic component of variation for the 10 traits was 23.44%, ranging from 5.26% for number of cysts to 44.87% for number of nauplii. Eco10gical and physio10gical differences have been previous1y reported among sev- era! populations of A. franciscana (Sorge- loos et al. 1975; Abreu-Grobois and Beard- more 1982; Bowen et al. 1985). These eco- 10gical and physio10gical differences have important impacts on Artemia populations by influencing reproductive strategies through biomass and cyst production. In saltWorks, Artemia occur abundant1y in the evaporation ponds at salinity levels betWeen 80 to 220 gIL (depending on the strain and/or species), and in rare occasions have been found at salinities up to 340 gIL (post and Youssef 1977). At these salinity levels high ionic concentrations are a lim- iting factor due to the energy requirements for osmoregu1ation. Artemia populations are favored by the absence of predators and food competitors and deve10p successfully in these extreme biotopes often at high den- sities. Further the dominance of these hy- persaline habitats is shared with archaeo- bacterial halophiles (Grant and Ross 1986) and halotolerant microalgae as the concen- tration of salts approaches saturation. Fur- ther, a re1ative1y smal1 number of these hal- obacterias and microalgae species might serve as a food source to this very efficient, non-se1ective filter feeder (Sorgeloos 1980). Some Artemia strains are predominantly oviparous at high salinities (i.e., Lake Ur- mia, Lake Grassmere, some strains from Spain, and Manaure strains-Berthé1émy- Okazaki 1986; Camargo et al. 2000), whi1e other strains primari1y opt for the ovovivip- arous mode of reproduction at high salini- ties (i.e., Great Salt Lake, Salina Cero strains-Amat 1982; Carnargo et al. 2000). Depending on the strain and the hydrobi0- logical conditions of the pond (i.e., high water retention, high evaporation, and pri- mary productivity), cyst production is in- duced by changes in brood number, salinity, photoperiod, temperature, hypoxia, and iron rich foods (Dutrieu 1960; Baker 1966; Bowen et al. 1969; Sorge1oos et al. 1975; Provasoli and Pintner 1980; Amat 1982; Berthé1émy-Okazaki 1986). Cyst produc- tion can be produced seasonaUy or annu- al1y. Studies in vitro revealed that optimum temperature and salinity varies between Ar- temia populations (Vanhaecke et al. 1984; Wear et al. 1986; Browne et al. 1988; Ca- margo, unpublished data). A. franciscana populations have a high reproductive ca- pacity up to senescence (Browne et al. 1984). Browne et al. (1988) further re- vealed that A. franciscana is the most plas- tic (flexible) Artemia species with respect to temperature followed by the partheno- genetic type. In nature, Artemia is found in neutral to alkaline waters. According to Vos (1979), nauplii growth decreases and the overall ap- pearance of adu1ts deteriorates with pH val- ues be10w 7.0, and they conc1uded that the optimum pH for Artemia growth ranges from 8.0 to 8.5. Additional1y, Sato (1967) determined that cyst hatching efficiency was greatly compromised at pHs be10w 8.0. The aim of this research was to identify physicochemica1 variables that influence monthly Artemia biomass production in vivo and monthly cyst production potential of some wild Co10mbian populations (Man- aure, Galerazamba, Tayrona, and Salina Cero) for potential use in aquaculture. Study Areas Manaure. Located west of the town of Manaure (l1046'32"N 72°29'27''W), in the
-
Upload
nguyenthuy -
Category
Documents
-
view
214 -
download
0
Transcript of Influence of Some Physicochemical Parameters on Artemia ... · o Copyrisbt by tbc Wodd AquacuI...

JOURNAL OF TUEWORLD AQUACULTURE SOCIETY
Vol. 35, No. 2June, 2004
Influence of Some Physicochemical Parameters on ArtemiaBiomass and Cyst Production in Some ThalassohalineAquatic Environments in the Colombian Caribbean
WILLIAM N. CAMARGO
Fisheries and l/Iinois Aquaculture Center. Southem l/Iinois University Carbondale.Carbondale. l/Iinois 62901 USA
JOE S. ELY
Departmentof Plant Biology.Southem l/Iinois Universityof Carbondale.Carbondale.l/linois 62901 USA
GABRIEL M. DURAN-COBO
Eseuela Superior Politeeniea del Utoral (ESPOL). Guayaquil. Eeuador
PATRICK SORGELOOS
Artemia Referenee Center and Laboratory of Aquaculture. Ghent University. Rover 44.Ghent B-9000 Belgium
Abstraet.-From July 1998 to June 2000 four thal-assohaline aquaûc environments along the ColombianCaribbean coast (Manaure. Galerazamba, Salina Cero,and Tayrona) wcre surveycd monthly to determine theinfluence of salinity, percent 0, saturaÛon. pH. tem-perature, and nutrients (NO,-, NO, - and PO.") on Ar-temia (Cmstacean, Anosuacan) biomass producûonand cyst producûon potenûal. The effects of the reg-ularly measurcd physicocbemical parameters on bic-mass and cyst producûon potenûal were anaIyzed us-ing univariate anaIysis of variance (SPSS VIO.O).TheinfIuence of physicochemical parameters on biomassproducûon was not significant (P > O.05). In contrast,there was a significant interacûon (P < O.05)of salin-ity, percent 0, saturaûon. and nitrate (uscd as a proxyfor chlorophyll a) on cyst producûon potenûal. In ad-diûon. for all four locaûons nitrate levels were directlypropotûonal to salinity. Tbis might be exp1ainedby thefact that in saltWorks numerous organisms are trappedand slowly die as salinity increases progressively inthe evaporaÛDgbasins; thus, organic matter accumu-lates and decomposes. Consequently, the concentrationof the nitrogenous compounds, first nitrite and laternitrate, increases through time as salinity increases.Moreover, decreasing nitrate levels seem toîiiereasecyst producûon potenûa1; thus supporûng the noûonthat when insufficient food is available cyst producûonincreases.
Increasing sampling efforts in LatinAmerica for potential Artemia (Crostacean,Anostracan) sites during the mid 1990shave resu1ted in numerous reports of Artem-
ia populations and the characterization of
Artemia habitats (Camargo et al. 2000; Ca-
margo 2002; Camargo et al. 2002a, 2003),particu1ar1y with respect to remote thalas-sohaline aquatic environments.
Lenz (1987) observed that zooplankton
population dynamics are influenced by abi-otic factors (salinity, temperature, and nu-trients concentration) and by bio10gical in-teractions (predation, competition, andgrazers). Further, Por (1980) and Lenz(1987) showed that bio10gical interactionsare limited in hypersaline communitieswhere species diversity is 10w, and abioticparameters, particular1y those that regu1ateseasonal characteristics, are dominant. Lenz
(1987) identified tWo critical factors thatdetermine the population dynamics of Ar-temia and they are: 1) habitat conditions al-10wing the survival of the animals through-ou~~ar; and 2)-p'~dicta'pi1ity of theseasonality of the environment.
Bisexual and parthenogenetic Artemia
species are characterized by an extensivegenetic variation resu1ting in difference intheir life history traits (Lenz and Browne1991). Under laboratory conditions, 10n-gevity, fecundity, and reproductive periodvary one order of magnitude among Artem-ia species and populations (Browne 1980,
o Copyrisbt by tbc Wodd AquacuI Sacicty 2004
274
ARTEMIA BIOMASS AND CYST PRODucnON IN COLOMBIA 275
.,~1.
1988; Lenz 1984, 1987; Dana and Lenz1986; Browne and Hoopes 1990). Browneet al. (2002) measured reproductive and lifespan traits for tWo obligate parthenogenetictypes and three sexual (tWo A. franciscanaand one A. sinica) brine shrimp populationsand determined that, for aU traits studied,the environmental component was greaterthan the genetic component measured.However, every trait has a genetic compo-nent that can potential1y be acted upon byselection. The average genetic componentof variation for the 10 traits was 23.44%,
ranging from 5.26% for number of cysts to44.87% for number of nauplii.
Eco10gical and physio10gical differenceshave been previous1y reported among sev-era! populations of A. franciscana (Sorge-loos et al. 1975; Abreu-Grobois and Beard-more 1982; Bowen et al. 1985). These eco-10gical and physio10gical differences haveimportant impacts on Artemia populationsby influencing reproductive strategiesthrough biomass and cyst production.
In saltWorks, Artemia occur abundant1yin the evaporation ponds at salinity levelsbetWeen 80 to 220 gIL (depending on thestrain and/or species), and in rare occasionshave been found at salinities up to 340 gIL(post and Youssef 1977). At these salinitylevels high ionic concentrations are a lim-iting factor due to the energy requirementsfor osmoregu1ation. Artemia populationsare favored by the absence of predators andfood competitors and deve10p successfullyin these extreme biotopes often at high den-sities. Further the dominance of these hy-persaline habitats is shared with archaeo-bacterial halophiles (Grant and Ross 1986)and halotolerant microalgae as the concen-tration of salts approaches saturation. Fur-ther, a re1ative1y smal1 number of these hal-obacterias and microalgae species mightserve as a food source to this very efficient,non-se1ective filter feeder (Sorgeloos 1980).
Some Artemia strains are predominantlyoviparous at high salinities (i.e., Lake Ur-mia, Lake Grassmere, some strains fromSpain, and Manaure strains-Berthé1émy-
Okazaki 1986; Camargo et al. 2000), whi1eother strains primari1y opt for the ovovivip-arous mode of reproduction at high salini-ties (i.e., Great Salt Lake, Salina Cerostrains-Amat 1982; Carnargo et al. 2000).Depending on the strain and the hydrobi0-logical conditions of the pond (i.e., highwater retention, high evaporation, and pri-mary productivity), cyst production is in-duced by changes in brood number, salinity,photoperiod, temperature, hypoxia, and ironrich foods (Dutrieu 1960; Baker 1966;Bowen et al. 1969; Sorge1oos et al. 1975;Provasoli and Pintner 1980; Amat 1982;Berthé1émy-Okazaki 1986). Cyst produc-tion can be produced seasonaUy or annu-al1y.
Studies in vitro revealed that optimumtemperature and salinity varies between Ar-temia populations (Vanhaecke et al. 1984;Wear et al. 1986; Browne et al. 1988; Ca-margo, unpublished data). A. franciscanapopulations have a high reproductive ca-pacity up to senescence (Browne et al.1984). Browne et al. (1988) further re-vealed that A. franciscana is the most plas-tic (flexible) Artemia species with respectto temperature followed by the partheno-genetic type.
In nature, Artemia is found in neutral toalkaline waters. According to Vos (1979),nauplii growth decreases and the overall ap-pearance of adu1ts deteriorates with pH val-ues be10w 7.0, and they conc1uded that theoptimum pH for Artemia growth rangesfrom 8.0 to 8.5. Additional1y, Sato (1967)determined that cyst hatching efficiencywas greatly compromised at pHs be10w 8.0.
The aim of this research was to identifyphysicochemica1 variables that influencemonthly Artemia biomass production invivo and monthly cyst production potentialof some wild Co10mbian populations (Man-aure, Galerazamba, Tayrona, and SalinaCero) for potential use in aquaculture.
Study Areas
Manaure. Located west of the town ofManaure (l1046'32"N 72°29'27''W), in the

:a, _,..,..
.:::::~!!f,... ", '.;;'-'"
276 CAMARoo ET AL.
center of La Guajira department, near thecity of Riohacha. This saltwork is a thal-assohaline shallow aquatic ecosystem thatextends over 4,000 ha. This zone was orig-inally a natural lagoon, still surrounded insome small areas by mangroves.
Galerazamba. Located approximately 20km to the north of the city of Cartagena(lOo47'38"N 75°14'48"W). This small thal-assohaline saltwork bas five ponds (three toobtain brine and two crystallizers) with atotal extension of 220 ha. This saltwork was
built in a natural saline lagoon, surroundedby some mangroves.
Tayrona or Chengue salt pond. Thissmall natural thalassohaline saltwork of ap-proximately 2.5 ha is situated (llOI9'03"N,74°08'13"W) approximately in the middleof the Tayrona National Natural Park. Tbesalt pond is ftooded, during most of theyear, by the often prolonged winter andserves as a saltwork during summer.
Salina Cero or Cienaga Prieto. Thissmall thalassohaline lagoon of only 18 hais located at only 3 km of Galerazamba(loo46'29"N, 75°15'55"W). For several de-cades, salt has been manually extracted oc-casionally once or twice per year.
Materials and Methods
Twenty Artemia samples were taken ran-domly in a given pond (Camargo et al.2000; Camargo 2002; Camargo et al.2oo2b) at each site (Manaure, Galerazamba,Tayrona, and Salina Cero) every monthwith a 14-L acrylic box (Haslen and Wear1985). Tbe samples were collected in bothshallow and deeper water sectors. Sampleswere preserved in a formalin-alcohol solu-tion (4% borax buffered), and maintainedin a cooler until they arrived at the labora-tory where they were counted and classifiedusing a stereoscope to determine populationdistribution (females, males, juveniles, andnauplius). .~
Samples containing a known number ofadults (males and females) were strainedand dried in an oven at 80 C for 24 h (Ca-margo et al. 2000; Camargo 2002). Iodivid-
ual average weight trom each of the fourlocations was calculated according to Ca-margo et al. (2000) by dividing the sampledry weight by the number of adults in eachsample. Finally, to calculate total dry bio-mass (producûon), the individu al dryweight was multiplied by the number ofadults counted at each of 20 stations in the
14-L box and finally multiplied by the totalrespective pond volume. Our assumptionfor the meao weight is that all individualstrom each population were the same weight(Camargo et al. 2000; Camargo 2002).
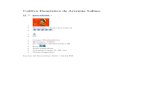
AdditionaUy, the foUowing physico-chemical parameters were determined, ateach staûon and location, simultaneouslywith the Artemia samples collection: salin-ity (temperature compensated refractome-ter); percent O2 saturation and temperature(Oxymeter WTW~ 330 calibrated prior touse, WTW Measurement Systems, Ioc., Ft.Myers, Florida, USA); pH (pHmeterWTW~ 330 calibrated prior to use, WTWMeasurement Systems, Inc., Ft. Myers,Florida, USA); nitrates, nitrites, and phos-phates (colorimetrie method within an hourof water sample collection using Hatch~DREL 2010 spectrophotometer, Hach Com-pany, Loveland, Colorado, USA); and chlo-rophyll a (we use the seston method, pre-serving samples in a cooler while arrivingto the laboratory; samples were then filteredusing Whatman~ GF/C filter papers and ac-etone for destruction and chlorophyll a ex-traction and finally read using a Hatch~DREL 2010 spectrophotometer, Hach Com-pany, Loveland, Colomdo, USA). We usedan indirect method to measure primary pro-ductivity, by correlating a limiting nutrient(nitrate)- to primaI'J-Productivity_Further,the N salt form (nitrate) was used ratherthan nitrites or phosphates, since nitrites asweIl as phosphates showed very low values,sometimes below detection levels (Fig. 1).Besides, no trend was established statisti-cally between nitrite or phosphate and cystand biomass production (Figs. 2a, 2b, 2h,2i).
Reproductive experiments were conduct-
ARTEMIA BIOMASS AND CYST PRODUcnON IN COLOMBIA 277
I) Mauawc, JuIy 199110 April 2000
"""'''''''ONO''''''A--'.SeplI99810 Juno 2000
-0'
...'11I A 11I.. .. A'ONO""IIIIA"--
c)T., (~~ -..,. 1999tGMaodo2000 :,""i...'1:1-,10I· f...'" , .. A A.ONO""..--
cl)__ JuIy199110J 2000
...j""i....I:1-I10I. f
= pol(ohn') dr'J_(ohn') ~I...IN03-I __"02_
FlOURE1. lnteroction of significant physleocMmleal parameters over monthly Artcmia biomass production andcyst production potent/al at the four locatlons.
..
1!, ed in the laboratory according to Vanhaecke
et al. (1984) under controlled conditions(salinity, temperature, DO, pH, illumina-tion, and supplying artificial food ad libi-tum) at two salinities (80 and 120 gIL) in
order to determine the mean cyst produc-tion potential for each site (Newmark 1988;Duran and Garcia 2001). Tbe density of fe-males in the pond was detennined by mul-tiplying the number of females counted in

-.18?;~~~-:; ~:~
278 CAMARGO Er AL.
\t..!
_ c...,.-- __ca-..-__
~8
I
I
- -..--Population
FIOURB2. Mean values wi/h s/andard errors for a) Artcmia c:ys/produc/ion po/en/ia/' b) Artcmia biomassproduc/ion (DW), c) pH. d) salinity. e) /empera/Ure, ft percent O2 sa/UratÎon. g) nitra/e, h) ni/rite, and i)phospha/es. (Chengue: Tayrona NNP).
the 20 samples by the pond volume at eachsite (Camargo et al. 2000; Camargo 2002).Pond volume was estimated from pondmean depth and surface area. Tota!cyst pro-duction potential was calculated at each re-spective site by multiplying the mean num-ber of cysts per female (previously obtained
in vitro) of a given population by the totalnumber of females in the pond.
Finally, the effects of the measured phys-icochemical parameters (Tabie 1) on bio-mass production and cyst production poten-tial were analyzed using univariate ANO-VA with Thkey post-hoc tests (SPSS VI0.0,
TABLE1. ResullS for /he field de/ermination of blotic and abiotic parameters of four Colomblon Artcmiapopulations. All values are rangesfor sampling period (July 1998-June 2(00) excep/ precipitation (maximum).
Parameters Manaure - -ÖaIeraiäffib-a-- - -layrona Salina Cero
Salinity (gIL) 148-275 65-295 34-330 19-204pH 7.6-7.9 7.2-8.1 7.9-11.8 6.7-11.6Percent 0. saL 56-99 70-150 23-131 53-131Temp. (C) 24.9-31.3 26.6-35.5 23.4-33.8 27.5-35.1N02- (mg/L) 0.005-0.025. 0.120-0.005 0.002-0.018 0.003-0.115NO,- (mg/L) 0.29-20.45 '. 1.4-33.7 2.15-22.1 0.4-18.75PO.-' (mg/L) 0.05-1.27 0.33-1.98 0.32-2.83 0.21-5.05P=ipitation (mm) 79.6 326.7 288.2 326.7ChI. a (mg/m') ND 0.01-0.11 0.01-0.39 0.09-3.04
ARTBMIA BIOMASS AND CYST PRODUcnON IN COLOMBIA 279
-....Z
SPSS Inc., Chicago, Dlinois, USA) amongpopulations. Variables (salinity, percent O2saturation, nitrate, and temperature) select-ed for the analyses were based on previ-ously observed relationships (Camargo etal. 2000) between physicochemical param-eters and dependent variables. Water tem-perature was excluded from the final anal-ysis because its correlation with cyst andbiomass production was insignificant. In or-der 10 apply the univariate statistical anal-ysis 10 the field data. we had to use a scor-ing method for physicochemical parametersranges, rather than reporting partial missingdata or deleting rows of incomplete data,because of the periodically partial or total(some or no water in the pond) crystalli-zation of some locations and its negativeeffect on the analysis. Dependent variables(biomass and cyst production) can have val-ues equal 10 zero, contrasting with indepen-dent variables (i.e., pH, temperature, salin-ity, and percent O2 saturation), which couldnot be measured upon crystallization of thepond, thus presenting a blank. Tbe valuesfor each variabie were scored consistentlyfrom 1 to 3, as follows: each physicochem-ical parameter analyzed was divided intothree data groups (containing 0 10 the max-imum value recorded in the field), wherecrystallization of the basin (0 or no data)was represented by rank No. 1 and the sub-sequent measured values within the tworanges were set to 2 and 3, respectively.Tbe scoring was done, rather than using theoriginal values, in order 10 determine pos-sible relationships. Tbe alpha (a) was set at0.05 for all analyses.
Results
Population Site Characteristics
At Manaure the overall distribution of fe-males (F), males (M), juveniles (1)and nau-plius (N) was 22:16:28:35 (F:M:J:N), witha female to male (F:M) ratio of 1:0.84.Bio-mass production in two evaporation areasof this very large saltwork occurred for 21mo (at two different time intervals) out of
the 24 surveyed (Fig. la), presenting thelowest mean biomass production (0.15 g/m3DW) of all four sites (Fig. 2b). Tbe highestmonthly biomass value of 1.33 glm3 (DW)was recorded in August 1998, when the sa-linity was 170 gIL, percent O2 saturationranged from 56 to 99%, and N03 - was11.75 mgIL (Tabie 1).
At Salina Cero the proportion of adults(males and females) to the juveniles andnauplii was lower than that of the four sites.From September 1998 to June 2000, meanpopulation composition was 11: 8:26:55 (F:M:J:N), and the F:M ratio was 1:0.84.Twenty-two inspections were conducted,collecting Artemia biomass in only three ofthem, although this site presented the high-est mean biomass production (0.28 glm3DW) of all sites surveyed (Fig. 2b). SalinaCero had the lowest monthly biomass pro-duction (Fig. lb), but reproduction experi-ments indicated that Salina Cero presentedthe highest mean cyst producûon of all foursites. In June 1999, monthly biomass pro-duction reached its maximum value with
0.53 glm3 (DW), salinity was 22 gIL, per-cent O2 saturation of 84%, and N03 - was4.85 mgIL (Tabie I).
At Tayrona the mean population distri-bution from November 1998 to June 2000
presented a high composition of adults(males and females) compared to that of ju-veniles and nauplius being 36:46:10:8 (F:M:J:N), but with a low F:M ratio of 0.88:I, contrary to the other three populations.Eighteen sampling campaigns were con-ducted, collecting Artemia biomass in onlysix occasions, and very rarely cysts. Tbissite presented the second highest mean bio-mass production (0.27 glm3 DW) of all sitessurveyed (Fig. 2b). Production reached itsmaximum value (Fig. Ic) in September1999 with a monthly biomass of 0.99 glm3(DW), salinity was 55 gIL, very low per-cent O2 saturation of 23.3% and N03 - was3.7 mgIL (Tabie 1).
At Galerazamba the population distribu-tion, between July 1998 and April 2000,was 20:16:26:38 (F:M:J:N) and F:M ratio
a
I
I., .
I
I
:, I \: I
: I,
b
, ! ! \ \
.
: h !
u I
u . .
I
. f I
. I.
.
.. i I.
.I I..

..
. (I
Vf'
~..../'
~.. J~~,::
~j:""_.
280 CAMAROO I!T AL
was 1:0.88. Artemia biomass was coUeeted
in the same pond, in 19 out of 24 samplingcampaigns realized in this saltwork; thus,achieving the most constant biomass pro-duction estimates of all sites. This site pre-sented a low mean biomass production(0.16 glm3 DW) of all sites sampled (Fig.2b). This saltwork showed an increase inbiomass and salt production in the seeondsemester of the seeond sampling year. InJanuary 2000, monthly biomass productionreached its maximum value (Fig. ld) with0.79 glm3 (DW), salinity was 120 gIL, per-cent O2 saturation of 91%, and N03- was9.8 mgIL (Tabie 1).
The results of the in vitro experiment forthe number of Artemia cysts produced perfemale at two salinities (80 and 120 gIL)showed that Galerazamba, Tayrona, and Sa-lina Cero females produced more cysts(14.9, 57.8, and 127.6 cystlfemale, respec-tively) at the higher salinity (120 gIL) thanManaure (34.0 cystlfemale at 80 gIL).
The inftuence of physicochemical param-eters (Fig. 2) on mean biomass productionwas not significant (P > 0.05). Conversely,the interaction of salinity, percent O2 satu-ration, and nitrate on cyst production wassignificant (P < 0.05). Salina Cero had thehighest mean cyst production potential atthe lowest salinity, percent O2 saturation,and nitrate concentration (Fig. 2) and Tay-rona was characterized by higher salinityand nitrate concentration but low percent O2saturation. In addition, Manaure and Gal-erazamba had a relatively low cyst produc-tion potential at higher salinity, percent O2saturation, and nitrate concentrations.
Discussion
The results of this study demonstratedthat variation in Artemia biomass amongdifferent thalassohaline aquatic environ-ments in the Colombian Caribbean were
less pronounced than estimates of variationsin cyst production potential derived-fromcontrolled laboratory measurements of re-production. Variation in physicochemicalconditions between the sites exarnined did
not appear to inftuence biomass production.However, the results of this study were con-sistent with previous observations that Ar-temia populations respond to food short-ages, associated with changes in physico-chemical conditions, by increasing cyst pro-duction (Ballardin and Metalli 1963;D' Agostino and Provasoli 1968; Amat1985; Román and Rodrlguez 1986).
The relatively low proportion of adultspresented at Salina Cero (19%) may havebeen due to the relatively low salinity (Ta-bIe 1) presented by this site compared toManaure (38%), Galerazamba (36%) andTayrona (82%). Further, the high propor-tions of adults at Tayrona may have beendue to poor recruitment (J:N) resulting froma possible environmental impact at this site.
Relatively stabie physicochemical con-ditions at Manaure may have suppressedcyst production compared to the other threesites (Galerazamba, Salina Cero, and Tay-rona) which presented less stabie condi-tions. According to the criteria of Wear etal. (1986); Lake Grassmere is indicative ofa relative environmental stability which re-sults in a low cyst production and a highovoviviparity level, which might be selee-tively advantageous for Artemia to be ableto maximize the success of intraspecificcompetence. Moreover, the encysting mech-anism might cause a delay in populationgrowth that could be disadvantageous underrelatively stabie conditions (Browne 1980).Further, in the Manaure saltwork, Colombia(Camargo et al. 2000) an abundant cyst pro-duction was reported for some years (Rozoand Pinzón 1983; Bengtson and Simpson1989) before Hurricane Joan in 1988. TheftQQ«!!caus~ bL the hurricane damaged thelevees considerabÏY; therefore, ]oinÏng sev-eral evaporation ponds in the salt produc-tion circuit and causing salinity and tem-perature to ftuctuate very little for severalyears. Consequently, Artemia primarily ex-hibited the ovoviviparous mode of repro-duction for several years. Only until re-cently, salinity has increased in the sameevaporating ponds, above the tolerance lim-
Iï
I
IARTEMIA BIOMASS AND CYST PRODUCTION IN COLOMBIA
.~
it for Artemia, thus producing appreciableamounts of cysts. These types of relativelystabie environments can result in a low butpermanent reproduction capacity and a mul-tiplicity of asynchronic generations eachyear (Lenz 1987).
The results of the reproductive experi-ment (mean cyst production per female) donot entirely agree with the estimated cystproduction potential at each site and maybe due to a combination of certain param-eters (i.e., salinity, percent O2 saturation,low nitrate, and/or starvation of the adult
Artemia population after reaching a highdensity, particularly of the female portion)on cyst production, particularly in the caseof Salina Cero. AD additional explanationto these contradictory results may he due tothe controlled conditions where Artemiawas fed ad libitum (constant food avail-ability). In contrast, these conditions do not
occur naturally in the field. The latter maybe supported, in part, by the observation oflow nitrate levels during the peak cyst pro-duction potential during this study. Further,nitrate levels and cyst production show anegative trend and do not add to cyst pro-duction, thus providing evidence that foodlimitation might contribute to cyst produc-tion. According to Amat (1985) at low sa-linities and percent O2 saturation, the factorinftuencing Artemia oviparity was invari-ably hypoalimentation, particularly pro-longed starvation periods (Ballardin andMetalli 1963). Román and Rodnguez(1986) affirm that cyst production in Cadizsaltworks (Spain) is given when the chIo-rophyll a and Artemia density ratio de-creased. Similarly, according to Newman(personal communication, 2001) food quan-tity (density) might he the determining fac-tor for Artemia to select an oviparous modeof reproduction rather than food quality.There are times wherein a particularly pooralgae strain dominates and Artemia cannotbreak down the cell wall and at that pointthey will most likely switch reproductivemodes, prior to their demise. However,D' Agostino and Provasoli (1968) also rec-
281
ognized that food quality and quantitycould induce oviparity. Moreover, accord-ing to Hemandonena (1974), with increas-ing salinity, Artemia energy requirementsdecrease and protein intake for albumin in-creases.
In addition, for all four locations nitrate
levels were directly proportional to salinity(Fig. 2d, 2g), this might he explained by thefact that in saltworks numerous organismsare trapped and slowly die as salinity in-creases progressively in the evaporating ba-sins; thus, organic matter accumulates anddecomposes, both of animal (i.e., crusta-ceans, fish, insects) and plant origin (i.e.,leaves, phytoplankton). Consequently, theconcentration of the nitrogenous com-pounds, first nitrite and later nitrate(through the process of nitrification), in-creases through time as salinity increases.Similarly, Nyonje et al. (1995) showed thatin the saltworks at Gongoni (Kenya) chlo-rophyll a increased with increasing salinityand related this to elevated levels of nitrates
in the evaporation ponds. The same phe-nomenon which occurs in saltworks (chIo-rophyll a being dependent on nitrate andsalinity) also is present in the sea as a cyc1e(summer vs. winter). According to Slinn(1974) in the North-West Irish Sea, nitratedisappeared during the summer and thuswould he the limiting nutrient. Even more,the lack of nitrate limits photosynthetic ac-tivity in the surface ocean layers (Knapp-USGS in Conomos and Gross 1968).
To summarize we were able to show a
relationship between three physicochemicalparameters (nitrate, percent O2 saturation,and salinity) and Artemia cyst productionpotential for the four locations surveyed. Incontrast, no significant relationship couldbe established for any physicochemical var-iable and Artemia biomass production. Fur-ther, nitrate (chIorophyll a) and salinitywere shown to be negatively correlated tocyst production potential. The previous ob-servation may be supported by indicationsthat insufficient food plays an importantrole on cyst production (food shortage hy-

...~
,",
282 CAMAROO BT AL.ARTBMIA BIOMASS AND CYST PRODUcnON IN COLOMBIA 283
pothesis) as shown by Ballardin 80d Metalli(1963), D' Agostino 80d Provasoli (1968),Amat (1985), 80d Román 80d Rodrlguez(1986).
Finally, the implications of these resultsmight be valuable to bal80ce the conser-vation and exploitation of natural popula-tions of Artemia to supply the demand ofthe local 80d global aquaculture industry.
Acknowledgments
This study was fin80ced by a doctoratescholarship 80d a research project "Eval-uación y aprovecharniento del recurso nat-ural Artemia en las salinas de M80aure yGalerazamba, Caribe colombiano, .. direct-ed by William Camargo (code 1116-09-343-97) 80d gr80ted by the Colombi8OCouncil of Science 80d Technology "Fran-cisco José de Caldas" (COLCIENCIAS)80d by the Universidad del Atlántico, Bar-ranquilla, Colombia. The sampling field-work during the explorations 80d monthlydata collection was possible thanks to thevery valuable cooperation received fromJu80 Carlos Linero, Licet Hem8Odez, J8O-dro Bolaiio, Javier Garcia, Tania Acufia,Karime Coha, Saul Pereira, Victor Escorcia,Orl8Odo Rada 80d all the members of the
Artemia Research Group (GIA), FundaciónUniversitaria San Martin 80d Uniatlántico.
We express our most sincere gratitude toChristopher Kohier, Els Vanden Berghe,80d to the anonymous evaluators for theirrecommendations on the preparation of thispaper.
Literature Clted
Abreu-Grobols, F. A. and J. A. Beardmore. 1982.GeneIic differenIiaIion and speciaIion in Ihe brineshrimp Artemia. Pages 345-376 in C. Barigozzi,editor. Meehanisms of speciaIion. Alan R. Liss,Inc., New York, USA.
Amat, F. 1985. Biologia de Artemia. Inf Teeni InstInv Pesq 126-127.
Amat, F. D. 1982. Diferenciaci6n y distribuci6n de laspoblaciones de Artemia de Espai!a. m. Oviparis-mo y ovoviviparismo. Estudio cualitaIivo y cuan-IitaIivo. Inv Pesq 46(1):3-13.
Baker, M. J. 1966. Autecology of Artemia: factoninftuencing hemoglobin synlhesis and cyst pro-
ducIion. San Francisco: San Francisco State Col-lege.
Ballardln, E. and P. Metalll. 1963. Osservazioni sullabiologia di Artemia saUna (L.) IsIituto Lombardo.Italia, Ace di Sci E lettere 97:194-254.
Bengtson, D. A. and K. L. Slmpsoo. 1989. FacIibi-lidad de explotacion comercial de Artemia enManaure. Colombia. Reporte para Proexpo. Bo-go~ Colombia. Univenity of Rhode Island.
Berthélémy-OkazakJ. N. J. 1986. Environmental. bio-cbemical. and geneIic facton regulaIing oviparityin Artemia. Doctorallhesis. Univenity of Califor-nia, Davis. California, USA.
Bowen, S. T., H. G. Lebben, M. C. Poon, H. S.Chow, and T. CrlgllattL 1969. Tbe hemoglobinsof Artemia saUna Linee. L DeterminaIion of pbe-notype by genotype and environmenL CompBiochem Physiol 31:733-747.
Bowen, S. T., EoA. Fogarlno, K. N. IDtchner, G. L.Dana, V. H. S. Cbow, M. R. BuoncrisdanJ, andJ. R. Carl. 1985. Ecologica1isolaIion in Artemia:populaIion differences in tolerance of anion con-centraIions. 1 Crost Biol 5(1):106-129.
Browne, R. A. 1980. ReproducIive pattem and modein Ihe brine shrimp. Ecology 61(3):466-470.
Browne, R. A. 1988. Ecological and geneIic diver-gence of sexual and asexual brine shrimp (Artem-ia) from Ihe Mediterranean basin. Nat Geog Res4(4):548-554.
Browne, R. A. and C. W. Hoopes. 1990. Genotypedivenity and seleeIion in asexual brine shrimp(Artemia). EvoluIion 44(4):1035-1051.
Browne, R. A., L. EoDavls, and S. EoSallee. 1988.Effects of temperature and relaIive fitness of sex-ual and asexual brine shrimp Artemia. 1 Exp MasBiol Beol 24:1-20.
Browne, R. A., V. Moller, V. EoForbes, Bnd M. H.Depledge. 2002. EsIimaIing geneIic and environ-mental components of variance using sexual andclonal Artemia. 1 Exp Mas Biol Beo 267(1):107-119.
Browne, R. A., S. Eo Sallee, D. S. Groscb, W. O.Segreti, and S. M. Purser. 1984. ParIiIioning ge-neIic and environmental components of reproduc-ûon and lifespan in Artemia. Beology 65(3):949-960.
Cam1ll1:0, W. N. 2002. Review of Ihe biogeograpbyof Ihe genus Artemia (CrustaceB, Anostraca) inColombia. Doctorallhesis. Faculty of Agricu1ture.Univenity of Gben!, Belgium.
Camargo, W. N., P. BossIer, P. Sorg.~loos, and Y.Suo. 2002a. Pre1iminasygenetic data on some Ca-ribbean Artemia franciscana strains based onRAPD's. Hydrobiologia 468:245-249.
Camargo, W. N., K. V. Coba, and G. C. Durán.2000. Reporte final del proyeeto de invesIigaci6n:"Evaluación y aproveehamiento del recuno nat-ural Artemla en las sa1inasde Manaure y Galera-
zamba, Caribe colombiano. .. Colciencias y Univ-enidad del AtJánIico, Colombia.
Camargo, W. N., J. S. Ely, Bnd P. Sorgeloos. 2003.Morphometric cbaracterizaIion of IhalassobalineArtemia populaIions from Ihe Colombian Carib-bean. 1 Biogeography 30:1-6.
Camargo, W. N., L. Van Vooren, and P. Sorgel00s.2002b. Effeets of lunas cycles on Artemia densityin bypersaline environments. Hydrobiologia 468:251-260.
Conomos, T. J. and M. G. Gross. 1968. Mixing ofColumbia river and ocean walen in summer.ASCE proc. 1 Sanit Eng Div. Vol 94. No SAS.Pap 6187. Oet 1968, p. 979-994.
D' Agosdno, A. S. and L. Provasoll. 1968. Effeets ofsalinity and nutrients on mono- and diaxenic cul-tures of two strains of Artemia saUna. The Bio-logical BulleIin 134(1):1-14.
Dans, G. L. and P. H. Leni. 1986. Effeets of increas-ing salinity on an Artemia populaIion from MonoLake. California. Oceanologia 68:428-436.
Durán, G. C. and J. N. Garda. 2001. Inftuencia dela salinidad y temperatura en el modo de reprod-ucei6n de la Artemia (Crustacea, Anostraca) devarias localizaciones del Caribe Colombiano. B.S.Ihesis. Universidad del AtJánIico. Facultad deBiologfa, Basranquilla, Colombia.
Dutrieu, J. 1960. ObservaIions biocbimiques et pby-siologiques sur le d6veloppement d'Artemia sali-na Leacb. Arcb Zool Exp Gt!n 99:1-134.
Grant, W. D. and H. N. M. Ross. 1986. Tbc eeologyand taxonomy of halobacteria. Pages 9-15 in W.D. Grant and M. Kagu!, editors. EMBO-FEMSWorkshop on Ihe Moleeulas Basis of Haloadap-taûon in Microorganisms. Obermaschtal (PRG).
Raslert, S. J. and R. G. Wear. 1985. Biomass esIi-maIion of Artemia at Lake Grassmere. Maslbor-ough, New Zea1and Aus. 1 Mas Fresh W Res 36:537-557.
Hernandonen&, A. 1974. Effeets of salinity on nutri-Iional requirements of Artemia salina. Biol Bull146(2):238.
Lenz, P. H. 1984. Life history analysis of an ArtemiapopulaIion in a changing environmenL 1 PlanktonRes 6(6):967-983.
Lenz, P. H. 1987. Beologica1 studies on Artemia: areview. Pages 5-18 in P. Sorgeloos, D. A. Bengt-$On,W. Decleir. and E. laspen, editon. Artemiareseascb and its BpplicaIions,volume 3. Beology,culturlng, use in aquaculture. Univena Press. Wet-teren, Belgium.
Lenz, P. H. Bnd R. A. Browne. 1991. Ecology ofArtemia. Pages 237-253 in R. A. Browne, P. Sor-gelIlOs, and C. N. A. 'IIotman, editon. ArtemiaBiology. CRC Press, Inc., Boca Raton. Florida,USA.
Newmark, F. U. 1988. Compasaci6n experimental dela producei6n de biomasa y quistes de las cepas
colombianas de Artemia (Galerazamba, Manaurey Pozos Colorados) con anotaeiones sobre sus es-trategias reproducIivas. MSc Ihesis. UnivenidadNacional de Colombia. Facultad de Ciencias, SedeSanta Marta, Colombia.
Nyol\Je, B. M., D. Mung&, and J. RadulL 1995. Ben-efits of Artemia producIion in 50las saltworks: acasestudy.FisberiesSocietyof Africa.Fint PanAfrican Fisberies Congress on Sustainable Devel-opment of Fisheries in Africa, Nairobi, Kenya.Por, F. D. 1980. A classificaIion of bypenalinewaten. based on trophic criteria. Mas Ecol ProgSer 1:121-131.
Post, F. J. and N. N. Yousser. 1977. A prokaryoIicintracellulas symbiont of Ihe Great Salt Lake brineshrimp Artemia saUna (Leacb). Can 1 Microbio23(9):1232-1236.
ProvasolI, L. and I. J. Plntner. 1980. Bipbasic pas-Iieulate media for!he culture of filter-feeden. Pag-es 47-54 in G. Penoone. P. Sorgeloos, O. RoeJs.and E. laspen. editon. Tbe brine shrimp Artemia,volume 3. Ecology. culturing. use in aquaculture.Univena Press. Wetteren, Belgium.
Román, M. and A. Rodriguez. 1986. Cultivo de Ar-temia en estanques de salinas de Cádiz (Espada).Inv Pesq 50(3):407-419.
Rozo, M. and O. Plnz6o. 1983. Artemicultura en lassa1inas de Manaure y Galerazamba. Univenidadde los Andes e InsÛlUtode Fomento Industrial(IFI). Bogo~ Colombia.
Salo, N. L. 1967. EnzymaIic contribuIion to Ihe en-cystment of Artemia salina. Sci Rep Toboku Univ33:319-327.
Sllnn, D. J. 1974. Water circulaIion and nutrients inIhe Norlh-West Irlsb Sea. Liverpool Univ. (Eng-land). DepL of Marine Biology. Estuarine andCoastal Marine Science 2(1):1-25.
SOl'&eloos,P., M. Baeza-Mesa, F. BenIJts, and G.Persoone. 1975. Reseasch on Ihe culturing of Ihebrine shrimp Artemia salina L. at Ihe state uni-venity of Ghent (Belgium).
Sorgeloos, P. 1980. Ufe history of Ihe brine shrimpArtemia. Pages 19-23 in G. Penoone. P. Sorge-loos, O. RoeIs. and E. laspen. editors. Tbe brineshrimp Artemia. volume 1. Univena Press, Wet-teren, Belgium.
Vanbaecke, P., S. E. Slddall, and P. SOl'&eloos.1984.InternaIional study on Artemia xxxn Combinedeffeets of Iemperature and salinity on Ihe survivalof Artemia of VariOU5geographical origin. 1 ExpMas Biol Beol 80:259-275.
Vos, J.1979. Brine shrimp Artemia salina inoculaIionin tropica1 salt ponds; A preliminasy guide for usein Ihailand. FAO report THA/75 WPI3, 43 p.
Wear, R. G., S. J. Haslelt, and N. L. Alexander.1986. Effeets of temperature and salinity on Ihebiology of Artemia franciscana Kellogg fromLake Grassmere, New Zealand. 1 Exp Mas BiolBeol 98:167-183.











![The occurrence of the brine shrimp, Artemia franciscana ... · America [25], other Artemia species include: Artemia persimilis of South America [26], Artemia salina in the Mediterranean](https://static.fdocuments.us/doc/165x107/5c4b6a7093f3c3117d72c1b0/the-occurrence-of-the-brine-shrimp-artemia-franciscana-america-25-other.jpg)