
Influence of ADAM28 on biological characteristics of human dental follicle cells
11
Influence of ADAM28 on biological characteristics of human dental follicle cells Zheng Zhao a,b , Hongchen Liu a, *, Yan Jin c , E. Lingling a a Institute of Stomatology, General Hospital of Chinese People’s Liberation Army, 28 Fuxing Road, Beijing, 100853, China b Department of Stomatology, The 401st Hospital of Chinese People’s Liberation Army, Qingdao, 266071, China c Research and Development Centre for Tissue Engineering, Fourth Military Medical University, Xi’an, Shaanxi 710032, China 1. Introduction The dental follicle (DF) is a loose ectomesenchymally derived, connective fibrous tissue sac surrounding the enamel organ and the dental papilla of the developing tooth germ prior to eruption. The DF cells are generally believed to contain precursor cells for cementoblasts, osteoblasts and periodontal ligament cells and further have the capability to differentiate into periodontium consisting of cementum, alveolar bone and periodontal ligament. 1 The development of the tooth root and periodontium is thought to derive from the DF cells through a mechanism of epithelial–mesenchymal or extracellular matrix-mesenchymal interaction. 2 Tooth eruption is a complex and tightly regulated process involved in both odontogenic and osteogenic cells, which requires alveolar bone resorption and formation, and appears to be regulated by the DF. 3,4 Furthermore, the DF cells of a developing tooth secrete a bone resorbing factor that can archives of oral biology 54 (2009) 835–845 article info Article history: Accepted 17 May 2009 Keywords: ADAM28 HDFCs Proliferation Differentiation Apoptosis abstract Objectives: The aim of this study was to investigate the effects of a disintegrin and metallo- proteinase 28 (ADAM28) on the biological characteristics of human dental follicle cells (HDFCs) and possible action mechanism. Methods: Eukaryotic expression plasmid containing ADAM28 coding region and ADAM28 antisense oligodeoxynucleotides (AS-ODN) with FITC labelling were constructed and synthesised by gene clone and recombination. Then we respectively transfected them into HDFCs by Lipofectamine 2000 system and detected their effects on proliferation, differ- entiation and apoptosis of HDFCs by MTT assay, cell cycle detection, ALP activity and Annexin V-FITC/PI analysis. Finally we observed the effects of ADAM28 AS-ODN on HDFCs expressing extracellular matrix (ECM) proteins by immunocytochemical staining. Results: ADAM28 eukaryotic plasmid was constructed and identified successfully, and could be correctly translated and expressed in HDFCs, furthermore overexpression of ADAM28 promoted the HDFCs proliferation and inhibited specific differentiation of HDFCs, while inhibition of ADAM28 exerted the opposite effects and induced apoptosis. Moreover ADAM28 could significantly inhibit the secretion of OPN and type III collagen of HDFCs. Conclusions: ADAM28 might actively participate in the network regulation which associates HDFCs proliferation, differentiation, apoptosis with matrix mineralisation during tooth development by interacting with multiple signal molecules. Crown Copyright # 2009 Published by Elsevier Ltd. All rights reserved. * Corresponding author. Tel.: +86 010 83345786; fax: +86 010 66936254. E-mail address: [email protected] (H. Liu). available at www.sciencedirect.com journal homepage: www.intl.elsevierhealth.com/journals/arob 0003–9969/$ – see front matter . Crown Copyright # 2009 Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.archoralbio.2009.05.013
-
Upload
zheng-zhao -
Category
Documents
-
view
212 -
download
0
Transcript of Influence of ADAM28 on biological characteristics of human dental follicle cells

Influence of ADAM28 on biological characteristics of humandental follicle cells
Zheng Zhao a,b, Hongchen Liu a,*, Yan Jin c, E. Lingling a
a Institute of Stomatology, General Hospital of Chinese People’s Liberation Army, 28 Fuxing Road, Beijing, 100853, ChinabDepartment of Stomatology, The 401st Hospital of Chinese People’s Liberation Army, Qingdao, 266071, ChinacResearch and Development Centre for Tissue Engineering, Fourth Military Medical University, Xi’an, Shaanxi 710032, China
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5
a r t i c l e i n f o
Article history:
Accepted 17 May 2009
Keywords:
ADAM28
HDFCs
Proliferation
Differentiation
Apoptosis
a b s t r a c t
Objectives: The aim of this study was to investigate the effects of a disintegrin and metallo-
proteinase 28 (ADAM28) on the biological characteristics of human dental follicle cells
(HDFCs) and possible action mechanism.
Methods: Eukaryotic expression plasmid containing ADAM28 coding region and ADAM28
antisense oligodeoxynucleotides (AS-ODN) with FITC labelling were constructed and
synthesised by gene clone and recombination. Then we respectively transfected them into
HDFCs by Lipofectamine 2000 system and detected their effects on proliferation, differ-
entiation and apoptosis of HDFCs by MTT assay, cell cycle detection, ALP activity and
Annexin V-FITC/PI analysis. Finally we observed the effects of ADAM28 AS-ODN on HDFCs
expressing extracellular matrix (ECM) proteins by immunocytochemical staining.
Results: ADAM28 eukaryotic plasmid was constructed and identified successfully, and could
be correctly translated and expressed in HDFCs, furthermore overexpression of ADAM28
promoted the HDFCs proliferation and inhibited specific differentiation of HDFCs, while
inhibition of ADAM28 exerted the opposite effects and induced apoptosis. Moreover
ADAM28 could significantly inhibit the secretion of OPN and type III collagen of HDFCs.
Conclusions: ADAM28 might actively participate in the network regulation which associates
HDFCs proliferation, differentiation, apoptosis with matrix mineralisation during tooth
development by interacting with multiple signal molecules.
Crown Copyright # 2009 Published by Elsevier Ltd. All rights reserved.
avai lab le at www.sc iencedi rect .com
journal homepage: www.intl.elsevierhealth.com/journals/arob
1. Introduction
The dental follicle (DF) is a loose ectomesenchymally derived,
connective fibrous tissue sac surrounding the enamel organ
and the dental papilla of the developing tooth germ prior to
eruption. The DF cells are generally believed to contain
precursor cells for cementoblasts, osteoblasts and periodontal
ligament cells and further have the capability to differentiate
into periodontium consisting of cementum, alveolar bone and
* Corresponding author. Tel.: +86 010 83345786; fax: +86 010 66936254.E-mail address: [email protected] (H. Liu).
0003–9969/$ – see front matter . Crown Copyright # 2009 Published bdoi:10.1016/j.archoralbio.2009.05.013
periodontal ligament.1 The development of the tooth root and
periodontium is thought to derive from the DF cells through a
mechanism of epithelial–mesenchymal or extracellular
matrix-mesenchymal interaction.2
Tooth eruption is a complex and tightly regulated process
involved in both odontogenic and osteogenic cells, which
requires alveolar bone resorption and formation, and appears
to be regulated by the DF.3,4 Furthermore, the DF cells of a
developing tooth secrete a bone resorbing factor that can
y Elsevier Ltd. All rights reserved.

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5836
enhance and maintain osteoclast formation, which is needed
for a tooth eruption pathway in the alveolar bone. It is thus
clear that DF cells play an essential role in the process of tooth
root development and tooth eruption.5 Previous studies have
indicated that the biological functions of DF cells are
controlled by a network of regulatory molecules including
growth factors, extracellular matrix proteins (ECM), cytokines
and developmental genes.6,7
ADAM28 gene is a newly discovered member of ADAM
family in humans and murine with proteolytic, adhesive
properties and autocatalytic activity,8,9 which screened from
patients with congenital hypoplasia of tooth root (CHTR) by
the authors.10 CHTR is a disease of tooth root physiological
development disorder resulting from ectodermal dysplasia
and the patients usually display tooth mobility, early slough
and atonia masticatoria. According to clinical situation, CHTR
is divided into maldevelopment of root cementum and dentin,
root paramorphia such as short cone anomaly which occurs
mostly in maxillary incisors, and root attachment organ
dysplasia.11,12 So far no effective therapy has been found in the
world, and its concrete pathopoiesis mechanism still remains
unclear. The human ADAM28 gene is located in chromosome
8p21.2 and is mainly expressed in dendrite-like cells,
lymphocytes and macrophages.13 The defining feature of
the ADAM28 is a conserved modular domain organisation
consisting of a N-terminal signal sequence, a prodomain, a
metalloprotease domain (MP) and a disintegrin-like domain, a
cysteine-rich domain, an epidermal growth factor (EGF)-like
domain, a transmembrane domain and a cytoplasmic
domain.14,15 Furthermore, the MP activity is of functional
importance for extracellular matrix modelling and in the
ectodomain processing of molecules.8
ADAM28 is known to have been implicated in several
crucial biological processes, including neurogenesis, muscle
development, protein ectodomain shedding, cell–cell adhe-
sion events and release of membrane-anchored proteins.16
Researches uncovering the overexpression of ADAM28 in
non-small cell lung carcinomas and breast carcinoma
cell implicated its involvement in the interplay of various
cell–cell and cell–matrix interactions, cell proliferation, and
cell motility.17,18 Recent studies displayed that ADAM28
could promote proliferation of human breast cancer cells by
the IGF signalling, and that the IGF system has a funda-
mental role in protecting cells from apoptosis.19 These
evidence suggested that ADAM28 might be a regulatory
factor closely related to organogenesis, cell proliferation and
apoptosis.
Furthermore, our findings have proved that ADAM28 was
expressed at almost each stage of human and murine tooth
germ development with different levels, and it may participate
in this network regulation to link cell proliferation, differ-
entiation with matrix synthesis.10 However, little attention
has hitherto been paid to the effects of ADAM28 on biological
features of HDFCs from molecular level and functional
mechanisms of relatively independent odontogenic capability
of HDFCs.
Therefore, to elucidate the effects of ADAM28 gene on
biological characteristics of HDFCs, we used immunocyto-
chemistry, molecular biology and cell biology techniques to
study the roles of ADAM28 in HDFCs development.
2. Materials and methods
The nucleotide sequence data about human ADAM28 in this
study appear in the GenBank Nucleotide Sequence Databases
under accession no. NM-014265. ADAM28 polyclonal antibody
was prepared in our laboratory and the specificity and titre of
polyclonal antibody were determined undoubtedly.10 The
protocol was approved by the Ethical Committee on Human
and Animal Research of General Hospital of Chinese PLA.
2.1. Cell culture and source identification
The dental follicles were physically isolated from the lower
third molar tooth germs (uneruption) of 12-year-old patients
under stereomicroscope (Olympus, Microsystems, Japan).
The surfaces of the follicles were cleaned, rinsed in
Dulbecco’s modified Eagle’s medium (DMEM, Gibco-BRL,
Grand Island, USA) containing penicillin G (100 U/mL,
Gibco-BRL, Grand Island, USA) and streptomycin (100 mg/
mL, Gibco-BRL, Grand Island, USA) and minced into pieces.
The follicles were then digested in a solution of dispase I
(Gibco-BRL, Grand Island, USA) and type I collagenase
(0.66 mg/mL; Sigma, St.Louis, MO) for 1 h at 37 8C.20 Single
cell suspensions were generated by filtration through a 70 mm
strainer, washed with DMEM supplemented with 10% (v/v)
fetal bovine serum (FBS, Gibco, USA), and then placed into
culture flasks and cultured in 5% CO2 at 37 8C. Through
repeated differential trypsinisation as reported,21 purified
HDFCs were obtained excluding the contamination from
epithelial cells. When the primary HDFCs arrived 90%
confluence, they were digested in 0.25% trypsin and
performed subculture. The fourth passage HDFCs were used
for the following experiments. The patients gave informed
consent and donation was voluntary.
Source identification: immunocytochemical staining of
cultured HDFCs with monoclonal antibody (mAb) against
vimentin (DAKO, USA) at 1:30 dilution and mAb against
cytokeratin (DAKO, USA) at 1:150 dilution were performed
respectively.
2.2. The construction and identification of ADAM28eukaryotic expression plasmid
In the present study, human ADAM28 was extracted and
cloned from lymphocytes. A PCR product (2,327 bp) corre-
sponding to total length of ADAM28 coding region (48–2375)
was generated using gene-specific primers:
Upstream primer: 50-ATGTTGCAAGGTCTCCTGCCAGT-
CAGTCTC-30 (amino acids 48–77); Downstream primer: 50-
TCATGCTTTTGGATTTGAGTCCTTAGGTGTAGACA-30 (amino
acids 2375–2341). PCR product was separated by 1% agarose
gel electrophoresis, visualised with ethidium bromide staining
and purified by NucleoTrap Gel Extraction Kit (Clontech, USA).
A 2327 bp fragment was cloned into the corresponding sites
of a pMD18-T vector (Takara, Japan) to construct pMD18-T-
ADAM28. The positive clones containing the target inserts
were sequenced prior to transformation into Escherichia coli
DH5a. DNA homology search was performed on the National
Center for Biotechnology Information (NCBI) server with
BLAST (Basic Local Alignment Search Tool) 2.0 program.

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5 837
Total length of human ADAM28 coding region was excised
from pMD18-T-ADAM28 using EcoR I and Sal I, and ligated into
pcDNA3.1(+) to construct the recombinant plasmid
pcDNA3.1(+)-ADAM28 with EcoR I and Not I digestion. The
eukaryotic expression plasmid was verified by PCR, restriction
endonuclease digestion identification and DNA sequencing.
Meanwhile, glyceraldehyde-3-phosphate dehydrogenase
(GAPDH, 900 bp) was used as a control, specific primers were
as follows: forward: 50-AGCCGCATCTTCTTTTGCGTC-30;
reverse: 50-TCATATTTGGCAGGTTTTTCT-30.
2.3. Transfection of HDFCs with ADAM28 eukaryoticplasmid and expression detections
Experimental cells were divided into three groups according to
pcDNA3.1(+)-ADAM28 group, pcDNA3.1(+) group with
30 mmol/L concentration and untransfected group.
Transient transfection was performed using Lipofecta-
mineTM 2000 system. At 1 day before transfection, the
fourth passage HDFCs were seeded in 24-well culture
plates (Nunc A/S, USA) at a cell density of 5 � 106 with
coverslips put in partial wells, 75 cm2 culture flasks (Gibco,
USA) and 96-well plates (Gibco, USA). Cells were cultivated
in the DMEM/F12 containing 10% FBS but not containing
penicillin G (100 U/mL) and streptomycin (100 mg/mL). At 4 h
before transfection, DMEM/F12 not containing FBS was
added into flasks. 1.0 mg DNA of each well was diluted with
50 mL DMEM/F12 not containing FBS, meanwhile 2 mL
LipofectamineTM 2000 (Invitrogen, USA) of each well
was diluted with 50 mL DMEM/F12 not containing FBS.
After 5 min of incubation at room temperature, diluted
DNA and LipofectamineTM 2000 reagent were combined
and incubated at room temperature for 20 min to form
the DNA-Lipofectamine 2000 complex. Then, 1 mL of
complex was added into 24-well culture plates and flasks
respectively. After 6 h of transfection, routine DMEM/F12
containing 10% FBS was changed into the plates and flasks.
After 48 h, cells on coverslips were washed with PBS, fixed in
4% paraformaldehyde for 2 h and then subjected to
immunofluorescence staining. The procedures were as
follows: After extensive washing with PBS, cells were
incubated with normal goat serum at 37 8C for 30 min and
then exposed to the ADAM28 antibody (1:200 dilution)
overnight at 4 8C. Thereafter, the cells were washed three
times with PBS and incubated with FITC-labelled goat anti-
rabbit IgG (1:50 dilution, Santa Cruz, USA) at 37 8C for
30 min. After being washed with PBS, fluorescence was
observed under microscope (Olympus, Microsystems,
Japan).
The other cells in 24-well plates and 75 cm2 flasks were
collected and used for RT-PCR, Western blot (Western blotting
kit, Chemicon International, Temecula, CA, USA) and cell cycle
detections. GAPDH was used as an endogenous control.
Meanwhile labworks software of UVP gel image analysis
system was used to detect the greyscales of all PCR bands.
Alpha ImagerTM 1220 image analysis system was used to
detect the greyscales of Western blot bands. Relative greyscale
analysis of gene expression was calculated by the Delta CT
method with ADAM28/GAPDH, which was normalised to the
GAPDH controls.
2.4. The design and transfection of ADAM28 AS-ODN
The nucleotides of 20nt specifically targeting human ADAM28
mRNA (GenBank no. NM-014265) were designed and synthe-
sised as ADAM28 AS-ODN (50-GG CAG GAG ACC TTG CAA CAT-
30, 67–48), meanwhile the sequences of 20nt were designed as
ADAM28 S-ODN (50-CAG TCT CCT CCT CTC TGT TG-30, 71–90).
The sequences were subjected to sulphur modification and
FITC fluorescence labelling (Takara, Japan), and confirmed to
have satisfactory specificity by NCBI/BLAST 2.0 database
search. Experimental concentration was 30 mmol/L. Trans-
fection procedures were the same as mentioned before.
Experimental cells were divided into three groups according
to the ADAM28 AS-ODN group, S-ODN group with 30 mmol/L
transfection concentration and untransfected group. Trans-
fection efficiency was observed under fluorescence micro-
scope after transfected 48 h, and transfection
efficiency = fluorocyte number at the same eyeshot/total
cellular score at the same eyeshot under inverted phase
contrast microscope � 100%. The inhibition effect of ADAM28
AS-ODN on HDFCs was determined by immunocytochemical
(LsABC kit, DAKO, USA) staining, RT-PCR and Western blot
assays.
Immunocytochemical staining and image analysis were
performed as described previously.10 The average greyscale
(means � S.D.) and relative greyscale from RT-PCR and
Western blot assays of each group were obtained, and the
results were subjected to statistical analysis.
2.5. Cell proliferation assay
The cell density was kept at 5 � 106 using for the detection of
MTT, cell cycle, apoptosis and ALP activity. Cell proliferation
assay was performed in all five groups of HDFCs using MTT [3-
(4,5-dimethylthiazol-2-yl)-2, 5-diphenyl-2H-tetrazoliumbro-
mide] method, respectively 20 mL (5 mg/mL) of MTT was
added into each well of 96-well plate for incubation 4 h at
37 8C, after 48 and 72 h of transfection. Then the supernatant
was removed and 150 mL of dimethylsulfoxide (DMSO) was
added following 10 min of oscillation. The optical density (OD)
value was determined with an enzyme-linked immunosor-
bent assay (ELISA) machine at a wavelength of 490 nm and the
assay was repeated for five times. The data were presented as
the means � S.D.
2.6. Cell cycle detection
The cells after 48 h and 72 h of transfection were performed
cell cycle analysis. The cellular DNA content was determined
by flow cytometry, as described previously.22 We collected the
floating and attached cells using trypsin-EDTA and resus-
pended them in DMEM (no FBS). The cells were fixed for 30 min
in an ice-cold 70% ethanol solution containing ribonuclease
(RNase; 2 mg/mL). We washed the cells in PBS, and then
stained them with propidium iodide (PI) for 30 min at room
temperature in the dark. The PI-elicited fluorescence was
measured for individual cells using a FACSCalibur flow
cytometer (Becton Dickinson, Tokyo, Japan) with laser excita-
tion at 488 nm. We analysed a total of 5 � 106 cells for each
sample and determined the percentages of cells in G0/G1, S and
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5838
G2/M phases using standard ModiFit and Cell Quest software
(Becton Dickinson Biosciences).
2.7. Apoptosis analysis by Annexin V-FITC/PI assay
The HDFCs of S-ODN group, AS-ODN group and untrasfected
group were cultured in DMEM (no FBS) at 4 h before
transfection procedure. After 48 or 72 h of transfection the
cells were harvested by trypsin-EDTA treatment, washed with
cold PBS and stained with PI and fluorescein isothiocyanate
(FITC)-conjugated Annexin V using an Annexin V-FITC
Apoptosis Detection Kit I (Becton Dickinson). Annexin V-FITC
identifies cells in early apoptosis by detecting externalised
phosphatidylserine, and PI identifies cells that have lost
plasma membrane integrity (i.e., necrotic or late apoptotic
cells). The cells were resuspended in 50 mL of 1� binding buffer
supplemented with 5 mL of Annexin V-FITC and 10 mL of PI,
and kept at room temperature in the dark for 15 min according
to the manufacturer’s instructions. Following the addition of
450 mL of 1� binding buffer, the stained cells were kept on ice
and subjected to fluorescence-activated cell sorter (FACS)
analysis using a FACSCalibur flow cytometer (BD Biosciences,
San Diego, CA, USA) with Cell Quest software (BD Biosciences).
We measured the FITC fluorescence between 515 and 545 nm
and the PI fluorescence between 564 and 606 nm.
2.8. Alkaline phosphatase (ALP) activity detection
After 48 and 72 h of transfection respectively, the cells of all
five groups were washed three times with 0.1 M PBS and 50 mL
of cold 10 mM Tris–HCl buffer (pH 7.4) containing 0.1%
TritonX-100 that was added before incubation at 4 8C over-
night. ALP substrate solution (100 mL) containing 2 mM MgCl2and 16 mM p-nitrophenyl phosphate was then mixed with
each sample. After incubation at 37 8C for 30 min, the reaction
was stopped by adding 50 mL of 0.2 M NaOH and the liberated
p-nitrophenol was measured spectrophotometrically at
410 nm. Each experiment was repeated for five times and
the data were presented as the means � S.D.
2.9. Effects of ADAM28 AS-ODN on HDFCs expressingECM proteins
Immunocytochemical (LsABC kit, DAKO, USA) staining and
image analysis were performed as described above. Polyclonal
antibody against bone sialoprotein (BSP), osteopontin (OPN)
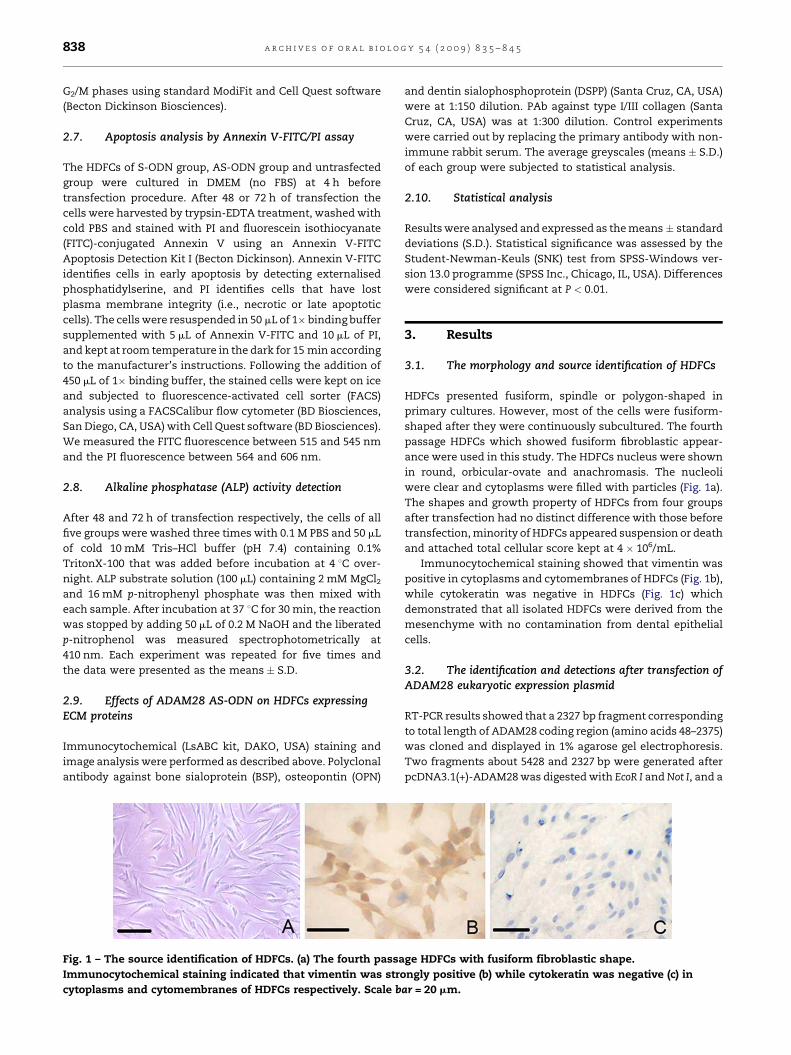
Fig. 1 – The source identification of HDFCs. (a) The fourth passa
Immunocytochemical staining indicated that vimentin was stro
cytoplasms and cytomembranes of HDFCs respectively. Scale b
and dentin sialophosphoprotein (DSPP) (Santa Cruz, CA, USA)
were at 1:150 dilution. PAb against type I/III collagen (Santa
Cruz, CA, USA) was at 1:300 dilution. Control experiments
were carried out by replacing the primary antibody with non-
immune rabbit serum. The average greyscales (means � S.D.)
of each group were subjected to statistical analysis.
2.10. Statistical analysis
Results were analysed and expressed as the means � standard
deviations (S.D.). Statistical significance was assessed by the
Student-Newman-Keuls (SNK) test from SPSS-Windows ver-
sion 13.0 programme (SPSS Inc., Chicago, IL, USA). Differences
were considered significant at P < 0.01.
3. Results
3.1. The morphology and source identification of HDFCs
HDFCs presented fusiform, spindle or polygon-shaped in
primary cultures. However, most of the cells were fusiform-
shaped after they were continuously subcultured. The fourth
passage HDFCs which showed fusiform fibroblastic appear-
ance were used in this study. The HDFCs nucleus were shown
in round, orbicular-ovate and anachromasis. The nucleoli
were clear and cytoplasms were filled with particles (Fig. 1a).
The shapes and growth property of HDFCs from four groups
after transfection had no distinct difference with those before
transfection, minority of HDFCs appeared suspension or death
and attached total cellular score kept at 4 � 106/mL.
Immunocytochemical staining showed that vimentin was
positive in cytoplasms and cytomembranes of HDFCs (Fig. 1b),
while cytokeratin was negative in HDFCs (Fig. 1c) which
demonstrated that all isolated HDFCs were derived from the
mesenchyme with no contamination from dental epithelial
cells.
3.2. The identification and detections after transfection ofADAM28 eukaryotic expression plasmid
RT-PCR results showed that a 2327 bp fragment corresponding
to total length of ADAM28 coding region (amino acids 48–2375)
was cloned and displayed in 1% agarose gel electrophoresis.
Two fragments about 5428 and 2327 bp were generated after
pcDNA3.1(+)-ADAM28 was digested with EcoR I and Not I, and a
ge HDFCs with fusiform fibroblastic shape.
ngly positive (b) while cytokeratin was negative (c) in
ar = 20 mm.
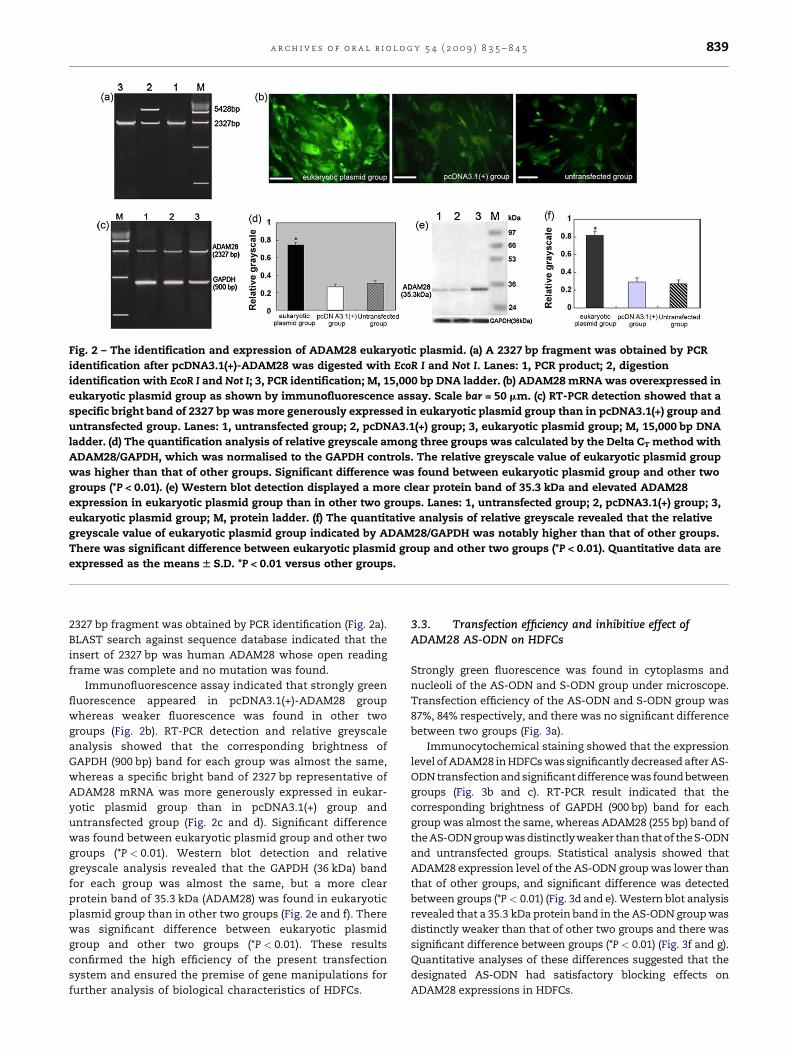
Fig. 2 – The identification and expression of ADAM28 eukaryotic plasmid. (a) A 2327 bp fragment was obtained by PCR
identification after pcDNA3.1(+)-ADAM28 was digested with EcoR I and Not I. Lanes: 1, PCR product; 2, digestion
identification with EcoR I and Not I; 3, PCR identification; M, 15,000 bp DNA ladder. (b) ADAM28 mRNA was overexpressed in
eukaryotic plasmid group as shown by immunofluorescence assay. Scale bar = 50 mm. (c) RT-PCR detection showed that a
specific bright band of 2327 bp was more generously expressed in eukaryotic plasmid group than in pcDNA3.1(+) group and
untransfected group. Lanes: 1, untransfected group; 2, pcDNA3.1(+) group; 3, eukaryotic plasmid group; M, 15,000 bp DNA
ladder. (d) The quantification analysis of relative greyscale among three groups was calculated by the Delta CT method with
ADAM28/GAPDH, which was normalised to the GAPDH controls. The relative greyscale value of eukaryotic plasmid group
was higher than that of other groups. Significant difference was found between eukaryotic plasmid group and other two
groups (*P < 0.01). (e) Western blot detection displayed a more clear protein band of 35.3 kDa and elevated ADAM28
expression in eukaryotic plasmid group than in other two groups. Lanes: 1, untransfected group; 2, pcDNA3.1(+) group; 3,
eukaryotic plasmid group; M, protein ladder. (f) The quantitative analysis of relative greyscale revealed that the relative
greyscale value of eukaryotic plasmid group indicated by ADAM28/GAPDH was notably higher than that of other groups.
There was significant difference between eukaryotic plasmid group and other two groups (*P < 0.01). Quantitative data are
expressed as the means W S.D. *P < 0.01 versus other groups.
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5 839
2327 bp fragment was obtained by PCR identification (Fig. 2a).
BLAST search against sequence database indicated that the
insert of 2327 bp was human ADAM28 whose open reading
frame was complete and no mutation was found.
Immunofluorescence assay indicated that strongly green
fluorescence appeared in pcDNA3.1(+)-ADAM28 group
whereas weaker fluorescence was found in other two
groups (Fig. 2b). RT-PCR detection and relative greyscale
analysis showed that the corresponding brightness of
GAPDH (900 bp) band for each group was almost the same,
whereas a specific bright band of 2327 bp representative of
ADAM28 mRNA was more generously expressed in eukar-
yotic plasmid group than in pcDNA3.1(+) group and
untransfected group (Fig. 2c and d). Significant difference
was found between eukaryotic plasmid group and other two
groups (*P < 0.01). Western blot detection and relative
greyscale analysis revealed that the GAPDH (36 kDa) band
for each group was almost the same, but a more clear
protein band of 35.3 kDa (ADAM28) was found in eukaryotic
plasmid group than in other two groups (Fig. 2e and f). There
was significant difference between eukaryotic plasmid
group and other two groups (*P < 0.01). These results
confirmed the high efficiency of the present transfection
system and ensured the premise of gene manipulations for
further analysis of biological characteristics of HDFCs.
3.3. Transfection efficiency and inhibitive effect ofADAM28 AS-ODN on HDFCs
Strongly green fluorescence was found in cytoplasms and
nucleoli of the AS-ODN and S-ODN group under microscope.
Transfection efficiency of the AS-ODN and S-ODN group was
87%, 84% respectively, and there was no significant difference
between two groups (Fig. 3a).
Immunocytochemical staining showed that the expression
level of ADAM28 in HDFCs was significantly decreased after AS-
ODN transfection and significant difference was found between
groups (Fig. 3b and c). RT-PCR result indicated that the
corresponding brightness of GAPDH (900 bp) band for each
group was almost the same, whereas ADAM28 (255 bp) band of
the AS-ODN group was distinctly weaker than that of theS-ODN
and untransfected groups. Statistical analysis showed that
ADAM28 expression level of the AS-ODN group was lower than
that of other groups, and significant difference was detected
between groups (*P < 0.01) (Fig. 3d and e). Western blot analysis
revealed that a 35.3 kDa protein band in the AS-ODN group was
distinctly weaker than that of other two groups and there was
significant difference between groups (*P < 0.01) (Fig. 3f and g).
Quantitative analyses of these differences suggested that the
designated AS-ODN had satisfactory blocking effects on
ADAM28 expressions in HDFCs.
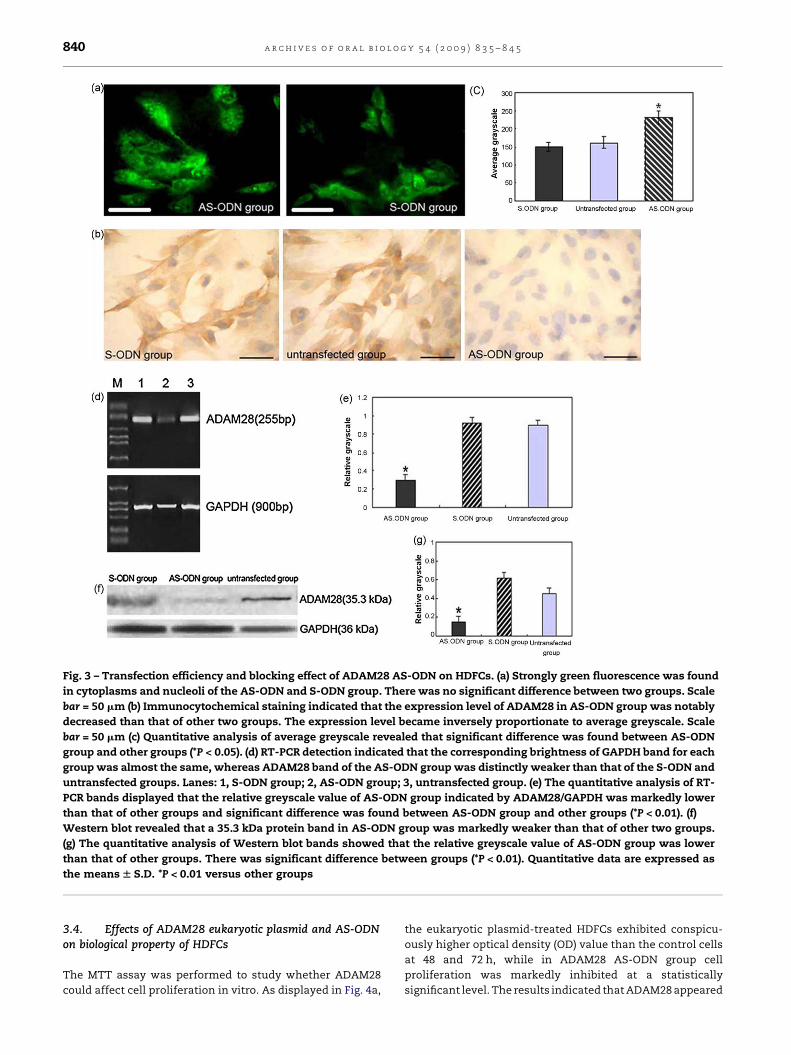
Fig. 3 – Transfection efficiency and blocking effect of ADAM28 AS-ODN on HDFCs. (a) Strongly green fluorescence was found
in cytoplasms and nucleoli of the AS-ODN and S-ODN group. There was no significant difference between two groups. Scale
bar = 50 mm (b) Immunocytochemical staining indicated that the expression level of ADAM28 in AS-ODN group was notably
decreased than that of other two groups. The expression level became inversely proportionate to average greyscale. Scale
bar = 50 mm (c) Quantitative analysis of average greyscale revealed that significant difference was found between AS-ODN
group and other groups (*P < 0.05). (d) RT-PCR detection indicated that the corresponding brightness of GAPDH band for each
group was almost the same, whereas ADAM28 band of the AS-ODN group was distinctly weaker than that of the S-ODN and
untransfected groups. Lanes: 1, S-ODN group; 2, AS-ODN group; 3, untransfected group. (e) The quantitative analysis of RT-
PCR bands displayed that the relative greyscale value of AS-ODN group indicated by ADAM28/GAPDH was markedly lower
than that of other groups and significant difference was found between AS-ODN group and other groups (*P < 0.01). (f)
Western blot revealed that a 35.3 kDa protein band in AS-ODN group was markedly weaker than that of other two groups.
(g) The quantitative analysis of Western blot bands showed that the relative greyscale value of AS-ODN group was lower
than that of other groups. There was significant difference between groups (*P < 0.01). Quantitative data are expressed as
the means W S.D. *P < 0.01 versus other groups
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5840
3.4. Effects of ADAM28 eukaryotic plasmid and AS-ODNon biological property of HDFCs
The MTT assay was performed to study whether ADAM28
could affect cell proliferation in vitro. As displayed in Fig. 4a,
the eukaryotic plasmid-treated HDFCs exhibited conspicu-
ously higher optical density (OD) value than the control cells
at 48 and 72 h, while in ADAM28 AS-ODN group cell
proliferation was markedly inhibited at a statistically
significant level. The results indicated that ADAM28 appeared
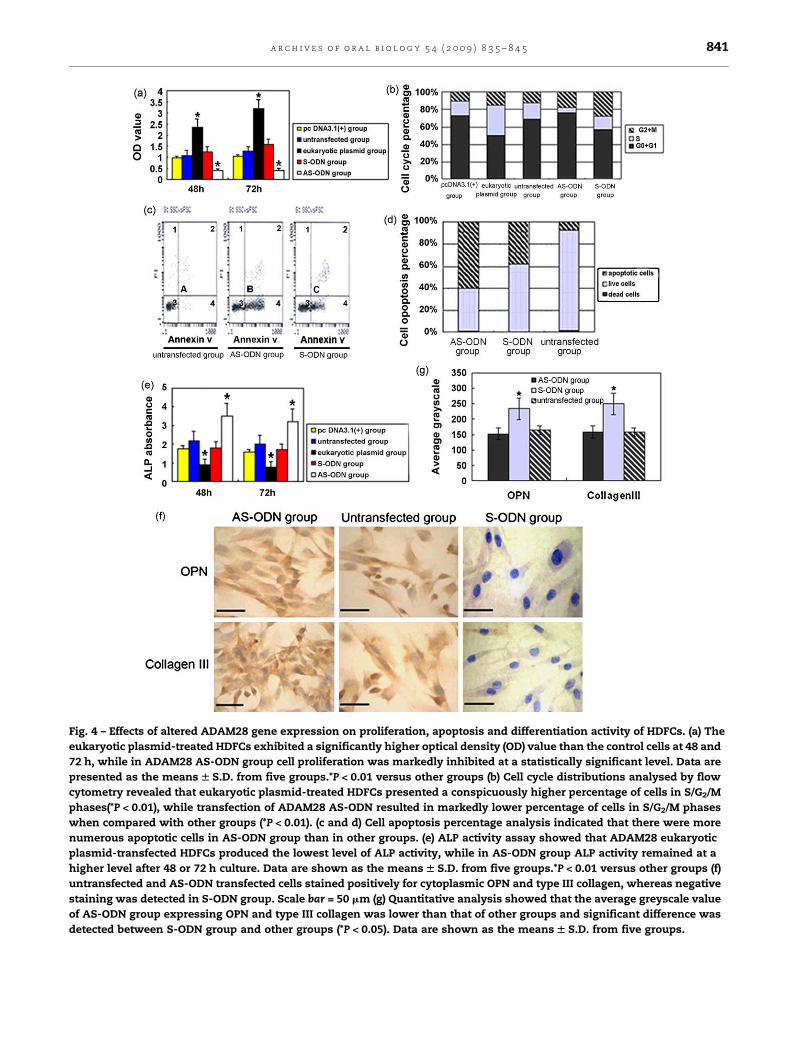
Fig. 4 – Effects of altered ADAM28 gene expression on proliferation, apoptosis and differentiation activity of HDFCs. (a) The
eukaryotic plasmid-treated HDFCs exhibited a significantly higher optical density (OD) value than the control cells at 48 and
72 h, while in ADAM28 AS-ODN group cell proliferation was markedly inhibited at a statistically significant level. Data are
presented as the means W S.D. from five groups.*P < 0.01 versus other groups (b) Cell cycle distributions analysed by flow
cytometry revealed that eukaryotic plasmid-treated HDFCs presented a conspicuously higher percentage of cells in S/G2/M
phases(*P < 0.01), while transfection of ADAM28 AS-ODN resulted in markedly lower percentage of cells in S/G2/M phases
when compared with other groups (*P < 0.01). (c and d) Cell apoptosis percentage analysis indicated that there were more
numerous apoptotic cells in AS-ODN group than in other groups. (e) ALP activity assay showed that ADAM28 eukaryotic
plasmid-transfected HDFCs produced the lowest level of ALP activity, while in AS-ODN group ALP activity remained at a
higher level after 48 or 72 h culture. Data are shown as the means W S.D. from five groups.*P < 0.01 versus other groups (f)
untransfected and AS-ODN transfected cells stained positively for cytoplasmic OPN and type III collagen, whereas negative
staining was detected in S-ODN group. Scale bar = 50 mm (g) Quantitative analysis showed that the average greyscale value
of AS-ODN group expressing OPN and type III collagen was lower than that of other groups and significant difference was
detected between S-ODN group and other groups (*P < 0.05). Data are shown as the means W S.D. from five groups.
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5 841

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5842
to have a positive regulatory effect on the proliferation of
HDFCs.
Cell cycle distributions analysed by flow cytometry
revealed that eukaryotic plasmid-treated HDFCs presented a
notably higher percentage of cells in S/G2/M phases (Fig. 4b,
*P < 0.01), while transfection of ADAM28 AS-ODN resulted in
markedly lower percentage of cells in S/G2/M phases when
compared with other groups (Fig. 4b, *P < 0.01), demonstrating
that the deletion of ADAM28 could inhibit the proliferation of
HDFCs.
To detect whether this effect was related to the changes in
cell survival, cell apoptosis assay was carried out which
suggested that there were constitutively more numerous
apoptotic cells in ADAM28 AS-ODN group (Fig. 4c and d).
In contrast to the cell proliferation assay, ALP activity
analysis displayed a reverse trend that ADAM28 eukaryotic
plasmid-transfected HDFCs produced the lowest value of ALP
absorbance while AS-ODN group kept in a higher level after 48
or 72 h culture (Fig. 4e). The data showed that ADAM28 could
significantly inhibit ALP activity of HDFCs.
3.5. Effects of ADAM28 AS-ODN on HDFCs expressingECM proteins
Immunocytochemical staining results indicated that OPN and
type III collagen were stained positively in cytoplasms of
untransfected and AS-ODN transfected cells, whereas nega-
tive staining was shown in S-ODN transfected cells (Fig. 4f).
Quantitative analysis revealed that significant difference was
found between S-ODN group and other groups (Fig. 4g,
*P < 0.05). These results indicated that ADAM28 could sig-
nificantly inhibit the expressions of OPN and type III collagen
and present negative correlation with them.
4. Discussion
The multiple functions possessed by ADAM family members
suggest many potential roles for ADAM28. Our recent
researches about human ADAM28 continuous expressions
in tooth germ, ameloblasts, dental papilla cells and dental
follicle cells suggested that ADAM28 was involved in crown
and root morphogenesis process.10 The initiation of root
formation is determined by Hertwig’s epithelial root sheath
(HERS), which induces the differentiation of odontoblasts
forming root dentine.11 Moreover HERS and dental follicle cells
were stained positively from bell stage, which suggested that
ADAM28 could play a major role in maintaining the structure
of the dental follicle and the inducement to cervical loop, both
as a substrate and as a reservoir of paracrine molecules.10
Consistent with above reports, our present study demon-
strates convincingly the expression and localisation of
ADAM28 in HDFCs and further validated that ADAM28 might
function as a potential regulator involved in the proliferation,
specific differentiation and apoptosis of HDFCs.
In the present study, ADAM28 eukaryotic expression
system and antisense oligodeoxynucleotides (AS-ODN)
technique with reverse function were applied successfully.
ADAM28 could be correctly transcribed, translated and
expressed after transfected into HDFCs. Antisense oligonu-
cleotide biotechnology provides an efficient and specific
method for inhibiting gene expression and exploring gene
function, and multiple application of antisense inhibitory
experimental strategies have been leveraged to facilitate the
understanding of the signals involved in inductive epithe-
lial–mesenchymal interactions during tooth morphogen-
esis.23–25
Then we demonstrated that ADAM28 had an up-regula-
tive effect on the proliferation of HDFCs by MTT assay and
cell cycle detection. Cell proliferative kinetics is an essential
feature during tooth morphogenesis since high cell prolif-
erative activity is desirable for rapid increase in volume of
the tooth germ and tooth cusp formation.26 Cell cycle
detection revealed that more cells accumulated in S phase
(DNA synthesis phase) of pcDNA3.1(+)-ADAM28 group,
which resulted in the elevation of total PI value (cell
proliferation index, S + G2 + M) distinguishable from those
of control groups. The critical step required for the
proliferation of cells is their transition from cell cycle arrest
to entry into the active phase of the cell cycle through the G1/
S restriction point, thus the fast transition can enhance
higher rates of cell proliferation.27 Above results would argue
that ADAM28 facilitate the HDFCs proliferation by promoting
the transition process from G1 to S phase, further accelerat-
ing DNA reproduction and protein synthesis of HDFCs. This
is probably due to the process that ADAM28 eukaryotic
plasmid might enhance its catalytic activity of metallopro-
teinase, cleave matrix protein and rebuild tissue structure to
regulate cell proliferation liveness.28 Moreover, ADAM28
comprises an EGF-like domain, which may possess certain
functions of EGF such as the stimulation or maintenance of
undifferentiated cell proliferation during embryonic devel-
opment.29 Thus, our results verified beyond doubt the causal
relationship between ADAM28 and HDFCs proliferation.-
Bone-specific alkaline phosphatase is regarded as a non-
collagenous protein secreted by osteoblasts that is essential
for bone mineralisation such as cementum and considered
to be a highly specific marker of osteoblast differentiation
function.30,31 Of particular importance in mineralisation are
the high concentrations of ALP found in matrix vesicles,
which in dentine are the sites of initial crystal formation,
proposed to be involved in the formation of mantle dentine
and also in maturation of the extracellular matrix.32 In
addition, during dentinogenesis, the extracellular matrix
synthesised by the odontoblasts coincides with their
expression level of ALP.33 Accordingly, ALP activity is
generally used for an important reference index in detecting
odontogenic mesenchymal cytodifferentiation and matrix
mineralisation.34
In this study, it seemed that the overexpression of ADAM28
inhibited the differentiation of HDFCs while the use of
ADAM28 AS-ODN produced opposite effect, as indicated by
ALP activity. These evidences suggested that ADAM28, as an
important regulator between the epithelium and mesench-
yme,10 might participate in early formation, proliferation and
differentiation start of HDFCs.10
Cytodifferentiation expressed by ALP activity, and cell
proliferation evaluated by the MTT assay and cell cycle
analysis, coincidentally demonstrated that ADAM28 could
promote HDFCs proliferation during the process of differ-

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5 843
entiation inhibition, which was in accordance with the
previous concept that proliferation and differentiation are
inversely correlated with each other due to the existence of
dual-function regulators participating in controlling both the
processes.35 Therefore, it is reasonably proposed that some
components of ADAM28 could act not only as promoters for
the proliferation of HDFCs but also as inhibitors for cytodiffer-
entiation.
Apoptosis is a key process in the embryological develop-
ment of the tooth, periodontal ligament and supporting oral
tissue in the progression of oral disease, bone resorption,
immunological response and inflammation, and in wound
healing and certain pharmacological effects.36 Cell apoptosis
and proliferation are interrelated and interact with each
other in tooth development, and they both participate in
sculpturing the shape of teeth. Previous work displayed that
epithelial–mesenchymal tissue interactions prevent apop-
tosis. Moreover, apoptosis represents an important process
in teeth morphogenesis and remodelling during tooth
development. Understanding of apoptosis regulation in
the vestigial tooth primordia can help to elucidate the
mechanism of their suppression during evolution and to
identify factors essential for tooth survival.37 Hence, apop-
tosis may be a general mechanism for the silencing of
embryonic signalling centres including odontogenesis signal
transmission.38
In this study, it was shown that transfection of AS-ODN
could notably induce apoptosis of HDFCs which strongly
suggested ADAM28 might actively participate in the negative-
feedback regulation mechanism of apoptotic event in the
mesenchyme during tooth morphogenesis. Nevertheless,
there is much to be learned about its relationship and
interaction with other signalling molecules relevant to
apoptosis in tooth development. Of particular interest has
been the application of antisense inhibitory experimental
strategies to advance a suggested causal relationship between
signal transduction and inductive epithelial–mesenchymal
interactions.23
The extracellular matrix (ECM) is a biologically active tissue
composed of a complex mixture of macromolecules, that in
addition to serving a structural function, also profoundly
affect the cellular physiology of an organism.39 In addition, the
ECM relays multiple signals from the cell microenvironment to
direct proliferation and differentiation during tissue develop-
ment,40 and plays significant roles in tissue physiology
through interaction with cells and interstitial fluid transport.
These roles include regulating cell morphology, growth and
intercellular signaling.41,42 Cell adhesion, migration, prolifera-
tion and differentiation are examples of biological processes
influenced by the composition and structural organisation of
surrounding extracellular matrices.43
It is well known that osteopontin, bone sialoprotein (BSP)
and type III collagen are the ECM molecules which are
thought to be closely related to cytodifferentiation, matrix
secretion, mineralisation and bone formation, OPN and BSP
have also been regarded as critical markers for osteogenic
differentiation.44 Bone sialoprotein and osteopontin (OPN)
are two major mineralised tissue-specific, non-collagenous
matrix glycoproteins that are almost expressed exclusively
in bone and other mineralised connective tissues, such as
dentine, cementum, alveolar bone and calcified cartilage
tissue.45–47 Subsequent studies have demonstrated that BSP
mRNA expression is associated with the onset of miner-
alisation and is essentially restricted to differentiated
osteoblasts, osteoclasts, cementoblasts, odontoblasts, ame-
loblasts and hypertrophic chondrocytes.48,49 OPN gene
expression with several proposed functions has been
associated with cell transformation and metastases in vivo
and in vitro.50,51 In bone formation studied in vitro, OPN is
expressed before the formation of a mineralised collagenous
matrix52 but at much higher concentrations after bone
formation, indicating a prominent role for OPN in both the
formation and resorption of bone.53 This divergent pattern
of OPN/BSP expression could be an important determinant
for the different characteristics of these two types of bone
metastasis, which is consistent with the proposed role of
OPN in differentiation and activation of osteoclasts and of
BSP as a stimulator of bone mineralisation.44 Thus, OPN and
BSP are believed to serve a vital role in osteogenic
differentiation, formation and remodelling of the miner-
alised tissue matrix.54
Type I and Type III collagen are regarded as important
factors mediating chemotaxis, cell attachment and migration.
They probably serve as developmental signals, which may be
critical to the regulation of interactions between periodontal
fibroblasts and the root surfaces, and thus influence the
regeneration and repair of the periodontium.54 Moreover, type
III collagen is present in reticular fibres which provide
elasticity to tissues and the characteristics that type III
collagen imparts to tissues in vivo make it a worthwhile
molecule to study for tissue engineering applications includ-
ing odontogenic tissue and cell regeneration.26
As mentioned above, the fact that DF cells finally generate
the three types of periodontal structure in a concomitant
manner renders it as a good model for study of proliferation,
differentiation and mineralisation process.7,54 Accordingly,
OPN, BSP and type III collagen were used for evaluation of
osteogenic differentiation and matrix formation in HDFCs in
this study. The fact that the expressions of OPN and type III
collagen were significantly enhanced due to transfection of
ADAM28 AS-ODN disclosed that the abolition of ADAM28
promoted lineage-specific differentiation of HDFCs. Maybe
there exists a negative correlation between ADAM28 and OPN/
type III collagen. As mentioned above, ADAM28 protein
contains a transmembrane domain, a cytoplasmic domain
and an EGF-like domain, which including components of the
extracellular matrix and thus might play an important role in
signalling transduction, intracellular protein maturation, or
localisation to sites of activity.16
In a word, this study provides comprehensive evidence
about expressions of ADAM28 in HDFCs and its influence on
biological features of HDFCs at transcription, translation and
protein levels, which demonstrate ADAM28, as a novel
regulator, might actively manipulate proliferation, differen-
tiation and apoptosis of HDFCs by interacting with multiple
signal molecules including ECM and other growth factors.
These data will not only be beneficial for the better prehension
and utilisation of HDFCs for regenerative dental therapy, but
also promising for developing feasible gene treatment against
congenital hypoplasia of tooth root.

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5844
Acknowledgements
This study was supported by a grant from the Nature Science
Foundation of China (Project No. 30572046).
Competing interests: None declared.
Ethical approval: Ethical Approval was given by the Ethical
Committee on Human and Animal Research of General
Hospital of Chinese PLA.
r e f e r e n c e s
1. Ten Cate AR, Sharpe PT, Roy S, Nanci A. Development of thetooth and its supporting tissues. Ten Cate’s oral histology.Development, structure, and function. sixth ed. Louis: Mosby;2003. pp. 79–110.
2. Thesleff I, Vaahtokari A, Vainio S, Jowett A. Molecularmechanisms of cell and tissue interactions during earlytooth development. Anat Rec 1996;245:151–61.
3. Cahill DR, Marks Jr SC. Tooth eruption: evidence forthe central role of the dental follicle. J Oral Pathol 1980;9:189–200.
4. Marks Jr SC, Cahill DR. Regional control by the dental follicleofalterations in alveolar bone metabolism during tootheruption. J Oral Pathol 1987;16:164–9.
5. Kawakami M, Kuroda S, Yoshida CA, Yamashita K, TakadaK. Dental follicle cell-conditioned medium enhances theformation of osteoclast-like multinucleated cells. Eur JOrthod 2000;22:675–82.
6. Thesleff I, Mikkola M. The role of growth factors in toothdevelopment. Int Rev Cytol Suppl 2002;217:93–135.
7. Sena K, Morotome Y, Baba O, Terashima T, Takano Y,Ishikawa I. Gene expression of growth differentiationfactors in the developing periodontium of rat molars. J DentRes 2003;82:166–71.
8. Primakoff P, Myles DG. The ADAM gene family: surfaceproteins with adhesion and protease activity. Trends Genet2000;16:83–7.
9. Blobel CP. ADAMs: key components in EGFR signaling anddevelopment. Nat Rev Mol Cell Biol 2005;6:32–43.
10. Zhao Z, Wen LY, Jin M, Deng ZH, Jin Y. ADAM28 participatesin the regulation of tooth development. Arch Oral Biol2006;51:996–1005.
11. Apajalahti S, Arte S, Pirinen S. Short root anomaly infamilies and its association with other dental anomalies.Eur J Oral Sci 1999;107:97–101.
12. Apajalahti S, Holtta P, Turtola L, Pirinen S. Prevalence ofshort-root anomaly in healthy young adults. Acta OdontolScand 2002;60:56–9.
13. Evans JP. Fertilin beta and other ADAMs as integrin ligands:Insights into cell adhesion and fertilization. Bioessays2001;23:628–39.
14. White JM. ADAMs: modulators of cell–cell and cell–matrixinteractions. Curr Opin Cell Biol 2003;15:598–606.
15. Bridges LC, Bowditch RD. ADAM-Integrin Interactions:potential integrin regulated ectodomain shedding activity.Curr Pharm Des 2005;11:837–47.
16. Howard L, Maciewicz RA, Blobel CP. Cloning andcharacterization of ADAM28: evidence for autocatalytic pro-domain removal and for cell surface localization of matureADAM28. Biochem J 2000;348:21–7.
17. Ohtsuka T, Shiomi T, Shimoda M, Kodama T, Amour A,Murphy G, et al. ADAM28 is overexpressed in human non-small cell lung carcinomas and correlates with cellproliferation and lymph node metastasis. Int J Cancer2006;118:263–73.
18. Mitsui Y, Mochizuki S, Kodama T, Shimoda M, Ohtsuka T,Shiomi T, et al. ADAM28 is overexpressed in human breastcarcinomas: implications for carcinoma cell proliferationthrough cleavage of insulin-like growth factor bindingprotein-3. Cancer Res 2006;66:9913–20.
19. Mochizuki S, Shimoda M, Shiomi T, Fujii Y, Okada Y.ADAM28 is activated by MMP-7 (matrilysin-1) and cleavesinsulin-like growth factor binding protein-3. Biochem BiophysRes Commun 2004;315:79–84.
20. Morsczeck C, Moehl C, Gotz W, Heredia A, Schaffer TE,Eckstein N, et al. In vitro differentiation of human dentalfollicle cells with dexamethasone and insulin. Cell Biol Int2005;29:567–75.
21. Jernvall J, Thesleff I. Reiterative signaling and patterningduring mammalian tooth morphogenesis. Mech Dev2000;92:19–29.
22. Otsubo A, Bhawal UK, Nomura Y, Mitani Y, Ozawa K,Kuniyasu H, et al. UCN-01(7-hydroxystaurosporine) inducesapoptosis and G1 arrest of both primary and metastatic oralcancer cell lines in vitro. Oral Surg Oral Med Oral Pathol OralRadiol Endo 2007;103:391–7.
23. Slavkin HC. Antisense oligonucleotides: an experimentalstrategy to advance a causal analysis of development. Int JDev Biol 1995;39:123–6.
24. Schmitt R, Fausser JL, Lesot H, Ruch JV. Effects of hepatocytegrowth factor anti-sense oligodeoxynucleotides or met D/Dgenotype on mouse molar crown morphogenesis. Int J DevBiol 2000;44:403–8.
25. Tabata MJ, Fujii T, Liu JG, Ohmori T, Abe M, Wakisaka S,et al. Bone morphogenetic protein 4 is involved in cuspformation in molar tooth germ of mice. Eur J Oral Sci2002;110:114–20.
26. Chai Y, Zhao J, Mogharei A, Xu B, Bringas PJr. Shuler C, et al.Inhibition of transforming growth factor-beta type IIreceptor signaling accelerates tooth formation in mousefirst branchial arch explants. Mech Dev 1999;86:63–74.
27. Santamaria D, Ortega S. Cyclins and CDKs in developmentand cancer: lessons from genetically modified mice. FrontBiosci 2006;11:1164–88.
28. Howard L, Zheng Y, Horrocks M, Maciewicz RA, Blobel C.Catalytic activity of ADAM28. FEBS Lett 2001;498:82–6.
29. Partanen AM, Thesleff I. Localization and quantitation of125I-epidermal growth factor binding in murine embryonictooth and other embryonic tissues at differentdevelopmental stages. Dev Biol 1987;120:186–97.
30. Groeneveld MC, Everts V, Beertsen W. Alkaline phosphataseactivity in the periodontal ligament and gingiva of the ratmolar: its relation to cementum formation. J Dent Res1995;74:1374–81.
31. Van Hoof VO, De Broe ME. Interpretation and clinicalsignificance of alkaline phosphatase isoenzyme patterns.Crit Rev Clin Lab Sci 1994;31:197–293.
32. Milan AM, Waddington RJ, Embery G. Fluoride alters caseinkinase II and alkaline phosphatase activity in vitro withpotential implications for dentine mineralization. Arch OralBiol 2001;46:343–51.
33. Wistuba J, Ehmcke J, Clemen G. Tooth development inAmbystoma mexicanum: phosphatase activities, calciumaccumulation and cell proliferation in the tooth-formingtissues. Ann Anat 2003;185:239–45.
34. Shiba H, Mouri Y, Komatsuzawa H, Mizuno N, Xu W,Noguchi T, et al. Enhancement of alkaline phosphatasesynthesis in pulp cells co-cultured with epithelial cellsderived from lower rabbit incisors. Cell Biol Int 2003;27:815–23.
35. Zhu L, Skoultchi AI. Coordinating cell proliferation anddifferentiation. Curr Opin Genet Dev 2001;11:91–7.
36. Satchell PG, Gutmann JL, Witherspoon DE. Apoptosis: anintroduction for the endodontist. Int Endod J 2003;36:237–45.

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 8 3 5 – 8 4 5 845
37. Peterkova R, Peterka M, Lesot H. The developing murinedentition: a new tool for apoptosis study. Ann N Y Acad Sci2003;1010:453–66.
38. Vaahtokari A, Aberg T, Thesleff I. Apoptosis in thedeveloping tooth: association with an embryonic signalingcenter and suppression by EGF and FGF-4. Development1996;122:121–9.
39. Terranova VP, Wikesjo UM. Extracellular matrices andpolypeptide growth factors a phogenesis. Curr OpinBiotechnol 2003;14:526–32.
40. Tsuchiya S, Honda MJ, Shinohara Y, Saito M, Ueda M.Collagen type I matrix affects molecular and cellularbehavior of purified porcine dental follicle cells. Cell TissueRes 2008;331:447–59.
41. Berthiaume F, Moghe PV, Toner M, Yarmush ML. Effect ofextracellular matrix topology on cell structure, function,and physiological responsiveness: hepatocytes cultured in asandwich configuration. FASEB J 1996;10:1471–84.
42. Borene ML, Barocas VH, Hubel A. Mechanical and cellularchanges during compaction of a collagen-sponge-basedcorneal stromal equivalent. Ann Biomed Eng 2004;32:274–83.
43. Kleinman HK, Philp D, Hoffman MP. Role of the extracellularmatrix in morphogenesis. Curr Opin Biotechnol 2003;14:526–32.
44. Carlinfante G, Vassiliou D, Svensson O, Wendel M,Heinegard D, Andersson G. Differential expression ofosteopontin and bone sialoprotein in bone metastasis ofbreast and prostate carcinoma. Clin Exp Metastasis2003;20:437–44.
45. Chen JK, Shapiro HS, Wrana JL, Reimers S, Heersche JN,Sodek J. Localization of bone sialoprotein (BSP)
expression to sites of mineralized tissue formation infetal rat tissues by in situ hybridization. Matrix 1991;11:133–43.
46. Chen JK, Shapiro HS, Sodek J. Developmental expression ofbone sialoprotein mRNA in rat mineralized connectivetissues. J Bone Mineral Res 1992;8:987–97.
47. Bianco P, Fisher LW, Young MF, Termine JD, Robey PG.Expression of bone sialoprotein (BSP) in developing humantissues. Calcif Tissue Int 1991;49:421–6.
48. Ganss B, Kim RH, Sodek J. Bone sialoprotein. Crit Rev Oral BiolMed 1999;10:79–98.
49. Chen JK, Shapiro HS, Sodek J. Developmental expression ofbone sialoprotein mRNA in rat mineralized connectivetissues. J Bone Miner Res 1992;7:987–97.
50. Giachelli CM, Steitz S. Osteopont: a versatile regulator ofinflammation and biomineralization. Matrix Biol2000;19:615–22.
51. Sodek J, Ganss B, McKee MD. Osteopontin. Crit Rev Oral BiolMed 2000;11:279–303.
52. Sodek J, Chen J, Nagata T, Kasugai S, Todescan Jr R, Li IW,et al. Regulation of osteopontin expression in osteoblasts.Ann N Y Acad Sci 1995;760:223–41.
53. Sodek J, Chen J, Kasugai S, Nagata T, Zhang Q, McKee MD,et al. Elucidating the functions of bone sialoprotein andosteopontin in bone formation. In: Slavkin H, Price P,editors. Chemistry and biology of mineralized tissues. ElsevierScience Publishers B.V; 1992. p. 297–306.
54. Hou LT, Liu CM, Chen YJ, Wong MY, Chen KC, Chen J, et al.Characterization of dental follicle cells in developing mousemolar. Arch Oral Biol 1999;44:759–70.


















