
inflamm10
8
Biol. Chem., Vol. 387, pp. 365–372, April 2006 • Copyright by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2006.049 2006/391 Article in press - uncorrected proof Minireview Oxidative and nitrative DNA damage in animals and patients with inflammatory diseases in relation to inflammation- related carcinogenesis Shosuke Kawanishi 1, *, Yusuke Hiraku 1 , Somchai Pinlaor 1,2,3 and Ning Ma 4 1 Department of Environmental and Molecular Medicine, Mie University Graduate School of Medicine, Mie 514- 8507, Japan 2 Department of Parasitology, Faculty of Medicine, Khon Kaen University, Khon Kaen, 40002, Thailand 3 Liver Fluke and Cholangiocarcinoma Research Center, Faculty of Medicine, Khon Kaen University, Khon Kaen, 40002, Thailand 4 Department of Anatomy, Mie University Graduate School of Medicine, Mie 514-8507, Japan * Corresponding author e-mail: [email protected] Abstract Infection and chronic inflammation are proposed to con- tribute to carcinogenesis through inflammation-related mechanisms. Infection with hepatitis C virus, Helicobac- ter pylori and the liver fluke, Opisthorchis viverrini (OV), are important risk factors for hepatocellular carcinoma (HCC), gastric cancer and cholangiocarcinoma, respec- tively. Inflammatory bowel diseases (IBDs) and oral dis- eases, such as oral lichen planus (OLP) and leukoplakia, are associated with colon carcinogenesis and oral squa- mous cell carcinoma (OSCC), respectively. We performed a double immunofluorescence labeling study and found that nitrative and oxidative DNA lesion products, 8-nitro- guanine and 8-oxo-7,8-dihydro-29-deoxyguanosine (8- oxodG), were formed and inducible nitric oxide synthase (iNOS) was expressed in epithelial cells and inflammatory cells at the site of carcinogenesis in humans and animal models. Antibacterial, antiviral and antiparasitic drugs dramatically diminished the formation of these DNA lesion markers and iNOS expression. These results sug- gest that oxidative and nitrative DNA damage occurs at the sites of carcinogenesis, regardless of etiology. There- fore, it is considered that excessive amounts of reactive nitrogen species produced via iNOS during chronic inflammation may play a key role in carcinogenesis by causing DNA damage. On the basis of our results, we propose that 8-nitroguanine is a promising biomarker to evaluate the potential risk of inflammation-mediated carcinogenesis. Keywords: carcinogenesis; DNA damage; inducible nitric oxide synthase; inflammation; 8-nitroguanine; 8-oxo-7,8-dihydro-29-deoxyguanosine. Introduction Recently, experimental and epidemiological evidence indicates that a variety of infectious agents constitute one of the main causes of cancer (Coussens and Werb, 2002; IARC, 2003). The International Agency for Research on Cancer (IARC) has estimated that approxi- mately 18% of cancer cases worldwide are attributable to infectious diseases (IARC, 2003). The burden of can- cer caused by infectious agents is shown in Table 1. Viruses, bacteria and parasites can cause chronic inflam- mation and contribute to 1.6 million cases of infection- related malignancies per year. Inflammation can be induced not only by chronic infection, but also by many other physical, chemical and immunological factors (Coussens and Werb, 2002; Ohshima et al., 2003). It has been hypothesized that many malignancies arise from areas of infection and inflammation (Balkwill and Man- tovani, 2001; Coussens and Werb, 2002). Reactive oxygen species (ROS) and reactive nitrogen species (RNS) are capable of causing damage to various cellular constituents, such as nucleic acids, proteins and lipids. ROS and RNS are considered to play an important role in carcinogenesis through oxidative and nitrative DNA damage (Hussain et al., 2003; Ohshima et al., 2003). ROS can induce the formation of oxidative DNA lesion products, including 8-oxo-7,8-dihydro-29-deoxyguano- sine (8-oxodG) (Wiseman and Halliwell, 1996; Burrows and Muller, 1998; Kawanishi et al., 2001; Evans et al., 2004), which is considered to be mutagenic. It has been reported that misincorporation of adenine occurs oppo- site 8-oxodG during DNA synthesis, leading to G™T transversions (Shibutani et al., 1991; Bruner et al., 2000). ROS are generated from multiple sources, including inflammatory cells, as well as carcinogenic chemicals and their metabolites and the electron transport chain in mitochondria. On the other hand, nitric oxide (NO) is generated spe- cifically during inflammation via inducible nitric oxide syn- thase (iNOS) in inflammatory and epithelial cells (Figure 1). Excess NO production plays a crucial role in an enor- mous variety of pathological processes, including cancer (Hussain et al., 2003; Ohshima et al., 2003). NO reacts with superoxide (O 2 •- ) to form peroxynitrite (ONOO - ), a highly reactive species causing nitrative and oxidative DNA damage. ONOO - can mediate the formation of 8- oxodG (Inoue and Kawanishi, 1995) and 8-nitroguanine, a marker of nitrative DNA damage (Yermilov et al., 1995). Akaike et al. (2003) demonstrated that 8-nitroguanine is formed via NO production associated with inflammation in mice with viral pneumonia. 8-Nitroguanine is consid-
-
Upload
rev-jeremy-d -
Category
Documents
-
view
212 -
download
0
description
Shosuke Kawanishi 1, *, Yusuke Hiraku 1 , Somchai Pinlaor 1,2,3 and Ning Ma 4 Keywords: carcinogenesis; DNA damage; inducible nitric oxide synthase; inflammation; 8-nitroguanine; 8-oxo-7,8-dihydro-29-deoxyguanosine. Article in press - uncorrected proof Abstract Introduction * Corresponding author e-mail: [email protected] Biol. Chem., Vol. 387, pp. 365–372, April 2006 • Copyright by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2006.049 •-
Transcript of inflamm10

Biol. Chem., Vol. 387, pp. 365–372, April 2006 • Copyright � by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2006.049
2006/391
Article in press - uncorrected proof
Minireview
Oxidative and nitrative DNA damage in animals and patientswith inflammatory diseases in relation to inflammation-related carcinogenesis
Shosuke Kawanishi1,*, Yusuke Hiraku1,Somchai Pinlaor1,2,3 and Ning Ma4
1 Department of Environmental and Molecular Medicine,Mie University Graduate School of Medicine, Mie 514-8507, Japan2 Department of Parasitology, Faculty of Medicine, KhonKaen University, Khon Kaen, 40002, Thailand3 Liver Fluke and Cholangiocarcinoma Research Center,Faculty of Medicine, Khon Kaen University, Khon Kaen,40002, Thailand4 Department of Anatomy, Mie University GraduateSchool of Medicine, Mie 514-8507, Japan
* Corresponding authore-mail: [email protected]
Abstract
Infection and chronic inflammation are proposed to con-tribute to carcinogenesis through inflammation-relatedmechanisms. Infection with hepatitis C virus, Helicobac-ter pylori and the liver fluke, Opisthorchis viverrini (OV),are important risk factors for hepatocellular carcinoma(HCC), gastric cancer and cholangiocarcinoma, respec-tively. Inflammatory bowel diseases (IBDs) and oral dis-eases, such as oral lichen planus (OLP) and leukoplakia,are associated with colon carcinogenesis and oral squa-mous cell carcinoma (OSCC), respectively. We performeda double immunofluorescence labeling study and foundthat nitrative and oxidative DNA lesion products, 8-nitro-guanine and 8-oxo-7,8-dihydro-29-deoxyguanosine (8-oxodG), were formed and inducible nitric oxide synthase(iNOS) was expressed in epithelial cells and inflammatorycells at the site of carcinogenesis in humans and animalmodels. Antibacterial, antiviral and antiparasitic drugsdramatically diminished the formation of these DNAlesion markers and iNOS expression. These results sug-gest that oxidative and nitrative DNA damage occurs atthe sites of carcinogenesis, regardless of etiology. There-fore, it is considered that excessive amounts of reactivenitrogen species produced via iNOS during chronicinflammation may play a key role in carcinogenesis bycausing DNA damage. On the basis of our results, wepropose that 8-nitroguanine is a promising biomarker toevaluate the potential risk of inflammation-mediatedcarcinogenesis.
Keywords: carcinogenesis; DNA damage; induciblenitric oxide synthase; inflammation; 8-nitroguanine;8-oxo-7,8-dihydro-29-deoxyguanosine.
Introduction
Recently, experimental and epidemiological evidenceindicates that a variety of infectious agents constituteone of the main causes of cancer (Coussens and Werb,2002; IARC, 2003). The International Agency forResearch on Cancer (IARC) has estimated that approxi-mately 18% of cancer cases worldwide are attributableto infectious diseases (IARC, 2003). The burden of can-cer caused by infectious agents is shown in Table 1.Viruses, bacteria and parasites can cause chronic inflam-mation and contribute to 1.6 million cases of infection-related malignancies per year. Inflammation can beinduced not only by chronic infection, but also by manyother physical, chemical and immunological factors(Coussens and Werb, 2002; Ohshima et al., 2003). It hasbeen hypothesized that many malignancies arise fromareas of infection and inflammation (Balkwill and Man-tovani, 2001; Coussens and Werb, 2002).
Reactive oxygen species (ROS) and reactive nitrogenspecies (RNS) are capable of causing damage to variouscellular constituents, such as nucleic acids, proteins andlipids. ROS and RNS are considered to play an importantrole in carcinogenesis through oxidative and nitrativeDNA damage (Hussain et al., 2003; Ohshima et al., 2003).ROS can induce the formation of oxidative DNA lesionproducts, including 8-oxo-7,8-dihydro-29-deoxyguano-sine (8-oxodG) (Wiseman and Halliwell, 1996; Burrowsand Muller, 1998; Kawanishi et al., 2001; Evans et al.,2004), which is considered to be mutagenic. It has beenreported that misincorporation of adenine occurs oppo-site 8-oxodG during DNA synthesis, leading to G™Ttransversions (Shibutani et al., 1991; Bruner et al., 2000).ROS are generated from multiple sources, includinginflammatory cells, as well as carcinogenic chemicalsand their metabolites and the electron transport chain inmitochondria.
On the other hand, nitric oxide (NO) is generated spe-cifically during inflammation via inducible nitric oxide syn-thase (iNOS) in inflammatory and epithelial cells (Figure1). Excess NO production plays a crucial role in an enor-mous variety of pathological processes, including cancer(Hussain et al., 2003; Ohshima et al., 2003). NO reactswith superoxide (O2
•-) to form peroxynitrite (ONOO-), ahighly reactive species causing nitrative and oxidativeDNA damage. ONOO- can mediate the formation of 8-oxodG (Inoue and Kawanishi, 1995) and 8-nitroguanine,a marker of nitrative DNA damage (Yermilov et al., 1995).Akaike et al. (2003) demonstrated that 8-nitroguanine isformed via NO production associated with inflammationin mice with viral pneumonia. 8-Nitroguanine is consid-

366 S. Kawanishi et al.
Article in press - uncorrected proof
Table 1 The burden of cancer caused by infectious agents worldwide (adapted from the IARC World Cancer Report;IARC, 2003).
Infectious agent IARC Cancer site Number of Percentage ofclassificationa cancer cases cancer cases
worldwide (%)
Bacterial infectionH. pylori 1 Stomach 490 000 5.4
Viral infectionHPV 1, 2A Cervix and other sites 550 000 6.1HBV, HCV 1 Liver 390 000 4.3EBV 1 Lymphoma, 99 000 1.1
nasopharyngeal carcinomaHHV-8 2A Kaposi sarcoma 54 000 0.6HTLV-1 1 Leukemia 9 000 0.1
Parasitic infectionSchistosoma haematobium 1 Bladder 2 700 0.1Liver flukesOpisthorchis viverrini 1 Intra- and extrahepatic 800Clonorchis sinensis 2A bile ducts
Total infection-related cancers 1 600 000 17.7Total cancers in 1995 9 000 000 100aGroup 1, carcinogenic to humans; Group 2A, probably carcinogenic to humans.
Figure 1 Proposed mechanism for mutation via 8-nitroguanine formation induced by inflammation.
ered to be not only a marker of inflammation, but also apotential mutagenic DNA lesion product, leading to car-cinogenesis. 8-Nitroguanine formed in DNA is chemicallyunstable, and thus can be spontaneously released,resulting in the formation of an apurinic site (Yermilovet al., 1995). The apurinic site can form a pair with ade-nine during DNA synthesis, leading to G™T transver-sions (Loeb and Preston, 1986; Figure 1). A recent studyhas revealed that the activity of APE1, the major apurinic-site endonuclease, was significantly increased in colonepithelium displaying elevated inflammation in patients
with ulcerative colitis (Hofseth et al., 2003a). Apurinicsites might represent the major lesion that requires error-prone DNA polymerase z for efficient trans-lesion DNAsynthesis. DNA polymerase z can efficiently bypass aba-sic sites by extending from nucleotides inserted oppositethe lesion by other DNA polymerase (Haracska et al.,2001). Cells deficient in subunits of DNA polymerase z
were hypersensitive to nitrative stress, and trans-lesionDNA synthesis past apurinic sites mediated by this poly-merase might contribute to extensive point mutations(Wu et al., 2006). Recently, it has been reported that in a

DNA damage in inflammation-related carcinogenesis 367
Article in press - uncorrected proof
Figure 2 Double immunofluorescent staining of 8-oxodG and 8-nitroguanine in the liver of OV-infected hamsters.Immunoreactivities of 8-oxodG and 8-nitroguanine were investigated in the same inflammatory cells and epithelium of bile ducts. Theimmunoreactivities of these DNA lesions remained in the epithelium of bile duct on day 90, and increased with the frequency of OVinfection. Inset: inflammatory cells in which 8-oxodG and 8-nitroguanine are colocalized on day 14. Arrow: multinuclear giant cell inwhich only 8-oxodG is formed. 8-NG, 8-nitroguanine. Scale bar represents 50 mm.Methods: To examine the distribution of 8-nitroguanine, a highly sensitive and specific anti-8-nitroguanine antibody without cross-reaction with other related compounds, such as 3-nitrotyrosine, 8-bromoguanine and 8-oxodG, was used (Pinlaor et al., 2004a). 8-Nitroguanine aldehyde-rabbit serum albumin conjugate was intracutaneously injected into a rabbit. The antibody was purified fromthe blood by affinity chromatography. The specificity of the purified antibody was examined by a dot immunobinding assay andabsorption test as previously described (Pinlaor et al., 2004a). Double immunofluorescence labeling studies of 8-nitroguanine and 8-oxodG were performed using tissues obtained from hamsters with OV infection. Deparaffinized paraffin sections were treated withrabbit polyclonal anti-8-nitroguanine antibody and mouse monoclonal anti-8-oxodG antibody. They were then incubated with Alexa594-labeled goat antibody against rabbit IgG and Alexa 488-labeled goat antibody against mouse IgG. The immunostained sectionswere examined by confocal microscopy.
cell-free system, adenine is preferentially incorporatedopposite 8-nitroguanine during DNA synthesis mediatedby DNA polymerase h and k associated with trans-lesionDNA synthesis, suggesting that G™T transversions canalso occur via this mechanism (Suzuki et al., 2005). Inthe ONOO--treated supF shuttle vector plasmid, whichwas then replicated in Escherichia coli, the majority ofmutations occurred at G:C base pairs, predominantlyinvolving G™T transversions (Juedes and Wogan, 1996;Kim et al., 2005). Therefore, 8-nitroguanine is a poten-tially mutagenic DNA lesion product leading to carcino-genesis, as well as 8-oxodG. In particular, we have pro-posed the possibility that 8-nitroguanine is a potentialbiomarker for evaluating the risk of inflammation-relatedcarcinogenesis. Here we discuss the role of oxidative andnitrative DNA damage in carcinogenesis caused bychronic inflammation.
DNA damage in animal models ofinflammation-related carcinogenesis
Liver fluke infection and cholangiocarcinoma
Infection with the liver fluke Opisthorchis viverrini (OV) isa major risk factor for cholangiocarcinoma, especially in
the north-eastern region of Thailand (Haswell-Elkinset al., 1994; IARC Working Group, 1994b). Approximately70% of OV-induced cholangiocarcinomas occur in theintrahepatic bile ducts, and the remainder occur in theextrahepatic duct (Uttararvichen et al., 1996). We inves-tigated DNA damage in the liver of hamsters with singleand repeated OV infection as a model of inflammation-related carcinogenesis in humans. We first demonstratedthat 8-nitroguanine is formed in relation to inflammation-related carcinogenesis using OV-infected hamsters (Pin-laor et al., 2003). Double immunofluorescence stainingrevealed that 8-oxodG and 8-nitroguanine were formedin inflammatory cells and epithelium of bile ducts (Pinlaoret al., 2004a,b). The immunoreactivities of 8-oxodG and8-nitroguanine in inflammatory cells were most promi-nently observed on days 21 and 30, respectively (Pinlaoret al., 2004a). Interestingly, these DNA lesion productsstill remained in the epithelium of bile ducts on day 180(Pinlaor et al., 2004a). The formation of 8-nitroguanineand 8-oxodG increased in the epithelium of bile ducts inthe order of triple infection ) double infection ) singleinfection (Pinlaor et al., 2004b; Figure 2). This may beexplained by the fact that repeated infection increasediNOS expression in the epithelium of bile ducts in thesame order (Pinlaor et al., 2004b). The treatment of
368 S. Kawanishi et al.
Article in press - uncorrected proof
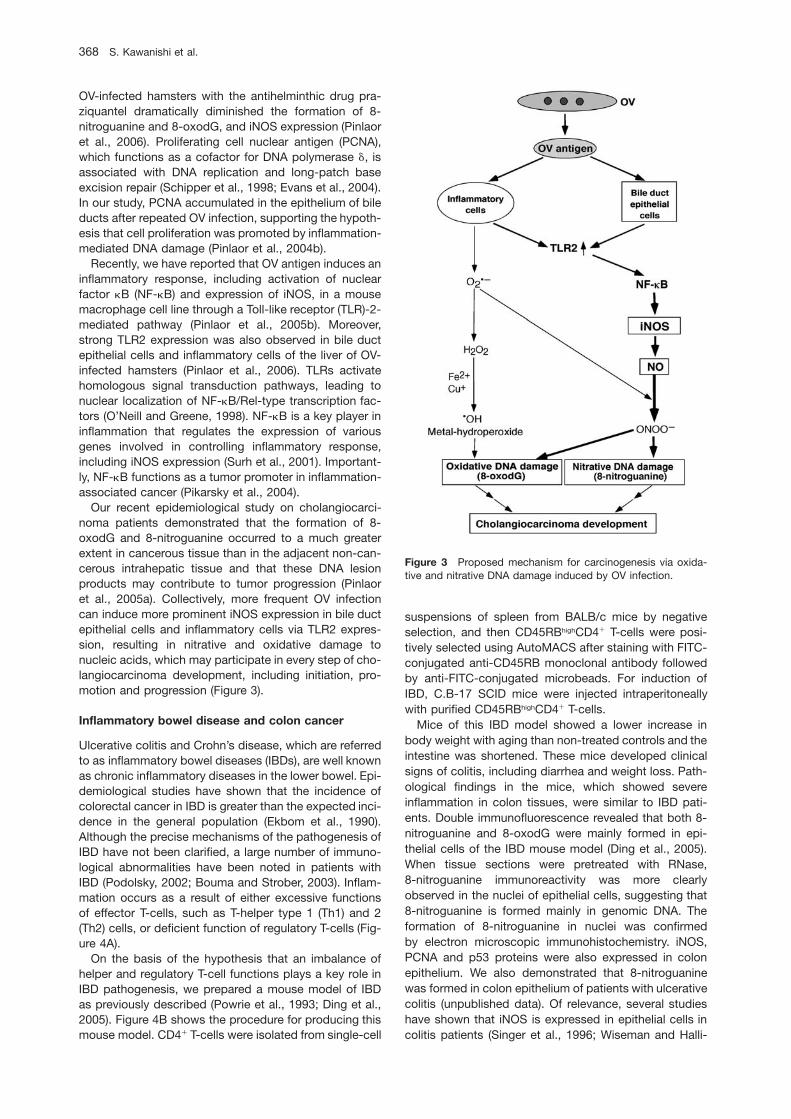
Figure 3 Proposed mechanism for carcinogenesis via oxida-tive and nitrative DNA damage induced by OV infection.
OV-infected hamsters with the antihelminthic drug pra-ziquantel dramatically diminished the formation of 8-nitroguanine and 8-oxodG, and iNOS expression (Pinlaoret al., 2006). Proliferating cell nuclear antigen (PCNA),which functions as a cofactor for DNA polymerase d, isassociated with DNA replication and long-patch baseexcision repair (Schipper et al., 1998; Evans et al., 2004).In our study, PCNA accumulated in the epithelium of bileducts after repeated OV infection, supporting the hypoth-esis that cell proliferation was promoted by inflammation-mediated DNA damage (Pinlaor et al., 2004b).
Recently, we have reported that OV antigen induces aninflammatory response, including activation of nuclearfactor kB (NF-kB) and expression of iNOS, in a mousemacrophage cell line through a Toll-like receptor (TLR)-2-mediated pathway (Pinlaor et al., 2005b). Moreover,strong TLR2 expression was also observed in bile ductepithelial cells and inflammatory cells of the liver of OV-infected hamsters (Pinlaor et al., 2006). TLRs activatehomologous signal transduction pathways, leading tonuclear localization of NF-kB/Rel-type transcription fac-tors (O’Neill and Greene, 1998). NF-kB is a key player ininflammation that regulates the expression of variousgenes involved in controlling inflammatory response,including iNOS expression (Surh et al., 2001). Important-ly, NF-kB functions as a tumor promoter in inflammation-associated cancer (Pikarsky et al., 2004).
Our recent epidemiological study on cholangiocarci-noma patients demonstrated that the formation of 8-oxodG and 8-nitroguanine occurred to a much greaterextent in cancerous tissue than in the adjacent non-can-cerous intrahepatic tissue and that these DNA lesionproducts may contribute to tumor progression (Pinlaoret al., 2005a). Collectively, more frequent OV infectioncan induce more prominent iNOS expression in bile ductepithelial cells and inflammatory cells via TLR2 expres-sion, resulting in nitrative and oxidative damage tonucleic acids, which may participate in every step of cho-langiocarcinoma development, including initiation, pro-motion and progression (Figure 3).
Inflammatory bowel disease and colon cancer
Ulcerative colitis and Crohn’s disease, which are referredto as inflammatory bowel diseases (IBDs), are well knownas chronic inflammatory diseases in the lower bowel. Epi-demiological studies have shown that the incidence ofcolorectal cancer in IBD is greater than the expected inci-dence in the general population (Ekbom et al., 1990).Although the precise mechanisms of the pathogenesis ofIBD have not been clarified, a large number of immuno-logical abnormalities have been noted in patients withIBD (Podolsky, 2002; Bouma and Strober, 2003). Inflam-mation occurs as a result of either excessive functionsof effector T-cells, such as T-helper type 1 (Th1) and 2(Th2) cells, or deficient function of regulatory T-cells (Fig-ure 4A).
On the basis of the hypothesis that an imbalance ofhelper and regulatory T-cell functions plays a key role inIBD pathogenesis, we prepared a mouse model of IBDas previously described (Powrie et al., 1993; Ding et al.,2005). Figure 4B shows the procedure for producing thismouse model. CD4q T-cells were isolated from single-cell
suspensions of spleen from BALB/c mice by negativeselection, and then CD45RBhighCD4q T-cells were posi-tively selected using AutoMACS after staining with FITC-conjugated anti-CD45RB monoclonal antibody followedby anti-FITC-conjugated microbeads. For induction ofIBD, C.B-17 SCID mice were injected intraperitoneallywith purified CD45RBhighCD4q T-cells.
Mice of this IBD model showed a lower increase inbody weight with aging than non-treated controls and theintestine was shortened. These mice developed clinicalsigns of colitis, including diarrhea and weight loss. Path-ological findings in the mice, which showed severeinflammation in colon tissues, were similar to IBD pati-ents. Double immunofluorescence revealed that both 8-nitroguanine and 8-oxodG were mainly formed in epi-thelial cells of the IBD mouse model (Ding et al., 2005).When tissue sections were pretreated with RNase,8-nitroguanine immunoreactivity was more clearlyobserved in the nuclei of epithelial cells, suggesting that8-nitroguanine is formed mainly in genomic DNA. Theformation of 8-nitroguanine in nuclei was confirmedby electron microscopic immunohistochemistry. iNOS,PCNA and p53 proteins were also expressed in colonepithelium. We also demonstrated that 8-nitroguaninewas formed in colon epithelium of patients with ulcerativecolitis (unpublished data). Of relevance, several studieshave shown that iNOS is expressed in epithelial cells incolitis patients (Singer et al., 1996; Wiseman and Halli-

DNA damage in inflammation-related carcinogenesis 369
Article in press - uncorrected proof
Figure 4 Hypothesis for the pathogenesis of IBD and preparation of a mouse model.It has been hypothesized that an imbalance of helper and regulatory T-cell functions plays a key role in IBD pathogenesis (A). Toprepare a mouse model of IBD, CD45RBhighCD4q T-cells (mainly consisting of Th1- and Th2-cells) were isolated from the spleen ofBALB/c mice, and then injected intraperitoneally into C.B-17 SCID mice.
well, 1996; Hofseth et al., 2003b). In non-cancerouscolon tissues from patients with ulcerative colitis, iNOSprotein levels were positively correlated with p53 serine15 phosphorylation levels (Hofseth et al., 2003b). Theseresults suggest that nitrative DNA damage, as well asoxidative DNA damage, participates in colon carcinogen-esis in patients with IBD.
DNA damage in patients with inflammatorydiseases
Helicobacter pylori infection and gastric cancer
Infection with the Gram-negative bacterium Helicobacterpylori is not only associated with chronic atrophicgastritis and peptic ulcer, but also with gastric adeno-carcinoma and non-Hodgkin’s lymphoma wmucosa-asso-ciated lymphoid tissue (MALT) lymphomax (Peek andBlaser, 2002). In 2005, Barry Marshall and Robin Warrenwere awarded with the Nobel Prize for discovery of H.pylori. To examine whether nitrative and oxidative DNAdamage plays a role in the carcinogenic process trig-gered by H. pylori infection, we performed a doubleimmunofluorescence labeling study using biopsy speci-mens of gastric mucosa. Intense immunoreactivities of8-nitroguanine and 8-oxodG were observed in both gas-tric gland epithelial cells and inflammatory cells inpatients with H. pylori infection (Ma et al., 2004). On theother hand, in gastritis patients without H. pylori infec-tion, these immunoreactivities were observed mainly ininflammatory cells, and not in gastric gland epithelialcells. This is the first report showing that 8-nitroguanineis formed in humans in relation to inflammation-related
carcinogenesis. Moreover, we found that 8-nitroguanineformation in patients with H. pylori infection was dra-matically decreased by eradication (unpublished data).The mechanisms by which H. pylori infection causes gas-tric cancer have been investigated. Lipopolysaccharide(LPS), a component of Gram-negative bacteria, includingH. pylori, is a ligand of TLR4. TLR4 is involved in acti-vation of the transcription factor NF-kB (Maeda et al.,2001), which mediates expression of iNOS and variousinflammatory cytokines. Alternatively, the Cag-positive H.pylori strain induces an intense inflammatory response,including interleukin (IL)-8 production by epithelial cellsand subsequent production of tumor necrosis factor(TNF)-a by inflammatory cells (Peek and Blaser, 2002).The host immune response to H. pylori mediated bycytokines, resulting in iNOS expression, may lead to anincrease in the accumulation of 8-nitroguanine and8-oxodG in gastric epithelium.
Several studies have demonstrated that PCNA is anindependent prognostic factor for gastric cancer inpatients with H. pylori infection (Schipper et al., 1998). Inour study, accumulation of PCNA was observed in gas-tric gland epithelial cells in patients with H. pylori infec-tion (Ma et al., 2004). These results suggest that nitrativeand oxidative DNA damage in gastric epithelial cells andtheir proliferation by H. pylori infection may lead to gas-tric carcinoma.
Hepatitis C virus infection and liver cancer
Hepatitis C virus (HCV) is a major cause of chronic hep-atitis, liver cirrhosis and hepatocellular carcinomathroughout the world (IARC Working Group, 1994a; Poy-nard et al., 2003). Hepatocellular carcinoma arises

370 S. Kawanishi et al.
Article in press - uncorrected proof
through a multistep process of genetic alterations inhepatocytes during chronic hepatitis C (CHC) (Bressacet al., 1991; Hsu et al., 1991; Oda et al., 1992; Casel-mann and Alt, 1996). However, the mechanism of HCVinfection-induced hepatitis followed by hepatocarcino-genesis via DNA damage is still unclear.
We investigated DNA damage in liver biopsy speci-mens from patients with CHC and the effect of interferontreatment. Immunoreactivities of 8-nitroguanine and 8-oxodG were strongly detected in liver from patients withCHC, but not in control livers (non-alcoholic fatty liver)(Horiike et al., 2005). 8-Nitroguanine formation was foundnot only in infiltrating inflammatory cells, but also in hepa-tocytes, particularly in the periportal area. The accumu-lation of 8-nitroguanine and 8-oxodG increased withinflammatory grade. iNOS expression was observed inthe cytoplasm of hepatocytes and Kupffer cells in CHCpatients. In the sustained virological responder groupafter interferon therapy, accumulation of 8-nitroguanineand 8-oxodG in the liver was markedly decreased(Horiike et al., 2005). Histological examination showedmore prominent inflammatory changes in CHC patientsthan in control patients, and these changes wereimproved after interferon therapy. Our results are consis-tent with previous reports showing that iNOS wasexpressed in hepatocytes of patients with chronic hep-atitis (McNaughton et al., 2002) and hepatocellular car-cinoma (Rahman et al., 2001). Taken together, thesefindings indicate that 8-nitroguanine is a useful biomarkerfor evaluating the severity of HCV-induced chronicinflammation leading to hepatocellular carcinoma and theefficacy of CHC treatment.
Oral lichen planus, oral cancer and leukoplakia
Oral lichen planus (OLP) is a chronic inflammatory muco-sal disease (Scully et al., 1998). Several pathological fea-tures indicate that OLP is an immunologically mediatedinflammatory response, including an intense, band-likeinfiltrate of predominantly T-lymphocytes subjacentto epithelium. Basal epithelial cells are the target forimmune destruction by cytotoxic T-lymphocytes (Tyldes-ley and Appleton, 1973; Dekker et al., 1997). The mostimportant complication of OLP is development of oralsquamous cell carcinoma (OSCC) (Rajentheran et al.,1999; Mignogna et al., 2004). Oral leukoplakia is a pre-cancerous lesion characterized by white plaque andhyperkeratosis (Neville and Day, 2002; Reibel, 2003), with5–15% of leukoplakia histologically classified as dyspla-sia (Suarez et al., 1998; Sudbo and Reith, 2005). A sub-stantial proportion of dysplasia is reported to developinto oral carcinoma (Lumerman et al., 1995; Sudbo andReith, 2005). However, DNA damage associated withthese oral diseases has not been investigated.
We demonstrated that accumulation of 8-nitroguanineand 8-oxodG was observed in oral epithelium of biopsyspecimens from patients with OLP and OSCC, whereasno immunoreactivity was observed in normal oral muco-sa (Chaiyarit et al., 2005). 8-Nitroguanine and 8-oxodGwere also observed in oral epithelium of patients withleukoplakia (Ma et al., 2006). Colocalization of 8-nitro-guanine and iNOS was found in oral epithelium ofpatients with OLP, OSCC and leukoplakia. Immunoreac-
tivity of 3-nitrotyrosine, which is formed by protein tyro-sine nitration and considered to be a biochemical markerfor inflammation, was also observed in oral epithelialcells. Accumulation of p53 was observed in oral epithe-lium in OLP and leukoplakia, and more prominentexpression of this protein was observed in OSCCpatients. Our findings demonstrate that iNOS-dependentDNA damage may lead to p53 accumulation not only inOLP and leukoplakia, but also in OSCC. It is concludedthat the formation of 8-nitroguanine and 8-oxodG maycontribute to the development of oral cancer from OLPand leukoplakia.
Conclusion
In relation to inflammation-related carcinogenesis, weexamined the formation of 8-nitroguanine and 8-oxodGin experimental animal models. DNA damage was spe-cifically induced at sites of carcinogenesis under variousinflammatory conditions. In hamsters infected with theliver fluke OV causing cholangiocarcinoma, 8-nitrogua-nine formation was induced in bile duct epithelium (Pin-laor et al., 2004b). Moreover, 8-nitroguanine was formedin colonic gland epithelial cells in a mouse model of IBD(Ding et al., 2005). It is noteworthy that 8-nitroguanineformation was also observed in human samples. 8-Nitro-guanine was formed in gastric gland epithelial cells ofpatients with H. pylori infection (Ma et al., 2004) and inhepatocytes of patients with chronic hepatitis C (Horiikeet al., 2005). 8-Nitroguanine was also formed in oral epi-thelium of patients with OLP, OSCC (Chaiyarit et al.,2005) and leukoplakia (Ma et al., 2006). Therefore, 8-nitroguanine could be used as a potential biomarker ofinflammation-related carcinogenesis. Establishment ofmethods for quantitative analysis of 8-nitroguanine inbiological or clinical specimens could be useful for eval-uating the risk of carcinogenesis. However, 8-nitrogua-nine formed in DNA is chemically unstable, and thischaracteristic may hamper its quantitative analysis.Therefore, free 8-nitroguanine released from DNA in urinemight be available for quantification as a biomarker. Inaddition, measurement of 8-nitroguanosine derived from8-nitroguanine-bound RNA in clinical specimens, includ-ing white blood cells, may be useful for evaluation of car-cinogenic potential.
Recently, 8-nitroguanosine has been reported to be ahighly redox-active molecule (Sawa et al., 2003; Zaki etal., 2005). More importantly, experimental evidence hassuggested that 8-nitroguanine is a mutagenic DNAlesion, which preferentially leads to G™T transversions(Yermilov et al., 1995; Suzuki et al., 2005), in addition to8-oxodG (Shibutani et al., 1991; Bruner et al., 2000).Indeed, G™T transversions have been observed in vivoin the ras gene (Bos, 1988) and the p53 tumor suppressorgene in lung and liver cancer (Takahashi et al., 1989; Hsuet al., 1991). These findings imply that DNA damagemediated by ROS and RNS may participate in carcino-genesis via activation of proto-oncogenes and inactiva-tion of tumor suppressor genes. In conclusion, oxidativeand nitrative DNA damage could represent promising

DNA damage in inflammation-related carcinogenesis 371
Article in press - uncorrected proof
biomarkers for evaluating the risk of inflammation-relatedcarcinogenesis.
References
Akaike, T., Okamoto, S., Sawa, T., Yoshitake, J., Tamura, F., Ichi-mori, K., Miyazaki, K., Sasamoto, K., and Maeda, H. (2003).8-Nitroguanosine formation in viral pneumonia and its impli-cation for pathogenesis. Proc. Natl. Acad. Sci. USA 100,685–690.
Balkwill, F. and Mantovani, A. (2001). Inflammation and cancer:back to Virchow? Lancet 357, 539–545.
Bos, J.L. (1988). The ras gene family and human carcinogenesis.Mutat. Res. 195, 255–271.
Bouma, G. and Strober, W. (2003). The immunological andgenetic basis of inflammatory bowel disease. Nat. Rev.Immunol. 3, 521–533.
Bressac, B., Kew, M., Wands, J., and Ozturk, M. (1991). Selec-tive G to T mutations of p53 gene in hepatocellular carci-noma from southern Africa. Nature 350, 429–431.
Bruner, S.D., Norman, D.P., and Verdine, G.L. (2000). Structuralbasis for recognition and repair of the endogenous mutagen8-oxoguanine in DNA. Nature 403, 859–866.
Burrows, C.J. and Muller, J.G. (1998). Oxidative nucleobasemodifications leading to strand scission. Chem. Rev. 98,1109–1151.
Caselmann, W.H. and Alt, M. (1996). Hepatitis C virus infectionas a major risk factor for hepatocellular carcinoma. J. Hepa-tol. 24, 61–66.
Chaiyarit, P., Ma, N., Hiraku, Y., Pinlaor, S., Yongvanit, P., Jinta-kanon, D., Murata, M., Oikawa, S., and Kawanishi, S. (2005).Nitrative and oxidative DNA damage in oral lichen planus inrelation to human oral carcinogenesis. Cancer Sci. 96,553–559.
Coussens, L.M. and Werb, Z. (2002). Inflammation and cancer.Nature 420, 860–867.
Dekker, N.P., Lozada-Nur, F., Lagenaur, L.A., MacPhail, L.A.,Bloom, C.Y., and Regezi, J.A. (1997). Apoptosis-associatedmarkers in oral lichen planus. J. Oral Pathol. Med. 26,170–175.
Ding, X., Hiraku, Y., Ma, N., Kato, T., Saito, K., Nagahama, M.,Semba, R., Kuribayashi, K., and Kawanishi, S. (2005). Induc-ible nitric oxide synthase-dependent DNA damage in mousemodel of inflammatory bowel disease. Cancer Sci. 96,157–163.
Ekbom, A., Helmick, C., Zack, M., and Adami, H.O. (1990).Increased risk of large-bowel cancer in Crohn’s disease withcolonic involvement. Lancet 336, 357–359.
Evans, M.D., Dizdaroglu, M., and Cooke, M.S. (2004). OxidativeDNA damage and disease: induction, repair and significance.Mutat. Res. 567, 1–61.
Haracska, L., Unk, I., Johnson, R.E., Johansson, E., Burgers,P.M., Prakash, S., and Prakash, L. (2001). Roles of yeast DNApolymerases d and z and of Rev1 in the bypass of abasicsites. Genes Dev. 15, 945–954.
Haswell-Elkins, M.R., Mairiang, E., Mairiang, P., Chaiyakum, J.,Chamadol, N., Loapaiboon, V., Sithithaworn, P., and Elkins,D.B. (1994). Cross-sectional study of Opisthorchis viverriniinfection and cholangiocarcinoma in communities within ahigh-risk area in northeast Thailand. Int. J. Cancer 59,505–509.
Hofseth, L.J., Khan, M.A., Ambrose, M., Nikolayeva, O., Xu-Wel-liver, M., Kartalou, M., Hussain, S.P., Roth, R.B., Zhou, X.,Mechanic, L.E., et al. (2003a). The adaptive imbalance inbase excision-repair enzymes generates microsatellite insta-bility in chronic inflammation. J. Clin. Invest. 112, 1887–1894.
Hofseth, L.J., Saito, S., Hussain, S.P., Espey, M.G., Miranda,K.M., Araki, Y., Jhappan, C., Higashimoto, Y., He, P., Linke,S.P., et al. (2003b). Nitric oxide-induced cellular stress and
p53 activation in chronic inflammation. Proc. Natl. Acad. Sci.USA 100, 143–148.
Horiike, S., Kawanishi, S., Kaito, M., Ma, N., Tanaka, H., Fujita,N., Iwasa, M., Kobayashi, Y., Hiraku, Y., Oikawa, S., et al.(2005). Accumulation of 8-nitroguanine in the liver of patientswith chronic hepatitis C. J. Hepatol. 43, 403–410.
Hsu, I.C., Metcalf, R.A., Sun, T., Welsh, J.A., Wang, N.J., andHarris, C.C. (1991). Mutational hotspot in the p53 gene inhuman hepatocellular carcinomas. Nature 350, 427–428.
Hussain, S.P., Hofseth, L.J., and Harris, C.C. (2003). Radicalcauses of cancer. Nat. Rev. Cancer 3, 276–285.
IARC (2003). Chronic infections. In: World Cancer Report, B.W.Stewart and P. Kleihues, eds. (Lyon, France: IARC Press), pp.56–61.
IARC Working Group (1994a). Hepatitis C virus. IARC Monogr.Eval. Carcinog. Risks Hum. 59, 165–221.
IARC Working Group (1994b). Infection with liver flukes (Opis-thorchis viverrini, Opisthorchis felineus and Clonorchis sinen-sis). IARC Monogr. Eval. Carcinog. Risks Hum. 61, 121–175.
Inoue, S. and Kawanishi, S. (1995). Oxidative DNA damageinduced by simultaneous generation of nitric oxide andsuperoxide. FEBS Lett. 371, 86–88.
Juedes, M.J. and Wogan, G.N. (1996). Peroxynitrite-inducedmutation spectra of pSP189 following replication in bacteriaand in human cells. Mutat. Res. 349, 51–61.
Kawanishi, S., Hiraku, Y., and Oikawa, S. (2001). Mechanism ofguanine-specific DNA damage by oxidative stress and itsrole in carcinogenesis and aging. Mutat. Res. 488, 65–76.
Kim, M.Y., Dong, M., Dedon, P.C., and Wogan, G.N. (2005).Effects of peroxynitrite dose and dose rate on DNA damageand mutation in the supF shuttle vector. Chem. Res. Toxicol.18, 76–86.
Loeb, L.A. and Preston, B.D. (1986). Mutagenesis by apurinic/apyrimidinic sites. Annu. Rev. Genet. 20, 201–230.
Lumerman, H., Freedman, P., and Kerpel, S. (1995). Oral epithe-lial dysplasia and the development of invasive squamous cellcarcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol.Endod. 79, 321–329.
Ma, N., Adachi, Y., Hiraku, Y., Horiki, N., Horiike, S., Imoto, I.,Pinlaor, S., Murata, M., Semba, R., and Kawanishi, S. (2004).Accumulation of 8-nitroguanine in human gastric epitheliuminduced by Helicobacter pylori infection. Biochem. Biophys.Res. Commun. 319, 506–510.
Ma, N., Tagawa, T., Hiraku, Y., Murata, M., Ding, X., and Kawa-nishi, S. (2006). 8-Nitroguanine formation in oral leukoplakia,a premalignant lesion. Nitric Oxide 14, 137–143.
Maeda, S., Akanuma, M., Mitsuno, Y., Hirata, Y., Ogura, K., Yos-hida, H., Shiratori, Y., and Omata, M. (2001). Distinct mech-anism of Helicobacter pylori-mediated NF-kB activationbetween gastric cancer cells and monocytic cells. J. Biol.Chem. 276, 44856–44864.
McNaughton, L., Puttagunta, L., Martinez-Cuesta, M.A., Knete-man, N., Mayers, I., Moqbel, R., Hamid, Q., and Radomski,M.W. (2002). Distribution of nitric oxide synthase in normaland cirrhotic human liver. Proc. Natl. Acad. Sci. USA 99,17161–17166.
Mignogna, M.D., Fedele, S., Lo Russo, L., Lo Muzio, L., andBucci, E. (2004). Immune activation and chronic inflammationas the cause of malignancy in oral lichen planus: is there anyevidence? Oral Oncol. 40, 120–130.
Neville, B.W., and Day, T.A. (2002). Oral cancer and precancer-ous lesions. CA Cancer J. Clin. 52, 195–215.
Oda, T., Tsuda, H., Scarpa, A., Sakamoto, M., and Hirohashi, S.(1992). p53 gene mutation spectrum in hepatocellular carci-noma. Cancer Res. 52, 6358–6364.
Ohshima, H., Tatemichi, M., and Sawa, T. (2003). Chemical basisof inflammation-induced carcinogenesis. Arch. Biochem.Biophys. 417, 3–11.
O’Neill, L.A., and Greene, C. (1998). Signal transduction path-ways activated by the IL-1 receptor family: ancient signalingmachinery in mammals, insects, and plants. J. Leukoc. Biol.63, 650–657.

372 S. Kawanishi et al.
Article in press - uncorrected proof
Peek Jr., R.M., and Blaser, M.J. (2002). Helicobacter pylori andgastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2,28–37.
Pikarsky, E., Porat, R.M., Stein, I., Abramovitch, R., Amit, S.,Kasem, S., Gutkovich-Pyest, E., Urieli-Shoval, S., Galun, E.,and Ben-Neriah, Y. (2004). NF-kB functions as a tumour pro-moter in inflammation-associated cancer. Nature 431,461–466.
Pinlaor, S., Yongvanit, P., Hiraku, Y., Ma, N., Semba, R., Oikawa,S., Murata, M., Sripa, B., Sithithaworn, P., and Kawanishi, S.(2003). 8-Nitroguanine formation in the liver of hamstersinfected with Opisthorchis viverrini. Biochem. Biophys. Res.Commun. 309, 567–571.
Pinlaor, S., Hiraku, Y., Ma, N., Yongvanit, P., Semba, R., Oikawa,S., Murata, M., Sripa, B., Sithithaworn, P., and Kawanishi, S.(2004a). Mechanism of NO-mediated oxidative and nitrativeDNA damage in hamsters infected with Opisthorchis viverrini:a model of inflammation-mediated carcinogenesis. NitricOxide 11, 175–183.
Pinlaor, S., Ma, N., Hiraku, Y., Yongvanit, P., Semba, R., Oikawa,S., Murata, M., Sripa, B., Sithithaworn, P., and Kawanishi, S.(2004b). Repeated infection with Opisthorchis viverrini induc-es accumulation of 8-nitroguanine and 8-oxo-7,8-dihydro-29-deoxyguanine in the bile duct of hamsters via inducible nitricoxide synthase. Carcinogenesis 25, 1535–1542.
Pinlaor, S., Sripa, B., Ma, N., Hiraku, Y., Yongvanit, P., Wongk-ham, S., Pairojkul, C., Bhudhisawasdi, V., Oikawa, S., Murata,M., et al. (2005a). Nitrative and oxidative DNA damage inintrahepatic cholangiocarcinoma patients in relation to tumorinvasion. World J. Gastroenterol. 11, 4644–4649.
Pinlaor, S., Tada-Oikawa, S., Hiraku, Y., Pinlaor, P., Ma, N., Sithi-thaworn, P., and Kawanishi, S. (2005b). Opisthorchis viverriniantigen induces the expression of Toll-like receptor 2 in mac-rophage RAW cell line. Int. J. Parasitol. 35, 591–596.
Pinlaor, S., Hiraku, Y., Yongvanit, P., Tada-Oikawa, S., Ma, N.,Pinlaor, P., Sithithaworn, P., Sripa, B., Marata, M., Oikawa, S.,and Kawanishi, S. (2006). iNOS-dependent DNA damage viaNF-kB expression in hamsters infected with Opisthorchisviverrini and its suppression by the antihelminthic drug pra-ziquantel. Int. J. Cancer, in press.
Podolsky, D.K. (2002). Inflammatory bowel disease. N. Engl. J.Med. 347, 417–429.
Powrie, F., Leach, M.W., Mauze, S., Caddle, L.B., and Coffman,R.L. (1993). Phenotypically distinct subsets of CD4q T cellsinduce or protect from chronic intestinal inflammation in C.B-17 scid mice. Int. Immunol. 5, 1461–1471.
Poynard, T., Yuen, M.F., Ratziu, V., and Lai, C.L. (2003). Viralhepatitis C. Lancet 362, 2095–2100.
Rahman, M.A., Dhar, D.K., Yamaguchi, E., Maruyama, S., Sato,T., Hayashi, H., Ono, T., Yamanoi, A., Kohno, H., and Naga-sue, N. (2001). Coexpression of inducible nitric oxidesynthase and COX-2 in hepatocellular carcinoma and sur-rounding liver: possible involvement of COX-2 in the angio-genesis of hepatitis C virus-positive cases. Clin. Cancer Res.7, 1325–1332.
Rajentheran, R., McLean, N.R., Kelly, C.G., Reed, M.F., andNolan, A. (1999). Malignant transformation of oral lichen pla-nus. Eur. J. Surg. Oncol. 25, 520–523.
Reibel, J. (2003). Prognosis of oral pre-malignant lesions: sig-nificance of clinical, histopathological, and molecular biolog-ical characteristics. Crit. Rev. Oral Biol. Med. 14, 47–62.
Sawa, T., Akaike, T., Ichimori, K., Akuta, T., Kaneko, K., Nakaya-ma, H., Stuehr, D.J., and Maeda, H. (2003). Superoxide gen-eration mediated by 8-nitroguanosine, a highly redox-activenucleic acid derivative. Biochem. Biophys. Res. Commun.311, 300–306.
Schipper, D.L., Wagenmans, M.J., Peters, W.H., and Wagener,D.J. (1998). Significance of cell proliferation measurement ingastric cancer. Eur. J. Cancer 34, 781–790.
Scully, C., Beyli, M., Ferreiro, M.C., Ficarra, G., Gill, Y., Griffiths,M., Holmstrup, P., Mutlu, S., Porter, S., and Wray, D. (1998).Update on oral lichen planus: etiopathogenesis and manage-ment. Crit. Rev. Oral Biol. Med. 9, 86–122.
Shibutani, S., Takeshita, M., and Grollman, A.P. (1991). Insertionof specific bases during DNA synthesis past the oxidation-damaged base 8-oxodG. Nature 349, 431–434.
Singer, I.I., Kawka, D.W., Scott, S., Weidner, J.R., Mumford, R.A.,Riehl, T.E., and Stenson, W.F. (1996). Expression of induciblenitric oxide synthase and nitrotyrosine in colonic epitheliumin inflammatory bowel disease. Gastroenterology 111, 871–885.
Suarez, P., Batsakis, J.G., and el-Naggar, A.K. (1998). Leukopla-kia: still a gallimaufry or is progress being made? – A review.Adv. Anat. Pathol. 5, 137–155.
Sudbo, J. and Reith, A. (2005). The evolution of predictive oncol-ogy and molecular-based therapy for oral cancer prevention.Int. J. Cancer 115, 339–345.
Surh, Y.J., Chun, K.S., Cha, H.H., Han, S.S., Keum, Y.S., Park,K.K., and Lee, S.S. (2001). Molecular mechanisms underlyingchemopreventive activities of anti-inflammatory phytoche-micals: down-regulation of COX-2 and iNOS through sup-pression of NF-kB activation. Mutat. Res. 480/481, 243–268.
Suzuki, N., Yasui, M., Geacintov, N.E., Shafirovich, V., and Shi-butani, S. (2005). Miscoding events during DNA synthesispast the nitration-damaged base 8-nitroguanine. Biochem-istry 44, 9238–9245.
Takahashi, T., Nau, M.M., Chiba, I., Birrer, M.J., Rosenberg, R.K.,Vinocour, M., Levitt, M., Pass, H., Gazdar, A.F., and Minna,J.D. (1989). p53: a frequent target for genetic abnormalitiesin lung cancer. Science 246, 491–494.
Tyldesley, W.R., and Appleton, J. (1973). Observations on theultrastructure of the epithelium in oral lichen planus. J. Oral.Pathol. 2, 46–57.
Uttararvichen, T., Buddhiswasdi, V., and Pairojkul, C. (1996). Bileduct cancer and the liver fluke: pathology, presentation andsurgical management. Asian J. Surg. 19, 267–270.
Wiseman, H., and Halliwell, B. (1996). Damage to DNA by reac-tive oxygen and nitrogen species: role in inflammatory dis-ease and progression to cancer. Biochem. J. 313, 17–29.
Wu, X., Takenaka, K., Sonoda, E., Hochegger, H., Kawanishi, S.,Kawamoto, T., Takeda, S., and Yamazoe, M. (2006). Criticalroles for polymerase z in cellular tolerance to nitric oxide-induced DNA damage. Cancer Res. 66, 748–754.
Yermilov, V., Rubio, J., Becchi, M., Friesen, M.D., Pignatelli, B.,and Ohshima, H. (1995). Formation of 8-nitroguanine by thereaction of guanine with peroxynitrite in vitro. Carcinogenesis16, 2045–2050.
Zaki, M.H., Akuta, T., and Akaike, T. (2005). Nitric oxide-inducednitrative stress involved in microbial pathogenesis. J. Phar-macol. Sci. 98, 117–129.


















