
Induction of teratogenesis of freshwater Snail ... fileInduction of teratogenesis of freshwater...
15
Sci-Afric Journal of Scientific Issues, Research and Essays Vol. 2 (6), Pp. 255-268, June, 2014. (ISSN 2311-6188) http://www.sci-africpublishers.org Research Paper Induction of teratogenesis of freshwater Snail (Biomphalaria alexandrina) using the molluscicide Niclosamide. Hoda K. Salem, Nahla E.E. Omran, *Samia H. Eissa, Manar A. Kandeil Zoology Department, Faculty of Science, Tanta University, Tanta, Egypt. *Corresponding author e-mail: [email protected] Accepted June 12 th , 2014 ------------------------------------------------------------------------------------------------------------------------------------------------------------ ABSTRACT The present work aims to study the teratogenic effect of Niclosamide on Biomphalaria snail embryo and determine the more sensitive embryonic stage to this molluscicide substance. The subsequent or co-treatment with recovery agents such as vitamin C, holothurin extract and folic acid were also investigated. The developmental stages (cleavages, blastula, gastrula, trochophore, veliger, hippo- veliger and prehatching stages) revealed many deformations when exposed to sublethal concentrations of Niclosamide including vaculation, dissociation and degeneration of embryonic cells, arrested development, intensive pigmentation, enlarged head and foot and lyses of cells. The blastula and veliger stages can be considered specific stages for Niclosamide. SDS-PAGE electrophoresis of Niclosamide treated veliger showed increase of the intensity of some bands, disappearance of the band 158.5 kDa and appearance of the band 26.9 kDa. Also, exposure of veliger larvae to sublethal concentration of Niclosamide induced high increase in the total protein level, insignificant increase in the glycogen level and an insignificant decrease in the total lipid. Results indicated that folic acid was the most effective recovery agents. It is able to reverse the negative effects of Niclosamide especially when used alone post to Niclosamide. Folic acid brought all or most of the embryos towards their normal state. Vitamin C was effective for induction of normal embryos. The percentage was significantly elevated from 24.9% (post-treated) to 66.67% embryos co-treated with vitamin C. Also, holothurin can reverse the harm action of Niclosamide and recover a high percentage of malformed embryos (69.39%) especially when used alone. Key words: Freshwater snails, Biomphalaria alexandrina, Embryogenesis, Niclosamide Teratogenecity, Recovery. --------------------------------------------------------------------------------------------------------------------------------------------------------------- INTRODUCTION Niclosamide is a relatively selective, molluscicide; principally used effectively against freshwater snails that are intermediate hosts for schistosomiasis and fascioliasis. It has also been used in the control of cestodes infecting man as an antiparasitic drug in human and as a veterinary medicine. It is of very low toxicity to mammals but toxic to aquatic vertebrates, e.g. fish (Evans, 1983, Gool et al. 1984), amphibians and crustaceans (Oliveira-Filho and Paumgartten, 2000). Niclosamide is non-persistent in the aquatic environment, has a slight effect on aquatic plants and zooplankton and it is not generally phytotoxic at field concentrations (WHO, 2008). The most used synthetic molluscicide Niclosamide or bayluscide proved to be effective against all developmental stages of snails and schistosome (Tchounwou et al., 1991, 1992; Abu-Elyazeed et al., 1993 and Lowe et al., 2005). Also Niclosamide is currently the only molluscicide recommended by WHO for snail control programmes (Sturrock 2001; WHO 1993, 2006). Oliveira-Filho et al. (2010) mentioned that Niclosamide induced embryo mortality of Biomphalaria glabrata during the first 96 hrs (exposure period) and the latency to death becomes shorter as Niclosamide concentration increases. Mohamed et al. (2012) mentioned that Juvenile snails of Biomphalaria alexandrina were dead post 3 weeks of exposure to the sub-lethal concentration LC 50 of bayluscide. They concluded that bayluscide has deleterious effects on snails’ reproductive rate, their eggs and the intensities of DNA and RNA in their ovotestis–digestive gland complex. The rapid action of Niclosamide in killing snails is due to its toxic effect on the respiratory function of the snails by acting as uncoupler of oxidative phosphorylation at the mitochondrial level (White House, 1964 and Andrews et al. 1983).
Transcript of Induction of teratogenesis of freshwater Snail ... fileInduction of teratogenesis of freshwater...

Sci-Afric Journal of Scientific Issues, Research and Essays Vol. 2 (6), Pp. 255-268, June, 2014. (ISSN 2311-6188)
http://www.sci-africpublishers.org
Research Paper
Induction of teratogenesis of freshwater Snail (Biomphalaria alexandrina) using the molluscicide Niclosamide.
Hoda K. Salem, Nahla E.E. Omran, *Samia H. Eissa, Manar A. Kandeil
Zoology Department, Faculty of Science, Tanta University, Tanta, Egypt.
*Corresponding author e-mail: [email protected]
Accepted June 12th, 2014
------------------------------------------------------------------------------------------------------------------------------------------------------------
ABSTRACT
The present work aims to study the teratogenic effect of Niclosamide on Biomphalaria snail embryo and determine the more sensitive embryonic stage to this molluscicide substance. The subsequent or co-treatment with recovery agents such as vitamin C, holothurin extract and folic acid were also investigated. The developmental stages (cleavages, blastula, gastrula, trochophore, veliger, hippo-veliger and prehatching stages) revealed many deformations when exposed to sublethal concentrations of Niclosamide including vaculation, dissociation and degeneration of embryonic cells, arrested development, intensive pigmentation, enlarged head and foot and lyses of cells. The blastula and veliger stages can be considered specific stages for Niclosamide. SDS-PAGE electrophoresis of Niclosamide treated veliger showed increase of the intensity of some bands, disappearance of the band 158.5 kDa and appearance of the band 26.9 kDa. Also, exposure of veliger larvae to sublethal concentration of Niclosamide induced high increase in the total protein level, insignificant increase in the glycogen level and an insignificant decrease in the total lipid. Results indicated that folic acid was the most effective recovery agents. It is able to reverse the negative effects of Niclosamide especially when used alone post to Niclosamide. Folic acid brought all or most of the embryos towards their normal state. Vitamin C was effective for induction of normal embryos. The percentage was significantly elevated from 24.9% (post-treated) to 66.67% embryos co-treated with vitamin C. Also, holothurin can reverse the harm action of Niclosamide and recover a high percentage of malformed embryos (69.39%) especially when used alone.
Key words: Freshwater snails, Biomphalaria alexandrina, Embryogenesis, Niclosamide Teratogenecity, Recovery. --------------------------------------------------------------------------------------------------------------------------------------------------------------- INTRODUCTION
Niclosamide is a relatively selective, molluscicide; principally used effectively against freshwater snails that are intermediate hosts for schistosomiasis and fascioliasis. It has also been used in the control of cestodes infecting man as an antiparasitic drug in human and as a veterinary medicine. It is of very low toxicity to mammals but toxic to aquatic vertebrates, e.g. fish (Evans, 1983, Gool et al. 1984), amphibians and crustaceans (Oliveira-Filho and Paumgartten, 2000). Niclosamide is non-persistent in the aquatic environment, has a slight effect on aquatic plants and zooplankton and it is not generally phytotoxic at field concentrations (WHO, 2008). The most used synthetic molluscicide Niclosamide or bayluscide proved to be effective against all developmental stages of snails and schistosome (Tchounwou et al., 1991, 1992; Abu-Elyazeed et al., 1993 and Lowe et al., 2005). Also Niclosamide is currently the only molluscicide recommended by WHO for snail control programmes (Sturrock 2001; WHO 1993, 2006).
Oliveira-Filho et al. (2010) mentioned that Niclosamide induced embryo mortality of Biomphalaria glabrata during the first 96 hrs (exposure period) and the latency to death becomes shorter as Niclosamide concentration increases. Mohamed et al. (2012) mentioned that Juvenile snails of Biomphalaria alexandrina were dead post 3 weeks of exposure to the sub-lethal concentration LC50 of bayluscide. They concluded that bayluscide has deleterious effects on snails’ reproductive rate, their eggs and the intensities of DNA and RNA in their ovotestis–digestive gland complex. The rapid action of Niclosamide in killing snails is due to its toxic effect on the respiratory function of the snails by acting as uncoupler of oxidative phosphorylation at the mitochondrial level (White House, 1964 and Andrews et al. 1983).

Sci-Afric J. Sci. Issues. Res. Essays Hoda et al 255
The relatively high toxicity of Niclosamide to freshwater snails is one of the reasons why it has been recommended by the World Health Organization as the molluscicide of choice for control of schistosome-bearing snails. In US, bayluscide (Niclosamide) is registered as a piscicide and used more to control sea lamprey and trash fishes than snails (Schreier et al. 2000). Even though reports are available on the toxicity of Niclosamide against some snail vectors, a detailed study on its teratogenic molluscicidal effects is lacking. So the study was designed to test the effect of lower doses of the drug on embryonic stages of snail hoping that these doses will not affect the non-target organisms.
Recovery study
Since the Niclosamide is used commonly in medical treatments for both human and cattles, the necessity for protection from its negative-effects has increased. It is thought that the harm effect of these chemicals could be tolerated by using some vitamins as vitamin C and folic acid .Vitamin C could exert chemo protective effects without apparent toxicity at doses higher than the current recommended dietary allowance of 60mg/day (Lee et al., 2003). Furthermore, vitamin C helps improve sperm count and motility and protects spermatozoa against endogenous oxidative DNA damage (Fraga et al., 1991).
According to Guillermo et al. (2007), consumption of antioxidants, such as folic acid, may mitigate the effects of the chemicals. Folic acid have the capacity to resist (or to neutralize) the effects of free radicals (Sai et al., 1992). The preclinical pharmacology of antitumor and cytotoxic marine compounds has been reported (Mayer, 1999; Mayer and Lehmann, 2001; Mayer and Gustafson, 2003, 2004, 2006, 2008). Holothurin has been reported to have some biological effects on free living as well as on parasitic protozoa (Styles, 1970; Sen et al., 1981). It has antiglycation, antioxidant activities and hypoglycemic activity (Francis et al., 2002; Abdel-Hassan et al., 2000; Rao and Gurfinkel, 2000). Indeed, these secondary metabolites have been reported to possess a wide spectrum of pharmacological effects including hemolytic, antitumoral, anti-inflammatory, antifungal, anti-bacterial, antiviral, ichthyotoxic, cytostatic and antineoplastic activities (Kerr and Chen 1995; Kalinin et al. 1996a, 1996b; Prokofieva et al. 2003). Many of these activities are the result of its active properties.
The present study investigated the efficacy of Biomphalaria alexandrina as a model species for environmental toxicology testing specifically, its developmental stages exposure to the common drug, Niclosamide. Also the study was aimed to elucidate the possible protective effects of some agents in alleviating the toxicity of Niclosdamide on snail embryo. MATERIALS AND METHODS
Experimental snails: Adult snails of Biomphalaria alexandrina (8-10 mm) were purchased from laboratory bred colony in Medical Malacology Department, Theodor Bilharz Research Institute (TBRI). The snails were kept in plastic containers (80 cm diameter and 20 cm depth) filled with dechlorinated tap water at 25 ± 3 °C with a 12/12 hours light/dark cycle. Snails were fed on fresh lettuce and water was renewed weekly.
Egg masses: The freshly egg masses of Biomphalaria alexandrina were collected daily on polyethylene sheets floating on each aquarium or from the wall of the glass using a spatula and were then maintained in climatic chambers at 25 °C (±3 °C) until the end of the experiments.
Experimental drug Niclosamide: (molluscicide with the common name bayluscide) was purchased from ADWIA (Egyptian Company for Chemicals and Pharmaceuticals), Cairo, Egypt. Chemical structure of Niclosamide is 2’5-dichloro-4' nitrosalicylanilide (C13H8Cl2N2O4). Molecular weight is 327.1. Niclosamide is a yellowish grey, odourless crystalline solid with melting point between 224-229 °C.
Preparation of Niclosamide: A series of freshly prepared concentrations (0.1, 0.2, 0.3, 0.4, 0.5 ppm) that would permit the computation of LC50 and LC90 values were prepared.
EXPERIMENTAL DESIGN
Three experiments were undertaken during this study, they are: Study of the ovicidal activity and the degree of toxicity of Niclosamide; Evaluating the teratogenic potency of Niclosamide by determinging the specific stage of development, its protein profile and organic contents and Study of the effect of folic acid, vitamin C and holothurin as recovery agents against teratogenic effect of Niclosamide after or during exposure periods. Study the ovicidal activity and the degree of toxicity of Niclosamide
Experiment with snail eggs were conducted in test container (Petri-dishes) filled with 50 ml of a specific concentration of the testing chemical. Different developmental stages were exposed to series of concentrations of Niclosamide

Hoda et al 256
(0.1-0.5 ppm) to determine the LC50 and LC90. The lethal concentrations were calculated using statistical probit analysis (Finney, 1971). The developmental stages which were used in the ovicidal activity test were 2-cell stage, 4-cell stage, blastula stage, gastrula stage, trochophore larva and veliger larva. Three replicates were used for each test concentration. Mortality of embryos was indicated by many observable facts as embryo immotile, decaying embryonic tissues or absence of heart beat. Evaluation of teratogenic potency of Niclosamide by determination the specific stage of development, its protein profile and organic contents
The freshly laid egg masses containing fertilized eggs were exposed to different sub-lethal concentrations of the tested drug. Control and treated samples were held at constant temperature at 25 ± 3 °C. At least three concentrations with 3 replicates were used for the drug. Criteria for egg viability were based on the motility of the embryo within the egg. This could be distinguished by dissecting microscope examination. The effect of the tested molluscicide on different developmental stages of B. alexandrina was observed and photographed using digital camera attached to the microscope unit (Olympus microscope CX 31; Tokyo, Japan) to detect the embryonic abnormalities and to determine the affected stage and the most effective concentration for using in biochemical assay.
To determine the teratogenicity of Niclosamide, the treatment was initiated at blastula, gastrula, trochophore and veliger stages after seven. This teratogenic test of Niclosamide against developmental stages of B. alexandrina involved utilization of control groups exposed to dechlorinated tap water only. The experimental samples composed of 5 egg masses each with about 25 individuals. The sub-lethal concentrations used in this experiment were 0.07-0.08 ppm for Niclosamide, according to the LC50 investigation. Preparation of samples for biochemical studies
To study the protein bands, total protein, total lipid and glycogen of both control and treated egg masses of B. alexandrina, numerous freshly laid egg masses were collected and divided into 2 groups. The first group was the control one which exposed to dechlorinated tab water until the formation of veliger stage. The second group was exposed to 0.08 ppm Niclosamide for 5 days until the formation of the Veliger stage. Electrophoretic study
Total protein contents were extracted from 0.02 g of either the exposed or the control trochophore embryo using 200 µl of 0.1 M Tris-buffer (pH= 7.5) containing EDTA (0.01 M), KCl (0.01 M) MgCl2 (0.1 M) and 4% polyvinyl pyrrolidine (PVP). Tissue homogenates were centrifuged at 8050g for 20 min at 4 °C (Sigma 3K18 cooling centrifuge). The protein extract was then transferred into a fresh eppendorff tube and used directly for electrophoresis or kept at -20 °C until use. SDS-PAGE was performed on 8% separating and 4% stacking gels according to the method of Laemmli (1970). The gels were stained with 2% Coomassie Brilliant Blue R-250. A similarity matrix was constructed on the basis of the presence / absence of bands according to the similarity coefficient as described by Dice (1945) using the formula: S= 2a/2a+b+c, where a = number of bands shared between samples 1 and 2, b= the number of bands present in 1 but not in 2 and c = number of bands present in 2 but not in one. Biochemical assay Total protein: Total protein was determined using a commercial kit (Biomed Diagnostics, 30175 Hannover, Germany) and bovine serum albumin as a standard.
Total lipid: was determined using a commercial kit purchased from Biodiagnostic (Egypt) which depends on the reaction of lipids with vanillin in a medium of sulphuric acid and phosphoric acid to form pink colored complex (Zollner & Kirsch, 1962). Olive oil standards ranging between 5 and 40 mg/dl were used for standard curves. Samples were homogenized in phosphate buffer (0.1 M Na2HPO4, pH 7. Extraction and determination of glycogen
For extraction of glycogen, 200 mg of sample of embryos ground with 20 ml of 5% trichloroacetic acid (TCA) in a mortar or preferably in a homogenizer (Potter & Elvehjem, 1936). The precipitate of proteins was filtered off and the clear filtrate submitted to analysis. In a colorimeter tube (1.2 cm diameter) 2 ml of sample extract is added to 3 ml of iodine reagent. After mixing, the optical density was read in a photometer at 650 nm. The blank obtained by adding 2 ml of 5% TCA to 3 ml of reagent in the same way. The amount of glycogen is read from a calibration curve, which must be prepared using glycogen of the same origin as the sample to be estimated.
Hoda et al 257
Study the effect of folic acid, vitamin C and holothurin as recovery agents against teratogenic effect of Niclosamide after or during exposure periods. Determination of sub-lethal concentration of vitamin C, folic acid and holothurin: Vitamin C and folic acid were purchased from the local pharmacy. Holothurin extracted from the sea cucumber Holothuria polii was obtained as gift from Dr. Nahla Omran (Professor of Invertebrates, Zoology Department, Faculty of Science, Tanta University). A series of concentrations of vitamin C, folic acid and holothurin has been prepared to calculate the mortality rate. The concentration at which no mortality was obtained is considered a sub-lethal concentration.
Recovery of malformed embryos post-treated with vitamin (C), folic acid and holothurin: to examine the recovery role of vitamin (C), folic acid and holothurin on Niclosamide teratogenicity, freshly laid egg masses of B. alexandrina were collected. These egg masses were divided into two groups, Niclosamide treated group (0.08 ppm) and control group received no treatment. When the control egg masses reached veliger stage, the malformed embryos from the treated group were removed from the toxic agent and washed through several changes of dechlorinated tab water and transferred for further development to three petri dishes. The first contained the recovery agent vitamin (C), the second contained the recovery agent folic acid and the third contained the recovery agent holothurin. The concentrations used for vitamin (C), folic acid and holothurin were (5, 0.5 and 17 ppm) respectively. Control egg masses remained always in dechlorinated tab water. When the control group reached pre-hatching stage, the post-treated embryos were examined for growth and development. The percentage of recovered embryos was determined after 7 days.
Recovery of malformed embryos co-treated with the drugs and recovery agents: freshly laid egg masses of B. alexandrina were collected. These egg masses were divided into three groups, each of them treated with Niclosamide (0.08 ppm). At the same time of Niclosamide addition, the recovery agents (vitamin C, folic acid and holothurin) were added to each of these groups. The concentrations of recovery agents were as recorded in the previous experiment. The percentage of recovered embryos which complete their growth and development was determined after seven days from recovery. Statistical analysis
To compare the malformation frequencies among the control and treated groups, abnormality values of Niclosamide were expressed as a mean ± S.D., and statistically analyzed by one-way ANOVA test. Results of biochemical assays were statistically analyzed using Student's t-test to determine significant differences between treated and control samples. Statistical analysis was performed with the aid of the SPSS computer program (version 17 windows). RESULTS
Niclosamide LC50 and LC90 determination in different developmental stages of Biomphalaria alexandrina
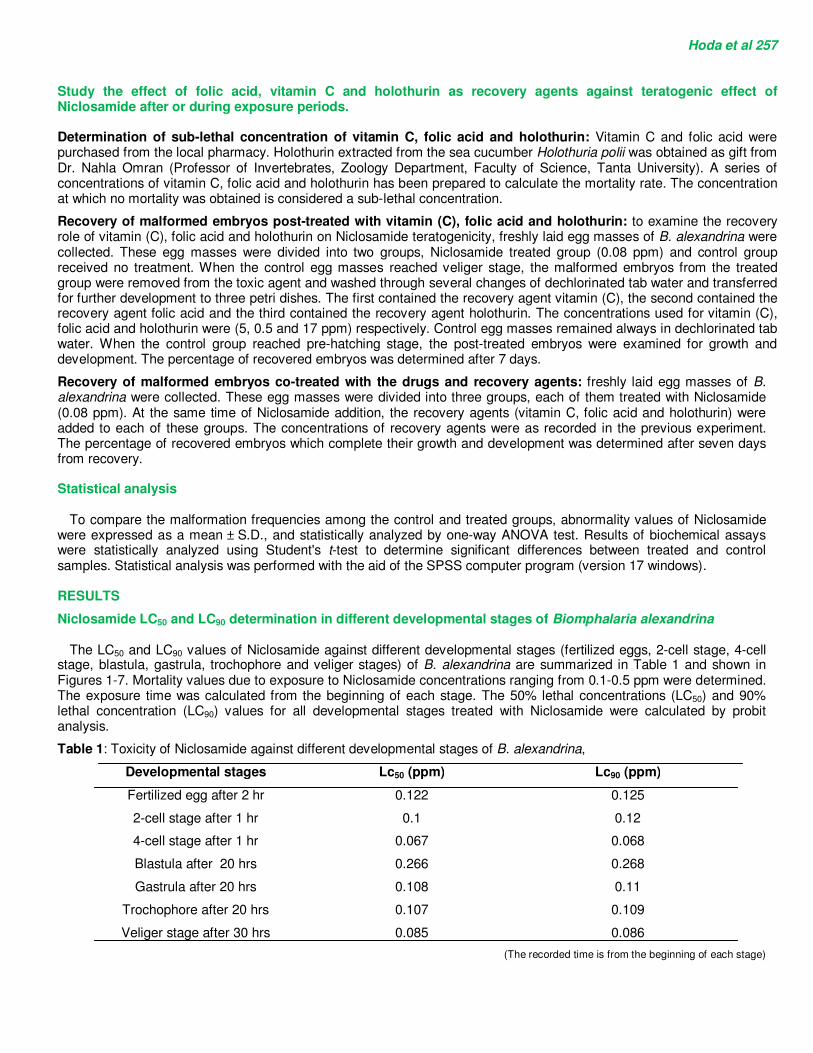
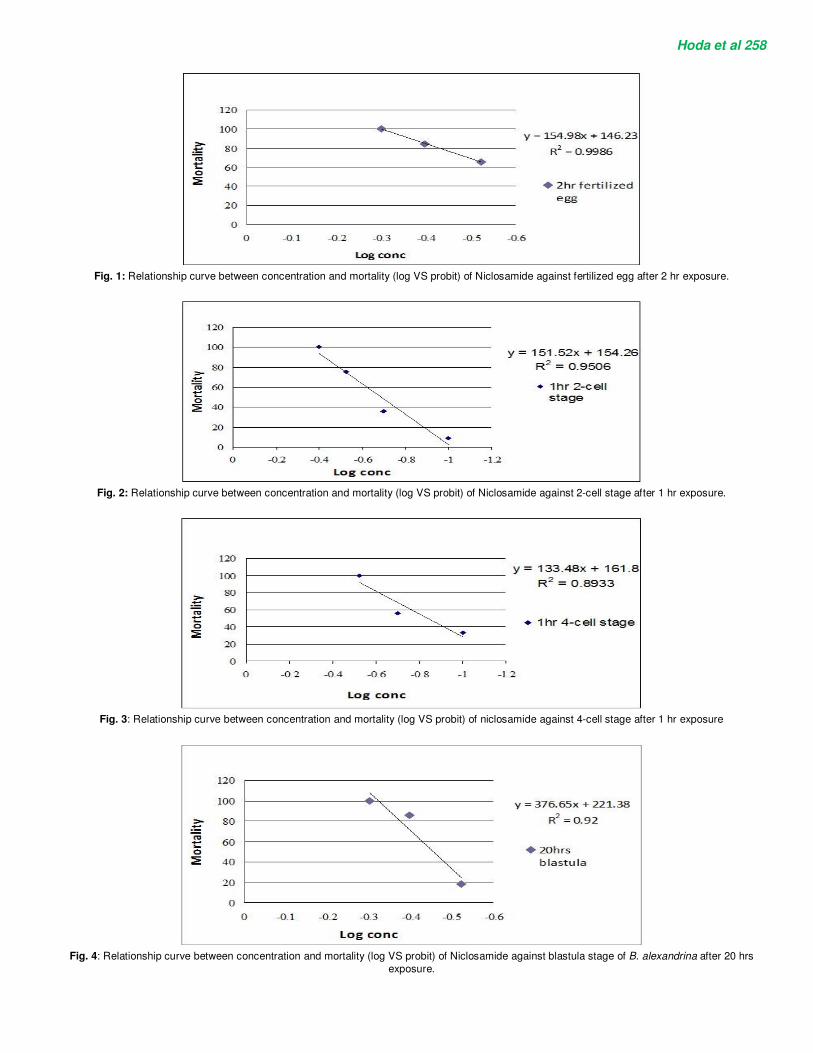
The LC50 and LC90 values of Niclosamide against different developmental stages (fertilized eggs, 2-cell stage, 4-cell stage, blastula, gastrula, trochophore and veliger stages) of B. alexandrina are summarized in Table 1 and shown in Figures 1-7. Mortality values due to exposure to Niclosamide concentrations ranging from 0.1-0.5 ppm were determined. The exposure time was calculated from the beginning of each stage. The 50% lethal concentrations (LC50) and 90% lethal concentration (LC90) values for all developmental stages treated with Niclosamide were calculated by probit analysis.
Table 1: Toxicity of Niclosamide against different developmental stages of B. alexandrina,
Developmental stages Lc50 (ppm) Lc90 (ppm)
Fertilized egg after 2 hr 0.122 0.125
2-cell stage after 1 hr 0.1 0.12
4-cell stage after 1 hr 0.067 0.068
Blastula after 20 hrs 0.266 0.268
Gastrula after 20 hrs 0.108 0.11
Trochophore after 20 hrs 0.107 0.109
Veliger stage after 30 hrs 0.085 0.086
(The recorded time is from the beginning of each stage)
Hoda et al 258
Fig. 1: Relationship curve between concentration and mortality (log VS probit) of Niclosamide against fertilized egg after 2 hr exposure.
Fig. 2: Relationship curve between concentration and mortality (log VS probit) of Niclosamide against 2-cell stage after 1 hr exposure.
Fig. 3: Relationship curve between concentration and mortality (log VS probit) of niclosamide against 4-cell stage after 1 hr exposure
Fig. 4: Relationship curve between concentration and mortality (log VS probit) of Niclosamide against blastula stage of B. alexandrina after 20 hrs
exposure.
Hoda et al 259
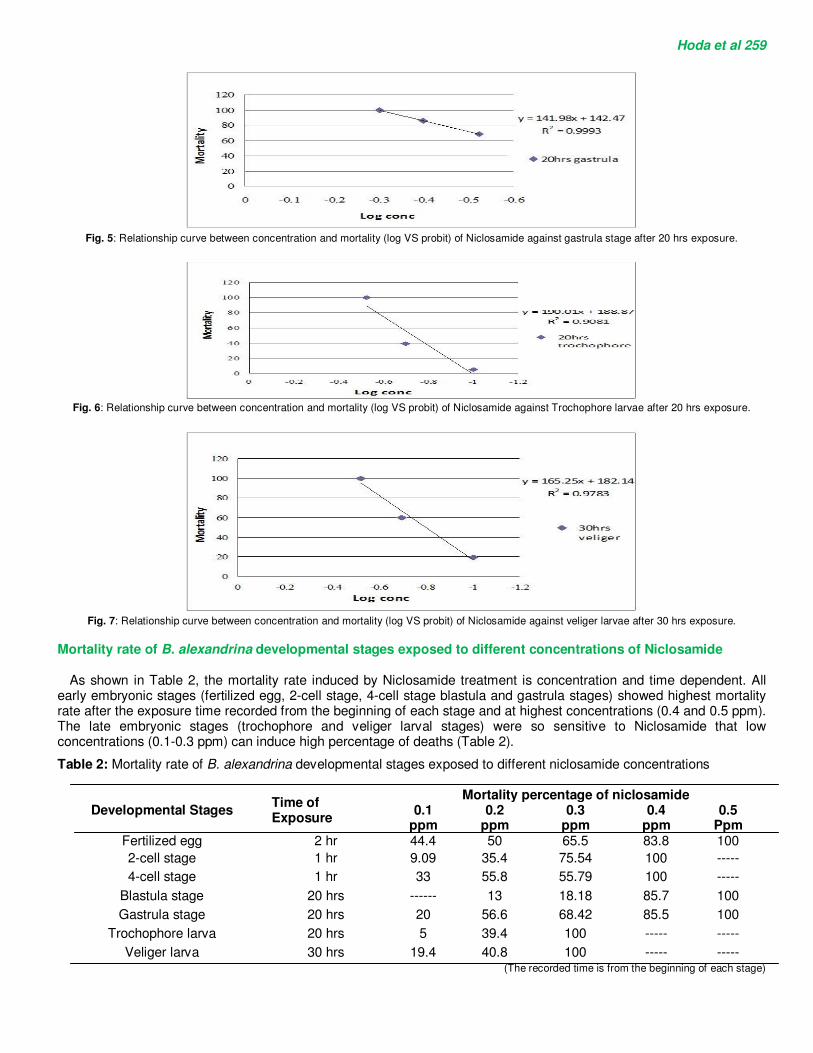
Fig. 5: Relationship curve between concentration and mortality (log VS probit) of Niclosamide against gastrula stage after 20 hrs exposure.
Fig. 6: Relationship curve between concentration and mortality (log VS probit) of Niclosamide against Trochophore larvae after 20 hrs exposure.
Fig. 7: Relationship curve between concentration and mortality (log VS probit) of Niclosamide against veliger larvae after 30 hrs exposure.
Mortality rate of B. alexandrina developmental stages exposed to different concentrations of Niclosamide
As shown in Table 2, the mortality rate induced by Niclosamide treatment is concentration and time dependent. All early embryonic stages (fertilized egg, 2-cell stage, 4-cell stage blastula and gastrula stages) showed highest mortality rate after the exposure time recorded from the beginning of each stage and at highest concentrations (0.4 and 0.5 ppm). The late embryonic stages (trochophore and veliger larval stages) were so sensitive to Niclosamide that low concentrations (0.1-0.3 ppm) can induce high percentage of deaths (Table 2).
Table 2: Mortality rate of B. alexandrina developmental stages exposed to different niclosamide concentrations
Developmental Stages Time of Exposure
Mortality percentage of niclosamide 0.1
ppm 0.2
ppm 0.3
ppm 0.4
ppm 0.5
Ppm Fertilized egg 2 hr 44.4 50 65.5 83.8 100
2-cell stage 1 hr 9.09 35.4 75.54 100 -----
4-cell stage 1 hr 33 55.8 55.79 100 -----
Blastula stage 20 hrs ------ 13 18.18 85.7 100
Gastrula stage 20 hrs 20 56.6 68.42 85.5 100
Trochophore larva 20 hrs 5 39.4 100 ----- -----
Veliger larva 30 hrs 19.4 40.8 100 ----- ----- (The recorded time is from the beginning of each stage)
Hoda et al 260 Determination of the specific stage (The stage at which high percentage of abnormal embryos was obtained)
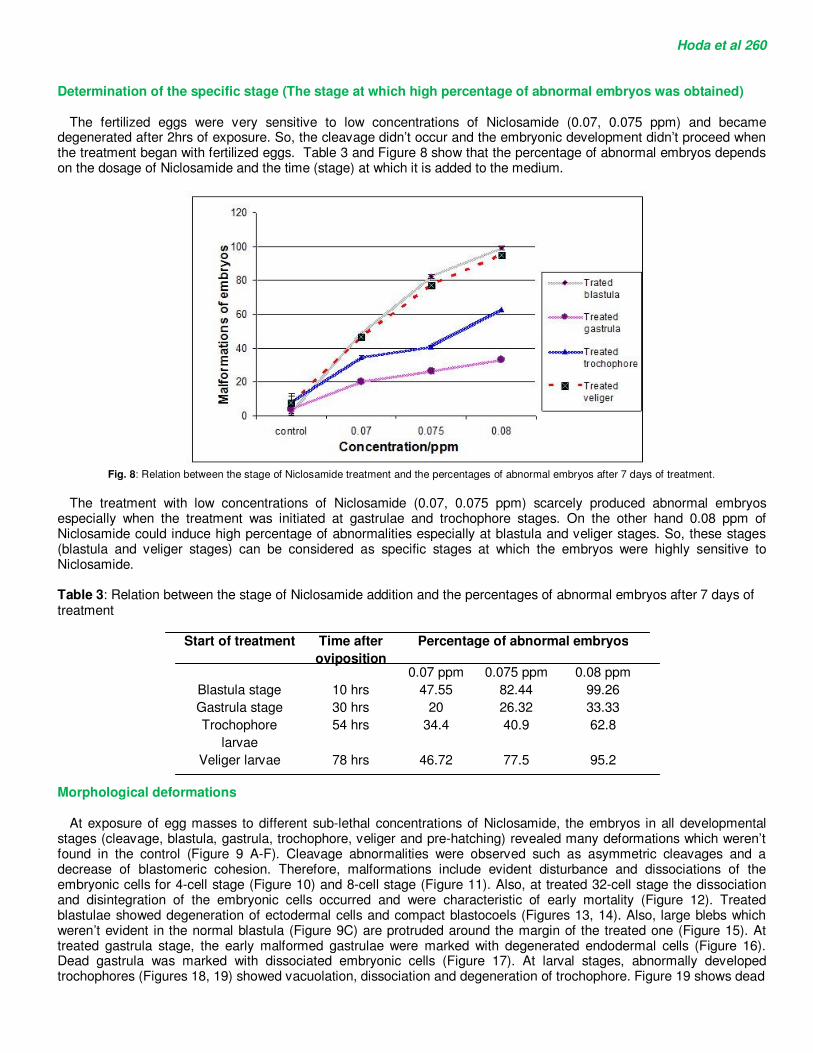
The fertilized eggs were very sensitive to low concentrations of Niclosamide (0.07, 0.075 ppm) and became degenerated after 2hrs of exposure. So, the cleavage didn’t occur and the embryonic development didn’t proceed when the treatment began with fertilized eggs. Table 3 and Figure 8 show that the percentage of abnormal embryos depends on the dosage of Niclosamide and the time (stage) at which it is added to the medium.
Fig. 8: Relation between the stage of Niclosamide treatment and the percentages of abnormal embryos after 7 days of treatment.
The treatment with low concentrations of Niclosamide (0.07, 0.075 ppm) scarcely produced abnormal embryos
especially when the treatment was initiated at gastrulae and trochophore stages. On the other hand 0.08 ppm of Niclosamide could induce high percentage of abnormalities especially at blastula and veliger stages. So, these stages (blastula and veliger stages) can be considered as specific stages at which the embryos were highly sensitive to Niclosamide. Table 3: Relation between the stage of Niclosamide addition and the percentages of abnormal embryos after 7 days of treatment
Percentage of abnormal embryos Time after
oviposition
Start of treatment
0.08 ppm 0.075 ppm 0.07 ppm
99.26 82.44 47.55 10 hrs Blastula stage
33.33 26.32 20 30 hrs Gastrula stage
62.8 40.9 34.4 54 hrs Trochophore
larvae
95.2 77.5 46.72 78 hrs Veliger larvae
Morphological deformations
At exposure of egg masses to different sub-lethal concentrations of Niclosamide, the embryos in all developmental stages (cleavage, blastula, gastrula, trochophore, veliger and pre-hatching) revealed many deformations which weren’t found in the control (Figure 9 A-F). Cleavage abnormalities were observed such as asymmetric cleavages and a decrease of blastomeric cohesion. Therefore, malformations include evident disturbance and dissociations of the embryonic cells for 4-cell stage (Figure 10) and 8-cell stage (Figure 11). Also, at treated 32-cell stage the dissociation and disintegration of the embryonic cells occurred and were characteristic of early mortality (Figure 12). Treated blastulae showed degeneration of ectodermal cells and compact blastocoels (Figures 13, 14). Also, large blebs which weren’t evident in the normal blastula (Figure 9C) are protruded around the margin of the treated one (Figure 15). At treated gastrula stage, the early malformed gastrulae were marked with degenerated endodermal cells (Figure 16). Dead gastrula was marked with dissociated embryonic cells (Figure 17). At larval stages, abnormally developed trochophores (Figures 18, 19) showed vacuolation, dissociation and degeneration of trochophore. Figure 19 shows dead
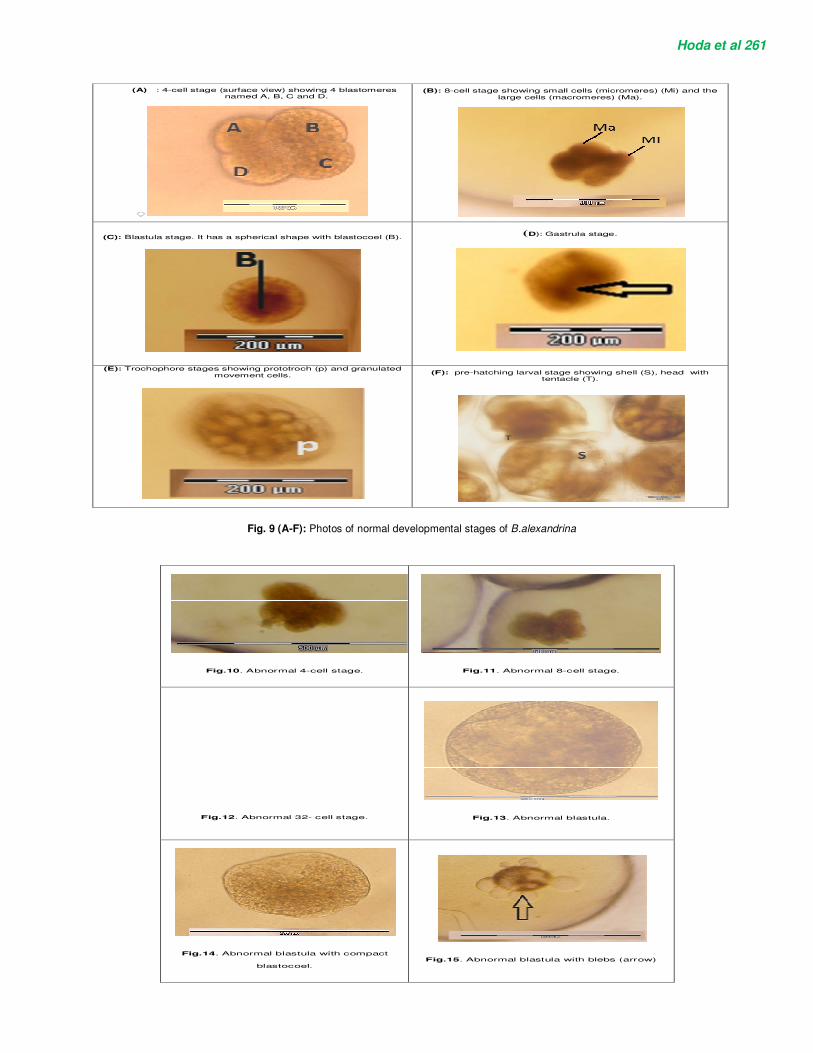
Hoda et al 261
(B): 8-cell stage showing small cells (micromeres) (Mi) and the
large cells (macromeres) (Ma).
(A) : 4-cell stage (surface view) showing 4 blastomeres named A, B, C and D.
◌
(D): Gastrula stage.
(C): Blastula stage. It has a spherical shape with blastocoel (B).
(F): pre-hatching larval stage showing shell (S), head with tentacle (T).
(E): Trochophore stages showing prototroch (p) and granulated movement cells.
Fig. 9 (A-F): Photos of normal developmental stages of B.alexandrina
Fig.10. Abnormal 4-cell stage.
Fig.11. Abnormal 8-cell stage.
Fig.12. Abnormal 32- cell stage.
Fig.13. Abnormal blastula.
Fig.14. Abnormal blastula with compact
blastocoel.
Fig.15. Abnormal blastula with blebs (arrow)
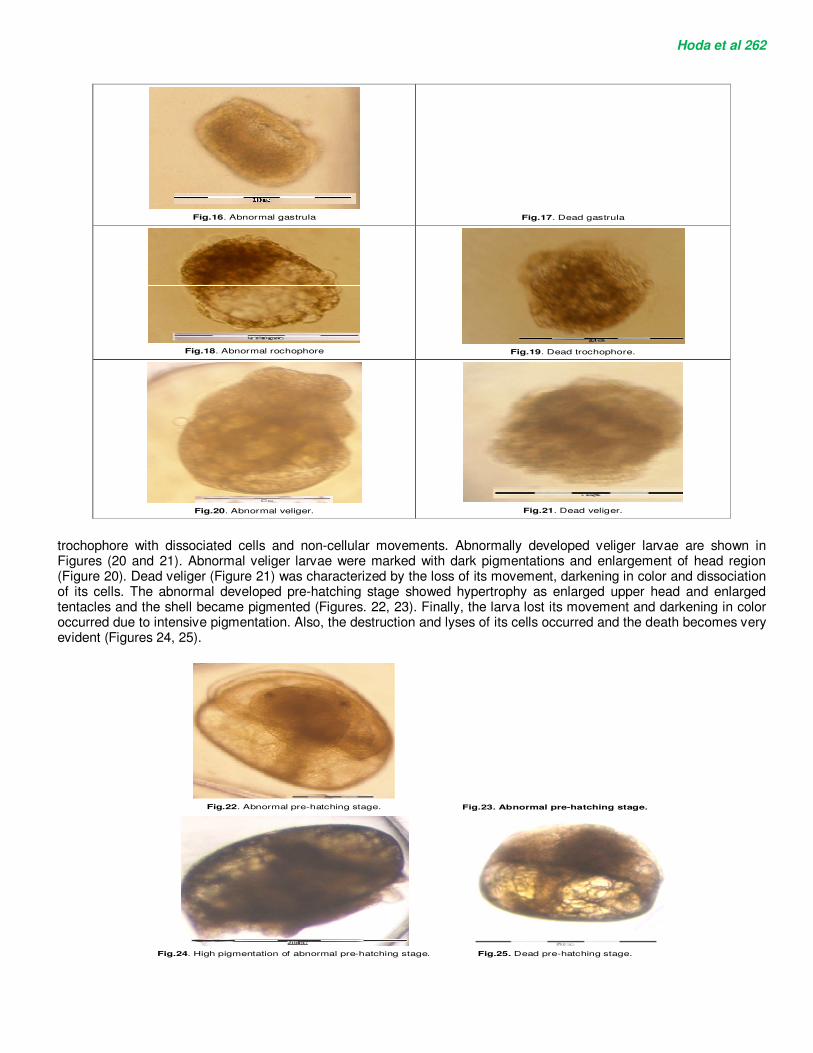
Hoda et al 262
Fig.16. Abnormal gastrula
Fig.17. Dead gastrula
Fig.18. Abnormal rochophore
Fig.19. Dead trochophore.
Fig.20. Abnormal veliger.
Fig.21. Dead veliger.
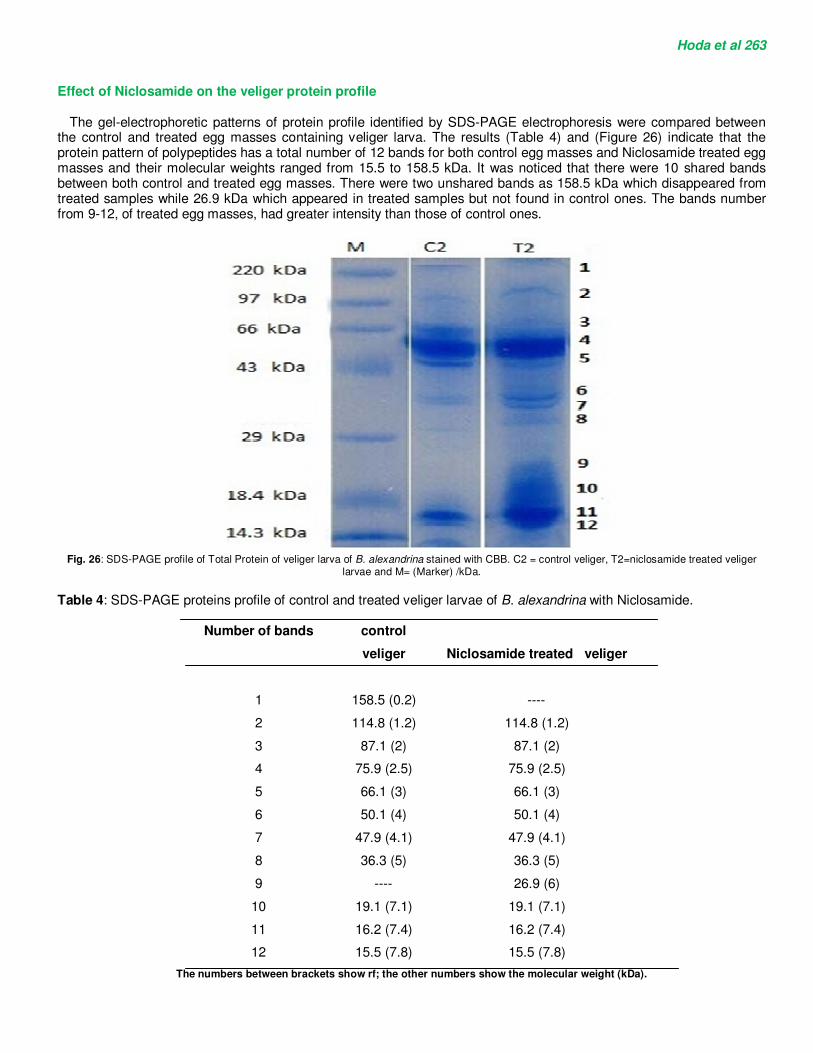
trochophore with dissociated cells and non-cellular movements. Abnormally developed veliger larvae are shown in Figures (20 and 21). Abnormal veliger larvae were marked with dark pigmentations and enlargement of head region (Figure 20). Dead veliger (Figure 21) was characterized by the loss of its movement, darkening in color and dissociation of its cells. The abnormal developed pre-hatching stage showed hypertrophy as enlarged upper head and enlarged tentacles and the shell became pigmented (Figures. 22, 23). Finally, the larva lost its movement and darkening in color occurred due to intensive pigmentation. Also, the destruction and lyses of its cells occurred and the death becomes very evident (Figures 24, 25).
Fig.22. Abnormal pre-hatching stage.
Fig.23. Abnormal pre-hatching stage.
Fig.24. High pigmentation of abnormal pre-hatching stage.
Fig.25. Dead pre-hatching stage.
Hoda et al 263 Effect of Niclosamide on the veliger protein profile
The gel-electrophoretic patterns of protein profile identified by SDS-PAGE electrophoresis were compared between the control and treated egg masses containing veliger larva. The results (Table 4) and (Figure 26) indicate that the protein pattern of polypeptides has a total number of 12 bands for both control egg masses and Niclosamide treated egg masses and their molecular weights ranged from 15.5 to 158.5 kDa. It was noticed that there were 10 shared bands between both control and treated egg masses. There were two unshared bands as 158.5 kDa which disappeared from treated samples while 26.9 kDa which appeared in treated samples but not found in control ones. The bands number from 9-12, of treated egg masses, had greater intensity than those of control ones.
Fig. 26: SDS-PAGE profile of Total Protein of veliger larva of B. alexandrina stained with CBB. C2 = control veliger, T2=niclosamide treated veliger
larvae and M= (Marker) /kDa.
Table 4: SDS-PAGE proteins profile of control and treated veliger larvae of B. alexandrina with Niclosamide.
Niclosamide treated veliger
control
veliger
Number of bands
---- 158.5 (0.2) 1
114.8 (1.2) 114.8 (1.2) 2
87.1 (2) 87.1 (2) 3
75.9 (2.5) 75.9 (2.5) 4
66.1 (3) 66.1 (3) 5
50.1 (4) 50.1 (4) 6
47.9 (4.1) 47.9 (4.1) 7
36.3 (5) 36.3 (5) 8
26.9 (6) ---- 9
19.1 (7.1) 19.1 (7.1) 10
16.2 (7.4) 16.2 (7.4) 11
15.5 (7.8) 15.5 (7.8) 12
The numbers between brackets show rf; the other numbers show the molecular weight (kDa).

Hoda et al 264
The similarity coefficient ″S″ was based on the number of protein profiles separated by SDS-PAGE (Table 5). The
results obtained revealed that the similarity between control and Niclosamide treated veligers was (0.91). Table 5: Matrix of similarity index (SD) of protein subunits in veliger larva of the freshwater B. alexandrina snails Effect of Niclosamide on organic content of veliger larvae
The low concentration of Niclosamide against egg masses containing veliger larvae induced many disturbances of organic substances when compared with the control. As shown in Table 6 there is a significant increase in the total protein level (3.76±0.13g/dl), with a ratio of increase reaching 15.3% (F = 0.459, df = 4 and P = 0.006). Also, there was an increase but insignificant in the glycogen level (4.66 ± 0.79 mg/100g of tissue), with a ratio of increase reaching 17.68% (F= 0.002, df = 4 and P = 0.34). In the case of lipid, the results revealed insignificant decrease in the total lipid (2.92 ± 0.73mg/dl), with a ratio of decrease reaching 14.37% (F = 0.495, df = 4 and P = 0.4). Table 6: The effect of sub-lethal concentration (0.08 ppm) of Niclosamide on biochemical parameters of egg masses of B. alexandrina containing veliger larvae
Groups Total protein
(g/dl) Percentage
change Total lipid
(mg/dl) Percentage
change
Glycogen mg/100g of
tissue
Percentage change
Control veliger
3.26 ± 0.10 3.14±0.42 3.96±0.77
Treated veliger
3.76±0.13* 15.34% ↑ 2.92±0.73 14.37% ↓ 4.66±0.79 17.68% ↑
*Significant (P<0.05)
Recovery study
The results of protective effect of some recovery agents (vitamin C, folic acid and holothurin) on Niclosamide-induced embryo injuries in the snail, Biomphalaria alexandrina are presented in Figure 27. Results indicated that folic acid was the most effective recovery agent. It could recover 99.5% of malformed embryos. It helps to reverse the negative effects of Niclosamide especially when used alone post to Niclosamide. The reversal effect of folic acid is complete and sustained and tended to bring all or most of the embryos towards their normal state. Therefore, folic acid (alone or concomitantly) afforded high protection against harm effects of Niclosamide. Also, vitamin C was effective for induction of normal embryos. The percentage was significantly elevated from 24.9% (post-treated) to 66.67% embryos co-treated with vitamin C. Also, holothurin can reverse the harm action of Niclosamide and recover a high percentage of malformed embryos (69.39%) especially when used alone.
Fig. 27: Percentage of recovered malformed embryos induced by Niclosamide.
2 1 Lan Number
1 Control veliger
1
1 0.91 Treated Niclosamide 2

Hoda et al 265 DISCUSSION Effect of Niclosamide on development
In this study, the molluscicidal effect of Niclosamide against embryonic development of B. alexandrina snails was tested. The results of mortality data of Niclosamide against all developmental stages pointed that 0.268 ppm Niclosamide is the highest LC90 values among all developmental stages. So, it is suggested to apply Niclosamide at 0.268 ppm in ponds. The study of Takougang et al. (2006) revealed that Niclosamide at concentration of 0.50 g/m
3
(ppm) applied in two rounds of 0.25 g/m3 resulted in high snail mortality and low lethality to fish, frogs, and tadpoles.
The obtained results showed that mortality rate induced by Niclosamide treatment is concentration and time-dependent. Also, the study of Oliveira-Filho et al. (2010) revealed that Niclosamide induced a concentration-dependent increase in embryo deaths. The study on the effect of Niclosamide against embryonic development showed that Niclosamide had adverse effect against the embryonic stages of B. alexandrina. This come in accordance with Sukumaran et al. (2004) who found that Niclosamide is toxic against eggs, immature, and adults of Lymnaea luteola.
Niclosamide is also reported as more toxic than nicotinanilide against young mature stage of L. auricularia (Parashar et al. 1995). The study of Mohamed et al. (2012) revealed that Niclosamide is more toxic to the snails than the pesticides Selecron and Basudin. This is parallel to the previous findings on the high toxicity of Niclosamide to B. alexandrina snails in comparison with some herbicides, pesticides and copper sulfate (El-Fiki and Mohamed, 1981 and El-Deeb and Ismail, 2003). The ovicidal action of Niclosamide against the eggs of B. alexandrina increase with the age of the fertilized eggs. The blastula and veliger stages can be considered as specific stages at which the embryos are highly sensitive to Niclosamide. This agrees with Gomot (1998) who found that pulmonate snail veliger stage was considered the most sensitive stage for toxicity studies. A similar observation was also reported by Parashar et al. (1990, 1995) where nicotinanilide affect 1 day and 4 to 5 days old eggs of L. auricularia. Also, Sukumaran et al. (2004) studied the ovicidal action of nicotinanilide against L. luteola and mentioned that the compound more toxic to the 72-96 hrs old eggs than to the 0-24 hrs old eggs. The ovicidal action of Niclosamide against the eggs of B. alexandrina increases with the age of the eggs due to protective covering of capsular jelly like material that covers and protects the freshly laid eggs from the external environment. On contrary the jelly like egg protectant covering ova in L. natalensis was reported to be easily penetrated by Niclosamide and produced 100% mortality (Gillet & Bruaux, 1962). Zhang and Guo (1992) reported that the eggs of Oncomelania snails exposed to bromoacetamide showed the deformation in the earlier stages but not in the later stages. Sukumaran et al. (1994, 1995, and 2002) also reported that the n-butanol extracts of some plant molluscicides like Sapindus trifoliatus, Agave americana, Balanites aegyptica, Jatropha gossypifolia and Vaccaria pyramidata is toxic against freshly laid eggs of L. luteola.
Concerning the effect of Niclosamide on protein synthesis, it was found that Niclosamide induced increase of the intensity of some bands of treated embryos, disappearance of one band from treated egg masses and appearance of one band which not found in the control. The electrophoretic pattern of the proteins of B. alexandrina snails treated with methanol extract from some plants revealed difference in the number and molecular weights of protein bands compared to the control snails (Bakry, 2009). He mentioned that differences indicated that these plant extracts caused intensive molluscicidal effects which induced fractionation of the native protein. He suggested that snails that gave similar type of protein bands to that of the control were able to resist molluscicidal effect. Protein changes due to treatment with some molluscicides were previously detected by Rawi et al. (1993), Aly et al. (2004) and El-Sayed (2006). Accordingly, the fractionation of native proteins into bands different from that of the control may be attributed to changes occurred in DNA of the treated snails. This suggestion is confirmed by the increase of the DNA concentration shown in the treated snails. The DNA intensity was obviously increased in the treated snails than that of control (El- Sayed, 2006 and Bakry, 2009).The present study indicated that some proteins synthesized in normal embryos were more prominent than those in treated embryos. This result suggests that these proteins can be regarded as the proteins involved in the cell differentiation of embryo. On the other hand, many of other proteins which were similarly synthesized in both types of embryos may be ″housekeeping″ proteins which are not necessary for cellular differentiation. Possible roles of these proteins which are responsible in the induction of malformations and in cell differentiation in snail embryos will become clearer by identification of these proteins.
With regard to organic content, total protein, total lipid and glycogen have been measured in this study. The low concentration of Niclosamide resulted in a significant increase in the total protein, insignificant decrease in the total lipid and insignificant increase in the glycogen level in the egg masses of B. alexandrina containing veliger larvae. Similar results regarding the increased protein level was reported by Mantawy et al. (2004) on testing the molluscicidal effects of Capparis spinosa and Acacia arabica leaves and Mantawy and Mahmoud (2002), on testing onion and garlic. They suggested that the snails produce additional proteins in response to the stressful conditions induced by the molluscicide. The rise in the content of total proteins in the snails could be related to the acceleration in the development of gluconeogenesis with the progressive level of intoxication caused by higher concentrations of E. splendens latex (Mello-Silva et al., 2006). Contradictory to these results are those obtained by Al-Sharkawy (1996) and Mantawy (2001), on

Hoda et al 266
investigating the molluscicidal effect of Ammi majus and Allium cepa & A. sativum, where marked reduction in protein was obtained and they referred this reduction to direct interference of active component of the extract with the protein synthesis. The decrease in the total lipid, in herein species, come in agreement with Adewunmi (1991) who observed that the development of Biomphalaria glabrata was arrested by Niclosamide (0.31-0.625 ppm). He concluded that the toxic effect of Niclosamide is due to the reduction in the energy content which is important for the development of the embryo and this indicated by a decrease in the total lipid content as a subsidiary source of energy on the stressed embryo. Recovery trials on malformed embryos The results of herein study indicated that niclosamide can be considered as a molluscicide and teratogen because it possesses developmental toxicity potential indicated by induction of snail embryo abnormalities (teratogenesis). The widespread use of this drug for pharmaceutical (medical) purposes by human and domestic animals has stimulated research for recovery agents against their negative effects. Also the great similarity between embryo cell and cancer cell (Andrews et al. 2005) provoke these recovery trials. According to Steinmetz and Potter (1996), phytochemicals have protective and recovery effects against many negative conditions. These substances also decrease significantly the percentage of formation of congenital malformations by affecting the cell differientation. The various phytochemicals have also important functions in DNA repair act, and a crucial role against the damages occurring in DNA. In addition folic acid also has a pivotal role in transporting the single carbon units especially in synthesis of DNA, RNA and proteins. Consequently, all these previous advantages of folic acid can be considered logic reasons for its strong positive action, which indicated in the present study, against niclosamide-induced malformations. Concerning the vitamin C, the present results revealed that it has a positive effect on niclosamide-induced abnormalities. The antiteratogenic capacity of vitamin C has been cited in many investigations; which strongly support our findings. Simán and Eriksson (1997) cleared that vitamin C reduced the rates of congenital malformations. Also, Rekka et al. (1992) referred to the ability of vitamin C of repair of tissue damage. They found that there is healing of tissue parenchyma and regeneration of renal tissue after administration of vitamin C. In addition, many investigators support the present results as Frei (2004); Ayo et al. (2006) and Ambali et al. (2007) who reported that vitamin C is an effective antioxidant in various biological systems. It was observed that, holothurin (saponin) can recover most of the niclosamide-induced malformations. Saponin can build up an organism's resistance to pernicious environmental factors and has special protective and repair functions. It has been reported that saponin can influence apoptosis by removing reactive oxygen species and by regulating the expression of proteins (Fang et al., 2002). The anti-teratogenic properties of saponin may operate by promoting the synthesis of nucleotides and proteins, enabling normal embryonic differentiation and development. The protective effects of saponin against niclosamide agree with Helian et al. (2003). They studied on murine fetal deformities caused by hyperthermia and found that saponin can restore embryos damaged by hyperthermia. The study elucidated the therapeutic effects of the recovery agents used as they caused significant improvements in teratogenicity and minimize the embryo toxicity. Their mechanisms are still obscure and need to be investigated. REFERENCES Abdel-Hassan IA, Abdel-Barry JA. and Mohammed ST. The hypoglycaemic and antihyperglycaemic effect of Citrullus colocynthis fruit aqueous extract
in normal and alloxan diabetic rabbits. Ethnopharmacol J 2000; 71: 325-330.
Abu-Elyazeed RR, Podgore JK, Mansour NS and Kilpatrick ME. Field trial of 1% niclosamide as a topical antipenetrant to Schistosoma mansoni
cercariae. Trop Med Hyg Am J 1993; 49: 403-409.
Adewunmi CO. Subchronic exposure of Biomphalaria glabrata eggs to aridanin and niclosamide. Ethnopharmacol J 1991; 31: 209-216.
Al-Sharakawy IM. Laboratory evaluation of the molluscicidal activity of Ammi majus (Umbilliferae) against Biomphalaria alexandrina snails. Ger Soc
Zool Egypt J 1996; 20: 227-244.
Aly SA, Aly HF, Saba-El-Rigal N and Sammou REM. Induced changes in biochemical parameters of the molluscan tissues non-infected using two
potent plants molluscicides. Parasitol Soc Egypt J 2004; 34(2): 527-542.
Ambali S, Akanbi D and Igbokwe N. Evaluation of subchronic chlorpyrifos poisoning on haematological and serum biochemical changes in mice and
protective effect of vitamin C. Toxicol Sc J 2007; 32: 111-120.
Andrews PW, Matin M M, Bahrami AR, Damjanov I, Gokhale P and Draper JS. Embryonic stem (ES) cells and embryonal carcinoma (EC) cells:
opposite sides of the same coin. Biochem Soc Trans 2005; 33(6): 1526-1530.
Andrews PW, Thyssen J and Lorke D. The biology and toxicology of molluscicides, Bayluscide®. Pharmacol Ther 1983; 19: 245-295.
Ayo JO, Minka NS and Mamman MM. Excitability scores of goats administered ascorbic acid and transported during hot-dry conditions. Vet Sc J 2006;
7: 127-131.
Bakry FA. Impact of some plant extracts on histological structure and protein patterns of Biomphalaria alexandrina snails. Mol Sci Global J 2009; 4:
34–41.
Dice LR. Measures of the amount of ecologic association between species. Ecol J 1945; 26: 297–302.
El-Deeb FA and Ismail NMM. Evaluation of some pesticides pre-exposure treatments on the efficiency of certain molluscicides against Biomphalaria
alexandrina snails. Appl Sci Egypt J 2003; 18 (5):618–624.

Hoda et al 267
El-Fiki SA and Mohamed AM. Effect of some herbicides on the toxicity of certain molluscicides against Biomphalaria alexandrina snails, Bilh Egypt J
1981; 5(1–2): 91–100.
El-Sayed KA. Effect of the plant Cupressus on some haematological and Biochemical parameters of Biomphalaria alexandrina snails. Parasitol Soc
Egypt J 2006; 36(3): 911-924.
Evans AC. Control of schistosomiasis in large irrigation schemes by use of niclosamide: a ten-year study in Zimbabwe. Trop Med Hyg Am J 1983;
32(5) 1029-1039.
Fang F, Chen XC and Zhu YG. Possible mechanism of mitochondria on anti-apoptotic effect of ginsenoside Rgl on MPP-induced cellular apoptosis.
Pharmal Ther Chin J 2002; 7: 412-416.
Finney D J. Probit Analysis. 3rd ed., Cambridge University Press, New Delhi 1971; 333 pp.
Fraga CG, Motchnik PA and Shigenaga MK. Ascorbic acid protects against endogenous oxidative DNA damage in human sperm. Proc Natl Acad Sci
USA 1991; 88:11003–11006.
Francis G, Kerem Z, Makkar HP and Becker K. The biological action of saponins in animal systems. Nutr Br J 2002; 88: 587-605.
Frei B. Efficacy of dietary antioxidants to prevent oxidative damage and inhibit chronic disease. Nut J 2004; 134: 3196- 3198.
Gillet J and Bruaux P. A laboratory and field testing of (Bayer 73). Pflanzenschutz-Nachrichten Bayer 1962; 15: 70-74 (English edition).
Gomot A. Toxic effects of cadmium on reproduction, development, and hatching in the freshwater snail Lymnaea stagnalis for water quality monitoring.
Ecotoxicol Environ Safe 1998; 41:288–297.
Gool P H, Wilkins H A and De Marshall T F. Dynamics of Schistosoma haematobium infection in a Gambian Community. II. The effect on transmission
of the control of Bulinus senegalensis by the use of niclosamide. Tran R Soc Trop Med Hyg 1984; 78: 222-226.
Guillermo JRA, Lucia NCL, Javier GE and Virginia RN Methotrexate-induced mucositis in acute leukemia patients is not associated with the MTHFR
677T allele in Mexico. Hematol 2007; 5: 387-391.
Helian LI, Bowei W, Dan Z and Liying H. Protective effects of vitamin B12, ginseng saponin, and folic acid against murine fetal deformities caused by
hyperthermia. Chin Med J 2003; 116(11): 1776-1778.
Kalinin VI, Anisimov MM, Prokofieva NG, Avilov SA, Afiyatullov SH SH and Stonik VA. Biological activities and biological role of triterpene glycosides
from holothuroids (Echinodermata). In: Jangoux M and Lawrence LM (eds). Echinoderm studies. Balkema, Rotterdam 1996a; 139–181.
Kalinin VI, Prokofieva NG, Likhatskaya GN, Schentsova EB, Agafonova IG, Avilov SA. and Drozdova OA. Hemolytic activities of triterpene glycosides
from the holothurian order dendrochirotida: Some trends in the evolution of this group of toxins. Toxicon 1996b; (34): 475–483.
Kerr RG and Chen Z. In vivo and in vitro biosynthesis of saponins in sea cucumbers. Nat Prod J 1995; 58:172–176.
Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature Lond J 1970; 227:680-685.
Lee SE, Hwang HJ, Ha JS, Jeong HS and Kim JH. Screening of medicinal plant extracts for antioxidant activity. Life Sci 2003; 73: 167-179.
Lowe D, Xi J, Meng X, Wu Z, Qiu D. and Spear R. Transport of Schistosoma japonicum cercariae and the feasibility of niclosamine for cercariae
control. Parasitol Int 2005; 54: 83-89.
Mantawy M I. Potential effect of Allium cepa and Allium sativum on haemolymph of Biomphalaria alexandrina, the intermediate host of Schistosoma
mansoni. Soc Parasitol Egypt J 2001; 31(3):711-723.
Mantawy MM, Hamed MA, Sammour EM and Sanad M. Influence of Capparis spinosa and Acacia arabica on certain biochemical haemolymph
parameters of Biomphalaria alexandrina. Parasitol Soc Egypt J 2004; 4: 659-677.
Mantawy MM and Mahmoud AH. Effect of Allium cepa and Allium sativum feeding on glucose, glycogen, protein bands profile and phenol oxidase
activity in Biomphalaria alexandrina. Parasitol Soc Egypt J 2002; 32(1): 271-83.
Mayer AMS. Marine Pharmacology in 1998 Antitumor and Cytotoxic Compounds. Pharmacol 1999; 41: 159–164.
Mayer AMS and Gustafson KR. Marine pharmacology in 2000: antitumor and cytotoxic compounds. Cancer Int J 2003; 105: 291–299.
Mayer AMS and Gustafson KR. Marine pharmacology in 2001–2: antitumour and cytotoxic compounds. Cancer Eur J 2004; 40: 2676–2704.
Mayer AMS and Gustafson KR. Marine pharmacology in 2003–2004: anti-tumour and cytotoxic compounds. Cancer Eur J 2006; 42: 2241–2270.
Mayer AMS and Gustafson KR. Marine pharmacology in 2005–2006: antitumour and cytotoxic compounds. Cancer Eur J 2008; 44: 2357–2387.
Mayer AMS and Lehmann VKB. Marine pharmacology in 1999: antitumor and cytotoxic compounds. Anticancer Res 2001; 21: 2489–2500.
Mello-Silva CC, Vasconcellos MC, Pinheiro J and Rodrigues MLA. Physiological changes in Biomphalaria glabrata Say, 1818 (Pulmonata:
Planorbidae) caused by sub-lethal concentrations of the latex of Euphorbia splendens var. hislopii N.E.B (Euphorbiaceae). Mem Ins.t Oswaldo
Cruz 2006; 101: 1-6.
Mohamed AM, El-Emam MA, Osman GY, Abdel-Hamid H and Ali REM. Effect of Basudin, Selecron and the phytoalkaloid Colchicine (pesticides) on
biological and molecular parameters of Biomphalaria alexandrina snails. Pestic Biochem Phys 2012; 102: 68–78.
Oliveira-Filho EC, Barbara R, Geraldino BR, Coelho DR, De- Carvalho RR and Paumgartten FJR. Comparative toxicity of Euphorbia milii latex and
synthetic molluscicides to Biomphalaria glabrata embryos. Chemosphere 2010; 81: 218–227.
Oliveira-Filho EC and Paumgartten FJR. Toxicity of Euphorbia milii Latex and Niclosamide to Snails and Nontarget Aquatic Species. Ecotoxicol
Environ Safe 2000; 46: 342-350.
Parashar BD, Kaushik MP, Gupta AK, Swamy RV and Rao KM. Toxicity of some molluscicids to fresh water snail Lymnaea auricularia the vector of
animal fasciolasis and to non-target organisms. Proc Acad Environ Biol 1995; 4: 183-187.
Parashar BD, Kaushik MP, Shriprakash and Rao KM. Toxicity of nicotinanilide and its analogues to the freshwater snail Indoplanorbis exustus,
mediator of animal schistosomiasis and to non-target organism. Appl Malacol Med J 1990; 2: 135-140.
Potter VR and Elvehjem CA. The effect of selenium on cellular metabolism. The rate of oxygen uptake by living yeast in the presence of sodium
selenite. Biochem J 1936; 30: 189–0.
Prokofieva NG, Chaikina E L, Kicha AA and Ivanchina, NV. Biological activities of steroid glycosides from starfish. Biochem Phys C. (2003) (134):
695–701.
Rao AV and Gurfinkel DM. The bioactivity of saponins: triterpenoid and steroid glycosides. Drug Metab Drug Interact 2000; 17:211–235.
Rawi SM, El-Gindy HI and Abd-Kader A. The effect of some molluscicides on total protein, albumin, globulin and total lipid of the snail Biomphalaria
alexandrina. Bull Fac Sci Cairo Univ 1993; 61: 87-101.

Hoda et al 268
Rekka E, Kourounakas P, Shahidi F, Janitha PK and Anasundara PD. Phenolic antioxidants. Critical Reviews in Food Science and Nutrition 1992; 32:
67-103.
Sai K, Hayashi M and Takagi A. Effects of antioxidants on induction of micronuclei in rat peripheral blood reticulocytes by potassium bromate. Mutat
Res 1992; 269: 113–118.
Schreier TM, Dawson VK, Choi Y, Spanjer NJ and Boogaard MA Determination of niclosamide residues in rainbow trout (Oncorhynchus mykiss) and
channel catfish (Ictalurus punctatus) fillet tissue by HPLC. Food Chem J Agric 2000; 48: 2212-2215.
Sen DK, Lankaranifard G and Narain NK. Effect of holothurin on Typanosoma musculi infection in three strains of mice. Protozool J 1981; 28: 167-170.
Simán CM and Eriksson UJ. Vitamin C supplementation of the maternal diet reduces the rate of malformation in the offspring of diabetic rats.
Diabetologia 1997; 40 (12): 1416–1424.
Steinmetz KA and Potter JD. Vegetables, fruit, and cancer prevention. Diet Assoc Am J 1996; 96: 1027–1039.
Sturrock RF Schistosomiasis epidemiology and control: How did we get here and where should we go? Mem Inst Oswaldo Cruz 2001; 96 (Suppl): 17-
27.
Styles T. Effect of holothurin on trypanosome lewisi infections in rats. Protozool J 1970; 17: 196-198.
Sukumaran D, Parashar BD, Gupta AK, Jeevaratnam K and Prakash S. Molluscicidal effect of nicotinanilide and its intermediate compounds against a
freshwater snail Lymnaea luteola, the vector of animal schistosomiasis. Mem Inst Oswaldo Cruz 2004; 99: 205–210.
Sukumaran D, Parashar, BD and Rao KM. Molluscicidal properties of Agave americana and Balanites aegyptica. Pharmacogn Int J 1994; 32: 232-
238.
Sukumaran D, Parashar BD and Rao KM. Toxicity of extracts Jatropha gossypifolia and Saponaria saponaria as molluscicide against freshwater snails
Lymnaea luteola and Indoplanorbis exustus, the vectors of animal schistosomiasis. Fitoterapia 1995; LXVI: 393-398.
Sukumaran D, Parashar BD and Rao KM. Evaluation of some plant molluscicides against a freshwater snail Lymnaea luteola, the vector of animal
schistosomiasis. Pharmaceutical Biol 2002; 40: 450-455.
Takougang I, Meli J and Angwafo F. Field trials of low dose Bayluscide on snail hosts of schistosome and selected non-target organisms in sahelian
Cameroon. Mem Inst Oswaldo Cruz, Rio de Janeiro 2006; 101(4): 355-358.
Tchounwou PB, Englande, Jr AJ and Malek EA. Toxicity Evaluation of Bayluscide ® and Malathion to Three Developmental Stages of Freshwater
Snails. Arch Environ Contam Toxicol 1991; 21:351-358.
Tchounwou PB, Englande Jr AJ and Malek EA. The influence of selected environmental parameters on the toxicity of Bayluscide to Schistosoma
mansoni miracidia. Arch Environ Contam Toxicol 1992; 23: 223-229.
White House MM. Report of salicylanilide as decouplers of oxidative phosphorylation in rat mitochondria. Biochem Pharmacol 1964; 13: 319.
WHO 1993: The control of schistosomiasis Second Report of the WHO Expert Committee. WHO Technical Report Series 830 Geneva.
WHO 2006: Pesticides and their application for the control ofvectors and pests of public health importance. WHO/CDS/NTD/
WHOPES/GCDPP/2006.1.
WHO 2008: Data sheet on pesticides No.63 niclosamide. Available at http://www.inchem.Org/documents/pds/pds/pest63_e.htm.WHO/VBC /DS/88.63.
Zhang Y and Guo YH. Study on the effect of bromoacetamide upon the development of snail eggs. Parasitol Parasitic Dis J 1992; 10: 258-262.
Zollner N and Kirsch K. Uber die quantitative Bestimmung von Lipoiden (Mikromethode) mittels der vielen natiirlichen Lipoiden (allen bekannten
Plasmalipoiden) gemeinsamen Sulphophosphovanillin Reaktion. Z ges exp Med 1962; 135: 545-561.


















