
Independent research paper SES 2012 Michael Marty-Rivera ...
16
1 Marine Biological Laboratory Ecosystems Center Semester in Environmental Science Effects of nutrients on palatability and concentration of anti-herbivory compounds in macroalgae Independent research paper SES 2012 Michael Marty-Rivera Universidad de Puerto Rico Recinto Río Piedras Avenida Juan Ponce De León, San Juan, PR 00931 Advisor: Dr. Joe Vallino Marine Biological Laboratory 7 MBL Street, Woods Hole, MA 02543
Transcript of Independent research paper SES 2012 Michael Marty-Rivera ...

1
Marine Biological Laboratory
Ecosystems Center
Semester in Environmental Science
Effects of nutrients on palatability and concentration of anti-herbivory compounds in macroalgae
Independent research paper SES 2012
Michael Marty-Rivera
Universidad de Puerto Rico Recinto Río Piedras
Avenida Juan Ponce De León, San Juan, PR 00931
Advisor: Dr. Joe Vallino
Marine Biological Laboratory
7 MBL Street, Woods Hole, MA 02543

2
Abstract:
Macroalgae are a key component in aquatic ecosystems providing a food source for many organisms.
Many marine macroalgae have developed defensive mechanisms, such as anti-herbivory compounds. In
this study I will test for differences in herbivory on macroalgae grown under different nutrient
conditions by two amphipods, Jassa falcata and Gammarus oceanicus. Algal anti-herbivory compounds
and palatability was assayed by constructing synthetic algae leaf grids made up with agar and
dried/ground algae (Fucus vesiculosus and Gracilaria spp.) and feeding these to amphipods given the
selection of algae coming from two different nutrient availability conditions. A bioassay showed
Amphipods preference for Fucus vesiculosus coming from Sage Lot Pond, a low nutrient site, with a
consumption of 46% over the Child’s river (high nutrient site) variety. Phenolic compounds found in
Fucus vesiculosus did not appear to inhibit amphipod grazing. No preference was observed for Gracilaria
spp. and this can serve as an indication of no chemical defenses against Jassa falcata and Gammarus
oceanicus.
Keywords: Macroalgae, Amphipod, herbivory, nutrients, Anti-herbivory compounds, bioassay.
Introduction:
Macroalgae form part of the trophic web and are an important primary producer in aquatic ecosystems,
therefore a key part of the ecosystem energy and nutrient cycle. Recent research has shown that
macroalgae can serve as a bioremediation agent via nutrient removal from eutrophic waters (Marinho-
Soriano 2009 et al.; Marinho-Soriano et al. 2011). There are also some commercial products derived
from algae such as carrageenan, agar, fertilizers (Chynoweth 2001). Algae biomass has also been found
to be a good candidate for biofuel production since the 1970’s (Roesijadi 2010).

3
Macroalgae have several needs for its optimal growth such as light, temperature, salinity, available
space and nutrients (Pederson et al. 1996) and have developed mechanisms to prevent herbivory, this
gives them an opportunity to thrive in ecosystems with many herbivores and be exploited for
anthropogenic benefits. The types of algae defenses can be separated as structural, chemical, seasonal
(Hay and Fenical 1988). This study will focus on the chemical defenses of macroalgae based on its
growing environment. Two sites in the Waquoit Bay area (Figure 1), Child’s River (CR) an estuary which
receives a nutrient loading of 407 kg N * Ha-1 * year-1, and Sage lot Pond (SLP) which receives only 7.6 kg
N * Ha-1 * year-1 (Hauxwell et al. 2003), were used to compare the effects of the brown algae Fucus
Vesiculosus, the red algae Gracilaria spp. and the green algae Ulva lactuca anti-herbivory compounds
against common herbivores in the area, the amphipods Gammarus oceanicus and Jassa falcata.
Macroalgae grown in a nutrient enriched area will have a higher growth rate than those grown in an
oligotrophic environment (Hemmi 2004) and ideally algae growth inhibited by a lack of nutrients will
have more resources to allocate into chemical defenses (Hemmi 2004). With those assumptions I
hypothesize that algae from the Child’s River site will be more susceptible to herbivory due to having a
greater nutrient availability for growth, therefore not needing to invest energy in defensive chemicals.
Methods:
I collected macroalgae from the coastlines of both study sites, cleaned with seawater to remove visible
epiphytes, oven-dried at 60°C and ground to a fine powder. I prepared an agar mixture using 1L filtered
seawater and 20g of Agar reagent and autoclaved for 3 minutes. Once the agar cooled down, I poured
50mL into a beaker along with 5g of ground species of algae and mixed thoroughly using a glass stir rod
and once homogeneous it was poured over a sheet of wax paper with four plastic grid squares in its
center. A new sheet of wax paper was placed on top of the mixture and a cinderblock was placed on top

4
to weigh down on the agar to force it into the plastic grids (Figure 2). With these, an incubation tank was
set up using algae square grids (AG) of two different species and 3 amphipods (either Jassa falcata or
Gammarus oceanicus) per AG. This was done for both Gracilaria spp. and Fucus Vesiculosus algae. In
order to test differences in amphipod selectivity of algae from the same site I set up another tank
experiment with two replicates of Gracilaria spp. and Fucus Vesiculosus and Ulva lactuca AG from
Child’s River and 6 Jassa falcata or Gammarus oceanicus per AG.
I performed an adaptation of Folin-Ciocalteau’s for determination of phenolic compounds, where tannic
acid was used instead of gallic acid as a standard. I extracted dried and ground algae with 40% Ethanol,
sonicated for 2 minutes and shaked for 2 hours. Then I filtered the extracted material using a 47mm
swinnex filter and analyzed in a Shimadzu UV-Vis spectrometer at a wavelength of 790nm.
I did the Elemental analysis using a Europa ANCA-SL elemental analyzer - gas chromatograph
preparation system attached to a continuous-flow Europa 20-20 gas source stable isotope ratio mass
spectrometer following standard operation procedures for plant material of the Ecosystems Center -
Marine Biological Laboratory Stable Isotope Laboratory. I analyzed Nitrate and Ammonia in water
samples from both CR and SLP following Soloranzo (1969) and Wood et al. (1967) procedures,
respectively.
Results:
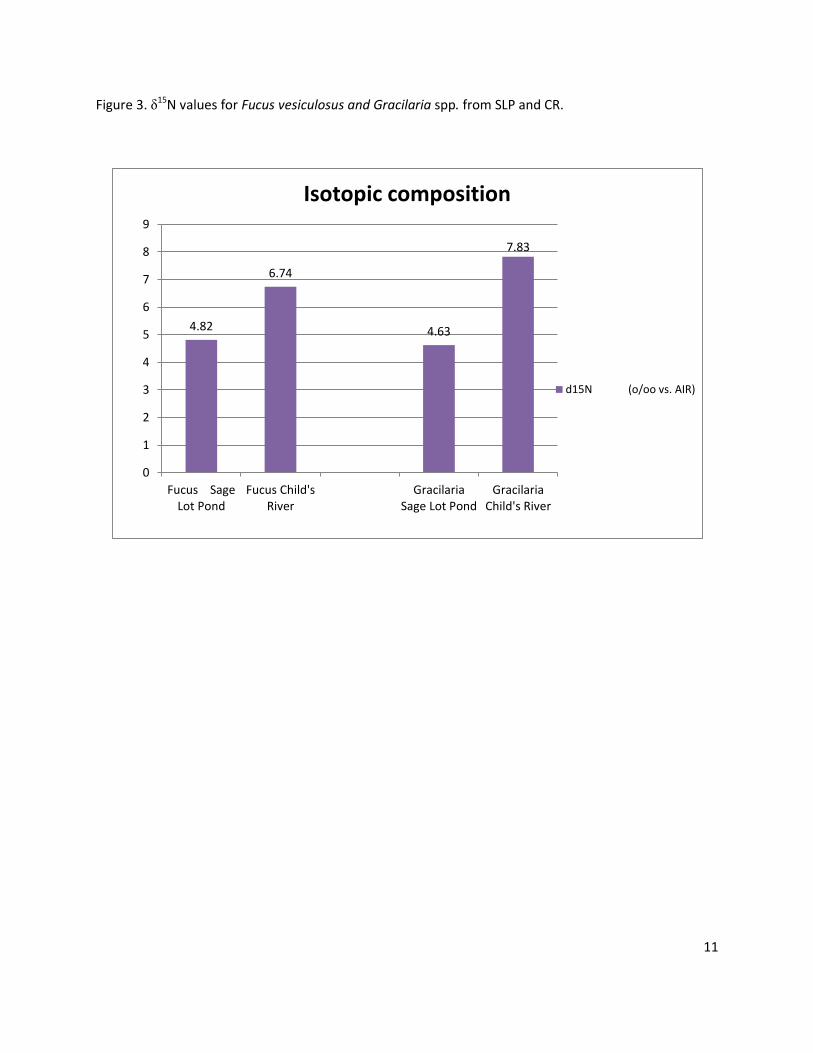
I performed several tests to look for differences between sites, algae and herbivory. Figure 3 shows δ15N
values of Fucus vesiculosus and Gracilaria spp. coming from both study sites where higher value can be
seen for both species at CR ( 6.74 and 7.38 0/00 vs Air respectively ). Figure 4 shows carbon and nitrogen
molar ratio (C:N) for Fucus vesiculosus and Gracilaria spp. where CR algae have a lower C:N than their

5
SLP counterpart, a lower value meaning less carbon per every nitrogen. Nitrate and ammonia were also
measured for SLP and CR (Table 1). Again CR has a higher value for both of these nutrients. With this
ongoing trend of data I assumed both sites to be considerably different and performed a series of
bioassays. The first assay is shown in Figure 5 where both amphipods consumed 40 to 50% more from
the CR Fucus spp. than the SLP variety. The Gracilaria spp. assay showed a small difference between
algae consumed of about 76 to 92% in favor of the CR algae (Figure 6). The third assay comparing algae
from CR showed both amphipods showing preference to the Ulva spp. over the other algae. In the assay
using Gammarus oceanicus the Gracilaria spp. was also heavily grazed on (Figure 7).
Discussion:
Isotope values from CR can serve as an indication of an anthropogenic input of nutrients to the system
because human generated wastes generally have a higher δ15N value (Valiela 2000). And the C:N ratio
can help confirm this idea since the algae from CR have a lower ratio, meaning there are less carbon per
mole of nitrogen. Inorganic nutrients analyzed give another solid confirmation of differences in nutrients
available in both sites, showing a greater amount at CR.
My results indicate that there is a preference for Fucus vesiculosus from the SLP site, contrary to my
expectations; the phenolic compounds, shown in figure 8, produced by the Fucus spp. appear to have no
detrimental effect against either Jassa falcata or Gammarus oceanicus and may even be a beneficial
compound for such amphipods; as other herbivores can benefit from chemical compounds produced by
plants. However another way of analyzing my data is that amphipods might be consuming less algae
from CR because of its higher nutritional content and do not need to feed that often.

6
No apparent preference was observed for either amphipod in the Gracilaria spp. bioassay as they both
consumed a similar amount of algae from both sites. During this experiment I observed amphipod’s
behavior and noticed how both species of amphipod were hasty towards hiding underneath AG, this
type of behavior leads me to think that amphipods will prioritize hiding over feeding preferences.
Results from CR bioassay show that amphipod’s were consuming algae and had a higher preference for
the Ulva spp. over both Gracilaria spp. and Fucus spp. which can be due to being as less palatable than
Ulva spp.; this was expected as Ulva spp. is generally consumed by many herbivores. The assay also
showed that amphipods have different feeding rates as each bioassay had either Gammarus or Jassa
outcompeting one another.
Acknowledgments: I thank my advisor Joe Vallino for all of his help and advice throughout this project. I
also thank the course TA’s Rich McHorney, Carrie Harris, and Alice Carter, Ivan Valiela and Scott Lindell
for advice on project ideas, Jimmy Nelson for advice on bioassay preparation and Ken Foreman for his
advice on chemical compounds tests. This project was funded by the Semester in Environmental Science
Program 2010 with lab space and equipment provided by the Marine Biological Laboratory.
References:
Chynoweth, D.P., C.E. Turick, J.M. Owens, D.E. Jerger, Peck M.W. 1993.Biochemical methane potential of
biomass and waste feedstocks. Biomass and Bioenergy. 5:95-111.
Geiselman, J.A. 1980. Ecology of Chemical Defenses of Algae Against the Herbivorous Snail, Littorina Littorea, In the New England Rocky Intertidal Community. PhD thesis, Massachusetts Institute of Technology/Woods Hole Oceanographic Institution, Massachusetts, USA.

7
Hauxwell, J., J. Cebrian, I. Valiela. 2003. Eelgrass Zostera marinaloss in temperate estuaries: relationship to land-derived nitrogen loads and effect of light limitation imposed by algae. Marine Ecology Process Series. 247:59-73
Hay, M.E., and W. Fenical. 1988. Marine Plant-herbivore Interactions: The Ecology of Chemical Defense.
Ann. Rev. Ecol. Syst. 19:111-45 Hay, M.E., J.J. Stachowicz, E. Cruz-Rivera, S. Bullard, M.S. Deal, and N. Lindquist. 1998. Bioassays with
marine and freshwater macrwrganisms. Pages 39-141 in K.F. Haynes and J.G. Millar (eds.) Methods in Chemical Ecology. Volume 2, Bioassay Methods, Chapman and Hall, New York.
Hemmi, A., T. Honkanen, V. Jormalainen. 2004. Inducible resistance to herbivory in Fucus vesiculosus – duration, spreading and variation with nutrient availability. Marine Ecology Progress Series. 273: 109-120.
Marinho-Soriano, E., R.A Panucci,. M.A.A. Carneiro, D.C. Pereira.2009. Evaluation of Gracilaria Caudata J. Agardh for bioremediation of nutrients from shrimp farming wastewater. Bioresource Technology. 100:6192-6198.
Marinho-Soriano, E., C.A.A. Azevedo, T.G. Trigueiro, D.C. Pereira, M.A.A. Carneiro, M.R. Camara. 2011. Bioremediation of aquaculture wastewater using macroalgae and artemia. International Biodeterioration & Biodegradation. 65:253-257.
Pedersen, M.F. and J. Borum.1996. Nutrient control of algal growth in estuarine waters. Nutrient limitation and the importance of nitrogen requirements and nitrogen storage among phytoplankton and species of macroalgae. Marine Ecology Progress Series. 142:261-272.
Roesijadi, G., S.B. Jones, L.J. Snowden-Swan, Y. Zhu. 2010. Macroalgae as a Biomass Feedstock: A Preliminary Analysis. Pacific Northwest National Laboratory. PNNL-19944.
Soloranzo, L. 1969. Determination of ammonia in natural waters by the phenolhypochlorite method.
Limnol. Oceanogr. 14:799.
Valiela I., M. Geist, J. McClelland and G. Tomasky. 2000. Nitrogen loading from watershed to estuaries:
Verification of the Waquoit Bay Nitrogen Loading Model. Biogeochemistry. 49:277-293.
Wood, E.D., F.A.G. Armstrong, and F.A. Richards. 1967. Determination of nitrate in seawater by
cadmium-copper reduction to nitrite. J.Mar. Biol. Assoc. U.K. 47:23

8
Tables:
Table 1. Dissolved Inorganic Nutrient analysis for CR and SLP sites.
Sage Lot Pond Child's River
NH4 0.063 0.048
NO3 0.060 3.440

9
Figures:
Figure 1. Study site. Point A is Child’s River area, point B is Sage lot Pond.
41°33’ N , 70°31’ W

10
Figure 2. Algae square grid used for the bioassay.
11
Figure 3. δ15N values for Fucus vesiculosus and Gracilaria spp. from SLP and CR.
4.82
6.74
4.63
7.83
0
1
2
3
4
5
6
7
8
9
Fucus SageLot Pond
Fucus Child'sRiver
GracilariaSage Lot Pond
GracilariaChild's River
Isotopic composition
d15N (o/oo vs. AIR)

12
Figure 4. Carbon and nitrogen ratio of Fucus vesiculosus and Gracilaria spp.
21.88
15.59
11.94
8.63
0
5
10
15
20
25
FucusSage Lot
Pond
FucusChild'sRiver
GracilariaSage Lot
Pond
GracilariaChild'sRiver
Carbon & nitrogen composition
Mole C:N

13
Figure 5. Bioassay for the algae Fucus vesiculosus from SLP and CR. Average of four replicates.

14
Figure 6. Bioassay for the algae Gracilaria spp. from SLP and CR. Average of four replicates.

15
Figure 7. Bioassay for Child’s River macroalgae. Two replicates per algae.
-10
0
10
20
30
40
50
60
Jassa falcata Gammarus oceanicus
% o
f al
gae
co
nsu
me
dChild's River Assay
Ulva
Gracilaria
Fucus

16
Figure 8. Phenolic compounds concentration in Fucus vesiculosus, Gracilaria spp. and Ulva lactuca from
SLP sites.
0
0.5
1
1.5
2
2.5
3
FucusSage Lot
Pond
FucusChildsRiver
GracilariaSage Lot
Pond
GracilariaChildsRiver
UlvaChildsRiver
Co
nce
ntr
atio
n (
mg/
L)
Phenolic compounds


















