
Incompatibility in Populus: Structural and cytochemical...
8
Incompatibility in Populus: Structural and cytochemical characteristics of the receptive stigmas of Populus alba and P. nigra M. VILLAR', M. GAGET 2 , C. SAID 2 , R. B. KNOX 3 and C. DUMAS 2 ^Station d'Ainelioration des Arbres Forestiers, lnra Ardon 45160 Olivet, France 2 Recotmaissatice Cellulaire et Amelioration des Plantes, De'partement de Biologie Yegetale, Universite de Lyon I, 69622 Vdleurbanne Cedes France and UM CNRS 380024, France 3 Plant Cell Biology Research Centre, School of Botany, University of Melbourne, Parkville, Victoria 3052, Australia Summary Structural and cytochemical characteristics of stigmas of Populus alba (section Leuce) and P. nigra (section Aigeiros) have been studied to pro- vide an understanding of the cell biology of the interspecific incompatibility reactions between these two species. In addition to specific morpho- logical characteristics, stigma surfaces differ in the presence of an exudate studied by ultrastruc- tural and stigma print techniques. P. nigra shows features of a dry-type stigma covered by a pel- licle, whereas P. alba has a copious exudate typi- cal of a wet-type stigma that contains unsaturated lipids. Key words: Populus, stigma, exudate, interspecific incompatibility. Introduction Interspecific incompatibility in Populus has been extensively studied with the aim of obtaining hybrids for forest tree breeding programs (Knox etal. 1972a,b; Willing & Pryor, 1976; Stettler & Guries, 1976; Stettler et al. 1980). Hybrids are readily obtained between species within a section, but not between sections, except by manipulative techniques. Pollen mixtures (Mentor pollen technique, Stettler, 1968) or solvent treatment of the stigma surface (Willing & Pryor, 1976) have been used to overcome incompati- bility barriers. In attempts to understand the nature of interspecific incompatibility in Populus, we have explored the interactions between the male and female partners during incompatible crosses. Structural and biochemi- cal characteristics of the pollen and pistil have provided the first data on the informational molecules involved (Knox et al. \911a; Ashford & Knox, 1980). A key feature of the stigma surface is the pellicle, the initial membrane-like layer that receives pollen, and may regulate hydration in Brassica (Mattsson et al. 1974). Observations of poplar pollen tube behaviour have shown that stigma organization is a major factor in the control of incompatibility (Knox et al. 19726; Gaget et Journal of Cell Science 87, 483-490 (1987) Printed in Great Britain © The Company of Biologists Limited 1987 al. 1984). A comparison is made here of some cyto- logical and cytochemical characteristics of receptive stigmas of Populus nigra (section Aigeiros) and P. alba (section Leuce). Major differences in these features are reported between these two species. Materials and methods Populus nigra and P. alba shoots from female trees were obtained from the INRA Station des Arbres Forestiers, Orleans and grown in a greenhouse at the University of Lyon, Villeurbanne. For scanning electron microscopy (SEM), stigmas were fixed in 3 % glutaraldehyde in 0-1 M-sodium cacodylate buffer (pH 7-4) and prepared by the critical-point drying method using equipment manufactured by Balzers (Liechtenstein) and observed in a JEOL CF 35 SEM at 15kV at the CMEABG, University of Lyon. Uncoated fresh pistil samples were observed directly after rapid liquid nitrogen freezing (Dumas & Lecocq, 1975), under 15 kV on a Cambridge S 600 SEM. For cytochemistry, pistil samples were fixed in 3 % glutar- aldehyde in 0-1 M-sodium cacodylate buffer (pH 7-4) for 1 h, post-fixed in 1 % osmium tetroxide and embedded in Epon resin, according to the method of Dumas et al. (1978). Semithin sections of 1 ^m were obtained and stained for polysaccharides (PAS, Jensen, 1962), for lignin and phenolic components (Toluidine Blue, Trump et al. 1961), for 483
Transcript of Incompatibility in Populus: Structural and cytochemical...

Incompatibility in Populus: Structural and cytochemical characteristics
of the receptive stigmas of Populus alba and P. nigra
M. VILLAR', M. GAGET2, C. SAID2, R. B. KNOX3 and C. DUMAS2
^Station d'Ainelioration des Arbres Forestiers, lnra Ardon 45160 Olivet, France2Recotmaissatice Cellulaire et Amelioration des Plantes, De'partement de Biologie Yegetale, Universite de Lyon I, 69622 Vdleurbanne CedesFrance and UM CNRS 380024, France3Plant Cell Biology Research Centre, School of Botany, University of Melbourne, Parkville, Victoria 3052, Australia
Summary
Structural and cytochemical characteristics ofstigmas of Populus alba (section Leuce) and P.nigra (section Aigeiros) have been studied to pro-vide an understanding of the cell biology of theinterspecific incompatibility reactions betweenthese two species. In addition to specific morpho-logical characteristics, stigma surfaces differ inthe presence of an exudate studied by ultrastruc-tural and stigma print techniques. P. nigra shows
features of a dry-type stigma covered by a pel-licle, whereas P. alba has a copious exudate typi-cal of a wet-type stigma that contains unsaturatedlipids.
Key words: Populus, stigma, exudate, interspecificincompatibility.
Introduction
Interspecific incompatibility in Populus has beenextensively studied with the aim of obtaining hybridsfor forest tree breeding programs (Knoxetal. 1972a,b;Willing & Pryor, 1976; Stettler & Guries, 1976;Stettler et al. 1980). Hybrids are readily obtainedbetween species within a section, but not betweensections, except by manipulative techniques. Pollenmixtures (Mentor pollen technique, Stettler, 1968) orsolvent treatment of the stigma surface (Willing &Pryor, 1976) have been used to overcome incompati-bility barriers.
In attempts to understand the nature of interspecificincompatibility in Populus, we have explored theinteractions between the male and female partnersduring incompatible crosses. Structural and biochemi-cal characteristics of the pollen and pistil have providedthe first data on the informational molecules involved(Knox et al. \911a; Ashford & Knox, 1980). A keyfeature of the stigma surface is the pellicle, the initialmembrane-like layer that receives pollen, and mayregulate hydration in Brassica (Mattsson et al. 1974).Observations of poplar pollen tube behaviour haveshown that stigma organization is a major factor in thecontrol of incompatibility (Knox et al. 19726; Gaget et
Journal of Cell Science 87, 483-490 (1987)Printed in Great Britain © The Company of Biologists Limited 1987
al. 1984). A comparison is made here of some cyto-logical and cytochemical characteristics of receptivestigmas of Populus nigra (section Aigeiros) and P. alba(section Leuce). Major differences in these features arereported between these two species.
Materials and methods
Populus nigra and P. alba shoots from female trees wereobtained from the INRA Station des Arbres Forestiers,Orleans and grown in a greenhouse at the University of Lyon,Villeurbanne.
For scanning electron microscopy (SEM), stigmas werefixed in 3 % glutaraldehyde in 0-1 M-sodium cacodylate buffer(pH 7-4) and prepared by the critical-point drying methodusing equipment manufactured by Balzers (Liechtenstein)and observed in a JEOL CF 35 SEM at 15kV at theCMEABG, University of Lyon. Uncoated fresh pistilsamples were observed directly after rapid liquid nitrogenfreezing (Dumas & Lecocq, 1975), under 15 kV on aCambridge S 600 SEM.
For cytochemistry, pistil samples were fixed in 3 % glutar-aldehyde in 0-1 M-sodium cacodylate buffer (pH 7-4) for 1 h,post-fixed in 1 % osmium tetroxide and embedded in Eponresin, according to the method of Dumas et al. (1978).Semithin sections of 1 ^m were obtained and stained forpolysaccharides (PAS, Jensen, 1962), for lignin and phenoliccomponents (Toluidine Blue, Trump et al. 1961), for
483

proteins (Coomassie Blue, Fisher, 1968) and lipids (SudanBlack B, Bronner, 1975).
A method for microscopic examination of the surfacecomponents of stigmas was developed. Stigma prints wereobtained by pressing fresh stigmas between two glass sides,then fixed with glutaraldehyde or osmium tetroxide vapourand stained by different cytochemical techniques (Said et al.1985). For SEM observations, stigma prints were first fixedand gold coated before observation.
For enzyme cytochemical studies with transmission elec-tron microscopy (TEM), stigmas from receptive flowers weretreated with the reaction mixture for non-specific esterase(Mattsson et al. 1974) using (r-naphthyl acetate as substrateand hexazotized pararosanilin as coupling agent. Treatedstigmas were post-fixed in glutaraldehyde as described above,thin sections cut and observed at 80 kV with Hitachi HU 12Aand Philips EM 300 microscopes at CMEABG, University ofLyon. Ultrathin sections were post-stained with uranylacetate (lOmin) and lead citrate (8min) following Reynolds'(1963) procedures.
Additional cytochemical tests of the stigma surface in-cluded the use of lanthanum nitrate to test for surfacemembrane permeability, and cationized ferritin for surfacepolyanions (methods described by Gaude & Dumas, 1986).
Results
The receptive stigma surface
The stigmas of P. nigra (Fig. 1) and P. alba (Fig. 2)show striking differences in morphological character-istics (Table 1). In spite of the smaller stigmatic area ofP. alba, the receptive surfaces in both P. nigra and P.alba inflorescences are equivalent, because of the largernumber of flowers per catkin in P. alba. On the otherhand, pollination of P. alba stigmas is partly obstructedby numerous bract hairs borne near the base of thepedicel. In both species, the stigma papillae arerounded and bullate (Figs 3, 4), covered with smallspherical surface wax deposits.
Semithin sections of the receptive stigmas of P. nigraand P. alba showed that the receptive cells are charac-terized by the presence of large polyphenol vacuolesthat stain green with Toluidine Blue. Stigma cellsare generally much larger than their non-receptivecounterparts. Comparison is rendered difficult by thethickness of the papillar cell walls of both species. Onthe outer surface a positively stained surface film isapparent in sections stained with Coomassie Blue for
Table 1. Morphological characteristics of the femalecatkins of P. alba and P. nigra
Table 2. Cytochemistry of fresh stigma prints ofP. alba and P. nigra
Characteristics P. alba P. nigra
No. of flowers per catkin 110 40Size (mm) of stigmatic lobe (shape) 1-0 2-0
(filiform, (triangular,erect) procumbent)
No. of lobes 4 4
Stains Specificity P. nigra P. alba
Coomassie Blue ProteinsPAS PolysaccharidesEsterase activity Substrate:
c-naphthyl acetateSudan Black B Lipids
NT, not tested.
NT
proteins. This surface layer presumably representsboth cuticle and pellicle, since it is positively stainedwith the PAS reaction for polysaccharides and SudanBlack B for lipids (including cutin). The wall of thereceptive cells is multi-layered, and the major part ofthe wall is stained red with PAS, especially the innerlayer adjacent to the cytoplasm.
Cytochemistry of stigma prints
P. alba stigma prints (see Table 2) are obtained fromstigmatic compounds mainly localized in the clefts ofthe stigma surface. Each print comprises a distinctexudate when detected by SEM (Fig. 5), and showspositive staining with Coomassie Blue (Fig. 6) andesterase activity (Fig. 7). On the other hand P. nigraprints represent the wax droplets of the receptive cellsurface (Fig. 8). There is no staining for polysacchar-ides and proteins (Table 2).
Ultrastructure of the receptive stigma cellsThe receptive surface of P. nigra stigmas is covered bya diffuse electron-dense layer of irregular thickness(approximately 30 nm) comprising the surface pellicle(Fig. 9). Immediately beneath this layer, and largelyoutside the cuticle, are wax platelets. These surfacelayers contain cytochemically detectable reaction prod-uct for esterase (not illustrated). The underlyingcuticle is a thin layer abutting directly against the cellwall surface (Fig. 9). The wall of the receptive cellappears bilayered and heterogeneous: a reticulate outerzone containing spherical electron-opaque droplets,and a fibrillar inner zone (Fig. 9).
Further resolution of the stigma surface is providedby using two cytochemical probes: lanthanum nitratetreatment gives deposits in the pellicle and cell wall(Fig. 10); cationized ferritin-labelled preparationsshow deposits in the pellicle only (Figs 11, 12), whichis clearly differentiated from the underlying cuticle.
The mature stigma cell contains a large centralvacuole that displaces that cytoplasm to the periphery
484 M. Villar et al.
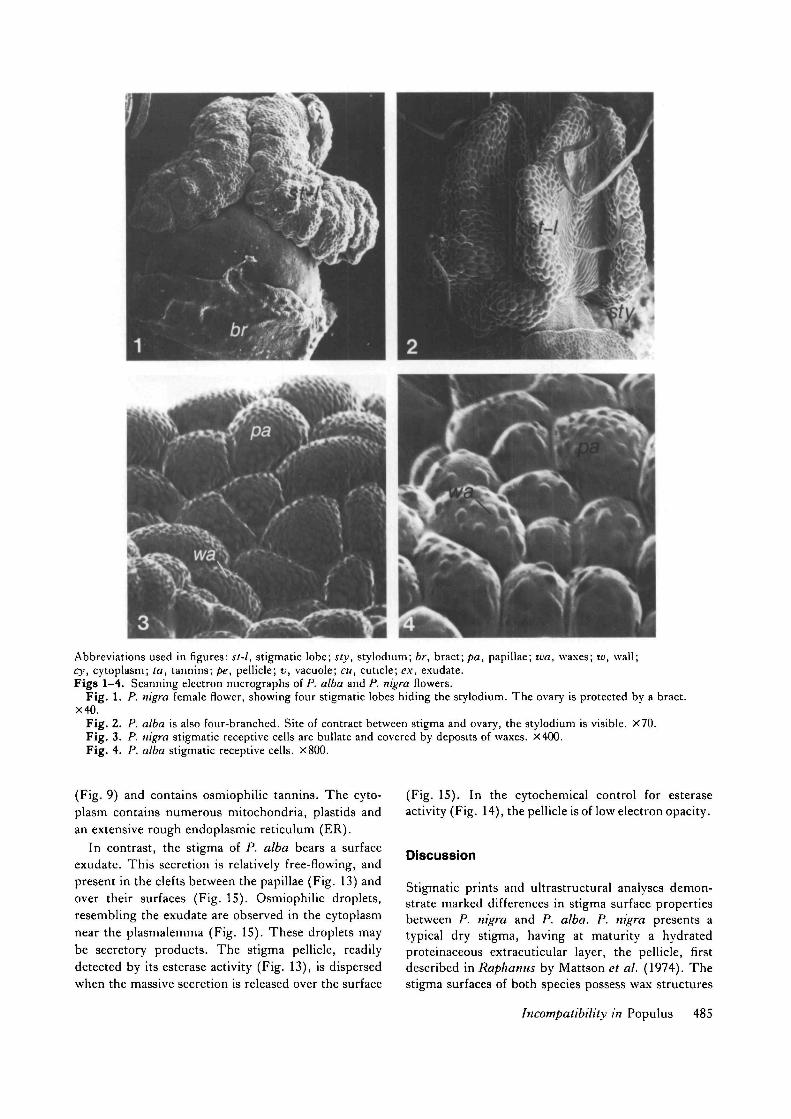
Abbreviations used in figures: st-l, stigmatic lobe; sty, stylodium; br, bract; pa, papillae; wa, waxes; w, wall;cy, cytoplasm; la, tannins; pe, pellicle; v, vacuole; cu, cuticle; ex, exudate.Figs 1-4. Scanning electron micrographs of P. alba and P. nigra flowers.
Fig. 1. P. nigra female flower, showing four stigmatic lobes hiding the stylodium. The ovary is protected by a bract.X40.
Fig. 2. P. alba is also four-branched. Site of contract between stigma and ovary, the stylodium is visible. X70.Fig. 3. P. nigra stigmatic receptive cells are bullate and covered by deposits of waxes. X400.Fig. 4. P. alba stigmatic receptive cells. X800.
(Fig. 9) and contains osmiophilic tannins. The cyto-plasm contains numerous mitochondria, plastids andan extensive rough endoplasmic reticulum (ER).
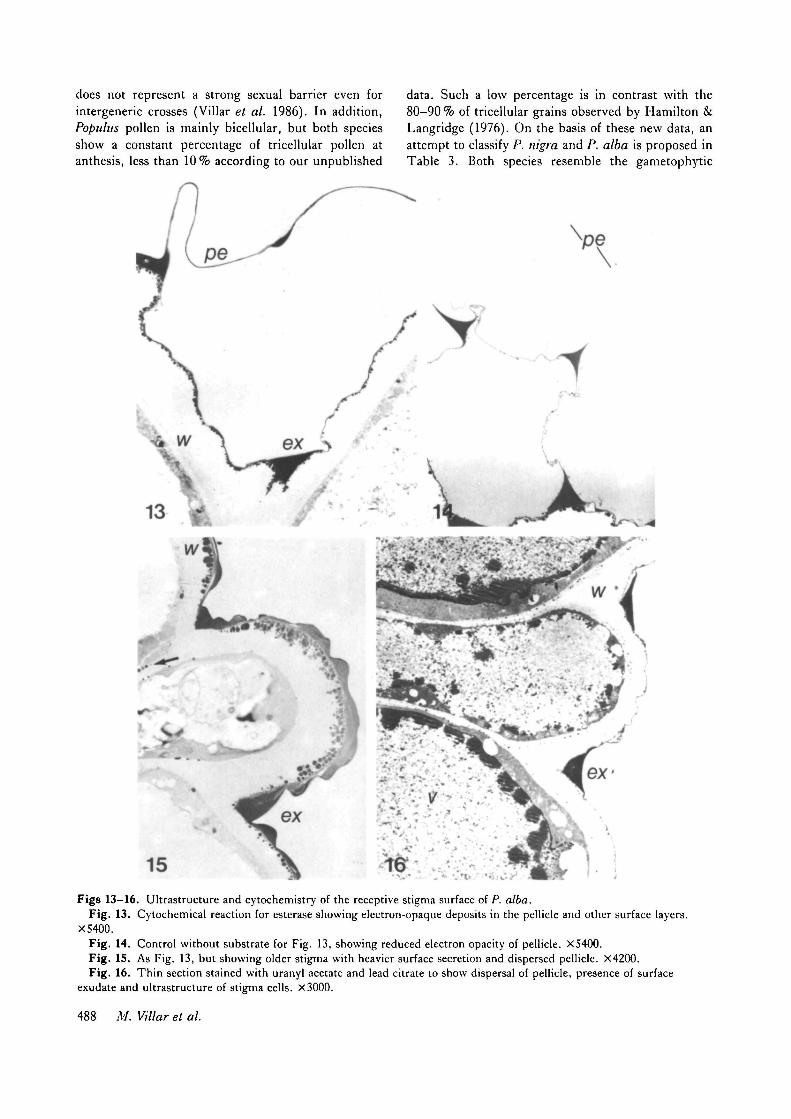
In contrast, the stigma of P. alba bears a surfaceexudate. This secretion is relatively free-flowing, andpresent in the clefts between the papillae (Fig. 13) andover their surfaces (Fig. 15). Osmiophilic droplets,resembling the exudate are observed in the cytoplasmnear the plasmalemma (Fig. 15). These droplets maybe secretory products. The stigma pellicle, readilydetected by its esterase activity (Fig. 13), is dispersedwhen the massive secretion is released over the surface
(Fig. 15). In the cytochemical control for esteraseactivity (Fig. 14), the pellicle is of low electron opacity.
Discussion
Stigmatic prints and ultrastructural analyses demon-strate marked differences in stigma surface propertiesbetween P. nigra and P. alba. P. nigra presents atypical dry stigma, having at maturity a hydratedproteinaceous extracuticular layer, the pellicle, firstdescribed in Raphanus by Mattson et al. (1974). Thestigma surfaces of both species possess wax structures
Incompatibility in Populus 485

Table 3. Characteristics of selected self- and interspecific incompatibility systems for comparison with data
presented in this paper concerning interspecific incompatibility in Populus
Species
Gamctophytic self-incompatible systemsLycopersicon penivianumPmnus avium
Sporophytic self-incompatible systemsBrassica oleraceaRaphanus sativus
Interspecific incompatible systemsP. albaP. nigraRJiododendmn 3pp.
Stigmatype
WetWet
DryDry
WetDryWet
Arrest site
StyleStyle
Stigma surfaceStigma surface
StyleStyleStigma, style or ovary
Pollen type
BicellularBicellular
TricellularTricellular
±Bicellular±BicellularBicellular
Reference
Dumas el al. (1978)Rait el al. (1983)
Knox el al. (1975); Gaude & Dumas (1986)Dickinson & Lewis (1973)
Gaget et al. (1984); this paperGaget el al. (1984); this paperWilliams el al. (1982)
s; • * . -
• • • .
* ;
•- • • •••.;.•.• • • : . ' • • • • • • • : •
• • •
* . . ; . . • •
Figs 5—8. Cytochemistry of stigma prints.
Fig. 5. SEM of P. alba stigmatic prints. X800.Fig. 6. P. alba stigmatic prints stained with Coomassie Blue. X1400.Fig. 7. P. alba stigmatic prints stained for esterase activity. X240.Fig. 8. Optical micrograph of P. nigra stigmatic pnnts stained with Sudan Black B. X1400.
486 M. Villar et al.

similar to the waxy substances observed on Brassicastigma papillae (Roggen, 1972) and detected in P.deltoides stigmas by Hamilton (1976). On the otherhand, the presence of a surface exudate on P. albareceptive cells, demonstrates that this species possessesa stigma of the wet-type, as defined by Heslop-Harrison & Shivanna (1977). The transport of se-cretory products through cytoplasm and cell wallto the stigmatic surface, suggest the occurrence ofeccrine secretion in P. alba. This secretion resemblesthe lipophilic stigmatic secretion of Lycopersicum
peruvianum (Dumas et al. 1978). Thus Populusappears as a heterogeneous genus in terms of stigmatype. Such striking differences have also been ob-served in the two morphs (pin and thrum) of LJnumgrandiflorum (Ghosh & Shivanna, 1980) and Primulaobconica (Schou, 1984).
Populus has been considered to have a surface(sporophytic) type of interspecific incompatibility sys-tem (Hamilton, 1976), but recently Gagetef al. (1984)demonstrated that pollen tube arrest in P. nigra X P.alba occurs in the style, and that the stigmatic surface
12Figs 9-12. Ultrastructure of the receptive stigma surface of P. nigra.
Fig. 9. Presence of bilayered cell wall, cuticle and pellicle, with mitochondria-rich peripheral cytoplasm. Uranylacetate/lead citrate staining. X10 000.
Fig. 10. Lanthanum nitrate as probe of surface, showing deposits in pellicle and cell wall. X27 500.Figs 11, 12. Cationized ferritin as probe of surface, showing deposits in the pellicle. Fig. 11, X52 500; Fig. 12, X39000.
Incompatibility in Populus 487
does not represent a strong sexual barrier even forintergeneric crosses (Villar et al. 1986). In addition,Populus pollen is mainly bicellular, but both speciesshow a constant percentage of tricellular pollen atanthesis, less than 10% according to our unpublished
data. Such a low percentage is in contrast with the80-90% of tricellular grains observed by Hamilton &Langridge (1976). On the basis of these new data, anattempt to classify P. nigra and P. alba is proposed inTable 3. Both species resemble the gametophytic
\ pe
15Figs 13-16. Ultrastmcture and cytochemistry of the receptive stigma surface of P. alba.
Fig. 13. Cytochemical reaction for esterase showing electron-opaque deposits in the pellicle and other surface layers.X5400.
Fig. 14. Control without substrate for Fig. 13, showing reduced electron opacity of pellicle. X5400.Fig. 15. As Fig. 13, but showing older stigma with heavier surface secretion and dispersed pellicle. X4200.Fig. 16. Thin section stained with uranyl acetate and lead citrate to show dispersal of pellicle, presence of surface
exudate and ultrastructure of stigma cells. X3000.
488 M. Villar et al.

incompatibility system (pollen tube arrest in the style),even though P. nigra possess a stigma of the dry-type,and bicellular pollen grains. These differences inclassification of Populus species in terms of the classicalincompatibility systems may provide an explanation forinterspecific incompatibility. Moreover, the significantdifferences between the stigmas of these two speciescould explain the unilateral hybridization phenomenareported in Populus (Ronald, 1982). In the literature,(see Willing & Pryor, 1976) only hybrids between a P.alba female partner and P. nigra have been reported.The reciprocal crosses have not succeeded. A similarsituation exists for P. deltoides. Natural hybrids be-tween P. deltoides X P. nigra produced P. eurameri-cana but P. nigra X P. deltoides has never beenobtained. Interspecific incompatibility is stronglyexpressed. Even breeding techniques such as theMentor effect have failed to overcome this incompati-bility in crosses between P. nigra and P. alba (M.Gaget et ai, unpublished data).
In this paper, we have demonstrated significantdifferences in the nature of the receptive stigma surfaceof these two species. The read-out system for pollentubes may differ in the two species, thus accounting forpollen tube failure in the style. Alternatively, thespecific pollen tube signal may interact with stylargene-products, that act to suppress foreign pollen tubegrowth in the style, but not self species tubes. Thisstudy presents the cytochemical features of P. nigraand P. alba stigmas, sites of pollen-pistil interactions.It represents a step towards the understanding ofinterspecific incompatibility in Populus. Biochemicalinvestigations are in progress to complement thiscytochemical study.
References
ASHFORD, A. E. & KNOX, R. B. (1980). Characteristics ofpollen diffusates and pollen wall cytochemistry inpoplars. J. CellSci. 44, 1-17.
BRONNER, R. (1975). Simultaneous demonstration of lipidsand starch in plant tissues. Slain Technol. 50, 1-4.
DICKINSON, H. G. & LEWIS, D. (1973). Cytochemical andultrastructural differences between intraspecificcompatible and incompatible pollinations in Raphanus.Proc. R. Soc. Land. B, 183, 21-38.
DUMAS, C. & LECOCQ, M. (1975). Mise en evidence etobservation de la secretion stigmatique au microscopeelectronique a balayage, a l'aide d'une technique adapteeau freeze-drying. C.r. hebd. Seanc. Acad. Sci., Paris 280,837-840.
DUMAS, C , ROUGIER, M., ZANDONELLA, P., CIAMPOLINI,
F., CRESTI, M. & PACINI, E. (1978). The secretorystigma in Lycopersicon peruvianum Mill. Ontogenesisand glandular activity. Protoplasma 96, 173-187.
FISHER, D. B. (1968). Protein staining of ribboned eponsections for light microscopy. Histochemie 16, 92-96.
GAGET, M., SAID, C , DUMAS, C. & KNOX, R. B. (1984).
Pollen-pistil interactions in interspecific crosses ofPopulus (Section Aigeiros and Leuce): Pollen adhesion,hydration, and callose responses, J. Cell Sci. 72,173-184.
GAUDE, T. & DUMAS, C. (1986). Organization of stigmasurface components in Brassica: a cytochemical study. J.Cell Sci. 82, 203-216.
GHOSH, S. & SHIVANNA, K. R. (1980). Pollen-pistil
interaction in LJlium grandiflorum: scanning electronmicroscopic observations and proteins of the stigmasurface. Planta 149, 257-261.
HAMILTON, D. (1976). Intersectional incompatibility inPopulus, Ph.D. thesis, Australian National University,Canberra.
HAMILTON, D. & LANGRIDGE, P. (1976). Trinucleate pollenin the Genus Populus. Experientia 32, 467-468.
HESLOP-HARRISON, Y. & SHIVANNA, K. R. (1977). The
receptive surface of the angiosperm stigma. Ann. Bot. 41,1233-1258.
JENSEN, W. A. (1962). Botanical Histochemistry. SanFrancisco, London: W. H. Freeman and Co.
KNOX, R. B. (1984). Pollen-pistil interactions. In CellularInteractions. Encycl. PI. Physiol. (ed. J. Heslop-Harrison& H. F. Linskens), vol. 17, pp. 508-608. Berlin:Springer.
KNOX, R. B., HESLOP-HARRISON, J. & HESLOP-HARRISON,
Y. (1975). Pollen-wall proteins: localization andcharacterization of gametophytic and sporophyticfractions. In Biology of the Male Gamete (ed. J. G.Duckett & P. Racey), pp. 177-187. London: AcademicPress.
KNOX, R. B., WILLING, R. R. & ASHFORD, A. E. (\972a).
Role of pollen wall proteins as recognition substances ininterspecific incompatibility in poplars. Xature, Loud.237, 381-383.
KNOX, R. B., WILLING, R. R. & PRYOR, L. D. (19726).
Interspecific hybridization in poplars using recognitionpollen. Silvae Genet. 21, 65-69.
MATTSON, O., KNOX, R. B., HESLOP-HARRISON, J. &
HESLOP-HARRISON, Y. (1974). Protein pellicle ofstigmatic papillae as a probable recognition site inincompatibility reactions. Nature, Land. 247, 298-300.
RAFF, J. W., PETrnr, J. M. & KNOX, R. B. (1983).
Cytochemistry of pollen tube growth in stigma and styleof Prunus avium. Phytomorphology, 1981, 31, 214—237.
REYNOLDS, E. S. (1963). The use of lead citrate at highpH as an electron opaque stain in electron microscopy.J. Cell Biol. 17, 208-212.
ROGGEN, H. P. J. R. (1972). Scanning electronmicroscopical observations on compatible andincompatible pollen-stigma interactions in Brassica.Euphytica 21, 1-10.
RONALD, W. G. (1982). Intersectional hybridization ofPopulus sections, Leuce-Aigeiros and Leuce-Tacamahaca.Silvae Genet. 31, 94-99.
SAID, C , ZANDONELLA, P., GAUDE, T. & DUMAS, C.
(1985). Cytochemistry of the stigma surface componentsusing the print technique. In Sexual Reproduction in
Incompatibility in Populus 489

Seed Plants, Ferns and Mosses. Proc. 8th Int. Symp. on
Sexual Reproduction in Seed Plants, Ferns and Mosses(ed. M. T. M. Willemse & J. L. van Went), p. 83.
Wageningen: Pudoc.
SCHOU, O. (1984). The dry and wet stigmas of Primula
obconica: Ultrastructural and cytochemical dimorphisms.Protoplasma 121, 99-113.
STETTLER, R. F. (1968). Irradiated mentor pollen: its usein remote hybridization of black cottonwood. Nature,
Land. 219, 746-747.
STETTLER, R. F. & GURIES, R. P. (1976). The mentor
pollen phenomenon in black cottonwood. Can.J. Bot.
54, 820-830.
STETTLER, R. F., KOSTER, R. & STEENACKERS, V. (1980).
Interspecific crossability studies in poplar. Theor. Appl.Genet. 58, 273-282.
TRUMP, B. F., SMUCKLER, E. A. & BENDITT, E. P. (1961).
A method for staining Epoxy sections for lightmicroscopy. J . Ultrastruct. Res. 5, 343-348.
VILLAR, M., GAGET, M. & DUMAS, C. (1986). Sexual
reproduction biology in Populus, compatibility andincompatibility. In Biotechnology and Ecology of Pollen(ed. D. L. Mulcahy, G. B. Mulcahy & E. Ottaviano),pp. 514—517. Berlin: Springer-Verlag.
WILLIAMS, E. G., KNOX, R. B. & ROUSE, J. L. (1982).
Pollination sub-systems distinguished by pollen tubearrest after incompatible interspecific crosses inRhododendron. J. Cell Sci. 53, 255-277.
WILLING, R. R. & PRYOR, L. D. (1976). Interspecifichybridization in popular. Theor. appl. Genet. 47,141-151.
{Received 11 April 1986 -Accepted, in revised form,30 January 1987)
490 M. Villar et al.


















