Immunological Mechanisms Affecting Angiogenesis and Their Relation to Porcine Pregnancy Success
20
Immunological Investigations, 37:611–629, 2008 Copyright © Informa Healthcare USA, Inc. ISSN: 0882-0139 print / 1532-4311 online DOI: 10.1080/08820130802191540 LIMM 0882-0139 1532-4311 Immunological Investigations, Vol. 37, No. 5-6, June 2008: pp. 1–30 Immunological Investigations Immunological Mechanisms Affecting Angiogenesis and Their Relation to Porcine Pregnancy Success Immune Mechanisms at the Maternal-Fetal Interface N. F. Linton et al. Nicola F. Linton, 1 Jocelyn M. Wessels, 1 Sonya A. Cnossen, 1 B. Anne Croy, 1,2 and Chandrakant Tayade 1 1 Department of Biomedical Sciences, University of Guelph, Guelph, Ontario, Canada 2 Department of Anatomy and Cell Biology, Queen’s University, Kingston, Ontario, Canada Prenatal mortality due to loss of lymphocyte-promoted endometrial angiogenesis is being investigated as a major cause of litter reductions during pregnancy in pigs. This review discusses immune mechanisms influencing porcine endometrial angiogenesis as well as additional signalling molecules that may play important roles in the compro- mise of peri-implantation and mid-gestation fetal pig survival. These include dendritic cells, signalling molecules such as toll-like receptors, chemokines and ficolins. Together these cells and molecules regulate immune responses and, ideally, protect the mother and prevent immune-based conceptus losses. Dendritic cells were recently shown to be angiogenic. Their tolerogenic role at the maternal-fetal interface coupled with the ability to secrete and respond to angiogenic factors suggests that dendritic cells are the key coordinators of angiogenesis at the porcine maternal-fetal interface. Chemokines coordinate the localization of immune effector and endothelial cells. The balance between pro-angiogenic and anti-angiogenic chemokines is addressed in rela- tion to conceptus viability. Ficolins, components of the lectin-mediated complement activation pathway, are used for self/non-self recognition. Together, these components of the immune system could regulate lymphocyte- and non-lymphocyte-promoted endometrial angiogenesis to determine conceptus survival. Keywords Dendritic cells, Chemokines, Ficolins, Pregnancy loss, Toll-like receptors, Uterine lymphocytes. Address correspondence to Chandrakant Tayade, Department of Biomedical Sciences, University of Guelph, Guelph, ON N1G 2W1 Canada; E-mail: [email protected] Immunol Invest Downloaded from informahealthcare.com by University of Calgary on 10/03/13 For personal use only.
-
Upload
chandrakant -
Category
Documents
-
view
212 -
download
0
Transcript of Immunological Mechanisms Affecting Angiogenesis and Their Relation to Porcine Pregnancy Success

Immunological Investigations, 37:611–629, 2008Copyright © Informa Healthcare USA, Inc.ISSN: 0882-0139 print / 1532-4311 onlineDOI: 10.1080/08820130802191540
LIMM0882-01391532-4311Immunological Investigations, Vol. 37, No. 5-6, June 2008: pp. 1–30Immunological InvestigationsImmunological Mechanisms Affecting Angiogenesis and Their Relation to Porcine Pregnancy SuccessImmune Mechanisms at the Maternal-Fetal InterfaceN. F. Linton et al.
Nicola F. Linton,1 Jocelyn M. Wessels,1 Sonya A. Cnossen,1 B. Anne Croy,1,2 and Chandrakant Tayade1
1Department of Biomedical Sciences, University of Guelph, Guelph, Ontario, Canada2Department of Anatomy and Cell Biology, Queen’s University, Kingston, Ontario, Canada
Prenatal mortality due to loss of lymphocyte-promoted endometrial angiogenesis isbeing investigated as a major cause of litter reductions during pregnancy in pigs. Thisreview discusses immune mechanisms influencing porcine endometrial angiogenesis aswell as additional signalling molecules that may play important roles in the compro-mise of peri-implantation and mid-gestation fetal pig survival. These include dendriticcells, signalling molecules such as toll-like receptors, chemokines and ficolins.Together these cells and molecules regulate immune responses and, ideally, protect themother and prevent immune-based conceptus losses. Dendritic cells were recentlyshown to be angiogenic. Their tolerogenic role at the maternal-fetal interface coupledwith the ability to secrete and respond to angiogenic factors suggests that dendriticcells are the key coordinators of angiogenesis at the porcine maternal-fetal interface.Chemokines coordinate the localization of immune effector and endothelial cells. Thebalance between pro-angiogenic and anti-angiogenic chemokines is addressed in rela-tion to conceptus viability. Ficolins, components of the lectin-mediated complementactivation pathway, are used for self/non-self recognition. Together, these componentsof the immune system could regulate lymphocyte- and non-lymphocyte-promotedendometrial angiogenesis to determine conceptus survival.
Keywords Dendritic cells, Chemokines, Ficolins, Pregnancy loss, Toll-like receptors,Uterine lymphocytes.
Address correspondence to Chandrakant Tayade, Department of Biomedical Sciences,University of Guelph, Guelph, ON N1G 2W1 Canada; E-mail: [email protected]
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

612 N. F. Linton et al.
INTRODUCTION ON PRENATAL MORTALITY IN MEAT PIGS
Prenatal mortality is a major economic concern for commercial swine produc-ers in North America. Approximately 20 to 30% of conceptuses die betweengestation days (gd) 12–30 (peri-implantation/attachment) and an additional10–15% are lost by mid-gestation (Pope, 1994). In commercial herds, littersizes would average 15 to 17 rather than the observed 10–11, if all fertilizedeggs developed to live born piglets (USDA, 2001). Maximum embryonicmortality occurs during ~gd11–15, the stage of blastocyst attachment, whenporcine embryos transform into long, thin, filamentous structures and startsecreting estrogen to promote endometrial attachment (Geisert et al., 1982).
Blastocyst-derived estrogen creates a hostile, inflammatory milieu formore slowly elongating littermates. Chinese Meishan pigs are a breed thatfarrows 4–5 more piglets per litter than European-derived North Americanpig breeds from a similar number of ovulated eggs. In comparison with NorthAmerican breeds, Chinese Meishan pre-implantation trophoectodermal cells,the cells that differentiate into the fetal part of the placenta, have lowermitotic rates, less elongation and less estrogen production. This results insmaller but more efficient placentae. These smaller placentae also have moreblood vessels in the outer placental membranes to meet the demands of therapidly growing, later gestation fetuses (Wilson et al., 1999).
Porcine blastocyst development depends on endometrial and conceptus-derived growth factors. Early trophoblast elongation uses histotrophic nutritionprovided by uterine gland secretions (Spencer and Bazer, 2004). Post-attachmentconceptus growth and development depend upon endometrial-placental inter-actions with robust increases in maternal blood supply required to sustainfetal nutrition. Major endometrial vascular remodelling and growth occur,particularly in the subepithelial plexus.
For more than two decades, genetic selection pressures have been appliedwithout success to commercial swine for traits like placental efficiency,uterine capacity or ovulation rate. Recently, from the largest genetic studiesconducted to date by the United States Department of Agriculture, the conclu-sion was that 11 generations of selection for uterine capacity and ovulationrate made no significant improvement in porcine litter size (Freking et al.,2007). Studies on nutrition and vitamin supplements or improved balance ofdiets also failed to boost litter sizes born (Foxcroft, 1997).
Our studies suggest that deficient endometrial angiogenesis causes earlyand mid-pregnancy conceptus loss. We used laser capture microdissection(LCM) and quantitative RNA studies to compare angiogenic gene expressionin endometrium, endometrial lymphocytes, endometrial endothelial cells andtrophoblasts collected from individual healthy or arresting littermate attach-ment sites. Loss of maternal lymphocyte expression of angiogenic moleculesand of tissue oxygen sensing was linked with porcine fetal growth arrest and
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Immune Mechanisms at the Maternal-Fetal Interface 613
subsequent death (Tayade et al., 2006, 2007a). In this review, we examinepotential regulatory pathways for angiogenic gene expression in endometriallymphocytes during pregnancy that may be altered in pregnancy failure. Ourexperimental approach is outlined in Figure 1.
IMPLANTATION SITE ANGIOGENESIS
Endometrial angiogenesis is essential during placental development to meet themetabolic demands of conceptuses. Angiogenesis is promoted by a variety ofgrowth factors, including but not limited to basic fibroblast growth factor(bFGF), vascular endothelial growth factor (VEGF), placental growth factor(PlGF) and tumour necrosis factor alpha (TNF-α) (Zygmunt et al., 2003). VEGFsignals through two of its receptors, VEGFRI and VEGFRII but also binds otherreceptors such as soluble fms-like tyrosine kinase receptor 1 (sFlt-1), a splicevariant of VEGFRI, which prevents VEGF signalling (Zygmunt et al., 2003).
Figure 1: A diagram of our protocol for tissue collection from pregnant porcine uteri for down-stream applications. RNA is extracted from whole tissue and reverse transcribed into cDNA.Relative real time PCR is performed for each target gene, using β-actin as the housekeepinggene. Porcine endometrial paraffin and frozen sections are prepared for staining using cross-reactive anti-human antibodies for cells and proteins of interest. A rapid staining protocol wasdevised when staining frozen sections to maintain RNA stability for LCM. After LCM wasperformed, RNA from captured cells was extracted. cDNA was synthesised, followed byamplification, due to low numbers of captured cells (Tayade et al., 2006; Linton et al., 2008manuscript in preparation). Relative real time PCR was performed as above. Whole tissue wasalso used to extract protein, which was subsequently analysed using the chemiluminescentprotein array by Searchlight (Thermofisher Scientific).
Collection of Endometrial and Trophoblast Samples (gd20, 50 and non-pregnantgilts)
RNA extractionfrom endometrialand trophoblastbiopsies
Quantitative realtime PCR forgenes of interest
Frozenendometrialsections
Relative quantification of target genes with β-actin asa housekeeper
Proteinextraction
Quantificationusing
Searchlight®protein array
(ThermofisherScientific)
cDNAsynthesis
LCM in frozen sections
RNA extraction, cDNAsynthesis and amplification
IHCusingDC-SIGN
RapidH&EStain
Captureof DCs
Capture of endometrial lymphocytes
and endothelium
Histology andIHC in parafiinembeddedsections
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

614 N. F. Linton et al.
Many studies of both humans and pigs have examined expression ofangiogenesis-related factors in healthy compared to complicated pregnancies.Two complications of pregnancy in humans, intrauterine growth restriction(IUGR) and pre-eclampsia (PE), have altered placental perfusion and dysregu-lation of angiogenic genes (Huppertz et al., 2007; McCarthy et al., 2007;Jarvenpaa et al., 2007). IUGR, characterized by a decrease in the rate of fetalgrowth resulting in an increased risk of perinatal mortality (McCarthy et al.,2007), is an outcome similar to that seen in arresting or newborn runt pigs.PE, a potentially fatal clinical syndrome in the last trimester of humanpregnancy, is caused by placental-induced hypertension that is relieved byplacental delivery (Kopcow and Karumanchi, 2007).
As in human IUGR, arresting pig conceptuses are characterized bydecreased growth and decreased placental vasculature (Tayade et al., 2006,2007a). We hypothesized that endometrial lymphocytes, specifically recruitedto the porcine maternal-fetal interface at the time of blastocyst attachment,are angiogenic and control conceptus development. In pigs, where no deciduaforms because trophoblast is not invasive, endometrial lymphocytes arerecruited to implantation sites by conceptus-derived signals that remainpoorly characterized (Engelhardt et al., 2002b; Croy et al., 2006; Tayade2007b). To understand the dynamic features of angiogenic and cytokine geneexpression at the porcine maternal-fetal interface, mRNA comparisons to vir-gin endometrium were made at gd 20 and 50 for endometrial and trophoblastbiopsies and for ˜500 endometrial lymphocytes or endometrial endothelialcells captured from frozen biopsy sections using laser capture microdissection(LCM). Profiles were derived for the relative expression of angiogenic andcytokine genes from quantitative real time PCR analyses (Figure 1).
VEGF expression was elevated in pregnant porcine endometrium at sitesassociated with healthy conceptus implantation compared to sites associatedwith arresting conceptuses. Pregnancy elevated VEGF expression in lympho-cytes captured from endometrium associated with a healthy conceptus.Lymphocytes from arresting sites either did not transcribe VEGF or had muchlower expression than healthy sites in the same uterus. Lymphocyte VEGFexpression exceeded that of total endometrium, endothelium or trophoblasts,indicating that lymphocytes are a dominant source of porcine implantationsite VEGF, at least to midpregnancy.
Hypoxia inducible factor (HIF)-1α, is a transcription factor expressed whentissue oxygen levels fall or there is a hypoxic environment. Its expressionsignals a requirement for angiogenesis and it is a key regulator of VEGF tran-scription (Fukushima et al., 2008). More endometrial HIF-1α mRNA expressionoccurs in healthy attachment sites than in age-matched arresting sites. HIF-1αmRNA expression in lymphocytes dissected from endometrium associated withhealthy conceptuses was higher than in lymphocytes dissected from virginendometrium or endometrium associated with arresting conceptuses.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Immune Mechanisms at the Maternal-Fetal Interface 615
Endometrium associated with arresting attachment sites had more pro-inflammatory cytokine gene expression (interferon γ (IFN-γ), TNF-α andinterleukin (IL)-1β) than virgin endometrium or endometrium associated withhealthy conceptuses (Tayade et al., 2006, 2007a). These data suggest thatimmune cells contribute to conceptus success or failure by influencing endome-trial angiogenesis. As discussed in the following sections, other elements of theimmune system must participate in gestational endometrial angiogenesis, byregulation of lymphocyte-promoted angiogenesis. They may also use more directmechanisms and we are evaluating these in our porcine pregnancy model.
ROLES FOR DENDRITIC CELLS AND TOLL-LIKE RECEPTORS IN ENDOMETRIAL ANGIOGENESIS DURING PREGNANCY
Dendritic Cells and the Immune SystemDendritic cells (DC) play central regulatory roles for both innate and
adaptive immunity and have pleiotropic interactions with other immune celltypes (Adams et al., 2005). Bidirectional communication between DCs andinnate immune cells, such as NK cells, influences overall innate immunity.For example, NK cells enhance the ability of DCs to produce IL-12 in responseto antigen (Ag) from Toxoplasma gondii (Guan et al., 2007), yet DCs prime NKcell responses by trans-presenting IL-15, a cytokine important for NK cellviability and function (Lucas et al., 2007; Zhang et al., in press). DCs alsopresent Ag to T cells, promoting tolerance or adaptive immunity.
After stimulation, DCs must mature and express major histocompatibiltycomplex II (MHCII) to enhance their Ag presentation function (Stingl andBergstresser, 1995). DCs use DC-SIGN (Dendritic Cell-Specific IntracellularCell Adhesion Molecule (ICAM)-Grabbing Nonintegrin) and ICAM-3 to formsynapses with lymphocytes and trigger cytokine production in these“educated” cells (Hartgers et al., 2000; Langenkamp et al., 2000). We haveidentified putative porcine endometrial DCs using anti-human DC-SIGNantibody (Figure 2). These cells are infrequent and we are investigating theirrelationships to other immune cells and their angiogenic potential at theporcine maternal-fetal interface. Others have shown the presence of majorhistocompatibility complex II positive (MHCII+) macrophages and dendritic-like cells in porcine gestational endometrium (Bischof et al., 1995).
Dendritic Cells and AngiogenesisThe pivotal role of DCs in immune responses led to numerous studies that
initially established their angiogenic potential in tumour development(reviewed in Noonan et al., 2007). These studies also established that angiogenicmolecules influence DC maturation. In a murine model of endometriosis,
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.
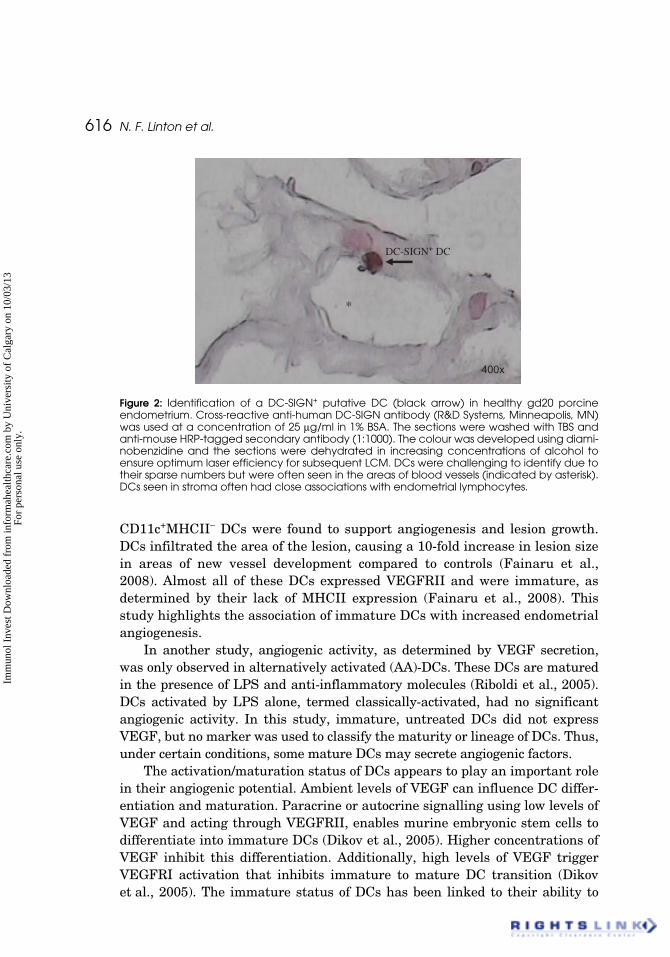
616 N. F. Linton et al.
CD11c+MHCII− DCs were found to support angiogenesis and lesion growth.DCs infiltrated the area of the lesion, causing a 10-fold increase in lesion sizein areas of new vessel development compared to controls (Fainaru et al.,2008). Almost all of these DCs expressed VEGFRII and were immature, asdetermined by their lack of MHCII expression (Fainaru et al., 2008). Thisstudy highlights the association of immature DCs with increased endometrialangiogenesis.
In another study, angiogenic activity, as determined by VEGF secretion,was only observed in alternatively activated (AA)-DCs. These DCs are maturedin the presence of LPS and anti-inflammatory molecules (Riboldi et al., 2005).DCs activated by LPS alone, termed classically-activated, had no significantangiogenic activity. In this study, immature, untreated DCs did not expressVEGF, but no marker was used to classify the maturity or lineage of DCs. Thus,under certain conditions, some mature DCs may secrete angiogenic factors.
The activation/maturation status of DCs appears to play an important rolein their angiogenic potential. Ambient levels of VEGF can influence DC differ-entiation and maturation. Paracrine or autocrine signalling using low levels ofVEGF and acting through VEGFRII, enables murine embryonic stem cells todifferentiate into immature DCs (Dikov et al., 2005). Higher concentrations ofVEGF inhibit this differentiation. Additionally, high levels of VEGF triggerVEGFRI activation that inhibits immature to mature DC transition (Dikovet al., 2005). The immature status of DCs has been linked to their ability to
Figure 2: Identification of a DC-SIGN+ putative DC (black arrow) in healthy gd20 porcineendometrium. Cross-reactive anti-human DC-SIGN antibody (R&D Systems, Minneapolis, MN)was used at a concentration of 25 μg/ml in 1% BSA. The sections were washed with TBS andanti-mouse HRP-tagged secondary antibody (1:1000). The colour was developed using diami-nobenzidine and the sections were dehydrated in increasing concentrations of alcohol toensure optimum laser efficiency for subsequent LCM. DCs were challenging to identify due totheir sparse numbers but were often seen in the areas of blood vessels (indicated by asterisk).DCs seen in stroma often had close associations with endometrial lymphocytes.
400x
DC-SIGN+ DC
*
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Immune Mechanisms at the Maternal-Fetal Interface 617
induce tolerance to a conceptus (Blois et al., 2004). The uterine microenviron-ment of pregnancy may therefore prevent DC maturation, direct their func-tions towards angiogenesis and provide immune protection for the conceptus.
Dendritic Cells and PregnancyStudies in humans identified a unique, pregnancy-specific population of
immature DC-SIGN+ DCs in decidua and showed that this populationremains stable to term (Kammerer et al., 2003 and this volume). These cellspreferentially surround spiral arteries, but are also found throughout thedecidua (Kammerer et al., 2003). Human decidual DC-SIGN+ DCs appear tointeract with CD56+ lymphocytes, via DC-SIGN and ICAM-3 (Kammereret al., 2003). CD56 marks uNK cells, cells known to secrete VEGF (Engelhardtet al., 2002b; Li et al., 2001). There are no reports indicating that uNK cellsexpress either VEGFRI or VEGFRII. Since NK cells do not express eitherVEGFR (Li et al., 2001), we hypothesize that endometrial DC-SIGN+ DCs andNK cells communicate via VEGF and its receptors, in addition to their inter-actions through ICAM-3 and DC-SIGN. This would predict co-localization ofthese cells to perivascular areas at sites of active endothelial cell proliferation(Grunewald et al., 2006). Communication through VEGF and both of itssignalling receptors may allow DCs to respond to and possibly control theangiogenic actions of NK cells.
In our preliminary studies, we found that DC-SIGN+ cells in gd20 and 50endometrium from healthy porcine attachment sites express not only VEGFbut also both VEGFRI and VEGFRII. DCs dissected from arresting conceptusimplantation sites do not have the same levels of expression of VEGF or itsreceptors (Linton et al., 2008, manuscript in preparation).
Toll-Like Receptors in the Immune SystemToll-like receptors (TLRs) are a subset of pattern recognition receptors,
responsible for recognizing a limited set of conserved structures found onviruses and bacteria. They are found on all cells of the immune system, partic-ularly in the innate compartment, and they function as a first line of defenceagainst potentially harmful pathogens (Abrahams and Mor, 2005; Wang et al.,2008, Abrahams, this volume). Immature DCs fulfilling an immune sentinelfunction at mucosal sites express most TLRs depending on the DC subset(Hartgers et al., 2000; Adams et al., 2005). In humans, trophoblast coveringthe peripheral chorionic villi and intermediate trophoblasts express TLR-2and -4 (Holmlund et al., 2002). Abrahams and Mor (2005) have also reportedTLR-2 and -4 expression in villous cytotrophoblast and extravillous tropho-blast in first trimester placenta.
TLR-1 and -6 co-localise with TLR-2 (Abrahams and Mor, 2005). TLRs areupstream mediators of inflammation, a primary response to blastocyst
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

618 N. F. Linton et al.
implantation and to pathogen invasion. The relationship between intrauterineinfections and preterm birth has led to the hypothesis that TLR functionsduring pregnancy can affect its outcome (reviewed in Patni et al., 2007). Thereis less focus on the potential for TLR recognition patterns displayed byhealthy or failing trophoblasts.
No published studies have addressed TLR expression in pigs. To enablecomparative study of TLR expression in porcine endometrium and trophoblastfrom healthy or arresting attachment sites, TLRs-1 through -10 were screenedin porcine endometrium and trophoblast at both gd20 and gd50. Sequenceswere cloned and submitted to GenBank (Table 1). Based on studies of thehuman maternal-fetal interface, expression of TLR-1, -2, -4 and -6, was exam-ined in the pig uterus. TLRs-1, -4 and -6 but not -2, were detected at sufficientlevels for further study and their relative expression patterns are understudy. Preliminary analysis indicated that these three TLRs were elevated ingd50 trophoblast recovered from an arresting conceptus compared withtrophoblast from its healthy littermate controls. No difference in TLR expres-sion was observed in endometrium from either healthy or arresting sites.Additionally TLR-4 expression in DCs captured from gd50 arrestingendometrium is elevated but statistical analysis has not been completed(Linton et al., 2008, manuscript in preparation).
VEGF expression in macrophages is augmented when TLR-2, -4, -7 and -9are activated by their respective agonists such as LPS (TLR-4 agonist). TLRactivation alone has no effect on VEGF production (Pinhal-Enfield et al.,2003) but, by modulating the function of other immune components, TLRsmay aid in angiogenesis.
CHEMOKINES AND ANGIOGENESIS
Chemokines are chemotactic cytokines secreted by a wide variety of cell types.Their main functions are to control and regulate the migration of motile cell
Table 1: List of porcine partial coding sequences submitted to the NCBI GenBank.
Gene Name Accession Number
Sus scrofa TLR1 EU180625Sus scrofa TLR2 EU180626Sus scrofa TLR3 EU180627Sus scrofa TLR4 EU180628Sus scrofa TLR5 EU180629Sus scrofa TLR6 EU180630Sus scrofa TLR7 EU180631Sus scrofa TLR8 EU180632Sus scrofa TLR9 EU189699Sus scrofa TLR10 EU180633
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Immune Mechanisms at the Maternal-Fetal Interface 619
populations (reviewed in Kelly et al., 2001; reviewed in Charo and Ransohoff,2006; Drake et al., 2002; Huang et al., 2006; McKimmie and Graham, 2006).Chemokines are classified structurally into four groups (CC, CXC, XC, andCX3C), based on the location of conserved cysteine residues (reviewed inCharo and Ransohoff, 2006). Chemoattraction, their main function, beginswhen the chemokine binds to a signalling chemokine receptor.
This interaction initiates a signal transduction cascade that induces thetranscription of genes responsible for cytoskeletal rearrangement, and hencecell movement. Both homeostatic and inflammatory chemokines areabundant at the maternal-fetal interface in humans and mice and have thepotential to act on an array of immune, endothelial and trophoblast celltypes. Differences in chemokine expression between healthy and arrestingporcine attachment sites are likely to be responsible, at least in part, forfetal outcomes.
Inflammatory chemokines, as well as their regulators, decoy receptors(see Borroni et al, this volume), have been hypothesized to be differentiallytranscribed and translated at healthy and arresting porcine attachmentsites (Wessels et al., 2007). These chemokines are thought to act as dangersignals that alert the maternal immune system to mount a localized immuneresponse against the newly developed endometrial vasculature supporting astressed or abnormal conceptus and to attract a different array of leukocytesto arresting conceptus sites (Wessels et al., 2007). Severing maternal angio-genic support for the pregnancy would eventually lead to fetal death(Wessels et al., 2007). In our ongoing studies, we are quantifying severalchemokines and chemokine decoy receptors at the porcine maternal-fetalinterface using real time PCR, and SearchLight chemiluminescent proteinarray.
Chemokines, especially CXC Chemokines, also act as positive and neg-ative mediators of angiogenesis, usually through a chemokine receptormediated pathway (reviewed in Mehrad et al., 2007). The CXC group ofchemokines can be subdivided into two additional subgroups. One groupcontains chemokines with an ELR (glutamic acid, leucine, and arginine)motif (ELR+), and the other group contains chemokines without this motif(ELR-) (Baggiolini et al., 1994). ELR+ chemokines are usually angiogeniceither by inducing the proliferation of endothelial cells, or by attractingendothelial cells via chemotaxis (Koch et al., 1992; Strieter et al., 1992,1995; Maione et al., 1990). On the other hand, ELR- chemokines areusually anti-angiogenic by inhibiting the ability of basic FGF (bFGF) toinduce proliferation of endothelial cells, or by inhibiting the migration ofendothelial cells (Koch et al., 1992; Strieter et al., 1992, 1995; Maione et al.,1990; Presta et al., 1998). Chemokines can positively or negatively affectangiogenesis through a receptor mediated pathway, or a receptor indepen-dent pathway.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

620 N. F. Linton et al.
While most chemokines have only one action on the angiogenic process,the ELR+ chemokine, CXCL8, is pro-angiogenic through the chemokine recep-tor mediated pathway and anti-angiogenic through a receptor independentpathway. The ability of CXCL8 to inhibit angiogenesis is dose dependent(Presta et al., 1998). As heterogeneity exists between blood vessels of differingtissues, calibers, or types, it is conceivable that chemokine effects on angio-genesis are highly variable (McCarthy et al., 1991). Other anti-angiogenicchemokines are ELR+ CXCL1, CXCL3, and ELR- CXCL4, and the CCchemokines CCL2, CCL3, and CCL5 (Presta et al., 1998).
A pro-angiogenic chemokine of great interest is CXCL12 (reviewed inCharo and Ransohoff, 2006), a molecule induced in human endometrium bydecidualization (Hanna et al., 2003; Wu et al., 2005). CXCL12 mediates dose-dependent angiogenesis by induction of endothelial cell proliferation via itsinteraction with CXCR4 on endothelial and vascular smooth muscle cells, andby attracting cells that express VEGFRI (Kollmar et al., 2007; Petit et al.,2007; Zheng et al., 2007; Zhou et al., 2007). Human uNK cells strongly expressCXCR4 and would be expected to co-localize at sites of CXCL12-promotedangiogenesis. CXCL12 through its interaction with CXCR4, activates thePI3K/Akt pathway to promote VEGF production (Kollmar et al., 2007; Lianget al., 2007; Zheng et al., 2007). Interestingly, VEGF acts in synergy withCXCL12 to attract endothelial cells by inducing the upregulation of CXCR4 onvascular endothelial cells (Kryczek et al., 2005).
CXCL12 transcription and translation were quantified at the porcinematernal-fetal interface using real time PCR and the SearchLight chemilumi-nescent protein array (ThermoFisher, Woburn, MA, USA; Figure 1 and 3).Three healthy and arresting littermate pairs from three mothers at bothgd20 and gd50 were used to compare gene transcription between healthstatuses. Porcine specific primers (F-CAGTGTCCCCAGTGTGTCAG, R-ACGATCTGAAGGGCACAGTT) were designed and used for PCR and theproduct was sequenced to ensure primer specificity (GenBank Accessionnumber: EU155483.1). Six healthy and arresting littermate pairs from fivemothers at gd20 and four mothers at gd50 were used to compare proteinexpression between conceptus health status conditions.
Protein was analysed using a cross-reactive human chemiluminescentprotein array. CXCL12 transcription was elevated at healthy attachment sitesin both endometrium and trophoblast and appeared to increase as pregnancyproceeded, although statistical significance was not found between the gd20and 50 time points or between healthy and arresting samples. The pattern ofCXCL12 protein expression seemed to be the exact inverse of its transcriptionpattern and lacked statistical significance. This permits us to conclude thatCXCL12 is stably transcribed and translated at gd20 and 50 in porcineendometrium and trophoblast associated with both healthy and arrestingconceptuses.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.
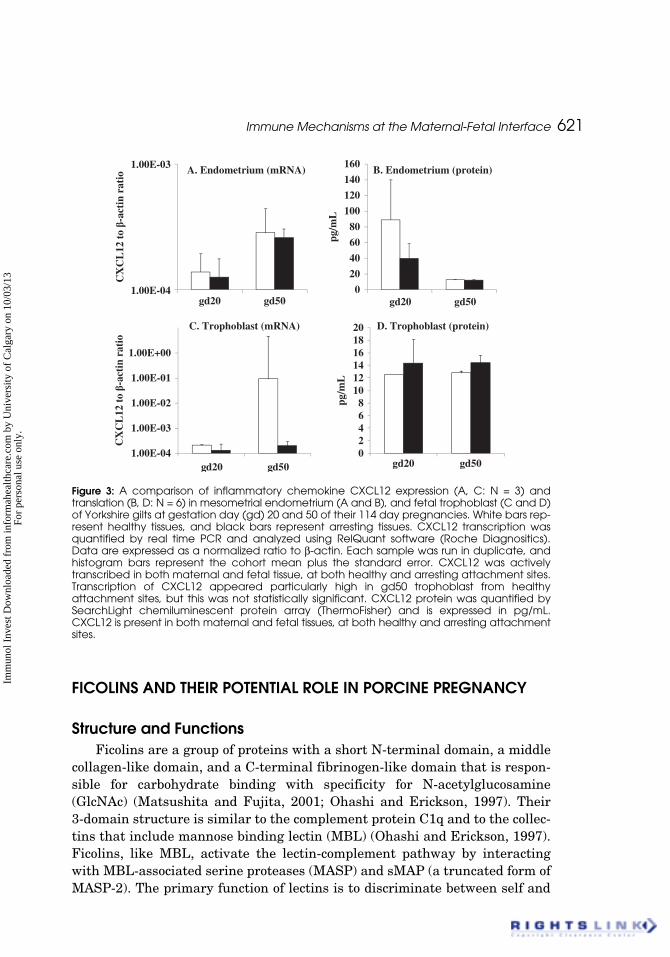
Immune Mechanisms at the Maternal-Fetal Interface 621
FICOLINS AND THEIR POTENTIAL ROLE IN PORCINE PREGNANCY
Structure and FunctionsFicolins are a group of proteins with a short N-terminal domain, a middle
collagen-like domain, and a C-terminal fibrinogen-like domain that is respon-sible for carbohydrate binding with specificity for N-acetylglucosamine(GlcNAc) (Matsushita and Fujita, 2001; Ohashi and Erickson, 1997). Their3-domain structure is similar to the complement protein C1q and to the collec-tins that include mannose binding lectin (MBL) (Ohashi and Erickson, 1997).Ficolins, like MBL, activate the lectin-complement pathway by interactingwith MBL-associated serine proteases (MASP) and sMAP (a truncated form ofMASP-2). The primary function of lectins is to discriminate between self and
Figure 3: A comparison of inflammatory chemokine CXCL12 expression (A, C: N = 3) andtranslation (B, D: N = 6) in mesometrial endometrium (A and B), and fetal trophoblast (C and D)of Yorkshire gilts at gestation day (gd) 20 and 50 of their 114 day pregnancies. White bars rep-resent healthy tissues, and black bars represent arresting tissues. CXCL12 transcription wasquantified by real time PCR and analyzed using RelQuant software (Roche Diagnositics).Data are expressed as a normalized ratio to β-actin. Each sample was run in duplicate, andhistogram bars represent the cohort mean plus the standard error. CXCL12 was activelytranscribed in both maternal and fetal tissue, at both healthy and arresting attachment sites.Transcription of CXCL12 appeared particularly high in gd50 trophoblast from healthyattachment sites, but this was not statistically significant. CXCL12 protein was quantified bySearchLight chemiluminescent protein array (ThermoFisher) and is expressed in pg/mL.CXCL12 is present in both maternal and fetal tissues, at both healthy and arresting attachmentsites.
A. Endometrium (mRNA) B. Endometrium (protein)
C. Trophoblast (mRNA) D. Trophoblast (protein)pg
/mL
pg/m
L
CX
CL
12 t
o β-
acti
n ra
tio
CX
CL
12 t
o β-
acti
n ra
tio
1.00E-04
1.00E-03
0
20
40
60
80
100
120
140
160
gd20 gd50 gd20 gd50
gd20 gd5002468
101214161820
1.00E-04
1.00E-03
1.00E-02
1.00E-01
1.00E+00
gd20 gd50
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

622 N. F. Linton et al.
non-self through initiation of aggregation and opsonization of pathogens andother foreign materials for phagocytosis (Hoffmann et al., 1999). Ficolins alsoactivate production of cytokines by cells of the innate immune system,particularly TNF-α (Wang et al., 2007).
Genetic Differences Between SpeciesFicolins were originally isolated as transforming growth factor beta
(TGFβ) binding proteins from pig uterine extracts (Ichijo et al., 1991) and theFicolin a and b genes (Fcna and Fcnb) were obtained by screening a piguterine cDNA library (Ichijo et al., 1993). More recently, various ficolinisoforms have been identified at cDNA and/or protein levels in vertebratesand invertebrates. The isoforms have different tissue distributions, suggest-ing ficolin functions may also vary between tissues (Matsushita and Fujita,2001). In humans, three types of ficolins have been identified. The corticoster-oid binding protein, termed hucolin (Edgar, 1995), and the elastin bindingprotein, termed EBP-37 (Harumiya et al., 1995), are expressed predominantlyin liver (Matsushita and Fujita, 2001; Ohashi and Erickson, 1997); while theGlcNAc binding lectin, termed P35 (Matsushita et al., 1996), is expressed bylung and monocytes (Matsushita, 1996). Mice have two ficolin genes, ficolin-Aand ficolin-B. Ficolin-A mRNA is highly transcribed in liver and spleen(Fujimori et al., 1998) while ficolin-B is predominantly expressed in bone mar-row and spleen (Ohashi, 1998). Ohashi and Erickson (1998) reported that thetranscription of ficolin a in pigs is high in lung, liver and bone marrow and lowin uterus and that ficolin b mRNA is transcribed at low levels in bone marrowand not at all in the uterus.
Ficolins and PregnancyWang et al. (2007) reported that in human pre-eclampsia, a disease of mid
to late pregnancy, ficolin circulation was depressed while MBL levels wereunchanged. Ficolins were abundant in the pre-eclamptic placenta bound todying (Fas+) regions of syncytiotrophoblast, the outer layer of the placenta.Ficolin binding in these placentae appeared to activate innate immune cytok-ines and chemokines such as TNF-α, which was shown earlier to be increasedat arresting porcine attachment sites at gd50 (Tayade et al., 2007a). Arraydata also indicated that ficolins suppressed interferons and cytokines involvedin adaptive immunity, including IFN-γ and TGF-β. The work by Wang et al.(2007) coupled with our observation that midpregnancy porcine fetal loss isaccompanied by elevated TNF-α and depressed IFN-γ, suggested thatincreased local ficolin expression may be a regulatory pathway elevated in the2nd wave of porcine fetal loss.
To investigate putative roles for ficolin α and β in pregnancy loss, theirtranscription was relatively quantified by real-time PCR in attachment site
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.
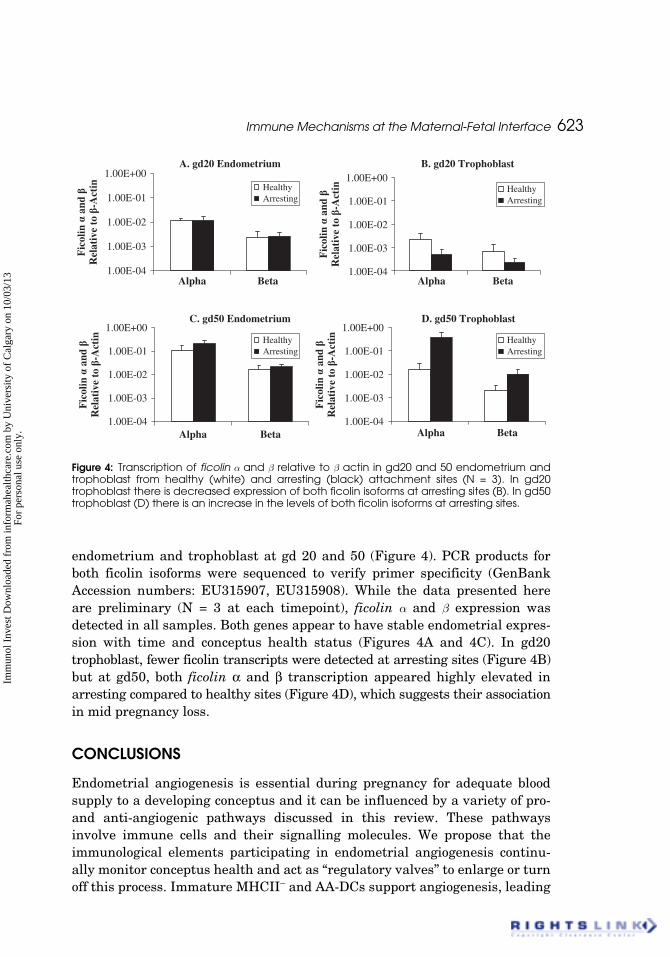
Immune Mechanisms at the Maternal-Fetal Interface 623
endometrium and trophoblast at gd 20 and 50 (Figure 4). PCR products forboth ficolin isoforms were sequenced to verify primer specificity (GenBankAccession numbers: EU315907, EU315908). While the data presented hereare preliminary (N = 3 at each timepoint), ficolin a and b expression wasdetected in all samples. Both genes appear to have stable endometrial expres-sion with time and conceptus health status (Figures 4A and 4C). In gd20trophoblast, fewer ficolin transcripts were detected at arresting sites (Figure 4B)but at gd50, both ficolin α and β transcription appeared highly elevated inarresting compared to healthy sites (Figure 4D), which suggests their associationin mid pregnancy loss.
CONCLUSIONS
Endometrial angiogenesis is essential during pregnancy for adequate bloodsupply to a developing conceptus and it can be influenced by a variety of pro-and anti-angiogenic pathways discussed in this review. These pathwaysinvolve immune cells and their signalling molecules. We propose that theimmunological elements participating in endometrial angiogenesis continu-ally monitor conceptus health and act as “regulatory valves” to enlarge or turnoff this process. Immature MHCII− and AA-DCs support angiogenesis, leading
Figure 4: Transcription of ficolin a and b relative to b actin in gd20 and 50 endometrium andtrophoblast from healthy (white) and arresting (black) attachment sites (N = 3). In gd20trophoblast there is decreased expression of both ficolin isoforms at arresting sites (B). In gd50trophoblast (D) there is an increase in the levels of both ficolin isoforms at arresting sites.
A. gd20 Endometrium
1.00E-04
1.00E-03
1.00E-02
1.00E-01
1.00E+00
Fic
olin
α a
nd β
Rel
ativ
e to
β-A
ctin Healthy
Arresting
Healthy Arresting
Healthy Arresting
Healthy Arresting
B. gd20 Trophoblast
C. gd50 Endometrium
1.00E-04
1.00E-03
1.00E-02
1.00E-01
1.00E+00
Fic
olin
α a
nd β
Rel
ativ
e to
β-A
ctin
1.00E-04
1.00E-03
1.00E-02
1.00E-01
1.00E+00F
icol
in α
and
βR
elat
ive
to β
-Act
in
1.00E-04
1.00E-03
1.00E-02
1.00E-01
1.00E+00
Fic
olin
α a
nd β
Rel
ativ
e to
β-A
ctin
D. gd50 Trophoblast
Alpha Beta Alpha Beta
Alpha BetaAlpha Beta
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

624 N. F. Linton et al.
to the proposal that the functions of DCs at the maternal fetal interface arehighly specialized and include secretion of VEGF and interactions with angio-genic NK cells to coordinate enlargement of the maternal vascular network.Chemokines with the CXC motif can be either ELR+ angiogenic or ELR- anti-angiogenic. The balance between these may determine fetal outcomes inindividual implantion sites. Ficolins, which stimulate some aspects of innateimmunity, also appear to contribute to complications seen in later pregnancyand may participate in trophoblast damage and/or its removal. Integration ofthese elements is extremely localized and, in litter-bearing species, is highlyefficient in selective removal of individuals without compromising allmembers of the litter.
ACKNOWLEDGMENTS
These studies were supported by awards from NSERC, OMAFRA, OntarioPork, Bioniche Life Sciences, Inc., and Agriculture and AgriFood Canada.
REFERENCES
Abrahams, V. M., Mor, G. (2005). Toll-like receptors and their role in the trophoblast.Placenta 26:540–547.
Adams, S., O’Neill, D. W., Bhardwa, N. (2005). Recent advances in dendritic cell biology.J. Clin. Immunol. 25:177–188.
Baggiolini, M., Dewald, B., Moser, B. (1994). Interleukin-8 and related chemotacticcytokines—CXC and CC chemokines. Adv Immunol 55:97–179.
Bischof, R. J., Brandon, M. R., Lee, C. S. (1995). Cellular immune responses in the piguterus during pregnancy. J. Reprod. Immunol. 29:161–178.
Blois, S. M., Alba Soto, C. D., Tometten, M., Klapp, B. F., Margni, R. A., Arck, P. C.(2004). Lineage, maturity, and phenotype of uterine murine dendritic cellsthroughout gestation indicate a protective role in maintaining pregnancy. Biol.Reprod. 70:1018–1023.
Charo, I. F., Ransohoff, R. M. (2006). The many roles of chemokines and chemokinereceptors in inflammation. N. Engl. J. Med. 354:610–621.
Croy, B. A., van den Heuvel, M. J., Borzychowski, A. M., Tayade, C. (2006). Uterinenatural killer cells: a specialized differentiation regulated by ovarian hormones.Immunol. Rev. 214:161–185.
Dikov, M. M., Ohm, J. E., Ray, N., Tchekneva, E. E., Burlison, J., Moghanaki, D.,Nadaf, S., Carbone, D. P. (2005). Differential roles of vascular endothelialgrowth factor receptors 1 and 2 in dendritic cell differentiation. J. Immunol.174: 215–222.
Douglas, G. C., Thirkill, T. L., Sideris, V., Rabieh, M., Trollinger, D., Nuccitelli, R.(2001). Chemokine receptor expression by human syncytiotrophoblast. J. Reprod.Immunol. 49:97–114.
Drake, P. M., Red-Horse, K., Fisher, S. J. (2002). Chemokine expression and functionat the human maternal-fetal interface. Rev. Endocr. Metab. Disord. 3:159–165.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Immune Mechanisms at the Maternal-Fetal Interface 625
Drake, P. M., Red-Horse, K., Fisher, S. J. (2004). Reciprocal chemokine receptor andligand expression in the human placenta: implications for cytotrophoblast differ-entiation. Dev. Dyn. 229:877–885.
Edgar, P. F. (1995). Hucolin, a new corticosteroid-binding protein from human plasmawith structural similarities to ficolins, transforming growth factor-beta 1-bindingproteins. FEBS Lett. 375:159–161.
Engelhardt, H., Croy, B. A., King, G. J. (2002a). Conceptus influences the distributionof uterine leukocytes during early porcine pregnancy. Biol. Reprod. 66:1875–1880.
Engelhardt, H., Croy, B. A., King, G. J. (2002b). Evaluation of natural killer cellrecruitment to embryonic attachment sites during early porcine pregnancy. Biol.Reprod. 66:1185–1192.
Engelhardt, H., Croy, B. A., King, G. J. (1997). Role of uterine immune cells in earlypregnancy in pigs. J. Reprod. Fertil. Suppl. 52:115–131.
Engert, S., Rieger, L., Kapp, M., Becker, J. C., Dietl, J., Kammerer, U. (2007). Profilingchemokines, cytokines and growth factors in human early pregnancy decidua byprotein array. Am. J. Reprod. Immunol. 58:129–137.
Fainaru, O., Adini, A., Benny, O., Adini, I., Short, S., Bazinet, L., Nakai, K., Pravda, E.,Hornstein, M. D., D’Amato, R. J., Folkman, J. (2008). Dendritic cells supportangiogenesis and promote lesion growth in a murine model of endometriosis.FASEB J. 22:522–529.
Foxcroft, G. R. (1997). Mechanisms mediating nutritional effects on embryonic survivalin pigs. J. Reprod. Fertil. Suppl. 52:47–61.
Freking, B. A., Leymaster, K. A., Vallet, J. L., Christenson, R. A., (2007). Number offetuses and conceptus growth throughout gestation in lines of pigs selected forovulation rate or uterine capacity. J. Anim. Sci. 85:2093–2103.
Fujimori, Y., Harumiya, S., Fukumoto, Y., Miura, Y., Yagasaki, K., Tachikawa, H.,Fujimoto, D. (1998). Molecular cloning and characterization of mouse ficolin-A.Biochem. Biophys. Res. Commun. 244:796–800.
Fukushima, K., Murata, M., Hachisuga, M., Tsukimori, K., Seki, H., Takeda, S.,Asanoma, K., Wake, N. (2008). Hypoxia inducible factor 1 Alpha regulates matrigel-induced endovascular differentiation under normoxia in a human extravilloustrophoblast cell line. Placenta 29:324–331.
Geisert, R. D., Brookbank, J. W., Roberts, R. M., Bazer. F. W. (1982). Establishment ofpregnancy in the pig: II. Cellular remodeling of the porcine blastocyst duringelongation on day 12 of pregnancy. Biol. Reprod. 27:941–955.
Grunewald, M., Avraham, I., Dor, Y., Bachar-Lustig, E., Itin, A., Jung, S., Chimenti, S.,Landsman, L., Abramovitch, R., Keshet, E. (2006). VEGF-induced adult neovascu-larization: recruitment, retention, and role of accessory cells. Cell 124:175–189.
Guan, H., Moretto, M., Bzik, D. J., Gigley, J., Khan, I. A. (2007). NK cells enhancedendritic cell response against parasite antigens via NKG2D pathway. J. Immu-nol. 179:590–596.
Hanna, J., Wald, O., Goldman-Wohl, D., Prus, D., Markel, G., Gazit, R., Katz, G.,Haimov-Kochman, R., Fujii, N., Yagel, S., Peled, A., Mandelboim, O. (2003).CXCL12 expression by invasive trophoblasts induces the specific migration ofCD16- human natural killer cells. Blood 102:1569–1577.
Hannan, N. J., Jones, R. L., White, C. A., Salamonsen, L. A. (2006). The chemokines,CX3CL1, CCL14, and CCL4, promote human trophoblast migration at the feto-maternal interface. Biol. Reprod. 74:896–904.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

626 N. F. Linton et al.
Hartgers, F. C., Figdor, C. G., Adema, G. J. (2000). Towards a molecular understandingof dendritic cell immunobiology. Immunol. Today 21:542–545.
Harumiya, S., Omori, A., Sugiura, T., Fukumoto, Y., Tachikawa, H., Fujimoto, D.(1995). EBP-37, a new elastin-binding protein in human plasma: structural simi-larity to ficolins, transforming growth factor-beta 1-binding proteins. J. Biochem.117:1029–1035.
He, Y. Y., Du, M. R., Guo, P. F., He, X. J., Zhou, W. H., Zhu, X. Y., Li, D. J. (2007).Regulation of C-C motif chemokine ligand 2 and its receptor in human decidualstromal cells by pregnancy-associated hormones in early gestation. Hum. Reprod.22:2733–2742.
Hirota, Y., Osuga, Y., Koga, K., Yoshino, O., Hirata, T., Morimoto, C., Harada, M.,Takemura, Y., Nose, E., Yano, T., Tsutsumi, O., Taketani, Y. (2006). The expres-sion and possible roles of chemokine CXCL11 and its receptor CXCR3 in thehuman endometrium. J. Immunol. 177:8813–8821.
Hoffmann, J. A., Kafatos, F. C., Janeway, C. A., Ezekowitz, R. A. (1999). Phylogeneticperspectives in innate immunity. Science 284:1313–1318.
Holmlund, U., Cebers, G., Dahlfors, A. R., Sandstedt, B., Bremme, K., Ekstrom, E. S.,Scheynius A. (2002). Expression and regulation of the pattern recognition recep-tors Toll-like receptor-2 and Toll-like receptor-4 in the human placenta. Immunol-ogy 107:145–151.
Huang, S. J., Schatz, F., Masch, R., Rahman, M., Buchwalder, L., Niven-Fairchild, T.,Tang, C., Abrahams, V. M., Krikun, G., Lockwood, C. J. (2006). Regulation ofchemokine production in response to pro-inflammatory cytokines in first trimesterdecidual cells. J. Reprod. Immunol. 72:60–73.
Huppertz, B., Abe, E., Murthi, P., Nagamatsu, T., Szukiewicz, D., Salafia, C. (2007).Placental angiogenesis, maternal and fetal vessels—a workshop report. Placenta28 Suppl A:S94-6.
Ichijo, H., Hellman, U., Wernstedt, C., Gonez, L. J., Claesson-Welsh, L., Heldin, C. H.,Miyazono, K. (1993). Molecular cloning and characterization of ficolin, a multimericprotein with fibrinogen- and collagen-like domains. J. Biol. Chem. 268:14505–14513.
Ichijo, H., Ronnstrand, L., Miyagawa, K., Ohashi, H., Heldin, C. H., Miyazono, K.(1991). Purification of transforming growth factor-beta 1 binding proteins fromporcine uterus membranes. J. Biol. Chem. 266:22459–22464.
Jarvenpaa, J., Vuoristo, J. T., Savolainen, E. R., Ukkola, O., Vaskivuo, T., Ryynanen, M.(2007). Altered expression of angiogenesis-related placental genes in pre-eclampsia associated with intrauterine growth restriction. Gynecol. Endo-crinol. 23:351–355.
Kammerer, U., Eggert, A. O., Kapp, M., McLellan, A. D., Geijtenbeek, T. B., Dietl, J.,van Kooyk, Y., Kampgen, E. (2003). Unique appearance of proliferating antigen-presenting cells expressing DC-SIGN (CD209) in the decidua of early human preg-nancy. Am. J. Pathol. 162:887–896.
Kelly, R. W., King, A. E., Critchley, H. O. (2001). Cytokine control in humanendometrium. Reproduction 121:3–19.
Koch, A. E., Polverini, P. J., Kunkel, S. L., Harlow, L. A., DiPietro, L. A., Elner, V. M.,Elner, S. G., Strieter, R. M. (1992). Interleukin-8 as a macrophage-derived media-tor of angiogenesis. Science 258:1798–1801.
Kollmar, O., Rupertus, K., Scheuer, C., Junker, B. Tilton, B., Schilling, M. K., Menger, M. D.(2007). Stromal cell-derived factor-1 promotes cell migration and tumor growth ofcolorectal metastasis. Neoplasia 9:862–870.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Immune Mechanisms at the Maternal-Fetal Interface 627
Kopcow, H. D., Karumanchi, S. A. (2007). Angiogenic factors and natural killer (NK)cells in the pathogenesis of preeclampsia. J. Reprod. Immunol. 76:23–29.
Kryczek, I., Lange,A., Mottram, P., Alvarez, X., Cheng, P., Hogan, M., Moons, L., Wei, S.,Zou, L., Machelon, V., Emilie, D., Terrassa, M., Lackner, A., Curiel, T. J.,Carmeliet, P., Zou, W. (2005). CXCL12 and vascular endothelial growth factor syner-gistically induce neoangiogenesis in human ovarian cancers. Cancer Res. 65:465–472.
Langenkamp, A., Messi, M., Lanzavecchia, A., Sallusto, F. (2000). Kinetics of dendriticcell activation: impact on priming of TH1, TH2 and nonpolarized T cells. Nat.Immunol. 1:311–316.
Li, X. F., Charnock-Jones, D. S., Zhang, E., Hiby, S., Malik, S., Day, K., Licence, D.,Bowen, J. M., Gardner, L., King, A., Loke, Y. W., Smith, S. K. (2001). Angiogenicgrowth factor messenger ribonucleic acids in uterine natural killer cells. J. Clin.Endocrinol. Metab. 86:1823–1834.
Liang, Z., Brooks, J., Willard, M., Liang, K., Yoon, Y., Kang, S., Shim, H. (2007).CXCR4/CXCL12 axis promotes VEGF-mediated tumor angiogenesis through Aktsignaling pathway. Biochem. Biophys. Res. Commun. 359:716–722.
Lucas, M., Schachterle, W., Oberle, K., Aichele, P., Diefenbach, A. (2007). Dendriticcells prime natural killer cells by trans-presenting interleukin 15. Immunity26:503–517.
Maione, T. E., Gray, G. S., Petro, J., Hunt, A. J., Donner, A. L., Bauer, S. I., Carson, H.F., Sharpe, R. J. (1990). Inhibition of angiogenesis by recombinant human plateletfactor-4 and related peptides. Science 247:77–79.
Matsushita, M., Endo, Y., Taira, S., Sato, Y., Fujita, T., Ichikawa, N., Nakata, M.,Mizuochi, T. (1996). A novel human serum lectin with collagen- and fibrinogen-likedomains that functions as an opsonin. J. Biol. Chem. 271:2448–2454.
Matsushita, M., Fujita, T. (2001). Ficolins and the lectin complement pathway. Immu-nol. Rev. 180:78–85.
McCarthy, C., Cotter, F. E., McElwaine, S., Twomey, A., Mooney, E. E., Ryan, F.,Vaughan, J. (2007). Altered gene expression patterns in intrauterine growthrestriction: potential role of hypoxia. Am. J. Obstet. Gynecol. 196:70.e1–70.e6.
McCarthy, SA., Kuzu, I., Gatter, K. C., Bicknell, R. (1991). Heterogeneity of the endot-helial cell and its role in organ preference of tumour metastasis. Trends Pharma-col. Sci. 12:462–467.
McKimmie, C. S., Graham, G. J. (2006). Leucocyte expression of the chemokine scaven-ger D6. Biochem. Soc. Trans. 34:1002–1004.
Mehrad, B., Keane, M. P., Strieter, R. M. (2007). Chemokines as mediators of angio-genesis. Thromb. Haemost. 97:755–762.
Moussa, M., Mognetti, B., Dubanchet, S., Menu, E, Roques, P., Dormont, D., Barre-Sinoussi, F., Chaouat, G. (2001). Expression of beta chemokines in explants and tropho-blasts from early and term human placentae. Am. J. Reprod. Immunol. 46:309–317.
Noonan, D. M., De Lerma Barbaro, A., Vannini, N., Mortara, L., Albini, A. (2008).Inflammation, inflammatory cells and angiogenesis: decisions and indecisions.Cancer Metastasis Rev. 27:31–40.
Ohashi, T., Erickson, H. P. (1998). Oligomeric structure and tissue distribution officolins from mouse, pig and human. Arch. Biochem. Biophys. 360:223–232.
Ohashi, T., Erickson, H. P. (1997). Two oligomeric forms of plasma ficolin havedifferential lectin activity. J. Biol. Chem. 272:14220–14226.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

628 N. F. Linton et al.
Patni, S., Flynn, L. P., Wynen, P., Seager, A. L., Morgan, G., White, J. O., Thornton, C.A. (2007). An introduction to Toll-like receptors and their possible role in theinitiation of labour. BJOG 114:1326–1334.
Petit, I., Jin, D., Rafii, S. (2007). The SDF-1-CXCR4 signaling pathway: a molecularhub modulating neo-angiogenesis. Trends Immunol. 28:299–307.
Pinhal-Enfield, G., Ramanathan, M., Hasko, G., Vogel, S. N., Salzman, A. L., Boons, G.J., Leibovich, S. J. (2003). An angiogenic switch in macrophages involving synergybetween Toll-like receptors 2, 4, 7, and 9 and adenosine A(2A) receptors. Am. J.Pathol. 163:711–721.
Pope, W. F. (1994). Embryonic mortality in swine. In: M. Zavy and R. Geiserteds. Embryonic Mortality in Domestic Species, CRC Press, Boca Raton, FL.pp. 53–77.
Presta, M., Belleri, M., Vecchi, A., Hesselgesser, J., Mantovani, A., Horuk, R. (1998).Noncompetitive, chemokine-mediated inhibition of basic fibroblast growth factor-induced endothelial cell proliferation. J. Biol. Chem. 273:7911–7919.
Red-Horse, K., Drake, P. M., Gunn, M. D., Fisher, S. J. (2001). Chemokine ligand andreceptor expression in the pregnant uterus: reciprocal patterns in complementarycell subsets suggest functional roles. Am. J. Pathol. 159:2199–2213.
Riboldi, E., Musso, T., Moroni, E., Urbinati, C., Bernasconi, S., Rusnati, M., Adorini, L.,Presta, M., Sozzani, S. (2005). Cutting edge: proangiogenic properties of alterna-tively activated dendritic cells. J. Immunol. 175:2788–2792.
Sato, Y., Higuchi, T., Yoshioka, S., Tatsumi, K., Fujiwara, H., Fujii, S. (2003). Tropho-blasts acquire a chemokine receptor, CCR1, as they differentiate towards invasivephenotype. Development 130:5519–5532.
Spencer, T. E., Bazer, F. W. (2004). Conceptus signals for establishment and mainte-nance of pregnancy. Reprod. Biol. Endocrinol. 2:49.
Stingl, G., Bergstresser, P. R. (1995). Dendritic cells: a major story unfolds. Immunol.Today 16:330–333.
Strieter, R. M., Kunkel, S. L., Elner, V. M., Martonyi, C. L., Koch, A. E., Polverini, P. J.,Elner, G. (1992). Interleukin-8. A corneal factor that induces neovascularization.Am. J. Pathol. 141:1279–1284.
Strieter, R. M., Polverini, P. J., Kunkel, S. L., Arenberg, D. A., Burdick, M. D., Kasper, J.,Dzuiba, J., Van Damme, J., Walz, A., Marriott, D. (1995). The functional role ofthe ELR motif in CXC chemokine-mediated angiogenesis. J. Biol. Chem. 270:27348–27357.
Tayade, C., Black, G. P., Fang, Y., Croy, B. A. (2006). Differential gene expression inendometrium, endometrial lymphocytes, and trophoblasts during successful andabortive embryo implantation. J. Immunol. 176:148–156.
Tayade, C., Fang, Y., Croy, B. A. (2007b). A review of gene expression in porcineendometrial lymphocytes, endothelium and trophoblast during pregnancy successand failure. J. Reprod. Dev. 53:455–463.
Tayade, C., Fang, Y., Hilchie, D., Croy, B. A. (2007a). Lymphocyte contributions toaltered endometrial angiogenesis during early and midgestation fetal loss. J. Leukoc.Biol. 82:877–886.
Wang, C. C., Yim, K. W., Poon, T. C., Choy, K. W., Chu, C. Y., Lui, W. T., Lau, T. K.,Rogers, M. S., Leung, T. N. (2007). Innate immune response by ficolin binding inapoptotic placenta is associated with the clinical syndrome of preeclampsia. Clin.Chem. 53:42–52.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Immune Mechanisms at the Maternal-Fetal Interface 629
Wang, R. F., Miyahara, Y., Wang, H. Y. (2008). Toll-like receptors and immune regula-tion: implications for cancer therapy. Oncogene 27:181–189.
Wessels, J. M. (in press). Chemokine and chemokine decoy receptor contributions tofetal success and failure during porcine pregnancy.
Wessels, J. M., Linton, N. F., Croy, B. A., Tayade, C. (2007). A review of molecularcontrasts between arresting and viable porcine attachment sites. Am. J. Reprod.Immunol. 58:470–480.
Wilson, M. E., Biensen, N. J., Ford, S. P. (1999). Novel insight into the control of littersize in pigs, using placental efficiency as a selection tool. J. Anim. Sci. 77:1654–1658.
Wu, X., Jin, L. P., Yuan, M. M., Zhu, Y., Wang, M. Y., Li, D. J. (2005). Human first-trimester trophoblast cells recruit CD56brightCD16- NK cells into decidua by wayof expressing and secreting of CXCL12/stromal cell-derived factor 1. J. Immunol.175:61–68.
Zhang, C., Zhang, J., Niu, J., Zhang, J., Tian, Z. (in press). Interleukin-15 improvescytotoxicity of natural killer cells via up-regulating NKG2D and cytotoxic effectormolecule expression as well as STAT1 and ERK1/2 phosphorylation. Cytokine
Zheng, H., G. Fu, T. Dai, and H. Huang (2007). Migration of endothelial progenitorcells mediated by stromal cell-derived factor-1alpha/CXCR4 via PI3K/Akt/eNOSsignal transduction pathway. J. Cardiovasc. Pharmacol. 50:274–280.
Zhou, B., Han, Z. C., Poon, M. C., Pu, W. (2007). Mesenchymal stem/stromal cells(MSC) transfected with stromal derived factor 1 (SDF-1) for therapeutic neovascu-larization: enhancement of cell recruitment and entrapment. Med. Hypotheses68:1268–1271.
Zygmunt, M., Herr, F., Munstedt, K., Lang, U., Liang, O. D. (2003). Angiogenesis andvasculogenesis in pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 110 Suppl1:S10–18.
Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.

Imm
unol
Inv
est D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Cal
gary
on
10/0
3/13
For
pers
onal
use
onl
y.