
Lesson 11 Adaptive Immunity “Specific Immunity” March 31,2015.

Immunity, Vol. 17, 549–560, November, 2002, Copyright 2002 by Cell Press
Invariant Chain-Induced B Cell DifferentiationRequires Intramembrane Proteolytic Releaseof the Cytosolic Domain
II �� dimers bind sequentially to a trimer of the Ii chainto form a nonameric complex (��Ii)3 (Anderson andCresswell, 1994; Marks et al., 1990; Roche et al., 1991),which then exits the ER. After being transported to thetrans Golgi, the ��Ii complex is diverted from the secre-
Didi Matza, Anat Kerem, Helena Medvedovsky,Frida Lantner, and Idit Shachar1
Department of Immunologythe Weizmann Institute of ScienceRehovot
tory pathway to the endocytic system (Bakke and Dob-Israel, 76100berstein, 1990; Lamb et al., 1991; Lotteau et al., 1990;Pieters et al., 1993) and ultimately to acidic endosome/lysosome-like structures called MHC class II compart-Summaryments (MIIC or CIIV; Stumptner-Cuvelette and Bena-roch, 2002). The N-terminal cytoplasmic tail of Ii containsImmature B cells differentiate in the spleen into maturetwo extensively characterized dileucine-based endoso-B cells, a process that is essential for their participa-mal targeting motifs (Lotteau et al., 1990; Odorizzi et al.,tion in the immune response. Previously, we showed1994; Pond et al., 1995). These motifs mediate internal-that the MHC class II chaperone, invariant chain (Ii),ization from the plasma membrane and from the transcontrols this differentiation to the mature stage. Ii cy-Golgi network. Once in this pathway, the Ii chain is pro-tosolic domain-induced B cell maturation involves ac-teolytically degraded, leaving only a small fragment, thetivation of transcription mediated by the NF-�B p65/class II-associated Ii chain peptide (CLIP), bound to theRelA homodimer and requires the B cell enriched co-released �� dimers. The final step requires interactionactivator, TAFII105. In this study we show that the cyto-of ��CLIP complexes in the specialized lysosome-likesolic region of Ii is cleaved within the plane of thecompartment with another class II-related �� dimer,membrane to generate a cytosolic fragment, which iscalled HLA-DM in the human system and H2-M in mice.essential for NF-�B activation and B cell differentia-This drives out the residual CLIP, rendering the �� di-tion. Our results suggest that Ii functions as a mem-mers ultimately competent to bind antigenic peptidesbrane-bound inactive inducer of NF-�B transcrip-that are mainly derived from internalized antigens andtion that is activated by intramembrane proteolyticare also delivered to the endocytic pathway (Neefjescleavage.et al., 1990; Roche and Cresswell, 1991). The peptide-loaded class II molecules then leave this compartment,Introductionby an unknown route, to be expressed on the cell surfaceand surveyed by CD4� T cells.B lymphocytes differentiate from hematopoietic stem
Previously, we have shown that besides its role as ancells in the bone marrow, resulting in formation of imma-MHC class II chaperonin molecule, Ii has an additionalture B cells. Immature cells leave the bone marrow androle in B cell maturation (Shachar and Flavell, 1996). Inmigrate to the spleen for their final differentiation intomice lacking Ii, the development from immature to ma-mature cells. The transition from immature to mature Bture B cells is impaired and B cells are arrested at ancells in the spleen is characterized by a series ofimmature stage characterized by low expression levelschanges in surface marker expression and in the activi-of IgD and CD23 and poor response to T-independentties of these cells. Recently, this transitional stage wasantigens (Kenty and Bikoff, 1999; Kenty et al., 1998;
further characterized and divided into two cell popula-Shachar and Flavell, 1996). At present, the molecular
tions. Transitional B cells of type 1 (T1) are the recentmechanism by which Ii controls B cell maturation is
immigrants from the bone marrow. These cells develop unknown. We have recently dissected the chaperonininto transitional B cells of type 2 (T2), which cycle and activity of Ii from its role in B cell maturation and demon-are found exclusively in the primary follicles of the spleen strated in vivo that Ii N-terminal domain is directly in-(Loder et al., 1999). The molecular mechanisms control- volved in the maturation of B cells and is sufficient toling the selection of immature B cells and especially promote B cell differentiation (Matza et al., 2002). More-their differentiation in the spleen are largely unknown. over, we have shown that Ii can act as a signaling mole-
MHC class II molecules are heterodimeric complexes cule and that its cytosolic domain induces B cell matura-that present foreign antigenic peptides on the cell sur- tion by activation of transcription mediated by the NF-�Bface of antigen-presenting cells (APCs) to CD4� T cells p65/RelA homodimer and requires the B cell-enriched(Harding and Unanue, 1990; Long, 1989; Unanue, 1984). coactivator, TAFII105 (Matza et al., 2001).MHC class II synthesis and assembly begins in the endo- In this study we followed the Ii cytosolic domain andplasmic reticulum (ER) with the noncovalent association the mechanism by which it transmits the signal to NF-of the � and � chains with trimers of invariant chain (Ii) �B. We show here that Ii cytosolic domain is released(Cresswell, 1994). Ii is a nonpolymorphic type II integral by proteolytic cleavage from the membrane, an essentialmembrane protein, in which the N-terminal constitutes event for NF-�B activation and B cell maturation.a short cytoplasmic tail of 30 amino acids (aa), followedby a single 24 aa transmembrane region and an approxi- Resultsmately 150 aa long lumenal domain. Three MHC class
Previously, we demonstrated that truncated Ii lackingits lumenal domain (aa 1-82) is sufficient to induce differ-1Correspondence: [email protected]

Immunity550
entiation of immature B cells (Matza et al., 2001, 2002). al., 1999) and separated on Tricine gels to visualize lowmolecular weight fragments. Western blot analysis withTo determine the mechanism by which this truncated Ii
activates NF-�B, we followed its localization in the cell. the IN1 antibody showed, besides the FL and 1-82 myctagged proteins, small bands (Figure 2D). Antibody, di-To this end, GFP chimeras linked to the N (GFP 1-82)
or C terminus (1-82 GFP) of 1-82 li were generated (Fig- rected against the Myc epitope, detected the FL and1-82 Myc proteins but not the cytosolic fragments, furtherures 1A and 1B). These constructs were transfected
into HEK 293 cells, which do not express endogenous indicating that these fragments were generated by N-ter-minal proteolytic cleavage (Figure 2D). To further dem-Ii or MHC II molecules. Moreover, Ii was previously
shown to activate NF-�B in these cells (Matza et al., onstrate that the cytosolic fragment indeed contains thecytosolic domain of Ii, we fused an Xpress epitope to2001). As expected, since the cytosolic domain of Ii is
known to contain targeting motifs for transport to the the N-terminal of FL Ii (Figure 1B). As shown in Figure2E, the cytosolic bands were detected by both IN1 andendosomal compartment, the chimeric GFP fused to
1-82 lumenal domain (1-82 GFP) was localized in the anti-Xpress antibodies, indicating that these fragmentsdo indeed represent a proteolytic product of the Ii cyto-endosomes (Figure 1C). However, the reciprocal 1-82
chimera in which GFP is fused to its cytosolic domain solic domain. Moreover, Xpress FL transfected cellswere fractionated and their membrane and cytosolic(GFP 1-82) was mostly distributed in the cytoplasm (Fig-
ure 1D). To verify the nature of these compartments, fractions were separated. As can be seen in Figure 2F,while the FL Ii could be detected only in the membranecells were transfected with the GFP full-length p31 Ii
(GFP FL) construct. As can be seen in Figure 1E, this fraction, indicating a proper insertion of the chimericprotein to the membrane, in the cytosolic fraction onlyprotein was mostly localized in the previously described
endocytic compartments (Pieters et al., 1993; Romag- the released Xpress-Ii fragment was found. These re-sults show that both the FL and the 1-82 truncated Iinoli et al., 1993; Stang and Bakke, 1997) as conferred
by specific staining of these compartments, while the release their cytosolic fragment.In order to localize the cytosolic fragment cleavagelocalization of the GFP 1-82 construct did not overlap
with these endosomal markers (Figure 1F). The overlap site, we constructed truncation mutants of Ii containingamino acids 1-60, 1-50, and 1-42 C-terminally fused tobetween the GFP and the endocytic marker may some-
times appear only partial due to the much stronger green the Myc epitope (Figure 1B). Extracts from 293 cellstransfected with these various Ii various constructs wereflouresence of the GFP-Ii as compared to the red marker.
These results indicate that although the endosomal tar- separated on Tricine gels and analyzed by Western blotanalysis with the IN1 antibody. As seen in Figure 3A,geting motifs in Ii 1-82 are functional, GFP, when linked
to the Ii cytosolic domain, demonstrate a cytoplasmic 1-60 and 1-50 were cleaved, although the latter to alesser extent, to generate the cytosolic fragments. How-appearance, suggesting a proteolytic release of the GFP
from the membrane. ever, the 1-42 Myc tagged molecule was not furthercleaved to the cytosolic fragment. Together, these re-To confirm these results, we searched for the exis-
tence of Ii cytosolic fragments in lysates of 293 cells sults suggest that the cytosolic domain of Ii is releasedfrom the Ii molecule by a proteolytic event that occurstransfected with the 1-82 Ii GFP, the GFP 1-82 Ii, as well
as the full-length p31 Ii (FL) linked to GFP in its cytosolic in the transmembrane domain in the region of aminoacids 42-50.domain (GFP FL) by Western blot analysis using the
IN1 antibody (which recognizes the Ii cytosolic domain) To analyze the specific amino acids involved in theproteolytic release of the Ii cytosolic domain, we mu-(Figure 1A). Interestingly, in addition to the Ii FL and
1-82 proteins, we detected smaller fragments (Figure tated amino acids that might be involved in this proteo-lytic process. Previously, Lipp and Dobberstein de-2A) when GFP was fused only to the N-terminal domain
of Ii. Using anti-GFP antibodies we could show that scribed a cleavage site in the transmembrane region ofhuman Ii at amino acid 42 (Lipp and Dobberstein, 1986).besides the cytosolic fragment of Ii, these smaller bands
contained the GFP molecules (Figure 2B). Moreover, We hypothesized that the cleavage that releases thecytosolic domain of Ii occurs at this amino acid. Toseparation between the membrane and the cytosolic
fractions revealed almost identical insertion of the three determine whether these amino acids may be importantfor both mouse and human Ii cleavage, we aligned theirconstructs to the membrane, while the released band
was detected mostly in the cytosolic fraction (Figure sequences (Figure 3B). Three leucines that were foundidentical in human and mouse at positions 42 to 44 were2C). Since the released fragment appeared roughly as
37–40 kDa and the GFP protein was estimated in our mutated to alanines in the 1-82 constructs (GFP 1-82LLL42-44AAA and 1-82 Myc LLL42-44AAA). Further-gels to run with an apparent molecular weight of about
33 kDa, the cytosolic fragment linked to the GFP was more, we identified in the Ii cytosolic domain a recentlydescribed RXXL motif, which serves as a degradationestimated to be roughly about 4–7 kDa in size.
To determine whether these small bands represent signal (d box) in proteins (Agami and Bernards, 2000;Cooper et al., 1997; Duncan et al., 1997; Zur and Bran-an artifact derived from purification of the GFP chimera
or are indeed specific degradation products of Ii, we deis, 2001). To destroy this putative d box, we mutatedthe amino acids at positions 28-30 (GFP 1-82 SRG28-analyzed the existence of these fragments in additional
Ii tagged constructs. Vector plasmids encoding Ii FL or 30AAA and 1-82 myc SRG28-30AAA). To verify insertionof the different constructs to the membrane, transfected1-82 molecules tagged C-terminally with a Myc epitope
and driven by an EF4 promoter (elongation factor 4) 293 cells were fractionated and their membrane andcytosolic fractions were separated. As can be seen in(illustrated in Figure 1B) were transfected into 293 cells.
The proteins were extracted using the hot-SDS method Figure 3C, the 1-82 chimera and the two 1-82 mutatedmyc tagged proteins (1-82 myc SRG28-30AAA and 1-82in order to preserve rapidly degraded proteins (Niwa et
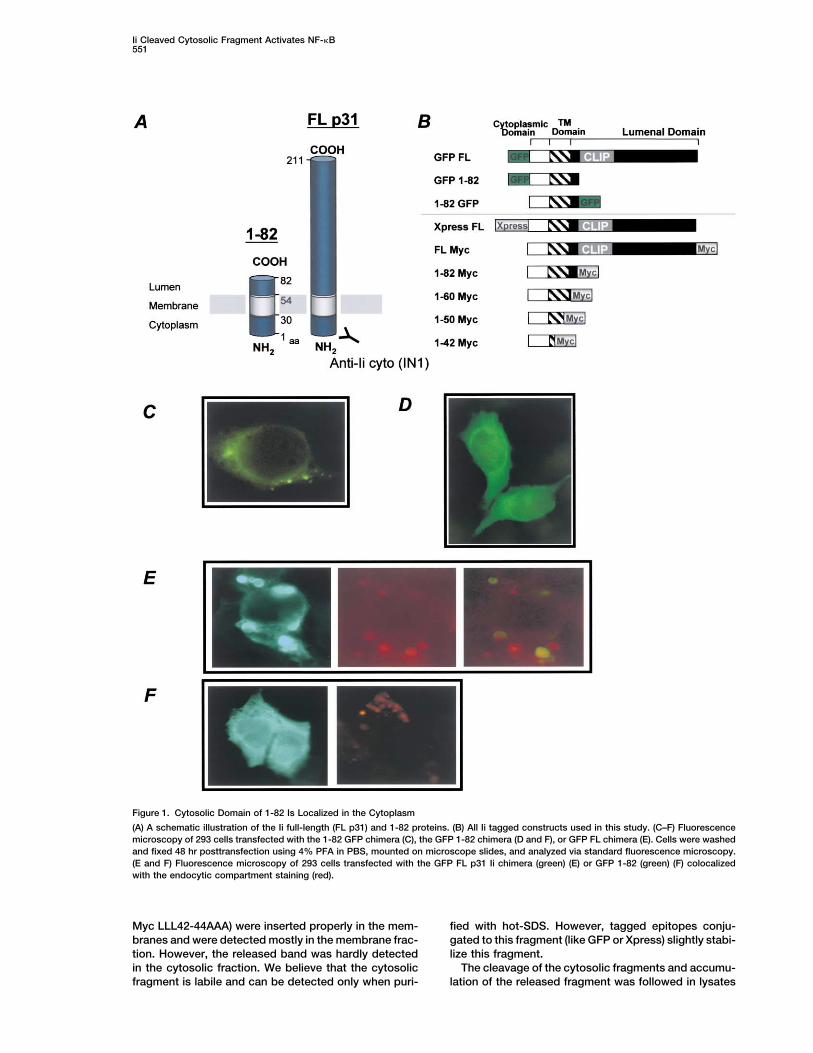
Ii Cleaved Cytosolic Fragment Activates NF-�B551
Figure 1. Cytosolic Domain of 1-82 Is Localized in the Cytoplasm
(A) A schematic illustration of the Ii full-length (FL p31) and 1-82 proteins. (B) All Ii tagged constructs used in this study. (C–F) Fluorescencemicroscopy of 293 cells transfected with the 1-82 GFP chimera (C), the GFP 1-82 chimera (D and F), or GFP FL chimera (E). Cells were washedand fixed 48 hr posttransfection using 4% PFA in PBS, mounted on microscope slides, and analyzed via standard fluorescence microscopy.(E and F) Fluorescence microscopy of 293 cells transfected with the GFP FL p31 Ii chimera (green) (E) or GFP 1-82 (green) (F) colocalizedwith the endocytic compartment staining (red).
Myc LLL42-44AAA) were inserted properly in the mem- fied with hot-SDS. However, tagged epitopes conju-gated to this fragment (like GFP or Xpress) slightly stabi-branes and were detected mostly in the membrane frac-
tion. However, the released band was hardly detected lize this fragment.The cleavage of the cytosolic fragments and accumu-in the cytosolic fraction. We believe that the cytosolic
fragment is labile and can be detected only when puri- lation of the released fragment was followed in lysates

Immunity552
Figure 2. The Cytosolic Domain of Ii Is Proteolytically Cleaved
Western blot analysis of 293 cell lysates transfected with various Ii tagged constructs. (A and B) After transfection with GFP FL, GFP 1-82,and 1-82 GFP, the cells were cultured for 48 hr and lysed; boiled total lysates were separated on 8% (w/v) SDS-PAGE and analyzed with IN1rat monoclonal antibodies that recognize the Ii cytosolic domain (A), or anti-GFP antibodies (B). (C) Membranes and cytosol of GFP FL, GFP1-82, and 1-82 GFP transfected cells were separated on 8% (w/v) SDS-PAGE and analyzed with anti-GFP antibodies. (D and E) Western blotanalysis of 293 cell lysates transfected with Myc tagged 1-82 (D) and FL (D and E) or Xpress tagged FL Ii (E). After transfection, the cells werecultured for 48 hr and lysed using the hot-SDS method; boiled total lysates were separated on Tricine gels and analyzed with IN1 ([D], lanes1 and 2 and [E], lanes 1 and 2) or anti-Myc mouse monoclonal antibodies ([D], lanes 3 and 4) or anti-Xpress mouse monoclonal antibody ([D],lanes 3 and 4). (F) Membranes and cytosol fractions of Xpress FL transfected cells were separated on Tricine gels and analyzed with IN1.
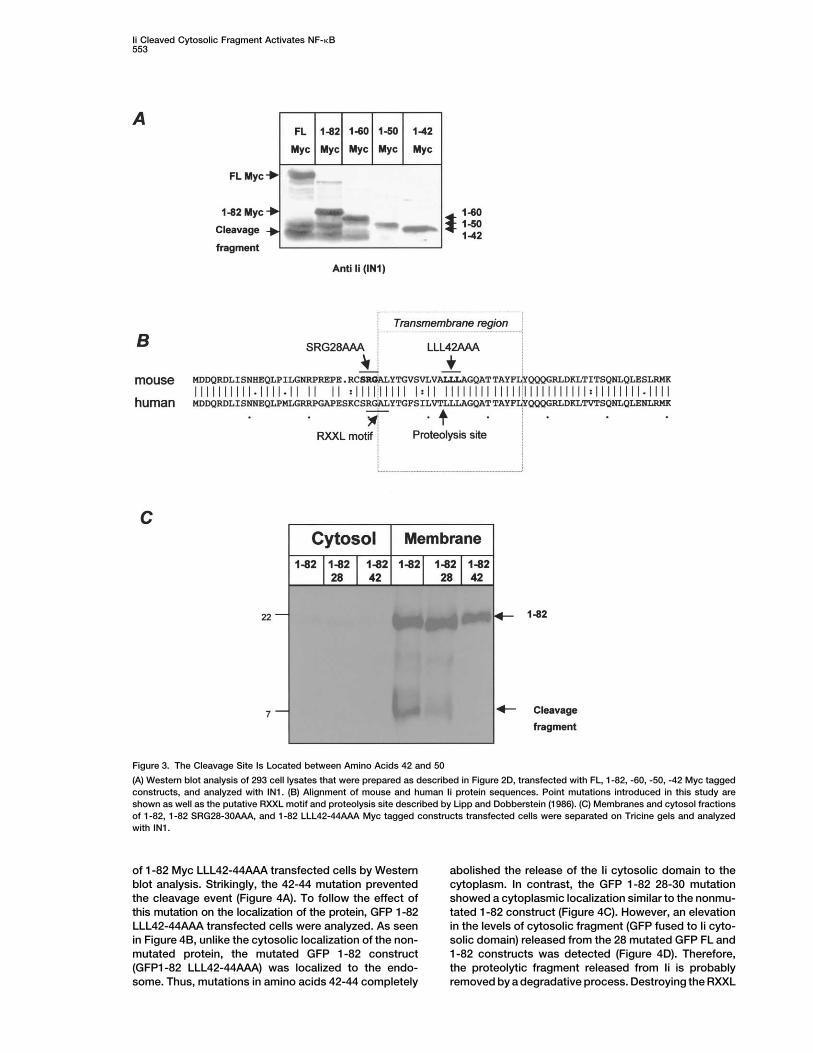
Ii Cleaved Cytosolic Fragment Activates NF-�B553
Figure 3. The Cleavage Site Is Located between Amino Acids 42 and 50
(A) Western blot analysis of 293 cell lysates that were prepared as described in Figure 2D, transfected with FL, 1-82, -60, -50, -42 Myc taggedconstructs, and analyzed with IN1. (B) Alignment of mouse and human Ii protein sequences. Point mutations introduced in this study areshown as well as the putative RXXL motif and proteolysis site described by Lipp and Dobberstein (1986). (C) Membranes and cytosol fractionsof 1-82, 1-82 SRG28-30AAA, and 1-82 LLL42-44AAA Myc tagged constructs transfected cells were separated on Tricine gels and analyzedwith IN1.
of 1-82 Myc LLL42-44AAA transfected cells by Western abolished the release of the Ii cytosolic domain to thecytoplasm. In contrast, the GFP 1-82 28-30 mutationblot analysis. Strikingly, the 42-44 mutation prevented
the cleavage event (Figure 4A). To follow the effect of showed a cytoplasmic localization similar to the nonmu-tated 1-82 construct (Figure 4C). However, an elevationthis mutation on the localization of the protein, GFP 1-82
LLL42-44AAA transfected cells were analyzed. As seen in the levels of cytosolic fragment (GFP fused to Ii cyto-solic domain) released from the 28 mutated GFP FL andin Figure 4B, unlike the cytosolic localization of the non-
mutated protein, the mutated GFP 1-82 construct 1-82 constructs was detected (Figure 4D). Therefore,the proteolytic fragment released from Ii is probably(GFP1-82 LLL42-44AAA) was localized to the endo-
some. Thus, mutations in amino acids 42-44 completely removed by a degradative process. Destroying the RXXL

Immunity554
Figure 4. Amino Acids 42-44 Are Essential for Ii Intramembrane Cleavage and the RXXL Motif Stabilizes the Cleaved Cytosolic Fragment
(A) Western blot analysis of 293 cell lysates transfected with 1-82 and 1-82 LLL42-44AAA Myc tagged constructs. After transfection, the cellswere cultured for 48 hr and lysed using the hot-SDS method; boiled total lysates were separated on a Tricine gel and analyzed with IN1.(B and C) 293 cells were transfected with GFP 1-82 or the GFP 1-82 LLL42-44AAA chimera, in which GFP is fused to the N-terminal cytosolicdomain (B) or with GFP 1-82 or the GFP 1-82 SRG28-30AAA chimera, in which GFP is fused to the N-terminal cytosolic domain (C). Cellswere washed and fixed 48 hr following transfection using 4% PFA in PBS, mounted on microscope slides, and analyzed by standardfluorescence microscopy. The dark part shows the flourecence staining while the light part shows the same cells by phase contrast.(D) Western blot analysis of GFP FL or 1-82 and their 28 mutated counterparts transfected 293 cell lysates. After transfection, the cells werecultured for 48 hr, lysed, and boiled total lysates were separated on 8% (w/v) SDS-PAGE and analyzed with IN1 Abs.
motif might have reduced this rapid degradation of the lyzed by Western blot using the IN1 antibody. As shownin Figure 5A, similarly to the 293 cells, Ii cytosolic frag-cytosolic fragment and allowed its accumulation in the
cells. ments were observed in primary B cells. In addition,primary B cells were pulse labeled and chased for vari-ous times. As can be seen in Figure 5B, the proteolyticCleavage of Invariant Chain Occurs in Primary B Cells
In an attempt to determine whether Ii is proteolytically fragment accumulated after 2 hr of chase. Thus, Ii isnaturally cleaved in primary B cells.cleaved naturally in primary B cells, we followed the
formation of the cytosolic fragments in these cells. Pro- To demonstrate the localization and behavior of Ii inprimary B cells, we transfected these cells with GFPteins from B splenocytes from C57BL/6 mouse were
extracted by using the hot-SDS method and were ana- constructs of Ii, as described above (Figure 1B). As can

Ii Cleaved Cytosolic Fragment Activates NF-�B555
Proteolytic Release of Ii Cytosolic Domain Is Requiredfor Its Function in NF-�B Activationand B Cell MaturationPreviously, we have shown that Ii induces B cell matura-tion by activating TAFII105-NF-�B-dependent transcrip-tion program (Matza et al., 2001). Our experiments dem-onstrated that one possible pathway utilized by Iiinvolves modulation of the p65/RelA activation domain.We speculate that this event augments the interactionof p65 with coactivator molecules such as TAFII105, re-sulting in activation of specific genes required for B cellmaturation. To determine whether the Ii cleaved peptide(1-42 peptide) is able to induce NF-�B activity, we uti-lized an expression plasmid that encodes a luciferasereporter plasmid containing two tandem NF-�B sitesand a minimal core promoter (Matza et al., 2001). Ascan be seen in Figure 7A, the released Ii peptide elevatedNF-�B activation similarly to Ii 1-82 molecule. In addi-tion, to investigate the mechanism by which Ii controlsNF-�B, we examined whether the effect of Ii is directedto the transcriptional activation function of p65/RelA. Forthis purpose a fusion of the C-terminal transactivationdomain of p65/RelA (TA1) with the DNA binding domainof the yeast transcription factor Gal4 was constructed.This plasmid was transfected along with a luciferasereporter containing the Gal4 binding sites in the pres-ence of various Ii plasmids (Matza et al., 2001). As canbe seen in Figure 7B, both 1-82 Myc and 1-42 Mycpeptides induced NF-�B activation. In addition, in orderto examine whether the proteolytic release of Ii cytosolicdomain is required for the induction of NF-�B activity,we studied the effect of various Ii expression constructson NF-�B transcription. Strikingly, as shown in Figure7C, the 42-44 mutation of 1-82 (1-82 Myc LLL42-44AAA)abolished 1-82 induction of NF-�B activation. However,the 1-82 28-30 mutation (1-82 Myc SRG28-30AAA) dra-matically increased 1-82 induced NF-�B activation.
Moreover, we have previously developed an in vitrosystem to analyze B cell maturation. The protocol weused for primary B cell transfection involves pretreat-ment of the cells with LPS. LPS is known as a potentFigure 5. Ii Cytosolic Domain Is Cleaved in Primary B Cellsinducer of NF-�B activity in B cells as well as in other(A) Western blot analysis of Ii in control C57BL/6 B cells. Purified Bcell types. However, Ii�/� primary B cells failed to differ-cells were lysed via the hot-SDS method, and boiled total lysates
were separated on 18% (w/v) SDS-PAGE and analyzed with IN1 Rat entiate in the presence of this compound whereas NF-monoclonal antibodies, which recognize the Ii cytosolic domain. A �B p65/RelA expression effectively induced maturationlysate of 293 cells transfected with the FL Myc construct, described (Matza et al., 2001). In this assay, primary immature Bin Figure 1B, was separated on the same gel and served as control cells from Ii-deficient mice can differentiate to maturefor the size of the cleaved cytosolic fragment.
cells in culture, by introducing Ii expression plasmid.(B) Pulse chase analysis of primary B cells. Purified B cells wereWe have found that IgD was the best marker to charac-pulse labeled with S35-Met for 10 min and chased for the indicatedterize this in vitro differentiation (Matza et al., 2001).time points. Immunoprecipitated labeled proteins were separated
on an 18% (w/v) SDS-PAGE gel. To directly show whether the proteolytic release of Iicytosolic domain is required for B cell differentiation,we have introduced the 1-42 Myc, 1-82 Myc, and the
be seen in Figure 6A, B full-length Ii was localized mostly 42-44 mutation of 1-82 (1-82 Myc LLL42-44AAA) andto the endosomes, while the GFP 1-82 construct was the 1-82 28-30 mutation (1-82 Myc SRG28-30AAA) to Ii-detected in the cytoplasm (Figure 6C). However, as was deficient primary B cells. As can be seen in Figure 7C,seen in the 293 cells, unlike the cytoplasmic appearance 1-82 Myc and 1-42 Myc induced differentiation of imma-of the 1-82 construct, the mutated GFP1-82 LLL42- ture B cells. While 1-82 Myc LLL42-44AAA was unable44AAA chimera was detected in the endosomes, more to induce B cell differentiation, the 1-82 Myc SRG28-profoundly than the FL construct (Figures 6D and 6E). 30AAA mutation dramatically elevated the differentiationThus, invariant chain cytosolic domain is cleaved and induced by the Ii 1-82 molecule. Thus, during matura-released from the membranes of primary B cells. As in tion, Ii cytosolic domain is cleaved and released to the293 cells, this release is inhibited by mutation of aa cytoplasm, a process that is essential for NF-�B activa-
tion. The released cytosolic segment subsequently re-residues 42-44.
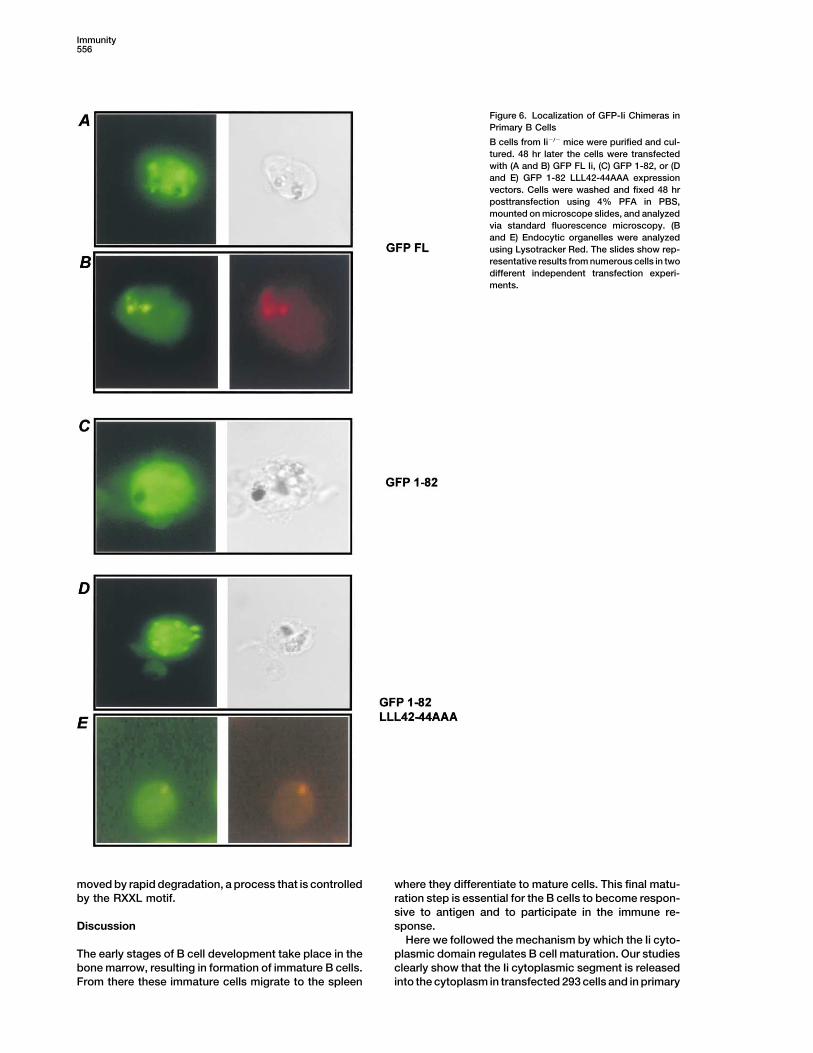
Immunity556
Figure 6. Localization of GFP-Ii Chimeras inPrimary B Cells
B cells from Ii�/� mice were purified and cul-tured. 48 hr later the cells were transfectedwith (A and B) GFP FL Ii, (C) GFP 1-82, or (Dand E) GFP 1-82 LLL42-44AAA expressionvectors. Cells were washed and fixed 48 hrposttransfection using 4% PFA in PBS,mounted on microscope slides, and analyzedvia standard fluorescence microscopy. (Band E) Endocytic organelles were analyzedusing Lysotracker Red. The slides show rep-resentative results from numerous cells in twodifferent independent transfection experi-ments.
moved by rapid degradation, a process that is controlled where they differentiate to mature cells. This final matu-ration step is essential for the B cells to become respon-by the RXXL motif.sive to antigen and to participate in the immune re-sponse.Discussion
Here we followed the mechanism by which the Ii cyto-plasmic domain regulates B cell maturation. Our studiesThe early stages of B cell development take place in the
bone marrow, resulting in formation of immature B cells. clearly show that the Ii cytoplasmic segment is releasedinto the cytoplasm in transfected 293 cells and in primaryFrom there these immature cells migrate to the spleen
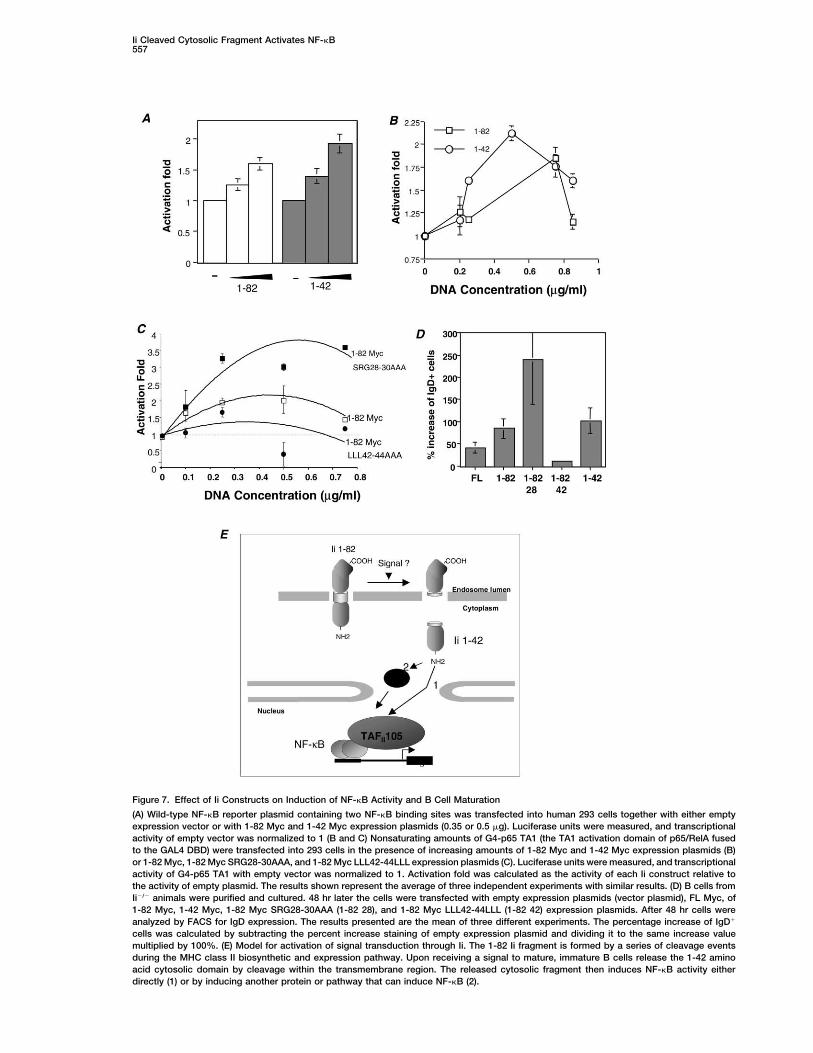
Ii Cleaved Cytosolic Fragment Activates NF-�B557
Figure 7. Effect of Ii Constructs on Induction of NF-�B Activity and B Cell Maturation
(A) Wild-type NF-�B reporter plasmid containing two NF-�B binding sites was transfected into human 293 cells together with either emptyexpression vector or with 1-82 Myc and 1-42 Myc expression plasmids (0.35 or 0.5 �g). Luciferase units were measured, and transcriptionalactivity of empty vector was normalized to 1 (B and C) Nonsaturating amounts of G4-p65 TA1 (the TA1 activation domain of p65/RelA fusedto the GAL4 DBD) were transfected into 293 cells in the presence of increasing amounts of 1-82 Myc and 1-42 Myc expression plasmids (B)or 1-82 Myc, 1-82 Myc SRG28-30AAA, and 1-82 Myc LLL42-44LLL expression plasmids (C). Luciferase units were measured, and transcriptionalactivity of G4-p65 TA1 with empty vector was normalized to 1. Activation fold was calculated as the activity of each Ii construct relative tothe activity of empty plasmid. The results shown represent the average of three independent experiments with similar results. (D) B cells fromIi�/� animals were purified and cultured. 48 hr later the cells were transfected with empty expression plasmids (vector plasmid), FL Myc, of1-82 Myc, 1-42 Myc, 1-82 Myc SRG28-30AAA (1-82 28), and 1-82 Myc LLL42-44LLL (1-82 42) expression plasmids. After 48 hr cells wereanalyzed by FACS for IgD expression. The results presented are the mean of three different experiments. The percentage increase of IgD�
cells was calculated by subtracting the percent increase staining of empty expression plasmid and dividing it to the same increase valuemultiplied by 100%. (E) Model for activation of signal transduction through Ii. The 1-82 Ii fragment is formed by a series of cleavage eventsduring the MHC class II biosynthetic and expression pathway. Upon receiving a signal to mature, immature B cells release the 1-42 aminoacid cytosolic domain by cleavage within the transmembrane region. The released cytosolic fragment then induces NF-�B activity eitherdirectly (1) or by inducing another protein or pathway that can induce NF-�B (2).

Immunity558
B cells. This cleavage is essential for NF-�B activation remarkable similarities to the behavior of li. In the caseand B cell differentiation, since the 1-42 fragment by of RIP, the intramembrane cleavage does not take placeitself can induce NF-�B activation and the mutation of until the bulk of the protein on the extracytosolic (lume-amino acids 42-44 (1-82 Myc LLL42-44AAA) completely nal or extracellular) face has been removed by primaryblocks the proteolytic release, NF-�B activation, and B processing events. We have not yet studied thesecell differentiation. Moreover, we have found the RXXL events; however, we believe that some unidentifiedmotif in the released fragment, which was characterized bands recognized by the IN1 antibody in our studiespreviously as a degradation signal. Mutation of this deg- might correspond to these intermediate removal stages.radation signal caused elevation in the appearance of This primary cleavage can occur in the ER, in a postthe released fragment in the cells. Moreover, this mu- ER compartment, or at the cell surface. The cleavagetated RXXL resulted in a dramatic induction of NF-�B shortens the extracytosolic segment to less that 30 aa,activation and B cell differentiation, demonstrating the which is prerequisite for the secondary intramembranerole of this motif in elimination of the active fragment. cleavage.Therefore, Ii is a signaling molecule that requires a pro- Ii is a type II transmembrane protein whose C-terminalcessing step to initiate differentiation of B cells. This domain was shown to be removed by various endocyticsignal is turned off by a degradation process that re- enzymes. We have previously demonstrated that the Iimoves the active protein from the cytoplasm. 1-82 molecule is as active as the full- length protein,
Invariant chain undergoes several proteolytic events indicating that the removal of the lumenal domain doesin the endocytic compartments, resulting in the forma- not interfere with the differentiation activity of Ii (Matzation of a membrane-bound 1-82 truncated protein. It is et al., 2001, 2002). We suggest that the removal of theplausible that these proteolytic cleavages are prerequi- Ii lumenal domain is the first step required for the releasesite for the truncation of the Ii cytosolic domain. There- of the cytosolic fragment. Moreover, amino acid 42 wasfore, the lumenal domain might serve as a negative regu- found to be essential for an intramembrane cleavagelator for the activity of Ii in B cell differentiation. that releases the Ii active fragment and NF-�B activation.Mutations of Ii residues 42-53 were recently shown not The identity of the protease facilitating the proteolysisto interfere with the Ii ER translocation, trimerization, or event and the degradation machinery of the releasedassociation with MHC class II molecules and had no fragment remain to be established. Moreover, we couldobvious effect on endosomal targeting of Ii. Such muta- not yet directly discriminate between the possible directtions did, however, increase the half-life of the molecule role of the cytosolic segment in transcription or, alterna-(Frauwirth and Shastri, 2001), supporting our findings tively, its ability to initiate a cascade in the cytoplasmthat mutation in this region may regulate li cleavage. We or the nucleus resulting in NF-�B induction.therefore believe that the luminal domain of Ii negatively The results presented in this study, as summarized incontrols the truncation of the cytosolic fragment. Figure 7E, suggest that Ii is a signaling molecule that
Quite a few regulatory proteins, including transcrip- induces B cell maturation by the release of its cytosoliction factors, are normally kept in a dormant state and are domain. This fragment induces probably the phosphory-activated by internal or environmental cues. Recently, a lation of the activation domain of p65 member of NF-novel strategy for the mobilization of dormant transcrip- �B and is rapidly degraded. This activity shows a strikingtion factors was discovered that requires proteolytic similarity to that of a family of membrane-bound signal-cleavage. These transcription factors are synthesized ing proteins that liberate their cytosolic domain to trans-initially as inactive, membrane-bound precursors. Once mit a signal that results in the regulation of transcription.triggered, these proteins are cleaved; this is consistentwith several systems, in which it was demonstrated that
Experimental Procedurestransmembrane proteins are cleaved and can migrateinto the nucleus to drive transcription. Two different Cellscleavage mechanisms have been identified, “RIP” (regu- Spleen cells were obtained from various strains of mice at 6–8 weekslated intramembrane proteolysis) and “RUP” (regulated of age as previously described (Shachar et al., 1995). The Animal
Research Committee at the Weizmann Institute approved all animalubiquitin/proteosome-dependent processing). RIP lib-procedures. B cells were enriched as previously described (Flaishonerates a cytosolic fragment that enters the nucleus toet al., 2000).control gene transcription within the plane of the mem-
brane (Hoppe et al., 2001). Examples include membraneConstructs and Molecular Cloningproteins such as Ire1 and ATF6, which release theirDNA constructs encoding Ii fragments and fusion proteins werecytosolic domains in the unfolded protein response inprepared by PCR employing the PfuTurbo DNA polymerase (Stra-the ER (Haze et al., 1999; Niwa et al., 1999), Notch, atagene) and using mouse Ii p31 cDNA (Matza et al., 2001) as a
plasma membrane receptor, whose cytosolic domain is template. The green fluorescent protein (GFP) sequence wasreleased throughout development (Weinmaster, 2000), attached at the N or C termini or to the 1-82 fragment using aand SREBPs (sterol regulatory element binding protein), combination of engineered PCR fragments that ensured readingwhose cytosolic transcription factor domains are liber- frame conservation.
For pEGFP-C1 (Clontech) cloning primer sequences were as fol-ated when cells are deprived of sterol (Brown andlows: 5� primer at position 1 of Ii, 5�-GCTTCGAATTCTATGGATGACGoldstein, 1997).CAACGCGACCTCATC-3�; 3� primer at the last amino acid of Ii (FL),Invariant chain was previously shown to initiate a sig-5�-CGGTGGATCCTCACAGGGTGACTTGACCCAG-3�; 3� primer at
naling cascade that is transmitted to the nucleus activat- position 82 of Ii, 5�-GGTGGATCCCGGAAGCTTCATGCGAAGGCing protein kinases and leading to modulation of either TCT-3�.the p65 activation domain or its coactivators (Matza et For pEGFP-N1 (Clontech) cloning primer sequences were as fol-
lows: 5� primer at position 1 of Ii, 5�-CTTCGAATTCTGATGGATGACCal., 2001). Some common features of RIP have shown

Ii Cleaved Cytosolic Fragment Activates NF-�B559
AACGCGACCTCATC-3�; 3� primer at position 82 of Ii, 5�- CGGTGGA anti-rat IgG (Jackson) or rabbit anti-mouse (Jackson) and peroxi-dase visualization by enhanced chemiluminescence (Amersham).TCCCGCGGAAGCTTCATGCGAAGGCTCT-3�.
For pEF4/Myc-HisA (Invitrogene) cloning primer sequences wereas follows: 5� primer at position 1 of Ii, 5�-GCTCGGATCCGGACAATG Tricine GelsGATGACCAACGCGACCTCAT-3�; 3� primer at the last amino acid of 16% (w/v) Tricine-SDS PAGE were performed as previously de-Ii (FL), 5�-TCTGCAGAATTCCCAGGCCAGAAGATAGGTCTTCCA-3�; scribed (Schagger and von Jagow, 1987).3� primer at position 82 of Ii, 5�-TCTGCAGAATTCTTTCGGAAGCTTCATGCGAAGGCT-3�; 3� primer at position 60 of Ii, 5�-TCTGCAGAAT Cell Lysates for Hot SDS MethodTCGCGGCCCTGTTGCTGGTACAG-3�; 3� primer at position 50 of Ii, Cells were collected 48 hr posttransfection and cell lysates were5�-TCTGCAGAATTCAGTGGTGGCCTGCCCAGCCAA-3�; 3� primer prepared as described previously (Erickson and Blobel, 1979).at position 42 of Ii, 5�-TCTGCAGAATTCCACCAGGACAGAGACACCGGTGTA-3�. Cell Fractionation
For pEF4/Xpress-HisA (Invitrogene) cloning primer sequences The method for membrane and cytosol separation was adoptedwere as follows: 5� primer at position 1 of Ii, 5�-CCTAGGATCCAT from Wang et al. (1994). Transfected 293 cells were disrupted byGATGACCAACGCGACCTCATCTCT-3�; 3� primer at the last amino incubation in 5 vol of low-salt buffer supplemented with proteaseacid of Ii (FL), 5�-TGC AGAATT CCTC ACAGG GTGACTTGACC inhibitors for 30 min, followed by passage 10 times through a 25GCAGTTC-3�. needle. The lysate was centrifuged at 1,000 � g for 10 min to precipi-
For all the plasmids constructed, the PCR products were analyzed tate the nuclei. The supernatant was further centrifuged at 100,000 �by agarose electrophoresis to confirm correct size. The product g for 1 hr to obtain a cytosolic fraction and a membrane pellet. Thiswas purified using Wizard PCR Preps (Promega), digested via pellet was resuspended in a high-salt buffer supplemented withBamHI (NEB) followed by EcoRI (Amersham Pharmacia Biotech), protease inhibitors and centrifuged at 100,000 � g for 30 min toand ligated into the appropriate plasmid vector. obtain a washed membrane pellet, which was then solubilized in 10
Clones were subjected to automated DNA sequencing by stan- mM Tris-HCl (pH 6.8), 0.1 M NaCl, and 1% (v/v) SDS. The cytosoldard protocols using an ABI377 machine (PE Biosystems). The full and the membrane fractions were supplemented with sample buffersequence of all Ii inserts was verified with no errors in the sequence. and boiled for 20 min and equal parts of each fraction were subjected
to immunoblot analysis.
Cell TransfectionFluorescence MicroscopyPrimary B and 293 cells were transfected as was previously de-293 cells were plated on coverslip. After 24 hr, the cells werescribed (Matza et al., 2001). For transfection of 293 cells, 293 cellstransfected as described above. 48 hr later the cells were fixed withwere seeded in a 10 cm2 dish. Transfections were performed using3% PFA and were mounted on glass coverslip with Mowiol 4-88the standard CaPO4 method. A total of 20 �g DNA was used per 10(CalBiochem). B cells were transfected as described above. Thecm2 dish. Cells were collected after 48 hr. Transient transfectionscells were plated on glass coverslips as previously described (Ham-for luciferase assay were performed as described previously (Matzamond et al., 1998). Staining of the endosomes was performed withet al., 2001). Primary B cell transfection: purified Ii�/� B cells wereLysoTracker Red (Molecular Probes) according to the manufactur-incubated with 50 �g/ml LPS from Salmonella typhosa (Sigma). Afterer’s directions.48 hr, the cells were washed with RPMI media and transfected with
TransFast transfection reagent (Promega) 12 �l/4 �g DNA (2 �g IiAcknowledgmentsconstruct � 2 �g empty vector or 4 �g empty vector) according to
the manufacturer’s directions. The cells were collected after 48 hrThe authors gratefully acknowledge members of the Shachar laband analyzed for their cell surface marker expression by FACSfor discussions and comments on this manuscript. This researchanalysis.was supported by The Israel Science Foundation founded by theStaining of the endocytic compartments: endocytic compartmentAcademy of Sciences and Humanities, and the German-Israeli Foun-staining of 293 cells was performed with Lysotracker Red (Moleculardation for Scientific Research and Development. I.S. is the incum-Probes) according to the manufacturer’s instructions.bent of the Alvin and Gertrude Levine Career Development Chair ofCancer Research.
Monitoring NF-�B Activation in a Luciferase AssayThis assay was performed as previously described (Matza et al., Received: April 24, 20022001). Revised: September 26, 2002
Metabolic Radiolabeling and Immunoprecipitation ReferencesFor metabolic labeling, primary B cells (2 � 107 /ml) were starvedfor 1 hr in methionine-free RPMI 1640 (Sigma), followed by a 10 min Agami, R., and Bernards, R. (2000). Distinct initiation and mainte-pulse with 100 �Ci (35 S) methionine (Amersham). The cells were nance mechanisms cooperate to induce G1 cell cycle arrest in re-recultured in full medium for the indicated times. Cell lysates were sponse to DNA damage. Cell 109, 55–66.prepared as described (Shachar et al., 1995). Protein G Sepharose Anderson, K.S., and Cresswell, P. (1994). A role for calnexin (IP90)(25 �l/sample) was conjugated to IN1 antibodies (1:20) for 2 hr (4C) in the assembly of class II MHC molecules. EMBO J. 13, 675–682.followed by 3� PBS washes, and the lysates were immunoprecipi-
Bakke, O., and Dobberstein, B. (1990). MHC class II-associatedtated overnight (4C) with 25 �l conjugate for each sample. The
invariant chain contains a sorting signal for endosomal compart-protein G-bound material was washed 3 times with PBS containing
ments. Cell 63, 707–716.0.1% SDS 0.5% NP-40 and subsequently analyzed by reducing
Brown, M.S., and Goldstein, J.L. (1997). The SREBP pathway: regu-SDS-PAGE. Gels were dried and visualized by phosphorimaginglation of cholesterol metabolism by proteolysis of a membrane-(Fuji).bound transcription factor. Cell 89, 331–340.
Cooper, K.F., Mallory, M.J., Smith, J.B., and Strich, R. (1997). StressPreparation of Cell Extracts and Western Blot Analysis
and developmental regulation of the yeast C-type cyclin Ume3pCells were pelleted and incubated with 50 �g/ml of Digitonin (Sigma)
(Srb11p/Ssn8p). EMBO J. 16, 4665–4675.and the pellet was then lysed as previously described (Shachar et
Cresswell, P. (1994). Assembly, transport, and function of MHC classal., 1995). Lysates were resolved by SDS-PAGE or Tricine gels andII molecules. Annu. Rev. Immunol. 12, 259–293.electroblotted onto nitrocellulose. The blots were blocked with 10%
(v/v) skim milk for 1 hr and then probed for 1 hr with IN1 (anti-Ii Duncan, E.A., Brown, M.S., Goldstein, J.L., and Sakai, J. (1997).Cleavage site for sterol-regulated protease localized to a leu-Serchain cytoplasmic tail mAb) antibodies, anti-GFP (Clontech), anti-
Myc (Invitrogene), or anti-xpress (Invitrogene), followed by washing bond in the lumenal loop of sterol regulatory element-binding pro-tein-2. J. Biol. Chem. 272, 12778–12785.and 1 hr incubation with horseradish peroxidase-conjugated goat

Immunity560
Erickson, A.H., and Blobel, G. (1979). Early events in the biosynthesis chain cytoplasmic tail and transmembrane region determine traffick-ing to an endocytic processing compartment. J. Cell Biol. 126,of the lysosomal enzyme cathepsin D. J. Biol. Chem. 254, 11771–
11774. 317–330.
Pieters, J., Bakke, O., and Dobberstein, B. (1993). The MHC class II-Flaishon, L., Hershkovitz, R., Lantner, F., Lider, O., Alon, R., Levo,associated invariant chain contains two endosomal targeting signalsY., Flavell, R.A., and Shachar, I. (2000). Autocrine secretion of inter-within its cytoplasmic tail. J. Cell Sci. 106, 831–846.feron negatively regulates homing of immature B cells. J. Exp.
Med. 192, 1381–1387. Pond, L., Kuhn, L.A., Teyton, L., Schutze, M.P., Tainer, J.A., Jackson,M.R., and Peterson, P.A. (1995). A role for acidic residues in di-Frauwirth, K., and Shastri, N. (2001). Mutation of the invariant chainleucine motif-based targeting to the endocytic pathway. J. Biol.transmembrane region inhibits II degradation, prolongs associationChem. 270, 19989–19997.with MHC class II, and selectively disrupts antigen presentation.
Cell. Immunol. 209, 97–108. Roche, P.A., and Cresswell, P. (1991). Proteolysis of the class II-associated invariant chain generates a peptide binding site in intra-Hammond, C., Denzin, L.K., Pan, M., Griffith, J.M., Geuze, H.J., andcellular HLA-DR molecules. Proc. Natl. Acad. Sci. USA 88, 3150–Cresswell, P. (1998). The tetraspan protein CD82 is a resident of3154.MHC class II compartments where it associates with HLA-DR, -DM,
and -DO molecules. J. Immunol. 161, 3282–3291. Roche, P.A., Marks, M.S., and Cresswell, P. (1991). Formation of anine-subunit complex by HLA class II glycoproteins and the invariantHarding, C.V., and Unanue, E.R. (1990). Cellular mechanisms of anti-chain. Nature 354, 392–396.gen processing and the function of class I and II major histocompati-
bility complex molecules. Cell Regul. 1, 499–509. Romagnoli, P., Layet, C., Yewdell, J., Bakke, O., and Germain, R.N.(1993). Relationship between invariant chain expression and majorHaze, K., Yoshida, H., Yanagi, H., Yura, T., and Mori, K. (1999).histocompatibility complex class II transport into early and late en-Mammalian transcription factor ATF6 is synthesized as a transmem-docytic compartments. J. Exp. Med. 177, 583–596.brane protein and activated by proteolysis in response to endoplas-
mic reticulum stress. Mol. Biol. Cell 10, 3787–3799. Schagger, H., and von Jagow, G. (1987). Tricine-sodium dodecylsulfate-polyacrylamide gel electrophoresis for the separation of pro-Hoppe, T., Rape, M., and Jentsch, S. (2001). Membrane-bound tran-teins in the range from 1 to 100 kDa. Anal. Biochem. 166, 368–379.scription factors: regulated release by RIP or RUP. Curr. Opin. Cell
Biol. 13, 344–348. Shachar, I., and Flavell, R.A. (1996). Requirement for invariant chainin B cell maturation and function. Science 274, 106–108.Kenty, G., and Bikoff, E.K. (1999). BALB/C invariant chain mutant
mice display relatively efficient maturation of CD4� T cell in the Shachar, I., Elliot, E.A., Chasnoff, B., Grewal, I.S., and Flavell, R.A.periphery and secondary proliferative responses elicited upon pep- (1995). Reconstitution of invariant chain function in transgenic micetide challenge. J. Immunol. 163, 232–242. in vivo by individual p31 and p41 isoform. Immunity 3, 373–383.
Kenty, G., Martin, W.D., Van Kaer, L., and Bikoff, E.K. (1998). MHC Stang, E., and Bakke, O. (1997). MHC class II-associated invariantclass II expression in double mutant mice lacking invariant chain chain-induced enlarged endosomal structures: a morphologicaland DM functions. J. Immunol. 160, 606–614. study. Exp. Cell Res. 235, 79–92.
Lamb, C.A., Yewdell, J.W., Bennink, J.R., and Cresswell, P. (1991). Stumptner-Cuvelette, P., and Benaroch, P. (2002). Multiple roles ofInvariant chain targets HLA class II molecules to acidic endosomes the invariant chain in MHC class II function. Biochim. Biophys. Actacontaining internalized influenza virus. Proc. Natl. Acad. Sci. USA 1542, 1–13.88, 5998–6002. Unanue, E.R. (1984). Antigen-presenting function of the macro-
phage. Annu. Rev. Immunol. 2, 395–428.Lipp, J., and Dobberstein, B. (1986). The membrane-spanning seg-ment of invariant chain (I gamma) contains a potentially cleavable Wang, X., Sato, R., Brown, M.S., Hua, X., and Goldstein, J.L. (1994).signal sequence. Cell 46, 1103–1112. SREBP-1, a membrane-bound transcription factor released by ste-
rol-regulated proteolysis. Cell 77, 53–62.Loder, F., Mutschler, B., Ray, R.J., Paige, C.J., Sideras, P., Torres,R., Lamers, M.C., and Carsetti, R. (1999). B cell development in the Weinmaster, G. (2000). Notch signal transduction: a real rip andspleen takes place in discrete steps and is determined by the quality more. Curr. Opin. Genet. Dev. 10, 363–369.of B cell receptor-derived signals. J. Exp. Med. 190, 75–90. Zur, A., and Brandeis, M. (2001). Securin degradation is mediatedLong, E.O. (1989). Intracellular traffic and antigen processing. Immu- by fzy and fzr, and is required for complete chromatid separationnol. Today 10, 232–234. but not for cytokinesis. EMBO J. 20, 792–801.Lotteau, V., Teyton, L., Peleraux, A., Nilsson, T., Karlsson, L., Schmid,S.L., Quaranta, V., and Peterson, P.A. (1990). Intracellular transportof class II MHC molecules directed by invariant chain. Nature 348,600–605.
Marks, M.S., Blum, J.S., and Cresswell, P. (1990). Invariant chaintrimers are sequestered in the rough endoplasmic reticulum in theabsence of association with HLA class II antigens. J. Cell Biol. 111,839–855.
Matza, D., Wolstein, O., Dikstein, R., and Shachar, I. (2001). Invariantchain induces B cell maturation by activating TAFII105-NF-kB-dependent transcription program. J. Biol. Chem. 276, 27203–27206.
Matza, D., Lantner, D., Bogoch, Y., Flaishon, L., Hershkoviz, R., andShachar, I. (2002). Invariant chain induces B cell maturation in aprocess which is independent of its chaperonic activity. Proc. Natl.Acad. Sci. USA 99, 3018–3023.
Neefjes, J.J., Stollorz, V., Peters, P.J., Geuze, H.J., and Ploegh, H.L.(1990). The biosynthetic pathway of MHC class II but not class Imolecules intersects the endocytic route. Cell 61, 171–183.
Niwa, M., Sidrauski, C., Kaufman, R.J., and Walter, P. (1999). Arole for presenilin-1 in nuclear accumulation of Ire1 fragments andinduction of the mammalian unfolded protein response. Cell 99,691–702.
Odorizzi, C.G., Trowbridge, I.S., Xue, L., Hopkins, C.R., Davis, C.D.,and Collawn, J.F. (1994). Sorting signals in the MHC class II invariant


















