IgG RECEPTOR OSN FOETAL CHICK YOLK SAC · 2005-08-21 · IgG RECEPTOR OSN FOETAL CHICK YOLK SAC...
12
J. Cell Set. 33, 317-328 (T 97 8) 3 x j Printed in Great Britain © Company of Biologists Limited 1978 IgG RECEPTORS ON FOETAL CHICK YOLK SAC CAROL D. LINDEN AND THOMAS F. ROTH Department of Biological Sciences, University of Maryland Baltimore County, Catonsville, Maryland 21228, U.S.A. SUMMARY In the chicken, maternal IgG is deposited in the oocyte yolk and subsequently transported intact by the foetal chick yolk sac into the chick circulation. Such transport appears to require a specific membrane-localized IgG receptor. Using the assay of 125 I-yolk IgG binding, we have investigated the existence and properties of such receptors in yolk sac tissue disks. We find that: (1) There are specific 125 I-IgG binding sites on yolk sac tissue; (2) specific yolk 125 I-IgG binding has a distinct pH optimum at pH 60-6-3; (3) the apparent equilibrium dissociation constant of specific 125 I-IgG binding is 2-3 x io~° M; and (4) heterologous proteins compete less efficiently than yolk IgG for 125 I-IgG binding. INTRODUCTION The transfer of maternal immunoglobulin to the neonate or foetus is essential for postnatal survival. Depending upon the species, maternal immunoglobulins are transferred primarily across the yolk sac, placenta or neonatal gut (Brambell, 1970). Immunological studies have revealed that immunoglobulin G (IgG) is transferred preferentially over other immunoglobulin species, i.e. IgA or IgM (Brambell, 1970). Brambell (1966) proposed a hypothesis, later modified by Wild (1975) to account for the selectivity of IgG transfer from maternal serum across a tissue barrier to the foetus. The modified hypothesis predicts (1) binding of IgG to (Fc) receptors localized in the surface membrane of the cells on the maternal side of the maternofoetal tissue barrier, (2) micropinocytosis of this region to form a vesicle containing IgG bound to its receptor, (3) diffusion (or transport) of the vesicle to the lateral or basal cell surface, and (4) discharge of the vesicle contents into the foetal circulation. Roth & Porter (1964) had demonstrated earlier that specialized 'coated' vesicles were the morphological correlates of protein uptake into developing oocytes. Since coated vesicle, which are distinguished by an electron-dense 'bristle-like' coating on their cytoplasmic surfaces, are found in nearly all eucaryotic cell types, it was proposed that a ubiquitous function of these organelles is transcellular transport of protein molecules. Coated vesicles, or 'micropinocytotic' vesicles, were incorporated into the model as the site of the IgG receptor and the carrier vesicles for protection of IgG (Wild, 1975, 1976). To date, specific IgG receptors have been characterized by studies of radiolabelled- IgG binding to the rabbit yolk sac (Tsay&Schlamowitz, 1975; Sonada & Schlamowitz, 1972; Hemmings, 1974), mouse placenta (Gitlin & Gitlin, 1974), human placenta 21 CEI. 33
Transcript of IgG RECEPTOR OSN FOETAL CHICK YOLK SAC · 2005-08-21 · IgG RECEPTOR OSN FOETAL CHICK YOLK SAC...

J. Cell Set. 33, 317-328 ( T 9 7 8 ) 3 x j
Printed in Great Britain © Company of Biologists Limited 1978
IgG RECEPTORS ON FOETAL CHICK
YOLK SAC
CAROL D. LINDEN AND THOMAS F. ROTHDepartment of Biological Sciences, University of Maryland Baltimore County,Catonsville, Maryland 21228, U.S.A.
SUMMARYIn the chicken, maternal IgG is deposited in the oocyte yolk and subsequently transported
intact by the foetal chick yolk sac into the chick circulation. Such transport appears to requirea specific membrane-localized IgG receptor.
Using the assay of 125I-yolk IgG binding, we have investigated the existence and propertiesof such receptors in yolk sac tissue disks. We find that: (1) There are specific 125I-IgG bindingsites on yolk sac tissue; (2) specific yolk 125I-IgG binding has a distinct pH optimum at pH60-6-3; (3) the apparent equilibrium dissociation constant of specific 125I-IgG binding is2-3 x io~° M; and (4) heterologous proteins compete less efficiently than yolk IgG for 125I-IgGbinding.
INTRODUCTION
The transfer of maternal immunoglobulin to the neonate or foetus is essential forpostnatal survival. Depending upon the species, maternal immunoglobulins aretransferred primarily across the yolk sac, placenta or neonatal gut (Brambell, 1970).Immunological studies have revealed that immunoglobulin G (IgG) is transferredpreferentially over other immunoglobulin species, i.e. IgA or IgM (Brambell, 1970).
Brambell (1966) proposed a hypothesis, later modified by Wild (1975) to account forthe selectivity of IgG transfer from maternal serum across a tissue barrier to the foetus.The modified hypothesis predicts (1) binding of IgG to (Fc) receptors localized in thesurface membrane of the cells on the maternal side of the maternofoetal tissue barrier,(2) micropinocytosis of this region to form a vesicle containing IgG bound to itsreceptor, (3) diffusion (or transport) of the vesicle to the lateral or basal cell surface,and (4) discharge of the vesicle contents into the foetal circulation. Roth & Porter (1964)had demonstrated earlier that specialized 'coated' vesicles were the morphologicalcorrelates of protein uptake into developing oocytes. Since coated vesicle, which aredistinguished by an electron-dense 'bristle-like' coating on their cytoplasmic surfaces,are found in nearly all eucaryotic cell types, it was proposed that a ubiquitous functionof these organelles is transcellular transport of protein molecules. Coated vesicles, or'micropinocytotic' vesicles, were incorporated into the model as the site of the IgGreceptor and the carrier vesicles for protection of IgG (Wild, 1975, 1976).
To date, specific IgG receptors have been characterized by studies of radiolabelled-IgG binding to the rabbit yolk sac (Tsay&Schlamowitz, 1975; Sonada & Schlamowitz,1972; Hemmings, 1974), mouse placenta (Gitlin & Gitlin, 1974), human placenta
21 CEI. 33

3i8 CD. Linden and T. F. Roth
(McNabb, Koh, Dorrington & Painter, 1976; Gitlin & Gitlin, 1974) and neonatalrat gut (Jones & Waldman, 1972). Electron-microscopic studies demonstrated thelocalization of ferritin or horseradish peroxidase-labelled IgG in coated vesicles insome of these tissues (Slade, 1975; Rodewald, 1973; Moxon, Wild & Slade, 1976;Roth, Cutting & Atlas, 1976).
We have investigated the existence of IgG receptors in the embryonic chick yolksac in order to elucidate the initial steps in maternofoetal IgG transport in this system.Previous immunological and biochemical studies showed that hen serum IgG isdeposited in the oocyte yolk and subsequently transported intact across the yolk sacinto the chick circulation (Buxton, 1952; Kramer & Cho, 1970; Holdsworth, Michell& Finean, 1974). Uptake of IgG from the yolk into the chick circulation begins bythe twelfth through fourteenth days of development (Buxton, 1952; Brierly &Hemmings, 1956). Other components of the oocyte yolk, primarily phosvitin andlipovitellin, are digested by enzymes of the yolk sac and are transported as smallmolecule nutrients (Willier, 1968; Lambson, 1970).
We present here initial studies of the properties of 125I-IgG binding to embryonicchick yolk sac tissue. These studies include characterization of the time course, pHdependence, equilibrium parameters and specificity of this binding.
MATERIALS AND METHODS
Carrier-free Na125I was obtained from the Amersham Radiochemical Centre, and [14C]inulin(sp. act. 2'44 mCi/mmol) from New England Nuclear. Cohn fraction V bovine serum albumin(BSA), bovine gamma globulins and phosvitin were from Sigma Co., and ovalbumin fromCalbiochem. Chromatographic resins were obtained from BioRad Laboratories. All otherchemicals were of reagent grade and obtained from commercial sources.
Hanks' balanced salts solution (HBSS) was formulated according to GIBCO and adjustedto pH 6-0-6-2. Bicarbonate and phenol red were omitted, and sodium azide added at a finalconcentration of <VO2%. MES (2-iV-morpholinoethane sulphonic acid) at o-oi M was substi-tuted for the phosphates in some instances. Unless otherwise specified, all buffer solutionscontained 1 mg/ml of BSA. For determination of the pH optimum of binding, all componentsof HBSS except the phosphates were added to 001 M sodium phosphate-citric acid buffersprepared at the pH values indicated.
IgG Purification. An in-house colony of laying hens provided the eggs used as the source ofIgG in these studies. IgG was isolated from yolks by a modification of the method of Bernardi& Cook (i960). Briefly, yolk granules were removed by low speed centrifugation (16000 g for20 min) of yolk diluted with an equal volume of 015 M NaCl. The supernatant was adjusted toi-o M NaCl and centrifuged at 100000 g for 15 to 19 h. The lipid layers resulting from thiscentrifugation were removed and the IgG-rich subnatant and pellet pooled. This crude IgGpreparation was precipitated by the addition of saturated ammonium sulphate (o °C) to a finalconcentration of 50 %. The precipitate was recovered and the preparation subjected to 3 suc-cessive ammonium sulphate precipitations at a final concentration of 33 %. The final precipitatewas collected, dissolved in a minimal volume of 0-15 M NaCl and applied to an agarose Ai '5-mcolumn. The column was eluted with PBS (o-oi M Na phosphate, 015 M NaCl, 0-02 % NaN3
at pH 7-0) and the fractions monitored for protein content by absorbance at 280 nm. Fractionseluting in the position of monomeric chicken IgG with an apparent molecular weight of179000 Daltons were pooled, precipitated at a final concentration of 33 % ammonium sulphateand stored at o °C. Prior to use, the precipitate was resuspended, dialysed against severalchanges of the appropriate experimental buffer and centrifuged at 106000 g for 1 h to removeaggregates. Protein concentration was determined by absorbance at 280 nm using a value

IgG receptors on yolk sac 319
of E^°,= 14. IgG prepared in this manner yielded a single precipitin line in immunodiffusioiiassays against rabbit anti-chicken globulin.
Radioiodination. IgG was iodinated by the procedure of Pressman & Eisin (1950). Typically,10 mg of IgG were exposed to 1-5 mCi of Na125I. After termination of the reaction, 125I-IgGwas separated from the reaction mixture by chromatography on a column of Bio Gel P-10which had been presaturated with unlabelled IgG. Fractions containing the radioactive proteinand eluting in the void volume were pooled and rcchromatographed on a column of agaroseAi'S-m immediately prior to use. Concentration of the fractions containing monomcric126I-IgG was achieved by ultrafiltration. These procedures routinely yielded 125I-IgG pre-parations with specific activities of approximately io8 cpm per mg.
Preparation of yolk sac disks. Fertile eggs aged 12-14 days were kindly provided by Dr M.O'Neill and used as the source of yolk sac tissue. The yolk sac was dissected free of the embryoand overlying membranes, drained of yolk and washed extensively in HBSS minus BSA at pH7-4 until microscopic examination showed no large bodies of yolk adherent to the tissue. Thetissue was further washed in HBSS minus BSA at pH 62 and subjected to mild fixation. Thefixation protocol was as follows: (1) 20 min incubation at o °C in 6 % freshly prepared formalde-hyde (in HBSS minus BSA at pH 62); (2) 10 min incubation at o °C in HBSS minus BSA;(3) two 10-min incubations at room temperature in HBSS (minus BSA) containing o-i Mglycine; and (4) 10 min incubation at o °C in HBSS containing BSA. Tissue disks for assaywere then cut with a no. 3 cork borer (7 mm dia.) from rugose areas of the yolk sac, blotted onpaper towelling and weighed in tared Siliclad-coated tubes containing 50—100/d of buffer.For use in an unfixed state, tissue was washed at pH 7-4, washed in HBSS plus BSA at pH 62,cut and weighed as just described.
Direct-binding studies. Siliclad-coated glass tubes, 12 x 75 mm, were used for all incubations.The final volume of the incubation mixtures was 250 fi\. Individual yolk sac disks were incubatedwith occasional agitation at 0-4 °C for the times indicated in each experiment. The incubationswere terminated by removal of the radioactive incubation mixture and addition of 4 ml of coldHBSS. The disks were then manually transferred to fresh tubes and washed with another 4 mlof buffer. This transfer and wash were repeated for a total of 4 washes, and the disks blottedand placed in counting tubes for determination of radioactivity by gamma counting. For calcu-lation of 125I-IgG bound, counts were normalized to the weight of each yolk sac disk.
In the direct-binding studies, specific 125I-IgG binding was defined as that which was dis-placed by a 30-50-fold molar excess of unlabelled yolk IgG, an amount sufficient to displaceapproximately 95 % of the IgG specifically bound. Each experiment was conducted usingparallel sets of incubations, the first containing only 125I-IgG plus yolk sac disk and the secondcontaining 125I-IgG, yolk sac disk and unlabelled IgG. The bound counts resulting from thefirst set of tubes represent total 125!-IgG binding, the counts in the second set represent non-specific 125I-IgG binding. By subtracting the non-specific from the total counts bound wearrived at values for 125I-IgG bound specifically.
The effect of the pH of the incubation mixture on specific 125I-IgG binding was studiedusing this protocol. The unlabelled IgG was dialysed before use against the buffers at each pHvalue to be tested, and all washes were performed with buffer of the appropriate pH. Yolk sacdisks used in these experiments were washed after fixation in 0-15 M NaCl, 0001 M phosphateat pH 6-o in order to minimize the possible effect of carryover of HBSS containing a higherbuffer concentration into the incubation mixtures.
Specific binding as a function of time was examined in parallel sets of assays where the incu-bations were terminated as described after 0-5, 1, 2 or 3 h.
Determination of entrapped volume using [^Cynulin
Sets of yolk sac disks were incubated at 0-4 °C with 100000 dpm of [14C]inulin for varioustimes up to 2 h. At each time point the disks were washed rapidly (four 4-ml buffer washes inless than 5 min) and their [14C]inulin content measured after dissolving the disks in NCSTissue Solubilizer (Amersham/Searle). The entrapped volume of an 'average' yolk sac diskwas calculated from the [14C]inulin retained by the disks after 2 h, a time equal to the usualincubation time for 125I-IgG plus yolk sac disks. The rate of loss of [14C]inulin from the diskswas determined using a set of yolk sac disks which had been incubated for 2 h and washed

320 C. D. Linden and T. F. Roth
rapidly. This set was washed further by sequential transfer of the disks into I ml of buffer andincubation for various times up to i h. The [14C]inulin content of each i ml of buffer was deter-mined and plotted as a function of time.
Eltttion studies. Yolk sac disks were incubated with io~6 M 125I-IgG for 3 h, washed once inthe incubation tube with 3 ml of buffer, then washed for 6-8 30-min periods in fresh tubescontaining 1 ml of buffer. After an overnight incubation in 1 ml of buffer, the disks were sub-jected to an additional 5-7 i-ml washes prior to beginning the washes with buffer containingthe materials to be tested for their ability to specifically elute 125I-IgG. Yolk IgG, phosvitin,ovalbumin and bovine gamma globulins were each tested at a concentration of io~4 M in aseries of ten 30-min incubations of 1 ml each. All wash and elution tubes were counted for125I-IgG content.
In studies where the effect of pH was compared to the effect of unlabelled IgG in elution of125I-lgG, yolk sac disks were washed as above prior to elution for 8 30-min periods with IgGor buffer at pH 6'8. In one set of elutions IgG was used in the first 8 elutions followed by aseries of 8 washes with buffer alone at pH 6-8. In a reciprocal set of elutions the pH 6-8 bufferwas used first followed by the IgG. Phosphate was used as the buffering species in this experi-ment.
RESULTS
Direct-binding studies
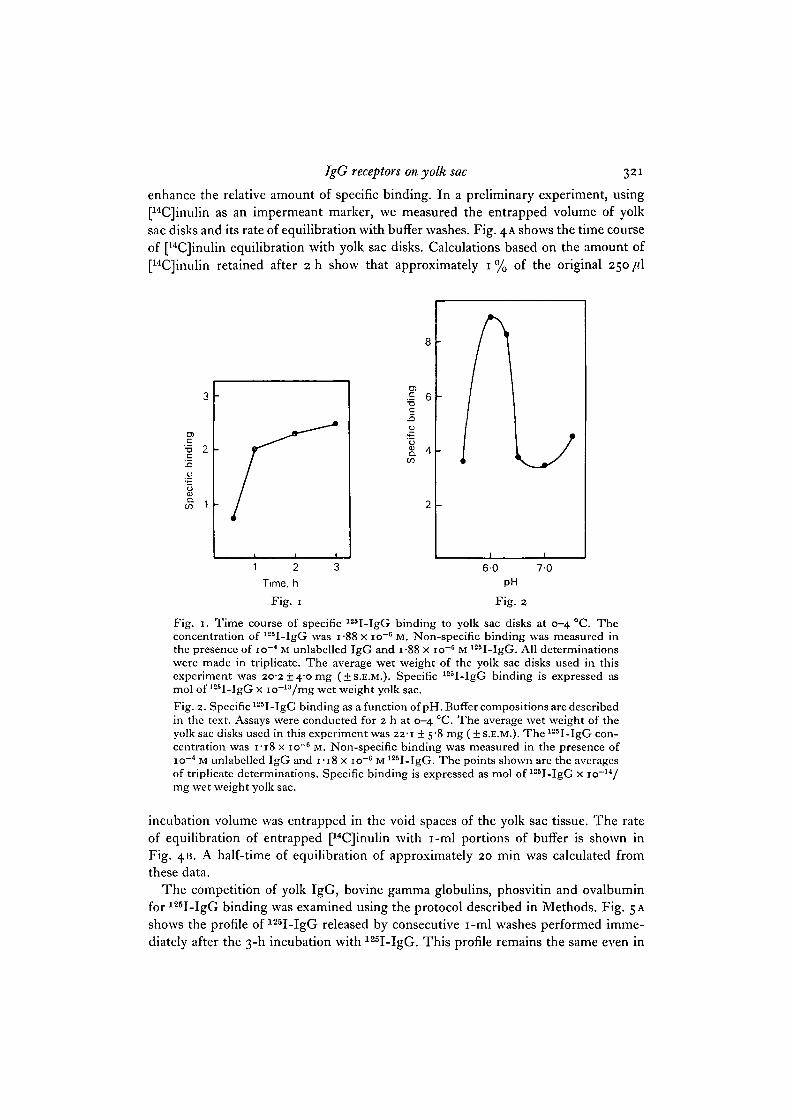
The time course of specific binding at io~G M 125I-IgG is shown in Fig. 1. Thereis a rapid and linear increase in specific binding during the first hour, followed by amore gradual increase during the following 2 h. We consistently observed 20-30%less specific binding at 22-26 °C than at 0-4 °C. Thus all experiments were conductedat 0-4 °C in order to maximize specific binding, inhibit phagocytosis and minimizeendogenous enzymic activities which would be enhanced at higher temperatures. A2-h period was chosen for a routine and convenient assay time. Although unfixedtissue gave qualitatively similar results to those obtained with fixed tissue, the latterappeared to bind slightly more IgG and give more consistent replicate samples. There-fore, fixed tissue was used in all subsequent studies.
Hanks' buffer adjusted to pH 6-o was chosen for use initially on the basis of previousstudies which showed that this is the pH of the yolk contents and also the pH optimumof phosvitin binding to the hen oocyte membrane (Yusko, S. & Roth, T. F., in pre-paration). A study of the pH optimum of 125I-IgG binding to yolk sac tissue confirmedthe validity of this choice. As shown in Fig. 2, the optimum pH for specific IgG bind-ing is pH 6-0-6-3. Binding below pH 6-o and above pH 6-3 is approximately 2-5-foldless than that at optimal pH values. Non-specific binding remained relatively constantover the pH range tested.
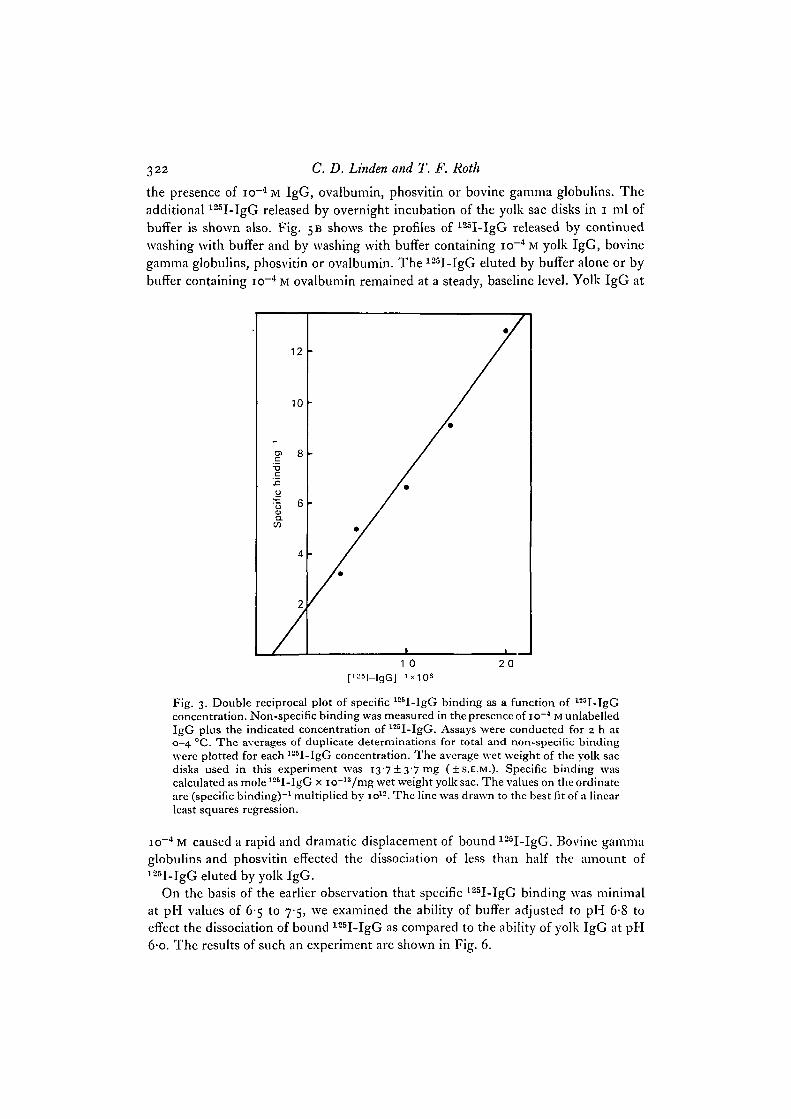
An apparent equilibrium dissociation constant (K^) for specific 125I-IgG bindingwas estimated from a study of binding as a function of 125I-IgG concentration. Suchexperiments yielded a value of 2-3 x io~6 M as determined from a double reciprocalplot of the data (Fig. 3).
Elution studies
Since, in the direct-binding studies, specific 125I-IgG binding accounted for ap-proximately 50% of the total 125I-IgG binding, we designed a protocol to reduce thecontributions of low affinity non-specific binding and entrapped 125I-IgG and thus
IgG receptors on yolk sac 321
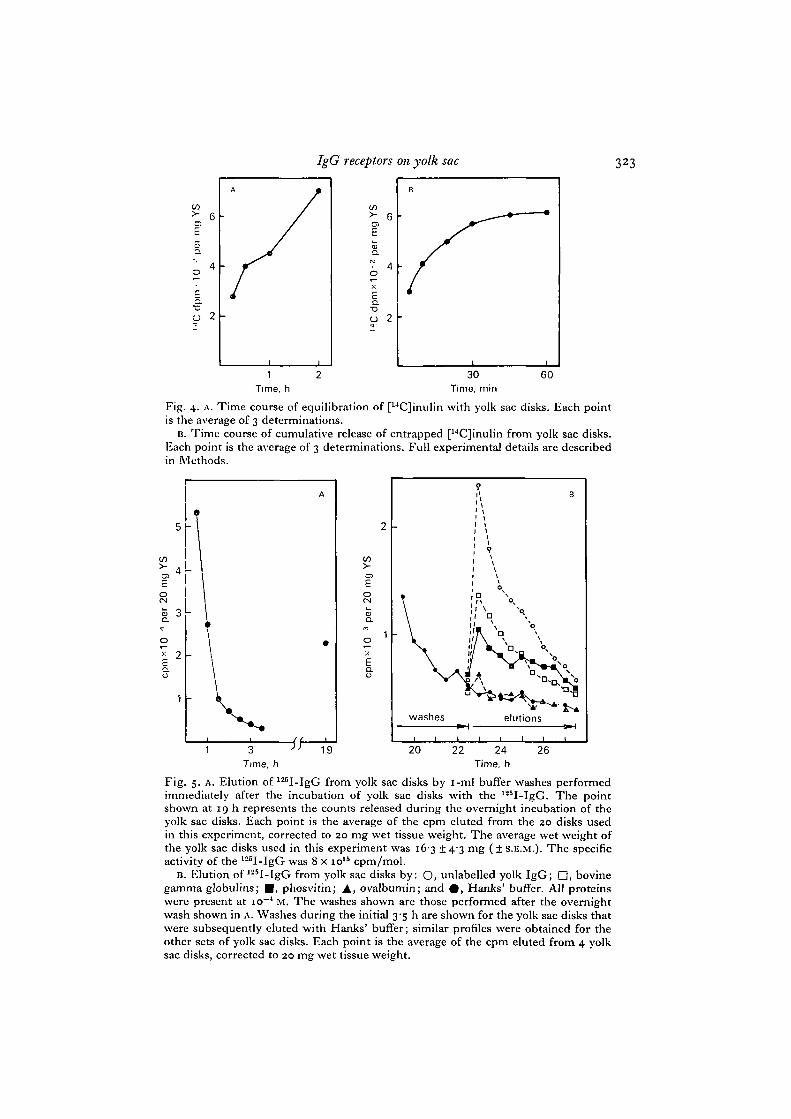
enhance the relative amount of specific binding. In a preliminary experiment, using[14C]inulin as an impermeant marker, we measured the entrapped volume of yolksac disks and its rate of equilibration with buffer washes. Fig. 4 A shows the time courseof [14C]inulin equilibration with yolk sac disks. Calculations based on the amount of[14C]inulin retained after 2h show that approximately 1% of the original 250 /(I
aa.
CO
c
a.CO
4 -
2 -
Fig. 2
Fig. 1. Time course of specific 125I-IgG binding to yolk sac disks at 0-4 °C. Theconcentration of 125I-IgG was i-88 x io~6 M. Non-specific binding was measured inthe presence of io~4 M unlabelled IgG and i-88 x io~" M 125I-IgG. All determinationswere made in triplicate. The average wet weight of the yolk sac disks used in thisexperiment was 20-2 + 40 mg (±S.E.M.). Specific 125I-IgG binding is expressed asmol of 125I-IgG x io~13/mg wet weight yolk sac.Fig. 2. Specific 125I-IgG binding as a function of pH. Buffer compositions are describedin the text. Assays were conducted for 2 h at 0-4 °C. The average wet weight of theyolk sac disks used in this experiment was 22-1 ± 5-8 mg ( + S.E.M.). The125I-IgG con-centration was 1-18 x io~° M. Non-specific binding was measured in the presence ofio~4 M unlabelled IgG and I - I 8 x io~6 M 125I-IgG. The points shown are the averagesof triplicate determinations. Specific binding is expressed as mol of 125I-IgG x io~14/mg wet weight yolk sac.
incubation volume was entrapped in the void spaces of the yolk sac tissue. The rateof equilibration of entrapped [14C]inulin with i-ml portions of buffer is shown inFig. 413. A half-time of equilibration of approximately 20 min was calculated fromthese data.
The competition of yolk IgG, bovine gamma globulins, phosvitin and ovalbuminfor 125I-IgG binding was examined using the protocol described in Methods. Fig. 5 Ashows the profile of 125I-IgG released by consecutive i-ml washes performed imme-diately after the 3-h incubation with 125I-IgG. This profile remains the same even in
3 2 2 C. D. Linden and T. F. Roth
the presence of io~4 M IgG, ovalbumin, phosvitin or bovine gamma globulins. Theadditional 125I-IgG released by overnight incubation of the yolk sac disks in i ml ofbuffer is shown also. Fig. 5B shows the profiles of 125I-IgG released by continuedwashing with buffer and by washing with buffer containing io~4 M yolk IgG, bovinegamma globulins, phosvitin or ovalbumin. The 125I-IgG eluted by buffer alone or bybuffer containing io~4 M ovalbumin remained at a steady, baseline level. Yolk IgG at
cT3C
loo
cif
0)Q.CO
/
12
10
8
6
4
2
/
/
//
//
/
1 0[125!-lgG] 1 *10 6
20
Fig. 3. Double reciprocal plot of specific 125I-IgG binding as a function of l25I-IgGconcentration. Non-specific binding was measured in the presence of io~4 M unlabelledIgG plus the indicated concentration of 125I-IgG. Assays were conducted for 2 h at0—4 CC. The averages of duplicate determinations for total and non-specific bindingwere plotted for each 125I-IgG concentration. The average wet weight of the yolk sacdisks used in this experiment was I 3 7 ± 3 7 m g ( + S.E.M.). Specific binding wascalculated as mole 125I-IgG x io~13/mg wet weight yolk sac. The values on the ordinateare (specific binding)"1 multiplied by io12. The line was drawn to the best fit of a linearleast squares regression.
io~4 M caused a rapid and dramatic displacement of bound 125I-IgG. Bovine gammaglobulins and phosvitin effected the dissociation of less than half the amount of125I-IgG eluted by yolk IgG.
On the basis of the earlier observation that specific 125I-IgG binding was minimalat pH values of 6-5 to 7-5, we examined the ability of buffer adjusted to pH 6-8 toeffect the dissociation of bound 125I-IgG as compared to the ability of yolk IgG at pH6-o. The results of such an experiment are shown in Fig. 6.
IgG receptors on yolk sac 32-
1Time, h
60
Fig. 4. A. Time course of equilibration of [14C]inulin with yolk sac disks. Each pointis the average of 3 determinations.
B. Time course of cumulative release of entrapped [14C]inulin from yolk sac disks.Each point is the average of 3 determinations. Full experimentaJ details are describedin Methods.
CO>
a.
washes elutions
20 22 24Time, h
26
Fig. 5. A. Elution of 125I-IgG from yolk sac disks by i-ml buffer washes performedimmediately after the incubation of yolk sac disks with the 125I-IgG. The pointshown at 19 h represents the counts released during the overnight incubation of theyolk sac disks. Each point is the average of the cpm eluted from the 20 disks usedin this experiment, corrected to 20 mg wet tissue weight. The average wet weight ofthe yolk sac disks used in this experiment was 16-3 ±4'3 mg (±S.E.M.). The specificactivity of the 125I-IgG was 8 x io15 cpm/mol.
B. Elution of 125I-lgG from yolk sac disks by: O, unlabelled yolk IgG; D, bovinegamma globulins; • , phosvitin; A, ovalbumin; and 0 , Hanks' buffer. All proteinswere present at io~4 M. The washes shown are those performed after the overnightwash shown in A. Washes during the initial 3'5 h are shown for the yolk sac disks thatwere subsequently eluted with Hanks' buffer; similar profiles were obtained for theother sets of yolk sac disks. Each point is the average of the cpm eluted from 4 yolksac disks, corrected to 20 mg wet tissue weight.
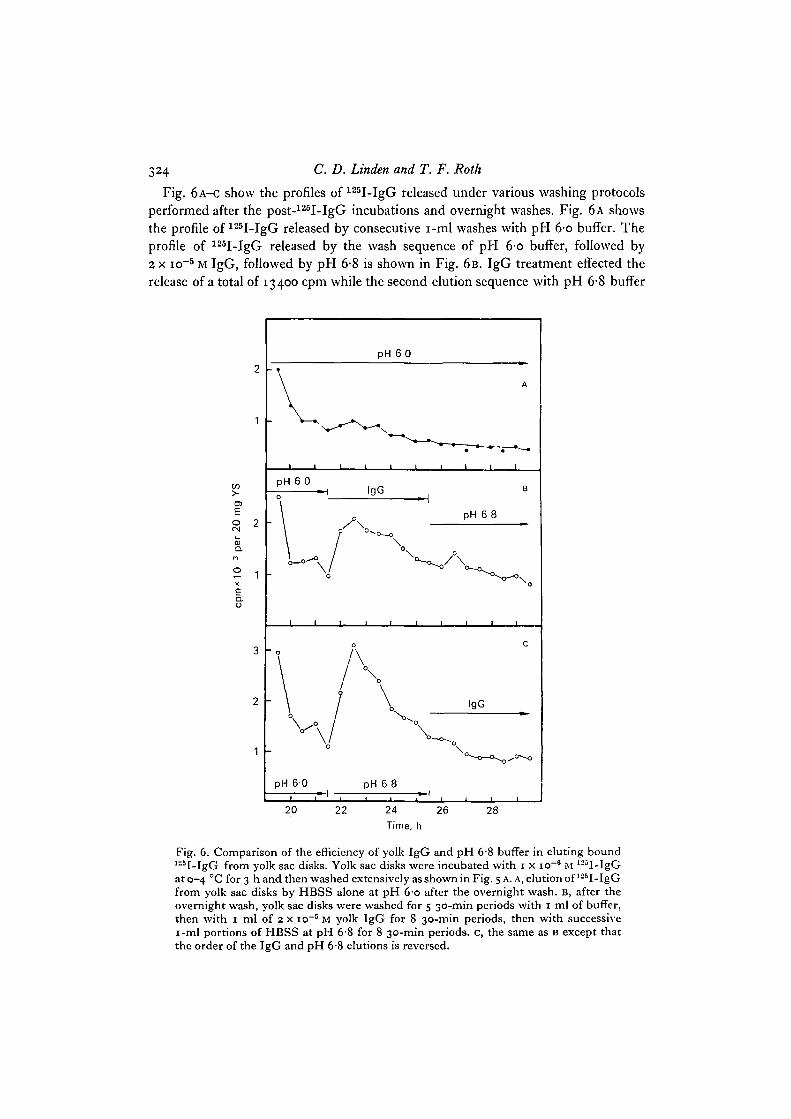
324 C. D. Linden and T. F. Roth
Fig. 6A-C show the profiles of 125I-IgG released under various washing protocolsperformed after the post-125I-IgG incubations and overnight washes. Fig. 6 A showsthe profile of 125I-IgG released by consecutive i-ml washes with pH 6-o buffer. Theprofile of 125I-IgG released by the wash sequence of pH 6-o buffer, followed by2 x io~5 M IgG, followed by pH 6-8 is shown in Fig. 6B. IgG treatment effected therelease of a total of 13400 cpm while the second elution sequence with pH 6-8 buffer
t/5
2 1X
a.o
2 -
1 -
pH 6 0
-I L
pH 6 0
IgG
V
pH 6 0 pH 6 8
20 22 24 26Time, h
28
Fig. 6. Comparison of the efficiency of yolk IgG and pH 6-8 buffer in eluting bound125I-IgG from yolk sac disks. Yolk sac disks were incubated with 1 x io"8 M 125I-IgGat 0-4 CC for 3 h and then washed extensively as shown in Fig. 5 A. A, elution of 125I-IgGfrom yolk sac disks by HBSS alone at pH 6-o after the overnight wash. B, after theovernight wash, yolk sac disks were washed for 5 30-min periods with 1 ml of buffer,then with 1 ml of 2 x io~5 M yolk IgG for 8 30-min periods, then with successivei-ml portions of HBSS at pH 6'8 for 8 30-min periods, c, the same as B except thatthe order of the IgG and pH 6-8 elutions is reversed.

IgG receptors on yolk sac 325
released 8500 cpm. The shape of the profile suggests that the125I-IgG released duringthe washes with pH 6-8 buffer resulted from the inherent slow and continuous releaseof 125I-IgG seen in the buffer control (Fig. 6 A). Fig. 6 c shows the profile of 125I-IgGreleased in the reciprocal elution sequences: pH 6-o, pH 6-8, 2 x io~5 M IgG. Bufferat pH 6-8 effected the release of 16700 cpm while subsequent elution with IgG released7600 cpm. Again, following the peak of 125I-IgG released by the first treatment, inthis case, pH 6-8 buffer, the profile of 125I-IgG released during the second series ofwashes in 2 x io~5 M IgG fell to a level similar to that seen in the pH 6-o buffer con-trol. The similarity in the amounts of 125I-IgG released by IgG at pH 6-o and bufferat pH 6-8, and the lack of elution of 'peaks' of 125I-IgG when IgG followed pH 6-8and vice versa, suggest that the same population of 12sI-IgG is released under these 2conditions.
DISCUSSION
On the basis of the data presented here, we postulate that there are specific IgGreceptors on embryonic chick yolk sac tissue. Identifying and characterizing such areceptor is the first step in determining whether or not a correlation exists betweenIgG binding to a membrane localized receptor and subsequent transport of IgG incoated vesicles. Thus far, the most convincing evidence of such a correlation comesfrom the LDL receptor system, where binding of LDL or cholesterol-LDL complexeshas been correlated with localization of the receptor in coated regions of the fibroblastcell membrane and uptake of the receptor-LDL complex in coated vesicles (Anderson,Goldstein & Brown, 1976). In addition, uptake of yolk proteins into developing oocytes(Roth et al. 1976) and of IgG by the absorptive epithelial cells of the neonatal ratintestine (Rodewald, 1973) has been localized to coated pits and coated vesicles. Inthe rabbit yolk sac, the reaction product of rabbit anti-horseradish peroxidase IgGwith horseradish peroxidase was localized in coated vesicles (Moxon et al. 1976;Slade, 1975).
While the results of the direct-binding studies allowed the preliminary characteriza-tion of the IgG receptor with respect to time course of binding, pH optimum andapparent Kd, the specific elution studies revealed most clearly that there is a compo-nent of specific IgG binding to the yolk sac tissue. This specificity was demonstratedby the relative efficiencies of yolk IgG, bovine gamma globulins, ovalbumin andphosvitin in eluting bound 125I-IgG from washed yolk sac disks. Yolk IgG wasover twice as effective as bovine gamma globulins and phosvitin, while ovalbuminwas completely ineffective. These findings are consistent with the properties of IgGreceptors in other maternofoetal transport systems, where heterologous IgGs maycompete to a certain extent with the homologous molecule for transport and/orbinding, and the homologous albumin has little or no effect in such competition (Wild,1971; Sonada & Schlamowitz, 1972; Jones & Waldman, 1972; Wild, Stauber &Slade, 1972; Rodewald, 1973; Hemmings, 1974; Gitlin & Gitlin, 1974; Hemmings,Jones & Faulk, 1975). The extent of phosvitin competition for 125I-IgG binding wassomewhat puzzling. Phosvitin is a yolk protein which contains 10% phosphoserine

326 C. D. Linden and T. F. Roth
and is highly charged (Clark, 1970, 1973; Joubert & Cook, 1958; Mecham & Olcott,1949). One possible explanation for its efficiency in displacing bound 125I-IgG is thatit interacts electrostatically with the receptor (Hillman, Schlamowitz & Shaw, 1977).However, in the oocyte yolk, phosvitin is found in yolk granules and not in the bulkfluid phase which contains IgG. Thus, its apparent competition for IgG binding invitro may not be important in vivo.
In addition to the component of specific IgG binding, low affinity nonspecificl25I-IgG binding and entrapped 125I-IgG contribute to the values obtained for totalbinding in the direct-binding studies. The amount of 125I-IgG arising from these 2sources was minimized by the extensive washing protocol used in the elution studies.Additional evidence that the low-affinity binding component lacks the specificityexpected of a receptor comes from earlier studies where we examined the dissociationrate of bound 125I-IgG at early times and under a variety of conditions. Immediatelyafter the initial incubation of yolk sac disks with 125I-IgG, elution of 125I-IgG in thepresence of buffer alone, or excess IgG, bovine gamma globulins, phosvitin and oval-bumin was monitored for 3 h. No differences in the dissociation rates of 125I-IgGwere observed using these 5 variables, and all results were similar to those obtainedby washing with buffer alone. If this low-affinity binding component were specific,a sharp difference in the dissociation rate of 125I-IgG in the presence of IgG versusthat in buffer alone should have been observed.
As shown in both the direct-binding studies and in the elution studies, IgG bindingto yolk sac disks displays a distinct pH optimum. The results of the elution studiessuggest also that manipulation of pH may be a useful tool in distinguishing specificfrom non-specific 125I-IgG binding. The pH dependence of IgG binding in thissystem is consistent with the phenomenon observed in some other maternofoetal IgGtransport systems. In the neonatal rat gut, for example, binding of IgG-horseradishpcroxidase conjugates to the intestinal epithelium occurred only at pH 6-0, the physio-logical pH of the proximal region which contains the absorptive cells, and not at pH7-4 (Rodewald, 1976). Since the transported IgG in all maternofoetal transport sys-tems is discharged into the foetal or neonatal blood where the pH is about 7-4, thein vitro data not only correlate with the physiology of the system but suggest a rea-sonable mechanism for the dissociation of IgG from its receptor at the discharge site(Rodewald, 1976; Jones & Waldman, 1972). It is likely that a similar mechanismoperates in this tissue, where the low pH of the yolk (pH 6) permits loading of thereceptor with IgG, and the pH of the plasma (pH 7-4) at the basal and lateral aspectsof the cell permits dissociation of the IgG-receptor complex and discharge of the IgGinto the circulation.
Since, in vivo, the chick yolk sac is exposed to the yolk (and thus, to IgG) only at theendodermal cell surface, one would predict that the bulk of the IgG receptors areassociated with the endodermal cell plasma membrane. Other investigators haveshown that this is indeed the site of the rabbit yolk sac IgG receptor (Wild & Dawson,1977; Sonada & Schlamowitz, 1972; Schlamowitz, Hillman, Lichtiger & Ahearn,1975). We have demonstrated that the IgG-binding properties of the chick yolk sacare quite similar to those of the rabbit yolk sac and the neonatal rat gut. Although these

IgG receptors on yolk sac 327
experiments do not exclude the possibility of IgG binding to mesothelial cells, itseems likely that the IgG receptor is associated with the endodermal cell surface.
Experiments designed to visualize the cellular localization of the chick yolk sac IgGreceptor are in progress.
This work was supported in part by grants HD11519 and HD09549 from the NationalInstitute of Child Health and Human Development, U.S. Public Health Service.
REFERENCES
ANDERSON, R. G. W., GOLDSTEIN, J. L. & BROWN, M. S. (1976). Localization of low densitylipoprotein receptors on plasma membrane of normal human fibroblasts and their absence incells from a familial hypercholesterolemia homozygote. Proc. natn. Acad. Sci. U.S.A. 73,2434-2438.
BERNARDI, G. & COOK, W. H. (i960). An electrophoretic and ultracentrifugal study on theproteins of the high density fraction of egg yolk. Biochim. biophys. Ada 44, 86-96.
BRAMBELL, F. W. R. (1966). The transmission of immunity from mother to young and thecatabolism of immunoglobulins. Lancet ii, 1087-1093.
BRAMBELL, F. W. R. (1970). The transmission of passive immunity from mother to young.Frontiers of Biology, vol. 18. Amsterdam: North-Holland.
BRIERLY, J. & HEMMINGS, W. A. (1956). The selective transport of antibodies from the yolksac to the circulation of the chick. J. Embryol. exp. Morph. 4, 34-41.
BUXTON, A. (1952). On the transference of bacterial antibodies from the hen to the chick. J. gen.Microbiol. 7, 268-286.
CLARK, R. C. (1970). The isolation and composition of two phosphoproteins from hen's egg.Biochem. J. 118, 537-542.
CLARK, R. C. (1973). Amino acid sequence of a cyanogen bromide cleavage peptide from hensegg phosvitin. Biochim. biophys. Ada 310, 174—187.
GITLIN, J. D. & GITLIN, D. (1974). Protein binding by specific receptors on human placenta,murine placenta, and suckling murine intestine in relation to protein transport across thesetissues. J. din. Invest. 54, 1155-1166.
HEMMINGS, W. A. (1974). Transport of the subclasses of human IgG across the yolk-sac ofthe foetal rabbit. Immunology 27, 693-697.
HEMMINGS, W. A., JONES, R. E. & FAULK, W. P. (1975). Transport across the rabbit foetalyolk-sac of fractions of IgG from several mammalian species. Immunology 28, 411-418.
HILLMAN, K., SCHLAMOWITZ, M. & SHAW, A. R. (1977). Characterization of IgG receptorsof the fetal rabbit yolk sac membrane: localization to subcellular fraction and effects ofchemical agents and enzymes on binding. J. Jmmun. 118, 782-788.
HOLDSWORTH, G., MICHELL, R. H. & FINEAN, J. B. (1974). Transfer of very low density lipo-protein from hen plasma into egg yolk. FEBS Letters, Amsterdam 39, 275—277.
JONES, E. A. & WALDMANN, T. A. (1972). The mechanism of intestinal uptake and transcellulartransport of IgG in the neonatal rat. J. clin. Invest. 51, 2916-2927.
JOUHERT, F. J. & COOK, W. H. (1958). Preparation and characterization of phosvitin from henegg yolk. Can.J. Biochem. Physiol. 36, 399-408.
KRAMER, T. T. & CHO, H. C. (1970). Transfer of immunoglobulins and antibodies in the hen'segg. Immunology 19, 157-167.
LAMBSON, R. O. (1970). An electron microscopic study of the entodermal cells of the yolk sacof the chick during incubation and after hatching. Am.jf. Anat. 129, 1-20.
MCNABB, T., KOH, T. Y., DORRINGTON, K. J. & PAINTER, R. H. (1976). Structure and func-tion of immunoglobulin domains v. binding of immunoglobulin G and fragments to placentalmembrane preparations.^. Itnmun. 117, 882-888.
MECHAM, D. K. & OLCOTT, H. S. (1949). Phosvitin, the principal phosphoprotein of egg yolk.J. Am. client. Soc. 71, 3670-3679.
MOXON, L. A., WILD, A. E. & SLADE, B. S. (1976). Localization of proteins in coated micro-pinocytotic vesicles during transport across rabbit yolk sac endoderm. Cell Tiss. Res. 171,175-193-

328 C. D. Linden and T. F. Roth
PRESSMAN, D. & EISIN, H. N. (1950). The zone localization of antibodies V. An attempt tosaturate antibody-binding sites in mouse kidney. J. Immiin. 64, 273-279.
RODEWALD, R. (1973). Intestinal transport of antibodies in the newborn rat. J. Cell Biol. 58,189-211.
RODEWALD, R. (1976). pH-dependent binding of immunoglobulins to intestinal cells of theneonatal rat. J. Cell Biol. 71, 666-670.
ROTH, T. F., CUTTING, J. A. & ATLAS, S. B. (1976). Protein transport: A selective membranemechanism. J. supramolec. Struct. 4, 527-548.
ROTH, T. F. & PORTER, K. R. (1964). Yolk protein uptake in the oocyte of the mosquito Aedesaegypti L,.jf. Cell Biol. 20, 313-332.
SLADE, B. (1975). The role of coated micropinocytosis vesicles in the transport of homologousimmunoglobulin across the rabbit yolk sac endoderm. IRCS Medical Science 3, 235.
SCHLAMOWITZ, M., HILLMAN, K., LICHTIGER, B. & AHEARN, M. J. (1975). Preparation of IgG-binding membrane vesicles from the microvillar brush border of the fetal rabbit yolk sac.J. Immun. 115, 296-302.
SONADA, S. & SCHLAMOWITZ, M. (1972). Specificity of the in vitro binding of IgG and rabbitserum albumin (RSA) to rabbit yolk sac membrane. J. Immun. 108, 1345-1352.
TSAY, D. D. & SCHLAMOWITZ, M. (1975). Comparison of the binding affinities of rabbit IgGfractions to the rabbit fetal yolk sac membrane: Use of 22Na to facilitate quantitation of126I-IgG binding. J. Immun. 115, 939-942.
WILD, A. E. (1971). Transmission of proteins from mother to conceptus in the grey squirrel.Immunology 20, 789-797.
WILD, A. E. (1975). Role of the cell surface in selection during transport of proteins from motherto foetus and newly born. Phil. Trans. R. Soc. Ser. B 271, 395-410.
WILD, A. E. (1976). Mechanism of protein transport across the rabbit yolk sac endoderm. InMaternofoetal Transmission of Immunoglobulins (ed. W. A. Hemmings), pp. 155-167. Londonand New York: Cambridge University Press.
WILD, A. E. & DAWSON, P. (1977). Evidence for Fc receptors on rabbit yolk sac endoderm.Nature, Lond. 268, 443-445.
WILD, A. E., STAUBER, V. V. & SLADE, B. S. (1972). Simultaneous localization of human-globulin-I125 and ferritin during transport across the rabbit yolk sac splanchnopleur. Z.Zelljorsch. mikrosk. Anat. 123, 168-177.
WILLIER, B. H. (1968). Glycogen synthesis, storage and transport mechanisms in the yolk-sacmembrane of the chick embryo. Wilhelm Roux Arch. EntioMech. Org. 161, 89-117.
{Received 31 January 1978)