
Identification of a yeast artificial chromosome spanning the 8q12 translocation breakpoint in...
6
GENES, CHROMOSOMES & CANCER 17:166171(l996) Identification of a Yeast Artificial Chromosome Spanning the 8q I 2 Translocation Breakpoint in Pleomorphic Adenomas With t(3;8)(p2 I ;q 12) Eva Roijer, Koen Kas, Ingrid Klawitz, Jorn Bullerdiek, Wim Van de Ven, and Goran Stenman Laboratory of Cancer Genetics, Department of Pathology, Goteborg University, Goteborg (E.R., G.S.). and Department of Cell Biology, Faculty of Health Sciences, Linkoping University, Linkoping (G.S.), Sweden: Center for Human Genetics, University of Leuven and Flanders Interuniversity Institute for Biotechnology. Leuven, Belgium (K.K., W.V.d.V.): Center for Human Genetics, Univenrty of Bremen, Bremen, Germany (I.K., J.B.) A subgroup of pleomorphic adenomas of the salivary glands is characterized by translocations involving chromosome 8, with consistent breakpoints at 8q 12. As part of a positional cloning effort to isolate the gene(s) affected by these translocations we now report the mapping of the 8q I 2 breakpoint in two primary pleomorphic adenomas with the recurrent t(3;8)(p2I ;q 12). Yeast artificial chromosome (YAC) clones corresponding to eight different loci in 8ql1-12 were isolated and mapped by fluorescence in situ hybridization (FISH). The t(3;8) breakpoint was mapped within a I Mb region flanked by MOS proximally and by the genetic marker D8S I66 distally. One YAC within this region was shown t o span the t(3;8) breakpoint in two tumors. This YAC will provide an excellent tool for isolating the gene@)at the breakpoint(@in adenomas with t(3;8). Genes Chromosom Cancer /7:/66-/7/ (1996). 0 1996 Wiley-Liss, Inc. INTRODUCTION Pleomorphic adenomas are benign epithelial tu- mors originating from the major and minor salivary glands. Extensive cytogenetic analyses have shown that they are characterized by, in particular, trans- locations involving chromosomes 3, 8, and 12, with consistent breakpoints at 3p21, 8q12, and 12q13-15 (see references in Sandros et al., 1990; Bullerdiek et al., 1993; Stenman et al., 1994). Rearrange- ments of chromosome 8 are most frequent, and are found in approximately 40% of the cases. T h e vast majority of these cases have translocations or inser- tions, although there are also a few cases reported with trisomy 8. T h e distribution of breakpoints on chromosome 8 is clearly non-random with a major cluster of breaks at 8q12 and two minor clusters at 8p12-23 and 8q21-22, respectively. T h e most common chromosome 8 abnormality is a t(3;8) (p21;qlZ) found in approximately 40% of the cases with 8q12 abnormalities. In the remaining cases, the translocation partners are variable. Transloca- tions involving 8q12 are often found as the sole deviations, indicating that they are primary cyto- genetic events of possible pathogenetic impor- tance. As part of a comprehensive search for genes in- volved in tumor-specific chromosome transloca- tions in benign tumors, we recently identified the HMGIC gene as the target for translocations involv- ing 12q13-15 in pleomorphic adenomas and in a variety of benign mesenchymal tumors, including lipomas, uterine leiomyomas, hamartomas of the breast and lung, fibroadenomas of the breast, an- giomyxomas, and endometrial polyps (Ashar et al., 1995; Schoenmakers et al., 1995). Recently, we initiated a positional cloning effort to isolate the gene(s) affected by the translocations involving 8q12 in pleomorphic adenomas. We now report the identification of a yeast artificial chromosome (YAC) spanning the 8q12 breakpoint in two pri- mary adenomas with t(3;8)( p2 1 ;ql2). MATERIALS AND METHODS Screening of YAC Libraries YAC clones were isolated from CEPH human genomic YAC libraries mark 1 and 3 (Albertsen et al., 1990; Chumakov et al., 1992) made available to us by the Centre d’atude du Polymorphisme Hu- main (CEPH). Polymerase chain reaction (PCR) screening was carried out as previously described using the primers listed in Table 1 (Schoenmakers et al., 1994). Contaminating Candida parapsylosis, which was sometimes encountered, was eradicated by adding terbinafin (kindly supplied by Dr. Dieter Romer, Sandoz Pharma L T D , Basel, Switzerland) to the growth medium (final concentration: 2.5 pg/ ml). The isolated YAC clones were characterized Received December 18, 1995; accepted May 31, 1996. Address reprint requests to Dr. Goran Stenman, Laboratory of Cancer Genetics, Department of Pathology, Goteborg University, Sahlgrenska University Hospital, S-413 45 Goteborg, Sweden. 0 1996 Wiley-Liss. Inc.
-
Upload
eva-roeijer -
Category
Documents
-
view
216 -
download
3
Transcript of Identification of a yeast artificial chromosome spanning the 8q12 translocation breakpoint in...

GENES, CHROMOSOMES & CANCER 17:166171( l996)
Identification of a Yeast Artificial Chromosome Spanning the 8q I2 Translocation Breakpoint in Pleomorphic Adenomas With t(3;8)(p2 I ;q 12) Eva Roijer, Koen Kas, Ingrid Klawitz, Jorn Bullerdiek, Wim Van de Ven, and Goran Stenman
Laboratory of Cancer Genetics, Department of Pathology, Goteborg University, Goteborg (E.R., G.S.). and Department of Cell Biology, Faculty of Health Sciences, Linkoping University, Linkoping (G.S.), Sweden: Center for Human Genetics, University of Leuven and Flanders Interuniversity Institute for Biotechnology. Leuven, Belgium (K.K., W.V.d.V.): Center for Human Genetics, Univenrty of Bremen, Bremen, Germany (I.K., J.B.)
A subgroup of pleomorphic adenomas of the salivary glands is characterized by translocations involving chromosome 8, with consistent breakpoints at 8q 12. As part of a positional cloning effort to isolate the gene(s) affected by these translocations we now report the mapping of the 8q I 2 breakpoint in two primary pleomorphic adenomas with the recurrent t(3;8)(p2I ;q 12). Yeast artificial chromosome (YAC) clones corresponding to eight different loci in 8ql1-12 were isolated and mapped by fluorescence in situ hybridization (FISH). The t(3;8) breakpoint was mapped within a I Mb region flanked by MOS proximally and by the genetic marker D8S I66 distally. One YAC within this region was shown to span the t(3;8) breakpoint in two tumors. This YAC will provide an excellent tool for isolating the gene@) at the breakpoint(@ in adenomas with t(3;8). Genes Chromosom Cancer /7 : /66- /7 / (1996). 0 1996 Wiley-Liss, Inc.
INTRODUCTION
Pleomorphic adenomas are benign epithelial tu- mors originating from the major and minor salivary glands. Extensive cytogenetic analyses have shown that they are characterized by, in particular, trans- locations involving chromosomes 3, 8, and 12, with consistent breakpoints at 3p21, 8q12, and 12q13-15 (see references in Sandros et al., 1990; Bullerdiek et al., 1993; Stenman et al., 1994). Rearrange- ments of chromosome 8 are most frequent, and are found in approximately 40% of the cases. T h e vast majority of these cases have translocations or inser- tions, although there are also a few cases reported with trisomy 8. T h e distribution of breakpoints on chromosome 8 is clearly non-random with a major cluster of breaks at 8q12 and two minor clusters at 8p12-23 and 8q21-22, respectively. T h e most common chromosome 8 abnormality is a t(3;8) (p21;qlZ) found in approximately 40% of the cases with 8q12 abnormalities. In the remaining cases, the translocation partners are variable. Transloca- tions involving 8q12 are often found as the sole deviations, indicating that they are primary cyto- genetic events of possible pathogenetic impor- tance.
As part of a comprehensive search for genes in- volved in tumor-specific chromosome transloca- tions in benign tumors, we recently identified the HMGIC gene as the target for translocations involv- ing 12q13-15 in pleomorphic adenomas and in a variety of benign mesenchymal tumors, including lipomas, uterine leiomyomas, hamartomas of the
breast and lung, fibroadenomas of the breast, an- giomyxomas, and endometrial polyps (Ashar et al., 1995; Schoenmakers et al., 1995). Recently, we initiated a positional cloning effort to isolate the gene(s) affected by the translocations involving 8q12 in pleomorphic adenomas. We now report the identification of a yeast artificial chromosome (YAC) spanning the 8q12 breakpoint in two pri- mary adenomas with t(3;8)( p2 1 ;ql2).
MATERIALS AND METHODS
Screening of YAC Libraries
YAC clones were isolated from CEPH human genomic YAC libraries mark 1 and 3 (Albertsen et al., 1990; Chumakov et al., 1992) made available to us by the Centre d’atude du Polymorphisme Hu- main (CEPH). Polymerase chain reaction (PCR) screening was carried out as previously described using the primers listed in Table 1 (Schoenmakers et al., 1994). Contaminating Candida parapsylosis, which was sometimes encountered, was eradicated by adding terbinafin (kindly supplied by Dr. Dieter Romer, Sandoz Pharma LTD, Basel, Switzerland) to the growth medium (final concentration: 2.5 pg/ ml). The isolated YAC clones were characterized
Received December 18, 1995; accepted May 31, 1996. Address reprint requests to Dr. Goran Stenman, Laboratory of
Cancer Genetics, Department of Pathology, Goteborg University, Sahlgrenska University Hospital, S-413 45 Goteborg, Sweden.
0 1996 Wiley-Liss. Inc.

8q12 TRANSLOCATION BREAKPOINT MAPPING 167
TABLE I. Primer Sequences Used for YAC Screening and Sequence Tagged Site (STS) Content Mapping"
Product STS Nucleotide sequence (5'-3') size (bp) Tmn ("C)
LYN GGAAGGAAAGGAAAGGAGA I94 60
MOS GAGCTCAACGTAGCAAGGCT 203 60
D8S I66 GATTGTGTCATTGCACTCCA I I 6 58
D8S I08 CAAACCTTGAA'TTACAAAACAG I I 6 58
PENK TAATAAAGGAGCCAGCTATG 75-83 58
GGTITGGGTGTITGGTGTG
AACTGTCCTCCAGTGCGG
ACAAGG AAGTTCClll lTGG
TGTTAATATTAGACCACCTITC
ACATCTGATGTAAATGCAAGT
AGCTTCAllllTCCCTCTAG
ATGGGTITATGGCCTITAC
085 I65 ACAAGAGCACACATITAGTCAG 138- I 52 58
D8S285 GCATCACACAGAATCmG 108-124 60
~ 8 ~ 2 6 0 AGGCTTGCCAGATAAGGTTG I 87-2 I 3 60 GCTGAAGGCTGTTCTATGGA
'Amplification product sizes and annealing temperatures (T-J are indi- cated.
by STS-PCR and fluorescence in situ hybridization (FISH) analysis.
PCR Analysis
PCR amplification was carried out using a Phar- macia LKB-Gene ATAQ Controller (Pharmacia/ LKB, Uppsala, Sweden) in final volumes of 100 pl containing 10 mM Tris-HC1, pH 8.3, 50 mM KCl, 1.5 mM MgCl,, 0.01% gelatin, 2 mM dNTPs, 20 pmol of each amplimer, 2.5 units of AmpliTaq (Perkin-Elmer Cetus, Nonvalk, CT), and 100 ng (for superpools) or 20 ng (for pools) of DNA. After initial denaturation for 5 min at 94"C, 35 amplifi- cation cycles were performed, each consisting of denaturation for 1 rnin at 94"C, annealing for 1 rnin a t the appropriate temperature (see Table l), and extension for 1 rnin at 72"C, and with a final ex- tension at 72°C for 10 min. Results were evaluated by analysis of 10 p1 of the reaction product on 2% agarose gels.
DNA Preparation
Agarose plugs containing high-molecular-weight yeast and YAC DNA (equivalent to 1 x lo9 cells/ ml) were prepared as described (Schoenmakers et al., 1994) and stored at room temperature in lysis buffer (1% lithium dodecyl sulfate, 100 mM EDTA, 10 mM Tris-HC1, pH 8.0). Plugs were thoroughly dialyzed against 4 changes of 25 ml T,,E, (10 mM Tris-HC1, 1 mM EDTA, pH 8.0) followed by 2 changes of 0.5 ml 1 x restriction buffer before they were subjected to YAC-end res- cue. YAC DNA (100 ng) was amplified by inter-Ah PCR (Pl: CTGCACTCCAGCCTGGG, P2: TC-
CCAAAGTGCTGGGATTACAG). After initial denaturation for 5 min at 94"C, 30 amplification cycles were performed, each consisting of denatur- ation for 1 rnin at 94"C, annealing for 30 sec at 37"C, and extension for 6 min at 72"C, and with a final extension at 72°C for 10 min. Amplified DNA was purified with a QIAQuick PCR purification kit (Qiagen, Chatsworth, CA).
Tumor Material and Cytogenetic Analysis
Fresh tumor specimens were obtained from pa- tients at the time of surgery. Primary cultures were established as described previously (Nordkvist et al., 1994). Chromosome preparations were made from exponentially growing primary cultures. Cells were harvested after Colcemid exposure followed by hypotonic treatment and fixation in methanol: acetic acid. Slides were subsequently G-banded and analyzed according to the recommendations of the ISCN (1991). Two adenomas, CG588 and CG644, showing the recurrent t(3;8)(p21;q12) as the sole anomaly, were selected for translocation breakpoint mapping. Slides for FISH were pre- pared from cells stored in fixative at -20°C.
FISH Analysis
Slides for FISH were aged for 2-10 days, or were treated with 2 X SSC (pH 7.0) at 37°C for 30 min, and subsequently denatured in 70% formamide in 2 x SSC (pH 7.0) at 75°C for 2-6 min. Phage and YAC DNAs (1 pg) were labeled with biotin-16- dUTP (Boehringer Mannheim, Indianapolis, IN) by nick translation. Labeled DNA was purified through a Sephadex G50 column (Pharmacia, Uppsala, Sweden), coprecipitated with 50 pg son- icated human placenta DNA (Sigma, St. Louis, MO), and dissolved in hybridization solution (50% formamide, 2 X SSC, pH 7.0, 50 mM sodium phosphate, pH 7.0, 10% dextran sulfate). Ten mi- croliters of probe mixture [25 ng biotin-labeled YAC probe, 5 ng digoxigenin-labeled alpha-satel- lite probe (D321 and D822, Oncor, Gaithersburg, MD), and if needed 1-10 pg sonicated human pla- centa DNA] was denatured at 75°C for 10 rnin fol- lowed by preannealing for 2 hr at 37°C. Hybridiza- tion was carried out for 16-20 hr at 37°C in a humidified chamber. After hybridization, the slides were washed 3 times in 50% formamide in 2 x SSC (pH 7.0) and 3 times in 2 X SSC (pH 7.0) at 42°C for 5 min each. Slides were subsequently preincubated with 40 p14 x SSC (pH 7.0)/5% non- fat dry milk (Semper AB, Stockholm, Sweden) for

I68 ROIjER ET AL.
10 min at room temperature. Detection of biotin- ylated probes was performed using fluorescein isothiocyanate (F1TC)-conjugated avidin (5 kg/ml; Sigma) followed by an amplification step using bio- tinylated mouse anti-avidin (5 pg/ml; Sigma) and another FITC-avidin step. For detection of digox- igenin-labeled alpha-satellite probes, rhodamine- conjugated sheep anti-digoxigenin (Oncor) was added to the last incubation. All incubations were performed at room temperature for 20 min and were followed by three 5 min washes in 4 x SSC (pH 7.0)/0.05% Tween-20. Chromosomes were counterstained with 4,6-diamino-Z-phenylindole (DAPI). Slides were examined in a Zeiss Axiophot epifluorescence microscope using the appropriate filter combinations. Fluorescence signals were dig- italized, enhanced, and analyzed using the Probe- Master FISH image analysis system (Perceptive Scientific Instruments, Houston, TX). Color prints were produced using a Kodak XL 7700 mono- chrome continuous printer.
RESULTS
Mapping of the Candidate Genes MOS, LYN, and CEBPD in Relation to the t(3;8) Breakpoint
A YAC corresponding to MOS (ICRFy900- G10157) was isolated from the ICRF YAC library no. 900 (Lehrach et al., 1990) using a MOS-specific probe (probe pHMZA, ATCC 41004). This clone also contained the YES-related proto-oncogene LYN. A phage clone (12K1) containing the entire human CEBPD gene was kindly provided by Dr. Cleutjens (Cleutjens et al., 1993). Hybridization of these clones to normal metaphase chromosomes confirmed their localization to 8qll-12 (Fig. 1A,B). CEBPD, which belongs to the family of leucine zipper (bZIP) transcription factors, has previously been mapped to 8 q l l (Cleutjens et al., 1993). However, during the course of our work, Wood et al. (1995) reported that CEBPD colocalizes with FGFRZ in the proximal short arm at 8p11.2 + p l l . 1. Our FISH mapping of CEBPD confirms that the gene indeed maps to the pericentromeric re- gion of chromosome 8. The signal from the gene overlaps with the signal from the alpha-satellite probe (Fig. lA), making it impossible to determine cytologically whether the gene is located in the proximal part of 8p or 8q. FISH mapping of these two clones on translocation chromosomes revealed that MOS, LYN, and CEBPD are proximal to the breakpoints in CG588 and CG644. Both cases showed hybridization signals on the normal 8 as well as on the der(8). Our mapping of LYN close to
MOS in 8q12 confirms the mapping by Corey and Shapiro (1994), who sublocalized LYN to 8qll-12 by FISH. LYN was previously assigned to 8q13 (Drayna and Wood, 1993).
Mapping of the t(3;8) Breakpoint to a I cM Interval Within 8q I2
According to previous radiation hybrid mapping data (Sapru et al., 1994), band 8q12 is flanked by the markers D8S165 and MOS proximally and by D8S260 distally. T h e estimated genetic distance between MOS and D8S260 is 9 cM (Gyapay et al., 1994). T o confirm that the breakpoints are located within this region we isolated two YACs (783H12 and 814A6) corresponding to D8S260. These YACs were shown by FISH to map distal to the t(3;8) breakpoint in both adenomas. T o sublocalize fur- ther the breakpoint region within band 8q12 we isolated YACs corresponding to the genetic mark- ers D8S108 (291D12) and D8S166 (388D6), which are located less than 1 Mb telomeric from MOS (Fig. 2). Both of these YACs mapped distal to the 8q12 breakpoint in both adenomas, demonstrating that the breakpoints are located within a 1 Mb re- gion flanked by MOS and D8S166.
Identification of a YAC That Spans the t(3;8) Breakpoint
T o fine map the breakpoints within the 1 Mb region flanked by MOS and D8S166, we initiated a chromosome walk starting from MOS. Two YACs corresponding to D8S285, a marker located less than 100 kb distal to MOS, were isolated. In addi- tion to D8S285, these YACs were shown to contain MOS and LYN (Y166F4) and D8S165, LYN, and PENK (Y935E9), respectively (Fig. 2). YAC 935E9 was shown to have an internal deletion encompass- ing MOS. FISH analysis using YAC 166F4 demon- strated that this YAC spans the t(3;8) translocation breakpoint in adenomas CG588 and CG644. Hy- bridization signals were observed on the normal 8 as well as on the der(8) and the der(3) in both cases (Fig. 1C). T h e intensity of the hybridization sig- nals on the two derivative chromosomes were equal, indicating that the breakpoints are similar in the two cases. T o confirm these observations the larger YAC 935E9, which completely overlaps with 166F4 (Fig. 2), was also hybridized to tumor meta- phases from CG588. As expected, this YAC also covered the t(3;8) translocation breakpoint.
DISCUSSION
In this paper we demonstrate that the break- points in two pleomorphic adenomas with the re-
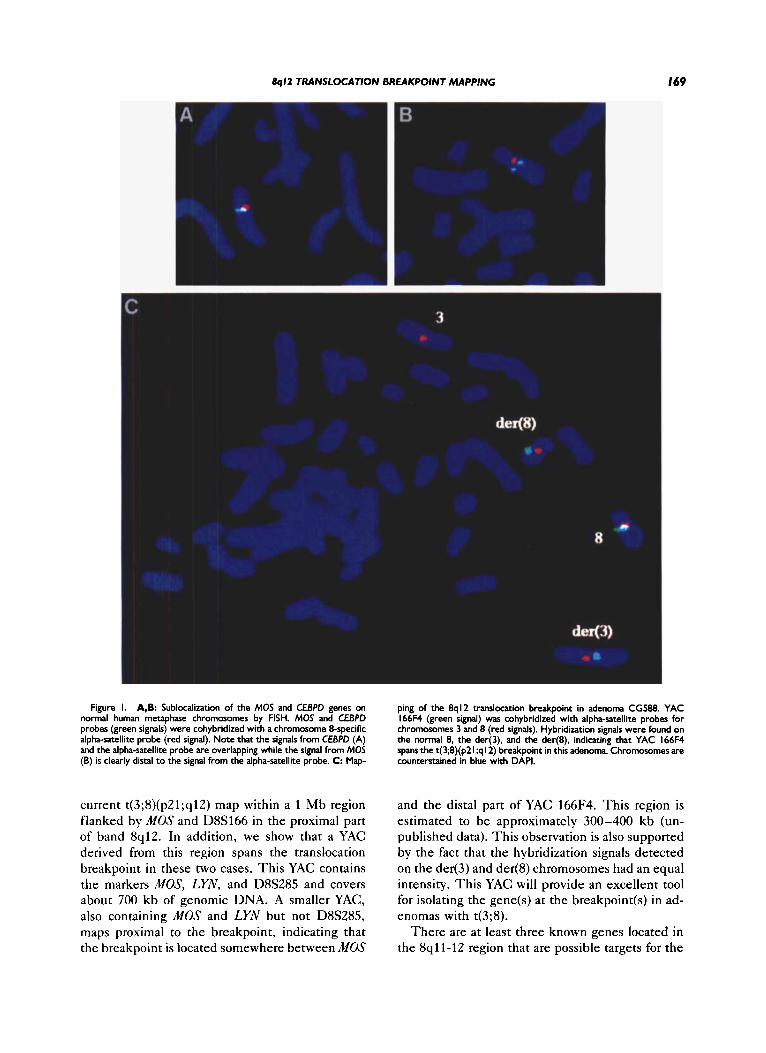
8ql2 TRANSLOCATION BREAKPOINT MAPPING I69
Figure I. A,B: Sublocalization of the MOS and CEBPD genes on normal human metaphase chromosomes by FISH. MOS and CEBPD probes (green signals) were cohybridized with a chromosome 8-specific alpha-satellite probe (red signal). Note that the signals from CEBPD (A) and the alpha-satellite probe are overlapping while the signal from MOS (B) is clearly distal to the signal from the alpha-satellite probe. C: Map-
ping of the 8q12 translocation breakpoint in adenoma CG588. YAC 166F4 (green signal) was cohybridized with alpha-satellite probes for chromosomes 3 and 8 (red signals). Hybridization signals were found on the normal 8, the der(3). and the der(8), indicating that YAC 166F4 spans the t(3;8)(p21;ql2) breakpoint in this adenoma. Chromosomes are counterstained in blue with DAPI.
current t(3;8)(p21;q12) map within a 1 Mb region flanked by MOS and D8S166 in the proximal part of band 8q12. In addition, we show that a YAC derived from this region spans the translocation breakpoint in these two cases. This YAC contains the markers MOS, LYN, and D8S285 and covers about 700 kb of genomic DNA. A smaller YAC, also containing MOS and LYN but not D8S285, maps proximal to the breakpoint, indicating that the breakpoint is located somewhere between MOS
and the distal part of YAC 166F4. This region is estimated to be approximately 300-400 kb (un- published data). This observation is also supported by the fact that the hybridization signals detected on the der(3) and der(8) chromosomes had an equal intensity. This YAC will provide an excellent tool for isolating the gene(s) at the breakpoint(s) in ad- enomas with t(3;8).
There are at least three known genes located in the 8qll-12 region that are possible targets for the

I 70 ROIjER ET AL.
I I I I I I I id cen
388D6 (430 kb) 90091 01 57 1 4 -
291D12 (410 kb) - c - 935E9 (1660 kb)
166F4’ (700 kb)
-
Figure 2. Map of the 8q I2 translocation breakpoint region. Gene symbols, genetic markers, addresses of individual YACs, and their sizes are shown above the lines. The YAC crossing the t(3;8) breakpoint is indicated by an asterisk
8ql2 translocations in pleomorphic adenomas, namely the proto-oncogenes MOS and LYN and the transcription factor gene CEBPD. Our finding that the chromosome 8 breakpoint in pleomorphic ad- enomas with t(3;8) maps close to MOS is of special interest since we have previously observed struc- tural rearrangements of MOS in two adenomas with translocation breakpoints at 8q12 (Stenman et al., 1991). However, in none of those cases did we find any evidence of translocation of MOS, thus further supporting the present observations. Collectively, the present and previous data exclude MOS as well as LYN and CEBPD as the target genes for the 8q12 translocations in pleomorphic adenomas.
Nearly half of the adenomas with translocations involving 8q12 show the typical t(3;8)(pZl;q12). In the remaining cases, the translocation partners are variable. In fact, most human chromosomes have acted as translocation partners with chromosome 8 (see references in Sandros et al., 1990; Bullerdiek et al., 1993). These observations raise the question whether the breakpoints in the variant transloca- tions occur in the same region as those in cases with t(3;8). Analysis of adenomas with different variant translocations involving 8q12 using YAC 166F4 will be useful to elucidate this question.
Interestingly, there is also another type of solid tumor with recurrent, structural rearrangements in- volving the 8qll-13 region, namely the lipoblas- toma (Sandberg et al., 1986; Ohjimi et al., 1992; Fletcher et al., 1993; Dal Cin et al., 1994; Sawyer et al., 1994). Lipoblastomas are benign pediatric tumors composed of lipoblasts in different stages of development (Enzinger and Weiss, 1995). They are often rapidly growing tumors, but have not been reported to metastasize. Among the seven cases so far analyzed cytogenetically, rearrange- ments involving 8qll-13 were observed in six cases. However, in all but two cases the transloca- tion partners were variable. The segments distal to 8qll-13 were translocated or inserted into the fol-
lowing chromosome bands: 3q12, 6p25, 7p22 (two cases), 9p22, and 14q24. A critical question in this context is, of course, whether the chromosome 8 translocations in pleomorphic adenomas and lipo- blastomas involve the same gene. Based on our knowledge about the HMGICgene in 12q15 (Wan- schura et al., 1995) and its involvement in a variety of benign tumors, it is not unreasonable to assume that the 8q12 translocations in pleomorphic ade- nomas and lipoblastomas might affect the same gene. Molecular cloning of the 8q12 translocation breakpoints in these tumors, which is now in progress, will provide the answer to this question.
ACKNOWLEDGMENTS
This work was supported by the Swedish Cancer Society, the IngaBritt and Arne Lundbergs Re- search Foundation, the EC through Biomed 1 pro- gram “Molecular Cytogenetics of Solid Tumours,” the “Geconcerteerde Onderzoekacties 1992- 1996,” the “National Fonds voor Wetenschappel- ijk Onderzoek” (NFWO), the “ASLK-Programma voor Kankeronderzoek,” the Flanders Institute for Biotechnology, and the Deutsche Forschungsge- meinschaft. K.K. is in a postdoctoral program of the NFWO.
REFERENCES Albertsen HM, Aberrahim H, Cann HM, Dausset J, Le Paslier D,
Cohen D (1990) Construction and characterization of a yeast ar- tificial chromosome library containing seven haploid genome equivalents. Proc Natl Acad Sci USA 87:4256-4260.
Ashar HR, Fejzo MS, Tkachenko A, Zhou X, Fletcher JA, Were- mowicz S, Morton CC, Chada K (1995) Disruption of the archi- tectural factor HMGI-C: DNA-binding AT hook motifs fused in lipomas to distinct transcriptional regulatory domains. Cell 8257- 65.
Bullerdiek J, Wobst G, Meyer-Bolte K, Chilla R, Haubrich J, Thode B, Bartnitzke S (1993) Cytogenetic subtyping of 220 sal- ivary gland pleomorphic adenomas: Correlation to occurrence, histological subtype, and in vitro cellular behavior. Cancer Genet Cytogenet 6527-31.
Chumakov I, Rigault P, Guillou S, Ougen P, Billaut A, Guasconi G, Gervy P, LeGall I, Soularue P, Grinas L, Bougueleret L, Bellane- Chantelot C, Lacroix B, Barillot E, Gesnouin P, Pook S, Vaysseix G, Frelat G, Schmitz A, Sambucy J-L, Bosch A, Estivill X, Weis- senbach J, Vignal A, Reithman H, Cox D, Patterson D, Gardiner

8q12 TRANSLOCATION BREAKPOINT MAPPING I71
K, Hattori M, Sakaki Y, Ichikawa H, Ohki M, Le Paslier D, Heilig R, Antonarakis S, Cohen D (1992) Continuum of overlap- ping clones spanning the entire human chromosome Zlq. Nature 359380-387.
Cleutjens CBJM, van Eekelen CCEM, van Dekken H, Smit EME, Hagemeijer A, Wagner MJ, Wells DE, Trapman J (1993) The human ClEBPS (CRP3KELF) gene: Structure and chromosomal localization. Genomics 16:520-523.
Corey SJ, Shapiro DN (1994) Localization of the human gene for src-related protein tyrosine kinase LYN to chromosome 8qll-12: A lymphoid signaling cluster? Leukemia 8~1914-1917.
Dal Cin P, Sciot R, De Wever I, Van Damme B, Van Den Berghe H (1994) New discriminative chromosomal marker in adipose tis- sue tumors. The chromosome 8qll-ql3 region in lipoblastoma. Cancer Genet Cytogenet 78:232-235.
Drayna D, Wood S (1993) Report of the committee on the genetic constitution of chromosome 8. Chromosome Coordinating Meet- ing (1992); Cuticchia AJ, Pearson PL, Klinger HP (eds.): Ge- nome Priority Reports, Vol l . Basel: Karger, pp 318-332.
Enzinger FM, Weiss SW (1995) Soft Tissue Tumors. 3rd Ed. St. Louis: C.V. Mosby, pp 401-405.
Fletcher JA, Kozakewich HP, Schoenberg ML, Morton CC (1993) Cytogenetic findings in pediatric adipose tumors: Consistent re- arrangement of chromosome 8 in lipoblastoma. Genes Chromo- som Cancer 6:24-29.
Gyapay G, Morissette J, Vignal A, Dib C, Fizaes C, Millasseau P, Marc S, BernaMi G, Lathrop M, Weissenbach J (1994) The 1993-94 Gtntthon human genetic linkage map. Nat Genet 7: 246-339.
ISCN (1991) Mitelman F (ed): Guidelines for Cancer Cytogenetics, Supplement to an International System for Human Cytogenetic Nomenclature. Basel: S. Karger.
Lehrach H, Drmanac R, Hoheisel J, Larin Z, Lennon G, Monaco AP, Nizetic D, Zehetner G, Poutska A (1990) Hybridization fin- gerprinting in genome mapping and sequencing. In Davies KE, Tilghman SM (eds): Genome Analysis. Vol 1: Genetic and Phys- ical Mapping. Cold Spring Harbor, Ny: Cold Spring Harbor Lab- oratory Press, pp 39-81.
Nordkvist A, Mark J, Gustavsson H, Bang G, Stenman G (1994) Nonrandom chromosome rearrangements in adenoid cystic carci- noma of the salivary glands. Genes Chromosom Cancer 1 0 115- 121.
Ohjimi Y, Iwasaki H, Kaneko Y, Ishiguro M, Ohgami A, Kikuchi M (1992) A case of lipoblastoma with t(3;8)(q12;q11.2). Cancer Genet Cytogenet 62: 103-105.
Sandberg AA, Gibas Z, Saren E, Li FP, Limon J, Tebbi CK (1986) Chromosome abnormalities in two benign adipose tumors. Cancer Genet Cytogenet 2255-61.
Sandros J, Stenman G, Mark J (1990) Cytogenetic and molecular observations in human and experimental salivary gland tumors. Cancer Genet Cytogenet 44:153-167.
Sapru M, Gu J, Gu X, Smith D, Yu C-E, Wells D, Wagner M (1994) A panel of radiation hybrids for chromosome 8. Genomics 21:
Sawyer JR, Parsons EA, Crowson ML, Smith S, Erickson S, Bell JM (1994) Potential diagnostic implications of breakpoints in the long arm of chromosome 8 in lipoblastoma. Cancer Genet Cytogenet 76~39-42.
Schoenmakers EFPM, Kools PFJ, Mols R, Kazmierczak B, Bart- nitzke S, BullerdiekJ, Dal Cin P, De Jong PJ, Van den Berghe H, Van de Ven WJM (1994) Physical mapping of chromosome 12q breakpoints in lipoma, pleomorphic salivary gland adenoma, uter- ine leiomyoma and myxoid liposarcoma. Genomics 20:210-222.
Schoenmakers EFPM, Wanschura S, Mols R, Bullerdiek J. Van den Berghe H, Van de Ven WJM (1995) Recurrent rearrangements in the high mobility group protein gene, HMGI-C, in benign mes- enchymal tumors. Nat Genet 10436-443.
Stenman G, Sahlin P, Mark J, Landys D (1991) Structural alter- ations of the c-mos locus in benign pleomorphic adenomas with chromosome abnormalities of 8q12. Oncogene 6 1105-1 108.
Stenman G, Bullerdiek J, Banniuke S, Sahlin P, Roijer E, Mark J (1994) Specificity and implications of chromosome 12 abnormal- ities in pleomorphic adenomas. In Bullerdiek J, Bartnitzke S (eds): Chromosome 12 Aberrations in Human Solid Tumors- Cytogenetics and Molecular Genetics. Berlin: Springer-Verlag,
Wanschura S, Kazmierczak B, Schoenmakers E, Meyen E, Bart- nitzke S, Van de Ven W, Bullerdiek J (1995) Regional mapping of the multiple-aberration region involved in uterine leiomyoma, lipoma and pleomorphic adenoma of the salivary gland to 12q15. Genes Chromosom Cancer 1468-70.
Wood S, Scheraer M, Yaremko ML (1995) Sequence identity lo- cates CEBPD and FGFRl to mapped human loci within proximal 8p. Cytogenet Cell Genet 70188-191.
208-216.
pp 3-15.


















