
Identification and immunological characterization of thioredoxin transmembrane-related protein from...
8
ORIGINAL PAPER Identification and immunological characterization of thioredoxin transmembrane-related protein from Clonorchis sinensis Chenhui Zhou & Meng Bian & Hua Liao & Qiong Mao & Ran Li & Juanjuan Zhou & Xiaoyun Wang & Shan Li & Chi Liang & Xuerong Li & Yan Huang & Xinbing Yu Received: 6 December 2012 / Accepted: 29 January 2013 / Published online: 13 February 2013 # Springer-Verlag Berlin Heidelberg 2013 Abstract Thioredoxin transmembrane related protein (TMX), a member of thioredoxin superfamily, is localized to the endoplasmic reticulum and possesses a thioredoxin-like do- main that plays an important role as an oxidoreductase. The functions of TMX in Clonorchis sinensis remain to be eluci- dated. In this study, we cloned and characterized a novel TMX of C. sinensis (CsTMX). The CsTMX cDNA sequence contained a 414-nucleotide open-reading frame encoding a protein of 137 amino acids. A thioredoxin domain was found in the position of aa 21–117 and contained the putative active-site motif Cys–Pro–Ala–Cys. BLASTx analysis showed that CsTMX shared 39–57 % amino acid identities with TMX of other organisms. Quantitative RT-PCR analysis demonstrated that CsTMX was differentially transcribed, with the highest level of expression in the adult worm stage and the lowest expression in egg stage. In addition, immunofluorescence assay showed CsTMX was localized in the tegument, vitelline gland, intestine, and intrauterine eggs of adult worm. Besides, immu- noblot assay revealed that the recombinant CsTMX (rCsTMX) could be recognized by the sera from rats infected with C. sinensis and the sera from rats immunized by excretory– secretory products. Furthermore, analysis of the antibody iso- type profile revealed that rats subcutaneously immunized with rCsTMX developed rCsTMX-specific antibody, which is dom- inance of IgG2a in sera. Meanwhile, production of IFN-γ was elevated strongly in the supernatants of spleen cell. The results collectively indicated that CsTMX might play an important role in the host–parasite interaction, as well as CsTMX probably involved in immunoregulation of host by inducing Th1-type dominated immune response in rats. Introduction Clonorchiasis are endemic in countries and regions of eastern Asia, including China, Taiwan, Northern Vietnam, and Korea. Globally, approximately 35 million people are infected with this food-borne trematode and more than 15 million people with Clonorchiasis in China (Chen et al. 2010; Lun et al. 2005). Clonorchis sinensis is a fish-borne zoonotic trematode that leads to clonorchiasis with pathological changes, includ- ing pyogenic cholangitis, cholelithiasis, cholecystitis, and he- patic fibrosis (Fried and Abruzzi 2010; Fried et al. 2011; Liao et al. 2006), even cholangiocarcinoma (Hong and Fang 2012; Fried et al. 2011). The C. sinensis metacercaria excyst in the duodenum, then migrate into the hepatic bile ducts where the flukes mature into adult worms (Hong et al. 1993). The fluke is continuously exposed to oxidizing molecules released by epithelial cells in the ductal system, and endogenous reactive oxygen species (ROS) generated by its own metabolic pro- cesses (Mkoji et al. 1988; Kaewkes 2003). Therefore, a series of antioxidant enzymes are of vital importance for C. sinensis against ROS derived from both the host cells and the normal cellular metabolism of the fluke. C. Zhou : M. Bian : H. Liao : Q. Mao : R. Li : J. Zhou : X. Wang : S. Li : C. Liang : X. Li : Y. Huang : X. Yu Department of Parasitology, Zhongshan School of Medicine, Sun Yat-sen University, Guangzhou 510080, People’ s Republic of China C. Zhou : M. Bian : H. Liao : Q. Mao : R. Li : J. Zhou : X. Wang : S. Li : C. Liang : X. Li : Y. Huang : X. Yu (*) Key Laboratory for Tropical Diseases Control, Ministry of Education, Sun Yat-sen University, Guangzhou 510080, People’ s Republic of China e-mail: [email protected] C. Zhou School of Nursing, Guangdong Medical College, Dongguan 523808, People’ s Republic of China Parasitol Res (2013) 112:1729–1736 DOI 10.1007/s00436-013-3331-5
Transcript of Identification and immunological characterization of thioredoxin transmembrane-related protein from...

ORIGINAL PAPER
Identification and immunological characterizationof thioredoxin transmembrane-related proteinfrom Clonorchis sinensis
Chenhui Zhou & Meng Bian & Hua Liao & Qiong Mao &
Ran Li & Juanjuan Zhou & Xiaoyun Wang & Shan Li &Chi Liang & Xuerong Li & Yan Huang & Xinbing Yu
Received: 6 December 2012 /Accepted: 29 January 2013 /Published online: 13 February 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Thioredoxin transmembrane related protein (TMX),a member of thioredoxin superfamily, is localized to theendoplasmic reticulum and possesses a thioredoxin-like do-main that plays an important role as an oxidoreductase. Thefunctions of TMX in Clonorchis sinensis remain to be eluci-dated. In this study, we cloned and characterized a novelTMX of C. sinensis (CsTMX). The CsTMX cDNA sequencecontained a 414-nucleotide open-reading frame encoding aprotein of 137 amino acids. A thioredoxin domain was foundin the position of aa21–117 and contained the putative active-sitemotif Cys–Pro–Ala–Cys. BLASTx analysis showed thatCsTMX shared 39–57 % amino acid identities with TMX ofother organisms. Quantitative RT-PCR analysis demonstratedthat CsTMX was differentially transcribed, with the highestlevel of expression in the adult worm stage and the lowestexpression in egg stage. In addition, immunofluorescence assayshowed CsTMXwas localized in the tegument, vitelline gland,intestine, and intrauterine eggs of adult worm. Besides, immu-noblot assay revealed that the recombinant CsTMX (rCsTMX)could be recognized by the sera from rats infected with
C. sinensis and the sera from rats immunized by excretory–secretory products. Furthermore, analysis of the antibody iso-type profile revealed that rats subcutaneously immunized withrCsTMX developed rCsTMX-specific antibody, which is dom-inance of IgG2a in sera. Meanwhile, production of IFN-γ waselevated strongly in the supernatants of spleen cell. The resultscollectively indicated that CsTMX might play an importantrole in the host–parasite interaction, as well as CsTMXprobably involved in immunoregulation of host by inducingTh1-type dominated immune response in rats.
Introduction
Clonorchiasis are endemic in countries and regions of easternAsia, including China, Taiwan, Northern Vietnam, and Korea.Globally, approximately 35 million people are infected withthis food-borne trematode and more than 15 million peoplewith Clonorchiasis in China (Chen et al. 2010; Lun et al.2005). Clonorchis sinensis is a fish-borne zoonotic trematodethat leads to clonorchiasis with pathological changes, includ-ing pyogenic cholangitis, cholelithiasis, cholecystitis, and he-patic fibrosis (Fried and Abruzzi 2010; Fried et al. 2011; Liaoet al. 2006), even cholangiocarcinoma (Hong and Fang 2012;Fried et al. 2011). The C. sinensis metacercaria excyst in theduodenum, then migrate into the hepatic bile ducts where theflukes mature into adult worms (Hong et al. 1993). The flukeis continuously exposed to oxidizing molecules released byepithelial cells in the ductal system, and endogenous reactiveoxygen species (ROS) generated by its own metabolic pro-cesses (Mkoji et al. 1988; Kaewkes 2003). Therefore, a seriesof antioxidant enzymes are of vital importance for C. sinensisagainst ROS derived from both the host cells and the normalcellular metabolism of the fluke.
C. Zhou :M. Bian :H. Liao :Q. Mao :R. Li : J. Zhou :X. Wang :S. Li : C. Liang :X. Li :Y. Huang :X. YuDepartment of Parasitology, Zhongshan School of Medicine,Sun Yat-sen University, Guangzhou 510080,People’s Republic of China
C. Zhou :M. Bian :H. Liao :Q. Mao :R. Li : J. Zhou :X. Wang :S. Li : C. Liang :X. Li :Y. Huang :X. Yu (*)Key Laboratory for Tropical Diseases Control, Ministryof Education, Sun Yat-sen University, Guangzhou 510080,People’s Republic of Chinae-mail: [email protected]
C. ZhouSchool of Nursing, Guangdong Medical College, Dongguan523808, People’s Republic of China
Parasitol Res (2013) 112:1729–1736DOI 10.1007/s00436-013-3331-5

The thioredoxin (TRX) superfamily, as one of the largestsuperfamilies of cellular redox homeostasis (Nakamura 2005),is extensively dispersed in the organism kingdom (Spindel etal. 2012; Holmgren and Sengupta 2010). Transmembranethioredoxin-related protein (TMX), a member of the TRXsuperfamily, is localized to the endoplasmic reticulum of theeukaryotes. TMX possesses a TRX-like domain with Cys–Pro–Ala–Cys (CPAC) active-site motif that is responsible forcatalysis of the thiol–disulfide exchange (Meng et al. 2003)and plays an important role in proper disulfide bond formationvia catalytic cysteine residues in the TRX-like domain(Sugiura et al. 2010; Haugstetter et al. 2005).
Recently, accumulate evidences demonstrate that TRXsuperfamily proteins play important roles not only in theparasite–host cell interaction but also in the protection ofadult parasite against oxidative stress (Liao et al. 2006;Williams et al. 2012; Maggioli et al. 2011). However, thecharacteristics of TMX in C. sinensis were not reported. Inthis study, TMX of C. sinensis was identified and isolated,as well as the spatiotemporal expression pattern of CsTMXwas examined at the RNA levels. Furthermore, immunolog-ical features of CsTMX were also characterized.
Material and methods
Preparation of parasites and C. sinensis excretory–secretoryproducts
Metacercariae ofC. sinensiswere isolated fromPseudorasboraparva digested with pepsin-HCl. Adult worms were freshlyextracted from the bile ducts of euthanized rats infected with C.sinensis for 8 weeks. Meanwhile, the C. sinensis excretory–secretory products (CsESPs) and eggs of C. sinensis wereobtained as described previously (Hu et al. 2007).
Sequence analysis of CsTMX
The complete encoding sequence of CsTMX was initiallyobtained from GenBank in the National Center forBiotechnology Information (http://www.ncbi.nlm.nih.gov).Proteomics and sequence analysis tools (http://www.expasy.org/) were applied to analyze protein characteristics such asmolecular weight (MW), theoretical isoelectric point (pI),and physicochemical parameters. Searching for CsTMXmotifs was performed by PROSITE programs. The homol-ogy was predicted by bioinformatic analysis software pack-ages PCgene and Vector NTI suite 8.0.
Transcript analysis of CsTMX gene
To analyze stage-specific expression at the level of transcrip-tion, total RNA were extracted from different stages of C.
sinensis. TRIZOL reagent (Invitrogen, USA) was applied forextraction following the manufacturer’s protocol. The cDNAswere obtained by reverse transcription polymerase chain re-action (PCR). Quantitative real-time PCR based on SYBR-Green I fluorescence (TaKaRa) was performed by usingRoche LigthCycler480. Primers (forward primer: 5′-GGCTTT ACA ATG GAT CAC GAA AC-3′ and the reverseprimer: 5′-TGG TGA GTA GTA CCA AGG GAT AGG-3′)of CsTMX were specifically designed by Premier 5.0. C.sinensis β-actin (GenBank accession number EU109284), asan internal control, was amplified using sense primer: 5′-ACCGTG AGA AGA TGA CGC AGA-3′ and antisense primer:5′-GCCAAGTCCAAACGAAGAATT-3′ (Yoo et al. 2009).The relative quantification analysis was carried out by calcu-lating the values of 2−ΔΔCT (Livak and Schmittgen 2001).
Cloning, expression, and production of the rCsTMX
To produce a recombinant CsTMX, the full-length cDNAsequence encoding the CsTMX was directionally clonedinto pET28a (+) (Novagen, USA), using sense primer:5′-TAGGATCCGACGTAATCAACATCAATTCC-3′ andantisense primer: 5′-GCTCTCGAGTTACTGAAATGTTGATGGTGAG-3′ (the restriction sites was underlined in pri-mers). The recombinant plasmid was confirmed by DNAsequencing, and then transformed into Escherichia coliBL21 (DE3) (Promega, USA). The protein expression wasinduced with 0.4 mM isopropyl-β-D-thiogalactopyranoside(IPTG) at 37 °C for 4 h in Luria–Bertani medium. TherCsTMX was purified by His·Bind Resin Kit (Novagen,USA) and assessed for identity and quality by 15 % sodiumdodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The final concentration of rCsTMX was detectedby the BCA Assay Kit (Novagen, USA).
Preparation of rat antisera
The purified rCsTMX (200 μg) and CsESPs (200 μg) wererespectively emulsified with equal volume of completeFreund’s adjuvant (CFA, Sigma-Aldrich, St. Louis, MO,USA) and subcutaneously injected into two 6-week-oldWistar rats. Two booster immunizations were carried out withrCsTMX or CsESPs (100 μg), respectively, mixed with equalvolume of incomplete Freund’s adjuvant at 2-week intervalsubsequently. Meanwhile, the naive sera were collected beforeinjection. The antisera were collected 2 weeks after the lastinjection. All sera samples were stored at −80 °C.
Western blotting analysis
The purified rCsTMX were separated by 15 % SDS-PAGEand subsequently transferred onto polyvinylidene difluoridemembrane (Whatman, USA) for 1 h at 100 V. The membrane
1730 Parasitol Res (2013) 112:1729–1736

was blocked with phosphate-buffered saline (PBS; pH7.4)containing 5% skimmilk for 2 h at 37 °C. After being washedfive times with PBS containing 0.05 % Tween 20 (PBS-T),strips were incubated respectively with His·Tag monoclonalantibody (mAb-His; Novagen, USA, 1:2,000 dilutions), anti-rCsTMX rat serum (1:1,000 dilutions),C. sinensis-infected ratserum (1:200 dilutions), anti-CsESPs rat serum (1:100 dilu-tions), or naive rat serum (1:100 dilutions) at 4 °C overnight.After rinsing five times with PBS-T, the strips were incubatedwith horseradish peroxidase-conjugated goat anti-rat IgG(ProteinTech Group, USA, 1:5,000 dilutions) at room temper-ature (RT) for 2 h. Blots were detected by enhanced chemilu-minescence method.
Immunofluorescence assay
Fresh C. sinensis adult worms were fixed in 4 % parafor-maldehyde overnight, then embedded with paraffin, andsliced into 4–5 μm sections. After dewaxing and dehydra-tion with graded ethanol, the samples were blocked withnormal goat serum for 2 h at RT, and incubated with anti-rCsTMX serum (1:200 dilutions) or naive rat serum (1:200dilutions) at 4 °C overnight, respectively. The samples werewashed three times with PBS-T and incubated with Cy™three-conjugated AffiniPure goat anti-rat IgG (1:400 dilu-tions) for 1 h in darkness at RT. The samples were viewedby fluorescence microscopy (Carl Zeiss, Germany).
Detection of CsTMX specific antibodies
Enzyme-linked immunosorbent assay (ELISA) was per-formed to detect specific IgG antibodies against rCsTMX(2 μg/well, pH9.6) coated on 96-well microtiter plates(Costar, USA) at 4 °C overnight. The wells were blockedwith 5 % skimmed milk in PBS-T for 2 h, and incubatedwith samples sera for 2 h at 37 °C (1:100 dilutions).Thereafter, the wells were respectively incubated with thegoat anti-rat IgG1- or IgG2a-conjugated horseradish perox-idase (1:2,500 dilutions; Sigma). Thorough washes weretaken after each antibody incubation step. Substrate solution3,3,5,5-tetramethyl-benzidine (100 μl , TMB, BDBiosciences) was added to each well. The plates were incu-bated in darkness for 10 min. The reaction was terminatedby 2 M sulfuric acid (50 μl/well), and the absorbance wasmeasured at 450 nm.
Cytokine analysis
To detect cytokines, splenocytes were isolated frommaceratedspleen of individual rat 2 weeks after the third immunizationwith rCsTMX or PBS. After removing the red blood cells byRBC lysis solution (Sigma), the splenocytes were cultured at aconcentration of 5×106 viable cells/well with 5 μg of
rCsTMX or ConA (Sigma), or PBS alone (unstimulated con-trol). The culture supernatants were collected (24 h for IL-4and 72 h for IFN-γ) and tested for the presence of cytokinesusing a commercial ELISA kit (BD Biosciences). All assayswere performed in triplicate. The concentrations of cytokineswere calculated using a linear regression equation obtainedfrom standard absorbance values.
Statistical analysis
Statistical analysis was performed using SPSS for Windowsversion 10.0 (SPSS Inc., Chicago, IL, USA). All resultswere presented as mean±SD and the statistical differencewas evaluated using variance (ANOVA). P<0.05 was con-sidered statistically significant.
Results
Sequence analysis of CsTMX
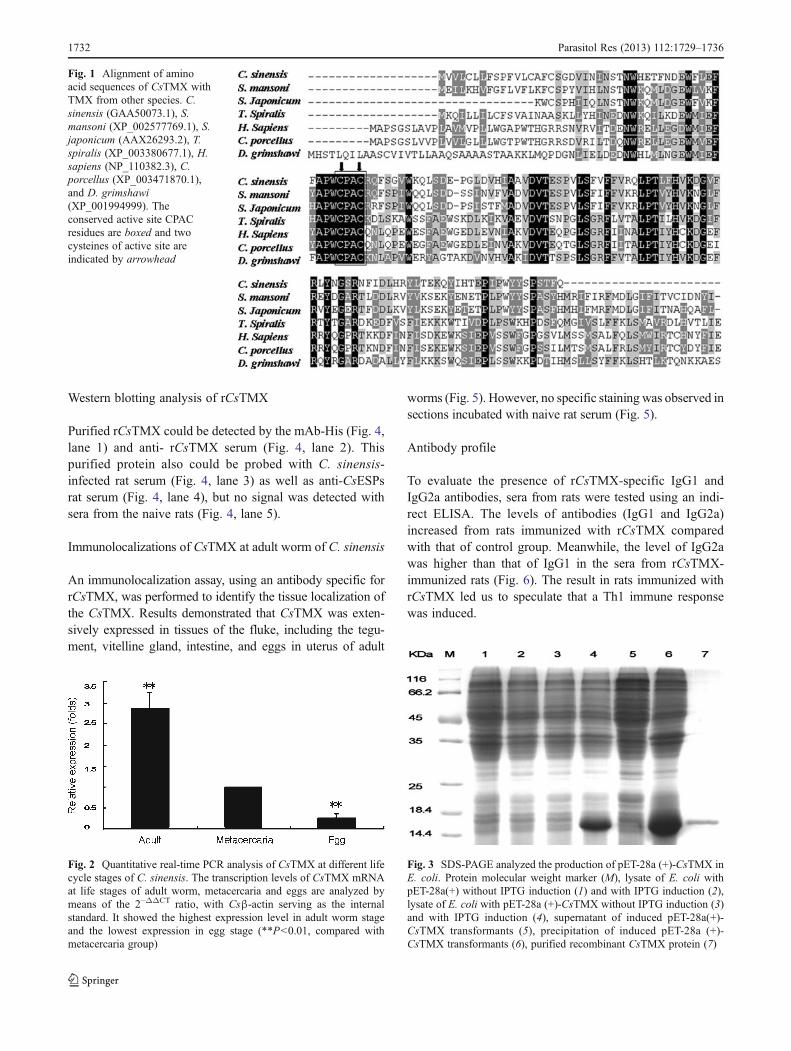
The CsTMX cDNA molecule contained a 414-nucleotideopen-reading frame encoding a protein of 137 amino acids(predicted MW, 16.037 KDa; pI, 5.16). BLASTx analysisshowed that the deduced amino acid sequence was homol-ogous to protein TMX from Schistosoma mansoni,Schistosoma japonicum, Trichinella spiralis, Homo sapiens,Drosophila grimshawi, and Cavia porcellus with 53, 57, 40,39, 45, and 39 % identities, respectively. SignalP analysisshowed that CsTMX has cleavage site between aa19 andaa20. A thioredoxin domain was found in the position ofaa21–117 with thioredoxin family conserved sites (aa38–56)which was characteristic in TMX from other species, andcontained the putative active-site motif CPAC (Meng et al.2003; Fig. 1).
Transcript analysis of CsTMX gene expression level
The results revealed that CsTMX mRNA was expressed inall developmental stages examined but with different levels.Interestingly, it showed the highest expression level in adultstage, in which the mRNA expression level of CsTMX wasabout threefold higher than that in metacercariae stage. Thetranscription level of C. sinensis in egg stage was the lowest(Fig. 2).
Expression and purification of rCsTMX
The recombinant CsTMX was expressed as inclusion bodieswith 6× His tag at the N-terminus in E. coli. The molecularweight of the purified protein was around 17.549 KDa,consistent with the predicted MW (Fig. 3), and the finalprotein concentration was up to 250 μg/ml.
Parasitol Res (2013) 112:1729–1736 1731
Western blotting analysis of rCsTMX
Purified rCsTMX could be detected by the mAb-His (Fig. 4,lane 1) and anti- rCsTMX serum (Fig. 4, lane 2). Thispurified protein also could be probed with C. sinensis-infected rat serum (Fig. 4, lane 3) as well as anti-CsESPsrat serum (Fig. 4, lane 4), but no signal was detected withsera from the naive rats (Fig. 4, lane 5).
Immunolocalizations of CsTMX at adult worm of C. sinensis
An immunolocalization assay, using an antibody specific forrCsTMX, was performed to identify the tissue localization ofthe CsTMX. Results demonstrated that CsTMX was exten-sively expressed in tissues of the fluke, including the tegu-ment, vitelline gland, intestine, and eggs in uterus of adult
worms (Fig. 5). However, no specific staining was observed insections incubated with naive rat serum (Fig. 5).
Antibody profile
To evaluate the presence of rCsTMX-specific IgG1 andIgG2a antibodies, sera from rats were tested using an indi-rect ELISA. The levels of antibodies (IgG1 and IgG2a)increased from rats immunized with rCsTMX comparedwith that of control group. Meanwhile, the level of IgG2awas higher than that of IgG1 in the sera from rCsTMX-immunized rats (Fig. 6). The result in rats immunized withrCsTMX led us to speculate that a Th1 immune responsewas induced.
Fig. 1 Alignment of aminoacid sequences of CsTMX withTMX from other species. C.sinensis (GAA50073.1), S.mansoni (XP_002577769.1), S.japonicum (AAX26293.2), T.spiralis (XP_003380677.1), H.sapiens (NP_110382.3), C.porcellus (XP_003471870.1),and D. grimshawi(XP_001994999). Theconserved active site CPACresidues are boxed and twocysteines of active site areindicated by arrowhead
Fig. 2 Quantitative real-time PCR analysis of CsTMX at different lifecycle stages of C. sinensis. The transcription levels of CsTMX mRNAat life stages of adult worm, metacercaria and eggs are analyzed bymeans of the 2−ΔΔCT ratio, with Csβ-actin serving as the internalstandard. It showed the highest expression level in adult worm stageand the lowest expression in egg stage (**P<0.01, compared withmetacercaria group)
Fig. 3 SDS-PAGE analyzed the production of pET-28a (+)-CsTMX inE. coli. Protein molecular weight marker (M), lysate of E. coli withpET-28a(+) without IPTG induction (1) and with IPTG induction (2),lysate of E. coli with pET-28a (+)-CsTMX without IPTG induction (3)and with IPTG induction (4), supernatant of induced pET-28a(+)-CsTMX transformants (5), precipitation of induced pET-28a (+)-CsTMX transformants (6), purified recombinant CsTMX protein (7)
1732 Parasitol Res (2013) 112:1729–1736

Cytokine profile
To examine the cell-mediated immune response induced bythe rCsTMX, the cytokine profile of spleen cells from ratsimmunized with rCsTMX (rCsTMX group) or PBS (controlgroup) was measured. The supernatants of spleen cell cul-tures from rCsTMX group contained higher level of IFN-γthan that of control group (Fig. 8). However, there was nosignificant difference in levels of IL-4 between two groups(Fig. 7). Taken together, these results indicate that immuni-zation with rCsTMX induced a specific Th1 dominate im-mune response in rats.
Discussion
The adult flukes of C. sinensis inhabited the bile duct andcontinuously encountered with lethal ROS released by theirown metabolic processes and the host effector cells. Tocontrol the redox balance, the parasites are equipped witha variety of antioxidants and redox enzymes to protect themfrom fatal radicals. TMX, as a member of TRX superfamily,
is an oxidoreductase with a rather unusual active site motif(CPAC) which could affect the redox potential and contrib-ute to its substrate specificity distinct from those of oxidor-eductases with the common CGHC motifs (Matsuo et al.2009). TMX are present mainly in the reduced form ineukaryotes (Ellgaard and Ruddock 2005), accumulated evi-dences suggest that TMX could work as a reductase andmight have a physiological role as an oxidase like proteindisulfide isomerase (PDI; Sugiura et al. 2010; Pasetto et al.2012; Matsuo et al. 2004). TMX, as an unconventionalinhibitor of major histocompatibility complex class I heavychain, might play a protective role in pathological condi-tions such as inflammation and infectious diseases (Matsuoet al. 2009).
In the present study, we isolated, cloned, and charac-terized a TMX of C. sinensis. The active site CPACresidues are completely conserved. Meanwhile, there are39–57 % amino acid identities CsTMX from TMX ofother organisms. SignalP analysis showed that CsTMX
Fig. 4 Immunoreactivity of rCsTMX was analyzed by western blot-ting. 1–4 Purified rCsTMX is probed with anti-His tag monoclonalantibody (1), sera from anti-rCsTMX (2), sera from C.sinensis-infectedrat (3), and sera from anti-CsESPs (4). rCsTMX cannot be blotted withsera from naive rats (5)
Fig. 5 Immunolocalization of CsTMX at adult worm of C. sinensis.Rat anti-rCsTMX serum was used as primary antibody (a, b, e and f)and goat anti-rat IgG was employed as secondary antibody. Sera fromrat immunized with PBS were used as negative control (c, d, g and h).
Sections were imaged under fluorescence light (a, c, e, g), or whitelight (b, d, f, h). CsTMX was present in tegument (t), intestine (i),vitellarium (v), and eggs (e). The sections were observed with ×5magnification
Fig. 6 IgG isotype in sera from rCsTMX immunized rats. Rats im-munized with PBS serve as control group. IgG1 (open bar) and IgG2a(black bar) specific antibodies were examined by ELISA. IgG2a levelsof rats immunized with rCsTMX were significantly higher than IgG1(*P<0.05)
Parasitol Res (2013) 112:1729–1736 1733

has cleavage site between aa19 and aa20 (Fig. 1). In addition,western blotting analysis revealed that rCsTMX can be probedwithC. sinensis-infected rat serum, and anti-CsESPs rat serum(Fig. 4). The ESPs of parasites often contained antioxidantswhich were shed from tegument, and proteases which weresecreted from intestine (Robinson et al. 2009). In C. sinensis,the CsTMX was located not only on the tegument, but also inthe intestine, which might help to explain why rCsTMX canbe probed by sera from rats immunized with CsESPs.Furthermore, TRX and PDI, as members of TRX super-family, have been reported as components of ESPs toprotect parasites in oxidative stress environment (Berndtet al. 2008; Suttiprapa et al. 2012; Hu et al. 2012).Therefore, further investigations are required to demon-strate whether CsTMX could work as a reductase toprotect parasites from lethal radicals.
CsTMX was expressed in developmental stages of C.sinensis, including adult worms, metacercariae, and eggsstage (Fig. 2). Interestingly, it showed the highest expressionlevel in adult worm stage and the lowest expression in eggstage. The expression of CsTMX was gradually increased inmore developed stages of C. sinensis. It was also clarified inthe immunolocalization assay that CsTMX was extensivelyexpressed in tissues of adult worms, including the tegument,vitelline gland, intestine, and eggs in uterus of adult worms(Fig. 5). These results suggested that CsTMX is essential forthe parasite throughout the life cycle. The results were alsoconsistent with the earlier reports that proteins of TRXsuperfamily are broadly distributed in several parasites, suchas TRX from Opisthorchis viverrini, PDI and peroxiredox-ins from C. sinensis (Hu et al. 2012; Bae et al. 2011). TRXfrom O. viverrini was detected in all developmental stagesand expressed in nearly all parasite tissue, suggesting itsmultiple biological functions including growth factor, struc-tural component of enzymes, redox regulation of transcrip-tion factor, and immune modulation (Suttiprapa et al. 2012).Recently, TMX was reported to play a major role in hostdefense under the type of inflammatory conditions andpotentially participate in the cellular protection from oxida-tive damage (Matsuo et al. 2012). Thus, we suspected thatthe protein might play important roles not only in the growthand development of C. sinensis but also in the protection ofC. sinensis against ROS generated from host defensivemechanisms.
The rCsTMX subcutaneous immunization elicited specif-ic humoral and cellular immune response in rats. The IgG1and IgG2a levels, especially IgG2a, increased significantlyin immunized group compared with control group (Fig. 6).Meanwhile, the cytokine profile induced by rCsTMX im-munization showed that IL-4 production was no significantchange between two groups (Fig. 7). Furthermore, the pro-duction of IFN-γ was strongly elevated in rCsTMX-immu-nized group compared with control group (Fig. 8). IFN-γ isthe key cytokine involved in Th1 cell development (Wang etal. 2009; Farias et al. 2010). These results illustrated thatrCsTMX could trigger a Th1-type predominant immuneresponse in rats. In the C. sinensis infected murine model,there are some evidences that Th1-type predominant im-mune response is related to protection (Wang et al. 2012;Lee et al. 2006; Lee and Yong 2004). Several reports havesuggested that IFN-γ-activated cells may be one mechanismof protective immunity (Dai et al. 2009; Lee et al. 2006).According to the immune responses elicited by rCsTMX,further studies about protection of the protein against clo-norchiasis are necessary to proceed.
In summary, we identified a novel CsTMX as the memberof TRX superfamily in C. sinensis and obtained the recombi-nant CsTMX. Moreover, CsTMX was widely expressed intissues of C. sinensis adult worms, and expressed across
Fig. 7 Quantitative ELISA analysis of IL-4 secreted by splenocytesafter PBS (open bar), rCsTMX (black bar 5 μg/ml) or ConA (hatchedbar 5 μg/ml) stimulation in the rats 2 weeks after the final vaccination
Fig. 8 IFN-γ profile secreted by splenocytes from the rats immunizedwith rCsTMX. Splenocytes were isolated from the rats 2 weeks afterthe final vaccination. Quantitative ELISA analysis of IFN-γ secretedby splenocytes after PBS (open bar), rCsTMX (black bar 5 μg/ml) orConA (hatched bar 5 μg/ml) stimulating for 72 h. The level of IFN-γin rCsTMX group was higher than that of control group (**P<0.01,compared with PBS group)
1734 Parasitol Res (2013) 112:1729–1736

developmental stages of the parasite. Furthermore, the im-mune responses that elicited by rCsTMX was a Th1-typepredominant immune response. Therefore, we speculatedCsTMX might play crucial roles in parasite survivaland parasite–host interactions. Further studies are stillneeded to confirm the biological functions of CsTMX,which are associated with invasion and survival of C.sinensis, and the potential value as a therapy target inC. sinensis infection.
Acknowledgments This work was supported by the National BasicResearch Program (973 project; No.2010CB530000), “Eleventh Five-year Plan” for Science & Technology Research of China (2008ZX1004-011) and the National Natural Science Foundation of China (81101270).
References
Bae YA, Kim SH, Lee EG, Sohn WM, Kong Y (2011) Identificationand biochemical characterization of two novel peroxiredoxins in aliver fluke, Clonorchis sinensis. Parasitol 138(9):1143–1153
Berndt C, Lillig CH, Holmgren A (2008) Thioredoxins and glutare-doxins as facilitators of protein folding. Biochim Biophys Acta1783(4):641–650
Chen D, Chen J, Huang J, Chen X, Feng D, Liang B, Che Y, Liu X,Zhu C, Li X, Shen H (2010) Epidemiological investigation ofClonorchis sinensis infection in freshwater fishes in the PearlRiver Delta. Parasitol Res 107(4):835–839
Dai Y, Zhu Y, Harn DA, Wang X, Tang J, Zhao S, Lu F, Guan X (2009)DNA vaccination by electroporation and boosting with recombi-nant proteins enhances the efficacy of DNA vaccines for Schisto-somiasis japonica. Clin Vaccine Immunol 16(12):1796–1803
Ellgaard L, Ruddock LW (2005) The human protein disulphide isom-erase family: substrate interactions and functional properties.EMBO Rep 6(1):28–32
Farias LP, Cardoso FC, Miyasato PA, Montoya BO, Tararam CA,Roffato HK, Kawano T, Gazzinelli A, Correa-Oliveira R, CoulsonPS, Wilson RA, Oliveira SC, Leite LC (2010) Schistosoma man-soni stomatin like protein-2 is located in the tegument and inducespartial protection against challenge infection. PLoS Negl Trop Dis4(2):e597
Fried B, Abruzzi A (2010) Food-borne trematode infections of humansin the United States of America. Parasitol Res 106(6):1263–1280
Fried B, Reddy A, Mayer D (2011) Helminths in human carcinogen-esis. Cancer Lett 305(2):239–249
Haugstetter J, Blicher T, Ellgaard L (2005) Identification and charac-terization of a novel thioredoxin-related transmembrane protein ofthe endoplasmic reticulum. J Biol Chem 280(9):8371–8380
Holmgren A, Sengupta R (2010) The use of thiols by ribonucleotidereductase. Free Radic Biol Med 49(11):1617–1628
Hong ST, Fang Y (2012) Clonorchis sinensis and clonorchiasis, anupdate. Parasitol Int 61(1):17–24
Hong ST, Kho WG, Kim WH, Chai JY, Lee SH (1993) Turnover ofbiliary epithelial cells in Clonorchis sinensis infected rats. KoreanJ Parasitol 31(2):83–89
Hu F, Yu X, Ma C, Zhou H, Zhou Z, Li Y, Lu F, Xu J, Wu Z, Hu X(2007) Clonorchis sinensis: expression, characterization, immu-nolocalization and serological reactivity of one excretory/secreto-ry antigen-LPAP homologue. Exp Parasitol 117(2):157–164
Hu Y, Huang L, Huang Y, He L, Zhang F, Li W, Liang P, Li R, Sun J,Wang X, Liang C, Li X, Yu X (2012) Molecular cloning,
expression, and immunolocalization of protein disulfide isomer-ase in excretory-secretory products from Clonorchis sinensis.Parasitol Res 111(3):983–989
Kaewkes S (2003) Taxonomy and biology of liver flukes. Acta Trop88:177–186
Lee JS, Yong TS (2004) Expression and cross-species reactivity offatty acid-binding protein of Clonorchis sinensis. Parasitol Res 93(5):339–343
Lee JS, Kim IS, Sohn WM, Lee J, Yong TS (2006) A DNA vaccineencoding a fatty acid-binding protein of Clonorchis sinensisinduces protective immune response in Sprague–Dawley rats.Scand J Immunol 63(3):169–176
Liao WC, Wang HP, Chiu HM, Chang CY, Lin JT (2006) Multiplehepatic nodules: rare manifestation of clonorchiasis. J Gastroen-terol Hepatol 21:1497–1500
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods 25(4):402–408
Lun ZR, Gasser RB, Lai DH, Li AX, Zhu XQ, Yu XB, Fang YY(2005) Clonorchiasis: a key foodborne zoonosis in China. LancetInfect Dis 5(1):31–41
Maggioli G, Silveira F, Martín-Alonso JM, Salinas G, Carmona C,Parra FA (2011) A recombinant thioredoxin-glutathione reductasefrom Fasciola hepatica induces a protective response in rabbits.Exp Parasitol 129(4):323–330
Matsuo Y, Nishinaka Y, Suzuki S, Kojima M, Kizaka-Kondoh S,Kondo N, Son A, Sakakura-Nishiyama J, Yamaguchi Y, MasutaniH, Ishii Y, Yodoi J (2004) TMX, a human transmembrane oxido-reductase of the thioredoxin family: the possible role in disulfide-linked protein folding in the endoplasmic reticulum. Arch Bio-chem Biophys 423(1):81–87
Matsuo Y, Masutani H, Son A, Kizaka-Kondoh S, Yodoi J (2009)Physical and functional interaction of transmembranethioredoxin-related protein with major histocompatibility com-plex class I heavy chain: redox-based protein quality control andits potential relevance to immune responses. Mol Biol Cell 20(21):4552–4562
Matsuo Y, Irie K, Kiyonari H, Okuyama H, Nakamura H, Son A,Lopez-Ramos DA, Tian H, Oka SI, Okawa K, Kizaka-Kondoh S, Masutani H, Yodoi J (2012) The protective roleof the transmembrane thioredoxin-related protein TMX ininflammatory liver injury. Antioxid Redox Signal (Epubahead of print)
Meng X, Zhang C, Chen J, Peng S, Cao Y, Ying K, Xie Y, Mao Y(2003) Cloning and identification of a novel cDNA codingthioredoxin-related transmembrane protein 2. Biochem Genet 41(3–4):99–106
Mkoji GM, Smith JM, Prichard RK (1988) Antioxidant systemsin Schistosoma mansoni: correlation between susceptibility tooxidant killing and the levels of scavengers of hydrogenperoxide and oxygen free radicals. Int J Parasitol 18(5):661–666
Nakamura H (2005) Thioredoxin and its related molecules: update.Antioxid Redox Signal 7(5–6):823–828
Pasetto M, Barison E, Castagna M, Della Cristina P, Anselmi C,Colombatti M (2012) Reductive activation of type 2 ribosome-inactivating proteins is promoted by transmembrane thioredoxin-related protein. J Biol Chem 287(10):7367–7373
Robinson MW, Menon R, Donnelly SM, Dalton JP, Ranganathan S(2009) An integrated transcriptomics and proteomics analysis ofthe secretome of the helminth pathogen Fasciola hepatica: pro-teins associated with invasion and infection of the mammalianhost. Mol Cell Proteomics 8(8):1891–1907
Spindel ON, World C, Berk BC (2012) Thioredoxin interacting pro-tein: redox dependent and independent regulatory mechanisms.Antioxid Redox Signal 16(6):587–596
Parasitol Res (2013) 112:1729–1736 1735

Sugiura Y, Araki K, Iemura S, Natsume T, Hoseki J, Nagata K (2010)Novel thioredoxin-related transmembrane protein TMX4 has re-ductase activity. J Biol Chem 285(10):7135–7142
Suttiprapa S, Matchimakul P, Loukas A, Laha T, Wongkham S,Kaewkes S, Brindley PJ, Sripa B (2012) Molecular expressionand enzymatic characterization of thioredoxin from the carcino-genic human liver fluke Opisthorchis viverrini. Parasitol Int 61(1):101–106
Wang X, Liang C, Chen W, Fan Y, Hu X, Xu J, Yu X (2009)Experimental model in rats for study on transmission dynamicsand evaluation of Clonorchis sinensis infection immunologically,morphologically, and pathologically. Parasitol Res 106(1):15–21
Wang X, Chen W, Lv X, Tian Y, Men J, Zhang X, Lei H, Zhou C, Lu F,Liang C, Hu X, Xu J, Wu Z, Li X, Yu X (2012) Identification andcharacterization of paramyosin from cyst wall of metacercariaeimplicated protective efficacy against Clonorchis sinensis infec-tion. PLoS One 7(3):e33703
Williams DL, Bonilla M, Gladyshev VN, Salinas G (2012) Thiore-doxin glutathione reductase-dependent redox networks in platy-helminth parasites. Antioxid Redox Signal (Epub ahead of print)
Yoo WG, Kim TI, Li S, Kwon OS, Cho PY, Kim TS, Kim K, Hong SJ(2009) Reference genes for quantitative analysis on Clonorchissinensis gene expression by real-time PCR. Parasitol Res 104(2):321–328
1736 Parasitol Res (2013) 112:1729–1736


















