
ICYSB, 2013Schaber 1/42 Introduction to Modelling Signalling Cascades in Yeast Jörg Schaber...
42
ICYSB, 2013 Schaber 1/42 Introduction to Modelling Signalling Cascades in Yeast Jörg Schaber Institute for Experimental Internal Medicine Otto-von-Guericke University Magdeburg www.sysbiolab.net
-
Upload
gregory-watkins -
Category
Documents
-
view
216 -
download
1
Transcript of ICYSB, 2013Schaber 1/42 Introduction to Modelling Signalling Cascades in Yeast Jörg Schaber...

ICYSB, 2013 Schaber 1/42
Introduction to Modelling Signalling Cascades in Yeast
Jörg Schaber
Institute for Experimental Internal Medicine
Otto-von-Guericke University
Magdeburg
www.sysbiolab.net

ICYSB, 2013 Schaber 2/42
• The modelling process• A simple step-by-step example
– The Sho1 branch of the HOG pathway• Basic concepts
– Signalling motifs– Model selection
Outline

ICYSB, 2013 Schaber 3/42
• Modelling requires verbal hypothesis be made specific and conceptually rigorous.
Why Models?
• Modelling highlights gaps in our knowledge.
• Modelling provides quantitative as well as qualitative predictions.
• Modelling is ideal for analysing complex interactions before experimental tests.
• Modelling is a low-cost, rapid test bed for candidate interventions.
• Well designed models are readily portable and adaptable for many purposes.

ICYSB, 2013 Schaber 4/42
ObservationPrior knowledge
• The observation of a natural phenomena drives a scientific question, which is represented by a testable hypothesis.
• This is the most important step as it sets the stage for the rest.
• Requires intuition and talent.
• The word model is the abstract representation of the processes that might explain data based on the hypothesis.
• Often represented by a diagram.
• Defines the components and their interactions.
• Defines systems boundaries.
• The math model is the formalised word model.
• The formalism depends on the processes and data, e.g.• dynamic systems:
ODEs• gene networks:
boolean models
• The verification is a first qualitative evaluation of the model.
• Checks whether the model is in principle able to explain the data, e.g.• if the data shows
oscillations, the model should be able to oscillate as well.
• The validation is a quantitative evaluation of the model.
• Checks whether the model is able to explain the data quantitatively.
• Fitting model to the data.
• Parameter optimization
• A successful validation consolidates our trust that we have captured the most important processes to explain the data.
• This justifies an analysis of model properties, e.g.• sensitivities• robustness
• The analysis may give indications for useful predictions.
• Sensitivity analysis may suggest a parameter with high impact on certain behaviour.
• We can define the ‘most informative’ experiment.
ObservationPrior knowledge
Math ModelMath Model
Analysis
Validation
Verification
Word Model
Hypothesis
Prediction
Analysis
Validation
Verification
Word Model
Hypothesis
The Modelling Process in 8 Steps
• A successful prediction consolidates our trust in the model and the hypothesis and that we have identified the most important processes to explain the data.
• Usually a prediction is not successful.
• Often we have to change our model/hypothesis in the course of the modelling process.
• Each modelling round deepens our understanding.

ICYSB, 2013 Schaber 5/42
A Step-by-Step Example
The Sho1-branch of the HOG pathway

ICYSB, 2013 Schaber 6/42
Oligomer-deficient
Oligomer-deficientPhospho-mimic
Hao et al. (2007) Curr. Biol. 17
sho1D ssk1D
The Data• It was shown that
a) Sho1 de-oligomerizes upon osmotic shockb) Hog1 phosphorylates de-oligomerized Sho1c) Phosphorylated Sho1 is less able to transmit
the signal

ICYSB, 2013 Schaber 7/42
The Hypothesis
– Phosphorylation of Sho1 by Hog1 constitutes a negative feedback loop.
– This negative feedback leads to the rapid attenuation of Hog1 signalling.
– Might be important to dampen crosstalk to pheromone signalling pathway.

ICYSB, 2013 Schaber 8/42
The Word Model(s)
• Signal: high osmolarity• Hog1 de-sensitizes Sho1
Hao et al. (2007) Curr. Biol. 17

ICYSB, 2013 Schaber 9/42
Formalising Word Models
Pbs2P
Hog1
‘Biologist’ notation
Not very useful (for modelling), becauseinteractions not clear.
Pbs2P
Hog1P
‘Systems Biologist’ notation
Hog1v1
v2
More useful, because each interaction is made specific.This facilitates mathematical formulation.

ICYSB, 2013 Schaber 10/42
Formalising Word Models
Pbs2P
Hog1PHog1v1
v2
• arrows between components indicate transformations, i.e. biochemical reactions, mass flows. They determine changes in concentrations, numbers, etc.
• Instead of giving explicit formulas for components, e.g. HogP(t)=f(v1,v2), we rather characterise the change of components over time.• Change = what goes in
– what goes out

ICYSB, 2013 Schaber 11/42
Formalising Word Models
Pbs2P
Hog1PHog1v1
v2
• That is one reason, why ordinary differential equation (ODE) models are so popular: easy to set up given a carefully set up wiring scheme:
21
21
1
1
vvdt
dHog
vvdt
PdHog
Note that Pbs2P does not change.

ICYSB, 2013 Schaber 12/42
Formalising Word Models
Pbs2P
Hog1PHog1v1
v2
• arrows on arrows indicate modifying interactions (enzymatic reactions), i.e. no (net) mass flows or concentrations changes involved from emanating components.
Biochemical notation
Hog1 + Pbs2P -> Hog1P + Pbs2P
Pbs2P neither consumed nor produced (netto-wise)

ICYSB, 2013 Schaber 13/42
Formalising Word Models
Pbs2P
Hog1PHog1v1
v2
Biochemical notation
v1: Hog1 + Pbs2P -> Hog1P + Pbs2P
v2: Hog1P -> Hog1
Most simple mathematical formulation: mass action kinetics, i.e. multiplication of substrates.
v1: k1·Hog1·Pbs2Pv2: k2·Hog1P
21
21
1
1
vvdt
dHog
vvdt
PdHog

ICYSB, 2013 Schaber 14/42
Formalising Word Models
• Set up wiring scheme, where each considered reaction with modifiers is made explicit.
• Formulate ODE system with balance equations.
• Choose kinetics rate formulation.
• Choose initial conditions.
Þ Theory tells us:a) There exists a solution.b) The solution is unique.c) The solution can be arbitrarily approximated.

ICYSB, 2013 Schaber 15/42
Formalising Word Models
Fus3
Fus3-Ste5
Ste5
v1
v2
Biochemical notation
v1: Fus3 + Ste5 -> Fus3-Ste5
v2: Fus3-Ste5 -> Fus3 + Ste5
Most simple mathematical formulation: mass action kinetics, i.e. linear multiplication of substrates.
v1: k1·Fus3·Ste5v2: k2·Fus3-Ste5
21
21
21
53
5
3
vvdt
StedFus
vvdt
dSte
vvdt
dFus

ICYSB, 2013 Schaber 16/42
Kinetic rate laws and signalling motifs
Pv1
Constant fluxand simple mass action degradation
v2
Pkkdt
dP21
P
v1
Modified constant fluxand simple mass action degradation
v2 PkSkdt
dP
Skdt
dS
2
3
1
S v3
2
2
1
10
k
kP
Pkk
2
1
k
k
0)0( tP
1)0(
0)0(
tS
tP

ICYSB, 2013 Schaber 17/42
Pv1
Modified conversion with signal degradationMass action kinetics
v2
1
)1( 2
3
1
PQ
PkPSkdt
dP
Skdt
dSS v3
Q
Pv1
Modified conversion with signal degradationMichaelis-Menten kinetics
v2
S v3
Q
PKm
Pk
PKm
PSk
dt
dP
SKm
Sk
dt
dS
2
2
1
3
3
1
)1(1
k3
k3/2
Km3
Kinetic rate laws and signalling motifs

ICYSB, 2013 Schaber 18/42
Pv1
Modified conversion with signal degradationHill kinetics
v2
S v3
Q
22
2
2
11
1
11
33
3
3
2
3
)1(
)1(hh
h
hh
h
hh
h
PKm
Pk
PKm
PSk
dt
dP
SKm
Sk
dt
dS
k3/2
Km3
k3
Kinetic rate laws and signalling motifs
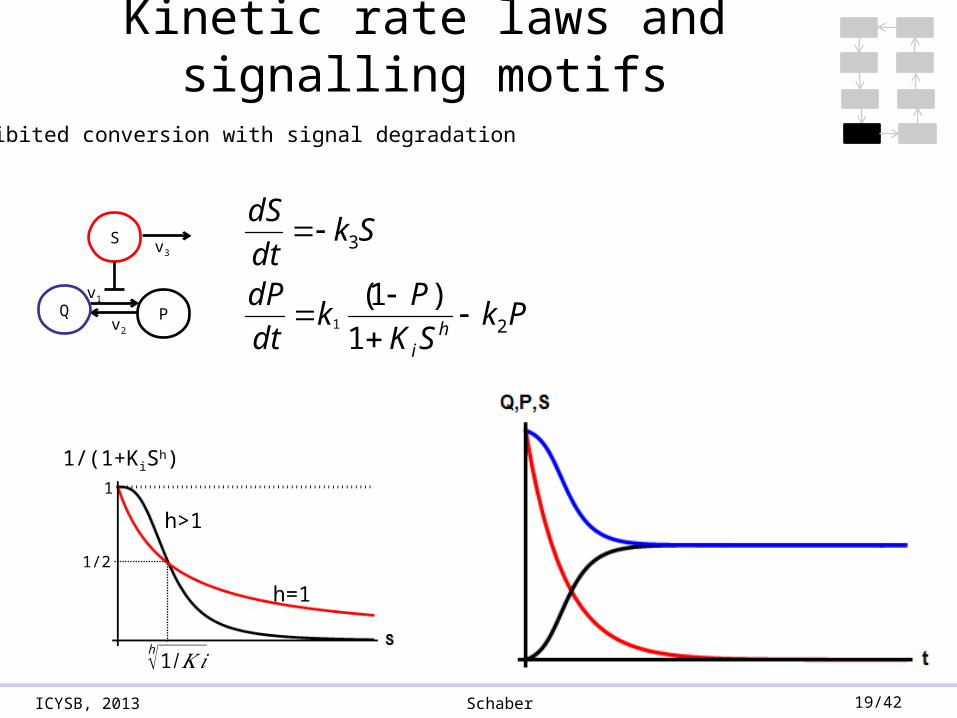
ICYSB, 2013 Schaber 19/42
Pv1
Inhibited conversion with signal degradation
v2
S v3
Q PkSK
Pk
dt
dP
Skdt
dS
hi
2
3
1
)1(1
Kinetic rate laws and signalling motifs
1/2
h√1/𝐾 𝑖
1
1/(1+KiSh)
h=1
h>1

ICYSB, 2013 Schaber 20/42
P
v1
Modified conversion with negative feedback
v2
S
Q
PkPK
PSk
dt
dPh
i21
)1(1
Observation:• System reaches new
steady state.• No ‘overshoot’
Possible explanation:• Feedback comes too
fast.
Kinetic rate laws and signalling motifs

ICYSB, 2013 Schaber 21/42
P
v1
Modified conversion with delayed negative feedback
v2
S
Q
PktPK
PSk
dt
dPh
i2)(1
)1(1
Observation:
• System ‘overshoots’, but still reaches new steady state.
• damped oscillations• The feedback to P
depends directly from P itself -> ‘auto-inhibitory feedback’
Kinetic rate laws and signalling motifs

ICYSB, 2013 Schaber 22/42
Modified conversion with negative feedback
Let’s do the math: Calculate steady-states of P(S) of a simplified system(all constants = 1)
PP
PS
dt
dP
1
)1(
261S-1-2
11
)1(0
SSP
PP
PS
In the steady-state:
Our analysis suggests that in a system with auto-inhibitory negative feedback, the steady-state depends on the input signal.

ICYSB, 2013 Schaber 23/42
The Mathematical Model
• Signal: high osmolarity• Hog1 de-sensitizes Sho1• Delayed auto-inhibitory
feedback• Michaelis-Menten kinetics (20
parameters)• Model was fitted to Hog1
activation data
Hao et al. (2007) Curr. Biol. 17

ICYSB, 2013 Schaber 24/42
Validation
Orig. 1 M KCl
Orig. 0.5 M KClOrig. 0.25 M KCl
Single shock Double shock
- Model fits data well. - Simulations show damped oscillations and increasing
steady states- possible spurious effects due to over-parameterization

ICYSB, 2013 Schaber 25/42
Predictions
Orig. P-Hog1
Orig. Sho1aOrig. Sho1i
0.4 M KCl tripple shock (0, 30, 60 min)

ICYSB, 2013 Schaber 26/42
Conclusion after first modelling round
• Auto-inhibitory feedback model fits single and double shock data well, but• shows increasing steady states with increasing
external osmolarity (not supported by data),• shows damped oscillations (not supported by data, but
might be due to over-parameterization)• Auto-inhibitory feedback model is not able to predict triple
shock experiment, because of desensitization.
Possible solution: a) the signal has to be removed by adaptation b) no desensitization

ICYSB, 2013 Schaber 27/42
Recalling the biology
Sln1Sho1
Pbs2
Hog1
P
P
Fps1
Glycerol
Hog1 P
Gpd1
Sln1Sho1
Hog1 P
• Increasing the ambientosmotic pressure leads to a rapid passive loss of water and cell skrinkage
• This leads to closure of the glycerol channel and activation of two parallel signaling branches that both activate Hog1.
• Activated Hog1 translocates to the nucleous and triggers production of enzyme that enhance glycerol prodction.• Increased glycerol equilibrates
water potential differences and forces water back into the cell leading to volume adaptation and HOG pathway deactivation.

ICYSB, 2013 Schaber 28/42
New Hypothesis
– The signal is the water potential difference (differences in osmolarity) rather than merely external osmolarity.
– The main feedback is via glycerol accumulation.

ICYSB, 2013 Schaber 29/42
The new word model
Signal
OuterOsmolarity
Hog1 P-Hog1
Glycerol
v3v1
v2
• Signal = OuterOmolarity-Glycerol
• 3 reactions with mass actions, i.e. 3 parameters

ICYSB, 2013 Schaber 30/42
The mathematical model
Signal
OuterOsmolarity
Hog1 P-Hog1
Glycerol
v3v1
v2
PHogkdt
dG
PHogkPHogGOkdt
PdHog
1
1)11)((1
3
21
– For simplicity we assume Hog1+Hog1P=1
– OuterOsmolarity (O) fixed input function.
– The feedback depends on increasing Glycerol, which stays even if Hog1P=0(𝐻𝑜𝑔1𝑃0 ,𝐺0 )=(0 ,𝑂0)

ICYSB, 2013 Schaber 31/42
Verification of the model
PHogk
PHogkPHogGOk
10
1)11)((0
3
21
Let’s do the math: Calculate steady-states
01
PHog
GO
In the steady-state:
1. In the steady state Hog1P=0 independent from the outer osmolarity.
Þ The system tracks the ‘desired’ steady state of Hog1P=0 independent from perturbations.
Þ Integral feedback property.Þ ‘Perfect adaptor’ (in Hog1P).
2. Internal glycerol concentration depends on the outer osmolarity.

ICYSB, 2013 Schaber 32/42
A block diagram of integral feedback control.
Yi T et al. PNAS 2000;97:4649-4653
©2000 by National Academy of Sciences
• Variable u is the input for a process with gain k.
• Difference between the actual output y1 and the steady-state, i.e. ‘desired’, output y0 represents the normalized output or error y.
• Integral control arises through the feedback loop in which the time integral of y, x, is fed back into the system.
• As a result, we have ẋ = y and y = 0 at steady-state for all u.
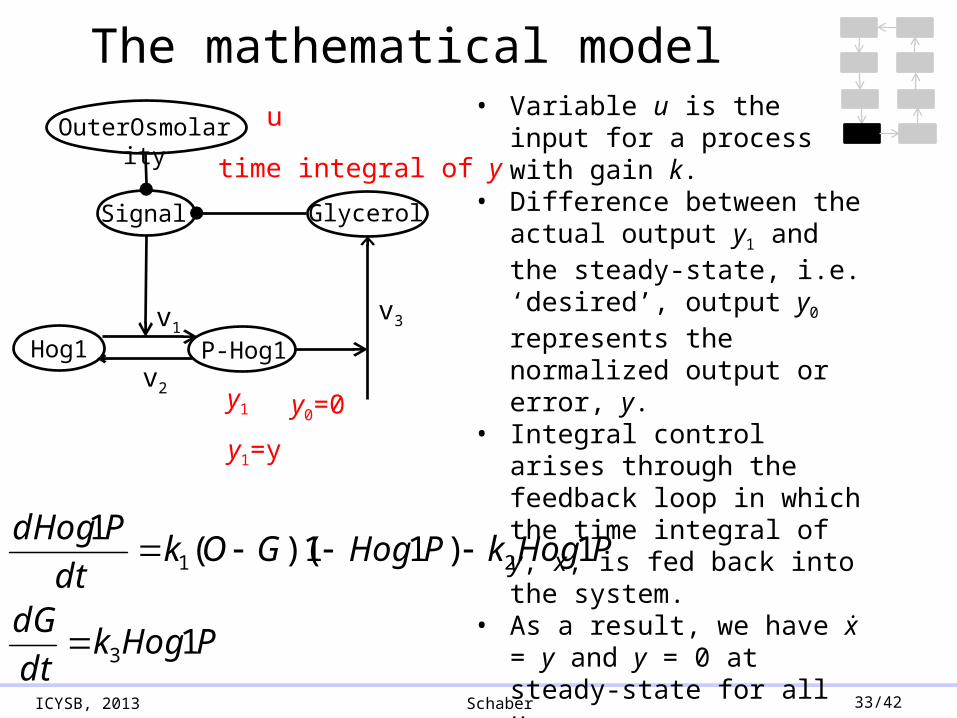
ICYSB, 2013 Schaber 33/42
The mathematical model
Signal
OuterOsmolarity
Hog1 P-Hog1
Glycerol
v3v1
v2
PHogkdt
dG
PHogkPHogGOkdt
PdHog
1
1)11)((1
3
21
• Variable u is the input for a process with gain k.
• Difference between the actual output y1 and the steady-state, i.e. ‘desired’, output y0 represents the normalized output or error, y.
• Integral control arises through the feedback loop in which the time integral of y, x, is fed back into the system.
• As a result, we have ẋ = y and y = 0 at steady-state for all u.
u
y1 y0=0
time integral of y
y1=y

ICYSB, 2013 Schaber 34/42
Validation of the model
Single shock
- Simulations show reasonable fits and perfect adaptation
Double shock
1 M KCl
0.5 M KCl0.25 M KCl

ICYSB, 2013 Schaber 35/42
Prediction
- Simple model reacts to triple shock and shows perfect adaptation.
P-Hog1l
InnerOsmolarityOuterOsmolarity
0.4 M KCl tripple shock (0, 30, 60 min)

ICYSB, 2013 Schaber 36/42
RemarksIntegral Feedback (3 parameters) Auto-inhibitory feedback (20 parameters)
• From the quality of the fit, the auto-inhibitory feedback model is much better than the integral feedback model.
• The quality of the fit is usually measured by the sum of squared residuals.
1 M KCl
0.5 M KCl0.25 M KCl
• If it weren’t for the prediction, which model is better?
n
ii
n
iii rptfyypSSR
1
2
1
2),((),(
r1
r2
r3

ICYSB, 2013 Schaber 37/42
RemarksIntegral Feedback (3 parameters) Transient feedback (20 parameters)
• Intuitively: both model capture the basic features of the data.
• In other words: The main information in the data is reproduced by the models.
• But: the information/parameter is much higher in the simpler model.
=> The parameters of the simpler model are more informative.
1 M KCl
0.5 M KCl0.25 M KCl

ICYSB, 2013 Schaber 38/42
The principle of parsimony• If we take the number of parameters as a measure for structural
properties of the data, we want to have few parameters, i.e. the most important structural features, and a good data representation.
“Everything should be madeas simple as possible, but not simpler.”
• Parsimony can be measured by the Akaike Information Criterion (AIC). nSSRnkAIC log2
• the lower AIC, the better the model approximates the data in terms of parsimony (k number of parameters, n number of data).
• We do not want to have additional parameter to ‘fit the errors’ or spurious effects.
• The simpler the model, the easier to analyse.
• Therefore, it is advisable to have a model that is as simple as possible and as complex as necessary: this is the principle of parsimony.
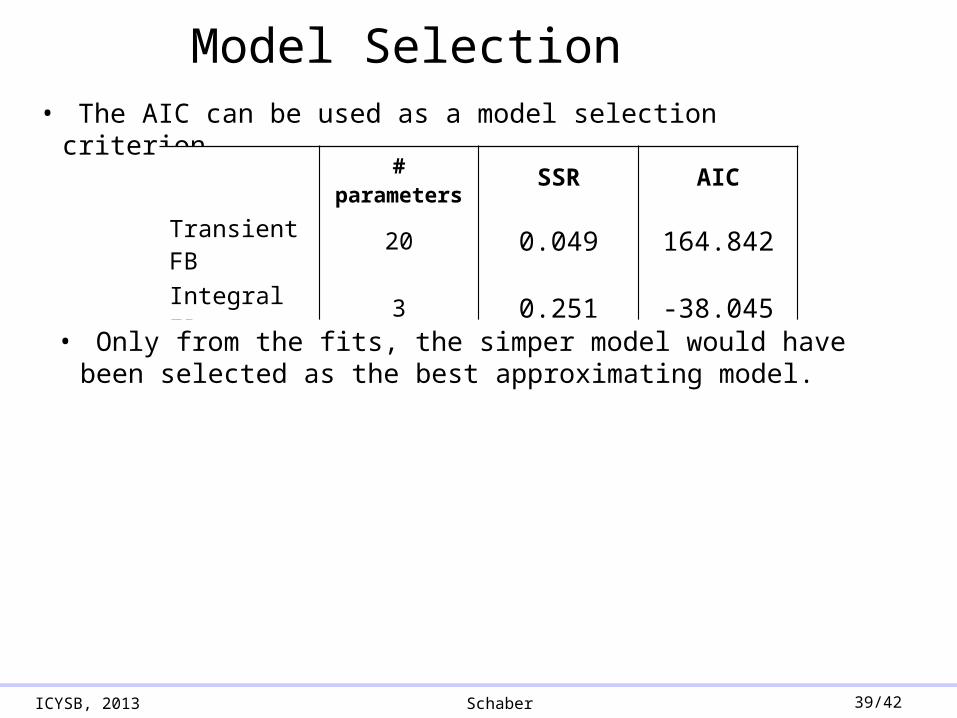
ICYSB, 2013 Schaber 39/42
Model Selection• The AIC can be used as a model selection criterion
# parameters SSR AIC
Transient FB 20 0.049 164.842
Integral FB 3 0.251 -38.045
• Only from the fits, the simper model would have been selected as the best approximating model.

ICYSB, 2013 Schaber 40/42
Conclusions concerning feedback in Sho1-branch
• According to the AIC, the data does not support a model with auto-inhibitory feedback, but rather a model with integral feedback.
• Therefore, for the adaptation and attenuation process, the proposed feedback of Hog1 on Sho1 is not necessary.
• The proposed feedback of Hog1 to Sho1 may modulate the signal and serve other purposes (crosstalk, stabilisation), but cannot explain adaptation to single and multiple shock.

ICYSB, 2013 Schaber 41/42
Final Remarks on Sho1-Modelling
• The integral feedback model has several shortcoming:
• glycerol is only accumulated, never lost.Þ Adaptation to a lowering in external osmolarity cannot be
modelled
• initial Hog1P is zero
All models are wrong, but some are useful.
• Our model was developed to address the question, whether or not the (multiple) osmotic shock data can be explained by a transient feedback, nothing else.
• All models are tailored to address specific questions. No model can explain everything.
• The clearer our hypotheses are formulated, the better models can be developed to address these.

ICYSB, 2013 Schaber 42/42
Final Remarks on Modelling
• The ‘truth’ (full reality) in biological sciences has infinite complexity and, hence, can never be revealed with only finite samples and a ‘model’ of those data.
• It is a mistake to believe that there is a simple “true model” and that during data analysis this model can be uncovered and its parameters estimated.
• We can only hope to identify a model that provides a good approximation to the data available.
• Uncertainty about the biology leads to multiple hypotheses about the underlying processes explaining a set of data.
• I recommend the formulation of multiple working hypotheses, and the building of a small set of models to clearly and uniquely represent these hypotheses.
• The best approximating model can then be identified by model selection criteria.


















