
Hybrid synapsesneurons: Amodelfor competitive reinnervation
5
Proc. Nati. Acad. Sci. USA Vol. 76, No. 9, pp. 4695-4699, September 1979 Neurobiology "Hybrid" synapses formed by foreign innervation of parasympathetic neurons: A model for selectivity during competitive reinnervation (regeneration/synaptic specificity/neuronal plasticity/cardiac ganglion) WILLIAM PROCTOR*, SAMY FRENK*, BARBARA TAYLOR*, AND STEPHEN ROPER*tt Departments of *Anatomy and tPhysiology, University of Colorado Health Sciences Center, Denver, Colorado 80262 Communicated by Stephen W. Kuffler, June 8, 1979 ABSTRACT Selectivity of synapse formation after nerve regeneration was tested in the parasympathetic cardiac gan- glion of frogs (Rana pipiens). First, we tested the ability of so- matic motor axons to establish synaptic connections with de- nervated ganglion cells by implanting the hypoglossus nerve into the vagotomized heart. After several weeks, stimulation of the implanted hy oglossus mediated a parasympathetic-like inhi- bition of the heart rate, and synaptic responses produced by hypoglossal stimulation were recorded intracellularly in gan- glion cells. Light and electron microscopy indicated that im- planted hypoglossal nerve terminals contacted parasympathetic ganglion cells only on their axons and not on the cell body (where most vagal synapses are found in control animals). Sec- ond, we tested whether regenerating vagal preganglionic axons would compete with foreign (hypoglossal) terminals for inner- vation of cardiac ganglion cells. We allowed the vagus nerve to regenerate in animals in which the implanted hypoglossus had established functional contacts with the cardiac ganglion. Vagal axons were able to reinnervate the heart and reestablish syn- aptic connections on the cell bodies of ganglion cells. Further- more, functional transmission at the foreign (hypoglossal) ter- minals disappeared concomitant with vagal reinnervation. One of the more salient problems concerning repair of damage in nervous tissue is the ability of regenerating axons to rees- tablish synaptic connections with appropriate postsynaptic cells. The mechanisms underlying the selectivity of synapse forma- tion during reinnervation are largely unknown, and as yet there is no explanation why regenerating axons reestablish appro- priate synaptic corrections in some cases but not in others. For example, we do not know why reinnervation of decentralized mammalian sympathetic ganglia accurately restores functional connections (1, 2), whereas reinnervation of mammalian skel- etal muscle is largely nonselective (3, 4). Although attempts have been made to explain selectivity during reinnervation, such as the existence of a specific "chemoaffinity" between nerve terminals and their appropriate targets (5), it has been difficult to test these ideas critically, and in brief, the cellular basis of selective synapse formation after nervous tissue damage still remains an enigma. We have been studying selectivity of synapse formation during reinnervation of parasympathetic neurons in the am- phibian cardiac ganglion to extend our understanding of neu- ronal interactions during regeneration. In particular, we have asked (i) whether a somatic motor nerve (i.e., a nerve supply quite foreign to parasympathetic ganglia) will establish func- tional connections with denervated autonomic neurons in the heart of the frog, and (ii), if so, whether regenerating vagal axons will successfully compete with foreign inputs to rein- nervate ganglion cells and thereby reveal specificity of synapse formation during this process. This report describes physiological and morphological characteristics of synaptic connections between parasympa- thetic neurons and regenerating preganglionic axons, and be- tween parasympathetic neurons and implanted somatic motor axons. METHODS Surgical Procedures. To implant a somatic motor nerve into the parasympathetic cardiac ganglion in the frog; we anasto- mosed the central end of the first spinal nerve, the hypoglossus, to the distal stump of the vagus nerve leading to the heart as follows: Frogs (Rana pipiens, northern variety) were anesthe- tized with tricaine (MS 222) (Sigma, 100 ,g/g of body weight, injected into the dorsal lymph sac) and the vagus nerves on both sides were exposed. We sectioned the vagus nerves approxi- mately 10 mm from their point of exit from the brain and su- tured the central ends into the skin to retard regeneration. The left hypoglossus was then sectioned about 5 mm from its exit at the spinal cord and the central end was sutured to the distal stump of the left vagus nerve with 10-0 monofilament nylon. In some animals, we reanesthetized the frog and sectioned the vagus nerves central to the original cut 5 days to 6 weeks before characterizing foreign innervation to exclude any spurious vagal reinnervation. Operated animals were returned to tanks with running water for recovery and kept for 8 to 52 weeks. During this time they were force-fed a slurry of liver, cooked lettuce, cod liver oil, and multivitamins. About half the animals survived the operation. Electrophysiological Recordings. The techniques for iso- lating the ganglion, impaling single cells, and measuring evoked intracellular responses are discussed more fully by Dennis et al. (6) and Roper and Ko (7) and will be described here only briefly. The heart, along with both vagus nerves, and the im- planted left hypoglossal nerve were dissected free of the animal, placed in Ringer's solution (112 mM NaCl/5 mM CaCl2/2 mM KC1/3 mM Hepes buffer, pH 7.2), and the interatrial septum containing the cardiac ganglion was exposed and pinned out in a shallow chamber. The vagal and implanted hypoglossal nerves were drawn into separate suction electrodes for stimu- lation. Neurons were impaled with fine-tipped microelectrodes (100-300 MQ resistance). In some experiments the heart rate was recorded in intact but pithed frogs by making a small incision in the skin overlying the heart and inserting a fine stainless steel wire, insulated ex- cept at its tip. Another stainless steel wire was inserted under the skin in one of the legs for a reference electrode. The elec- trodes were fed into a high gain ac amplifier and the heart rate was recorded on a pen recorder. f To whom reprint requests should be addressed. 4695 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "ad- vertisement" in accordance with 18 U. S. C. §1734 solely to indicate this fact.
Transcript of Hybrid synapsesneurons: Amodelfor competitive reinnervation

Proc. Nati. Acad. Sci. USAVol. 76, No. 9, pp. 4695-4699, September 1979Neurobiology
"Hybrid" synapses formed by foreign innervation ofparasympathetic neurons: A model for selectivityduring competitive reinnervation
(regeneration/synaptic specificity/neuronal plasticity/cardiac ganglion)
WILLIAM PROCTOR*, SAMY FRENK*, BARBARA TAYLOR*, AND STEPHEN ROPER*ttDepartments of *Anatomy and tPhysiology, University of Colorado Health Sciences Center, Denver, Colorado 80262
Communicated by Stephen W. Kuffler, June 8, 1979
ABSTRACT Selectivity of synapse formation after nerveregeneration was tested in the parasympathetic cardiac gan-glion of frogs (Rana pipiens). First, we tested the ability of so-matic motor axons to establish synaptic connections with de-nervated ganglion cells by implanting the hypoglossus nerve intothe vagotomized heart. After several weeks, stimulation of theimplanted hy oglossus mediated a parasympathetic-like inhi-bition of the heart rate, and synaptic responses produced byhypoglossal stimulation were recorded intracellularly in gan-glion cells. Light and electron microscopy indicated that im-planted hypoglossal nerve terminals contacted parasympatheticganglion cells only on their axons and not on the cell body(where most vagal synapses are found in control animals). Sec-ond, we tested whether regenerating vagal preganglionic axonswould compete with foreign (hypoglossal) terminals for inner-vation of cardiac ganglion cells. We allowed the vagus nerve toregenerate in animals in which the implanted hypoglossus hadestablished functional contacts with the cardiac ganglion. Vagalaxons were able to reinnervate the heart and reestablish syn-aptic connections on the cell bodies of ganglion cells. Further-more, functional transmission at the foreign (hypoglossal) ter-minals disappeared concomitant with vagal reinnervation.
One of the more salient problems concerning repair of damagein nervous tissue is the ability of regenerating axons to rees-tablish synaptic connections with appropriate postsynaptic cells.The mechanisms underlying the selectivity of synapse forma-tion during reinnervation are largely unknown, and as yet thereis no explanation why regenerating axons reestablish appro-priate synaptic corrections in some cases but not in others. Forexample, we do not know why reinnervation of decentralizedmammalian sympathetic ganglia accurately restores functionalconnections (1, 2), whereas reinnervation of mammalian skel-etal muscle is largely nonselective (3, 4). Although attemptshave been made to explain selectivity during reinnervation,such as the existence of a specific "chemoaffinity" betweennerve terminals and their appropriate targets (5), it has beendifficult to test these ideas critically, and in brief, the cellularbasis of selective synapse formation after nervous tissue damagestill remains an enigma.We have been studying selectivity of synapse formation
during reinnervation of parasympathetic neurons in the am-phibian cardiac ganglion to extend our understanding of neu-ronal interactions during regeneration. In particular, we haveasked (i) whether a somatic motor nerve (i.e., a nerve supplyquite foreign to parasympathetic ganglia) will establish func-tional connections with denervated autonomic neurons in theheart of the frog, and (ii), if so, whether regenerating vagalaxons will successfully compete with foreign inputs to rein-nervate ganglion cells and thereby reveal specificity of synapseformation during this process.
This report describes physiological and morphologicalcharacteristics of synaptic connections between parasympa-thetic neurons and regenerating preganglionic axons, and be-tween parasympathetic neurons and implanted somatic motoraxons.
METHODSSurgical Procedures. To implant a somatic motor nerve into
the parasympathetic cardiac ganglion in the frog; we anasto-mosed the central end of the first spinal nerve, the hypoglossus,to the distal stump of the vagus nerve leading to the heart asfollows: Frogs (Rana pipiens, northern variety) were anesthe-tized with tricaine (MS 222) (Sigma, 100 ,g/g of body weight,injected into the dorsal lymph sac) and the vagus nerves on bothsides were exposed. We sectioned the vagus nerves approxi-mately 10 mm from their point of exit from the brain and su-tured the central ends into the skin to retard regeneration. Theleft hypoglossus was then sectioned about 5 mm from its exitat the spinal cord and the central end was sutured to the distalstump of the left vagus nerve with 10-0 monofilament nylon.In some animals, we reanesthetized the frog and sectioned thevagus nerves central to the original cut 5 days to 6 weeks beforecharacterizing foreign innervation to exclude any spurious vagalreinnervation. Operated animals were returned to tanks withrunning water for recovery and kept for 8 to 52 weeks. Duringthis time they were force-fed a slurry of liver, cooked lettuce,cod liver oil, and multivitamins. About half the animals survivedthe operation.
Electrophysiological Recordings. The techniques for iso-lating the ganglion, impaling single cells, and measuring evokedintracellular responses are discussed more fully by Dennis etal. (6) and Roper and Ko (7) and will be described here onlybriefly. The heart, along with both vagus nerves, and the im-planted left hypoglossal nerve were dissected free of the animal,placed in Ringer's solution (112 mM NaCl/5 mM CaCl2/2 mMKC1/3 mM Hepes buffer, pH 7.2), and the interatrial septumcontaining the cardiac ganglion was exposed and pinned outin a shallow chamber. The vagal and implanted hypoglossalnerves were drawn into separate suction electrodes for stimu-lation. Neurons were impaled with fine-tipped microelectrodes(100-300 MQ resistance).
In some experiments the heart rate was recorded in intact butpithed frogs by making a small incision in the skin overlyingthe heart and inserting a fine stainless steel wire, insulated ex-cept at its tip. Another stainless steel wire was inserted underthe skin in one of the legs for a reference electrode. The elec-trodes were fed into a high gain ac amplifier and the heart ratewas recorded on a pen recorder.
f To whom reprint requests should be addressed.
4695
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.

4696 Neurobiology: Proctor et al.
A
I30r 20O/ I 0 V
B
TI
0 l
C{
C
vI5E I( V GO sec
FIG. 1. Electrocardiograms from three frogs, showing the effectof stimulating vagal and hypoglossal nerve roots. Trains of stimuli(2.5 msec duration pulses, 10 Hz) were applied to the nerves with silverwire bipolar hook electrodes near the brain and spinal cord. The in-tensity (volts) and duration of stimulation are shown below the re-
cordings. The animals were pithed prior to exposure of the nerve roots.(A) Control unoperated frog. Vagal stimulation immediately stoppedthe heart, whereas hypoglossal stimulation had no measurable effect.(B) Experimental animal in which the hypoglossus had been surgicallyredirected to the denervated heart 12 weeks before. Hypoglossalstimulation produced a parasympathetic-like bradycardia. Vagalregeneration had been prevented in this animal (see text). (C) Ex-perimental animal in which the hypoglossal nerve had been implantedin the heart and vagal axons had been allowed to regenerate (see text).The surgical operation had been performed 29 weeks before.
Morphological Techniques. For light microscopy, the tissuewas stained with a modified zinc iodide/osmium technique (M.Letinsky, personal communication) as follows: equal amountsof zinc iodide solution (3 g of zinc powder plus 1 g of iodinecrystals in 40 ml of distilled water) and 0.2 M sodium acetatebuffer (pH 4.0) were mixed, and 0.5 ml of 4% osmium tetroxide(Polysciences, Warrington, PA) was added to 2 ml of the buf-fered zinc iodide solution. The mixture was then immediatelyadded to the tissue and left for 60-90 min. Last, the tissue wasrinsed with Ringer's solution, dehydrated, cleared, andmounted. Whole-mount preparations were observed andphotographed.
Procedures for ultrastructural analysis of the cardiac gangliawere similar to those in McMahan and Kuffler (8). Ganglia werefixed in glutaraldehyde and phosphate buffer (pH 7.2) andpostfixed in osmium tetroxide. The tissue was dehydrated andmounted in thin wafers of Araldite 6005. Thin sections werecollected on slot grids and stained with lead citrate and uranylacetate and viewed with a Philips 300 electron microscope.
RESULTSIn frogs in which the hypoglossal nerve was directed to thedenervated heart, we tested whether somatic motor axons hadformed functional synapses with parasympathetic cardiacganglion cells. Eight to 52 weeks after the anastomosis, we
studied the extent of foreign innervation in the heart by twomethods.
FIG. 2. Intracellularly recorded synaptic responses from para-sympathetic neurons in the isolated frog cardiac ganglion. Threedifferent preparations are shown. (A) Postsynaptic potentials evokedby stimulating the preganglionic (vagus) nerve in control unoperatedanimals. Two traces are superimposed. Increasing the stimulus in-tensity to the vagus produced two synaptic responses, a subthresholdand a suprathreshold one, indicating that at least two separate pre-ganglionic fibers innervated this neuron. The residual depolarizationafter the action potential is caused by the intense and prolonged actionof the transmitter at vagal synapses and is typical for these neurons(see ref. 6). (B) Synaptic potentials produced by stimulating a hy-poglossal nerve that had been surgically redirected into the cardiacganglion 16 weeks prior to the experiment. Two traces are superim-posed. Both sub- and suprathreshold responses were evoked byvarying the stimulus intensity to the implanted hypoglossus. (C)Postsynaptic responses in a neuron innervated by a regenerating vagalaxon (upper trace) and an implanted hypoglossal axon (lower trace).The anastomosis had been performed 31 weeks before, and the vagushad been allowed to regenerate (see text). The conduction velocityof the vagal fibers was consistently slower than that of implantedhypoglossal axons (the nerve lengths were similar in all the tracingsshown in this figure).
First, animals were pithed, the vagal nerves and left hypo-glossal nerve were exposed near their exits from the brain andspinal cord, respectively, and the nerve roots were stimulatedwith fine hook electrodes while the electrocardiogram wasmonitored. In control unoperated animals, weak stimulationapplied to either vagus nerve (5-10 V at 10 Hz) readily causedthe heart to slow and stop beating (Fig. 1A). In experimentalanimals, vagal stimulation central to the transection had noeffect on the heart rate, consistent with complete vagotomy ofthe heart. However, stimulation of the implanted left hypo-glossal nerve slowed and stopped the heartbeat, indicating thatmotor axons had formed functional synapses with the dener-vated heart (Fig. 1B).
Second, to examine hypoglossal innervation more precisely,
A
I1
20 msec
20 mV
:; :i..:..;.I;.., I.- I-1: ;::;!..
Proc. Natl. Acad. Sci. USA 76 (1979)
Ik; :I .I; ;;
Neurobiology: Proctor et al.
we isolated the cardiac ganglion for intracellular recording fromindividual parasympathetic neurons. Fig. 2B shows intracellularresponses from parasympathetic neurons, produced by stimu-lating the implanted hypoglossus. In 76% of the neurons (359neurons from 15 ganglia) we recorded intracellular synapticresponses produced by stimulating the hypoglossal nerve,showing that somatic motor axons were indeed capable of in-nervating autonomic neurons, and, furthermore, that they es-tablished functional connections with a majority of the ganglioncells. Typically the responses evoked by stimulating the hypo-glossus were less than 5 mV and in only 9% of the innervatedneurons were the responses large enough to reach threshold andproduce an action potential.
Proc. Natl. Acad. Sci. USA 76 (1979) 4697
In contrast, when we recorded synaptic responses producedby preganglionic stimulation in unoperated control animals weobserved postsynaptic responses in 99% of the cells; 86% of thesecontrol neurons were innervated by one or more suprathresholdinputs (Fig. 2A). Similarly, reinnervation of denervated gan-glion cells by vagal preganglionic axons rapidly and completelyrestored ganglionic transmission (9, 10).We examined control and experimental ganglia with light
and electron microscopy to characterize the morphology of theforeign (hypoglossal) nerve terminals and to compare these withvagal endings in unoperated animals. Light microscopy of zinciodide/osmium-stained preparations revealed darkly stainedunmyelinated and varicose nerve terminals that contacted
4Lttr .
tF..4CrW hi -v -. Vae *e *WWsSax:'
44)~~~~~~~~~~~~~~4
IA~~~~~~A
-IC9
4>0# / r 2, - 4j~i * It 9 * J'/ *4crv--t-lF,>g o < S * 9 x ; 08 ftt;;/*9
r ~~~~S . i ~ _ _ .- ' iP~~~- t .s*; ^@ww- s~*4 Wf 963
'O - a .
to- 2!. w.-15~~~~~~
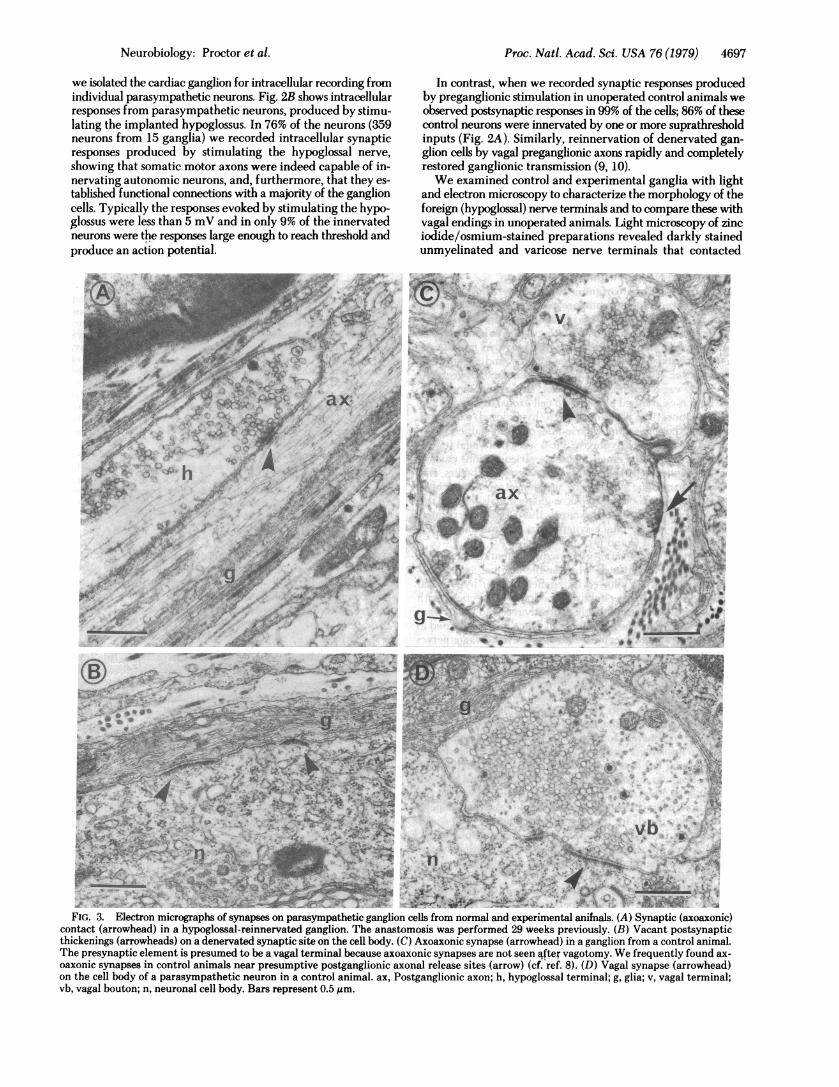
FIG.- 3. Electron micrographs of synapses on parasympathetic ganglion cells from normal and experimental aniMals. (A) Synaptic (axoaxonic)contctarrwhed) n ahypoglossal-reinnervated ganglion. The anastomosis was performed 29 weeks previously. (B) Vacant postsynaptic
thickenings (arrowheads) on a denervated synaptic site on the cell body. (C) Axoaxonic synapse (arrowhead) in a ganglion from a control animal.The presynaptic element is presumed to be a vagal terminal because axoaxonic synapses are not seen q~ftervagotomy. We frequently found ax-oaxonic synapses in control animals near presumptive postganglionic axonal release sites (arrow) (cf. ref. 8). (D) Vagal synapse (arrowhead)on the cell body of a parasympathetic neuron in a control animal. ax, Postganglionic axon; h, hypoglossal terminal; g, glia; v, vagal terminal;vb, vagal bouton; n, neuronal cell body. Bars represent 0.5 pum.
folb.itl10.

4698 Neurobiology: Proctor et al.
postganglionic axons, but did not form boutons on the cellbodies such as those found in control ganglia. Often, the hy-poglossal terminals would branch extensively and some col-laterals appeared to contact surrounding cardiac muscle fiberstrands.
At the electron microscope level, we found only axoaxonicsynapses on parasympathetic neurons in hypoglossal-reinner-vated ganglia (Fig. 3A). These synapses contained clusters ofpresynaptic vesicles and pre- and postsynaptic membranethickenings. No synapses were found on cell bodies even thoughvacant postsynaptic thickenings were present there (Fig. 3B).On the other hand, vagal boutons in control unoperated gangliacontacted cell bodies as well as axons (Fig. 3 C and D). In nor-mal animals, complete vagotorny eliminated both axosomaticand axoaxonic synapses within at least 10 days.
As a further control to examine the characteristics of regen-erating synapses and to compare reinnervation by foreignterminals to reinnervation by original ones, we inspected gan-glia from animals in which only the vagus nerves had been cutor crushed and allowed to return to the heart. As mentionedabove, functional transmission was rapidly and completelyrestored after vagal regeneration. Initial contact between re-generating vagal terminals and denervated parasympatheticneurons was established on ganglion cell axons, as was also thecase for connections between foreign (hypoglossal) axons andparasympathetic neurons. Zinc iodide/osmium-stained prep-arations during the early stages (3 to 5 weeks) of vagal regen-eration showed many small-diameter varicose axons passingclose to ganglion cell axons without contacting cell bodies, andwe found axoaxonic synapses by using electron microscopy.However, within about 10 weeks, vagal terminals reached thecell bodies and reestablished synapses there; these regeneratedsynapses resembled intact synapses in control unoperatedganglia (see Fig. 3 C and D).To examine the specificity of synapse formation during re-
generation, we next tested whether the original axons (pre-ganglionic vagal), if allowed to regenerate after hypoglossalimplantation, would reestablish functional connections withparasympathetic neurons that had already received inputs fromthe foreign nerve. The same surgical procedures were used asbefore, except that the central ends of the severed vagus nerveswere not tied to the skin, but instead were left in place and al-lowed to regenerate.Our preliminary test for vagal or foreign innervation of the
heart again consisted of stimulating vagal or hypoglossal nerves,or both, near their exits from the central nervous system inpithed, but otherwise intact, frogs. When the vagal axons wereallowed to regenerate for several weeks after the hypoglossushad been implanted, vagal stimulation was effective in slowingand stopping the heart beat. On the other hand, hypoglossalstimulation was no longer as effective as before and had onlya slight or no effect on the heart rate (Fig. IC). This was our firstindication that regenerating vagal axons successfully competedwith foreign inputs for innervation of the heart.
Next, cardiac ganglia were dissected free and placed in achamber for intracellular recordings. It was immediately clearthat regenerating vagal axons reestablished synaptic connectionswith ganglion cells, even if the neurons were also contacted byhypoglossal axons (Fig. 2C). Furthermore, as the percentageof cells innervated by the regenerating vagus nerves increasedwith time, the percent innervated by foreign, hypoglossal, axonscorrespondingly decreased. This relationship is shown in Fig.4.
Using electron and light microscopy as before, we then de-tected synaptic endings located on the cell bodies. These closelyresembled vagal boutons from control animals in which the
75
CC
c 500)WAn0
0 8-20 21-30 31-52 31-52weeks weeks weeks weeks
(5) (7) (3) (3)
FIG. 4. Competition between foreign (hypoglossal) inputs andregenerating vagal preganglionic fibers for parasympathet'i neuronsin the frog cardiac ganglion. To obtain these data, the hypoglossuson one side was anastomosed to the distal stump of the vagus (seetext). At the same time, the contralateral vagus was resected to delay,but not prevent, its regeneration. Open bars show the percentage ofneurons innervated by the hypoglossus and hatched bars show thepercentage innervated by the vagus. Experiment A, data from an earlystage of vagal reinnervation (8-20 weeks after the operation) but ata time when hypoglossal innervation had been established. Experi-ment B, middle stage. Experiment C, late stage. In experiment C' theregenerating vagus was resected in a second operation 1-6 weeks be-fore recording ganglionic innervation to demonstrate that hypoglossalinput did not decline even at long intervals after the anastomosis(31-52 weeks) unless vagal axons were allowed to reinnervate theparasympathetic neurons. Thus, the decline of foreign innervationshown in experiments A-C is truly correlated with vagal reinnervation.The numbers of animals are shown in parentheses below each dataset, and standard errors are indicated above each bar.
vagus nerve regenerated (without implantation of the hypo-glossus). We have not yet determined the fate of the axoaxonichypoglossal synapses by either light or electron microscopy andthus do not know whether foreign nerve terminals become in-active, withdraw, or degenerate.
DISCUSSIONThese data show that the hypoglossus can establish functionalsynapses with denervated parasympathetic neurons. We willrefer to these as "hybrid" synapses because the presynapticelement is a somatic motor axon and the postsynaptic cell is anautonomic neuron. Most of the responses produced by the hy-brid contacts recorded intracellularly from the cell bodies weresmall.§ Other investigators have found that motor axons caninnervate autonomic neurons in mammals, and that synapticinteractions at these sites are weak also (11, 12).Two important findings from our experiments are (i) foreign
(hypoglossgl) inputs contact the neurons only on their axons,and not on the cell body region, and (ii) regenerating vagalaxons successfully compete for parasympathetic neurons andforeign innervation is lost.
These two observations may be related; the finding that
§ Nevertheless, even though most of the synaptic responses were sub-threshold, hypoglossal innervation of the heart in our experimentswas capable of mediating a parasympathetic-like inhibition of theheart beat. We suspect that hypoglossal motor axons, in addition toinnervating parasympathetic ganglion cells, also influenced cardiacmuscle fibers directly, and this could explain the ability of the im-planted foreign nerve to slow and stop the heart. Alternatively, thesmall proportion of suprathreshold hypoglossal inputs to the cardiacganglion (average = 9% of the innervated neurons) may have beensufficient to mediate reflex cardiac inhibition.
Proc. Nati. Acad. Sci. USA 76 (1979)

Proc. Natl. Acad. Sci. USA 76 (1979) 4699
FIG. 5. Schematic drawing of a parasympathetic neuron in theheart of the frog, based on light microscopy of zinc iodide/osmium-stained preparations and on electron microscopy of glutaraldehyde-stained ganglia, and summarizing the sequence of events that occurafter denervation and reinnervation by appropriate (vagal) axons(A-B-C2) or by foreign (hypoglossal) nerve fibers (A-B-Cl). A glialcell (g) envelops the entire cell body and forms a complex trabecularnetwork around the axon hillock. One or more preganglionic vagalaxons in a control animal (A) spiral around the axon hillock, betweenthe glial processes, before depositing synaptic boutons on the cellbody. In B, regenerating nerve terminals first establish synapticcontact with the parasympathetic neuron along its axon, at somedistance from the cell body. C1 shows the subsequent step in the re-generation of foreign axons: terminals with varicosities and growthswellings branch and proliferate within the ganglion but do not enterthe glial trabecular network at the axon hillock. C2 illustrates the finalregeneration sequence for appropriate (vagal) axons. They grow
through the glial trabeculae and reinnervate the cell body, in markedcontrast to regenerating foreign nerve axons (CI). g, Glial cell (stip-pled); n, nucleus of glial cell; a, axon of preganglionic nerve; r, regen-
erating axon.
foreign inputs establish only axoaxonic synapses might be im-portant to understanding selective reinnervation. Although wedo not know why inappropriate (hypoglossal) axons do not gainaccess to denervated cell bodies in the frog cardiac ganglion andwhy regenerating appropriate ones (vagus nerves) do reestablishsynapses on the soma, we speculate that access to postsynapticsites on the cell body is some measure of the compatibility ofpresynaptic axon and postsynaptic target neuron. Access to thesoma, for example, may be controlled by the glial cell that in-vests each ganglion cell (8) and that forms a complex trabecularnetwork at the axon hillock (13, 14). The glial cell trabeculae
that surround the emergence of the ganglion cell axon arecovered by a cell coat that resembles a thick basal lamina. Incontrol and reinnervated ganglia, axons spiral around the axonhillock within these glial trabeculae before they deposit boutonson the cell body and therefore the vagal terminals are in closeapproximation to the glial processes and their cell coats (refs.8 and 13; unpublished observations). Foreign somatic axons(hypoglossal) do not enter this glial trabecular complex and donot contact the cell body. Others have shown the importanceof basal lamina in guiding regenerating nerve terminals backto the appropriate postsynaptic sites and in directing theirsubsequent maturation (15, 16), and we speculate that the glialtrabecular complex along with its cell coat allows appropriate(vagal) regenerating axons, but not foreign ones, access to thecell body. This is schematized in Fig. 5.
In summary, the cellular events that take place during re-generation of axons to the parasympathetic cardiac ganglionin the frog appear to be as follows: somatic motor axons or vagalpreganglionic fibers reach the heart via the original vagal nervestumps, which guide regenerating axons into the heart near thesinus venosus. When the hypoglossal axons reach the ganglionthey form many unmyelinated varicose branches, and some ofthese terminals contact ganglion cell axons. The motor terminalsremain functional unless vagal axons regenerate. When vagalaxons regenerate to the denervated heart, they also establishinitial contact with ganglion cell axons. However, in contrastto foreign inputs, they continue to grow to the cell body andpenetrate the glial trabecular complex to deposit boutons on thecell body and restore vagal function. At the same time, trans-mission from foreign innervation is lost. We do not as yet knowthe fate of the foreign nerve terminals when they cease trans-mitting.
We thank Drs. T. Finger, A. Herrera, and D. Purves for their assis-tance in preparing the manuscript, and we acknowledge the supportof the National Institutes of Health (Grants 5 R01 NS11505, 5 K04NS00257, and 1 F32 NS05283).
1. Langley, J. N. (1897) J. Physiol. 22, 215-230.2. Nja, A. & Purves, D. (1977) J. Physiol. 272,633-651.3. Bernstein, J. J. & Guth, L. (1961) Exp. Neurol. 4, 262-275.4. Kimura, J., Rodnitzky, R. L. & Okawara, S.-H. (1975) Neurology
25,989-993.5. Sperry, R. W. (1963) Proc. Natl. Acad. Sci. USA 50,703-710.6. Dennis, M. J., Harris, A. J. & Kuffler, S. W. (1971) Proc. R. Soc.
London Ser. B 177,509-539.7. Roper, S. & Ko, C.-P. (1978) in Neuronal Plasticity, ed. Cotman,
C. W. (Raven, New York), pp. 1-25.8. McMahan, U. J. & Kuffler, S. W. (1971) Proc. R. Soc. London Ser.
B 177, 485-508.9. Roper, S. (1976) Nature (London) 261, 148-149.
10. Ko, C.-P. & Roper, S. (1978) Nature (London) 274, 286-288.11. McLachlan, E. M. (1974) J. Physiol. 237, 217-242.12. Ostberg, A.-J. C., Raisman, G., Field, P. M., Iversen, L. L. &
Zigmond, R. E. (1976) Brain Res. 107, 445-470.13. Pick, J. (1963) J. Comp. Neurol. 120,409-462.14. Taxi, J. (1976) in Frog Neurobiology, ed. Llinas, R. & Precht, W.
(Springer, Berlin), pp. 93-150.15. Letinsky, M., Fischbeck, K. & McMahan, U. F. (1976) J. Neu-
rocytol. 5, 691-718.16. Sanes, J. R., Marshall, L. M. & McMahan, U. J. (1978) J. Cell Biol.
78, 176-198.
Neurobiology: Proctor et al.










![05[2] Strategy competitors, competitive rivalry, competitive behavior, and competitive dynamics · PDF fileDefine competitors, competitive rivalry, competitive behavior, and competitive](https://static.fdocuments.us/doc/165x107/5aa5b2eb7f8b9ac8748d8018/052-strategy-competitors-competitive-rivalry-competitive-behavior-and-competitive.jpg)







