bounded gaps between primes - D©partement de math©matiques et

Human intestinal epithelial cell survival:differentiation state-specific control mechanisms
REMY GAUTHIER,1 CHARLENE HARNOIS,1 JEAN-FRANCOIS DROLET,1
JOHN C. REED,2 ANNE VEZINA,1 AND PIERRE H. VACHON1,3
1Canadian Institutes of Health Research Group on the Functional Development and Physiopathologyof the Digestive Tract, Departement d’Anatomie et de Biologie Cellulaire, Faculte de Medecine,Universite de Sherbrooke, Sherbrooke, Quebec, Canada J1H 5N4; 2The Burnham Institute,La Jolla Cancer Research Center, La Jolla, California 92037; and 3Thematique de Rechercheen Physiopathololgie Digestive du Centre de Recherches Cliniques du CHUS,Fleurimont, Quebec, Canada J1H 5N4Received 5 December 2000; accepted in final form 17 January 2001
Gauthier, Remy, Charlene Harnois, Jean-FrancoisDrolet, John C. Reed, Anne Vezina, and Pierre H. Va-chon. Human intestinal epithelial cell survival: differentia-tion state-specific control mechanisms. Am J Physiol CellPhysiol 280: C1540–C1554, 2001.—To investigate whetherhuman intestinal epithelial cell survival involves distinctcontrol mechanisms depending on the state of differentiation,we analyzed the in vitro effects of insulin, pharmacologicalinhibitors of Fak, MEK/Erk, and PI3-K/Akt, and integrin (b1,b4)-blocking antibodies on the survival of the well-estab-lished human Caco-2 enterocyte-like and HIEC-6 cryptlikecell models. In addition, relative expression levels of six Bcl-2homologs (Bcl-2, Bcl-XL, Mcl-1, Bax, Bak, and Bad) andactivation levels of Fak, Erk-2, and Akt were analyzed.Herein, we report that 1) the enterocytic differentiation pro-cess results in the establishment of distinct profiles of Bcl-2homolog expression levels, as well as p125Fak, p42Erk-2, andp57Akt activated levels; 2) the inhibition of Fak, of the MEK/Erk pathway, or of PI3-K, have distinct impacts on entero-cytic cell survival in undifferentiated (subconfluent Caco-2,confluent HIEC-6) and differentiated (30 days postconfluentCaco-2) cells; 3) exposure to insulin and the inhibition of Fak,MEK, and PI3-K resulted in differentiation state-distinctmodulations in the expression of each Bcl-2 homolog ana-lyzed; and 4) Fak, b1 and b4 integrins, as well as the MEK/Erk and PI3-K/Akt pathways, are distinctively involved incell survival depending on the state of cell differentiation.Taken together, these data indicate that human intestinalepithelial cell survival is regulated according to differentia-tion state-specific control mechanisms.
anoikis; apoptosis; gut; intestine; signal transduction
PROGRAMMED CELL DEATH, or apoptosis, is a complex andtightly regulated process that performs crucial func-tions in tissue homeostasis (1, 25). The Bcl-2 family ofproteins constitutes a critical decisional checkpoint incell survival, regulating the common downstream ef-fector pathway of apoptosis (1, 47). At least 15 familymembers have been identified so far, functioning either
as anti-apoptotic (e.g., Bcl-2, Bcl-XL, Mcl-1) or pro-apop-totic (e.g., Bax, Bad, Bak) regulators (1, 20, 48). It isnow well acknowledged that regulation of cell survivalin different cell types does not depend on the activity ofsingle Bcl-2 homologs but, rather, on a balance of anti-and pro-apoptotic activities from multiple homologs (1,47). To this effect, extracellular signals determine inlarge part if a cell lives or dies, involving signalingevents that ultimately affect the expression and/orfunctions of multiple anti- and pro-apoptotic Bcl-2 ho-mologs (1, 4, 13, 15, 19–21). Such survival signalsoriginate from growth factors (e.g., insulin) and celladhesion, often implicating similar signaling pathways(4, 13, 15, 19–21, 25). In this respect, the roles of focaladhesion kinase (p125Fak), the phosphatidylinositol 39-kinase (PI3-K)/Akt (PKB; p57Akt) pathway, and theMEK [mitogen-activated protein kinase (MAPK) ki-nase]/Erk (extracellular regulated kinases 1 and 2;p42Erk-2/p44Erk-1 MAPK) pathway have received muchattention in recent years (4, 11, 13, 15, 21). However, itis becoming increasingly evident that the involvementof these signaling molecules/pathways in cell survivalvaries depending on the cell type that is receiving suchsignals, thus underlying the complexity in the regula-tion of apoptosis among various tissues (1, 4, 11, 13, 15,19–21, 25).
The intestinal epithelium is a useful model for thestudy of the working mechanics of tissue renewal pro-cesses, including apoptosis. Its rapid, continuous cellrenewal consists of spatially separated stem cells, pro-liferative and differentiated compartments located, re-spectively, in the lower regions of the crypts and on thevilli (26, 45). Such a “gradient” of cell differentiation isfurther defined by functional properties of the fullydifferentiated villus enterocytes, which distinguishthem from crypt cells (26, 37, 41, 45). Hormonal re-sponses as well as cell adhesion components, such asintegrins, underlie additional distinctions between
Address for reprint requests and other correspondence: P. H.Vachon, Departement d’Anatomie et de Biologie Cellulaire, Facultede Medecine, Universite de Sherbrooke, Sherbrooke, Quebec, Can-ada J1H 5N4 (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
Am J Physiol Cell Physiol280: C1540–C1554, 2001.
0363-6143/01 $5.00 Copyright © 2001 the American Physiological Society http://www.ajpcell.orgC1540
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

crypt and villus enterocytes (7, 16, 37, 41, 43–45).Although the predominant means to remove obsoleteenterocytes is through apoptosis and shedding at thevillus apex, “spontaneous” crypt cell apoptosis is a lessfrequent process that serves to remove defective/in-jured progeny cells (22, 24, 26, 40, 44, 45, 53). Inaddition to this apparent “duality of fate” betweenundifferentiated and differentiated intestinal cells,some Bcl-2 homologs have been shown individually toexhibit gradients of expression along the crypt-villusaxis (29–32, 38–40, 44, 45, 59, 60). By analyzing sixhomologs at the same time in the proximal-distal axisof the developing human gut (jejunum, ileum, andcolon), we have previously shown that villus cells grad-ually come to exhibit a Bcl-2 homolog expression profilethat clearly differs from the one observed in crypt cellsby midgestation, once the crypt-villus axis has ma-tured (52, 53). These observations altogether suggestthat intestinal epithelial cell survival may be regulateddistinctively according to the state of cell differentia-tion.
In the present study, we investigated this hypothesisby using the human enterocyte-like cell line Caco-2, awidely used model of intestinal epithelial cell matura-tion and functions (8, 27, 37, 41, 42, 50). Althoughcancerous in origin, these cells are unique in theirproperty to undergo a gradual differentiation processthat takes place spontaneously once confluence hasbeen reached and that is completed after 25–30 days ofpostconfluent culture (8, 27, 37, 41, 42, 50). Conse-quently, Caco-2 cells acquire a morphological polarityand show enzymatic activities as well as protein andmRNA levels of brush-border membrane enzymes thatare highly comparable to those of mature enterocytes(8, 27, 37, 41, 42, 50). In this respect, Caco-2 cells as asingle cell culture system have provided an importanttool to study and further understand the regulation ofvarious human intestinal epithelial cellular processes,such as enterocytic differentiation (2, 12, 37, 41, 42, 50,51), proliferation (2, 17, 41–43), digestive functions (8,12, 27, 37, 41), and cell adhesion (5–7, 35, 41, 51, 61).We also used the normal human fetal cryptlike HIEC-6cells (41), which do not differentiate even when inpostconfluent culture, as an additional model of undif-ferentiated intestinal epithelial cells to complementthe one represented by undifferentiated Caco-2 cells.
Herein, Caco-2 and HIEC-6 cells were exposed togrowth factors (namely, insulin) or pharmacologicalcompounds that inhibit signal transduction molecules(namely, tyrosine kinases, Fak, PI3-K/Akt, and/orMEK/Erk) to evaluate their impact on cell survival andBcl-2 homolog expression. We find that undifferenti-ated and differentiated cells display strikingly distinctBcl-2 homolog expression profiles, such profiles beingestablished gradually during the enterocytic differen-tiation process. In addition, we find that exposure toinsulin, or the inhibition of signaling molecules/path-ways, modulates the expression of Bcl-2 homologs re-gardless of the state of differentiation but that themodulatory effects and homologs affected vary betweenundifferentiated and differentiated cells. Furthermore,
we find that b1 and b4 integrins, as well as the PI3-K/Akt and MEK/Erk pathways, are distinctively involvedin the survival of undifferentiated (subconfluentCaco-2, confluent HIEC-6) and differentiated (30 dayspostconfluent Caco-2) enterocytes. Altogether, thesedata indicate that human intestinal cell survival issubject to distinct regulatory mechanisms according tothe state of differentiation.
MATERIALS AND METHODS
Cell culture. The Caco-2/15 cell line, a stable clone of theparent Caco-2 cells (42), has been characterized elsewhere (8,50). Caco-2/15 cells are known to be poor secretors of growthfactors (2, 8, 12, 17). Cells between passages 53 and 70 werecultured in plastic dishes (60 or 100 mm; Falcon Plastics, LosAngeles, CA), or on 13-mm coverslips, at 37°C in an atmo-sphere of 95% air-5% CO2. The medium used was Dulbecco’smodified Eagle’s medium (DMEM) supplemented with 10%fetal bovine serum (FBS; GIBCO BRL, Burlington, ON, Can-ada), 4 mM glutamine, 20 mM HEPES (pH 7.5), 50 U/mlpenicillin, and 50 mg/ml streptomycin. Cultures were refedevery 48 h and subcultured serially as described previously(8, 50). Studies were performed on cultures at 22 days(subconfluence; 50–70% confluence), 0 days (confluence),and/or 5–30 days postconfluence. The expression of sucrase-isomaltase, a major enterocytic differentiation marker, wasmonitored by Western blot to ensure proper differentiation ofcultures (not shown) as previously described (8, 50, 51). Insome experiments, the intestinal cryptlike human fetalHIEC-6 cells (41) were used in parallel with undifferentiatedCaco-2 cell cultures; HIEC-6 cells do not differentiate onreaching confluence, and their undifferentiated “cryptlike”status does not change with postconfluence (41).
For treatments, cell cultures were rinsed with DMEMwithout serum and then maintained 48 h in DMEM withoutserum with one of the following: 10 mg/ml human recombi-nant insulin (GIBCO BRL); 150–300 mM genistein (Sigma,St. Louis, MO), a wide-spectrum tyrosine kinase activityinhibitor (4); 0.3–1 mM cytochalasin D (Sigma), which inhib-its Fak within the concentration range used (36, 58); 20–60mM PD-98059 (Calbiochem, San Diego, CA), a specific inhib-itor of MEK, the upstream activator of Erk-1/Erk-2 (2, 3, 10,14, 33–35, 48, 49, 61); or 5–30 mM Ly-294002 (Calbiochem),a specific inhibitor of PI3-K (9, 14, 33, 34, 48, 49, 57). In someexperiments, treatments consisted instead of inhibiting thebinding activities of the b1 or b4 integrin subunits with themonoclonal antibodies P4C10 and 3E1, respectively, for 24 has previously described (5, 6, 55). For undifferentiated anddifferentiated cells, determination of the working concentra-tions of the compounds used led us to settle with the follow-ing: 300 mM genistein, 1 mM cytochalasin D, 20 mM PD-98059, and 30 mM Ly-294002. Note that maintenance of cellsin absence of serum for 48 h did not impact on their survivalcompared with exposure to 10% FBS (undifferentiated cells:see Fig. 2A, lane 1 vs. 2; differentiated cells: see Fig. 2B, lane1 vs. 2; Table 1) and did not influence significantly theirexpression levels of Bcl-2 homologs (not shown; Table 1).Henceforth, cultures maintained in DMEM without serumwere considered as “controls” for the experimental treat-ments used.
Antibodies. Primary rabbit polyclonal antibodies used inthe present study were Ab1682, directed against humanMcl-1 (29); Ab1695, directed to human/mouse Bcl-XL (28, 32);Ab1701 (29, 31) and AbPC68 (Calbiochem), both directedagainst human Bcl-2; Ab1712 (31) and AbPC66 (Calbio-chem), both directed to human Bax; Ab1764, directed against
C1541DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from
human Bak (28, 30); AbI-19 (Santa Cruz Biotechnology,Santa Cruz, CA), directed to human/mouse Bak; AbPC67(Calbiochem), directed to human Bcl-XL; AbK-20 (Santa CruzBiotechnology), directed against human/mouse Mcl-1; andAb9292 (New England Biolabs, Beverly, MA) and AbR-20(Santa Cruz Biotechnology), both directed to human Bad.Primary mouse monoclonal antibodies used were mAbCY-90(Sigma-Aldrich Canada, Oakville, ON), directed against hu-man cytokeratin 18 (K18).
Note that antibodies Ab1682, Ab1695, Ab1701, Ab1712,and Ab1764 were developed in the laboratory of one of theauthors (J. C. Reed) of the present study; these, as well as thecommercial antibodies used herein, have been characterizedin previous studies (10, 23, 28–32, 39, 52, 53).
In situ detection of apoptosis-associated DNA strandbreaks. For some experiments, in situ terminal deoxynucleo-tidyl transferase-mediated dUTP nick-end labeling (ISEL)was carried out as already described (52, 54, 55) on coverslip-grown cultures after treatments, using the ApopTag apopto-sis detection kit (Oncor, Gaithersburg, MD). Preparationswhere then counterstained with Evans blue and mountedand viewed with a Reichart Polyvar 2 microscope (Leica, St.Laurent, QC, Canada) equipped for epifluorescence. Evalua-tion of ISEL-positive cells was performed as previously de-scribed (54, 55). A minimum of 300 cells was counted in atleast three (n $ 3) separate experiments and/or cultures. Theapoptotic indexes were expressed as the percentage of apop-totic (ISEL-positive) cells over the total number of cellscounted (6SE); statistically significant (0.001 # P # 0.01)differences were determined with Student’s t-test.
DNA laddering assays. After treatments (see above), DNAwas isolated according to the modified method of Frisch (18)as previously described (54). DNA contents of all sampleswere estimated by optical density at 260 nm. For the visual-ization of apoptosis-associated DNA internucleosomal frag-mentation (DNA laddering), each sample was then resolvedby electrophoresis (20 mg DNA/lane) on 2% agarose gelsstained with ethidium bromide, using a 100-bp ladder
(GIBCO BRL) as standard. Note that the method used forDNA extraction uses Triton rather than SDS, thus leavingbehind intact genomic DNA (18); consequently, nonapoptoticcell cultures produce near-empty lanes on the gel as a result(for example, see Fig. 2, A and B, lanes 1 and 2).
Protein expression levels. For analyses of protein expres-sion levels of Bcl-2 homologs, total protein from cell lysateswas obtained after medium removal, PBS washes, and scrap-ping of cells in 13 solubilization buffer [2.3% SDS, 10%glycerol, and 0.001% bromphenol blue in 62.5 mM Tris zHCl(pH 6.8) containing 5% b-mercaptoethanol]. Samples werethen boiled (105°C, 5 min), cleared by centrifugation (15,000g, 5 min, room temperature), and processed for storage asdescribed previously (51–55). SDS-PAGE on 15% acrylamideTris zHCl gels (Bio-Rad, Hercules, CA) was performed asdescribed previously (51–55). Broad-range molecular massmarkers (6.8–209 kDa range; Bio-Rad) were used as stan-dards. Total proteins (50 mg/well) were separated by electro-phoresis and then electrotransferred to nitrocellulose mem-branes (Supported NitroCellulose-1; GIBCO BRL) forsubsequent immunoblotting (51–55). Rabbit antisera wereused at 1:200–1:2,000 dilutions, and mouse monoclonal an-tibodies were used at 1:5,000 dilutions. Immunoreactivebands were visualized by the enhanced chemiluminescencemethod (ECL system; Amersham/Pharmacia Biotechnology,Baie D’Urfe, QC, Canada) according to the manufacturer’sinstructions. Band intensities were quantified by laser den-sitometry using an Alpha Imager 1200 documentation andanalysis system (Alpha Innotech, San Leondro, CA).
To analyze the expression profiles of Bcl-2 homologs inrelation to the enterocytic differentiation process, relativeexpression levels were determined by comparison with cyto-keratin 18 (K18) as a reference protein. This epithelial cy-toskeletal component is expressed at stable levels throughoutthe enterocytic differentiation process (Fig. 1A) (51–53). To-tal peak areas [arbitrary units (AU) 3 mm] were determinedat 22, 0, 5, 10, 15, 20, and 30 days postconfluence to establishthe ratios of homolog to K18 for each molecule studied. For
Table 1. Differentiation state-specific controls of intestinal epithelial cell survival
Treatment StateApoptotic
Index
Homolog
Bcl-2 Bcl-XL Mcl-1 Bax Bak Bad
Control (no serum) U 7.961.8D 11.362.4
1FBS (10%) U 4.161.5D 5.961.7
1I (10 mg/ml) U 5.561.9 1 1 2 2D 8.262.0 2 1 2
1G (300 mM) U 63.867.7* 2 2 2 1 2D 83.168.1*† 2 2 1 1
1PD (20 mM) U 3.861.7 2 2D 39.565.1*† 2 2 2 2 2
1Ly (30 mM) U 85.765.4* 2 2 1D 67.267.1*† 2 2 2
1CD (1 mM) U 58.569.9* 2 1 1D 85.3612.7*† 2 2 2 1 1 2
Values for the apoptotic indexes are means 6 SE expressed as the percentage of apoptotic cells over the total number of cells counted.Caco-2/15 cells were maintained 48 h in DMEM medium without serum (control) or in the presence of serum [fetal bovine serum (FBS)],growth factors [insulin (I)], or inhibitors of signaling molecules/pathways (1G, genistein; 1PD, PD-98059; 1Ly, Ly-294002; 1CD,cytochalasin D) as specified; U, undifferentiated cultures; D, differentiated cultures. In situ TdT-mediated dUTP nick-end labeling (ISEL)was performed on coverslip-grown cell cultures to establish the apoptotic indexes after treatments. A minimum of 300 cells was counted inat least 3 (n $ 3) separate experiments and/or cultures. Homolog, qualitative assessment of significant modulations in the expression of Bcl-2homologs (Bcl-2, Bcl-XL, Mcl-1, Bax, Bak, Bad) in treated cultures, in comparison to control cultures, as observed from Western blot analyses(see Figs. 7 and 8); 2, downmodulation; 1, upregulation; lack of symbol indicates no significant modulation or control steady-state levels.*Statistically significant (0.001 # P # 0.01) differences between treatments and their respective control cultures. †Statistically significant(0.001 # P # 0.01) differences between differentiated and undifferentiated cells.
C1542 DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

treatments, the relative expression levels of molecules ana-lyzed were evaluated compared with that of control cultures(DMEM without serum) by determining total peak areas(AU 3 mm) to establish the ratios of treatment to control 3100 (expressed as “% of control”). In all cases, values shownrepresent means 6 SE for at least three (n $ 3) separateexperiments and/or cultures.
Immunoprecipitation. For some experiments, cell cultureswere harvested in cold RIPA buffer [50 mM Tris zHCl (pH7.2), 150 mM NaCl, 1 mM dithiothreitol, 0.5 mM EDTA, 1%Nonidet P-40, 0.5% sodium deoxycholate, 0.1% SDS, 100 mMNa3VO4, 1 mM phenylmethylsulfonyl fluoride, 0.5 mg/ml leu-peptin, 0.5 mg/ml aprotinin, 0.7 mg/ml pestadin, 40 mMb-glycerophosphate, and 10 mM Na2P4O7] and lysed in thebuffer (30 min on ice). Lysates were cleared by centrifugation(15,000 rpm, 15 min, 4°C) and aliquoted for storage as pre-viously described (6, 54). Immunoprecipitation of Fak wascarried out according to the protocol already described (54),using 1 mg of the rabbit polyclonal AbC-903 (Santa CruzBiotechnology) directed against human/mouse/rat p125Fak.
Assay of Fak, Erk-1/Erk-2, and Akt relative activationlevels by immunoblotting. Total proteins were resolved bySDS-PAGE and electrotransferred as described above. As-says of p125Fak, p42Erk-2/p44Erk-1, or p57Akt relative activa-tion levels were performed as described elsewhere (2, 10, 33,34, 61). For Fak assays, membranes were first probed withthe rabbit polyclonal Ab07-012 (Upstate Biotechnology, LakePlacid, NY) directed to the activated phospho-Tyr-397 form ofFak (11, 21, 61) and then reprobed with the AbC-903 fornormalization purposes. For some experiments, Fak immu-noprecipitates were used instead (see above). For Erk-1/Erk-2 assays, membranes were probed with the rabbit poly-clonal Ab9101s (New England Biolabs) directed to the doublyphosphorylated (activated) forms of Erk-1/Erk-2 (2, 4, 10, 34)and then reprobed with the rabbit polyclonal Ab9102 (NewEngland Biolabs) directed to total Erk-1/Erk-2. Finally, as-says of p57Akt relative activation levels were performed byfirst probing membranes with the rabbit polyclonal Ab9271s(New England Biolabs) directed to the activated phospho-Ser-473-Akt form (4, 15, 33, 34, 48, 56) and then reprobedwith the rabbit polyclonal Ab9272 (New England Biolabs)directed to total Akt.
The relative activation levels of p125Fak, p42Erk-2/p44Erk-1,or p57Akt were evaluated by determining the total peak areas(AU 3 mm) for the phosphorylated forms and for the corre-sponding total protein, to establish the ratios pp125Fak/p125Fak, pp42Erk-2/p42Erk-2, and pp57Akt/p57Akt. In the caseof Erk-1/Erk-2, analyses focused on p42Erk-2 (2, 10). Ratioswere in turn compared with those of control cultures, 3100(expressed as % of control). Values shown represent means 6SE for at least three (n $ 3) separate experiments and/orcultures.
RESULTS
Establishment of distinct Bcl-2 expression profilesduring enterocytic differentiation. To determine whetherhuman intestinal epithelial cell survival is subject todifferentiation state-specific regulatory mechanisms,we first investigated the steady-state expression levelsof six Bcl-2 homologs (Bcl-2, Bcl-XL, Mcl-1, Bax, Bak,Bad) during the enterocytic differentiation process ofCaco-2/15 cells. Immunoblot analyses of cell lysatesdemonstrated the protein expression of all moleculesanalyzed herein (Fig. 1A). Thus Bcl-2 (;26 kDa),Bcl-XL (;28–30 kDa), Mcl-1 (;39–42 kDa), Bax (;21
Fig. 1. Establishment of distinct Bcl-2 homolog expression profilesduring the enterocytic differentiation process of Caco-2/15 cells. A:representative Western blot analyses of Bcl-2, Bcl-XL, Mcl-1, Bax,Bak, Bad, and K18 steady-state expression levels from Caco-2/15 cellmonolayers at 22 (subconfluence; lane 1), 0 (confluence; lane 2), 5(lane 3), 10 (lane 4), 15 (lane 5), 20 (lane 6), and 30 (lane 7) dayspostconfluence. Total proteins (50 mg/well) were separated by SDS-PAGE under reducing conditions, electrotransferred onto nitrocellu-lose membranes, and then probed with specific antibodies for thedetection of Bcl-2 (;26 kDa), Bcl-XL (;28–30 kDa), Mcl-1 (;39–42kDa), Bax (;21 kDa), Bak (;25–28 kDa), or Bad (;28–32 kDa).Detection of K18 was used as standard of protein quantities ana-lyzed. B: relative expression levels of anti-apoptotic Bcl-2 homologsduring Caco-2/15 cell differentiation. Total proteins were separatedand probed as represented in A and then scanned by laser densitom-etry. Relative expression levels of Bcl-2, Bcl-XL, and Mcl-1 wereassessed at 22, 0, 5, 10, 15, 20, and 30 days postconfluence comparedwith that of K18 by determining total peak areas [arbitrary units(AU) 3 mm] to establish the ratios of homolog to K18 (homolog/K18).C: relative expression levels of proapoptotic Bcl-2 homologs duringCaco-2/15 cell differentiation. Total proteins were separated, probed,and scanned by laser densitometry to assess the relative expressionlevels of Bax, Bak, and Bad, as in B. Data in B and C representmeans 6 SE for at least 3 (n $ 3) separate and independent culturesfor each culture stage studied.
C1543DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

kDa), Bak (; 25–28 kDa), and Bad (;28–32 kDa) weredetected at all differentiation stages studied as proteinbands migrating at their previously reported relativemolecular weights (1, 10, 28–32, 39, 47–49, 50, 52, 53).
To examine the modulations of Bcl-2 homolog ex-pression in relation to Caco-2/15 cell enterocytic differ-entiation, the relative expression levels of Bcl-2 ho-mologs were determined by comparison with areference protein, K18. The densitometric data pre-sented in Fig. 1, B and C, show that the relativeexpression levels of all Bcl-2 homologs analyzed weredistinctively modulated in parallel to the differentia-tion process of Caco-2/15 cells. In the case of anti-apop-totic homologs (Fig. 1B), the steady-state levels of Bcl-2gradually increased between 0 and 15 days postconflu-ence, to stabilize more or less thereafter as the cellscompleted their differentiation (Fig. 1A, Bcl-2; Fig. 1B,open squares). Similarly, Mcl-1 levels gradually in-creased throughout the differentiation process of Caco-2/15 cells (Fig. 1A, Mcl-1; Fig. 1B, filled triangles). Onthe other hand, Bcl-XL levels decreased during entero-cytic differentiation, particularly between the 10- and30-day postconfluent stages (Fig. 1A, Bcl-XL; Fig. 1B,filled circles).
In the case of proapoptotic homologs (Fig. 1C), thesteady-state levels of Bax were found to increase grad-ually during Caco-2/15 cell differentiation (Fig. 1A,Bax; Fig. 1C, filled squares). On the other hand, Baklevels decreased sharply between the 22- and 0-daypostconfluent stages, to more or less stabilize at lowlevels thereafter (Fig. 1A, Bak; Fig. 1C, open triangles),whereas Bad levels decreased sharply between 5 and10 days postconfluence, also to remain more or lessstable at low levels thereafter (Fig. 1A, Bad; Fig. 1C,filled circles).
These gradual changes in Bcl-2 homolog steady-statelevels were found not to be a product of postconfluentculture but, instead, to be clearly associated with theenterocytic differentiation process of Caco-2/15 cells.First, these changes in Bcl-2 homolog expression levelswere parallel to the appearance and gradual increaseof sucrase-isomaltase expression (not shown; 8, 12, 27,37, 41, 42, 50, 51), as well as gradual changes in theactivation levels of signaling molecules (see below),during the differentiation of Caco-2/15 cells. Second,the steady-state levels of all Bcl-2 homologs analyzedherein did not change significantly in the undifferenti-ated cryptlike human HIEC-6 cells while maintainedin postconfluent culture (not shown); these cells remainundifferentiated regardless of their state of postconflu-ent culture (41).
It is noteworthy that the Caco-2/15 cell undifferen-tiated-to-differentiated expression profiles of the sixBcl-2 homologs analyzed herein do not correspond tothe in vivo crypt-villus gradients already reported forthese homologs in the adult (26, 28–32, 39, 45) and/ormidgestation (52, 53) human small intestine but corre-spond instead to those already observed along the fullyfunctional and jejunal-like crypt-villus axis of themidgestation human fetal colon (52). Nonetheless, ourobservations altogether indicate that the steady-state
expression levels of Bcl-2 homologs undergo majormodulations during the enterocytic differentiation pro-cess, resulting in expression profiles of differentiatedcells that are distinct from those of undifferentiatedones.
Enterocytes display distinct susceptibilites to apopto-sis according to their differentiation state. Because un-differentiated and differentiated Caco-2/15 cells ex-hibit distinct Bcl-2 homolog expression profiles, wethen sought to ascertain whether this translated intodifferential susceptibilities to apoptosis as well. UsingDNA laddering assays to visualize the internucleosomalDNA fragmentation associated with apoptosis, we eval-uated the presence of apoptosis in undifferentiated anddifferentiated cultures maintained for 48 h with or with-out FBS, or without serum but with the addition ofinsulin or pharmacological inhibitors of signal transduc-tion molecules/pathways. Apoptotic indexes in treatedcultures were also determined using the ISEL method.
We first found that maintenance of Caco-2/15 cells inthe absence of serum for 48 h did not impact signifi-cantly on their survival compared with their mainte-nance in the presence of FBS. Indeed, no DNA ladder-ing was evidenced in either culture condition forundifferentiated (Fig. 2A, lane 1 vs. 2) and differenti-ated cells (Fig. 2B, lane 1 vs. 2), whereas the “basic”apoptotic indexes found in the presence of FBS (Table1, 1FBS) did not differ significantly from those ob-tained in absence of serum (Table 1, control) for eitherdifferentiation states. Incidentally, we (51) and others(27, 41) have previously reported that absence of serumfor 24–48 h was not detrimental to enterocytic func-tions of Caco-2 cells.
As expected from the lack of differences between theabsence and presence of serum, the exposure to insulinfor 48 h had no significant influence on the survival ofeither undifferentiated (Fig. 2A, lane 3; Table 1, 1I) ordifferentiated (Fig. 2B, lane 3; Table 1, 1I) Caco-2/15cell cultures. In contrast, the inhibition of tyrosinekinase activities greatly impacted on cell survival inboth differentiation states (;64% apoptosis in undif-ferentiated cells and ;83% apoptosis in differentiatedcells; Table 1, 1G) (Fig. 2, A and B, lane 4). Likewise,the specific inhibition of PI3-K (Fig. 2, A and B, lane 6;Table 1, 1Ly) and Fak (Fig. 2, A and B, lane 7; Table 1,1CD) resulted in significant increases in apoptosis inboth states of differentiation.
However, stark differences were noted on the respec-tive impacts of some of these treatments on cell sur-vival between undifferentiated and differentiatedCaco-2/15 cells (Table 1). For instance, the inhibition oftyrosine kinase activities qualitatively resulted inmore abundant DNA laddering in differentiated Caco-2/15 cells than in undifferentiated ones (Fig. 2B, lane 4vs. A, lane 4), which translated into significant differ-ences in the corresponding apoptotic indexes (Table 1,1G). Likewise, the inhibition of Fak produced moreDNA laddering in differentiated Caco-2/15 cells thanundifferentiated ones (Fig. 2B, lane 7 vs. A, lane 7),which translated as well into a significantly greaterapoptotic index in differentiated cells than undifferen-
C1544 DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

tiated ones (;85% vs. ;59%; Table 1, 1CD). Morestriking was the inhibition of MEK/Erk, which did notimpact on the survival of undifferentiated Caco-2/15cells (Fig. 2A, lane 5; Table 1, 1PD) but did so indifferentiated ones (Fig. 2B, lane 5; Table 1, 1PD). Incontrast, the inhibition of PI3-K resulted in a signifi-cantly greater impact on the survival of undifferenti-ated Caco-2/15 cells than on differentiated ones (Fig.2B, lane 6 vs. A, lane 6; Table 1, 1Ly).
Hence, these results altogether indicate that entero-cytes exhibit distinct susceptibilities to apoptosis ac-cording to their state of differentiation. It is of notethat the results obtained herein with undifferentiatedCaco-2/15 cells were highly similar, if not identical, tothose obtained in parallel experiments with confluentcultures of undifferentiated cryptlike HIEC-6 cells (notshown), thus further underlying the differences be-tween undifferentiated (subconfluent Caco-2/15, con-fluent HIEC-6) and differentiated (30 days postconflu-ent Caco-2/15) intestinal epithelial cells in theregulation of their survival.
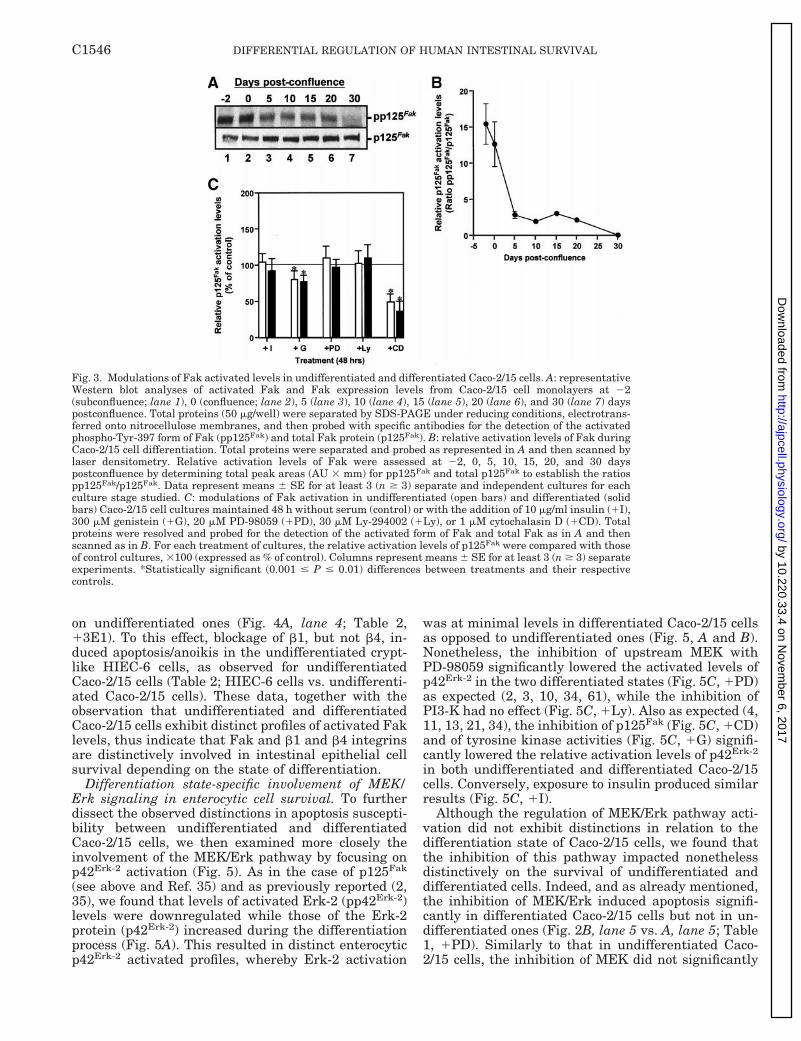
Differentiation state distinct involvement of Fak andintegrins in enterocytic cell survival. To verify whetherthe observed differentiation state distinctions in apop-tosis susceptibility were linked to a differential in-volvement of signaling pathways, we first investigatedspecifically the role of cell adhesion signaling in Caco-2/15 cell survival, focusing on p125Fak activation(Fig. 3). As previously reported in the parental Caco-2cell line (35), we found that levels of activated Fak(pp125Fak) were downregulated while those of the pro-tein (p125Fak) increased during the differentiation pro-cess (Fig. 3A). This resulted in distinct enterocyticp125Fak activation profiles, whereby Fak activationwas at minimal levels in differentiated cells as opposedto undifferentiated ones (Fig. 3, A and B). Nonetheless,relative p125Fak activated levels were equally inhibitedin the two differentiated states when either genistein(Fig. 3C, 1G) or cytochalasin D (Fig. 3C, 1CD) wasused as a treatment. The nonspecific inhibition ofp125Fak activation by genistein, a wide-spectrum in-hibitor of tyrosine kinases, was expected (35, 36, 58,61), whereas cytochalasin D can act as a more specificinhibitor of p125Fak at the concentration range used(36, 58). We also observed that the inhibition of thedownstream MEK/Erk pathway (Fig. 3C, 1PD) orPI3-K (Fig. 3C, 1Ly), as well as exposure to insulin(Fig. 3C, 1I), did not impact on the relative activationlevels of p125Fak itself in either undifferentiated ordifferentiated Caco-2/15 cells, as expected from previ-ous studies in other cell types and tissues (4, 11, 33, 34,36, 58).
Because Fak is largely responsible for b1 integrinsignaling (11, 19, 21), we then analyzed the impact ofthe inhibition b1 integrin activity on Caco-2/15 cellsurvival and immunoprecipitated p125Fak relative ac-tivation levels. In parallel, the potential involvement ofb4 integrins was analyzed as well; it is noteworthy thatenterocytes, including Caco-2/15 and HIEC-6 cells, ex-press the b4 integrin subunit (5, 7). As expected fromother studies (11, 18, 19, 21, 33, 55, 58), the blockage ofb1 integrins induced apoptosis/anoikis in undifferenti-ated (Fig. 4A, lane 3) and differentiated (Fig. 4A, lane7) Caco-2/15 cells, in addition to causing sharp reduc-tions in immunoprecipitable p125Fak activated levelsin both differentiation states (Fig. 4B, lanes 5 and 6;Fig. 4C, 1P4C10). However, and as already notedwhen Fak is inhibited by cytochalasin D (Table 1,1CD), the impact of b1 integrin inhibition on cellsurvival was significantly greater in differentiatedCaco-2/15 cells (;78% apoptosis; Table 2, 1P4C10)than in undifferentiated ones (;55% apoptosis; Table2, 1P4C10); this was likewise apparent from the in-tensity of the DNA laddering observed (Fig. 4A, lane 3vs. 7). Our b4 integrin inhibition experiments producedeven more striking results between undifferentiatedand differentiated Caco-2/15 cells. Although the inhi-bition of the b4 subunit did not influence p125Fak
relative activation levels (Fig. 4B, lanes 7 and 8; Fig.4C, 13E1), as expected (11, 21), it did nonethelessimpact significantly on the survival of differentiatedCaco-2/15 cells (Fig. 4A, lane 8; Table 2, 13E1) but not
Fig. 2. Differentiation state-distinct susceptibilities to apoptosis inintestinal epithelial cells. A and B: representative DNA ladderingassays from undifferentiated (22 days postconfluence; A) and differ-entiated (30 days postconfluence; B) Caco-2/15 cell cultures main-tained 48 h without serum (control, C, lane 1) or with the addition of10% fetal bovine serum (1FBS; lane 2), 10 mg/ml insulin (1I; lane 3),300 mM genistein (1G; lane 4), 20 mM PD-98059 (1PD; lane 5), 30mM Ly-294002 (1Ly; lane 6), or 1 mM cytochalasin D (1CD; lane 7).DNA (20 mg/lane) were isolated from treated cultures and thenseparated by agarose gel electrophoresis and stained with ethidiumbromide for the visualization of apoptosis/anoikis-associated inter-nucleosomal DNA fragmentation (“DNA laddering”). Results illus-trated here for undifferentiated Caco-2/15 cells were similar, if notidentical, to those obtained with the undifferentiated cryptlikeHIEC-6 cells (not shown). L, 100-bp DNA size markers.
C1545DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from
on undifferentiated ones (Fig. 4A, lane 4; Table 2,13E1). To this effect, blockage of b1, but not b4, in-duced apoptosis/anoikis in the undifferentiated crypt-like HIEC-6 cells, as observed for undifferentiatedCaco-2/15 cells (Table 2; HIEC-6 cells vs. undifferenti-ated Caco-2/15 cells). These data, together with theobservation that undifferentiated and differentiatedCaco-2/15 cells exhibit distinct profiles of activated Faklevels, thus indicate that Fak and b1 and b4 integrinsare distinctively involved in intestinal epithelial cellsurvival depending on the state of differentiation.
Differentiation state-specific involvement of MEK/Erk signaling in enterocytic cell survival. To furtherdissect the observed distinctions in apoptosis suscepti-bility between undifferentiated and differentiatedCaco-2/15 cells, we then examined more closely theinvolvement of the MEK/Erk pathway by focusing onp42Erk-2 activation (Fig. 5). As in the case of p125Fak
(see above and Ref. 35) and as previously reported (2,35), we found that levels of activated Erk-2 (pp42Erk-2)levels were downregulated while those of the Erk-2protein (p42Erk-2) increased during the differentiationprocess (Fig. 5A). This resulted in distinct enterocyticp42Erk-2 activated profiles, whereby Erk-2 activation
was at minimal levels in differentiated Caco-2/15 cellsas opposed to undifferentiated ones (Fig. 5, A and B).Nonetheless, the inhibition of upstream MEK withPD-98059 significantly lowered the activated levels ofp42Erk-2 in the two differentiated states (Fig. 5C, 1PD)as expected (2, 3, 10, 34, 61), while the inhibition ofPI3-K had no effect (Fig. 5C, 1Ly). Also as expected (4,11, 13, 21, 34), the inhibition of p125Fak (Fig. 5C, 1CD)and of tyrosine kinase activities (Fig. 5C, 1G) signifi-cantly lowered the relative activation levels of p42Erk-2
in both undifferentiated and differentiated Caco-2/15cells. Conversely, exposure to insulin produced similarresults (Fig. 5C, 1I).
Although the regulation of MEK/Erk pathway acti-vation did not exhibit distinctions in relation to thedifferentiation state of Caco-2/15 cells, we found thatthe inhibition of this pathway impacted nonethelessdistinctively on the survival of undifferentiated anddifferentiated cells. Indeed, and as already mentioned,the inhibition of MEK/Erk induced apoptosis signifi-cantly in differentiated Caco-2/15 cells but not in un-differentiated ones (Fig. 2B, lane 5 vs. A, lane 5; Table1, 1PD). Similarly to that in undifferentiated Caco-2/15 cells, the inhibition of MEK did not significantly
Fig. 3. Modulations of Fak activated levels in undifferentiated and differentiated Caco-2/15 cells. A: representativeWestern blot analyses of activated Fak and Fak expression levels from Caco-2/15 cell monolayers at 22(subconfluence; lane 1), 0 (confluence; lane 2), 5 (lane 3), 10 (lane 4), 15 (lane 5), 20 (lane 6), and 30 (lane 7) dayspostconfluence. Total proteins (50 mg/well) were separated by SDS-PAGE under reducing conditions, electrotrans-ferred onto nitrocellulose membranes, and then probed with specific antibodies for the detection of the activatedphospho-Tyr-397 form of Fak (pp125Fak) and total Fak protein (p125Fak). B: relative activation levels of Fak duringCaco-2/15 cell differentiation. Total proteins were separated and probed as represented in A and then scanned bylaser densitometry. Relative activation levels of Fak were assessed at 22, 0, 5, 10, 15, 20, and 30 dayspostconfluence by determining total peak areas (AU 3 mm) for pp125Fak and total p125Fak to establish the ratiospp125Fak/p125Fak. Data represent means 6 SE for at least 3 (n $ 3) separate and independent cultures for eachculture stage studied. C: modulations of Fak activation in undifferentiated (open bars) and differentiated (solidbars) Caco-2/15 cell cultures maintained 48 h without serum (control) or with the addition of 10 mg/ml insulin (1I),300 mM genistein (1G), 20 mM PD-98059 (1PD), 30 mM Ly-294002 (1Ly), or 1 mM cytochalasin D (1CD). Totalproteins were resolved and probed for the detection of the activated form of Fak and total Fak as in A and thenscanned as in B. For each treatment of cultures, the relative activation levels of p125Fak were compared with thoseof control cultures, 3100 (expressed as % of control). Columns represent means 6 SE for at least 3 (n $ 3) separateexperiments. *Statistically significant (0.001 # P # 0.01) differences between treatments and their respectivecontrols.
C1546 DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from
induce apoptosis in the undifferentiated cryptlikeHIEC-6 cells (not shown). These data, together withthe observation that undifferentiated and differenti-ated Caco-2/15 cells exhibit distinct profiles of acti-
vated Erk-2 levels, thus indicate that the MEK/Erkpathway is distinctively involved in intestinal epithe-lial cell survival, depending on the state of differenti-ation.
Differentiation state-distinct regulation of the PI3-K/Akt pathway and involvement in enterocytic cell sur-vival. Because the MEK/Erk pathway displayed differ-entiation state distinctions in its involvement inenterocytic cell survival but not in its regulation ofactivation, we then verified whether this was also thecase for the PI3-K/Akt pathway by focusing on p57Akt
activation (Fig. 6). As in the case of p125Fak (see aboveand Ref. 35) and p42Erk-2 (see above and Refs. 2 and35), we found that levels of activated Akt (pp57Akt)were downregulated while those of the Akt protein(p57Akt) increased during the differentiation process ofCaco-2/15 cells (Fig. 6A). This resulted in distinctp57Akt activated profiles, whereby Akt activation wasat minimal levels in differentiated Caco-2/15 cells asopposed to undifferentiated ones (Fig. 6, A and B).Conversely, we found that the inhibition of p125Fak
(Fig. 6C, 1CD) and tyrosine kinase activities (Fig. 6C,1G) lowered p57Akt relative activation levels in bothundifferentiated and differentiated Caco-2/15 cells,also as observed for p42Erk-2 (see above) and as ex-pected from previous studies in other cell types andtissues (11, 15, 21, 56).
Although the inhibition of MEK/Erk did not impacton p57Akt activated levels (Fig. 6C, 1PD), insulin wasfound to exert distinct influences on p57Akt activationdepending on the state of differentiation: hence, rela-tive p57Akt activated levels were increased by insulinin undifferentiated Caco-2/15 cells but were not af-fected significantly in differentiated ones (Fig. 6C, 1I).Further distinctions were noted in the regulation of thePI3-K/Akt pathway between undifferentiated and dif-
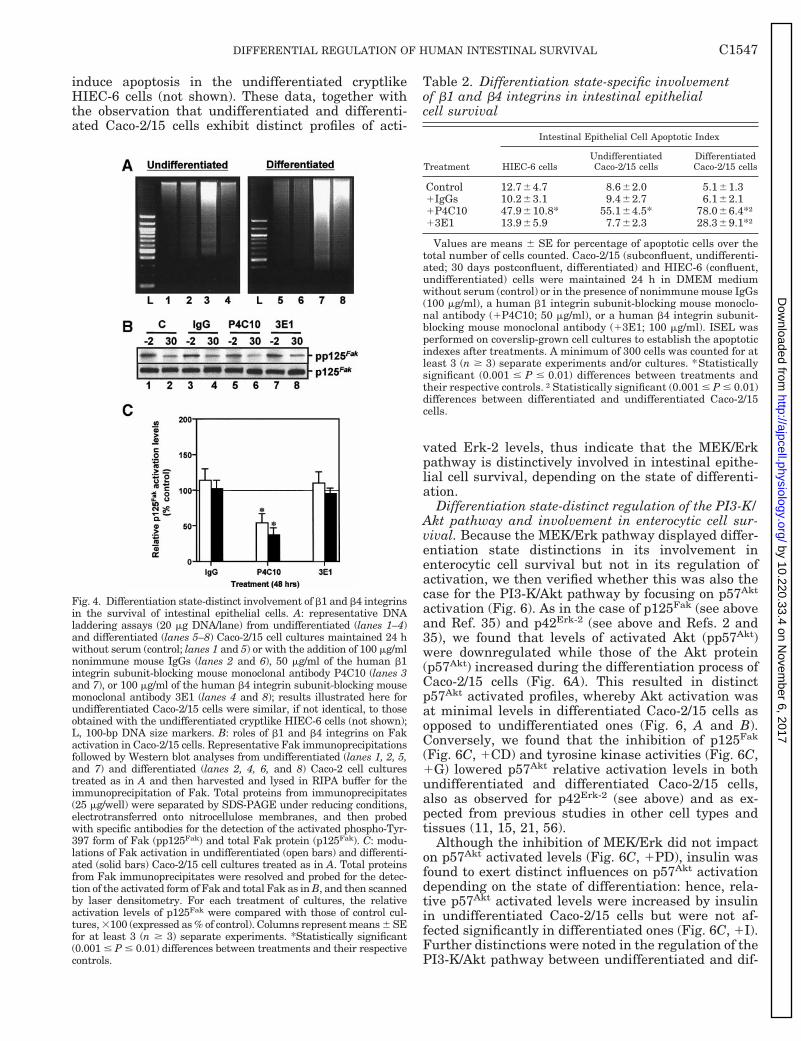
Fig. 4. Differentiation state-distinct involvement of b1 and b4 integrinsin the survival of intestinal epithelial cells. A: representative DNAladdering assays (20 mg DNA/lane) from undifferentiated (lanes 1–4)and differentiated (lanes 5–8) Caco-2/15 cell cultures maintained 24 hwithout serum (control; lanes 1 and 5) or with the addition of 100 mg/mlnonimmune mouse IgGs (lanes 2 and 6), 50 mg/ml of the human b1integrin subunit-blocking mouse monoclonal antibody P4C10 (lanes 3and 7), or 100 mg/ml of the human b4 integrin subunit-blocking mousemonoclonal antibody 3E1 (lanes 4 and 8); results illustrated here forundifferentiated Caco-2/15 cells were similar, if not identical, to thoseobtained with the undifferentiated cryptlike HIEC-6 cells (not shown);L, 100-bp DNA size markers. B: roles of b1 and b4 integrins on Fakactivation in Caco-2/15 cells. Representative Fak immunoprecipitationsfollowed by Western blot analyses from undifferentiated (lanes 1, 2, 5,and 7) and differentiated (lanes 2, 4, 6, and 8) Caco-2 cell culturestreated as in A and then harvested and lysed in RIPA buffer for theimmunoprecipitation of Fak. Total proteins from immunoprecipitates(25 mg/well) were separated by SDS-PAGE under reducing conditions,electrotransferred onto nitrocellulose membranes, and then probedwith specific antibodies for the detection of the activated phospho-Tyr-397 form of Fak (pp125Fak) and total Fak protein (p125Fak). C: modu-lations of Fak activation in undifferentiated (open bars) and differenti-ated (solid bars) Caco-2/15 cell cultures treated as in A. Total proteinsfrom Fak immunoprecipitates were resolved and probed for the detec-tion of the activated form of Fak and total Fak as in B, and then scannedby laser densitometry. For each treatment of cultures, the relativeactivation levels of p125Fak were compared with those of control cul-tures, 3100 (expressed as % of control). Columns represent means 6 SEfor at least 3 (n $ 3) separate experiments. *Statistically significant(0.001 # P # 0.01) differences between treatments and their respectivecontrols.
Table 2. Differentiation state-specific involvementof b1 and b4 integrins in intestinal epithelialcell survival
Treatment
Intestinal Epithelial Cell Apoptotic Index
HIEC-6 cellsUndifferentiatedCaco-2/15 cells
DifferentiatedCaco-2/15 cells
Control 12.764.7 8.662.0 5.161.31IgGs 10.263.1 9.462.7 6.162.11P4C10 47.9610.8* 55.164.5* 78.066.4*†13E1 13.965.9 7.762.3 28.369.1*†
Values are means 6 SE for percentage of apoptotic cells over thetotal number of cells counted. Caco-2/15 (subconfluent, undifferenti-ated; 30 days postconfluent, differentiated) and HIEC-6 (confluent,undifferentiated) cells were maintained 24 h in DMEM mediumwithout serum (control) or in the presence of nonimmune mouse IgGs(100 mg/ml), a human b1 integrin subunit-blocking mouse monoclo-nal antibody (1P4C10; 50 mg/ml), or a human b4 integrin subunit-blocking mouse monoclonal antibody (13E1; 100 mg/ml). ISEL wasperformed on coverslip-grown cell cultures to establish the apoptoticindexes after treatments. A minimum of 300 cells was counted for atleast 3 (n $ 3) separate experiments and/or cultures. *Statisticallysignificant (0.001 # P # 0.01) differences between treatments andtheir respective controls. †Statistically significant (0.001 # P # 0.01)differences between differentiated and undifferentiated Caco-2/15cells.
C1547DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from
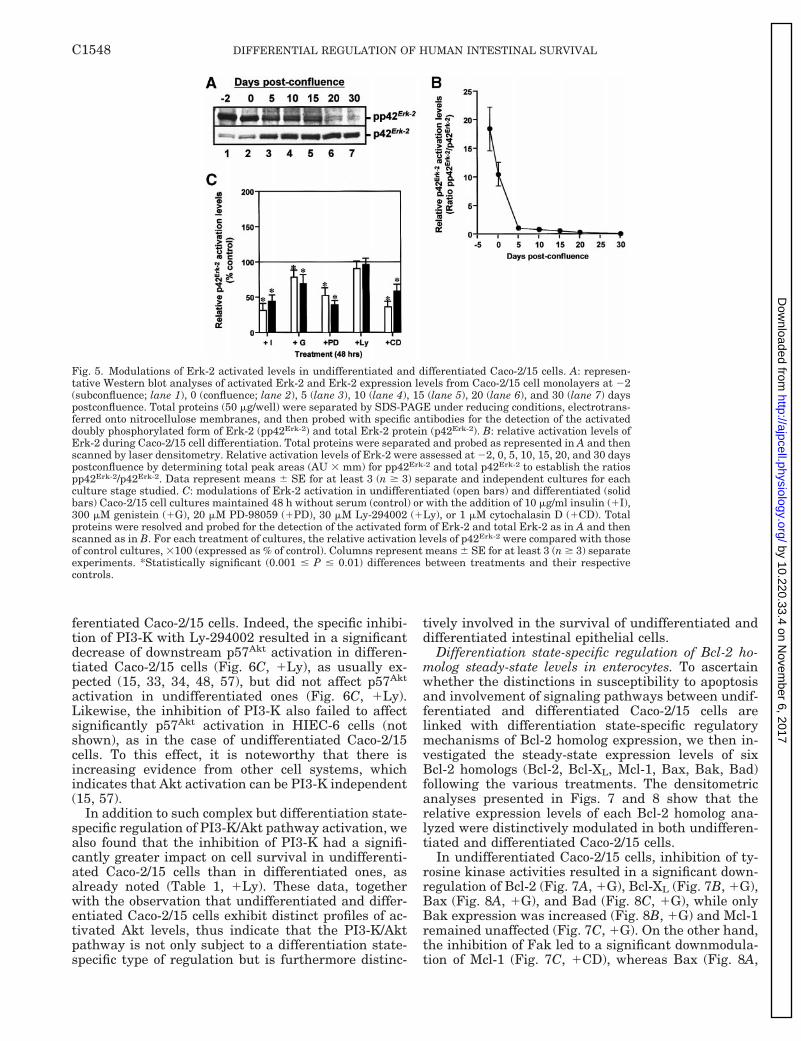
ferentiated Caco-2/15 cells. Indeed, the specific inhibi-tion of PI3-K with Ly-294002 resulted in a significantdecrease of downstream p57Akt activation in differen-tiated Caco-2/15 cells (Fig. 6C, 1Ly), as usually ex-pected (15, 33, 34, 48, 57), but did not affect p57Akt
activation in undifferentiated ones (Fig. 6C, 1Ly).Likewise, the inhibition of PI3-K also failed to affectsignificantly p57Akt activation in HIEC-6 cells (notshown), as in the case of undifferentiated Caco-2/15cells. To this effect, it is noteworthy that there isincreasing evidence from other cell systems, whichindicates that Akt activation can be PI3-K independent(15, 57).
In addition to such complex but differentiation state-specific regulation of PI3-K/Akt pathway activation, wealso found that the inhibition of PI3-K had a signifi-cantly greater impact on cell survival in undifferenti-ated Caco-2/15 cells than in differentiated ones, asalready noted (Table 1, 1Ly). These data, togetherwith the observation that undifferentiated and differ-entiated Caco-2/15 cells exhibit distinct profiles of ac-tivated Akt levels, thus indicate that the PI3-K/Aktpathway is not only subject to a differentiation state-specific type of regulation but is furthermore distinc-
tively involved in the survival of undifferentiated anddifferentiated intestinal epithelial cells.
Differentiation state-specific regulation of Bcl-2 ho-molog steady-state levels in enterocytes. To ascertainwhether the distinctions in susceptibility to apoptosisand involvement of signaling pathways between undif-ferentiated and differentiated Caco-2/15 cells arelinked with differentiation state-specific regulatorymechanisms of Bcl-2 homolog expression, we then in-vestigated the steady-state expression levels of sixBcl-2 homologs (Bcl-2, Bcl-XL, Mcl-1, Bax, Bak, Bad)following the various treatments. The densitometricanalyses presented in Figs. 7 and 8 show that therelative expression levels of each Bcl-2 homolog ana-lyzed were distinctively modulated in both undifferen-tiated and differentiated Caco-2/15 cells.
In undifferentiated Caco-2/15 cells, inhibition of ty-rosine kinase activities resulted in a significant down-regulation of Bcl-2 (Fig. 7A, 1G), Bcl-XL (Fig. 7B, 1G),Bax (Fig. 8A, 1G), and Bad (Fig. 8C, 1G), while onlyBak expression was increased (Fig. 8B, 1G) and Mcl-1remained unaffected (Fig. 7C, 1G). On the other hand,the inhibition of Fak led to a significant downmodula-tion of Mcl-1 (Fig. 7C, 1CD), whereas Bax (Fig. 8A,
Fig. 5. Modulations of Erk-2 activated levels in undifferentiated and differentiated Caco-2/15 cells. A: represen-tative Western blot analyses of activated Erk-2 and Erk-2 expression levels from Caco-2/15 cell monolayers at 22(subconfluence; lane 1), 0 (confluence; lane 2), 5 (lane 3), 10 (lane 4), 15 (lane 5), 20 (lane 6), and 30 (lane 7) dayspostconfluence. Total proteins (50 mg/well) were separated by SDS-PAGE under reducing conditions, electrotrans-ferred onto nitrocellulose membranes, and then probed with specific antibodies for the detection of the activateddoubly phosphorylated form of Erk-2 (pp42Erk-2) and total Erk-2 protein (p42Erk-2). B: relative activation levels ofErk-2 during Caco-2/15 cell differentiation. Total proteins were separated and probed as represented in A and thenscanned by laser densitometry. Relative activation levels of Erk-2 were assessed at 22, 0, 5, 10, 15, 20, and 30 dayspostconfluence by determining total peak areas (AU 3 mm) for pp42Erk-2 and total p42Erk-2 to establish the ratiospp42Erk-2/p42Erk-2. Data represent means 6 SE for at least 3 (n $ 3) separate and independent cultures for eachculture stage studied. C: modulations of Erk-2 activation in undifferentiated (open bars) and differentiated (solidbars) Caco-2/15 cell cultures maintained 48 h without serum (control) or with the addition of 10 mg/ml insulin (1I),300 mM genistein (1G), 20 mM PD-98059 (1PD), 30 mM Ly-294002 (1Ly), or 1 mM cytochalasin D (1CD). Totalproteins were resolved and probed for the detection of the activated form of Erk-2 and total Erk-2 as in A and thenscanned as in B. For each treatment of cultures, the relative activation levels of p42Erk-2 were compared with thoseof control cultures, 3100 (expressed as % of control). Columns represent means 6 SE for at least 3 (n $ 3) separateexperiments. *Statistically significant (0.001 # P # 0.01) differences between treatments and their respectivecontrols.
C1548 DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

1CD) and Bak (Fig. 8B, 1CD) were significantly in-creased. The inhibition of the MEK/Erk pathway de-creased the expression of Bcl-2 (Fig. 7A, 1PD) and Bad(Fig. 8C, 1PD) without affecting the other homologsstudied. Last, the inhibition of PI3-K resulted in asignificant decrease of Bcl-XL (Fig. 7B, 1Ly) and Mcl-1(Fig. 7C, 1Ly), while upregulating the expression ofBax only (Fig. 8A, 1Ly).
In the case of differentiated Caco-2/15 cells, the in-hibition of tyrosine kinase activities led to a significantdownregulation of Bcl-2 (Fig. 7A, 1G) and Bcl-XL (Fig.5B) and to an increase of Bak (Fig. 8B, 1G) and Bad(Fig. 8C, 1G). The inhibition of Fak resulted in asignificant decrease of all anti-apoptotic homologs an-alyzed (Fig. 7, A–C, 1CD) and of Bad (Fig. 8C, 1CD),while increasing Bax (Fig. 8A, 1CD) and Bak (Fig. 8B,1CD). The inhibition of the MEK/Erk pathway led to asignificant decrease of Bcl-2 (Fig. 7A, 1PD), Bcl-XL(Fig. 7B, 1PD) and all proapoptotic homologs analyzed(Fig. 8, A–C, 1PD). Last, the inhibition of PI3-K re-sulted in a decrease of Bcl-XL (Fig. 7B, 1Ly), Mcl-1(Fig. 7C, 1Ly), and Bax (Fig. 8A, 1Ly) without affect-ing the other homologs analyzed.
In addition to such complex modulations of Bcl-2homolog expression in both undifferentiated and dif-
ferentiated Caco-2/15 cells, we also observed stark dif-ferences between the two differentiation states (sum-marized in Table 1). For instance, although insulinincreased Mcl-1 levels in both undifferentiated anddifferentiated cells (Fig. 7C, 1I), the same treatmentfurther resulted in an increase of Bcl-2 (Fig. 7A, 1I)and decrease of Bax (Fig. 8A, 1I) and Bak (Fig. 8B, 1I)in undifferentiated cells (Table 1, 1I), while furtherresulting instead in a decrease of Bcl-XL (Fig. 7B, 1I)and Bad (Fig. 8C, 1I) in differentiated cells (Table 1,1I). Similarly, the inhibition of tyrosine kinase activi-ties affected Bcl-2 (Fig. 7A, 1G), Bcl-XL (Fig. 7B, 1G),Mcl-1 (Fig. 7C, 1G), and Bak (Fig. 8B, 1G) in the samemanner in both differentiation states (Table 1, 1G),while having opposing effects on Bad expression (Fig.8C, 1G) and causing a decrease of Bax (Fig. 8A, 1G) indifferentiated cells only (Table 1, 1G). Further exam-ples of differentiation state-distinct modulations ofindividual Bcl-2 homologs were noted as well whenFak, MEK/Erk, or PI3-K were inhibited (Table 1; Figs.7 and 8).
It is noteworthy that the relationship between thebalance of anti- vs. proapoptotic homologs and theimpact on enterocytic cell survival was not alwaysclear from the experiments performed herein (Table 1).
Fig. 6. Differentiation state-distinct modulations of Akt activated levels in intestinal epithelial cells. A: represen-tative Western blot analyses of activated Akt and Akt expression levels from Caco-2/15 cell monolayers at 22(subconfluence; lane 1), 0 (confluence; lane 2), 5 (lane 3), 10 (lane 4), 15 (lane 5), 20 (lane 6), and 30 (lane 7) dayspostconfluence. Total proteins (50 mg/well) were separated by SDS-PAGE under reducing conditions, electrotrans-ferred onto nitrocellulose membranes, and then probed with specific antibodies for the detection of the activatedphospho-Ser-473 form of Akt (pp57Akt) and total Akt protein (p57Akt). B: relative activation levels of Akt duringCaco-2/15 cell differentiation. Total proteins were separated and probed as represented in A, and then scanned bylaser densitometry. Relative activation levels of Akt were assessed at 22, 0, 5, 10, 15, 20, and 30 dayspostconfluence by determining total peak areas (AU 3 mm) for pp57Akt and total p57Akt, to establish the ratiospp57Akt/p57Akt. Data represent means 6 SE for at least 3 (n $ 3) separate and independent cultures, for eachculture stage studied. C: modulations of Akt activation in undifferentiated (open bars) and differentiated (solidbars) Caco-2/15 cell cultures maintained 48 h without serum (control) or with the addition of 10 mg/ml insulin (1I),300 mM genistein (1G), 20 mM PD-98059 (1PD), 30 mM Ly-294002 (1Ly), or 1 mM cytochalasin D (1CD). Totalproteins were resolved and probed for the detection of the activated form of Akt and total Akt as in A, and thenscanned as in B. For each treatment of cultures, the relative activation levels of p57Akt were compared with thoseof control cultures, 3100 (expressed as % of control). Columns represent means 6 SE for at least 3 (n $ 3) separateexperiments. *Statistically significant (0.001 # P # 0.01) differences between treatments and their respectivecontrols. Results illustrated here for undifferentiated Caco-2/15 cells were similar, if not identical, to thoseobtained with the undifferentiated cryptlike HIEC-6 cells (not shown).
C1549DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

For instance, the inhibition of MEK/Erk in differenti-ated Caco-2/15 cells resulted in a balance seemingly infavor of anti-apoptotic homologs (Table 1, 1PD), even ifthe treatment induced apoptosis (Fig. 2B, lane 5; Table
1, 1PD). Considering that cell survival implicates notjust one homolog but rather complex interactions be-tween numerous Bcl-2 homologs, as well as posttrans-lational modifications to further modulate their func-
Fig. 7. Differentiation state-specific modulations ofanti-apoptotic Bcl-2 homologs in Caco-2/15 cells. Rel-ative expression levels of Bcl-2 (A), Bcl-XL (B), andMcl-1 (C) in undifferentiated (open bars) and differ-entiated (solid bars) Caco-2/15 cell cultures main-tained 48 h without serum (control) or with the addi-tion of 10 mg/ml insulin (1I), 300 mM genistein (1G),20 mM PD-98059 (1PD), 30 mM Ly-294002 (1Ly), or1 mM cytochalasin D (1CD). Total proteins were sep-arated and probed as represented in Fig. 1, and thenscanned by laser densitometry. For each treatment ofcultures, the relative expression levels of each ho-molog were compared with those of control cultures,3100 (expressed as % of control). Columns representmeans 6 SE for at least 3 (n $ 3) separate experi-ments. *Statistically significant (0.001 # P # 0.01)differences between treatments and their respectivecontrols.
Fig. 8. Differentiation state-specific modulations ofproapoptotic Bcl-2 homologs in Caco-2/15 cells. Rela-tive expression levels of Bax (A), Bak (B), and Bad (C)in undifferentiated (open bars) and differentiated(solid bars) Caco-2/15 cell cultures maintained 48 hwithout serum (control) or with the addition of 10mg/ml insulin (1I), 300 mM genistein (1G), 20 mMPD-98059 (1PD), 30 mM Ly-294002 (1Ly), or 1 mMcytochalasin D (1CD). Total proteins were separatedand probed as represented in Fig. 1, and then scannedby laser densitometry. For each treatment of cultures,the relative expression levels of homolog were com-pared with those of control cultures, 3100 (expressedas % of control). Columns represent means 6 SE for atleast 3 (n $ 3) separate experiments. *Statisticallysignificant (0.001 # P # 0.01) differences betweentreatments and their respective controls.
C1550 DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

tions (1, 4, 20, 25, 47), such apparent discrepancies arelikely due to the involvement of additional homologsother than those analyzed herein (e.g., Bcl-w, Bid, etc.)and/or to Bcl-2 homolog phosphorylation events (1, 20,25, 45, 47, 59) not focused on in the present study.Nonetheless, these data altogether indicate that thesteady-state expression levels of Bcl-2 homologs arenot only subject to complex regulatory mechanisms inboth undifferentiated and differentiated intestinal ep-ithelial cells but that such mechanisms exhibit distinc-tions related to the state of differentiation.
DISCUSSION
In this study, we investigated the question ofwhether the regulation of human intestinal epithelialcell survival involves distinct control mechanisms de-pending on the state of differentiation, using the well-established human Caco-2/15 enterocyte-like andHIEC-6 cryptlike in vitro models. We found that theenterocytic differentiation process of Caco-2/15 cellsresults in the gradual establishment of differentiationstate-distinct profiles of 1) Bcl-2 homolog steady-statelevels and 2) p125Fak, p42Erk-2, and p57Akt activatedlevels. Accordingly, we found that the inhibition oftyrosine kinase activities, Fak, the MEK/Erk pathway,or PI3-K have distinct impacts on enterocytic cell sur-vival in undifferentiated (subconfluent Caco-2/15, con-fluent HIEC-6) and differentiated (30 days postconflu-ent Caco-2/15) cells. We also observed that exposure toinsulin and the inhibition of these various signalingmolecules/pathways modulated distinctively the ex-pression of each Bcl-2 homolog analyzed in both undif-ferentiated and differentiated Caco-2/15 cells; how-ever, sharp distinctions were noted between the twostates of differentiation in the resulting effects of thesame treatments on the expression of Bcl-2 homologs.Furthermore, we found that the PI3-K/Akt pathway isdistinctively regulated in undifferentiated and differ-entiated enterocytes. Finally, we have shown that Fakand b1 and b4 integrins, as well as the MEK/Erk andPI3-K/Akt pathways, are distinctively involved in en-terocytic cell survival, depending on the state of differ-entiation. Therefore, these data altogether indicatethat human intestinal epithelial cell survival is char-acterized by differentiation state-specific susceptibili-ties to apoptosis, which in turn are linked with distinc-tions in both the regulation of Bcl-2 homologs and theinvolvement of signaling molecules/pathways.
The intestinal crypt-villus axis is defined by prolif-erative and functional properties of the crypt cells thatdistinguish them from the fully differentiated villuscells (26, 37, 41, 44, 45). Previous in vivo studies in theadult rodent (26, 29–32, 38–40, 44, 45, 60) and human(26, 28–32, 39, 44, 45, 52, 53, 59) intestine have re-ported that Bcl-2 homologs exhibit gradients of expres-sion along this crypt-villus axis of enterocytic differen-tiation. To this effect, the data reported herein show forthe first time that such distinct Bcl-2 homolog expres-sion profiles are gradually established during the en-terocytic differentiation process. Because Bcl-2 ho-
mologs constitute a critical checkpoint in theregulation of apoptosis, it has been suggested thatintestinal epithelial cell survival may be regulateddistinctively according to the state of cell differentia-tion (22, 24, 26, 40, 44–46, 53). In support of this, it isnow well established that the predominant means toremove obsolete differentiated enterocytes is throughapoptosis and shedding at the villus apex, whereasspontaneous crypt cell apoptosis, a rarer (less frequent)process, serves to remove defective/injured progenycells (22, 26, 40, 44, 45, 53). Hence, in vivo studies haveconsistently reported a dramatic increase of apoptosisin crypt cells, but little or no increase in villus cells,after irradiation or chemotherapeutic drug exposure(24, 38, 44–46). Likewise, analyses of enterocytic apop-tosis in bcl-22/2 and bax2/2 knockout mice have re-ported differential consequences for crypt and villuscells, with regard to resistance and/or susceptibility toapoptosis after irradiation (44–46, 60). Incidentally,the present study provides further documentation onthe distinct susceptibilities to apoptosis between undif-ferentiated and differentiated enterocytes in vitro. Inaddition, our data demonstrate for the first time thatsuch distinctions are linked with a differentiationstate-specific regulation of Bcl-2 homolog expressionand involvement of integrins and signal transductionpathways. Finally, it has been recently reported (22)that villus cells exhibit distinct expression and activa-tion profiles of caspases, the major components of theapoptotic effector machinery (1, 25, 47). Consequently,and in light of these considerations, our data alto-gether clearly establish that human intestinal epithe-lial cell survival is subject to differentiation state-specific control mechanisms in vitro.
External stimuli responsible for the promotion of cellproliferation, differentiation, and/or survival largelycome from hormones/growth factors and cell adhesion(1, 4, 11, 13, 15, 19–21, 56). As with other cell types (4,10, 13, 19, 21, 25, 54, 55), the differentiation process ofenterocytes is accompanied by the establishment ofdifferentiation state distinctions in hormonal re-sponses (12, 16, 17, 27, 37, 43–45) as well as in theexpression of cell cycle regulators (2, 17) and cell ad-hesion components, such as integrins (7, 35, 37, 41,43–45), in addition to morphological and functionaldifferentiated characteristics. For example, while bothcrypt and villus cells express b1 integrins, differentia-tion state-distinct profiles of specific ab1 integrins arenonetheless expressed (6–7, 33, 35, 51); also, the b4integrin subunit is functional in differentiated entero-cytes but not in crypt cells (5, 7). In addition, signalingmolecules/pathways have been shown to play dualroles in enterocytes: for instance, the MEK/Erk path-way is required for proliferation of undifferentiatedcells and the onset of enterocytic differentiation (2).Similarly, PI3-K performs a major role in enterocyticde novo DNA synthesis and differentiation (9, 35).Taking these observations into consideration, it there-fore follows that 1) undifferentiated and differentiatedintestinal cells exhibit distinct profiles of activatedlevels of Fak (this study and Ref. 35), Erk-1/Erk-2 (this
C1551DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

study and Refs. 2 and 35) and Akt (this study) which, inthe case at least of the PI3-K/Akt pathway (this studyand Refs. 9 and 35), underlie as well distinct modes ofregulation depending on the differentiation state; and2) Fak and integrins b1 and b4, as well as the MEK/Erk and PI3-K/Akt pathways, exert distinct influenceson intestinal epithelial cell survival depending on thestate of cell differentiation (this study). In this respect,the establishment of differentiation state-specific pro-files of MEK/Erk and PI3-K/Akt activation has beenobserved in other cell types (10, 13, 15, 56). Further-more, a differentiation state-distinct involvement ofintegrins and/or signaling pathways in cell survivalhas been demonstrated in other cell types as well (4,11, 13, 15, 19, 21), such as neural (4, 13) and skeletalmuscle (54, 55) cells.
In the past few years, it has become increasinglyevident that the regulation of individual Bcl-2 ho-mologs can involve numerous pathways acting in syn-ergy or independently and that the specific pathwaysinvolved in the regulation of a single homolog can differdepending on the cell type and the differentiation statestudied (1, 4, 10, 11, 13–15, 19–21, 47, 49, 56). It nowappears that intestinal epithelial cells are no exceptionto this (this study and Refs. 23, 33, 39, 48). Indeed,exposure to butyrate induces apoptosis in undifferen-tiated intestinal cells by decreasing Bcl-2 levels with-out affecting those of Bax or Bak (48), whereas thesame treatment induces apoptosis in differentiatedcells by increasing Bak levels (39, 48). Similarly, rastransformation of rat crypt cells stimulates the PI3-K/Akt pathway but not the MEK/Erk pathway, resultingin decreased Bak levels (33). In addition to these, ourfindings that Fak, MEK/Erk, and PI3-K/Akt play dis-tinct roles in the modulation of individual Bcl-2 ho-molog expression either within undifferentiated anddifferentiated intestinal cells or between the two dif-ferentiation states illustrate well the complexity that isrequired in the regulation of the expression and func-tions of Bcl-2 homologs, depending on the cell type andstate of cell differentiation. This in turn furtherstresses the concept that cell survival is not regulatedby the activity of single Bcl-2 homologs but, rather, bya balance of activities from numerous homologs thatarises from the input of multiple stimuli and pathways(1, 4, 13, 14, 21, 25, 34, 47, 49).
It is now well acknowledged that signaling pathwayscan cross talk with each other to act in tight coordina-tion, although the exact mechanisms involved remainunclear (4, 13, 21, 56). For example, a close cooperationhas been reported between the MEK/Erk and PI3-K/Akt pathways in suppressing apoptosis in some celltypes, even to a point where both pathways can berequired to effectively sustain cell survival (4, 13, 14,21, 49, 56). On the other hand, it is becoming increas-ingly evident that cell survival is furthermore regu-lated through cell type-specific machineries of integra-tion and fine-tuning of signaling pathways, whichremain to be fully understood (1, 4, 13, 15, 20, 21, 56).For instance, recent observations indicate that theMEK/Erk and PI3-K/Akt pathways can also act coinde-
pendently in maintaining cell survival, with one beingable to compensate for the inhibition of the other (4,13–15, 34, 49, 56). To this effect, several lines of evi-dence now indicate that the relationship between theMEK/Erk and PI3-K/Akt pathways, as well as theirintegration and fine-tuning, are distinct depending onthe state of enterocytic differentiation: 1) the inhibitionof Fak impacts equally on the MEK/Erk and PI3-K/Aktpathways in undifferentiated and differentiated en-terocytes, and yet Fak and these two pathways none-theless display striking differentiation state-specificdistinctions in their involvement in enterocytic cellsurvival (this study); 2) the inhibition of the MEK/Erkpathway does not impact on the survival of undiffer-entiated cells (this study and Ref. 33) but does so fordifferentiated cells (this study); 3) the inhibition ofPI3-K causes apoptosis in both undifferentiated (thisstudy and Refs. 9 and 33) and differentiated (thisstudy) cells, albeit to a greater extent in the formerdifferentiation state (this study); 4) the inhibition of b1integrins causes a downactivation of Fak in both un-differentiated and differentiated enterocytes but, as forthe inhibition of Fak (this study), causes apoptosis to agreater extent in the latter cell state (this study); 5) theb1 integrin-mediated survival signaling in undifferen-tiated enterocytes appears to be PI3-K/Akt dependentbut not MEK/Erk dependent (33); 6) overexpression ofras protects crypt cells from anoikis by stimulating thePI3-K/Akt pathway and not the Raf/MEK/Erk pathway(33), even though ras is well known for its role inusually stimulating the latter pathway (2, 4, 11, 13, 21,34); 7) b4 integrins, which are known for their role inmaintaining the activation of the MEK/Erk pathwaythrough the recruitment of Shc and Grb2-SOS (7, 11,21), are involved in enterocytic cell survival only in thedifferentiated state (this study); and 8) p57Akt activa-tion appears to be PI3-K independent in undifferenti-ated enterocytes only (this study), an uncoupling situ-ation already shown to exist in other cell systems (13,15, 56). Consequently, these considerations altogethersuggest that both the MEK/Erk and PI3-K/Akt path-ways are required for the survival of differentiatedenterocytes, whereas PI3-K (with or without p57Akt) issufficient and able to compensate for the inhibition ofthe MEK/Erk pathway in the survival of undifferenti-ated enterocytes. Alternately, the MEK/Erk pathwaymay not play any role in the survival of undifferenti-ated intestinal cells.
In conclusion, the present findings provide new in-sights into the complex regulatory mechanisms thatare responsible for the survival of human intestinalepithelial cells. Such mechanisms are likely to varysomewhat in their specifics along the proximal-distalaxis of the gut, considering the differences in Bcl-2homolog expression profiles (28–32, 38, 40, 52, 53, 60),susceptibilities to apoptosis (24, 26, 38, 40, 44–46), andregulation of cellular functions (7, 16, 37, 41, 43) be-tween jejunum, ileum, and colon enterocytes in vivoand in vitro. Nonetheless, it is now clear that intestinalepithelial cells exhibit a differentiation state-specificsusceptibility to apoptosis through distinctions in the
C1552 DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

involvement of signaling molecules/pathways, such asFak, MEK/Erk, and PI3-K/Akt, which in turn impactdistinctively on the expression of Bcl-2 homologs. Ac-cordingly, cell adhesion components such as b1 and b4integrins also participate in the regulation of intestinalcell survival in a differentiation state-specific manner.However, the exact molecular processes responsible forsuch distinct control mechanisms of survival betweenundifferentiated and differentiated cells remain to befully understood. For example, the question is open asto why Akt is seemingly independent of PI3-K for itsactivation in undifferentiated intestinal cells. Con-versely, further analyses are required to dissect theexact molecular relationships and functions of theMEK/Erk and PI3-K/Akt pathways in the regulation ofenterocytic cell survival and Bcl-2 homolog expression.Increasing our knowledge on the specific roles of thesepathways in intestinal epithelial cell survival shouldprovide a better understanding of the role of apoptosis/anoikis in the maintenance and repair of the intestinalepithelium, as well as in the pathogenesis of intestinaldisorders with dysregulation of apoptosis, such as can-cer.
We thank Drs. E. Ruoslahti and E. Engvall (The Burnham Insti-tute, LaJolla, CA) for the kind gifts of the P4C10 and 3E1 antibodies,Dr. J.-F. Beaulieu for the gift of Caco-2/15 and HIEC-6 cells anduseful discussions, and Dr. N. Rivard also for useful discussions.
This work was supported by Canadian Institutes of Health Re-search Grant MGC-15186.
P. H. Vachon is a Chercheur-Boursier du Fonds de la Rechercheen Sante du Quebec and a Chercheur de la Fondation CanadiennePour l’Innovation.
REFERENCES
1. Adams JM and Cory S. The Bcl-2 protein family: arbiters ofcell survival. Science 281: 1322–1326, 1998.
2. Aliaga JC, Deschenes C, Beaulieu J-F, Calvo EL, and Ri-vard N. Requirement of the MAP kinase cascade for cell cycleprogression and differentiation of human intestinal cells. Am JPhysiol Gastrointest Liver Physiol 277: G631–G641, 1999.
3. Allesi DR, Cuenda A, Cohen P, Dudley DT, and Saltiel AR.PD098059 is a specific inhibitor of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J Biol Chem270: 27489–27495, 1995.
4. Anderson P. Kinase cascades regulating the entry into apopto-sis. Microbiol Mol Rev 61: 33–46, 1997.
5. Basora N, Herring-Gillam FE, Boudreau F, Perreault N,Pageot L-P, Simoneau M, Bouatrouss Y, and Beaulieu J-F.Expression of functionally distinct variants of the b4A integrinsubunit in relation to the differentiation state in human intesti-nal cells. J Biol Chem 274: 29819–29825, 1999.
6. Basora N, Vachon PH, Herring-Gillam FE, Perreault N,and Beaulieu J-F. Relation between integrin a7Bb1 expressionin human intestinal cells and enterocytic differentiation. Gastro-enterology 113: 1510–1521, 1997.
7. Beaulieu J-F. Integrins and human intestinal cell functions.Front Biosci 4: 310–321, 1999.
8. Beaulieu J-F and Quaroni A. Clonal analysis of sucrase-isomaltase expression in the human colon adenocarcinoma cellsCaco-2. Biochem J 280: 599–608, 1991.
9. Benistant C, Chapuis H, and Roche S. A specific function forphosphatidylinositol 3-kinase a (p85a-p110a) in cell survival andfor phosphatidylinositol 3-kinase b (p85a-p110b) in de novo DNAsynthesis of human colon carcinoma cells. Oncogene 19: 5083–5090, 2000.
10. Boucher M-J, Morisset J, Vachon PH, Reed JC, Laine J,and Rivard N. MEK/ERK signaling pathway regulates theexpression of Bcl-2, Bcl-XL, and Mcl-1 and promotes survival of
human pancreatic cancer cells. J Cell Biochem 79: 355–369,2000.
11. Coppolino MG and Dedhar S. Bi-directional signal transduc-tion by integrin receptors. Int J Biochem Cell Biol 32: 171–188,2000.
12. Cross HS and Quaroni A. Inhibition of sucrase-isomaltaseexpression by EGF in the human colon adenocarcinoma cellsCaco-2. Am J Physiol Cell Physiol 261: C1173–C1183, 1991.
13. Cross TG, Scheel-Toellner D, Henriquez NV, Deacon E,Salmon M, and Lord JM. Serine/threonine protein kinasesand apoptosis. Exp Cell Res 256: 34–41, 2000.
14. Danilkovitch A, Donley S, Skeel A, and Leonard EJ. Twoindependent signaling patways mediate the antiapoptotic actionof macrophage-stimulating protein on epithelial cells. Mol CellBiol 20: 2218–2227, 2000.
15. Datta SR, Brunet A, and Greenberg ME. Cellular survival: aplay in three Akts. Genes Dev 13: 2905–2927, 1999.
16. Drucker DJ. Epithelial cell growth and differentiation I. Intes-tinal growth factors. Am J Physiol Gastrointest Liver Physiol273: G3–G6, 1997.
17. Evers BM, Ko TC, Li J, and Thompson A. Cell cycle proteinsuppression and p21 induction in differentiating Caco-2 cells.Am J Physiol Gastrointest Liver Physiol 271: G722–G727, 1996.
18. Frisch SM. Methods for studying anoikis. In: Methods in Mo-lecular Biology: Integrin Protocols, edited by Howlett AR. To-towa, NJ: Humana, 1999, vol. 129, p. 251–256.
19. Frisch SM and Ruoslahti E. Integrins and anoikis. Curr OpinCell Biol 9: 701–706, 1997.
20. Gajewski TF and Thompson CB. Apoptosis meets signaltransduction: elimination of a BAD influence. Cell 87: 589–592,1996.
21. Giancotti FG and Ruoslahti E. Integrin signaling. Science285: 1028–1032, 1999.
22. Grossmann J, Mohr S, Lapetina EG, Fiocchi C, and LevineAD. Sequential and rapid activation of select caspases duringapoptosis of normal intestinal epithelial cells. Am J PhysiolGastrointest Liver Physiol 274: G1117–G1124, 1998.
23. Hague A, Dıaz GD, Hicks DJ, Krajewski S, Reed JC, andParaskeva C. Bcl-2 and Bak may play a pivotal role in sodiumbutyrate-induced apoptosis in colonic epithelial cells; however,overexpression of Bcl-2 does not protect against Bak-mediatedapoptosis. Int J Cancer 72: 898–905, 1997.
24. Hall PA, Coates PJ, Ansari B, and Hopwood D. Regulationof cell number in the mammalian gastrointestinal tract: theimportance of apoptosis. J Cell Sci 107: 3569–3577, 1994.
25. Jacobson MD, Weil M, and Raff MC. Programmed cell deathin animal development. Cell 88: 347–354, 1997.
26. Jones BA and Gores GJ. Physiology and pathophysiology ofapoptosis in epithelial cells of the liver, pancreas, and intestine.Am J Physiol Gastrointest Liver Physiol 273: G1174–G1188,1997.
27. Jumarie C and Malo C. Caco-2 cells cultured in serum-freemedium as a model for the study of enterocytic differentiation invitro. J Cell Physiol 149: 24–33, 1991.
28. Krajewska M, Moss SF, Krajewski S, Song K, Holt PR, andReed JC. Elevated expression of Bcl-X and reduced Bak inprimary colorectal adenocarcinomas. Cancer Res 56: 2422–2427,1996.
29. Krajewski S, Bodrug S, Krajewska M, Shabaik A, Gas-coyne R, Berean K, and Reed JC. Immunohistochemicalanalysis of Mcl-1 protein in human tissues: differential regula-tion of Mcl-1 and Bcl-2 protein production suggests a unique rolefor Mcl-1 in control of programmed cell death in vivo. Am JPathol 146: 1309–1319, 1995.
30. Krajewski S, Krajewska M, and Reed JC. Immunohisto-chemical analysis of in vivo patterns of Bak expression, a proapop-totic member of the Bcl-2 protein family. Cancer Res 56: 2849–2855, 1996.
31. Krajewski S, Krajewska M, Shabaik A, Miyashita T, WangHG, and Reed JC. Immunohistochemical determination of invivo distribution of Bax, a dominant inhibitor of Bcl-2. Am JPathol 145: 1323–1336, 1994.
32. Krajewski S, Krajewska M, Shabaik A, Wang H-G, Irie S,Fong L, and Reed JC. Immunohistochemical analysis of in
C1553DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from

vivo patterns of Bcl-X expression. Cancer Res 54: 5501–5507,1994.
33. Lee JW and Juliano RL. a5b1 Integrin protects intestinalepithelial cells from apoptosis through a phosphatidylinositol3-kinase and protein kinase B-dependent pathway. Mol Biol Cell11: 1973–1987, 2000.
34. Le Gall M, Chambard J-C, Breittmayer J-P, Grall D,Poussyegur J, and Van Obberghen-Schilling E. The p42/p44 MAP kinase pathway prevents apoptosis induced by anchor-age and serum removal. Mol Biol Cell 11: 1103–1112, 2000.
35. Levy P, Robin H, Kornprobst M, Capeau J, and Cherqui G.Enterocytic differentiation of the human Caco-2 cell line corre-lates with alterations in integrin signaling. J Cell Physiol 177:618–627, 1998.
36. Lipfert L, Haimovitch B, Schaller MD, Cobb JS, ParsonsJT, and Brugge JS. Integrin-dependent phosphorylation andactivation of the protein tyrosine kinase pp125FAK in platelets.J Cell Biol 119: 905–912, 1992.
37. Menard D, and Beaulieu J-F. Human intestinal brush bordermembrane hydrolases. In: Membrane Physiopathology, edited byBkaily G. Norwell: Kluwer Academic, 1994, p. 319–341.
38. Merritt AJ, Potten CS, Watson AJM, Yoh DY, NakayamaK-I, Nakayama K, and Hickman JA. Differential expressionof Bcl-2 in intestinal epithelia. Correlation with attenuation ofapoptosis in colonic crypts and the incidence of colonic neoplasia.J Cell Sci 108: 2261–2271, 1995.
39. Moss SF, Agarwal B, Arber N, Guan RJ, Krajewska M,Krajewski S, Reed JC, and Holt PR. Increased intestinal Bakexpression results in apoptosis. Biochem Biophys Res Commun223: 199–203, 1996.
40. Moss SF and Holt PR. Apoptosis in the intestine. Gastroenter-ology 111: 567–568, 1996.
41. Pageot L-P, Perreault N, Basora N, Francoeur C, Magny P,and Beaulieu J-F. Human cell models to study small intestinalfunctions: recapitulation of the crypt-villus axis. Microsc ResTech 49: 394–406, 2000.
42. Pinto M, Robine-Leon S, Appay MD, Kedinger M, TriadouN, Dussaulx E, Lacroix B, Simon-Assmann P, Haffen K,Fogh J, and Zweibaum A. Enterocyte-like differentiation andpolarization of the human colon carcinoma cell line Caco-2 inculture. Biol Cell 47: 323–330, 1983.
43. Podolsky DK. Regulation of intestinal epithelial proliferation: afew answers, many questions. Am J Physiol Gastrointest LiverPhysiol 264: G179–G186, 1993.
44. Potten CS. The significance of spontaneous and induced apop-tosis in the gastrointestinal tract of mice. Cancer Metastasis Rev11: 179–195, 1992.
45. Potten CS. Epithelial cell growth and differentiation II. Intes-tinal apoptosis. Am J Physiol Gastrointest Liver Physiol 273:G253–G257, 1997.
46. Pritchard DM, Potten CS, Korsmeyer SJ, Roberts S, andHickman JA. Damage-induced apoptosis in intestinal epitheliafrom Bcl-2-null and bax-null mice: investigations of the mecha-nistic determinants of epithelial apoptosis in vivo. Oncogene 18:7287–7293, 1999.
47. Reed JC, Miyashita T, Takayama S, Wang H-G, Sato T,Krajewski S, Aime-Sempe C, Bodrug S, Kitada S, andHanada M. Bcl-2 family proteins: regulators of cell death in-volved in the pathogenesis of cancer and resistance to therapy.J Cell Biochem 60: 23–32, 1996.
48. Rosen K, Rak J, Jin J, Kerbel RS, Newman MJ, and FilmusJ. Downregulation of the pro-apoptotic protein Bak is requiredfor the ras-induced transformation of intestinal epithelial cells.Curr Biol 8: 1331–1334, 1998.
49. Scheid MP, Schubert KM, and Duronio V. Regulation of Badphosphorylation and association with Bcl-XL by the MAPK/Erkkinase. J Biol Chem 274: 31108–31113, 1999.
50. Vachon PH, and Beaulieu J-F. Transient mosaic patterns ofmorphological and functional differentiation in the Caco-2 cellline. Gastroenterology 103: 414–423, 1992.
51. Vachon PH, and Beaulieu J-F. Extracellular heterotrimericlaminin promotes differentiation in human enterocytes. Am JPhysiol Gastrointest Liver Physiol 268: G857–G867, 1995.
52. Vachon PH, Cardin E, Harnois C, Reed JC, Plourde A, andVezina A. Early acquisition of bowel segment-specific Bcl-2homolog expression profiles during development of the humanileum and colon. Histol Histopathol 16: 497–510, 2001.
53. Vachon PH, Cardin E, Harnois C, Reed JC, and Vezina A.Early establishment of epithelial apoptosis in the developinghuman small intestine. Int J Dev Biol 44: 891–898, 2000.
54. Vachon PH, Loechel F, Xu H, Wewer UM, and Engvall E.Merosin and laminin in myogenesis: specific requirement formerosin in myotube stability and survival. J Cell Biol 134:1483–1497, 1996.
55. Vachon PH, Xu H, Liu L, Loechel F, Hayashi Y, Arahata K,Reed JC, Wewer UM, and Engvall E. Integrins (a7b1) inmuscle function and survival: disrupted expression in merosin-deficient congenital muscular dystrophy. J Clin Invest 100:1870–1881, 1997.
56. Vanhaesebroeck B and Alessi DR. The PI3K-PDK1 connec-tion: more than just a road to PKB. Biochem J 346: 561–576,2000.
57. Vlahos CJ, Matter WF, Hui KY, and Brown RF. A specificinhibitor of phosphatidylinositol 3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002). J Biol Chem 269:5241–5248, 1994.
58. Wary KK, Dans M, Mariotti A, and Giancotti FG. Biochem-ical analysis of integrin-mediated Shc signaling. In: Methods inMolecular Biology: Integrin Protocols, edited by Howlett AR.Totowa, NJ: Humana Press, 1999, vol. 129, p. 35–49.
59. Wilson JW, Nostro MC, Balzi M, Faraoni P, Cianchi F,Becciolini A, and Potten CS. Bcl-w expression in colorectaladenocarcinoma. Br J Cancer 82: 178–185, 2000.
60. Wilson JW and Potten CS. Immunohistochemical localizationof Bax and Bad in the normal and Bcl-2-null gastrointestinaltract. Apoptosis 1: 183–190, 1996.
61. Yu CF, Sanders MA, and Basson MD. Human Caco-2 motilityredistributes FAK and paxillin and activates p38 MAPK in amatrix-dependent manner. Am J Physiol Gastrointest LiverPhysiol 278: G952–G966, 2000.
C1554 DIFFERENTIAL REGULATION OF HUMAN INTESTINAL SURVIVAL
by 10.220.33.4 on Novem
ber 6, 2017http://ajpcell.physiology.org/
Dow
nloaded from